THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc Vol. 267, No. 19, Issue of July 5, pp. 13528-13534,1992 Printed in U. S. A. Identification and Localization of Multiple Forms of Serine Hydroxymethyltransferase in Pea (Pisum sativum) and Characterization of a cDNA Encoding a Mitochondrial Isoform* (Received for publication, December 21, 1991) Simon R. Turner$, Robert Ireland$, Colin Morgan, and Stephen Rawsthorne From The Cambridge Laboratorv. AFRC Institute of Plant Science Research, John Innes Centre, Colney Lane, Norwich NR4 7UJ,-United Kingdom Serine hydroxymethyltransferase (SHMT) has been purified from the mitochondria of green pea leaves. Activity can be fractionated into two distinct peaks by ion exchange chromatography. While these two forms of the enzyme are immunologically indistinguishable, immunoinhibition experiments show the presence of a distinct non-mitochondrial third form of the enzyme to also be present in green pea leaves. While this mito- chondrial form ofSHMT is abundant in leaves it is absent from roots, although the two tissues have com- parable SHMT activity. An antibody raised to purified mitochondrial SHMT was used to screen a cDNA expression library. The sequence of one of the isolated positive clones contained an open reading frame, which encoded a sequence that matched the amino acid se- quence determined from the N terminus of the mature protein. The open reading frame encodes a mature protein of 487 amino acids with a M, of 54,000, to- gether with a 27-31 amino acid serine-rich leader sequence, presumably required for mitochondrial tar- geting. The cDNA hybridizes to a small multigene fam- ily of 2-3 genes, which appear to be expressed predom- inantly in leaves. Comparison of the deduced amino acid sequence with the amino acid sequences of the rabbit mitochondrial and cytoplasmic SHMT, show that pea mitochondrial SHMT is equally similar to both of these enzymes. In addition, the rabbitsequences are more like one another than they are to the pea se- quence, suggesting an interesting evolutionary rela- tionship for these proteins. The enzyme serine hydroxymethyltransferase (SHMT)’ *This work forms part of the Agricultural and Food Research Council’s Plant Molecular Biology Program and was funded by the Cambridge Laboratory andNatural Science and EngineeringRe- search Council, Canada. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. The nucleotide sequencefs) reported in this paper has been submitted M87645. to the GenBankTM/EMBL Data Bank with accession numberfs) $ To whom correspondence should be addressed. Tel: 0603-52571; Fax: 0603-56844. 5 Permanent address: Mount Allison University, Dept. of Biology, Sackville, New Brunswick EOA 3C0, Canada. Tel.: 506-364-2504; Fax: 506-364-2505. ’ The abbreviations used are: SHMT, serine hydroxymethyltrans- ferase; THF, tetrahydrofolate; Rbu-Pn carboxylase, ribulose 1,5-bis- phosphate carboxylase/oxygenase; Hepes, 4-(2-hydroxyethyl)-l-pi- perazineethanesulfonic acid; EGTA, [ethylenebis(oxyethylene- nitri1o)ltetraacetic acid; SDS, sodium dodecyl sulfate. catalyzes the interconversion of glycine and serine, as well as a number of other reactions (for review, see Schirch (1982)). The enzyme has been purified from a number of sources including mammalian liver (Fujioka 1969; Palekar et al., 1973), Escherichia coli (Schirch et al., 1985), and plants (Rao and Rao, 1982). As well as using THF as a methyl acceptor, the enzyme from all these sources also uses the cofactor pyridoxal phosphate. The enzyme has been widely studied in mammalian liver and different forms of the enzyme from both cytosol and mitochondria have been purified and character- ized (Fujioka, 1969; Schirch and Peterson, 1980). Detailed work on the mechanism and structure of the cytosolic enzyme from rabbit liver has been carried out indicating that reaction specificity maybe determined by the conformation of the enzyme (Schirch et al., 1991). A gene for the E. coli SHMT has been cloned by its ability to complement the gly A mutant (Stauffer et al., 1981; Pla- mann et al., 1983) and site-directed mutagenesis of this se- quence has allowed the role of certain amino acids in the catalytic activity to be evaluated (Hopkins and Schirch, 1986). In addition, the amino acid sequence of both the rabbit mitochondrial and cytosolic enzymes has been determined directly (Martini et al., 1987; Martini et al., 1989), allowing the identification of several conserved regions, which may be important in the enzyme structure and function. In both E. coli and mammals, SHMT is considered to be of primary importance for the production of 1 carbon units, which are used in methionine and purine metabolism (for review, see Appling (1991)). Therefore the reaction which has been pri- marily studied is the production of glycine and 5,lO-methyl- ene-THF from serine. Indeed, Pizer (1965) has demonstrated that in E. coli, the only metabolic pathway for glycine synthe- sis is from serine via SHMT. Serine hydroxymethyltransferase has been purified from a number of different plant sources. The enzyme from soybean nodules is essential for supplying glycine and 1 carbon units, which together with ammonia from the bacteroids are required for purine biosynthesis (Mitchell et al., 1986). In green tissue, however, SHMT is a component of the mitochondrial glycine cleavage complex. Metabolism of phosphoglycollate, which is produced following the oxidative reaction of Rbu-Pz carbox- ylase, leads to the conversion of two molecules of glycine to serine, ammonia, and carbon dioxide in the mitochondria by the combined actions of the enzymes glycine decarboxylase and SHMT. Consequently, SHMT accumulates to high con- centrations in the mitochondria of green leaves (Bourguignon et al., 1988) and catalyzes the reverse reaction to that normally described for the mammalian and bacterial enzyme. Studies on a mutant deficient in mitochondrial SHMT activity (SO- merville and Ogren, 1981) suggest that this pathway is the 13528

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1992 by The American Society for Biochemistry and Molecular Biology, Inc
Vol. 267, No. 19, Issue of July 5, pp. 13528-13534,1992 Printed in U. S. A.
Identification and Localization of Multiple Forms of Serine Hydroxymethyltransferase in Pea (Pisum sativum) and Characterization of a cDNA Encoding a Mitochondrial Isoform*
(Received for publication, December 21, 1991)
Simon R. Turner$, Robert Ireland$, Colin Morgan, and Stephen Rawsthorne From The Cambridge Laboratorv. AFRC Institute of Plant Science Research, John Innes Centre, Colney Lane, Norwich NR4 7UJ,-United Kingdom
Serine hydroxymethyltransferase (SHMT) has been purified from the mitochondria of green pea leaves. Activity can be fractionated into two distinct peaks by ion exchange chromatography. While these two forms of the enzyme are immunologically indistinguishable, immunoinhibition experiments show the presence of a distinct non-mitochondrial third form of the enzyme to also be present in green pea leaves. While this mito- chondrial form of SHMT is abundant in leaves it is absent from roots, although the two tissues have com- parable SHMT activity. An antibody raised to purified mitochondrial SHMT was used to screen a cDNA expression library. The sequence of one of the isolated positive clones contained an open reading frame, which encoded a sequence that matched the amino acid se- quence determined from the N terminus of the mature protein. The open reading frame encodes a mature protein of 487 amino acids with a M, of 54,000, to- gether with a 27-31 amino acid serine-rich leader sequence, presumably required for mitochondrial tar- geting. The cDNA hybridizes to a small multigene fam- ily of 2-3 genes, which appear to be expressed predom- inantly in leaves. Comparison of the deduced amino acid sequence with the amino acid sequences of the rabbit mitochondrial and cytoplasmic SHMT, show that pea mitochondrial SHMT is equally similar to both of these enzymes. In addition, the rabbit sequences are more like one another than they are to the pea se- quence, suggesting an interesting evolutionary rela- tionship for these proteins.
The enzyme serine hydroxymethyltransferase (SHMT)’
*This work forms part of the Agricultural and Food Research Council’s Plant Molecular Biology Program and was funded by the Cambridge Laboratory and Natural Science and Engineering Re- search Council, Canada. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
The nucleotide sequencefs) reported in this paper has been submitted
M87645. to the GenBankTM/EMBL Data Bank with accession numberfs)
$ To whom correspondence should be addressed. Tel: 0603-52571; Fax: 0603-56844.
5 Permanent address: Mount Allison University, Dept. of Biology, Sackville, New Brunswick EOA 3C0, Canada. Tel.: 506-364-2504; Fax: 506-364-2505.
’ The abbreviations used are: SHMT, serine hydroxymethyltrans- ferase; THF, tetrahydrofolate; Rbu-Pn carboxylase, ribulose 1,5-bis- phosphate carboxylase/oxygenase; Hepes, 4-(2-hydroxyethyl)-l-pi- perazineethanesulfonic acid; EGTA, [ethylenebis(oxyethylene- nitri1o)ltetraacetic acid; SDS, sodium dodecyl sulfate.
catalyzes the interconversion of glycine and serine, as well as a number of other reactions (for review, see Schirch (1982)). The enzyme has been purified from a number of sources including mammalian liver (Fujioka 1969; Palekar et al., 1973), Escherichia coli (Schirch et al., 1985), and plants (Rao and Rao, 1982). As well as using THF as a methyl acceptor, the enzyme from all these sources also uses the cofactor pyridoxal phosphate. The enzyme has been widely studied in mammalian liver and different forms of the enzyme from both cytosol and mitochondria have been purified and character- ized (Fujioka, 1969; Schirch and Peterson, 1980). Detailed work on the mechanism and structure of the cytosolic enzyme from rabbit liver has been carried out indicating that reaction specificity may be determined by the conformation of the enzyme (Schirch et al., 1991).
A gene for the E . coli SHMT has been cloned by its ability to complement the gly A mutant (Stauffer et al., 1981; Pla- mann et al., 1983) and site-directed mutagenesis of this se- quence has allowed the role of certain amino acids in the catalytic activity to be evaluated (Hopkins and Schirch, 1986). In addition, the amino acid sequence of both the rabbit mitochondrial and cytosolic enzymes has been determined directly (Martini et al., 1987; Martini et al., 1989), allowing the identification of several conserved regions, which may be important in the enzyme structure and function. In both E. coli and mammals, SHMT is considered to be of primary importance for the production of 1 carbon units, which are used in methionine and purine metabolism (for review, see Appling (1991)). Therefore the reaction which has been pri- marily studied is the production of glycine and 5,lO-methyl- ene-THF from serine. Indeed, Pizer (1965) has demonstrated that in E. coli, the only metabolic pathway for glycine synthe- sis is from serine via SHMT.
Serine hydroxymethyltransferase has been purified from a number of different plant sources. The enzyme from soybean nodules is essential for supplying glycine and 1 carbon units, which together with ammonia from the bacteroids are required for purine biosynthesis (Mitchell et al., 1986). In green tissue, however, SHMT is a component of the mitochondrial glycine cleavage complex. Metabolism of phosphoglycollate, which is produced following the oxidative reaction of Rbu-Pz carbox- ylase, leads to the conversion of two molecules of glycine to serine, ammonia, and carbon dioxide in the mitochondria by the combined actions of the enzymes glycine decarboxylase and SHMT. Consequently, SHMT accumulates to high con- centrations in the mitochondria of green leaves (Bourguignon et al., 1988) and catalyzes the reverse reaction to that normally described for the mammalian and bacterial enzyme. Studies on a mutant deficient in mitochondrial SHMT activity (SO- merville and Ogren, 1981) suggest that this pathway is the
13528

SHMT f rom Pea Mitochondria 13529
sole source of photorespiratory COz release and that mito- chondrial SHMT is not essential for any other function within the plant.
Recent work on the plant glycine cleavage reaction has focused on the interactions of the different subunits (Oliver et al., 1990) and the control of expression of these light- induced polypeptides. To this end sequences of the H (Kim and Oliver, 1990; Macherel et al., 1990), P (Turner et al., 1992a), and L (Turner et al., 1992b) subunits of glycine decarboxylase from pea have recently been published. The aim of the work reported here is to examine the distribution of mitochondrial SHMT in the pea plant and to obtain a cDNA clone encoding SHMT, both to allow the comparison of a plant SHMT with that from other sources and as a prerequisite for the study of the expression and organization of the genes encoding SHMT.
EXPERIMENTAL PROCEDURES
Plant Material, Western Blotting, Amino Acid Sequencing, Isolation of Clone, and Nucleic Acid Manipulation-Growth of plant material (Pisurn satiuum, cv. Rirte). isolation of clones from Apt11 cDNA libraries, Western, Northern, and Southern hlotting and N-terminal amino acid sequencing were performed as described previously (Turner et al., 1992a).
Purification of SHMT-Mitochondrial protein extracts were pre- pared from pea leaves as described by Rourguignon et al. (1988), and loaded onto a Mono-Q HR5/5 column (Pharmacia LKR Biotechnol- ogy Inc.) equilibrated with 10 mM potassium phosphate, pH 6.8, containing 1 mM EGTA, 1 mM 2-mercaptoethanol, and 1 mM serine (huffer A). The column was washed with 5 volumes of buffer A, and the proteins eluted with a linear gradient (30 ml) of 0-400 mM potassium phosphate containing the concentrations of EGTA, serine, and 2-mercaptoethanol as ahove. The flow rate was 1 ml.min", and 1 -ml fractions were collected. Fractions containing activity (peaking at 130 mM phosphate) were pooled and diluted with 4 volumes of 20 mM Trig-HCI, pH 8.0, containing 1 mM serine, 1 r n M EGTA, and 1 mM 2-mercaptoethanol (buffer R). This pool was loaded onto a Mono- Q HR5/5 column which had been equilibrated with huffer R, and the proteins eluted using a linear gradient (A0 ml) of 0-0.5 M KC1 in huffer R. Again the flow rate was 1 ml .min", and 1-ml fractions were collected. SHMT activity eluted as two peaks, the first (designated SHMT I ) a t ahout 120 mM KCI, and the second (designated SHMT 11) a t about 200 mM KCI. Fractions containing activity from each peak were combined into two separate pools which were diluted with 5 (SHMT I) or 8 (SHMT 11) volumes of 20 mM potassium phosphate, p H 7.4, containing 1 mM serine, 1 mM EGTA, and 1 mM 2-mercap- toethanol (buffer C). These pools were (separately) loaded onto a column (8.0 X 1.0 cm) of Blue Sepharose CL-6H (Pharmacia) that had been equilibrated with huffer C. Elution was with a linear gradient (30 ml) of 0-0.5 M KC1 in buffer C, a t 1 ml.min", and 1-ml fractions were collected. SHMT activity (forms I and 11) eluted as a broad peak, centered around 300 mM KCI. Fractions containing activity were pooled (2 pools, SHMT I and SHMT II ) , dialyzed against water, and freeze-dried to concentrate the protein prior to preparative gel electrophoresis (Moos et al., 1988).
Assay of SHMT Actiuity-Activity of SHMT was determined by measuring the formation of N5,N1"-['"C]methylene-THF from [3-"C] serine using the method of Mitchell et al. (1986). Activities were assayed at, 30 "C for 15 min in 50 mM N,N,-bis(2-hydroxy- ethyl)glycine, pH 8.5, containing 10 mM 2-mercaptoethanol, 5 mM [3-"'C]serine.O.l pCi pmol", 1 mM THF, 250pM pyridoxal phosphate, and were started by the addition of enzyme extract.
Production of Antibody-Antibodies were produced in rahhits as described previously (Turner et al., 1992a). Two injections containing 75 pg each for SHMT I or 18 and 35 pg for SHMT I1 were used.
Tissue Preparation and Immunolnheling-Pea leaves (approxi- mately 80-100% fully expanded) and root segments (the apical 5 mm) were harvested from seedlings and treated for immunocytological investigations as follows. Small pieces of tissue were embedded as described by Rawsthorne et al. (1988), except that each step was performed a t 4 "C. Sections (100 nm thick) were collected on 200- mesh gold grids which had heen coated with Parlodion and carhon. The sect.ions were incuhated for 30 min in buffer containing 3% bovine serum alhumin, 10 mM Tris, pH 7.4, and 0.9% (w/v) NaCl
(buffer D). The sections were then incubated for a further 120 rnin in the same buffer containing appropriate dilutions of the SHMT I antihody. The sections were then incuhated for a futher 60 min in the buffer containing a 1:20 dilution of 1.5 nm of goat anti-rahhit gold (EM grade; Jansen Life Science Products, N'antage Oxfordshire. United Kingdom). The sections were stained for 60 min in uranyl acetate and 45 s in lead citrate. Sections were washed in deionized water hetween each stage of the procedure. The sect ions were viewed using a .Jeol EX electron microscope.
I '
. , 0 LLL-"..". -. .".
10 15 20 25 30 0.0
Fracilon number
b
1 II I II *130
*!x *39 c 2 7
4 1 7
anti-I anti-It FIG. 1. Mult iple forms of pea mitochondrial SHMT. a, elution
profile of Mono-9 ion exchange column during thr purification of SHMT. Proteins were eluted with a continuous gradient of 0-0.4 M KCI. The positions of the two penks of SHMT artivitv referred to in the text are indicated. Protein concmtrntions were monitored hy ahsorhance at 280 nm. h, N'estern hlot of pnrtiallv pure S H V T I (leff-hand side of each hlot) and SHMT 11 separated on an SDS- polyacrylamide gel probed with either anti-SHMT I (lcff-hand sidr) or anti-SHMT 11 (right-hand .si&). There were eqrrnl loadings of SHMT I and I1 (100 ng) in each track. Protein concentrations were estimated following the densitometric scanning of Coomnssie Rr i l - liant Blue R-stained gels.
S O 100 150 200 250 3b0
Anti-SHMT IgG (pg)
FIG. 2. Immunoinhibit ion of SHMT activity in mitochon- d r i a l a), leaf (O), a n d root (0) extracts . Inhihit ion was cnlct~inted by measuring thv activity in the supernatant following immunoprp- cipitation of SHMT and is expressed relative to the control experi- ment in which bovine serum albumin was atlded instend of I d ; . Incubations contained hetween 1.05 antl 1.21 nmol min of SHJIT activity with specific activities of 19.6 (lean. 22.0 (root). antl 110 (mitochondria) nmol min-l mg of protein I . J'oints rrprwent the average of two independent experiments, replicates vnrierl hy le95 t,han 5% of the mean.
13530 SHMT from Pea Mitochondria TABLE I
Srrinr hydrox~mrthyltmnsf~rase activit ies in total protein extracts of p a tissues
Pea leaves (approximately 80-100% fully expanded) and 5-mm segments from the apical or mature regions of the roots were ground in ice-cold extraction huffer (50 mM Hepes, pH 7.5, 1 mM dithiothre- itol, 1 mM serine, 1 mM EGTA) with 0.15 g of polyvinylpol.ypyrroli- done g" of fresh weight of tissue using a chilled mortar and pestle followed by an all-glass homogenizer. Extracts were spun a t 12,000 X c: for 20 min a t 4 "C. The supernatants were desalted on a PD-10 column (Pharmacia) equilihrated with 20 mM Hepes. pH 7.5, 1 mM dithiothreitol, 0.5 mM serine, and pyridoxal phosphate added to give a concentration of 50 pg ml". Values represent the mean ? S.E. of three separate extractions except for mature root segments where the values for two extracts are given.
Tissue Enzyme activity (nmol min") m f ' Jrrsh wright ma" prolrin
Leaves 600 -c 25 19.6 ? 1.0 Apical root segments 326 rf: 3 22.0 +. 1.5 Mature root segments 125, 124 20.0, 17.8
RESULTS
SHMT activity routinely eluted from the second Mono-Q column as two distinct peaks (Fig. la). Two peaks of SHMT activity also eluted a t identical positions when total protein extract from whole leaves was fractionated in the same way. Antibodies raised to both SHMT I and SHMT I1 appeared to recognize only a single protein of M , - 53,000 when used as a probe against a Western blot of pea leaf extract separated on an SDS-polyacrylamide gel (see, for example, Fig. 6). In addition, as judged by Western blot analysis, the SHMT I
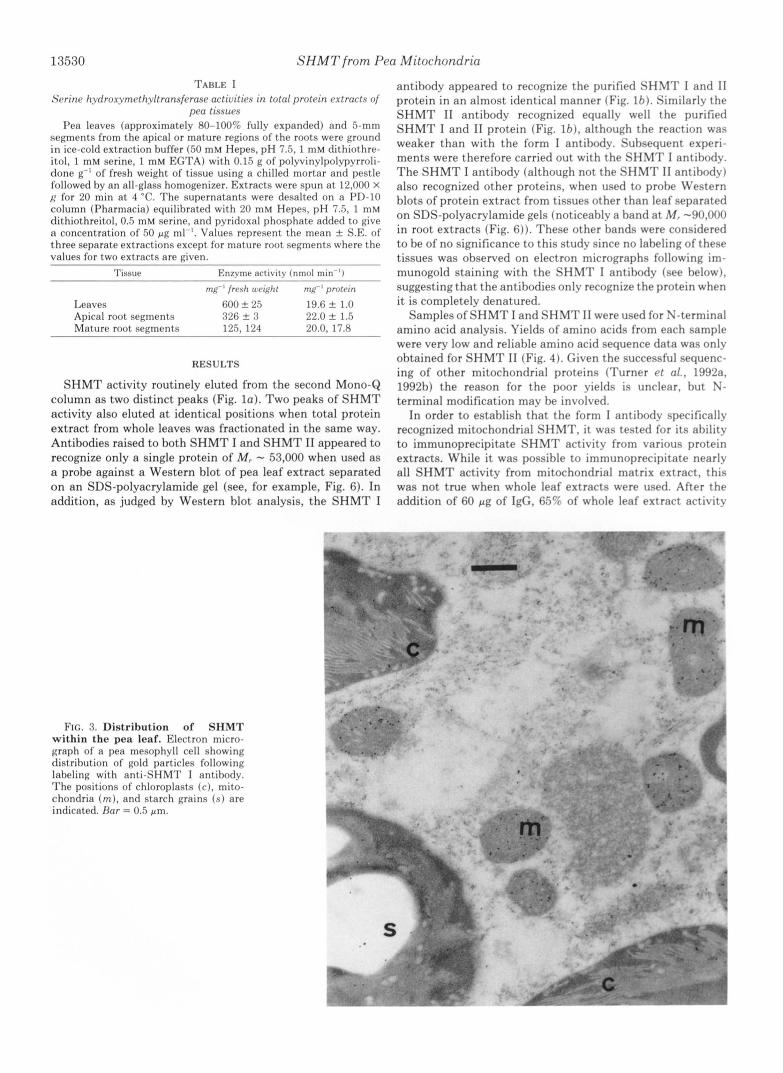
FIG. 3. Distribution of SHMT within the pea leaf. Electron micro- graph of a pea mesophyll cell showing distrihution of gold particles following laheling with anti-SHMT I antibody. The positions of chloroplasts ( c ) , mito- chondria ( m ) , and starch grains (s ) are indicated. Bar = 0.5 pm.
antibody appeared to recognize the purified SHMT I and I1 protein in an almost identical manner (Fig. I h ) . Similarly the SHMT I1 antibody recognized equally well the purified SHMT I and I1 protein (Fig. lh ) , although the reaction was weaker than with the form I antibody. Subsequent experi- ments were therefore carried out with the SHMT I antibody. The SHMT I antibody (although not the SHMT 11 antibody) also recognized other proteins, when used to probe Western blots of protein extract from tissues other than leaf separated on SDS-polyacrylamide gels (noticeably a band at M , -90,000 in root extracts (Fig. 6)). These other bands were considered to be of no significance to this study since no labeling of these tissues was observed on electron micrographs following im- munogold staining with the SHMT I antibody (see below), suggesting that the antibodies only recognize the protein when it is completely denatured.
Samples of SHMT I and SHMT I1 were used for N-terminal amino acid analysis. Yields of amino acids from each sample were very low and reliable amino acid sequence data was only obtained for SHMT I1 (Fig. 4) . Given the successful sequenc- ing of other mitochondrial proteins (Turner rt al., 199223, 1992b) the reason for the poor yields is unclear, but N- terminal modification may be involved.
In order to establish that the form I antibodv specifically recognized mitochondrial SHMT, it was tested for its ability to immunoprecipitate SHMT activity from various protein extracts. While it was possible to immunoprecipitate nearly all SHMT activity from mitochondrid matrix extract, this was not true when whole leaf extracts were used. After the addition of 60 pg of IgG, of whole leaf extract activity

1
61
121
181
241
301
361
421
481
541
601
661
721
781
841
901
961
1021
1081
1141
1201
1261
1321
1381
1441
1501
1561 1621 1681 1741 1801
SHMT from PC
ATGGACCTTTCTATCTCTAACATGGCTATGGCAATGGCGCTTCGTAAGCTTTCATCTTCC H A H A H A L R K L S S S 2 0
V N K S S R P L F S A S S L Y Y K S 8 1 40 GTCMCAMTCTTCACGTCCTCTCTTTTCTGCTTCATCTCTTTACTACAAGTCTTCTTTG
CCTGATOAAGCTGTTTACGACRRAGAGMTCCACGTGTTACGTGGC~GCAATTGAAT P D E A V Y D X E N P R V T Y P K Q L N 60
S P L E V I D P E I A D I I E L E K A P Rn
A V G S V H T N R Y S E G Y P G A R Y Y GCTGTTGGCTCTGTCATGACAI\ACAI\ATACAGTGAAGGTTATCC~CAAGATATTAT G G N E Y I D H A E T L C Q K R A L E A GGAGGAI\ATGAGTATATTACATGGCAGAI\ACACTATGCCAGAAGCGTGCCTTGGAAGCA F R L D P A K W G V N V Q P L S G S P S
N F Q V Y T A L L X P N D R I H A L D L TTTCGGTTGGATCCTOCTTGGGGAGTOMTGTGCAGCCTCTGTCTGGTTCGCCTTCA
AACTTTCMGTTTACACTGCATTGTTGAIUCCTCATGATAGAATCATGGCACTTGATCTC P H G G H L S H G Y Q T D T K K I S A V C C T C A T G G A G G G C A T C T T T C T C A T G G A T A C C A G A C C G A C T C S I F F E T H P Y R L D E S T G Y I D Y
D Q L E K S A T L F R P K L I V A G A S TCTATATTTTTTGAGACMTGCCATATAGGTTGGATGRRAGCACAGG~TACATTGACTAT
GACCAGCTAGAGAAATCGGCTACACTCTTCAGGCCRRAGTTAATAGTTGCCGGTGCTAGT A Y A R L Y D Y A R I R K V C D K Q K A G C C T A T G C A A G G C T G T A T G A T T A T G C A T A C O C A A G G C T V L L A D H A H I S Q L V A A G V I P S GTGTTGTTAGCAGATATGGCACATATCMTOOATTGGATTGGTTGCAGCTGGTGTTATCCCTTCA P F D Y A D V V T T T T H X S L R G P R CCTTTTGATTATGCAGATGTAGTGA~CCACAACTCACAAGTCACTTCGCGGCCCACGT G A H I F F R X G L K E V N K Q Q K E V
F Y D Y E D K I N Q A V F P G L Q G G P GGAGCTATGATATTTTTCAGGAA~ATTAGAAGTCAACRRACAAGG~GAGGTG
TTCTATGACTATGAGGACATCAAT~GCTGTCTTCCC~GACTGCAAGGTGGTCCT N N H T I T G L A V A L K Q A T T P E Y CACAACCATACCATTACAGGTTTAGCTGTTGCATTGAAGCAGGCTA~CTCCAGAGTAT
EAGTGGAAGGGGCTGGMCTGATACCATCAGAGAATTTTACTTCTTTATCTGTTATG~A ~ ~ ..
R A Y Q E Q V L S N S S K F A X A L S E AGAGCATATCAAGAGCAGGTTCTCAGCAATAGCTCRRAGTTTGCAAAGGCTCTGAGTGAG X Q Y D L V S G G T E N N L V L V N L K RRAGGCTATGATCTTGTTTCCGGTGGGACTGAGAATCATTTAGTTTTGGTGAATCTGAAG N K G I D G S R V E X V L E L V H I A A AACAAGGGTATTGATGGGTCCAGAGTTGAGAAAGTGTTGGAATTAGTACATATTGCTGCT N K N T V P G D V S A H V P Q G I R H G AATARAAACACAGTTCCAGGAGATGTGTCTGCTATGGTTCCTGGTGGCATCAGGATGGGA T P A L T S R G F V E E D F V K V A E Y
120
140
160
1 80
200
220
240
260
280
300
320
340
360
380
400
420
440
460
F D A A V S L A L K V K A E S K G ~ T ~ L ~ 480 TTTGATGCAGCTGTGWTTTGGCCTTGAAGGTTAAGGCAGRRAGC~GGAAC~TTA K D F V E A L Q T S S Y V Q S E I S X L 500 AAGGACTTCGTGGAAGCATTGCAGACATCCTCTTACGTGCAATCAGAGATTTCCAAGCTT K H D V E E F A K Q F P T I G F E K A T 520
H K Y N X * AAGCATGRTGTTGAGGAGTTTG~CAATTTCCAACCATTGGTTTTGARAAAGCAACC
FIG. 4. Sequence of the cDNA clone pSH1, together with the translation of the open reading frame. Amino acid sequence underlined and in bold represents the sequence matching that deter- mined directly from the N terminus of the mature protein.
could be precipitated (Fig. 2). However, very little further activity could be precipited despite a 4-fold increase in the amount of IgG added. In addition, the antibody had little or no effect on the SHMT activity of root extracts (Fig. 2), even though SHMT specific activities in roots and leaves were similar (Table I).
When the antibody was used as a probe in immunogold labeling experiments, the only compartment within the me- sophyll cells of a pea leaf which showed any significant labeling was the mitochondrial (Fig. 3). The antibody did not label any part of the cells from either dividing or mature regions of the pea root to any greater extent than preimmune IgG.'
The SHMT I antibody was used to screen a cDNA expres- sion library constructed from poly(A+) pea leaf mRNA in Xgtll. Several positive clones were isolated and five of these were selected for further analysis. Sequence from the ends of these clones revealed the presence of three different polyad- enylation sites.' The clone most extended in the 5' direction (pSH1) was sequenced completely on both strands (Fig. 4). Translation of a large open reading frame revealed a sequence which matched the N-terminal amino acid sequence deter- mined directly from the mature protein. Upstream (5') of this is the sequence MAMAMA and it seems likely that one of these methionine residues is the translation start site. This would give a 27-31 amino acid serine-rich (-30%) leader sequence presumably required for mitochondrial targeting, and a mature protein of 487 amino acids of M, 54,000, which
S. R. Turner, R. Ireland, C . Morgan, and S. Rawsthorne, unpub- lished experiment.
la Mitochondria 13531
agrees well with previous estimates from SDS denaturing gels of M, 53,000 (Bourguignon et al., 1988; Henricson and Ericson, 1988).
Alignment of the deduced amino acid sequence with previ- ously published bacterial and mammalian SHMT sequences (Fig. 5), shows considerable conservation along most of the length of the clone. The pea sequence is more similar to the rabbit cytosolic and mitochondrial sequences than it is to any bacterial sequence. Interestingly, however, the pea sequence is about equally similar to both the rabbit mitochondrial (74% similarity, 60% identity) and cytosolic sequences (72% simi- larity, 60% identity). Similarity at the N terminus of these sequences is comparatively poor, suggesting that although the N-terminal amino acid sequence that was obtained directly from the pea mitochondrial SHMT (Fig. 4) is relatively short (7 amino acids), it is likely to be unique to mitochondrial SHMT and not conserved in any other forms of SHMT which may be present within other cellular compartments of pea.
When pSHl was used to probe a Northern blot of RNA from various pea tissues, a strong signal (at -1,800 nucleo- tides) was obtained with RNA from leaf, but not root or embryo (Fig. 6). The level of SHMT mRNA corresponding to pSHl in roots or embryos is less than 1% that in green leaves. This closely matches the pattern of the SHMT polypeptide of M, -53,000 detected on Western blots using the anti- SHMT I antibody (Fig. 6), indicating that the expression of SHMT is controlled primarily by the level of the correspond- ing mRNA. The SHMT clone pSHl was also used as a probe to measure the number of SHMT-encoding genes. Compari- son of the signal obtained with pea DNA digested with various restriction enzymes with that of various concentrations of standard plasmid DNA suggests that there are 2-3 copies of a gene corresponding to pSHl (Fig. 7). If the stringency of the washing step is lowered to 2 X SSC at 65 "C, several other bands are also seen on the autoradiograph.'
DISCUSSION
We have purified SHMT from the mitochondria of green pea leaves and have resolved the activity into two forms on a Mono-Q ion exchange column. Mitochondrial SHMT has previously been purified from both pea (Bourguignon et al., 1988) and spinach leaf (Henricson and Ericson, 1988)) but in neither case has the presence of more than a single form been reported. Two peaks of activity are not only seen with partially pure SHMT (Fig. l ) , both of these peaks of SHMT activity have also been observed when crude leaf extracts have been separated by ion exchange chromatography.' This suggests that the presence of the two forms is not an artifact produced during the purification of SHMT. It seems likely that the presence of two forms has not been apparent in previous purifications, but has been observed here, because of the superior resolution of the Mono-Q column eluted with a KC1 gradient in Tris buffer. The significance of the two forms of SHMT isolated from peas and the relationship between the two forms remains unclear. No reliable N-terminal amino acid sequence information was obtained from SHMT I, and therefore no sequence comparison can be made between the two forms. However, as judged from the cross-reaction of the antibodies against Western blots of the two purified forms, there is no immunological difference between them (Fig. lb) , suggesting that the structure and primary sequence is identi- cal or very similar. Multiple forms of rabbit liver cytosolic SHMT have been resolved on ion exchange columns (Schirch et al., 1984; Artigues et al., 1990). This observation may be explained, in part at least, by structural changes in an aspar- aginyl residue at position 5 (Artigues et al., 1990). While there

13532
FIG. 5. Alignment of different SHMT amino acid sequences. Iden- tity with the deduced pea amino acid sequence is indicated by dots. Rahhit mi- tochondrial (Martini et al., 1989) and rabbit cytoplasmic (Martini et a!., 1987) sequences are directly determined amino acid sequences. The E. coli amino acid sequence was deduced from the sequence of the gly A gene (Plamann et al., 1983).
SHMT from Pea Mitochondria
Rabbitmito. ~QTQTGEASRGWTGQE-----S.SDT HWELLQR..D..CR.....A....C.RAALE Pea mito. SLPDEAVYDKENPRVTWPKQLNSPLEVIDPEIADIIELE~Q~GLELIPSENFTSLS~Q
E . c o l i H.KR.M-----------------NIADY.A.LWQAn.Q..V..EEHI...A...YT.PRV .. Pea mito. AVGSVMTNKYSEGYPGARYYGGNEYIDMAETLCQKRALEAFRLDPAKWGVNVQPLSGSPSNF Rabbitmito. .L..CLN.........K.....A.W.EI.L..QR......D....Q.......Y....A.L Rabbitcyt . .L..CLN.........Q.....T.H..EL....QK...Q.YG...QC.......Y....A.. E . c o l i .Q..QL....A.....K.....C..V.IV.Q.AID..K.L.GA.YA----....H...QA..
Pea mito. Q V Y T A L L K P H D R I ) . I A L D L P H G G H L S H G Y Q T D T K K I S A V D E S T G Y I D Y D Q L E Rabbitmito. AA..... Q......G....D....T...HS.V.RV..T.....S...K.NPQ..L...E..A Rabbit c y t . A.....VE..G...G....D....T..FH..K..I..T.....S.A.KVNPD.......R.. E . col i A.... .. E.G.TVLGHN.AH....T..SP-----VNFSGKLYNIV..GIDATGHI-..RD.. Pea mito. KSATLFRPKLIVAGASAYARLYDYARIRKVCDKQKAVLLADHAHISGLVAAGVIPSPFDYAD Rabbitmito. LT.R....R..I..T......I....H.E...EV..H... ........... K......KH.. Rabbitcyt . EN.R..H....I..T.C.S.N...G,L..IA.ENG.Y.H..........V...V....EHCH E . c o l i KQ.KEHK..H.IG.F ... SGW.W.KH.EIA.SIG.Y.FV....VA ....... Y.N.VPH.H
Rabbitmito. ........ T...A.SGL..Y...VRT.DP.T.Q.IP.TF.DR..€ .... S ..... H.. A.AA Pea mito. WTTTTHKSLRGPRGAnIFFRKGLKEVN-KQGKEVFYDYEDKINQAVFPGLQCGPHNHTITG
Rabbitcyt . ........ T . A . C . A G . . . Y . R . V R S . D P . T . .. 1L.NL.SL .. S.............A. A. E . c o l i ........ T...P.GGL.L--------- AKGGS.EL.K---KL.S.....G....LH.V.A.
Pea mito. LAVALKQATTPEYRAYQEQVLSNSSKF~SEKGYDLVSGGTENHLVLVNLKNKGIDGSRV
Rabbitcyt . V.......H...€KE..R..VA.CRALSA..V.L..KI.T..SD...I..D.RS..T..G.A E . c o l i K.....E.ME..FKT..Q..AK.AKAMVEVFL.R..KV.....D...€..D.VD.NLT.KEA
Pea mito. EKVLELVHIAANKNTVPGD-VSAHVPGGIRnGTPALTSRGWEEDFVKVAEYFDAAVSLALK Rabbitmito. .R.....S.T.....C...-R..IT...L.L.A......QFR.D..RR.MFI.EG.NIGLE
E . c o l i DAA.GRAN.TV...SV.N.PK.PF.TS...V.T..I.R...K.REAKEL.CWHCDVLD---S
Pea mito. VKAESKG-TKLKDFVEALQTSSYVQSEISKLKHDVEEFAKQFPTIGFEKATHKYNK Rabbit mito. ..RKT---A..Q..KSF.LKDPETSQRLAD.RRR.QQ..RA..MP.FP------EH Rabbit c y t . IQDDTGPRAT..E.K.K.AGDEKHQRAVRA.RQE..S..AL..LP.LP------GF E . c o l i IND.A--------------------- V.ERI.CK.LDICARY.VYA-----""-
... Rabbit c y t . ATAVNGAPRDAALWSSHEQH.AQ..KDS.A.VYD.IKK.SN..RV ..... A....A.RA.LE
Rabbitmito. V.......C..HF.E.SL...K.ARAH.D..L.R..S ...... D ...... D.RP..L.. A.A
Rabbit c y t . ..... ACS..C .... C...-K..LR.S.L.L... ...... LL.K..Q ... HFIHRGIE-TVQ
"130 "75
Protein '50 "39
RNA
FIG. 6. Tissue specific expression of SHMT. Western blot ( top) of total protein extracts from various pea tissues separated on an SDS-polyacrylamide gel, prohed with anti-SHMT I antibody. Figures on the right-hand side represent sizes of standard molecular weight markers (M, X lo-"). The corresponding Northern blot of a gel containing 1 pg of poly(A+) RNA from the same tissues, prohed with pSH1, is shown underneath.
is no asparagine residue a t position 5 in the SHMT sequence from pea reported here, it is possible that other asparagine residues, near the N terminus or elsewhere in the sequence, may behave in a similar way to that in the rabbit liver enzyme.
Incubation of the SHMT I antibody with mitochondrial matrix extract from green leaves leads to immunoprecipita- tion of nearly all the SHMT activity (Fig. 2). When the same experiment is performed on extracts from root tips, however, little or no SHMT activity is precipitated. This result is consistent with the work of Walton and Woolhouse (1986) who found that the majority of root SHMT activity was associated with plastids. This plastid form of SHMT is pre- sumably sufficiently different from the mitochondrial form of the enzyme, that it is not recognized by the SHMT I antibody. Most interestingly, however, the antibody is capable of im- munoprecipitating only 65% of SHMT activity from the whole leaf extracts, despite a 4-fold increase in the amount of antibody added (Fig. 2). This result indicates that there may
7"
5"
3" -
1"
FIG. 7. Copy number determination for SHMT genes. A Southern blot containing 10 pg of pea 1)NA rligcstcrl with various restriction enzymes. The appropriate conrentrat ion o f pSH 1 digested with KpnI was also loaded to give a signal corresponding to 0 . 5 4 copies per haploid genome. Following hyhridization with pSHI. hlots were washed with 0.2 X SSC at 65 ' C . Fimren on the I~ / t -hand PidP represent size of standard molecular weiKht markers ( h a w pairs X
10"').
be a non-mitochondrial form of SHMT in green leaves. Pre- vious reports (Henricson and Ericson, 1988), based on differ- ential centrifugation techniques, have suggested that R H green leaf SHMT activity is localized in the mitochondria. However, given the incomplete recovery of marker enzymes and the complications of interpretation of the data i f an enzyme is present in more than one compartment, it is possible that R
minor form of SHMT present in another compartment WRS

SHMT from Pea Mitochondria 13533
overlooked. Since both mitochondrial SHMT and the corre- sponding mRNA are expressed predominantly only in leaves (Fig. 6), it seems likely that the sole function of leaf mito- chondrial SHMT is for glycine decarboxylation during the photorespiratory cycle. It appears, however, that at least one other form of SHMT is found in both roots and leaves, presumably required for other “housekeeping” 1-carbon me- tabolism within the cell. Consistent with this idea, Somerville and Ogren (1981) have shown that a mutant lacking any mitochondrial SHMT activity still has SHMT activity in whole leaf extracts and that these mutants are able to grow quite normally under conditions which suppress photorespir- ation. In plants at least one form of non-mitochondrial SHMT is likely to be located in the plastid (Kisaki et al., 1971; Gardestrom et al., 1985; Shingles et al., 1984; Walton and Woolhouse, 1986).
Several reasons demonstrate that the clone pSHl encodes mitochondrial SHMT and not a form of SHMT from another cellular compartment. When used in immunogold labeling experiments with pea leaf sections the antibody used to isolate the clone will specifically label only mitochondria (Fig. 3). The amino acid sequence deduced from the open reading frame of the cDNA clone contains a sequence which exactly matches a sequence determined directly from the N terminus of the mature SHMT which we purified from isolated mito- chondria. Alignment of the mature N terminus with the open reading frame suggests the presence of a serine-rich leader sequence likely to be required for mitochondrial targeting. Two additional observations confirm that the clone is nuclear encoded. First, the cDNA clone contains a poly(A) tail, absent from mitochondrially encoded genes. Second, we have located the corresponding genes to a position on the pea chromosomal genetic map.’ There has been a report that pea leaf SHMT is mitochondrially encoded (Combette et at., 1991) and it is possible that a form of SHMT encoded in the mitochondria may also exist.
The open reading frame starts with the unusual sequence MAMAMA. This is interesting since the sequence of another polypeptide from the glycine cleavage reaction, that of the L subunit of glycine decarboxylase, starts with the sequence MAMA (Turner et al., 1992a). The significance of this, how- ever, is unclear. Although the second and third methionine residues match most closely to the plant initiation consensus of AACAAUGGC (Lutcke et al., 1987), the first methionine contains an A at -3 identified as being important in both plant and animal consensus sequences.
There is no obvious sequence near the 3’ end of the clone which corresponds to the AAATAA consensus sequence for a polyadenylation signal. Such an observation has also been made for the H, P, and L subunits of glycine decarboxylase (Macherel et al., 1990; Turner et al., 1992a, 1992b) and may explain why all these mRNAs apparently have multiple pol- yadenylation sites.
There appears to be a small multigene family of 2-3 genes corresponding to the cDNA pSHl (Fig. 7). The high strin- gency used to wash the blots indicates that these genes are closely related. It is therefore possible that the two forms of SHMT resolved by ion exchange chromatography may be encoded by different members of this gene family. Whether the genes which appear to hybridize a t lower strigency’ encode other non-mitochondrial forms of SHMT remains to be elu- cidated.
AS deduced from sequence homology the pyridoxal phos- phate cofactor attaches to Lys294 in the pea sequence, and around this residue are a number of amino acids conserved in both procaryotic and eucaryotic SHMT (Fig. 5). In addition
to the run of hydrophobic amino acids on the carboxy side of Lys294 previously identified as a hydrophobic pocket involved in the substrate-binding site in a number of pyridoxal phos- phate-containing amino acid-metabolizing enzymes (Morris and Boeker, 1983), is a run of 4 consecutive threonine residues followed by 2 valine residues on the amino side of LysZg4. Since this sequence is found only in SHMT (Vaaler and Snell, 1989) and not in the P subunit of glycine decarboxylase (Turner et at., 1992a), also a glycine metabolizing enzyme, it is tempting to speculate that this VVTTTT sequence may be involved in THF binding.
Alignment of known SHMT amino acid sequences (Fig. 5) clearly shows that the eucaryotic sequences are more like one another than they are like any of the procaryotic sequences. Purely on the basis of sequence identity (or similarity) the pea mitochondrial SHMT sequence is slightly more like the rabbit mitochondrial sequence than it is like the rabbit cyto- solic sequence. Other features, however, such as a 5-amino acid insertion near the N terminus common to both the pea mitochondrial and rabbit cytosolic sequences suggest the op- posite relationship. In addition, the two rabbit sequences are much more closely related to one another than they are to the pea sequence. The sequences of various enzymes, such as malate dehydrogenase, where both the cytosolic and mito- chondrial forms of the enzyme have been studied, provide good evidence for the symbiotic origin of mitochondria (McAlister-Henn, 1988). The relationship between the pea and rabbit sequences described above, however, is more dif- ficult to explain. Assuming a common origin for all mitochon- dria it would be expected that pea sequence would be consid- erably more like that of the rabbit mitochondrial enzyme than that of the rabbit cytosolic form. In addition, Cybulski and Fisher (1976) have suggested that in mammalian livers, mi- tochondrial SHMT acts as part of a shuttle to export l-carbon units to the cytoplasm and works in the direction of serine production from glycine, as for the pea mitochondrial enzyme. Thus, on a functional basis it would also be expected that the two mitochondrial enzymes would be more similar to one another than to a cytosolic form. Since this is clearly not the case, two possible explanations can be envisaged. First, it may be that the cytosolic form of the rabbit SHMT arose following a gene duplication event which occurred after the divergence of the lines from which animals and plants arose. To support this idea there seems to be no evidence for a cytosolic form of SHMT in plants. Alternatively, Gray et al. (1989) have sug- gested that rRNA genes of plant mitochondria are from a different origin to the rRNA genes of other organisms sup- porting a polyphyletic origin of mitochondria. If this is the case then the relationship between pea and rabbit SHMT genes may be affected by the differing origins of plant and animal mitochondrial DNA.
Acknowledgments-We thank Dr. Pat Barker (Institute of Animal Production and Grassland Research, Babraham, Cambridge) for per- forming the N-terminal amino acid sequencing, and Drs. A. Smith and D. Murphy for comments on the manuscript.
REFERENCES Appling, D. R. (1991) FASEB J. 5, 2645-2651 Artigues, A., Birkett, A,, and Schirch, V. (1990) J . Biol Chem. 265,4853-4858 Bourgulgnon, J., Neuburger, M., and Douce, R. (1988) Biochem. J . 255, 169-
178 Combette, B., Bonnard, G., Neuburger, M., Douce, R., and Grienenberger, J.
M. (1991) Abstract from the Third Plant Respiration Meeting, July 14-19, 1991 Utrecht, Holland
Fujioka, M. (1969) Biochem. Biophys. Acta 185,338-349 Cybulski, R. R., and Fisher, R. R. (1976) Biochemistry 15,3183-3187
Gardestrom, P., Edwards, G. E., Henricson, P., and Ericson, 1. (1985) Physiol.
Gray, M. W., Cedergren, R., Abel, Y., and Sankoff, D. (1989) Proc. Natl. Acad.
Henricson, D., and Ericson, I. (1988) Physiol. Plant ‘74,602-606
Plant 64,29-33
Sci. U. S. A. 8 6 , 2267-2271

13534 SHMT from Pea Mitochondria Hopkins, S., and Schirch, V. (1986) J. Biol. Chem. 261,3363-3369 Kim, Y., and Oliver, D. J. (1990) J. Biol. Chem. 265,848-853. Kisaki, T., Imai, A., and Tolbert, N.E. (1971) Plant Cell Physzol. 12,267-273 Lutcke, H. A,, Chow, K. C., Mickel, F. S., Moss, K. A,, Kern, H. F., and Scheele,
Macherel, D., Lebrun, M., Gagnon, J., Neuhurger, M., and Douce, R. (1990)
Martini, F., Angelaccio, S., Pascarella, S., Barra, D., Bossa, F., and Schirch, V.
Martinl, F., Maras, B., Tanci, P., Angelaccio, S., Pascarella, S., Barra, D.,
McAlister-Henn, L. (1988) Trends Biochem. Sci. 1 3 , 178-181 Mitchell, M. K., Reynolds, P. H. S., and Blevin, D. G. (1986) Plant Physiol.
Morris, D. R., and Boeker, E. A. (1983) Methods Enzymol. 9 4 , 125-134 Moos, M., Nguyen, N.Y., and Liu, T-Y. (1988) J. Biol. Chem. 263,6005-6008 Oliver, D. J., Neuburger, M., Bourguignon, J., and Douce, R. (1990) Plant
Palekar, A. G., Tate, S. S., and Meister, A. (1973) J. Biol. Chem. 2 4 8 , 1158-
G. A. (1987) EMBO J. 6,43-48
Biochem. J. 2 6 8 , 783-789
(1987) J. Biol. Chem. 262,5499-5509
Bossa, F., and Schirch, V. (1989) J. Biol. Chem. 264,8509-8519
81,553-557
Physiol. 94,833-839
1167 Plamann, M., Stauffer, L., Urbanowski, M., and Stauffer, G. (1983) Nucleic
Rao, D. N., and Rao, N. A. (1982) Plant Physiol. 6 9 , l l - 1 8 Acids Res. 11,2065-2075
Pizer, L. I. (1965) J. Bacteriol. 89, 1145-1150 Rawsthorne, S., Hylton, C. M., Smith, A. M., and Woolhouse, H. W. (1988)
Schirch, L. (1982) Adu. Enzymol. 53,83-112 Schlrch, L. and Peterson, D. (1980) J. Biol. Chem. 255 , 7801-7806 Schirch, V., Gavilanes, F., Peterson, D., Bullis, B., Barra, D., and Bossa, F.
(1984) Chemical and Biological Aspects of Vitamin Be Catalysls: Part A, pp.
Schirch, V., Hopkins, S., Villar, E., and Angelaccio, S. (1985) J. Bacteriol. 163, 301-388, Alan R. Liss Inc., New York
1-7 Schirch, V., Shostak, K., Zamora, M., and Gautam-Basak, M. (1991) J. Biol.
Chem. 266 , 759-764 Shingles, R., Woodrow, L., and Grodzinski, B. (1984) Plant Physiol. 7 4 , 705-
71 n
Planta (Heidelb.) 175,452-459
Somerville, C. R., and Ogren, W. L. (1981) Plant Physiol. 67,666-671 Stauffer, G. V., Plamann, M. D., and Stauffer, L.T. (1981) Gene (Amst.) 14 , 62-79
Turner, S. R., Ireland, R., and Rawsthorne, S. (1992a) J. Biol. Chem. 2 6 7 ,
Turner, S. R., Ireland, R., and Rawsthorne, S. (199213) J. Biol. Chem. 2 6 7 , 5355-5360
7 7 A L 7 7 K I l . ." , ,"" Vaaler, G. L., and Snell, E. E. (1989) Biochemistry 28, 7306-7313 Walton, N., and Woolhouse, H. W. (1986) Planta (Heidelb.) 167,119-128
Related Documents











