Vol. 46, No. 4, November 1998 BIOCHEMISTRY and MOLECULARBIOLOGYINTERNATIONAL Pages 775-785 IDENTIFICATION AND CHARACTERIZATION OF THE SMT3 cDNA AND GENE ENCODING UBiQUITIN-LIKE PROTEIN FROM DROSOPHILA MELANOGASTER 1 , , and Hung-Wen Huang , Stephen C.-M. Tsoi 2 Y. Henry Sun 3 1,2,4 Steven S.-L. Li 1 Department of Biological Sciences, National Sun Yat-Sen 2 , University, Kaohsiung, Taiwan 80424, ROC, Dlvislon of Intramural Research, National Institute of Environmental Health Sciences, National Institutes of Health, Research Triangle Park, North Carolina 27709, USA, and 3Institute of Molecular Biology, Academic Sinica, Taipei, Taiwan 11529, ROC. Received June 18, 1998 ReceivedaRerrevision, August3,1998 A SMT3 cDNA encoding ubiquitin-like protein from Drosophila melanogaster was isolated and sequenced. Drosophila SMT3 genomic DNA was amplified by polymerase chain reaction, and its nucleotide sequence was found to be identical to that of the cDNA, indicating the absence of intron in its protein coding region. The sequence of 90 amino acids of Drosophila SMT3 exhibited 55%, 73%, 70% and 52% identity to yeast SMT3, human SMT3A, SMT3B and SMT3C protein The nucleotide sequence has been deposited in the GenBank database under accession no. AF053083. 4To whom all correspondence should be addressed at NIEHS, NIH, RTP, NC27709, USA. Tel. 1-919-541-4253; FAX 1-919-541-7593; e-mail: LiSNIEHS.NIH.GOV 1039-9712/98/160775-11505.00/0 Copyright 1998 by Academic Press Australia. 775 All rights of reproduction in any farm reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Vol. 46, No. 4, November 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL Pages 775-785
IDENTIFICATION AND CHARACTERIZATION OF THE SMT3 cDNA AND GENE ENCODING UBiQUITIN-LIKE PROTEIN FROM DROSOPHILA MELANOGASTER
1 , , and Hung-Wen Huang , Stephen C.-M. Tsoi 2 Y. Henry Sun 3
1,2,4 Steven S.-L. Li
1 Department of Biological Sciences, National Sun Yat-Sen
2 ,
University, Kaohsiung, Taiwan 80424, ROC, Dlvislon of
Intramural Research, National Institute of Environmental
Health Sciences, National Institutes of Health, Research
Triangle Park, North Carolina 27709, USA, and 3Institute of
Molecular Biology, Academic Sinica, Taipei, Taiwan 11529,
ROC.
Received June 18, 1998 ReceivedaRerrevision, August3,1998
A SMT3 cDNA encoding ubiquitin-like protein from
Drosophila melanogaster was isolated and sequenced.
Drosophila SMT3 genomic DNA was amplified by polymerase chain
reaction, and its nucleotide sequence was found to be
identical to that of the cDNA, indicating the absence of
intron in its protein coding region. The sequence of 90 amino
acids of Drosophila SMT3 exhibited 55%, 73%, 70% and 52%
identity to yeast SMT3, human SMT3A, SMT3B and SMT3C protein
The nucleotide sequence has been deposited in the GenBank
database under accession no. AF053083.
4To whom all correspondence should be addressed at NIEHS, NIH, RTP, NC27709, USA. Tel. 1-919-541-4253; FAX 1-919-541-7593; e-mail: LiSNIEHS.NIH.GOV
1039-9712/98/160775-11505.00/0 Copyright �9 1998 by Academic Press Australia.
775 All rights of reproduction in any farm reserved.

Vol. 46, No. 4, 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL
sequences, respectively. Two Drosophila SMT3 transcripts of
2.5 kb and 1.5 kb were shown to be abundantly expressed in
embryo, larvae and adult tissues. The structural and
evolutionary relationships among ii SMT3 proteins from human,
mouse, Xenopus, Drosophila, nematode, Arabidopsis, rice,
Cicer, and yeast were also analyzed.
A family of ubiquitin-like proteins, including yeast
SMT3, human SMT3A, SHT3B and SMT3C, was recently discovered.
The yeast SMT3 gene was originally identified as a suppressor
of mutations in MIF2 gene, which encodes an essential protein
binding to the A+T-rich CDEII region of centromere DNA (i) .
Studies using temperature-sensitive mutants showed that the
loss of yeast Mif2p function results in chromosomes
missegregation, mitotic delay, and aberrant microtubule
morphologies (2). The Yeast Mif2 protein shares at least two
regions of similarity with mammalian centromere protein CENP-
C, an integral component of active kinetochores (3,4).
Human SMT3A gene was identified from genome project of
chromosome 21 (5). We have cloned human SMT3B (formerly
designated as HSMT3) CDNA (6). Human SMT3C protein was
independently isolated by several groups and denoted as
SU-MO-I (7), GMPI (8), PICI (9), UBLI (i0), sentrin (ii).
SUMO-I/GMPI was found to be covalently linked to the Ran
GTPase-activating protein RanGAPI, and attachment of SUMO-I
targets the otherwise cytosolic RanGAPI to the nuclear pore
complex. The modified form of RanGAPI also appeared to
associate with the mitotic spindle apparatus during mitosis
(7,8). PICI was shown to interact with the PML component of
nuclear multiprotein complex that is disrupted in acute
776

Vol. 46, No. 4, 1998 BIOCHEMISTRYond MOLECULAR BIOLOGY INTERNATIONAL
promyelocytic leukemia (9). UBLI was found to associate with
human RAD51/RAD52 proteins involved in DNA recombination and
DNA double-strand break repair (i0). Sentrin was shown to
interact with Fas/APO-I or TNF receptor i death domains, and
the overexpression of sentrin provided protection against
both anti-Fas/AP0-1 and TNF-induced cell death (ii) o
Here we report the cloning of a SMT3 cDNA, mRNA
expression, and characterization of genomic DNA from
Drosophila melanogater. We have also analyzed the structural
and evolutionary relationships of ii SMT3 protein sequences
from human, mouse, Xenopus, Drosophila, nematode,
Arabidopsis, rice, Cicer, and yeast.
MATERIALS AND METHODS
Isolation and characterization of SMT3 cDNA and gene.
Human SMT3C protein sequence was used as query to blast the
Drosophila EST database, and an EST (GenBank accession no.
AA264131) was found to contain homologous sequence. The two
polymerase-chain-reaction (PCR) primers based on the EST
sequence were: forward 5'-CATGTCTGACGAAAAGAAGGGAGG-3' and
reverse 5'-GTGGCGCTCATAAGATTACTTAF-3'. The mixed cDNAs
prepared from Drosophila adult cDNA library (Stratagene, La
Jolla, CA) were used as templates for PCR with the Expand
High Fidelity (Boehringer Mannheim) containing a mixture of
thermostable Taq and Pwo DNA polymerase. PCR conditions were:
1 cycle of 2 min. denaturation, 5 cycles (denaturation, 30
sec at 94~ annealing, 30 sec at 62~ and elongation, 2
min. at 68~ and 25 cycles (denaturation, 30 sec at 94~
annealing and elongation, 2 min. at 68~ The PCR-amplified
777

Vol. 46, No. 4, 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL
fragment was sequenced and labeled using PCR-Digoxigenin
(DIG) labeling kit (Boehringer Mannheim). This labeled DNA
probe was then used to screen the cDNA library, and a
positive cDNA clone was isolated. Two additional PCR primers
indicated in Fig. 1 were used to amplify SMT3 DNA using
templates of genomic DNA (BIOS laboratories, New Haven, CT,
USA) and cloned SMT3 cDNA. The sizes of PCR-amplified
fragments from SMT3 cDNA and genomic DNA were compared on
1.0% agarose gel. The purified DNA fragments amplified from
these two templates were labeled with the Dye Terminator kit
(Perkin-Elmer, Foster City, CA, USA), and both strands of the
amplified DNAs were completely sequenced using automatic DNA
sequencer (Applied Biosystem, USA, model 377).
Northern blot analysis. The poly(A§ from embryo,
larvae and adult were obtained from Clontech (Palo Alto, CA).
Approximately 2 ~g of each poly(A+)-RNAs were ran on a 1.2%
formaldehyde agarose gel for 3 hours at i00 volts and
transferred upward to a Hybond-N § membrane (Amersham Life
Science, Cleveland, OH) by capillary action for an hour. The
membrane was fixed by UV crosslinking and then hybridized
with DIG-labeled probe (25 ng/ml) in DIG Easy hybridization
solution. Prehybridization and hybridization conditions were
strictly followed according to the Genius System User's guide
(version 3.0) except the temperature was 45~ The specific
SMT3 transcripts were detected by DIG luminescent detection
kit.
Analyses of structural and evolutionary relationships
among SMT3 proteins. The complete amino-acid sequence of
778

Vol. 46, No. 4, 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL
Drosophila SMT3
CACGCCCGGCATTCGACGCTCCGCAAAAGAAAAAAAAACTTTTTTGACCACTTA GCAGC TTCAACAAGCAAC CAAAAAATCAACATGTCTGACGAAAAGAAGGGAGGT
M S D E K K G G 8
GAGACCGAGCACATCAACCTGAAGGTCCTCGGCCAGGACAACGCCGTCGTCCAG E T E H I N L K V L G Q D N A V V Q 26 TTCAAGATCAAGAAGCACACACCCTTGAGGAAGCTGATGAACGCCTACTGCGAC F K I K K H T P L R K L M N A Y C D 44 CGTGCCGGACTCTC CATGCAGGTGGTGCGCTTCCGTTTCGACGGACAGCCCATC R A G L S M Q V V R F R F D G Q P I 62 AACGAGAACGACAC TCCGACCTCGCTGGAGATGGAGGAGGGCGACACCATCGAG N E N D T P T S L E M E E G D T I E 80 GTTTACCAGCAGCAGAC TGGTGGCGCTCCATAAGATTACTTAGTTAAGTTAGTT V Y Q Q Q T G G A P * 90 AC TC CTC TTACAAC TACACAC TTAAAACAAAAAAGAAAAAAAATACAAGAAAAA C CACAAAAGCAAAAACACAACAACAACAACATGAAGAATCCAACAAACCAGGC C CTAAGAATCGATTGAATATGCTTTTAGTACAACTGTAGATTCTAAATGCGTCTG TGTGCGTTAATAACAAAAACATTTGCAGACAAGAAAATGGT
Fig. i. The nucleotide and deduced amino acid sequences of
Drosophila SMT3 cDNA.
The stop codon TAA is indicated by an asterisk. Two PCR primers used are underlined.
Drosophila melanogaster SMT3 protein was deduced from the
cDNA and gene sequences determined in this investigation
(Genbank accession no. AF053083). The names of the organisms
and the Genbank accession numbers of other reported SMT3
proteins are as follows: Human, Homo sapiens (SMT3A, X99584;
SMT3B, L76416; SMT3C, X99586); mouse, Mus musculus (SMT3C,
AF033353), African frog, Xenopus laevis (SMT3C, Z97073);
nematode, Caenorhabditis elegans (U94830); Thale cress,
Arabidopsis thaliana (X99609); Rice, Oryza sativa (X99608);
chickpea, Cicer arietinum (AJ001901) and yeast, Saccharomyces
cerevisiae (U27233). The amino-acid sequences were aligned
using the multiple alignment program in GENEWORK computer
package based on the method of Feng and Doolittle (12). The
structural and evolutionary relationships were analyzed using
UPGMA method from the same computer program (13).
779

Vol. 46, No. 4, 1998 BIOCHEMISTRYond MOLECULAR BIOLOGY INTERNATIONAL
RESULTS AND DISCUSSION
Characterization of SMT3 cDNA and gene. A Drosophila SMT3
cDNA clone was isolated and its sequence of 581 nucleotides
determined (GeneBank accession no. AF053083). This SMT3 cDNA
contains a protein-coding sequence of 270 nucleotides, 5' (85
nucleotides) and 3' (226 nucleotides) untranslated regions
(Fig. i) . The sizes of PCR-amplified fragments, including
protein-coding region, from SMT3 cDNA and genomic DNA were
found to be the same on 1.0% agarose gel (Fig. 2), and their
sequences of 463 nucleotides were determined to be identical.
These results indicated the absence of intron in the protein-
coding region of Drosophila SMT3 gene. It is of interest that
the protein-coding sequence of nematode SMT3 gene is
interrupted by two small introns of 56 and 50 bp at coden
nos. 22-23 and 56, respectively (14).
Northern blot Analysis. The results of Northern blot
analysis indicated that two SMT3 transcripts of 2.5 kb and
1.5 kb were expressed in embryo, larvae and adult tissues,
although the transcript of 1.5 kb is most abundantly present
in larvae sample (Fig. 3). These size differences may be due
to either different polyadenylation sites used or alternative
splicing of the same SMT3 gene. It is also possible that they
represent different transcripts of two similar SMT3 genes.
Further, one might wonder why the small SMT3 protein of 90
amino acids requires such large transcripts.
Amino acid sequence comparison among SMT3 proteins. The
sequence of 90 amino acids deduced from the Drosophila SMT3
780

Vol. 46, No. 4, 1998 BIOCHEMISTRYand MOLECULAR BIOLOGY INTERNATIONAL
1 2 3 Kb b p
4 6 3 ~
I 1.0
0.5
Fig. 2. Size comparison of PCR:amplified fragments using
Drosophila genomic DNA and SMT3 cDNA as templates.
Lane i, genomic DNA as template; lane 2, SMT3 cDNA as
template; lane 3, size marker of 1 kb DNA ladder (BRL Life
Technologies).
I 2 3 Kb
2.5
1.5
Fig. 3. Northern blot analysis of Drosophila SMT3
transcripts.
Lane i, larvae; lane 2, adult; lane 3, embryo.
781
Vol. 46, No. 4, 1998 BIOCHEMISTRYand MOLECULAR BIOLOGY INTERNATIONAL
~ A
O o o o o ~ o o o ~
H H H > ~ H H H H
~ > ) H H H ~ H
Z ~ N N N M Z ~ N H H H H H H H H H H H
I N I M M M ~ O N ~
I l l ~ l l l ~ I l l ~ l l l ~
o ~l o m tn co m
0 ~ 0 0 0 0 ~ 0 0 0 0
o
Q o
~ N N N N ~ N N
N N N N N N N N N N m
m m m H m m H ~ ~ ~ H
R R R R R R R R R R R
N N ~ N ~ N N ~ N
O m ~
R -,4
~ H N "~ m r �9 0 Q ~ -~
~ ~ o u,q �9 c,q .,-i o
4J o o �9
(1) -,-I ~1 H 4J o ~ ~ o
�9 o ~-~ m +J N ~ o
-~ @ ~ �9 o o ~ o ~ oh
o ~ ~ o o
o �9 �9 a ~ rn ~ x
.~ o ' ~ ~ o o
�9 o ~
O~ -~ o -,~ o (.9 q5
o~ Oh
q3 R ~3
CO Oh
O
-,--I o
o
.r-,i
782
Vol. 46, No. 4, 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL
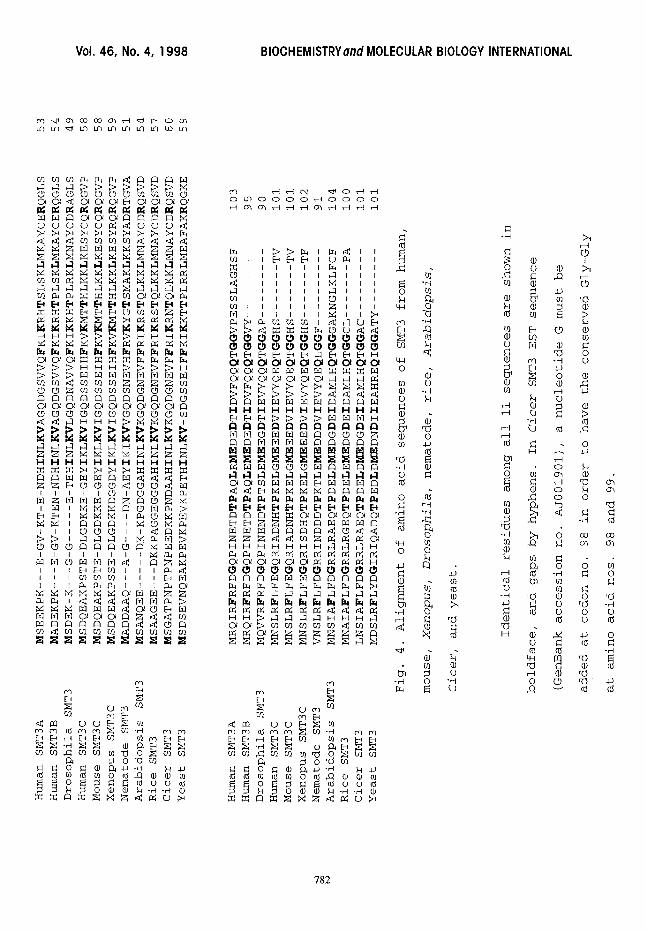
cDNA and gene sequences determined in this investigation was
aligned with the SMT3 protein sequences from human, mouse,
Xenopus, Drosophila, nematode, Arabidopsis, rice, Cicer and
yeast available in the databases (Fig. 4). The deduced
sequence of 90 amino acids from the Drosophila SMT3 cDNA and
gene exhibited 55%, 73%, 70% and 52% identity to the yeast
SMT3, human SMT3A, SMT3B and SMT3C protein sequences,
respectively. Human and mouse SMT3C proteins have identical
sequences. There are 21 identical residues out of Iii
positions compared among these ii SMT3 sequences. The
Human SMT3A
Human SMT3B
Drosophila SMT3
Human SMT3C
Mouse SMT3C
- - Xenopus SMT3C
Nematode SMT3
Arabidopsis SMT3
Rice SMT3
Cicer SMT3
Yeast SMT3
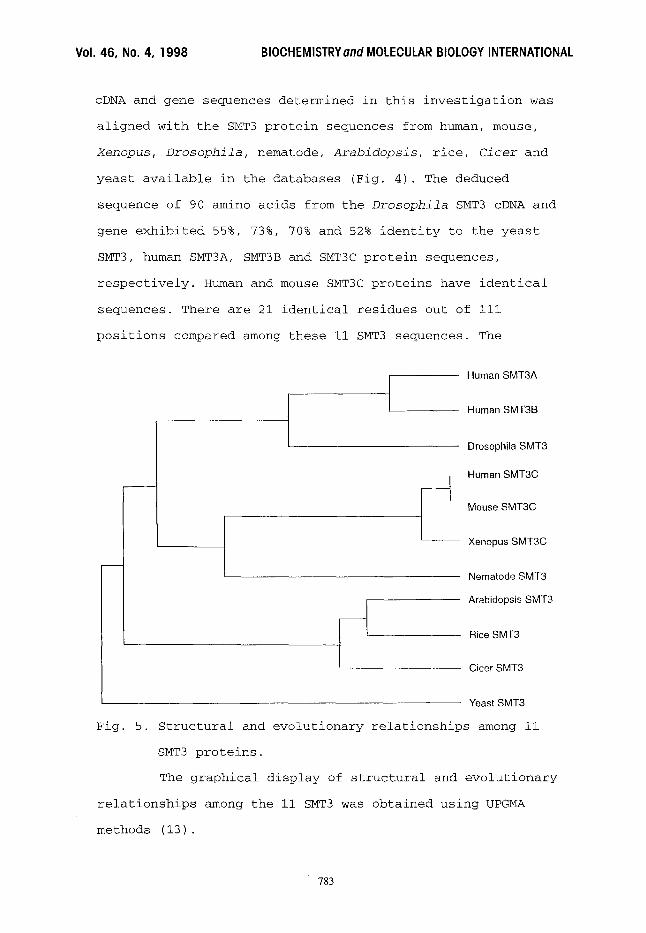
Fig. 5. Structural and evolutionary relationships among ii
SMT3 proteins.
The graphical display of structural and evolutionary
relationships among the ii SMT3 was obtained using UPGMA
methods (13).
783

Vol. 46, No. 4, 1998 BIOCHEMISTRYond MOLECULAR BIOLOGY INTERNATIONAL
significant amino acid identity and similarity of these SMT3
proteins among different species indicate their functional
importance. The SMT3C proteins from human, mouse and Xenopus
were reported to exhibit diverse functions (7-11, 15-16).
However, the exact function(s) of human SMT3A and SMT3B,
Drosophila, nematode and plant SMT3 proteins remain to be
elucidated experimentally. The structural relationships among
these ii SMT3 proteins from human, mouse, Xenopus,
Drosophila, nematode, Arabidopsis, rice, Cicer and yeast were
analyzed and the evolutionary tree constructed using UPGMA
method (13). Results are presented in Fig. 5. Drosophila SMT3
is clustered with human SMT3A and SMT3B, while nematode SMT3
is clustered with human, mouse and Xenopus SMT3C. Three plant
SMT3 are clustered into a separate group, and the yeast SMT3
is the out group.
ACKNOWLEDGMENTS
We thanks Drs. Frank Johnson and Po-Chuen Chan for
reading the manuscript. This investigation was supported in
part by grants NSC85-2732-B-II0-002 and NSC86-2313-B-II0-002
from National Science Council of Taiwan, ROC.
REFERENCES
i. Meluh, P.B., and Koshland, D. (1995) Mol. Biol. Cell
6, 794-807.
2. Brown, M. T., Goetsch, L., and Hartwell, L.H. (1993) J.
Cell. Biol. 9, 2544-2550.
3. Saitoh, H., Tomkiel, J., Cooke, C.A., Ratrie, H. III,
Maurer, M., Rothfield, N.F., and Earnshaw, W.C. (1992)
Cell 70, 115-125.
784

Vol. 46, No. 4, 1998 BIOCHEMISTRY and MOLECULAR BIOLOGY INTERNATIONAL
4. Brown, M. T. (1995) Gene 160, 111-116.
5. Lapenta, V., Chiurazzi, P., van der Spek, P., Pizzuti,
A., Hanooka, F. and Brahe, C. (1997) Genomics 40, 362-
366.
6. Mannen, H., Tseng, H.-M., Cho, C.-L., and Li, S.S.-L.
(1996) Biochem. Biophys. Res. Common. 222, 178-180.
7. Mahajan, R., Delphin, C., Quan, T., Gerace L., and
Melchior, F. (1997) Cell 88, 97-107.
8. Matunis, M.J., Coutava, E. and Blobel, G. (1996) J. Cell
Biol. 135, 14~7-1470.
9. Boddy, M.N., Howe, K., Ettkin, L.D., Solomon, E. and
Freemont, P.S. (1996) Oncogene 13, 971-982.
i0. Shen, Z., Pardington-Purtymun, P.E., Comeaux, J.C.,
Moyzis, R.K. and Chen, D.J. (1996) Genomics 36, 271-279.
ii. Okura. T., Gong, L., Kamitani, T., Wada, T., Okura, I.,
Wei, C.-F., Chang, H.-M. and Yeh, E.T.H. (1996) J.
Immunol. 15, 4277-4281.
12. Feng, D.F. and Doolittle, R.F. (1987) J. Mol. Evol. 25,
351-360.
13. Swofford, D.L. and Olsen, G.J. (1990) in Molecular
Systemics (Hillis, D.M. and Moritz, C. Eds.), pp. 411-
501, Sinauer, Sunderland, FLA.
14. Choudhury, B.K. and Li, S. S.-L. (1997 Biochem. Biophys.
Res. Commmen. 234, 788-791.
15. Howe, K., Willisaon, J., Boddy, N., Sheer, D., Freemont,
P.S. and Solomon, E. (1998) Genomics 47, 92-100.
16. Saitoh, H., Sparrow, D.B., Shiomi, T., Pu, R.T.,
Nishimoto, T., Mohun, T.J. and Dasso (1998) Current Biol.
8, 121-124.
785
Related Documents



![Ubiquitin and Ubiquitin-like Modifications in Viral ...1].pdf · Ubiquitin and Ubiquitin-like Modifications in Viral Infection and Immunity Abstracts of papers presented at the AUGUST](https://static.cupdf.com/doc/110x72/5e2d68ba2a69b505b71e58fa/ubiquitin-and-ubiquitin-like-modifications-in-viral-1pdf-ubiquitin-and-ubiquitin-like.jpg)







