Hypothalamic–Pituitary–Thyroid Axis Hormones Stimulate Mitochondrial Function and Biogenesis in Human Hair Follicles Silvia Vidali 1,9 , Jana Knuever 1,9,10 , Johannes Lerchner 2,11 , Melanie Giesen 3,11 , Tama ´s Bı ´ro ´ 4 , Matthias Klinger 5 , Barbara Kofler 6 , Wolfgang Funk 7 , Burkhard Poeggeler 1,12 and Ralf Paus 1,8 Thyroid hormones regulate mitochondrial function. As other hypothalamic–pituitary–thyroid (HPT) axis hormones, i.e., thyrotropin-releasing hormone (TRH) and thyrotropin (TSH), are expressed in human hair follicles (HFs) and regulate mitochondrial function in human epidermis, we investigated in organ-cultured human scalp HFs whether TRH (30 nM), TSH (10 mU ml 1 ), thyroxine (T 4 ) (100 nM), and triiodothyronine (T 3 ) (100 pM) alter intrafollicular mitochondrial energy metabolism. All HPT-axis members increased gene and protein expression of mitochondrial-encoded subunit 1 of cytochrome c oxidase (MTCO1), a subunit of respiratory chain complex IV, mitochondrial transcription factor A (TFAM), and Porin. All hormones also stimulated intrafollicular complex I/IV activity and mitochondrial biogenesis. The TSH effects on MTCO1, TFAM, and porin could be abolished by K1-70, a TSH-receptor antagonist, suggesting a TSH receptor–mediated action. Notably, as measured by calorimetry, T 3 and TSH increased follicular heat production, whereas T 3 /T 4 and TRH stimulated ATP production in cultured HF keratinocytes. HPT-axis hormones did not increase reactive oxygen species (ROS) production. Rather, T 3 and T 4 reduced ROS formation, and all tested HPT-axis hormones increased the transcription of ROS scavengers (catalase, superoxide dismutase 2) in HF keratinocytes. Thus, mitochondrial biology, energy metabolism, and redox state of human HFs are subject to profound (neuro-)endocrine regulation by HPT-axis hormones. The neuroendocrine control of mitochondrial biology in a complex human mini-organ revealed here may be therapeutically exploitable. Journal of Investigative Dermatology advance online publication, 15 August 2013; doi:10.1038/jid.2013.286 INTRODUCTION The hypothalamic–pituitary–thyroid (HPT) axis controls multi- ple metabolic processes in vertebrate organisms (Bassett and Williams, 2008; Costa-E-Sousa and Hollenberg, 2012), including human skin (Paus, 2010, 2011; Slominski et al., 2012). Stimulation of the pituitary gland by hypothalamus- derived thyrotropin-releasing hormone (TRH) leads to thyrotropin (TSH) secretion. TSH then stimulates the thyroid gland to synthesize and secrete the thyroid hormones (THs), thyroxine (T 4 ), and triiodothyronine (T 3 ) (Zoeller et al., 2007). Circulating T 4 is thought to largely act after deiodination into T 3 in peripheral organs, including human hair follicles (HFs) (Kaplan et al., 1988; van Beek et al., 2008; Safer et al., 2009). Several elements of the HPT axis are expressed in human skin in situ and/or some of its constituent cell populations in vitro: cultured human dermal and HF fibroblasts transcribe TRH (Slominski et al., 2002), whereas TSH mRNA and protein are found in normal human epidermis (Bodo ´ et al., 2010) and TSH receptor (TSHR) transcripts and protein in human skin and HF mesenchyme (Bodo ´ et al., 2009; Cianfarani et al., 2010). Both TRH and its receptor are expressed in the epithe- lium of normal human scalp HFs (Ga ´spa ´r et al., 2010, 2011). ORIGINAL ARTICLE 1 Department of Dermatology, University of Luebeck, Luebeck, Germany; 2 Institute of Physical Chemistry, TU Bergakademie, Freiberg, Germany; 3 Henkel Beauty Care, Henkel AG and Co. KGaA, Dusseldorf, Germany; 4 Department of Physiology, University of Debrecen, Debrecen, Hungary; 5 Department of Anatomy, University of Luebeck, Luebeck, Germany; 6 Research Program for Receptor Biochemistry and Tumor Metabolism, Department of Pediatrics, Paracelsus Medical University, Salzburg, Austria; 7 Klinik Dr Koslowski, Munich, Germany and 8 Institute of Inflammation and Repair, University of Manchester, and The Dermatology Centre, Salford Royal NHS Trust, Manchester, UK Correspondence: Ralf Paus, Department of Dematology, University of Luebeck, Ratzeburger Allee 160, 23538 Luebeck, Germany. E-mail: [email protected] or [email protected] 9 These authors contributed equally to this work. 10 Current address: Department of Dermatology, University of Cologne, Cologne, Germany 11 These authors contributed equally to this work. 12 Current address: QUIRIS Healthcare, Guetersloh, Germany Received 15 November 2012; revised 29 April 2013; accepted 31 May 2013; accepted article preview online 27 June 2013 Abbreviations: HF, hair follicle; HPT, hypothalamic–pituitary–thyroid; IR, immunoreactivity; MTCO1, mitochondrial-encoded subunit 1 of cytochrome c oxidase; ORS, outer root sheath; ROS, reactive oxygen species; SOD2, superoxide dismutase 2; TFAM, mitochondrial transcription factor A; TSH, thyrotropin; TSHR, thyrotropin receptor; TRH, thyrotropin-releasing hormone; T 3 , triiodothyronine; T 4 , thyroxine & 2013 The Society for Investigative Dermatology www.jidonline.org 1

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hypothalamic–Pituitary–Thyroid Axis HormonesStimulate Mitochondrial Function and Biogenesisin Human Hair FolliclesSilvia Vidali1,9, Jana Knuever1,9,10, Johannes Lerchner2,11, Melanie Giesen3,11, Tamas Bıro4, Matthias Klinger5,Barbara Kofler6, Wolfgang Funk7, Burkhard Poeggeler1,12 and Ralf Paus1,8
Thyroid hormones regulate mitochondrial function. As other hypothalamic–pituitary–thyroid (HPT) axishormones, i.e., thyrotropin-releasing hormone (TRH) and thyrotropin (TSH), are expressed in human hairfollicles (HFs) and regulate mitochondrial function in human epidermis, we investigated in organ-cultured humanscalp HFs whether TRH (30 nM), TSH (10 mU ml� 1), thyroxine (T4) (100 nM), and triiodothyronine (T3) (100 pM) alterintrafollicular mitochondrial energy metabolism. All HPT-axis members increased gene and protein expression ofmitochondrial-encoded subunit 1 of cytochrome c oxidase (MTCO1), a subunit of respiratory chain complex IV,mitochondrial transcription factor A (TFAM), and Porin. All hormones also stimulated intrafollicular complex I/IVactivity and mitochondrial biogenesis. The TSH effects on MTCO1, TFAM, and porin could be abolished by K1-70,a TSH-receptor antagonist, suggesting a TSH receptor–mediated action. Notably, as measured by calorimetry, T3
and TSH increased follicular heat production, whereas T3/T4 and TRH stimulated ATP production in cultured HFkeratinocytes. HPT-axis hormones did not increase reactive oxygen species (ROS) production. Rather, T3 and T4
reduced ROS formation, and all tested HPT-axis hormones increased the transcription of ROS scavengers(catalase, superoxide dismutase 2) in HF keratinocytes. Thus, mitochondrial biology, energy metabolism, andredox state of human HFs are subject to profound (neuro-)endocrine regulation by HPT-axis hormones. Theneuroendocrine control of mitochondrial biology in a complex human mini-organ revealed here may betherapeutically exploitable.
Journal of Investigative Dermatology advance online publication, 15 August 2013; doi:10.1038/jid.2013.286
INTRODUCTIONThe hypothalamic–pituitary–thyroid (HPT) axis controls multi-ple metabolic processes in vertebrate organisms (Bassett andWilliams, 2008; Costa-E-Sousa and Hollenberg, 2012),including human skin (Paus, 2010, 2011; Slominski et al.,2012). Stimulation of the pituitary gland by hypothalamus-derived thyrotropin-releasing hormone (TRH) leads tothyrotropin (TSH) secretion. TSH then stimulates the thyroidgland to synthesize and secrete the thyroid hormones (THs),thyroxine (T4), and triiodothyronine (T3) (Zoeller et al., 2007).Circulating T4 is thought to largely act after deiodinationinto T3 in peripheral organs, including human hair follicles(HFs) (Kaplan et al., 1988; van Beek et al., 2008; Safer et al.,2009).
Several elements of the HPT axis are expressed in humanskin in situ and/or some of its constituent cell populationsin vitro: cultured human dermal and HF fibroblasts transcribeTRH (Slominski et al., 2002), whereas TSH mRNA and proteinare found in normal human epidermis (Bodo et al., 2010) andTSH receptor (TSHR) transcripts and protein in human skinand HF mesenchyme (Bodo et al., 2009; Cianfarani et al.,2010). Both TRH and its receptor are expressed in the epithe-lium of normal human scalp HFs (Gaspar et al., 2010, 2011).
ORIGINAL ARTICLE
1Department of Dermatology, University of Luebeck, Luebeck, Germany;2Institute of Physical Chemistry, TU Bergakademie, Freiberg, Germany;3Henkel Beauty Care, Henkel AG and Co. KGaA, Dusseldorf, Germany;4Department of Physiology, University of Debrecen, Debrecen, Hungary;5Department of Anatomy, University of Luebeck, Luebeck, Germany;6Research Program for Receptor Biochemistry and Tumor Metabolism,Department of Pediatrics, Paracelsus Medical University, Salzburg, Austria;7Klinik Dr Koslowski, Munich, Germany and 8Institute of Inflammation andRepair, University of Manchester, and The Dermatology Centre, Salford RoyalNHS Trust, Manchester, UK
Correspondence: Ralf Paus, Department of Dematology, University ofLuebeck, Ratzeburger Allee 160, 23538 Luebeck, Germany.E-mail: [email protected] or [email protected]
9These authors contributed equally to this work.10Current address: Department of Dermatology, University of Cologne,Cologne, Germany11These authors contributed equally to this work.12Current address: QUIRIS Healthcare, Guetersloh, Germany
Received 15 November 2012; revised 29 April 2013; accepted 31 May 2013;accepted article preview online 27 June 2013
Abbreviations: HF, hair follicle; HPT, hypothalamic–pituitary–thyroid; IR,immunoreactivity; MTCO1, mitochondrial-encoded subunit 1 of cytochrome coxidase; ORS, outer root sheath; ROS, reactive oxygen species; SOD2,superoxide dismutase 2; TFAM, mitochondrial transcription factor A; TSH,thyrotropin; TSHR, thyrotropin receptor; TRH, thyrotropin-releasing hormone;T3, triiodothyronine; T4, thyroxine
& 2013 The Society for Investigative Dermatology www.jidonline.org 1

Human skin may therefore possess a peripheral equivalent ofthe central HPT axis (Slominski et al., 2002, Bodo et al., 2010;Paus, 2010).
THs directly alter important human HF functions, e.g., theyprolong anagen, stimulate hair matrix keratinocyte proli-feration and pigmentation, and modulate intrafollicularkeratin expression (van Beek et al., 2008). Although TSHregulates the expression of selected keratins in human HFs(Ramot et al., 2011), its functions in HF biology remainunclear (Bodo et al., 2009; Paus, 2010). Although THs andTSH do not significantly modulate new hair shaft forma-tion in vitro (van Beek et al., 2008; Bodo et al., 2009), TRHpotently stimulates hair shaft production, HF pigment-ation, anagen duration (Gaspar et al., 2010, 2011), andintrafollicular prolactin and prolactin-receptor expressionin situ (Langan et al., 2010).
THs greatly affect mitochondria (Harper and Seifert, 2008,Weitzel and Iwen, 2003, 2011), whose activities controlhuman energy metabolism and homeostasis (Scheffler, 2008,Portman, 2008; Kharlip and Cooper, 2009). Mitochondriaprocess glycolysis and fatty acid oxidation products togenerate ATP, contain enzymes critical for multiple bio-synthetic processes, contribute to the amino-acid metabolismand ion homeostasis, regulate cell death pathways by reactiveoxygen species (ROS) and Ca2þ signaling, and are involvedin aging, cancer, and the pathogenesis of an ever-increasinglist of human diseases (Birch-Machin, 2006; Scheffler, 2008;Wallace and Fan, 2009; Larsson, 2010; Wallace, 2010).Therefore, fully understanding the hormonal controls ofmitochondrial biology is of major biological and clinicalinterest also in the context of skin biology.
However, mainstream mitochondrial research has largelyignored human skin as a research model, even though it hasrecently turned out to serve as a most instructive model systemfor identifying novel, clinically relevant hormonal controls ofmitochondrial function and biogenesis: both TRH and TSHare powerful neuroendocrine regulators of mitochondrialbiology in human epidermis (Poeggeler et al., 2010a;Knuever et al., 2012). However, whether THs, TRH, and/orTSH exert a similar function in human HFs is unknown, asHF mitochondrial biology has remained almost completelyunexplored.
As hair growth is a highly energy-consuming process(Kealey et al., 1994; Stenn and Paus, 2001), we hypo-thesized that HPT-axis hormones may also regulate humanHF energy metabolism and mitochondrial function.
Therefore, we asked whether and how TRH, TSH, and THsinfluence intrafollicular mitochondrial biology in the absenceof neural, vascular, and extrafollicular hormonal inputs inserum-free organ culture of healthy human scalp HFs. Eachhormone was tested in a selected concentration that we hadpreviously documented to significantly alter defined humanHF biology parameters (see Supplementary Text S4 online).The list of mitochondrial read-out parameters that we hadpreviously established for organ-cultured human epidermis(Poeggeler et al., 2010a; Knuever et al., 2012) was furtherextended, including porin expression as an additional indi-cator of mitochondrial biogenesis (Feichtinger et al., 2011)
and calorimetry as an indicator of mitochondrial energymetabolism–dependent heat production (Lerchner et al.,2008a, b). Furthermore, in cultured human outer root sheath(ORS) keratinocytes, ATP and ROS production and theexpression of important ROS scavenger enzymes (catalase,superoxide dismutase 2 (SOD2)) were measured.
RESULTSHPT-axis elements increase MTCO1 and TFAM proteinexpression in situ, whereas T3/T4 stimulate MTCO1 and TFAMtranscription
Previously, we had shown that TSH upregulates MTCO1(mitochondrial-encoded subunit 1 of cytochrome c oxidase)gene and protein expression in human HFs (Bodo et al., 2009)and human epidermis, where it also increases the expres-sion of mitochondrial transcription factor A (TFAM) (Poeggeleret al., 2010a). TFAM controls mtDNA synthesis (Maniura-Weber et al., 2004; Scarpulla, 2008; Vina et al., 2009;Hallberg and Larsson, 2011), maintains and stabilizesmtDNA (Larsson et al., 1998), and is thus indispensable fora functional electron transport chain (Baris et al., 2011).
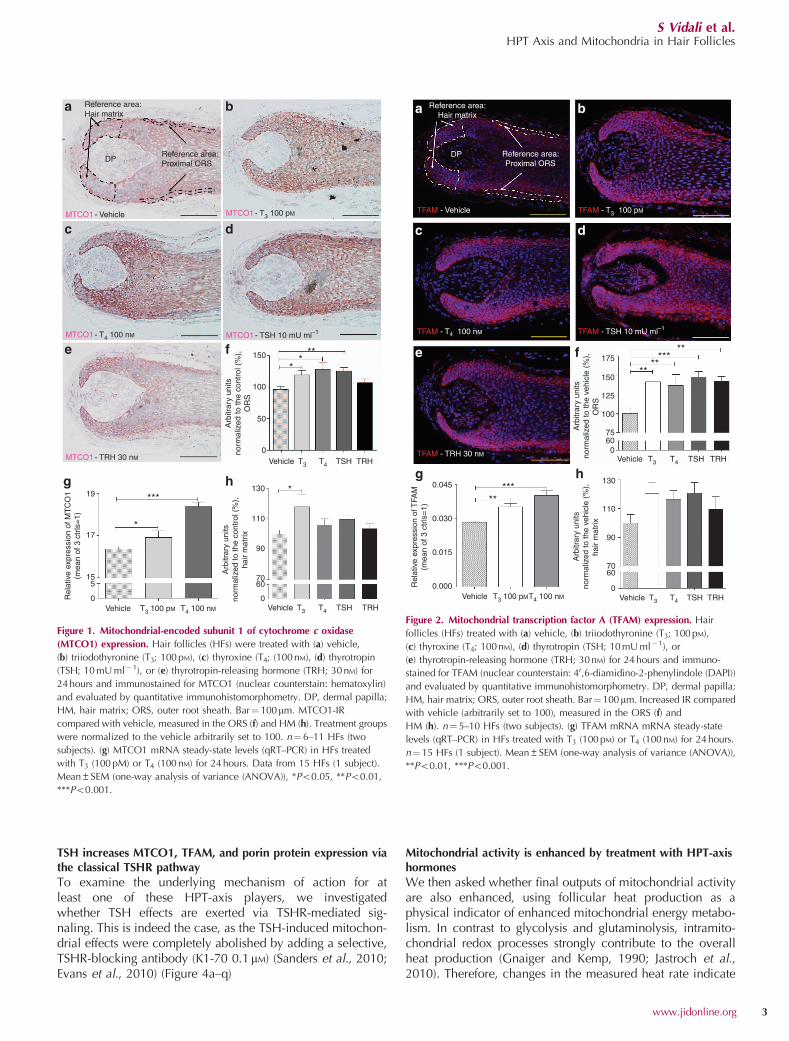
Therefore, we first assessed the impact of HPT-axishormones on MTCO1 and TFAM expression in human scalpHFs. This showed that T3, T4, and TSH significantly increasedMTCO1 immunoreactivity (IR) in the proximal ORS in situ,where TRH showed only a slight increase in IR (Figure 1a–f).TH treatment also increased intrafollicular MTCO1 mRNAtranscription (Figure 1g). TFAM protein expression in situwas also significantly increased after 24 hours of treatmentwith TRH, TSH, and THs (Figure 2a–f); T3 and T4 alsostimulated TFAM mRNA expression (Figure 2g). MTCO1 andTFAM-IR were analyzed in the hair matrix as well (Figures 1hand 2h). Interestingly, although T3 significantly increasedMTCO1-IR (Figure 1h), the other HPT hormones neitheraffected MTCO1 nor TFAM-IR (for further qPCR results, seeSupplementary Text S1 online, Supplementary Figure S1online).
T3/T4, TRH, and TSH treatment increases mitochondrialbiogenesis
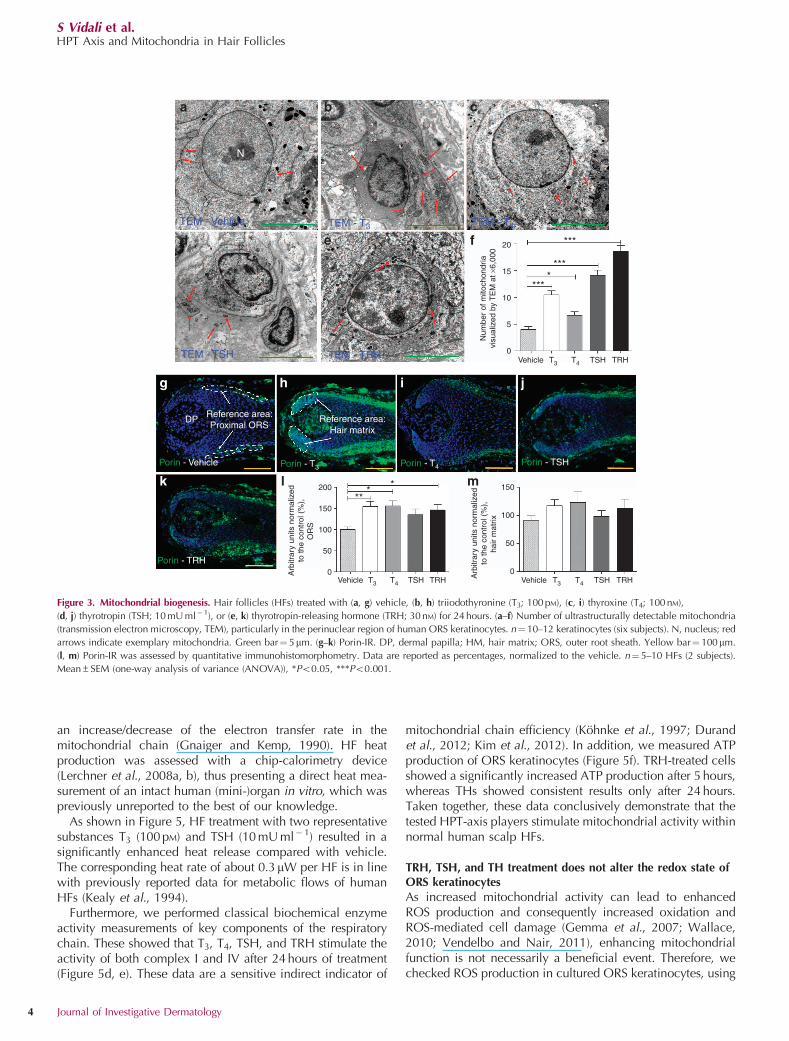
This raised the question whether HPT-axis hormones canstimulate intrafollicular mitochondrial biogenesis, as wehave already shown for TSH and TRH in human epidermis(Poeggeler et al., 2010a; Knuever et al., 2012). Transmissionelectron microscopy showed that the number of ultrastruc-turally detectable perinuclear mitochondria was upregulatedby all HPT-axis players in human ORS keratinocytes in situ(Figure 3a–f). To confirm this, porin-IR was assessed. Porinis located in the mitochondrial membrane, acts as achannel that enables small-molecule diffusion, and is wellestablished as a marker for mitochondrial mass (Feichtingeret al., 2011). Treatment for 24 hours with T3 and TRHsignificantly increased ORS porin-IR in situ, whereas T4 andTSH showed just a stimulatory trend (Figure 3g–l). Asporin-IR in the hair matrix was not significantly affected(Figure 3m), all tested HPT-axis elements stimulate mito-chondrial biogenesis in the HF epithelium, but preferentiallyin the ORS.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
2 Journal of Investigative Dermatology
TSH increases MTCO1, TFAM, and porin protein expression viathe classical TSHR pathway
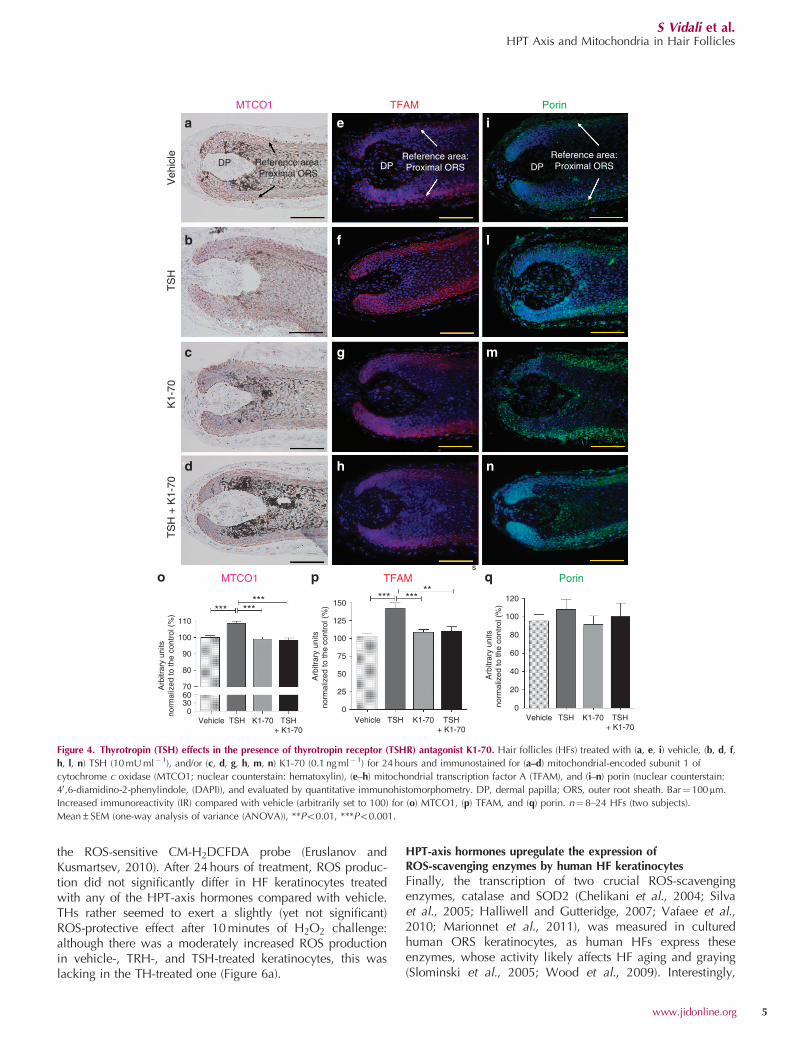
To examine the underlying mechanism of action for atleast one of these HPT-axis players, we investigatedwhether TSH effects are exerted via TSHR-mediated sig-naling. This is indeed the case, as the TSH-induced mitochon-drial effects were completely abolished by adding a selective,TSHR-blocking antibody (K1-70 0.1mM) (Sanders et al., 2010;Evans et al., 2010) (Figure 4a–q)
Mitochondrial activity is enhanced by treatment with HPT-axishormones
We then asked whether final outputs of mitochondrial activityare also enhanced, using follicular heat production as aphysical indicator of enhanced mitochondrial energy metabo-lism. In contrast to glycolysis and glutaminolysis, intramito-chondrial redox processes strongly contribute to the overallheat production (Gnaiger and Kemp, 1990; Jastroch et al.,2010). Therefore, changes in the measured heat rate indicate
DP
a b
c d
e f
g h***
*
*19
17
155
0 0
Arb
itrar
y un
itsno
rmal
ized
to th
e co
ntro
l (%
),O
RS
6070
90
110
130
150
100
50
0
Rel
ativ
e ex
pres
sion
of M
TC
O1
(mea
n of
3 c
trls
=1)
Vehicle VehicleT3 100 pM T3 T4 TSH TRH
Vehicle T3 T4 TSH TRH
T4 100 nM
MTCO1 - Vehicle MTCO1 - T3 100 pM
MTCO1 - T4 100 nM
MTCO1 - TRH 30 nM
MTCO1 - TSH 10 mU ml–1
Reference area:Hair matrix
Reference area:Proximal ORS
**
**
Arb
itrar
y un
itsno
rmal
ized
to th
e co
ntro
l (%
),ha
ir m
atrix
Figure 1. Mitochondrial-encoded subunit 1 of cytochrome c oxidase
(MTCO1) expression. Hair follicles (HFs) were treated with (a) vehicle,
(b) triiodothyronine (T3; 100 pM), (c) thyroxine (T4; (100 nM), (d) thyrotropin
(TSH; 10 mU ml�1), or (e) thyrotropin-releasing hormone (TRH; 30 nM) for
24 hours and immunostained for MTCO1 (nuclear counterstain: hematoxylin)
and evaluated by quantitative immunohistomorphometry. DP, dermal papilla;
HM, hair matrix; ORS, outer root sheath. Bar¼ 100mm. MTCO1-IR
compared with vehicle, measured in the ORS (f) and HM (h). Treatment groups
were normalized to the vehicle arbitrarily set to 100. n¼6–11 HFs (two
subjects). (g) MTCO1 mRNA steady-state levels (qRT–PCR) in HFs treated
with T3 (100 pM) or T4 (100 nM) for 24 hours. Data from 15 HFs (1 subject).
Mean±SEM (one-way analysis of variance (ANOVA)), *Po0.05, **Po0.01,
***Po0.001.
TFAM - Vehicle
DP
a b
c d
e f
g h
Reference area:Hair matrix
Reference area:Proximal ORS
TFAM - T3 100 pM
TFAM - T4 100 nM
TFAM - TRH 30 nM
TFAM - TSH 10 mU ml–1
0.045 130
110
90
7060
0Vehicle T3 T4 TSH TRH
0.030
0.015
0.000Rel
ativ
e ex
pres
sion
of T
FAM
(mea
n of
3 c
trls
=1)
Arb
itrar
y un
itsno
rmal
ized
to th
e ve
hicl
e (%
),ha
ir m
atrix
Vehicle T3 100 pMT4 100 nM
Vehicle T3 T4 TSH TRH
*****
****
*****
175
150
125
100
75600
Arb
itrar
y un
itsno
rmal
ized
to th
e ve
hicl
e (%
),O
RS
Figure 2. Mitochondrial transcription factor A (TFAM) expression. Hair
follicles (HFs) treated with (a) vehicle, (b) triiodothyronine (T3; 100 pM),
(c) thyroxine (T4; 100 nM), (d) thyrotropin (TSH; 10 mU ml�1), or
(e) thyrotropin-releasing hormone (TRH; 30 nM) for 24 hours and immuno-
stained for TFAM (nuclear counterstain: 40,6-diamidino-2-phenylindole (DAPI))
and evaluated by quantitative immunohistomorphometry. DP, dermal papilla;
HM, hair matrix; ORS, outer root sheath. Bar¼ 100mm. Increased IR compared
with vehicle (arbitrarily set to 100), measured in the ORS (f) and
HM (h). n¼5–10 HFs (two subjects). (g) TFAM mRNA mRNA steady-state
levels (qRT–PCR) in HFs treated with T3 (100 pM) or T4 (100 nM) for 24 hours.
n¼ 15 HFs (1 subject). Mean±SEM (one-way analysis of variance (ANOVA)),
**Po0.01, ***Po0.001.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
www.jidonline.org 3
an increase/decrease of the electron transfer rate in themitochondrial chain (Gnaiger and Kemp, 1990). HF heatproduction was assessed with a chip-calorimetry device(Lerchner et al., 2008a, b), thus presenting a direct heat mea-surement of an intact human (mini-)organ in vitro, which waspreviously unreported to the best of our knowledge.
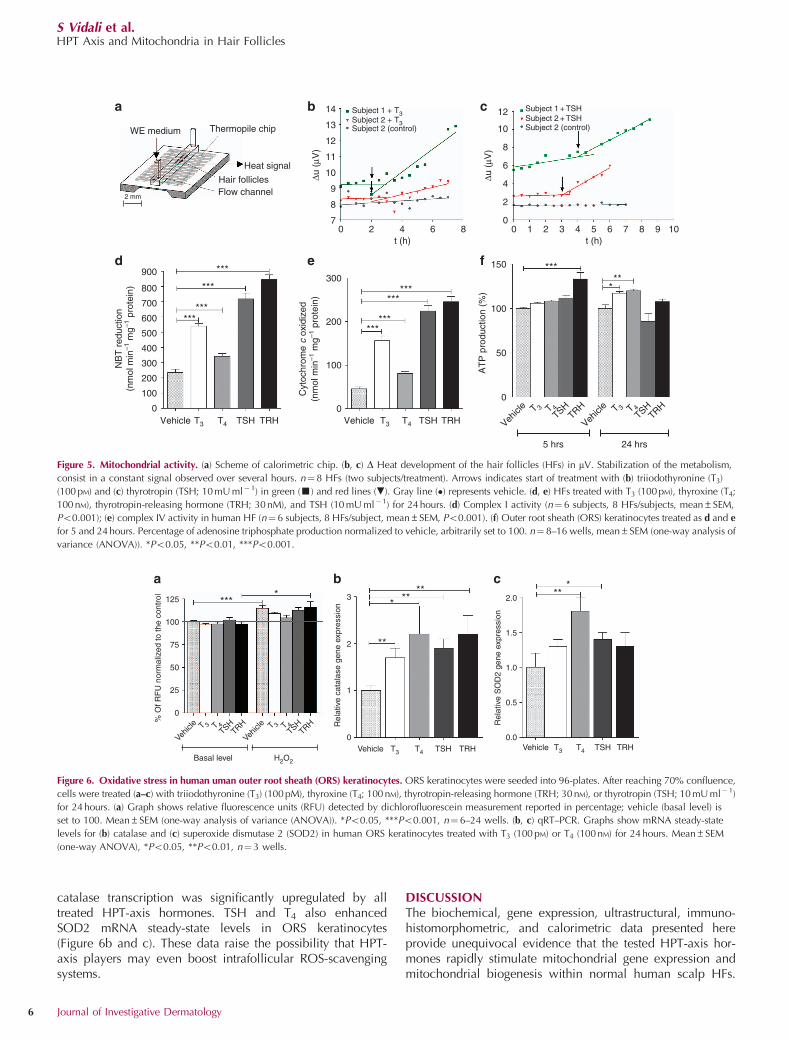
As shown in Figure 5, HF treatment with two representativesubstances T3 (100 pM) and TSH (10 mU ml�1) resulted in asignificantly enhanced heat release compared with vehicle.The corresponding heat rate of about 0.3mW per HF is in linewith previously reported data for metabolic flows of humanHFs (Kealy et al., 1994).
Furthermore, we performed classical biochemical enzymeactivity measurements of key components of the respiratorychain. These showed that T3, T4, TSH, and TRH stimulate theactivity of both complex I and IV after 24 hours of treatment(Figure 5d, e). These data are a sensitive indirect indicator of
mitochondrial chain efficiency (Kohnke et al., 1997; Durandet al., 2012; Kim et al., 2012). In addition, we measured ATPproduction of ORS keratinocytes (Figure 5f). TRH-treated cellsshowed a significantly increased ATP production after 5 hours,whereas THs showed consistent results only after 24 hours.Taken together, these data conclusively demonstrate that thetested HPT-axis players stimulate mitochondrial activity withinnormal human scalp HFs.
TRH, TSH, and TH treatment does not alter the redox state ofORS keratinocytes
As increased mitochondrial activity can lead to enhancedROS production and consequently increased oxidation andROS-mediated cell damage (Gemma et al., 2007; Wallace,2010; Vendelbo and Nair, 2011), enhancing mitochondrialfunction is not necessarily a beneficial event. Therefore, wechecked ROS production in cultured ORS keratinocytes, using
20
200 150
100
50
Arb
itrar
y un
its n
orm
aliz
edto
the
cont
rol (
%),
hair
mat
rix
0Vehicle
****
150
100
50
Arb
itrar
y un
its n
orm
aliz
edto
the
cont
rol (
%),
OR
S
0Vehicle T3 T4 TSH TRH T3 T4 TSH TRH
15
10
***
***
***
N
a b
d
g
k l m
h i j
e f
c
DP
Porin - Vehicle
Porin - TRH
Porin - T3 Porin - T4 Porin - TSH
Reference area:Proximal ORS
Reference area:Hair matrix
TEM - Vehicle TEM - T3
TEM - TSH TEM - TRH
TEM - T4
*
Num
ber
of m
itoch
ondr
iavi
sual
ized
by
TE
M a
t ×6,
000
5
0Vehicle T3 T4 TSH TRH
Figure 3. Mitochondrial biogenesis. Hair follicles (HFs) treated with (a, g) vehicle, (b, h) triiodothyronine (T3; 100 pM), (c, i) thyroxine (T4; 100 nM),
(d, j) thyrotropin (TSH; 10 mU ml� 1), or (e, k) thyrotropin-releasing hormone (TRH; 30 nM) for 24 hours. (a–f) Number of ultrastructurally detectable mitochondria
(transmission electron microscopy, TEM), particularly in the perinuclear region of human ORS keratinocytes. n¼ 10–12 keratinocytes (six subjects). N, nucleus; red
arrows indicate exemplary mitochondria. Green bar¼ 5mm. (g–k) Porin-IR. DP, dermal papilla; HM, hair matrix; ORS, outer root sheath. Yellow bar¼ 100mm.
(l, m) Porin-IR was assessed by quantitative immunohistomorphometry. Data are reported as percentages, normalized to the vehicle. n¼ 5–10 HFs (2 subjects).
Mean±SEM (one-way analysis of variance (ANOVA)), *Po0.05, ***Po0.001.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
4 Journal of Investigative Dermatology
the ROS-sensitive CM-H2DCFDA probe (Eruslanov andKusmartsev, 2010). After 24 hours of treatment, ROS produc-tion did not significantly differ in HF keratinocytes treatedwith any of the HPT-axis hormones compared with vehicle.THs rather seemed to exert a slightly (yet not significant)ROS-protective effect after 10 minutes of H2O2 challenge:although there was a moderately increased ROS productionin vehicle-, TRH-, and TSH-treated keratinocytes, this waslacking in the TH-treated one (Figure 6a).
HPT-axis hormones upregulate the expression ofROS-scavenging enzymes by human HF keratinocytes
Finally, the transcription of two crucial ROS-scavengingenzymes, catalase and SOD2 (Chelikani et al., 2004; Silvaet al., 2005; Halliwell and Gutteridge, 2007; Vafaee et al.,2010; Marionnet et al., 2011), was measured in culturedhuman ORS keratinocytes, as human HFs express theseenzymes, whose activity likely affects HF aging and graying(Slominski et al., 2005; Wood et al., 2009). Interestingly,
MTCO1
Veh
icle
DP DP DPReference area:Proximal ORS
Reference area:Proximal ORS
Reference area:Proximal ORS
TS
HK
1-70
TS
H +
K1-
70
TFAM Porin
110
s
Arb
itrar
y un
itsno
rmal
ized
to th
e co
ntro
l (%
)
Arb
itrar
y un
itsno
rmal
ized
to th
e co
ntro
l (%
)
Arb
itrar
y un
itsno
rmal
ized
to th
e co
ntro
l (%
) *** ****** *** ***
**
100
90
80
7060300
Vehicle TSH K1-70 TSH+ K1-70
150
125
100
75
50
25
0
120
100
80
60
40
20
0
Vehicle TSH K1-70 TSH+ K1-70
Vehicle TSH K1-70 TSH+ K1-70
MTCO1 TFAM Porin
a
b
c
d
e i
l
m
n
f
g
h
o p q
Figure 4. Thyrotropin (TSH) effects in the presence of thyrotropin receptor (TSHR) antagonist K1-70. Hair follicles (HFs) treated with (a, e, i) vehicle, (b, d, f,
h, l, n) TSH (10mU ml� 1), and/or (c, d, g, h, m, n) K1-70 (0.1 ng ml�1) for 24 hours and immunostained for (a–d) mitochondrial-encoded subunit 1 of
cytochrome c oxidase (MTCO1; nuclear counterstain: hematoxylin), (e–h) mitochondrial transcription factor A (TFAM), and (i–n) porin (nuclear counterstain:
40,6-diamidino-2-phenylindole, (DAPI)), and evaluated by quantitative immunohistomorphometry. DP, dermal papilla; ORS, outer root sheath. Bar¼ 100mm.
Increased immunoreactivity (IR) compared with vehicle (arbitrarily set to 100) for (o) MTCO1, (p) TFAM, and (q) porin. n¼8–24 HFs (two subjects).
Mean±SEM (one-way analysis of variance (ANOVA)), **Po0.01, ***Po0.001.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
www.jidonline.org 5
catalase transcription was significantly upregulated by alltreated HPT-axis hormones. TSH and T4 also enhancedSOD2 mRNA steady-state levels in ORS keratinocytes(Figure 6b and c). These data raise the possibility that HPT-axis players may even boost intrafollicular ROS-scavengingsystems.
DISCUSSIONThe biochemical, gene expression, ultrastructural, immuno-histomorphometric, and calorimetric data presented hereprovide unequivocal evidence that the tested HPT-axis hor-mones rapidly stimulate mitochondrial gene expression andmitochondrial biogenesis within normal human scalp HFs.
14
13
12
11
10
9
8
70
a b c
d e f
2 4 6 8t (h)
Subject 1 + T3Subject 2 + T3Subject 2 (control)
Δu (
μV)
12
10
8
6
4
2
00 1 2 3 4 5 6 7 8 9 10
t (h)
Subject 1 + TSHSubject 2 + TSHSubject 2 (control)
Δu (
μV)
900
800
700
600
500
400
300
200
100
0Vehicle T3 T4 TSH TRH Vehicle T3 T4 TSH TRH
NB
T r
educ
tion
(nm
ol m
in–1
mg–1
pro
tein
)
***
***
******
300
200
100
0
Cyt
ochr
ome
c ox
idiz
ed(n
mol
min
–1 m
g–1 p
rote
in)
******
******
150
100
50
0
Vehicl
e T 3 T 4TSH
TRH
Vehicl
e T 3 T 4TSH
TRH
5 hrs 24 hrs
*****
*
AT
P p
rodu
ctio
n (%
)
2 mm
WE medium Thermopile chip
Heat signal
Hair folliclesFlow channel
Figure 5. Mitochondrial activity. (a) Scheme of calorimetric chip. (b, c) D Heat development of the hair follicles (HFs) in mV. Stabilization of the metabolism,
consist in a constant signal observed over several hours. n¼8 HFs (two subjects/treatment). Arrows indicates start of treatment with (b) triiodothyronine (T3)
(100 pM) and (c) thyrotropin (TSH; 10 mU ml� 1) in green (’) and red lines (.). Gray line (�) represents vehicle. (d, e) HFs treated with T3 (100 pM), thyroxine (T4;
100 nM), thyrotropin-releasing hormone (TRH; 30 nM), and TSH (10 mU ml�1) for 24 hours. (d) Complex I activity (n¼ 6 subjects, 8 HFs/subjects, mean±SEM,
Po0.001); (e) complex IV activity in human HF (n¼ 6 subjects, 8 HFs/subject, mean±SEM, Po0.001). (f) Outer root sheath (ORS) keratinocytes treated as d and e
for 5 and 24 hours. Percentage of adenosine triphosphate production normalized to vehicle, arbitrarily set to 100. n¼ 8–16 wells, mean±SEM (one-way analysis of
variance (ANOVA)). *Po0.05, **Po0.01, ***Po0.001.
125 ****
*
***
** **
**
100
75
% O
f RF
U n
orm
aliz
ed to
the
cont
rol
50
25
0
0 0.0
1
0.5
1.5
2.0
1.0
2
3
Rel
ativ
e ca
tala
se g
ene
expr
essi
on
Rel
ativ
e S
OD
2 ge
ne e
xpre
ssio
n
Vehic
le T 3 T 4TSH
TRH
Vehic
le T 3 T 4TSH
TRH
Basal level H2O2
Vehicle T3 T4 TSH TRH Vehicle T3 T4 TSH TRH
a b c
Figure 6. Oxidative stress in human uman outer root sheath (ORS) keratinocytes. ORS keratinocytes were seeded into 96-plates. After reaching 70% confluence,
cells were treated (a–c) with triiodothyronine (T3) (100 pM), thyroxine (T4; 100 nM), thyrotropin-releasing hormone (TRH; 30 nM), or thyrotropin (TSH; 10 mU ml� 1)
for 24 hours. (a) Graph shows relative fluorescence units (RFU) detected by dichlorofluorescein measurement reported in percentage; vehicle (basal level) is
set to 100. Mean±SEM (one-way analysis of variance (ANOVA)). *Po0.05, ***Po0.001, n¼6–24 wells. (b, c) qRT–PCR. Graphs show mRNA steady-state
levels for (b) catalase and (c) superoxide dismutase 2 (SOD2) in human ORS keratinocytes treated with T3 (100 pM) or T4 (100 nM) for 24 hours. Mean±SEM
(one-way ANOVA), *Po0.05, **Po0.01, n¼ 3 wells.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
6 Journal of Investigative Dermatology

Moreover, TRH, TSH, and THs stimulate mitochondrialactivity. These findings are well in line with the long-recognized stimulatory role of THs on mitochondrial biologyand with the recent discovery of TRH and TSH as neuro-endocrine regulators of human epidermal mitochondrialfunction in situ (Poeggeler et al., 2010a; Knuever et al.,2012). Importantly, major mitochondrial stimulation wasexerted on ORS keratinocytes, and not in the hair matrix,the germinative part of the HF epithelium. The ORS might thusbe viewed as the energy storage compartment of the HF,perhaps to spare the germinative compartment from sourcesof oxidative stress (for detailed discussion, see SupplementaryText S2 online).
The fact that TSH action is completely abolished byantagonizing TSHR signaling strongly suggests that TSH actson HF mitochondria via its receptor rather than via alternativepathways. That TSH affects HF mitochondrial biology viaTSHR also correlates with the fact that TSHR, when bound byits ligand, activates a cyclic adenosine monophosphate/PKAcascade. This cascade not only promotes the release of THsbut also influences many other pathways, including mito-chondrial function and biogenesis as well as apoptosis(Antoni, 2012; Papa et al., 2002; Feliciello et al., 2005).Therefore, it is conceivable that TSH affects human HFmitochondrial biology mainly through cyclic adenosinemonophosphate release.
As the human epidermis expresses TSH mRNA and protein(Bodo et al., 2010) and human skin also expresses TSHR(Bodo et al., 2009, Cianfarani et al., 2010), we expected thelevels of TFAM, MTCO1, and porin to be modulated by theTSHR antagonist alone. Instead, these remained unchanged(Figure 4). This suggests that, in the absence of specific stimuli,intrinsic production of TSH in human HFs is low anddispensable for maintaining baseline TFAM, MTCO1, andporin levels.
Given that increased mitochondrial protein expression doesnot necessarily imply increased mitochondrial energy produc-tion, the latter was investigated by chip calorimetry andclassical enzyme activity assays. These experiments clearlydemonstrate that both T3 and TSH enhance HF heat pro-duction, although both hormones activate very distinctsignal transduction pathways. Furthermore, TRH, TSH, andTHs stimulate mitochondrial chain complex I and IV activityin human HFs and slightly increase ATP production inORS keratinocytes, well in line with our previous findings inthe epidermis and brain tissue (Poeggeler et al., 2010a, b;Knuever et al., 2012). As T3 already stimulates heat produc-tion after only 2 hours and slightly increases ATP productionafter 5 hours, its action on mitochondrial activity may not beexerted via the classical nuclear receptor pathway. It remainsto be determined whether the observed T3 effects operate viaintramitochondrial receptor isoforms (e.g., p43) (Wrutniaket al., 1995; Casas et al., 1999).
The current study represents a systematic characterizationof HF mitochondrial biology and encourages one to explorethe as under-investigated energy metabolism of humanHFs (Williams et al., 1993; Kealey et al., 1994). Our studyalso underscores that, contrary to conventional wisdom in
mainstream mitochondrial research, not only humanepidermis (Birch-Machin, 2006; Poeggeler et al., 2010a;Knuever et al., 2012) but also the metabolically highlyactive HF offers excellent model systems for dissecting thefunction, biogenesis, and controls of human mitochondriain situ. Human HF chip calorimetry even permits to directlymeasure heat production of a living human (mini-)organin vitro. Moreover, our data show that, besides THs, theintrafollicularly produced neurohormones, TSH and TRH,profoundly affect human HF energy metabolism.
Our findings underscore the importance of neuroendocrinecontrols not only in general mitochondrial biology (Poeggeleret al., 2010a, b; Knuever et al., 2012), but also in human skinand HF biology (Slominski and Wortsman, 2000; Arck et al.,2006; Slominski et al., 2002, 2007; Paus, 2010, 2011;Zmijewski and Slominski, 2011). The lessons learned fromstudying organ-cultured human HFs may well be transferrableto other, less accessible human tissues.
The current data also provide clinically relevant pointers tohow HF aging and disease correlated with declining mito-chondrial function might be effectively counteracted in thefuture by endogenous neurohormones produced in the humanepithelium, e.g., TRH (Gaspar et al., 2010) and TSH (Bodoet al., 2010). This also applies to THs, which have long beenknown to modulate hair shaft quality and/or pigmentation(Messenger, 2000; van Beek et al., 2008). Both TRH and T4
are administered routinely in thyroid medicine and areFDA-approved agents with a well-known toxicity profile.Therefore, regulatory hurdles to reposition these hormonesfor novel ‘‘mitochondrial hair medicine’’ approaches arerelatively low.
Current ‘‘mitochondrial medicine’’ concepts (e.g., Wallaceet al., 2010; Wallace, 2011) assume that, in systems withdeclining mitochondrial function, the therapeutic increaseof mitochondrial activity generally exerts beneficial effects.However, increased mitochondrial activity could also haveundesirable sequelae, such as increased ROS production(Gemma et al., 2007; Vendelbo and Nair, 2011). However,our HF keratinocyte data suggest that HPT-axis hormonesstimulate HF mitochondria without increasing ROS formationor affecting HF vitality (for details, Supplementary Text S3online). These hormones may even promote intrafollicularROS scavenging by upregulating catalase and SOD2 transcrip-tion. Evidently, only long-term stimulation experiments andin-depth analyses of ROS production, ROS damage indi-cators, and scavenging system efficiency can dissipateremaining concerns that long-term administration of HPT-axishormones might tip the balance of desired toward adversemitochondrial effects. Such follow-up studies are needed toinform future clinical applications of HPT-axis hormones ingeneral ‘‘mitochondrial medicine’’ and gerontobiology.
In summary, we show that mitochondrial function, energymetabolism, and biogenesis of human HFs are all subject toprofound (neuro-)endocrine regulation by TRH, TSH, andTHs. This makes it even more pressing to further explore theHPT-axis equivalent system of human skin and its appendages(Paus, 2010; Slominski et al., 2002, 2012), and to explore howthis system can be therapeutically targeted.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
www.jidonline.org 7

MATERIALS AND METHODSSkin samples, human HF organ culture
Human scalp skin samples were obtained after written informed
consent and institutional research board ethics license (University of
Luebeck) from 14 healthy middle-aged women undergoing routine
cosmetic plastic surgery, adhering to Helsinki guidelines. The varying
numbers of HFs and subjects are stated in each figure legend.
HF organ culture was performed in serum-free Williams’ E medium
supplemented with glutamine, insulin, hydrocortisone, and an anti-
biotic/antimycotic mixture as described before (Philpott et al., 1990;
Bodo et al., 2009; Kloepper et al., 2010; Gaspar et al., 2011). After
pre-culture of microdissected HFs for 24 hours to minimize
dissection-associated disturbances, vehicle, T3, T4, TRH, TSH
(Sigma-Aldrich, Schnelldorf, Germany), and/or the TSHR antagonist
K1-70 (RSR, Cardiff, UK) were added for an additional 24 hours.
After treatment, the HF samples were shock-frozen in liquid nitrogen
and stored at � 80 1C. Sections (6mm thick) were obtained for
immunostainings. As very limited human HFs were available, one
carefully selected concentration was assessed for each hormone,
based on prior published results: T3 (100 pM), T4 (100 nM) (van Beek
et al., 2008), TRH (30 nM) (Gaspar et al., 2010, 2011), and TSH
(10 mU ml� 1) (Bodo et al., 2009; Poeggeler et al., 2010a). The
concentration of 0.1mg ml� 1 K1-70 was chosen according to the
manufacturer’s protocols (Evans et al., 2010; Sanders et al., 2010)
(further details, Supplementary Text S4 online).
Human ORS keratinocyte culture
Human ORS keratinocyte cultures were obtained from face-lift
surgery samples of healthy donors. Primary cultures of ORS keratino-
cytes were established from plucked anagen HFs, as described
(Philpott and Kealey, 1994).
Immunohistochemistry, immunofluoresescence, and quantitativeimmunohistomorphometry
MTCO1-IR was determined as previously described (Poeggeler et al.,
2010a, b; Knuever et al., 2012). TFAM- and porin-IR were investi-
gated by fluorescence microscopy. For the mitochondrial transcrip-
tion factor TFAM, a rabbit polyclonal antiserum (kindly provided by
Rudolf Wiesner, University of Cologne, Germany) was used (1:200,
O/N at þ 4 1C). As a secondary antibody, we used a goat anti-rabbit
Rhodamine (Jackson Immuno Research, Newmarket, Suffolk, UK)
(1:200, 2 hours at RT).
VDAC1/porin was used to depict mitochondrial biogenesis
(Feichtinger et al., 2011), using a rabbit polyclonal antibody (1:600,
O/N at þ 4 1C). As a secondary antibody, we used a goat anti-rabbit
biotinylated antibody (Jackson Immuno Research) (1:200, 45 minutes
at RT). The TSA method was used for detection (TSA kit, FITC, Perkin
Elmer, Rodgau, Germany). Omission of the primary antibody served
as negative control. Cell nuclei were demarcated by 40,6-diamidino-
2-phenylindole, and images were obtained by digital microscopy
(Keyence, Neu-Isenburg, Germany). Quantitative immunohisto-
morphometry was performed by assessing the relative intensity
of IR in precisely defined HF reference areas with ImageJ (NIH,
Bethesda, MD).
Transmission electron microscopy
Transmission electron microscopy was performed as previously
described (Kramer et al., 2005; Knuever et al., 2012). Briefly, after
2 hours of fixation with 2.5% glutaraldehyde, HFs were washed in
0.1 M cacodylate buffer, pH 7.2. Using 1% osmiumtetroxide, samples
were postfixed, dehydrated in ethanol, and embedded in araldite
(Fluka, Buchs, Switzerland). Ultrathin sections were stained with
uranyl acetate and lead citrate, and then viewed with an electron
microscope 400 (Philips, Hamburg, Germany).
Chip calorimetry
Heat measurements as a reliable indicator of follicular mitochondrial
activity/energy metabolism were performed using an innovative
prototype chip calorimeter (Lerchner et al., 2008a, b) (for methodol-
ogical details on Calorimetry and prototype chip functionality, see
Supplementary Text S5 online).
Quantitative real-time PCR (qRT–PCR)
Steady-state mRNA levels from organ-cultured human HFs were
studied by qRT–PCR after 24 hours of incubation with the HPT-axis
hormones for the following key genes of mitochondrial biogenesis
and/or energy metabolism: MTCO1, TFAM, heat-shock protein 60,
peroxisome proliferator–activated receptor gamma coactivator-
1alpha, and brain and muscle aryl hydrocarbon receptor nuclear
translocator-like protein 1 (Cappello et al., 2008; Jung-Hynes et al.,
2010; Safdar et al., 2011). Gene expression was assessed by quantita-
tive real-time PCR performed on an ABI PRISM 7000 Sequence
Detection System (Applied Biosystems, Life Technologies, Darmstadt,
Germany) according to the published protocol, and normalized to
one of three tested housekeeping genes that proved to be least
regulated by the test hormone (Knuever et al., 2012).
The qRT–PCR in ORS keratinocytes for SOD2 and catalase was
performed as previously described in Giesen et al., 2011 (further
details, Supplementary Text S1, S6 online).
Biochemical assays
Complex I and IV activity were analyzed in HF homogenates as
described by (Poeggeler et al., 2010a, b), according to the protocols
of Mazzio and Soliman, 2004, Dabbeni-Sala et al., 2001, and Rustin
et al., 1994. Both experiments were performed with eight HFs each
and were repeated multiple times, with HFs from six different
subjects.
ATP quantification was performed on ORS cells seeded in a black
96-well plate (25,000 cells per well) with fully supplemented medium
(Limat and Noser, 1986). After first 24 hours of incubation, treatments
were added for further 24 hours (substances dissolved in medium
without FCS, T3, and Ham F12). The ATP Assay was performed
according to the manufacturer’s instructions (ATPlite, Perkin Elmer).
ROS production in ORS keratinocytes
Production of ROS was detected fluorometrically using the specific
dye CM-H2DCFDA (Life Technologies) according to the manufac-
turer’s protocol (further details, Supplementary Text S7 online).
Statistical analysis
If not stated otherwise, all data were analyzed using one-way analysis
of variance (Tukey’s test) by using the Graph Pad Prism software
(Graph Pad Software, La Jolla, CA).
CONFLICT OF INTERESTThe authors state no conflict of interest.
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
8 Journal of Investigative Dermatology

ACKNOWLEDGMENTSWe gratefully appreciate Swantje Haeger, Antje Winter-Keil, andGabriele Scheel for excellent technical support and Dr Balazs I. Toth andDr Arno Kromminga for PCR analyses. This work was supported in part by afaculty grant from the University of Luebeck to RP, and by the ManchesterHealth Sciences Centre.
SUPPLEMENTARY MATERIAL
Supplementary material is linked to the online version of the paper at http://www.nature.com/jid
REFERENCES
Antoni FA (2012) New paradigms in cAMP signalling. Mol Cell Endocrinol353:3–9
Arck PC, Slominski A, Theoharides TC et al. (2006) Neuroimmunology ofstress: skin takes center stage. J Invest Dermatol 126:1697–704
Baris OR, Klose A, Kloepper JE et al. (2011) The mitochondrial electrontransport chain is dispensable for proliferation and differentiation ofepidermal progenitor cells. Stem Cells 29:1459–68
Bassett JH, Williams GR (2008) Critical role of the hypothalamic-pituitary-thyroid axis in bone. Bone 3:418–26
Birch-Machin MA (2006) The role of mitochondria in ageing and carcinogen-esis. Clin Exp Dermatol 31:548–52
Bodo E, Kromminga A, Bıro T et al. (2009) Human female hair follicles are adirect, nonclassical target for thyroid-stimulating hormone. J InvestDermatol 129:1126–39
Bodo E, Kany B, Gaspar E et al. (2010) Thyroid-stimulating hormone, a novel,locally produced modulator of human epidermal functions, is regulatedby thyrotropin-releasing hormone and thyroid hormones. Endocrinology151:1633–42
Cappello F, Conway de Macario E, Marasa L et al. (2008) Hsp60 expression,new locations, functions and perspectives for cancer diagnosis andtherapy. Cancer Biol Ther 7:801–9
Casas F, Rochard P, Rodier A et al. (1999) A variant form of the nucleartriiodothyronine receptor c-ErbAlpha 1 plays a direct role in regulation ofmitochondrial RNA synthesis. Mol Cell Biol 19:7913–24
Cianfarani F, Baldini E, Cavalli A et al. (2010) TSH receptor and thyroid-specific gene expression in human skin. J Invest Dermatol 130:93–101
Chelikani P, Fita I, Loewen PC (2004) Diversity of structures and propertiesamong catalases. Cell Mol Life Sci 61:192–208
Costa-E-Sousa RH, Hollenberg AN (2012) Minireview: the neural regu-lation of the hypothalamic-pituitary-thyroid axis. Endocrinology 153:4128–35
Dabbeni-Sala F, Di Santo S, Franceschini D et al. (2001) Melatonin protectsagainst 6-OHDA-induced neurotoxicity in rats: a role for mitochondrialcomplex I activity. FASEB J 15:164–70
Durand G, Poeggeler B, Ortial S et al. (2012) Amphiphilic amide nitrones:a new class of protective agents acting as modifiers of mitochondrialmetabolism. J Med Chem 53:4849–611
Eruslanov E, Kusmartsev S (2010) Identification of ROS using oxidized DCFDAand flow-cytometry. In: Methods Mol Biol 594:57–72
Evans M, Sanders J, Tagami T et al. (2010) Monoclonal autoantibodies to theTSH receptor, one with stimulating activity and one with blocking activity,obtained from the same blood sample. Clin Endocrinol 73:404–12
Feichtinger RG, Neureiter D, Mayr JA et al. (2011) Loss of mitochondria inganglioneuromas. In: Front Biosci 3:179–86
Feliciello A, Gottesman ME, Avvedimento EV (2005) cAMP-PKA signalling tothe mitochondria: protein scaffolds, mRNA and phosphatases. Cell Signal17:279–87
Gaspar E, Hardenbicker C, Bodo E et al. (2010) Thyrotropin releasing hormone(TRH): a new player in human hair-growth control. FASEB J 24:393–403
Gaspar E, Nguyen-Thi KT, Hardenbicker C et al. (2011) Thyrotropin-releasinghormone selectively stimulates human hair follicle pigmentation. J InvestDermatol 131:2368–77
Gemma C, Vila J, Bachstetter A et al. (2007) Oxidative stress and the agingbrain: from theory to prevention. Brain aging Models Methods, andMechanisms, Frontiers in Neuroscience. David R Riddle (ed). Boca Raton:FL, USA. chapter 15
Giesen M, Gruedl S, Holtkoetter O et al. (2011) Ageing processes influencekeratin and KAP expression in human hair follicles. Exp Dermatol20:759–61
Gnaiger E, Kemp RB (1990) Anaerobic metabolism in aerobic mammaliancells: information from the ratio of calorimetric heat flux and respiro-metric oxygen flux. Biochim Biophys Acta 1016:328–32
Hallberg BM, Larsson NG (2011) TFAM forces mtDNA to make a U-turn.Nat Struct Mol Biol 18:1179–81
Halliwell B, Gutteridge JMC (2007) Free Radicals in Biology and Medicine,4th edn. Oxford Univ Press
Harper ME, Seifert EL (2008) Thyroid hormone effects on mitochondrialenergetics. Thyroid 18:145–56
Jastroch M, Divakaruni AS, Mookerjee S et al. (2010) Mitochondrial proton andelectron leaks. Essays Biochem 47:53–67
Jung-Hynes B, Reiter RJ, Ahmad N (2010) Sirtuins, melatonin and circadianrhythms: building a bridge between aging and cancer. J Pineal Res 48:9–19
Kaplan MM, Pan CY, Gordon PR, Lee JK et al. (1988) Human epidermalkeratinocytes in culture convert thyroxine to 3,5,3’-triiodothyronine bytype II iodothyronine deiodination: a novel endocrine function of the skin.J Clin Endocrinol Metab 66:815–22
Kealey T, Williams R, Philpott MP (1994) The human hair follicle engages inglutaminolysis and aerobic glycolysis: implications for skin, splanchnicand neoplastic metabolism. Skin Pharmacol 7:41–6
Kharlip J, Cooper DS (2009) Recent developments in hyperthyroidism. Lancet373:1930–2
Kim KH, Song K, Yoon SH et al. (2012) Rescue of PINK1 protein null-specificmitochondrial complex IV deficits by ginsenoside Re activation of nitricoxide signalling. J Biol Chem 287:44109–20
Kloepper JE, Sugawara K, Al-Nuaimi Y et al. (2010) Methods in hair research:how to objectively distinguish between anagen and catagen in humanhair follicle organ culture. Exp Dermatol 19:305–12
Knuever J, Poeggeler B, Gaspar E et al. (2012) Thyrotropin-releasing hormonecontrols mitochondrial biology in human epidermis. J Clin EndocrinolMetab 97:978–86
Kohnke D, Schramm M, Daut J (1997) Oxidative phosphorylation in myocar-dial mitochondria ’in situ’: a calorimetric study on permeabilized cardiacmuscle preparations. Mol Cell Biochem 174:101–13
Kramer J, Klinger M, Kruse C et al. (2005) Ultrastructural analysis of mouseembryonic stem cell-derived chondrocytes. Anat Embryol 210:175–85
Langan EA, Ramot Y, Hanning A et al. (2010) Thyrotropin-releasing hormoneand oestrogen differentially regulate prolactin and prolactin receptorexpression in female human skin and hair follicles in vitro. Br J Dermatol162:1127–31
Larsson NG, Wang J, Wilhelmsson H et al. (1998) Mitochondrial transcriptionfactor A is necessary for mtDNA maintenance and embryogenesis inmice. Nat Genet 18:231–6
Larsson NG (2010) Somatic mitochondrial DNA mutations in mammalianaging. Annu Rev Biochem 79:683–706
Lerchner J, Wolf A, Buchholz F et al. (2008a) Miniaturized calorimetry - a newmethod for real-time biofilm activity analysis. J Microbiol Methods 74:74–81
Lerchner J, Wolf A, Schneider HJ et al. (2008b) Nano-calorimetry of small-sized biological samples. Thermochim Acta 477:48–53
Limat A, Noser FK (1986) Serial cultivation of single keratinocytes from theouter root sheath of human scalp hair follicles. J Invest Dermatol 87:485–8
Maniura-Weber K, Goffart S, Garstka HL et al. (2004) Transient overexpressionof mitochondrial transcription factor A (TFAM) is sufficient to stimulatemitochondrial DNA transcription, but not sufficient to increase mtDNAcopy number in cultured cells. Nucleic Acid Res 32:6015–27
Marionnet C, Grether-Beck S, Seite S et al. (2011) A broad-spectrum sunscreenprevents UVA radiation-induced gene expression in reconstructed skinin vitro and in human skin in vivo. Exp Dermatol 20:477–82
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
www.jidonline.org 9

Mazzio EA, Soliman KFA (2004) Effects of enhancing mitochondrial oxi-dative phosphorylation with reducing equivalents and ubiquinone on1-methyl-4-phenylpyridinium toxicity and complex I-IV damage inneuroblastoma cells. Biochem Pharmacol 67:1167–84
Messenger AG (2000) Thyroid hormone and hair growth. Br J Dermatol142:633–4
Papa S, Scacco S, Sardanelli AM et al. (2002) Complex I cAMP cascade inhuman physiopathology. Biosci Rep 22:3–16
Paus R (2010) Exploring the "thyroid-skin connection": concepts, questions,and clinical relevance. J Invest Dermatol 130:7–10
Paus R (2011) A neuroendocrinological perspective on human hair folliclepigmentation. Pigment Cell Melanoma Res 24:89–106
Philpott MP, Green MR, Kealey T (1990) Human hair growth in vitro. J Cell Sci97(Pt 3):463–71
Philpott MP, Kealey T (1994) Culture of human philosebaceous units.In: Leigh Irene, Watt Fiona (eds). Keratinocyte Methods. CambridgeUniversity Press: Cambridge, pp, 37–44
Poeggeler B, Knuever J, Gaspar E et al. (2010a) Thyrotropin powers humanmitochondria. FASEB J 24:1525–31
Poeggeler B, Sambamurti K, Siedlak SL et al. (2010b) A novel endogenousindole protects rodent mitochondria and extends rotifer lifespan.PLoS ONE 5:e10206
Portman MA (2008) Thyroid hormone regulation of heart metabolism. Thyroid18:217–25
Ramot Y, Zhang G, Bıro T et al. (2011) TSH is a novel neuroendocrineregulator of selected keratins in the human hair follicle. J Dermatol Sci64:67–70
Rustin P, Chretien D, Bourgeron T et al. (1994) Biochemical and molecularinvestigations in respiratory chain deficiencies. Clin Chim Acta 228:35–51
Safdar A, Little JP, Stokl AJ et al. (2011) Exercise increases mitochondrialPGC-1alpha content and promotes nuclear-mitochondrial cross-talk tocoordinate mitochondrial biogenesis. J Biol Chem 286:10605–17
Safer JD, Persons K, Holick MF (2009) A thyroid hormone deiodinase inhibitorcan decrease cutaneous cell proliferation in vitro. Thyroid 19:181–5
Sanders J, Nunez R, Furmaniak J et al. (2010) TSH receptor mono-clonal antibodies with agonist, antagonist, and reverse agonist activities.Methods Enzymol 485:393–420
Scarpulla RC (2008) Transcriptional paradigms in mammalian mitochondrialbiogenesis and function. Physiol Rev 88:611–38
Scheffler IE (2008) Mitochondria, 2nd edn. Wiley-Liss: Hoboken, NJ, USA
Silva JP, Shabalina IG, Dufour E et al. (2005) SOD2 overexpression: enhancedmitochondrial tolerance but absence of effect on UCP activity. EMBO J24:4061–70
Slominski A, Wortsman J (2000) Neuroendocrinology of the skin. EndocrineRev 21:457–87
Slominski A, Wortsman J, Kohn L et al. (2002) Expression of hypothalamis-pituitary-thyroid axis related genes in the human skin. J Invest Dermatol119:1449–55
Slominski A, Wortsman J, Plonka PM et al. (2005) Hair follicle pigmentation.J Invest Dermatol 124:13–21
Slominski A, Wortsman J, Tuckey RC et al. (2007) Differential expres-sion of HPA axis homolog in the skin. Mol Cell Endocrinol265-266:143–9
Slominski AT, Zmijewski MA, Skobowiat C et al. (2012) Sensing theinviroment: regulation of local and global homeostasis by theskin’s neuroendocrine system. Adv Anat Embryol Cell Biol 212:1–115
Stenn KS, Paus R (2001) Controls of hair follicles cycling. Physiol Rev 81:449–94
Vafaee T, Rokos H, Salem MM et al. (2010) In vivo and in vitro evidence forepidermal H2O2-mediated oxidative stress in piebaldism. Exp Dermatol19:883–7
van Beek N, Bodo E, Kromminga A et al. (2008) Thyroid hormones directlyalter human hair follicle functions: anagen prolongation and stimulationof both hair matrix keratinocyte proliferation and hair pigmentation. J ClinEndocrinol Metab 93:4381–8
Vendelbo MH, Nair KS (2011) Mitochondrial longevity pathways. BiochimBiophys Acta 1813:634–44
Vina J, Gomez-Cabrera MC, Borras C et al. (2009) Mitochondrial biogenesis inexercise and in ageing. Adv Drug Deliv Rev 61:1369–7
Wallace DC, Fan W (2009) The pathophysiology of mitochondrial disease asmodeled in the mouse. Genes Dev 23:1714–36
Wallace DC (2010) Bioenergetics and the epigenome: interface between theenvironment and genes in common diseases. Dev Disabil Res Rev16:114–9
Wallace DC, Fan W, Procaccio V (2010) Mitochondrial eneregetics andtherapeutics. Annu Rev Pathol 5:297–348
Wallace DC (2011) Bioenergetic origins of complexity and disease. ColdSpring Harb Symp Quant Biol 76:1–16
Weitzel JM, Iwen KA, Seitz HJ (2003) Regulation of mitochondrial biogenesisby thyroid hormone. Exp Physiol 88:121–8
Weitzel JM, Iwen KA (2011) Coordination of mitochondrial biogenesis bythyroid hormone. Mol Cell Endocrinol 342:1–7
Williams R, Philpott MP, Kealey T (1993) Metabolism of freshly isolatedhuman hair follicles capable of hair elongation: a glutaminolytic, aerobicglycolytic tissue. J Invest Dermatol 100:834–40
Wood JM, Decker H, Hartmann H et al. (2009) Senile hair graying: H2O2-mediated oxidative stress affects human hair color by blunting methioninesulfoxide repair. FASEB J 23:2065–75
Wrutniak C, Cassar-Malek I, Marchal S et al. (1995) A 43-kDa protein relatedto c-Erb A a1 is located in the mitochondrial matrix of rat liver. J BiolChem 270:16347–54
Zmijewski MA, Slominski AT (2011) Neuroendocrinology of the skin: anoverview and selective analysis. Dermatoendocrinol 3:3–10
Zoeller RT, Tan WS, Tyl RW (2007) General background on the hypothalamic-pituitary-thyroid (HPT) Axis. Crit Rev Toxicol 37:11–52
S Vidali et al.HPT Axis and Mitochondria in Hair Follicles
10 Journal of Investigative Dermatology
Related Documents











