G Academic des sciences / Elsevier, Paris Anthropology / Anfhropologie Identification of Mycobacferium DNA in an Egyptian Pott’s disease of 5 400 years old Identificdtion dilDN de Mycobacterium dims un mul de POE &-yptien de 5400 ans Eric Cru b6zya*, Bertrand Ludes”, Jean-Dominique Povedac, John Claytond, Brigitte Crouau-Royd, Daniel Montagnone a Fkderation d’anthropologie, UMR 150 du CNRS, universite Toulouse-Ill, 39, allee Jules-Guesdes, 31000 Toulouse, France b lnstitut de medecine Iegale, 11, rue Humann, 67085 Strasbourg cedex, France ’ Centre de biologie medicale specialisee, lnstitut Pasteur, 28, rue du Docteur-Roux, 75724 Paris cedex 15, France d CNRS, CIGH UPR 8291, CHU Purpan, 31300 Toulouse, France e lnstitut d’embryologie, 1 1, rue Humann, 67085 Strasbourg cedex, France (Received 27 March 1997; accepted after revision 12 October 1998) Abstract - The antiquity of tuberculosis in the Old World is controversial because the morphology of the lesion in skeletal remains is non-specific. We report the recovery of a DNA fragment from a 5 400-year-old Predynastic Egyptian skeleton that exhibits a kyphotic, ‘hunchback’ spinal deformity consistent with Pott’s disease and suggestive of tuberculous vertebral involvement. The recovered DNA fragment was sequenced and is consistent with an original Mycobacterium sequence. We cannot prove that it is M. tuberculosis, M. bovis or an ancient mycobacteria resembling the two current forms because the observed modifications in the sequence could be attributed to the antiquity of Mycobacterium and/or to the effects of Taq polymerase. This provides the most specific evidence for the antiquity of human Mycobacterium disease in the world. (0 Academic des sciences / Elsevier, Paris.) ancient DNA / paleopathology / tuberculosis / anthropology Resume - L’anciennete de la tuberculose dans le Vieux Monde est controversee car la morphologie des lesions osseuses est non specifique. Nous presentons la dkouverte de fragments d’ADN prove- nant d’un squelette egyptien predynastique date de 5400 ans dont la colonne vertebrale presente des lesions et des deformations hocatrices dun ma1 de Pott et de tuberculose osseuse. Le sequen- Gage des fragments d’ADN est compatible avec celle d’une sequence de Mycobacterium. Nous ne pouvons pas savoir si c’est M. tuberculosis, M. bovis ou une ancienne mycobacterie qui leur ressem- blerait car les modifications de la sequence pourraient tout aussi bien etre attribuees a l’anciennete de la mycobacterie etlou aux effets de la Taq polymhase. Cela apporte la preuve formelle de l’anciennete des atteintes osseuses par Mycobacterium dans le monde. (0 Academic des sciences / Elsevier, Paris.) ADN an&n I p&opatbologie I tuberculose / anthropologic Note communicated by Yves Coppens *Correspondence and reprints E-mail: [email protected] C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

G Academic des sciences / Elsevier, Paris
Anthropology / Anfhropologie
Identification of Mycobacferium DNA in an Egyptian Pott’s disease of 5 400 years old
Identificdtion dilDN de Mycobacterium dims un mul de POE &-yptien de 5400 ans
Eric Cru b6zya*, Bertrand Ludes”, Jean-Dominique Povedac, John Claytond, Brigitte Crouau-Royd, Daniel Montagnone
a Fkderation d’anthropologie, UMR 150 du CNRS, universite Toulouse-Ill, 39, allee Jules-Guesdes, 31000 Toulouse, France
b lnstitut de medecine Iegale, 11, rue Humann, 67085 Strasbourg cedex, France
’ Centre de biologie medicale specialisee, lnstitut Pasteur, 28, rue du Docteur-Roux, 75724 Paris cedex 15, France
d CNRS, CIGH UPR 8291, CHU Purpan, 31300 Toulouse, France
e lnstitut d’embryologie, 1 1, rue Humann, 67085 Strasbourg cedex, France
(Received 27 March 1997; accepted after revision 12 October 1998)
Abstract - The antiquity of tuberculosis in the Old World is controversial because the morphology of the lesion in skeletal remains is non-specific. We report the recovery of a DNA fragment from a 5 400-year-old Predynastic Egyptian skeleton that exhibits a kyphotic, ‘hunchback’ spinal deformity consistent with Pott’s disease and suggestive of tuberculous vertebral involvement. The recovered DNA fragment was sequenced and is consistent with an original Mycobacterium sequence. We cannot prove that it is M. tuberculosis, M. bovis or an ancient mycobacteria resembling the two current forms because the observed modifications in the sequence could be attributed to the antiquity of Mycobacterium and/or to the effects of Taq polymerase. This provides the most specific evidence for the antiquity of human Mycobacterium disease in the world. (0 Academic des sciences / Elsevier, Paris.)
ancient DNA / paleopathology / tuberculosis / anthropology
Resume - L’anciennete de la tuberculose dans le Vieux Monde est controversee car la morphologie des lesions osseuses est non specifique. Nous presentons la dkouverte de fragments d’ADN prove- nant d’un squelette egyptien predynastique date de 5400 ans dont la colonne vertebrale presente des lesions et des deformations hocatrices dun ma1 de Pott et de tuberculose osseuse. Le sequen- Gage des fragments d’ADN est compatible avec celle d’une sequence de Mycobacterium. Nous ne pouvons pas savoir si c’est M. tuberculosis, M. bovis ou une ancienne mycobacterie qui leur ressem- blerait car les modifications de la sequence pourraient tout aussi bien etre attribuees a l’anciennete de la mycobacterie etlou aux effets de la Taq polymhase. Cela apporte la preuve formelle de l’anciennete des atteintes osseuses par Mycobacterium dans le monde. (0 Academic des sciences / Elsevier, Paris.)
ADN an&n I p&opatbologie I tuberculose / anthropologic
Note communicated by Yves Coppens
*Correspondence and reprints E-mail: [email protected]
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951

6. Crubky et al.
Version abrCgCe
1. Introduction
Alors que la tuberculose est restee d’actualite, ses origines font toujours I’objet de discussions car aucune l&ion osseuse n’esr pathognomonique de la maladie, Toutefois, son dia- gnostic par amplification de fragments de 1’ADN de la myco- bacterie extraits des l&ions osseuses esr possible. Nous prCsentons I’amplification et le sequenqage de 1’ADN d’une mycobacr&ie extrait d’un ma1 de Pott - tuberculose osseuse
vertebrale d’un squelette d’enfant egyptien, d’epoque predy- nastique (Nagada IId2, environ 3400 ans av. J.-C.).
2. Mat&iel et mCthodes
2.1. Extraction de 1’ADN
Au total, 1,45 g de cBte et 2,5 g de vertPbre provenant du mal de Pott furent abras& en surface puis Ccrases en petits fragments (0,2-0,3 cm3) et pilts dans un mortier en utilisant de l’azote liquide comme solution de refroidissement. La poudre d’os a Ctk peste sur un papier aluminium et placCe dans un tube sttrile etiquete de 50 mL en polycarbonate avec un bouchon visse. Les tchantillons furent incubts dans 40 mL de tampon (100 mM Tris-HCl, pH 7,6, 5 mM EDTA, 2 % SDS, 0,3 M acetate de sodium, 800 pg pro&- nase K/mL ) g 42 “C durant 21 h. AprPs dialyse au Tris- EDTA (0,l M, pH 8) durant 29 h, les echantillons furent centrifuges (3 000 tours/min pendant 10 min) puis precipi- t& dans l’bthanol absolu a -20 “C pendant 12 h. Le pr&ipitC fut remis en solution dans l’eau distillee et centrifugt g 13 000 tours/min durant 45 min, purifie dans une solution organique (phenol/ chloroforme/ isoamyl alcool, 25/24: 1, v/v) puis precipite avec 0,l vol de 3 M d’ac&ate de sodium B pH 8 et 3 vol d’ethanol absolu. Sur 10 % de l’extrait obtenu, une Clectrophorbe fut rtalisie dans un gel contenant 1 % SeaKem et des fragments d’ADN de tailles diffbrentes qui ser- vaienc de marqueurs et du bromide d’tthidium afin de pou- voir visualiser 1’ADN sous U.V.
Deux sCquences furent amplifiees : a) celle codant pour le gene de l’antigene 65 kD des mycobacteries ; b) l’&ment d’insertion IS6110, dont lo-15 copies existent chez M. tuberculosis et I-4 copies chez M. bovis. Les amorces TBl/TB2 furent utilisCes pour l’amplification du gene 65 kD. Celle-ci fut suivie d’une hybridation avec une sonde interne oligonu&otide marquee au gamma- 32P. Une ampli- fication nichee fut aussi rtalisee avec comme amorces exter- nes TBl/TB2, comme amorces internes TB28/TB29, et avec TBlO comme sonde interne Pour l’amplification de l’&- merit d’insertion IS 6110, trois types d’amorces et de sondes furent utilists : a) les amorces ISTB2/ISTB7 suivit d’une hybridation avec une sonde oligonuclCotide marquee IS2, b) les amorces ISl/IS2 suivit d’une hybridation avec une sonde oligonucleotide BKl marquee au gamma-32P et c) une PCR nichee dans un seul tube en utilisant comme amorces exter- nes J et K comme amorces internes ISI /IS2 et comme sonde interne pour l’hybridation BKl. Le sequenqage fut r&&se
grfce au kit Dye Deoxy Terminator d’Applied Biosystem@ et au sequenceur Applied Biosystem 373A selon les recomman- dations du fabricant. Pour tviter tout risque de contamina- tion par de I’ADN moderne, de multiples p&cautions ont et6 prises et des contrales negatifs (sans ADN) ont eW realists B chaque &tape. Les sequences obrenues furent comparees g cel- les des souches de rt!f&ence de M. tuberculosis H37Rv, M. bovis var. BCG et celles du gene de 65 kD disponible pour d’autres mycobact&ies ainsi qu’8 celles des plus proches, chez les champignons, les batteries et les hommes (HSP 70). L’idenrite des sequences proteiques fut aussi recherchee. L’ensemble des arbres phylog&&iques racines possibles a iti examine et, pour chacun, la sequence ancestrale, qui mini- mise le nombre de mutations et la variance du nombre de mutation, a et6 dCfinie.
3. Rkltats
Au total, 50 ng d’ADN ont et& extraits du fragment de c&e et 10 ng de celui de la vertkbre. L’klectrophorPse des ampli- ficats rtalises avec les amorces TBlITB2, pour le gtne de l’antigtne 65 kD des mycobact&ies, montre des fragments de la longueur attendue (343 bp) mais le signal apres hybrida- tion avec la sonde TBlO est faible. Sur les neuf PCR nichees rPali&es B partir de la msme PCR avec les amorces internes TB28/TB29,4 (no 7,9, 10, 17) prCsentent un signal B I’&?- trophorkse (exemple 2c de lajgure 2) de la longueur atten- due (133 bp) mais l’hybridation avec la sonde TBlO n’a don& aucun rCsultat. L’ADN amplifie fut s&quenc& Le frag- ment de la PCR no 7 est celui qui ressemble le plus aux sCquences Mycobacterium de refkrence, pour la longueur (I33 bp), comme pour le nombre de transitions et de trans- versions Les arbres racinis prCsentant le nombre minimum de mutations (73) et une variance faible (1,2) moncrent que I’ADN provenant de la PCR no 7 se regroupe avec les autres sCquences de type mycobacteries, la sbquence H37 &ant tou- jours plus tloignte. Les arbres suivants presentent essentiel- lement un changemenr au niveau du regroupement BCG, 65K et H37RV.
Seul I’ADN extrait de la vertebre a permit une amplifica- tion detectable sur gel avec les amorces ISTBZ/ISTB-/. 11 s’hybride avec la sonde IS2 mais le signal est plus petit que ce qui etait attendu. La PCR rCalis&e sur 1’ADN extrait de la ver- tebre avec les amorces IS 1 /IS2 et la PCR nichee utilisant J/K et les amorces ISI/IS2 a fournit un fragment de la longueur attendue, mais il ne s’est pas hybrid6 avec la sonde BKl et son sCquen$age tchoua.
4. Discussion
Le fragment provenant de la PCR no 7 a une sequence pro- the de celle attendue. Les diffirences et nommment les tran- sitions entre AT er GC pourraient Ctre reliCes a) aux effets de la Taq polym&ase lors de son utilisation avec de 1’ADN ancien, done degrade mais il faudrait alors cloner, b) etlou B l’anciennete de la mycobactCrie qui pourrait avoir CtC une espece intermediaire en M bovis et Ad. tuberculosis. 11 en est de m?me pour l’impossibilite d’hybrider correclement la
942 C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951

5 400-year-old Mycobacferium DNA
sonde TBlO aux fragments amplifies. MCme en augmentant le nombre de cycles de I’amplification, lors de la PCR nichee avec les amorces TB28/TB29, le nombre de fragments ampli- fies visibles sur le gel de l’electrophoreseest plus important mais aucune hybridation n’est obtenue. 11 faut signaler qu’apres amplification avec les amorces TBl/TB2 la sonde oligonucltotide TBlO utilisee dans l’hybridation peut diffe- rencier des molecules d’ADN de mycobacteries d’especes t&s proches done les sequences ne different que de quelques nuclCotides.
L’aspect anatomique des pieces est tout B fait similaire a celui des tuberculoses modernes d’avant l’ere des antibioti- ques et il signe mCme la presence d’un ou deux abets para- vertebraux et une survie assez longue du sujet entre le debut
et la fin de sa maladie. Un autre cas de ma1 de Pott, cicatrise celui la, a et6 signal6 dans la necropole. Precedemment, un bacille tuberculeux avait et-6 reconnu histologiquement dans un poumon de momie tgyptienne da&e entre 1000 B.C. et AD 400 et la sequence codant pour le gene de l’antigirne 65 kD a et& amplifiee dans I’ADN extrait d’un autre poumon de momie date de 1550 a 1080 BC. Le cas decrit ici est done le plus vieux cas de tuberculose formellement identifit, il est le resultat, soit d’une infection a M. bovis ou M. tuberculosis,
soit d’une infection par une mycobacterie plus proche de M. tuberculosis que de M. bovis. Cette dtcouverte est en accord avec les differences genetiques entre les mycobacteries, qui suggerent que la tuberculose aurait pu apparaitre il y a plus de 15 000 ans.
1. Introduction
Tuberculosis not only remains a major cause of mor- bidity and mortality in developing countries, but recent years have witnessed a resurgence in the incidence of tuberculosis in the United States and Europe with recent strains highly resistant to drug treatment [l]. It is a chronic infectious disease caused by Mycobacterium tuberculosis
of the human or bovine type, with visceral and sometimes skeletal lesions. Tuberculosis is associated with high morbidity and mortality, particularly among populations suffering from malnutrition and immunosuppression, characteristics of a large percentage of the third world. It is a leading cause of death due to an infectious agent world- wide and estimates suggest that as much as one third of the world’s population may be infected [I]
The origin of tuberculosis as a human disease has been the topic of speculation [2]. Reliable reconstruction of infectious disease patterns in antiquity depends on the recovery and identification of the infectious agent directly from human remains 131. This has only recently been made possible, modern methods ]4] based on DNA amplification by polymerase chain reaction [5] permitting the effective diagnosis of tuberculosis directly from archaeological tissues [3]. While DNA normally degrades rapidly after death [6] owing to the action of DNAses released at this time, the cell walls of mycobacteria pro- tect the bacterial DNA until these enzymes become inac- tivated. Salo et al. [3] have recently been successful in extracting, amplifying and sequencing ancient DNA of Mycobacterium tuberculosis from the lung tissue of a spontaneously mummified pre-Columbian Peruvian body about 1 000 years old. Using identical primers, Spigel- man and Lemma [7] were able to extract DNA from apparently tuberculosis-infected bones from medieval Europe, Turkey and pre-European-contact Borneo, but were unable to sequence these fragments.
We had the opportunity to attempt the recovery of DNA from a 5 400-year-old skeleton of Predynastic Egypt
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951
showing classical morphological indications suggestive of tuberculous involvement. The precise identification of tuberculosis in the skeletal record is hampered by the ambiguous nature of the morphological diagnostic crite- ria available in the archaeological record. For example, the spinal deformity called ‘Pott’s disease’, secondary to tuberculous vertebral involvement, which has typically been used to identify the most ancient cases of skeletal tuberculosis in the Old World (circa 4 500 BC) [81 can be caused by a compression fracture, brucellosis, osteomy- elitis or by a variety of fungi, such as Coccidioides immitis
[3, 91. Causal can only be ascertained by direct evidence of the presence of Mycobacterium.
2. Materials and methods
2.1. Skeleton remains
In the present study, we had the opportunity to examine the nearly complete skeleton of a 12-14-year-old child showing classical evidence of Pott’s disease (figure 7).
This individual was exhumed from a tomb (T 35) in the Predynastic necropolis of Ada’ima, located in the district of Esna (Upper-Egypt), where more than 200 graves have been excavated since 1990 [lo]. Associated with the skel- eton was pottery typical of Nagada IID2, dating between 3400 and 3300 BC. This period was a time of increasing urbanization, as the settlement centres of Upper-Egypt, such as Ada’ima, expanded into large towns or villages with considerable use of mud-brick architecture. Three asymmetrically collapsed vertebral bodies of the eighth to tenth thoracic vertebrae and the proximal part of the eighth left rib with periosteal new bone formation were the focus of the study.
2.2. DNA Extraction
Samples of 1.45 g of rib and 2.5 g of the vertebral frag- ment containing the lesion were cleaned with fine emery paper to remove as much material as possible from the
943

E. Crubky et al
Figure 1. The vertebral lesion is centred between T8 (at the top of the
photograph) and Tl 0, with fusion at their apophyses.
Severe kyphosis and scoliosis is evident along the vertebral column.
Despite these lesions, the neural canal is not affected. The eighth to
tenth thoracic vertebrae exhibit significant destruction. The body of
T8 is collapsed with erosion of the pedicles. The superior plateau is
intact and has a normal vertical anatomical orientation. There is a
complete loss of the body ofT9 with only the left pedicle and lamina
remaining intact. Finally, there is massive destruction of the body of
TlO with assymetric collapse of the anterior part, more pronounced
on the left side than on the right. Evidence of lesions are found
throughout the vertebrae from T6 to L4. These include an anterior
cavitation of T7, an extensive destruction without collapse, of the
body of Tl 1 and T12. In T6 and Ll, periosteal new bone formation
covers part of the anterior and lateral faces of the bodies including
the proximal part of the twelth right rib. Finally, a periosteal reaction on the proximal inferior part of the twelth right rib is evident.
outside surface of the bones to minimize contamination. After cleaning, the specimens were pulverized into small fragments (0.2-0.3 cm3) and ground in a mortar using liq- uid nitrogen as a freezing solution to avoid melting any DNA that might be present. The bone powder was weighed on disposable aluminium foil and placed in a labelled, sterile, 50 mL polycarbonate tube with a screw- cap. The samples were incubated in 40 mL of lysis buffer (100 mM Tris-HCl, pH 7.6, 5 mM EDTA, 2 % SDS, 0.3 M sodium acetate, 800 pg proteinase K/mL ) at 42 “C for 21 h. After dialysis with Tris-EDTA (0.1 M, pH: 8) for 29 h, the
samples were centrifuged (3 000 rpm/lO min) and pre- cipitated in absolute ethanol at -20 “C for 12 h. The pre- cipitate was resuspended in distilled water and centrifuged at 13 000 rpm for 45 min, and then purified with an organic procedure (phenol/chloroform/isoamyl alcohol, 25:24: 1, v/v) followed by precipitation with 0.1 volume of 3 M sodium acetate pH 8 and 3 volumes of absolute ethanol. Ten per cent of the extracts were elec- trophoresed through 1 % SeaKem gel with DNA size marker gels and stained with ethidium bromide to visu- alize the DNA under UV light. In addition to fragments of expected size, cf. infra, two other observations were nota- ble: a) infrequently, a blue fluorescence was noted in ethidium bromide gels examined under UV transillumi- nation, thought to be caused by soil-derived humic acids in the bone DNA extracts [l 11; b) light weight molecular DNA (I 14 bp) is observed on the electrophoresis gels of bone extracts. These have been attributed to micro-organ- isms [12] and the specificity of the sequence was per- formed with the sequencing.
2.3. PCR
The DNA extract was then subjected to PCR. Two mycobacterial sequences were targeted: i) the sequence coding for the mycobacterial 65 kD antigen [13] (figure 2), present in a single copy in the mycobacterial genome; ii) the insertion element IS61 10 [14], present as 1 O-l 5 copies in M. tuberculosis and 1-4 copies per cell in M. bovis [I 51 (figure 3).
For the amplification of the 65 kD protein gene, T&l/ TB2 primers [16] were used, followed by hybridization with an internal gamma-32P labelled oligonucleotide probe TBlO. In addition, nested amplification was also used with TBl/TB2 as external primers and TB28/TB29 as internal primers, with TBlO as an internal probe [17] (figure 4).
For the amplification of the IS61 10 insertion element, three sets of primers and probes were used: a) ISTB2/ ISTB7 primers followed by hybridization with a peroxi- dase labelled IS2 oligonucleotide probe [4], b) ISl/lS2 primers followed by hybridization with a gamma-32P labelled oligonucleotide BKl probe 1181, and c) nested PCR in a single tube [I 91 using J and K as external primers [201, ISl/lS2 as internal primers and BKl as an internal probe for hybridization.
2.4. Sequencing
PCR products were bidirectionally sequenced using an automatic sequencer (Applied Biosystem 3734) and Taq Dye Deoxy TM terminator cycle sequencing and protocols from Perkin Elmer. The primers used for the sequencing are those used for the amplification reaction. Templates were prepared from gel purified fragments and were sin- gle stranded. The nested PCR products as well as the cor- responding sequences from &CC and H37 reference strains were directly sequenced using the two primers TB28 and TB29 separately (each sequencing was per-
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998. 321,941-951

5 400.year-old Mycobacterium DNA
2a
1 7
; 4 5 6 7 8 9 10 11 12 13 14 15
2b
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
2c
1 2 3 4 5 6 7 8 9 10 11 12 13
2d
4
5
6
7
8
9
10
11
12
13
Figure 2. Amplification of the gene encoding the 65 kD antigen
PCR was performed in a 100 pL reaction mixture containing 100 pg/mL bovine serum albumine per pL, 10 mM Tris-HCI (pH 8.31, 1.5 mM M&I,,
1 pL:mL gelatin, 0.250 nM of TBl/TBZ primers, 2.5 units of Taq polymerase (Perkin-Elmer) and 200 PM of each dNTP.
The 40 cycle programme was performed at 95 “C for 120 s., 60 “C for 120 s., 72 “C for 120 s on a Perkin-Elmer DNA Thermal cycler. For nested
PCR 12~ and 2dl, internal primers were used to re-amplify 1 pL of the first stage reaction with the same programme conditions. Fragments were
visualized after electrophoresis (2a and 2c) and hybridization (2b and 2d) with the “P labelled T&l 0 probe.
2a (Fragments after electrophoresis) and 2b (fragment aiter hybridization for 7 h). Lane 4: vertebra fragment DNA, lane 1: positive control M. bovis
BCG DNA, lanes 2, 3, 5, 6, 7: 8, 9, 10, 11, 12, 13, 14, 15: negative PCR controls.
2c (Nested PCR, fragment after electrophoresis) and 2d (nested PCR, fragment after hybridization). The high concentrations of primer-dimers can
be seen (according to the 200 uM of each dNTPi. Length fragment on the gel is 133 bp. Lanes 3, 4, 9 and 10: vertebra iragment DNA (lane 3:
PCR no. 10; lane 4: PCR no. 9; lane 9: PCR no. 8; lane 10: PCR no. 7), lanes 1, 2, 6, 7, 12 and 13: positive controls, M. bovis BCC or ,%1. tuber-
culosis H37Rv DNA, lanes 5, 8 and 1 1: nested PCR from negative controls.
formed twice). The obtained sequences were compared using the BESTFIT and PILEUP programs (KG Software Package) and aligned with the region nt. 650-786 from the sequence register at the bank EMBL (Ml 5467), cor- responding to a portion of cDNA of the 65 kD antigen from M. tuberculosis (figure 4). We took care to avoid contamination of the lanes with the positive controls
2.5. Phylogenetic analyses
To further investigate the relationship of the studied DNA sequence to contemporary Mycobacterium DNA variations, phylogenetic tree reconstructions were per- formed {figure 5). All possible rooted trees were examined and, for each, the ancestral sequence was defined that
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321.941-951

i. Crub&y et al
3a 3b 3c 3d
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
1 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
2
3
II
4
5
6
7
8
9
10
11
12
13
14
15
Figure 3. Amplification of the insertion element IS61 10.
PCR (3a and 3b, DNA of the vertebra) was performed with ISTB2/lSTB7 primers. For nested PCR (3c and 3d! J/K were used as external primers
and ISl/lS2 as internal primers and reactions were conducted in a single tube with a drop of mineral oil separating the two reaction mixtures [18].
PCR programmes were as previously described. Fragments were visualized after electrophoresis (3a and 3~) and hybridization with the peroxidase
labelled IS2 probe (3b) or the 32P labelled BKl probe (3d).
3a and 3b. Lanes 2 and 5: positive controls: M. bovis BCC DNA, lanes 10 and 11: vertebra fragment DNA, lanes 1, 3, 4, 6, 7, 8, 9, 12, 13, 14,
15: negative controls.
3c and 3d. Lane 2: positive control: M. bovis BCG DNA, lanes 9 and 10: vertebra fragment DKA (5 and 10 ng), other lanes: negative controls.
minimized both the number of mutational events and the
variance of the number of mutations between the ances-
tral sequence and the observed sequences. The programs
to perform this analysis were written in C++.
2.6. General laboratory procedures
Several precautions were taken to guard against con-
tamination from contemporary DNA: a) the sampling dis-
sections were carried out in a room with no previous
exposure to mycobacteria; b) the extractions were per-
formed in another separate room with similar history; c)
all reusable labware (mortar and pestle) were soaked
overnight in 0.6 M HCI, thoroughly rinsed with Milli Q
treated (Millipore) water, wrapped in aluminium foil and
baked at 180 “C for 12 h; d) sterile disposable labware
was used whenever possible, including positive displace-
ment pipettes; e) various components of the extraction
buffers were autoclaved and blank extractions and rea-
gent blanks (negative controls) were run alongside the tis-
sue extracts. The PCR was handled in two separate
stations: one for the set up, one for the PCR itself,
equipped with a hood for the work-up of the reactions.
Each station was equipped with its own set of essential
equipment and these were not interchanged. Gloves were
worn at all times and always changed before going from
one area to the next. Contemporary M. tuberculosis
H37Rv and M. bovis var. BCG were prepared in a sepa-
C. R Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951
5 400.year-old Mycobacterium DNA
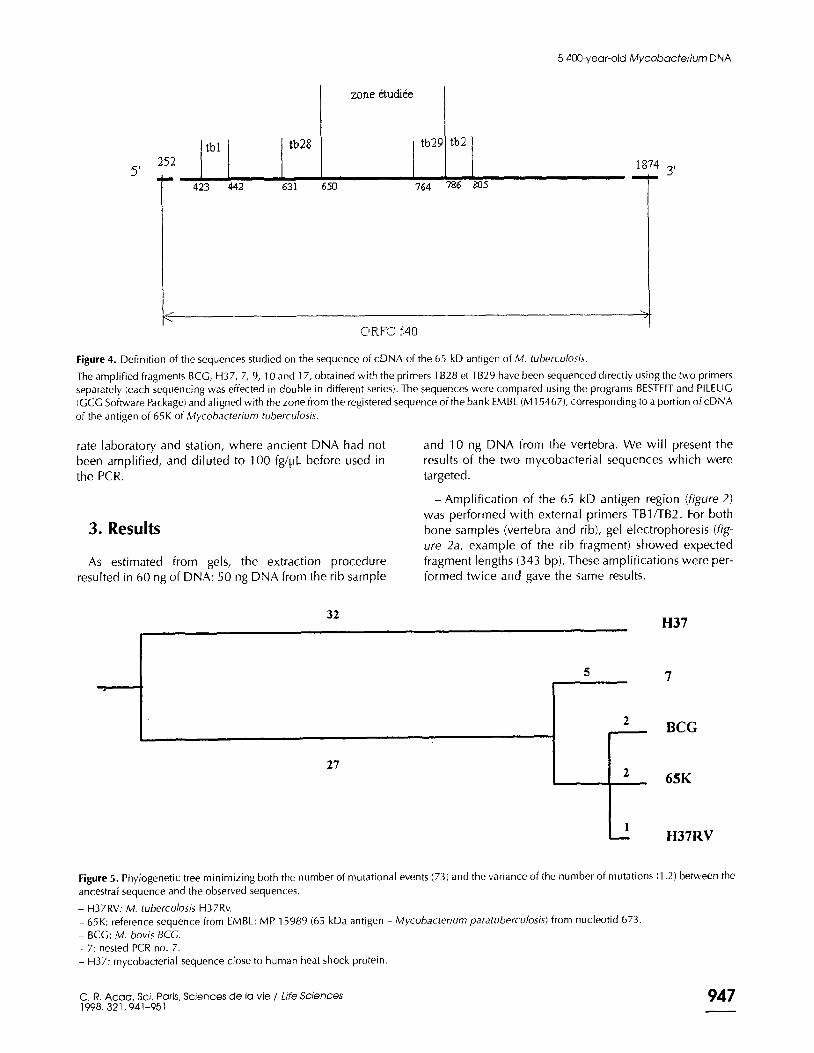
Figure 4. Definition of the sequences studied on the sequence of cDNA of the 65 kD antigen of M. tuberculosis.
The amplified fragments BCG, H37, 7, 9, 10 and 17, obtained with the primers TB28 et TB29 have been sequenced directly using the two primers
separately (each sequencing was effected in double in different series). The sequences were compared using the programs BESTFIT and PILEUC
(GCC Software Package! and aligned with the zone from the registered sequence of the bank EMBL (Ml 54671, corresponding to a portion of cDNA
of the antigen of 65K of Mycobacterium tuberculosis.
rate laboratory and station, where ancient DNA had not been amplified, and diluted to 100 fg/pL before used in the PCR.
3. Results
As estimated from gels, the extraction procedure resulted in 60 ng of DNA: 50 ng DNA from the rib sample
and 10 ng DNA from the vertebra. We will present the results of the two mycobacterial sequences which were targeted.
- Amplification of the 65 kD antigen region (figure 2) was performed with external primers TBl/TB2. For both bone samples (vertebra and rib), gel electrophoresis (fig- ure 2a, example of the rib fragment) showed expected fragment lengths (343 bp). These amplifications were per- formed twice and gave the same results.
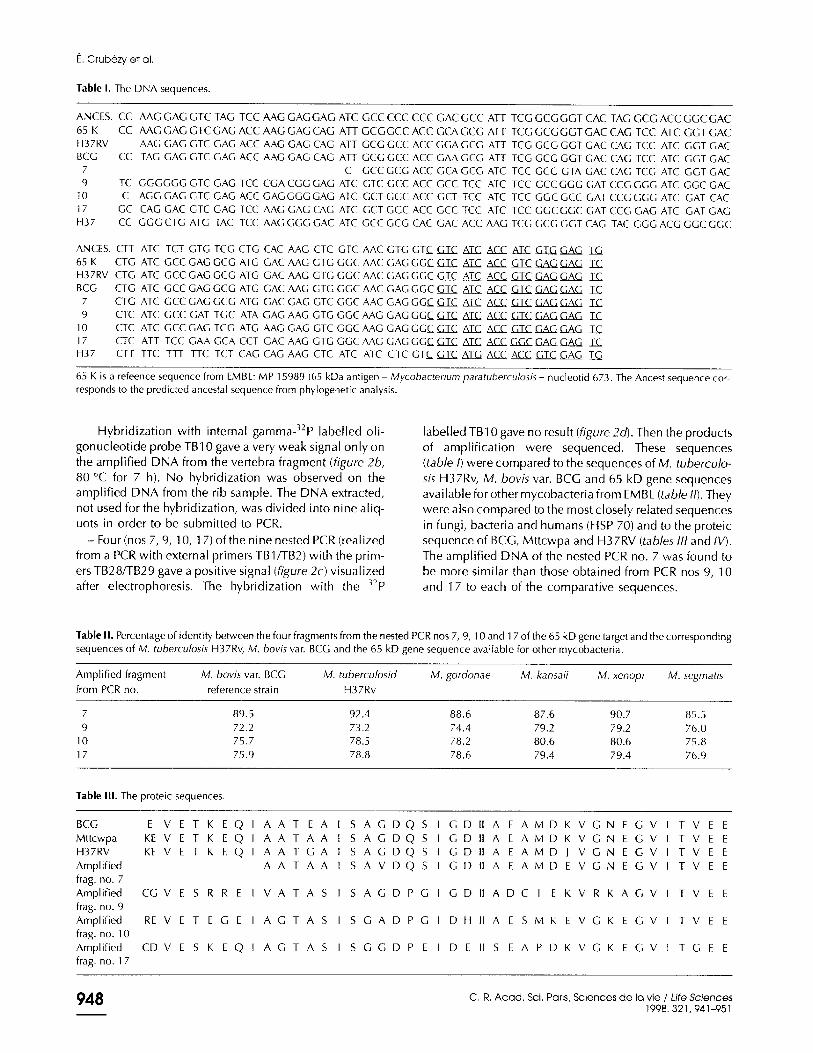
32 H37
5 7
2 BCG
27 2 65K
L 1 H37RV
Figure 5. Phylogenetic tree minimizing both the number of mutational events (73) and the variance of the number of mutations (1.2) between the
ancestral sequence and the observed sequences.
- H37RV: M. tuberculosis H37Rv.
- 65K: reference sequence from EMBL: MP 15989 (65 kDa antigen - Mycobacterium paratuberculosis) from nucleotid 673. - BCG: M. bovis BCG. - 7: nested PCR no. 7. - H37: mycobacterial sequence close to human heat shock protein.
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 19X321,941-951
i. Crubhy et al.
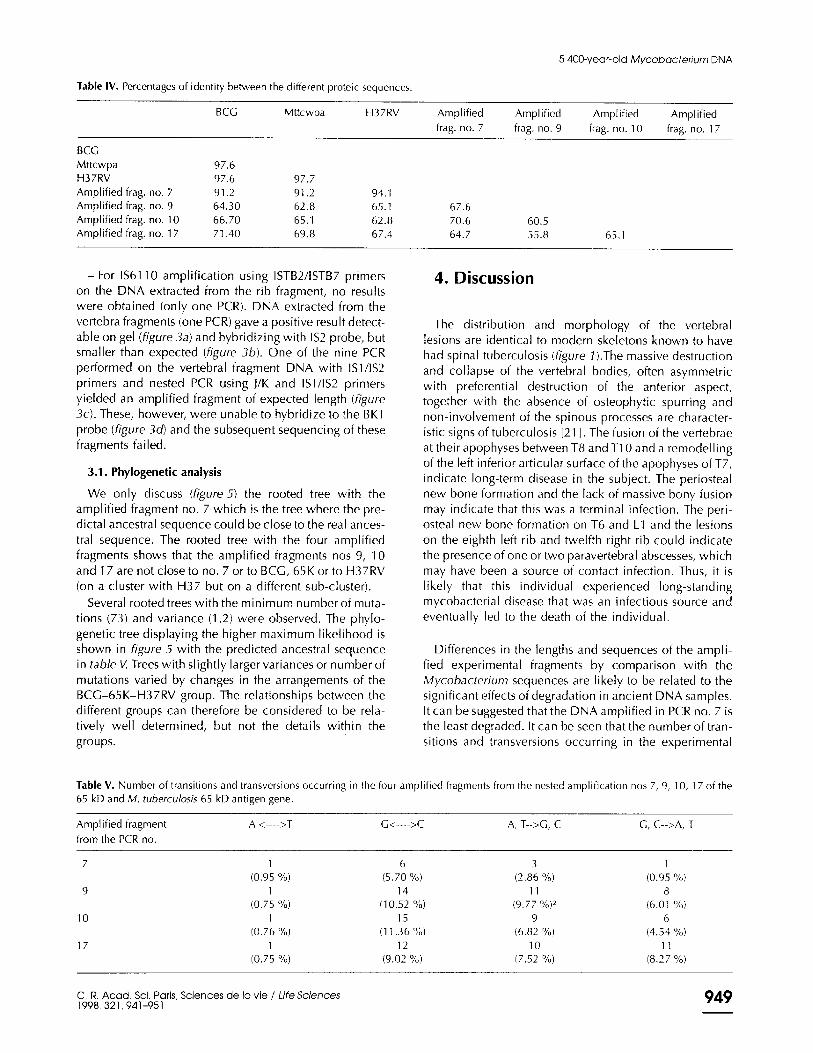
Table I. The DNA sequences.
ANCES. CC AAC GAG GTC TAG TCC AAG GAGGAG ATC CCC CCC CCC GAC CCC ATT TCC GCGGGT CAC TAG KG ACC GGCGAC 65 K CC AAG GAG GTC GAG ACC AAG GAG CAG ATT GCCGCC ACC GCA CCC ATT TCG GCC GGT GAC CAG TCC ATC GCT GAC H37RV AAG GAG GTC GAG ACC AAG GAG CAG ATT CCC CCC ACC CGA CCC ATT TCG CCC GGT GAC CAG TCC ATC GGT GAC BCG CC TAG GAG GTC GAG ACC AAG GAG CAG ATT CCC CCC ACC CAA GCG ATT TCG GCG GGT GAC CAG TCC ATC GGT CAC
7 C CCC CCC ACC GCA CCC ATC TCC CCC GTA GAC CAG TCG ATC GCT GAC 9 TC GGGGGG GTC GAG TCC CGA CGG GAG ATC GTC CCC ACC CCC TCC ATC TCC CCC CCC CAT CCC CCC ATC CCC GAC
10 C ACG GAG CTC GAG ACC GAGGGG GAG ATC GCT GGC ACC GCT TCC ATC TCC CCC GCC CAT CCC GGG ATC CAT CAC 17 CC CAC GAC GTC GAG TCC AAG GAG CAG ATC GCT CGC ACC CCC TCC ATC TCC GGCGGC CAT CCC GAG ATC CAT GAG H37 CC GGG CTG ATG TAC TCC AAG GGG GAC ATC GCC GCG CAC GAC ACC AAG TCC GCG GGT CAG TAC GGG ACG CCC GGC
ANCES. CTT ATC TCT GTG TCG CTG CAC AAG CTC GTC AAC GTG GTL: !ZIC BI(1 ACf ATC !ZLCU.Ci II; 65 K CTG ATC CCC GAG GCG ATG GAC AAG GTG GGC AAC GAG GGCGIC: 81% AX !Z.lImGBG II: H37RV CTG ATC CCC GAG CCC ATG GAC AAG GTG CCC AAC GAG GGT c;Ic && &xX GI(: !Z,!&GBG Ic BCG CTG ATC CCC GAG CCC ATG GAC AAG GTG GGC AAC GAG GGC GIc ATC A= GII: GAG GAG Iz
7 CTG ATC CCC GAG GCG ATG GAC GAG GTC GGC AAC GAG CCC fJ.C AIC 81l; GIG GAG&&Z III 9 CTC ATC GCC GAT TCC ATA GAG AAC GTG GGCAAG GAGGGCCJ..C BIG: ACC GICf,AfC& IC
10 CTC ATC CCC GAG TCG ATG AAG GAG GTC GGC AAG GAG CC&&ICC ATC ACC GICGA.Cf&Z IC 17 CTC ATT TCC GAA GCA CCT GAC AAG GTG GGCAAG GAGGGTm 81% &X!X.C&UXI&Z II; H37 CTT TTC TTT TTC TCT CAG CAG AAG CTC ATC ATC GTC GTC GTI: 9u; &X Au; &IC &I& I&
65 K is a refeence sequence from EMBL: MP 15989 (65 kDa antigen - Mycobacterium paratubercu/osis - nucleotid 673. The Ancest sequence cor- responds to the predicted ancestal sequence from phylogenetic analysis.
- Hybridization with internal gamma-32P labelled oli- gonucleotide probe TBlO gave a very weak signal only on the amplified DNA from the vertebra fragment (figure 2b, 80 “C for 7 h). No hybridization was observed on the amplified DNA from the rib sample. The DNA extracted, not used for the hybridization, was divided into nine aliq- uots in order to be submitted to PCR.
- Four (nos 7, 9, 10, 17) of the nine nested PCR (realized from a PCR with external primers TBlTTB2) with the prim- ers TB28/TB29 gave a positive signal (figure 2c) visualized after electrophoresis. The hybridization with the 32P
labelled TBI 0 gave no result (figure 24. Then the products of amplification were sequenced. These sequences (table I) were compared to the sequences of M. tuberculo-
sis H37Rv, M. bovk var. BCC and 65 kD gene sequences available for other mycobacteria from EMBL (table II). They were also compared to the most closely related sequences in fungi, bacteria and humans (HSP 70) and to the proteic sequence of BCG, Mttcwpa and H37RV (tables 111 and Iv). The amplified DNA of the nested PCR no. 7 was found to be more similar than those obtained from PCR nos 9, 10 and 17 to each of the comparative sequences.
Table II. Percentage of identity between the four fragments from the nested PCR nos 7, 9, 10 and 17 of the 65 kD gene target and the corresponding sequences of M. tuberculosis H37Rv, M. bovis var. BCG and the 65 kD gene sequence available for other mycobacteria.
Amplified fragment M. bovis var. BCG
from PCR no. reference strain
M. tuberculosid
H37Rv
M. gordonae M. kansaii M. xenopi M. segmatis
7 89.5 92.4 88.6 87.6 90.7 85.5 9 72.2 73.2 74.4 79.2 79.2 76.0
10 75.7 78.5 78.2 80.6 80.6 75.8 17 75.9 78.8 78.6 79.4 79.4 76.9
Table Ill. The proteic sequences.
BCC EVETKEQIAATEAISAGDQSICDIIAEAMDKVGNEGVITVEE Mttcwpa KEVETKEQIAATAAISAGDQSICDllAEAMDKVGNECVITVEE H37RV KEVETKEQIAATGAISACDQSICDllAEAMDJVGNECVITVEE Amplified AATAAISAVDQSIGDIIAEAMDEVGNEGVITVEE frag. no. 7 Amplified CGVESRREIVATASISAGDPGIGDIIADCIEKVRKAGVITVEE frag. no. 9 Amplified REVETEGEIAGTASISCADPGIDHllAESMKEVGKEGVITVEE frag. no. 10 Amplified CDVESKEQIAGTASISGGDPEIDEIISEAPDKVGKEGVITGEE frag. no. 17
948 C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321,941-951
5 4OOyear-old Mycobacterium DNA
Table IV. Percentages of identity between the different proteic sequences.
BCG Mttcwpa H37RV Amplified Amplified Amplified Amplified
frag. no. 7 frag. no. 9 frag. no. 10 frag. no. 17
BCC
Mttcwpa 97.6
H37RV 97.6 97.7
Amplified frag. no. 7 91.2 91.2 94.1
Amplified frag. no. 9 64.30 62.8 65.1 67.6
Amplified frag. no. 10 66.70 65.1 62.8 70.6 60.5
Amplified frag. no. 17 71.40 69.8 67.4 64.7 55.8 65.1
- For IS61 10 amplification using ISTB2/1STB7 primers on the DNA extracted from the rib fragment, no results were obtained (only one PCR). DNA extracted from the vertebra fragments (one PCR) gave a positive result detect- able on gel (figure 3a) and hybridizing with IS2 probe, but smaller than expected (figure 3b). One of the nine PCR performed on the vertebral fragment DNA with ISl/lS2 primers and nested PCR using J/K and ISl/lS2 primers yielded an amplified fragment of expected length (figure
3~). These, however, were unable to hybridize to the BKI probe (figure 3$) and the subsequent sequencing of these fragments failed.
3.1. Phylogenetic analysis
We only discuss (figure 5) the rooted tree with the amplified fragment no. 7 which is the tree where the pre- dicta1 ancestral sequence could be close to the real ances- tral sequence. The rooted tree with the four amplified fragments shows that the amplified fragments nos 9, 10 and 17 are not close to no. 7 or to BCC, 6.5K or to H37RV (on a cluster with H37 but on a different sub-cluster).
Several rooted trees with the minimum number of muta- tions (73) and variance (1.2) were observed. The phylo- genetic tree displaying the higher maximum likelihood is shown in figure 5 with the predicted ancestral sequence in table V. Trees with slightly larger variances or number of mutations varied by changes in the arrangements of the BCG-65K-H37RV group. The relationships between the different groups can therefore be considered to be rela- tively well determined, but not the details within the groups.
4. Discussion
The distribution and morphology of the vertebral lesions are identical to modern skeletons known to have had spinal tuberculosis (figure I).The massive destruction and collapse of the vertebral bodies, often asymmetric with preferential destruction of the anterior aspect, together with the absence of osteophytic spurring and non-involvement of the spinous processes are character- istic signs of tuberculosis [21]. The fusion of the vertebrae at their apophyses between T8 and TIO and a remodelling of the left inferior articular surface of the apophyses of T7, indicate long-term disease in the subject. The periosteal new bone formation and the lack of massive bony fusion may indicate that this was a terminal infection. The peri- osteal new bone formation on T6 and Ll and the lesions on the eighth left rib and twelfth right rib could indicate the presence of one or two paravertebral abscesses, which may have been a source of contact infection. Thus, it is likely that this individual experienced long-standing mycobacterial disease that was an infectious source and eventually led to the death of the individual.
Differences in the lengths and sequences of the ampli- fied experimental fragments by comparison with the Mycobacterium sequences are likely to be related to the significant effects of degradation in ancient DNA samples. It can be suggested that the DNA amplified in PCR no. 7 is the least degraded. It can be seen that the number of tran- sitions and transversions occurring in the experimental
Table V. Number of transitions and transversions occurring in the four amplified fragments from the nested amplification nos 7, 9, 10, 17 of the
65 kD and M. tuberculosis 65 kD antigen gene.
Amplified fragment
from the PCR no.
A <---->T G<---->C A, T-->G, C G, C-->A, T
7 1
(0.95 %j
9 1
10.75 %j
10 (0.7; %j
17 1
io.75 %)
6 3 1
(5.70 %) (2.86 %j (0.95 “/o)
14 8
(1 0.52 %)
(9.77 %)2
(6.01 %)
15 9 6
(1 1.36 “1’) (6.82 %I (4.54 %)
12 10 11
(9.02 %) (7.52 %) (8.27 %)
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998.321.941-951

i. Crubky et al.
amplification samples in comparison with the Mycobac-
terium 65 kDa antigen gene are also minimized in this amplified DNA from PCR no. 7 (table v). This sequence from PCR no. 7 appears to have been best preserved, yielding a PCR product with the closest approximation to an original Mycobacterium sequence and more than 94 % homology with the proteic sequence of H37RV. The observed modifications can be attributed to the antiquity of the Mycobacterium and/or to the effects of Taq polymerase which propagates transitions, particularly between AT and GC [3, 221. The inability to correctly hybridize to TBlO probe of the amplified fragments (figure 2) can also be explained by the same reasons. This is even clearer after more cycles of amplification with nested PCR using TB28rTB29 primers. The amount of amplified fragment, as seen on electrophoresis gel (figure 2), is higher, but no more hybridization is obtained. The high stringency conditions used in the hybridization reaction with an oligonucleotide probe such as TBlO can differentiate mycobacterial DNA molecules from closely related species after amplification with TBl/TBZ primers even though their sequences contain only a few nucle- otides differences [16]. These differences in sequence were higher here after amplification of ancient DNA and did not allow a correct hybridization. This lack of hybricl- ization of amplified fragment also eliminates the possi- bility of a laboratory contamination with contemporary DNA, which would have strongly hybridized with the probe used.
Two hypotheses can be invoked to explain the unusual diversity of this DNA sequence of Mycobacteria.
- a) The large amount of damage present in ancient DNA, and the presence of modified nucleotides at ran- dom positions increases the error rate during PCR because the natural polymerase errors are supplemented by misreadings. In order to increase the statistical likeli- hood of overcoming random error in direct sequencing, a larger number of fragments must be sequenced when employing ancient DNA. This last step was not performed in our experiment.
- b) This case is 5 400 years old. At this period Myco-
bacterial sequences are supposed to be more close to atypical mycobacteria than to present day M. tuberculosis
or M. bovis [23]. In fact, the obtained sequence from PCR no. 7 is closer to M. tuberculosis than to M. bovis. This last result is also consistent with the following.
a) The speculation that the agent of human tuberculosis arose from a very closely related cattle pathogen M. bovis
by host specialization [23] several thousand years before the predynastic case of Ada’ima 121. At this period, the evolution of the host specialization could have been closer to M. tuberculosis than to M. bovis. Even at the present time, in vivo, an important number of species between M. bovis and M. tuberculosis exist, especially in Africa [24].
b) The healed cases of bone tuberculosis (Pott’s disease) in this necropolis could prove that one part of the pop- ulation was already immunized against Mycobacterium
and that the disease already had a long evolution. Our extraction and identification of mycobacterial
DNA from the bone lesions of this 5 400-year-old Predy- nastic burial is the oldest evidence at the present time for Mycobacterium as a human disease. Previously, an acid- fast bacilii had been identified histologically from lung tis- sue of a mummified specimen dated to circa 1 000 BC to AD 400 [25] and 65 kD antigen has been amplified in the DNA extracted from another lung tissue of a mummified specimen dated to circa 1 550 to 1 080 BC [261. The tech- nological advances in recent years in molecular biology allow the further investigation of this disease from skeletal remains alone, and the more specific, genetic identifica- tion of disease agents. Our results are intriguing regarding the origins of human-hosted tuberculosis: we cannot be sure whether the lesions in the present individual were due to M. tuberculosis, M. bovis or an ancient Mycobac-
terium resembling the present two. It is possible, that if more of the original DNA were recoverable, the sequence similarities could be clarified. Our results agree with recent research in molecular biology 121 that tuberculosis is a human disease likely to be not less than 15 000 years old.
Acknowledgements: This work was supported by the lnstitut frangais d’archbologie orientale. We thank C. Chureau, B. Gicquel, M. Lampl, 8. Midant-Reynes, S. Cole, T. Janin, N. Crimal and S. Vicaire for their generous intellectual and material support of this work. We thank A. Leclerc for photography.
5. References [41 Brisson-Noel A., Aznar C., Chureau C., Nguyen 5.. Pierre C., Bartoli M., Bonete M., Pialoux G., Cicquel B., Garrigue G.. Diagnosis of tuber- culosis by DNA amplification in clinical practice. Evaluation, Lancet 338 (1991 i 364-366.
[l I Bloom B.R., Murray C.L.J., Tuberculosis: commentary on a reemer- gent killer, Science 257 (1992) 1055-1064.
[51 Mullis K.B., Faloona F., Specific synthesis of DNA in vitro via a polymerase-catalysed chain reaction, Methods in Enzymology 155
(21 Kapur V., Whittam T.S., Musser J.M., Is Mycobacterium tubercu- losisl5,OOO years, j. Infect. Dis. 170 (1994) 1348-l 349.
131 Salo W.L., Aufderheide A.C., Buikstra J.. Holcomb T.A., Identification of Mycobacterium tuberculosis DNA in a pre-Columbian Peruvian mummy, Proc. Natl. Acad. Sci. USA 91 (1994) 2091-2094.
(1987) 335-350.
[6] PZibo S.. Ancient DNA, SCI. Am. 269 15) 11993) 86-92.
[71 Spigelman M., Lemma E., The use of the polymerase chain reaction iPCRj to detect Mycobacterium tuberculosis in ancient skeletons, Int. 1. Osteoarchaeol. 3 (1994) 137-l 43.
C. R. Acad. Sci. Paris, Sciences de la vie / Life Sciences 1998. 321,941-951

5 400.year-old Mycobacterium DNA
[81 Buikstra I.E., Baker B.J., Cook D.C., What diseases plagued ancient Egyptians? A century of controversy considered, in: Biological Anthropol- ogy and the Study of Ancient Egypt, British Museum Press, London, 1993, pp. 24-30.
191 Ortner D., Putschar W., Identification of Pathological Conditions in Human Skeletal Remains, Smithsonian Institution Press, 1985.
[lo] Midant-Reynes B., Crubezy E., Janin T.. van heer W., Le site Predy- nastique d’Adaima. Rapport prGliminaire de la quatriPme campagne de fouille. Bul. Instit. Fran$. Archbo. Orient 93 119933 349-370.
[l 11 Tuross N., The biochemistry of ancient DNA in bone, Experienta 50 119941530-53s.
[121 Handt O., H&s M., Krings M.. P%bo S.. Ancient DNA: methodoiog- ical challenges, Experienta 50 (1994) 524-529.
[13] Shinnick T.M., The 65.kilodalton antigen of mycobacterium-tuber- culosis, J. Bacterial. 169 (1987) 1080-l 088.
[14] Thierry D., Cave M.D., Eisenach K.D., Crawford J.T., Bates J.H., Cic- quel B., Guesdon J.L., IS61 10 an IS like element of Mycobacterium tuber- culosis complex, Nucleic Acids Res. 18 I 1990’1 188.
[15] van Soolingen D., Hermans P.WM., De Haas P.E.W., Soll D.R., ban Embden J.D.A., Occurrence and stability of insertion sequences in myco- bacterium-tuberculosis complex strains evaluation of an inseitlon sequence-dependent DNA polymorphism as a tool in the epidemiology of tuberculosis, J. Clin. Microbial. 29 (1991) 2578-2586.
1161 Hance A.J., Grandchamp B.. Levy-Frebault V,, Lecossier D., Rauzier J., Bocart D., Gicquel B., Detection and identification of myco- bacteria by amplification of mycobacterial DNA, Mol. Microbial. 37 (1989) 843-849.
[l 71 Pierre C., Lecossier D., Boussougant Y. Bocart D., Joly V., Yeni P., Hance A.J., Use of a reamplification protocol improves sensitivity of detection of Mycobacterium tubercoiosis in clinical samples by amplifi- cation of DNA, J. Clin. Microbial. 29 (1991) 712-71 7.
1181 Eisenach K.D., Donald Cave M., Bates J.H., Crawford J.T., Polymer- ase Chain reaction amplification of a repetitive DNA sequence specific for Mycobacterium tuberculosis, J.lnfect. Diseases 161 (1990’1 977-981.
[19] Feray C., Samuel D., Thiers V., Cigou M.. Pichon F., Bismuth A., Rey- nes M., Maisonneuve P., Bismuth H., Brechot C., J. Clin. Invest. 89 (1992) 1361-1365.
1201 Cuesdon J.L., Thierry D., French Patent 9007192 and application European Patent 91401501 .l
1211 Sorrel E., Sorrel-Dejerine E., Tuberculose osseuse et osteo-articu- laire,Masson, Paris, 1932.
1221 P%ibo S., Irwin D., Wilson A.C, DNA damage promotes jumping between templates during enzymatic amplification, J. Biol. Chem. 265 11990)4718-3721.
1231 Crmek M., Les maladies B I’aube de la civilisation occidentaie, Payot, Paris, 1983.
1241 Le Miror L., Veron M.. Bact&iologie medicale, Flammarion, M@de- tine-Sciences, 1994, 986 p.
[25J Zimmerman M.R., Pulmonary and osseous tuberculosis in an Egyp- tian mummy, Bull. NY Acad. Med. 5S (19791 604-608.
[261 Nerlich A.C., Haas C.. I., Zink A., Szeimies U., Agedorn H.C., Molecular evidence for tuberculosis in an ancient Egyptian mummy, Lancet 350 (19971 1404.
C. R. Acad. Sci, Paris, Sciences de la we / Life Sciences 1998.321,941-951
951
Related Documents











