Hypothalamic ependymal-glial cells express the glucose transporter GLUT2, a protein involved in glucose sensing Marı ´a de los Angeles Garcı ´a,* Carola Milla ´n, Carolina Balmaceda-Aguilera, Tamara Castro, Patricia Pastor, Herna ´n Montecinos, Karin Reinicke, Felipe Zu ´n ˜iga,§ Juan Carlos Vera,§ Sergio A. On ˜ateà and Francisco Nualart *Departamento de Biologı ´a Molecular, Laboratorio de Neurobiologı ´a Celular, Departamento de Histologı ´a y Embriologı ´a, §Departamento de Fisiopatologı ´a, Facultad de Ciencias Biolo ´ gicas, Universidad de Concepcio ´n, Chile àDepartment of Cell Biology and Physiology, University of Pittsburgh, Pittsburgh, Pennsylvania, USA Abstract The GLUT2 glucose transporter and the K-ATP-sensitive potassium channels have been implicated as an integral part of the glucose-sensing mechanism in the pancreatic islet b cells. The expression of GLUT2 and K-ATP channels in the hypothalamic region suggest that they are also involved in a sensing mechanism in this area. The hypothalamic glial cells, known as tanycytes a and b, are specialized ependymal cells that bridge the cerebrospinal fluid and the portal blood of the median eminence. We used immunocytochemistry, in situ hybridization and transport analyses to demonstrate the glu- cose transporters expressed in tanycytes. Confocal micros- copy using specific antibodies against GLUT1 and GLUT2 indicated that both transporters are expressed in a and b tanycytes. In addition, primary cultures of mouse hypotha- lamic tanycytes were found to express both GLUT1 and GLUT2 transporters. Transport studies, including 2-deoxy- glucose and fructose uptake in the presence or absence of inhibitors, indicated that these transporters are functional in cultured tanycytes. Finally, our analyses indicated that tany- cytes express the K-ATP channel subunit Kir6.1 in vitro. As the expression of GLUT2 and K-ATP channel is linked to glucose-sensing mechanisms in pancreatic b cells, we pos- tulate that tanycytes may be responsible, at least in part, for a mechanism that allows the hypothalamus to detect changes in glucose concentrations. Keywords: glia, glucose sensing, glucose transporter 2, hypothalamus, K-ATP-sensitive potassium channels, tany- cytes. J. Neurochem. (2003) 86, 709–724. The hypothalamus is thought to be involved in modulating feeding behavior and corporal growth through its ability to detect changes in circulating glucose (Schwartz et al. 2000). The hypothesis that the hypothalamus is able to detect changes in glucose requires the identification of cells involved in this process, as well as the expression and secretion of the key molecules that participate in the hypothalamic glucose-sensing mechanism (Oomura et al. 1969; Levin et al. 2001). The glucose-sensing mechanism by pancreatic b cells involves a number of molecules, including GLUT2, glucokinase, glucagon-like peptide-1 receptors and the ATP-sensitive K + channels (Guillam et al. 1997; Schuit et al. 2001). The finding that ventricular hypothalamic glial cells (tanycytes) express glucose sensor molecules suggests that they may be responsible, at least in part, for glucose sensing by the hypothalamus (Alvarez et al. 1996; Navarro et al. 1996; Thomzig et al. 2001). The hypothalamic peri-ventricular neurons are grouped in nuclei (arcuate nucleus) that are in close contact with highly elongated ependymal cells, namely tanycytes (Flament- Durand and Brion 1985; Chauvet et al. 1995) (Fig. 1). Received March 6, 2003; revised manuscript received April 29, 2003; accepted April 29, 2003. Address correspondence and reprint requests to Dr Francisco Nualart, Laboratorio de Neurobiologı ´a Celular, Departamento de Histologı ´a y Embriologı ´a, Facultad de Ciencias Biolo ´gicas, Universidad de Concep- cio ´ n, casilla 160C, Chile. E-mail: [email protected] Abbreviations used: 2-DOG, 2-deoxy-glucose; bNOS, nitric oxide synthase-brain; CK-HMW, cytokeratin of high molecular weight; GFAP, glial-fibrillary acidic protein; GLUT, glucose transporter; HVWF, von Willebrand factor; LH-RH, luteinizing hormone-releasing hormone; K-ATP, channels, K-ATP-sensitive potassium channels; MBP, myelin basic protein; p75NGFr, neurotrophic growth factor receptor; TTR, transthyretin. Journal of Neurochemistry , 2003, 86, 709–724 doi:10.1046/j.1471-4159.2003.01892.x Ó 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724 709

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hypothalamic ependymal-glial cells express the glucose
transporter GLUT2, a protein involved in glucose sensing
Marıa de los Angeles Garcıa,* Carola Millan,� Carolina Balmaceda-Aguilera,� Tamara Castro,�Patricia Pastor,� Hernan Montecinos,� Karin Reinicke,� Felipe Zuniga,§ Juan Carlos Vera,§
Sergio A. Onate� and Francisco Nualart�
*Departamento de Biologıa Molecular, �Laboratorio de Neurobiologıa Celular, Departamento de Histologıa y Embriologıa,
§Departamento de Fisiopatologıa, Facultad de Ciencias Biologicas, Universidad de Concepcion, Chile
�Department of Cell Biology and Physiology, University of Pittsburgh, Pittsburgh, Pennsylvania, USA
Abstract
The GLUT2 glucose transporter and the K-ATP-sensitive
potassium channels have been implicated as an integral part
of the glucose-sensing mechanism in the pancreatic islet
b cells. The expression of GLUT2 and K-ATP channels in the
hypothalamic region suggest that they are also involved in a
sensing mechanism in this area. The hypothalamic glial cells,
known as tanycytes a and b, are specialized ependymal cells
that bridge the cerebrospinal fluid and the portal blood of the
median eminence. We used immunocytochemistry, in situ
hybridization and transport analyses to demonstrate the glu-
cose transporters expressed in tanycytes. Confocal micros-
copy using specific antibodies against GLUT1 and GLUT2
indicated that both transporters are expressed in a and b
tanycytes. In addition, primary cultures of mouse hypotha-
lamic tanycytes were found to express both GLUT1 and
GLUT2 transporters. Transport studies, including 2-deoxy-
glucose and fructose uptake in the presence or absence of
inhibitors, indicated that these transporters are functional in
cultured tanycytes. Finally, our analyses indicated that tany-
cytes express the K-ATP channel subunit Kir6.1 in vitro. As
the expression of GLUT2 and K-ATP channel is linked to
glucose-sensing mechanisms in pancreatic b cells, we pos-
tulate that tanycytes may be responsible, at least in part, for a
mechanism that allows the hypothalamus to detect changes in
glucose concentrations.
Keywords: glia, glucose sensing, glucose transporter 2,
hypothalamus, K-ATP-sensitive potassium channels, tany-
cytes.
J. Neurochem. (2003) 86, 709–724.
The hypothalamus is thought to be involved in modulating
feeding behavior and corporal growth through its ability to
detect changes in circulating glucose (Schwartz et al. 2000).
The hypothesis that the hypothalamus is able to detect
changes in glucose requires the identification of cells
involved in this process, as well as the expression and
secretion of the key molecules that participate in the
hypothalamic glucose-sensing mechanism (Oomura et al.
1969; Levin et al. 2001). The glucose-sensing mechanism by
pancreatic b cells involves a number of molecules, including
GLUT2, glucokinase, glucagon-like peptide-1 receptors and
the ATP-sensitive K+ channels (Guillam et al. 1997; Schuit
et al. 2001). The finding that ventricular hypothalamic glial
cells (tanycytes) express glucose sensor molecules suggests
that they may be responsible, at least in part, for glucose
sensing by the hypothalamus (Alvarez et al. 1996; Navarro
et al. 1996; Thomzig et al. 2001).
The hypothalamic peri-ventricular neurons are grouped in
nuclei (arcuate nucleus) that are in close contact with highly
elongated ependymal cells, namely tanycytes (Flament-
Durand and Brion 1985; Chauvet et al. 1995) (Fig. 1).
Received March 6, 2003; revised manuscript received April 29, 2003;
accepted April 29, 2003.
Address correspondence and reprint requests to Dr Francisco Nualart,
Laboratorio de Neurobiologıa Celular, Departamento de Histologıa y
Embriologıa, Facultad de Ciencias Biologicas, Universidad de Concep-
cion, casilla 160C, Chile. E-mail: [email protected]
Abbreviations used: 2-DOG, 2-deoxy-glucose; bNOS, nitric oxide
synthase-brain; CK-HMW, cytokeratin of high molecular weight; GFAP,
glial-fibrillary acidic protein; GLUT, glucose transporter; HVWF, von
Willebrand factor; LH-RH, luteinizing hormone-releasing hormone;
K-ATP, channels, K-ATP-sensitive potassium channels; MBP, myelin
basic protein; p75NGFr, neurotrophic growth factor receptor; TTR,
transthyretin.
Journal of Neurochemistry, 2003, 86, 709–724 doi:10.1046/j.1471-4159.2003.01892.x
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724 709

Different studies using the Golgi impregnation method, or
analysis by electron microscopy (Akmayev and Fidelina
1974), identified two main types of tanycytes, b1 and b2.The b1 tanycytes are present in the lateral lower part of the
third ventricle and they are capable of developing cell
processes that contact neurons of the arcuate nucleus as well
as the blood capillary vessels in the hypothalamus (Chauvet
et al. 1995; Peruzzo et al. 2000; Garcıa et al. 2001). The end
feet of the cells processes reach the lateral sulcus of the
infundibular region contacting luteinizing hormone-releasing
hormone (LH-RH) terminals, which are involved in hormone
release to the hypophyseal portal vessels.
The b2 tanycytes are located in the floor of third ventricle
lining the median eminence (Fig. 1). The proximal part of the
cells is in contact with the cerebrospinal fluid of the third
ventricle, while the dorsal part of the cells forms processes in
which the end feet reaches the pial surface of the brain or the
local capillary plexus in the median eminence (Chauvet et al.
1995; Peruzzo et al. 2000; Garcıa et al. 2001). These cells
develop tight-junctions that form the cerebrospinal fluid–
median eminence barrier. The existence of two additional
types of tanycytes, a1 and a2, has also been proposed
(Akmayev and Fidelina 1974) (Fig. 1). These cells line the
dorsal and lateral walls of the third ventricle mainly facing the
ventromedial nucleus of the hypothalamus. The functions of
tanycytes remain a matter of controversy and speculation.
Originally, it was suggested that they might represent a link
between the cerebrospinal fluid and the portal vessels, and that
they could be involved in regulating neuroendocrine function
(Rodrıguez et al. 1985). It has also been suggested that
tanycytes are involved in uptake and transport but the nature of
thematerial that might be transported remains to be discovered
(Flament-Durand and Brion 1985; Garcıa et al. 2001).
Differential gene expression of facilitative glucose trans-
porters (GLUT1–13) mediates the uptake of hexoses in
mammalian cells (Joost et al. 2002). Experiments designed
to identify the precise cellular localization of these transporters
indicate that GLUT1 and GLUT3 are the main isoforms
expressed in brain (Kalaria et al. 1988; Vannucci 1994;Gerhart
et al. 1995; Nualart et al. 1999). GLUT2 is a low-affinity
transporter for glucose and fructose expressed at low levels in
different regions of the brain (Brant et al. 1993). Because of
this low affinity for glucose, but high transport capacity,
GLUT2 is believed to play a major role in glucose-sensing
mechanisms (Guillam et al. 1997, 2000). In situ hybridization
data suggest that GLUT2 is present in hypothalamus (Navarro
et al. 1996). Immunohistochamical analysis indicates GLUT2
expression in astrocytes-like cells (Leloup et al. 1994), how-
ever, GLUT2 has also been detected in ependymal cells
(Ngarmukos et al. 2001).Maekawa et al. (2000) confirmed the
expression of GLUT2 in ependymal cells of the dorsal third
ventricle and cerebral aqueduct, but the hypothalamic ependy-
mal cells (tanycytes) were negative. Thus, the exact localiza-
tion of the specific glial cell types that express GLUT2 in the
hypothalamus remains to be determined.
Here, we report that tanycytes are the main glial cells in
the periventricular zone of the hypothalamus. Expression
analysis of the glucose transporters in mouse hypothalamic
tanycytes revealed that GLUT1 and GLUT2 are expressed in
both a and b tanycytes. The functional properties of GLUT1
and GLUT2 transporters were demonstrated in tanycytes
isolated from mouse hypothalamus. In addition to GLUT1
and GLUT2, we observed that tanycytes in culture express
the pore-forming subunit Kir6.1, an essential molecule in the
formation of K-ATP-sensitive potassium (K-ATP) channels.
Materials and methods
Immunocytochemistry and confocal microscopy
Mice (C57BL/J6) brains were dissected and fixed immediately by
immersion in Bouin’s solution. Fixation in situ was performed by
vascular perfusion (Nualart et al. 1991). Samples were dehydrated in
graded alcohol solutions and embedded in paraffin. Frontal sections
Fig. 1 Schematic representation of mouse hypothalamus. The cells
that are directly identified are: 1, ciliated ependymocytes lining the
rostral wall of the third ventricle, 2, a tanycytes located in the dorsal
lateral wall of the third ventricle, 3, b1 tanycytes located in the lower
lateral wall of the third ventricle, 4, b2 tanycytes located in the median
eminence. The cells form the median eminence–cerebrospinal fluid
barrier (thick line). The projections of b1 and b2 tanycytes contact the
portal blood vessels of the median eminence and pars tuberalis that
are characterized by the absence of a blood–brain barrier. There is
experimental evidence that tanycytes and neurons of the lower lateral
wall of the third ventricle express glucokinase, glucagon-like peptide-1
receptor (GLP-1), and ATP-sensitive K+ channels; molecules involved
in the glucose sensing mechanism. V-, portal blood vessels without
blood–brain barrier.
710 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

(5 lm) of the hypothalamic area were mounted on poly L-lysine-
coated glass slides.
For immunohistochemical analyses, we used a panel of affinity-
purified antibodies raised against synthetic peptides encompassing the
last 10–13 carboxy terminal amino acid of each isoform of the human
facilitative hexose transporters (GLUT1 to GLUT5, Alpha Diagnos-
tic, San Antonio, TX, USA). Sections were incubated, overnight at
room temperature (22�C) in a humid chamber, with anti-GLUT
polyclonal antibodies (1 : 200–1 : 1000) diluted in a Tris-HCl buffer
(pH 7.8) containing 8.4 mM sodium phosphate, 3.5 mM potassium
phosphate, 120 mM NaCl and 1% bovine serum albumin. After
washing extensively, the sections were incubated for 2 h with Cy2-
conjugated affinity-purified donkey anti-rabbit IgG (1 : 200; Jackson
Immuno-Research, West Grove, PA, USA) at room temperature.
Alternatively, anti-mouse IgG (1 : 50; Dako, Carpinteria, CA, USA)
labeled with fluorescein isothiocyanate (1 : 30; Dako) was used as a
secondary antibody. For confocal laser microscopy analysis, the tissue
sections were incubated with propidium iodine in the absence of
RNAase for cellular staining. As negative controls for GLUT1 and
GLUT2, we utilized both primary antibodies pre-absorbed with the
respective peptides used to elicit them, and pre-immune serum. To
characterize glial cell distribution in the hypothalamic area, serial
tissue sections were immunostained using an anti-glial fibrillary acidic
protein (GFAP) polyclonal antibody (1 : 100; Dako).
In situ hybridization
A cDNA of approximately 2.2 kb subcloned in pGEM-4Z
(Clontech, Palo Alto, CA, USA) and encoding the human GLUT2
was used to generate sense and anti-sense digoxigenin-labeled
riboprobes. RNA probes were labeled with digoxigenin-UTP by
in vitro transcription with SP6 or T7 RNA polymerase following the
manufacturer’s procedure (Boehringer Mannheim, Mannheim,
Germany). In situ hybridization was performed on hypothalamic
frontal sections mounted on poly L-lysine-coated glass slides. The
sections were baked at 60�C for 1 h, deparaffinized in xylene, and
rehydrated in graded ethanol. Following proteinase K treatment
(5 min at 37�C in PBS, 1 lg/mL proteinase K), the tissue sections
were fixed with 4% p-formaldehyde for 5 min at 4�C, washed in
cold PBS and then acetylated in 0.1 M triethanol amine-HCl
(pH 8.0) at room temperature for 10 min. After a brief wash, the
sections were incubated in pre-hybridization solution for 15 min at
37�C, and then 25 lL of hybridization mix [50% formamide, 0.6 M
NaCl, 10 mM Tris-HCl (pH 7.5), 1 mM EDTA, 1 · Denhart’s
solution, 10% PEG 8000, 10 mM DTT, 500 lg yeast tRNA/mL,
50 lg/mL heparin, 500 lg/mL DNA carrier, and 1 : 20–1 : 100
diluted riboprobe] were added to each slide. The slides were
covered with glass coverslips and placed in a humidified chamber at
42�C overnight. After removal of the coverslip, the slides were
rinsed in 4 · SSC and washed twice for 30 min at 42�C. The slideswere washed at 37�C for 30 min each in 2 · SSC, 1 · SSC and
0.3 · SSC. Visualization of digoxigenin was performed by incuba-
tion with a monoclonal antibody coupled to alkaline phosphatase
(anti-digoxigenin-alkaline phosphatase Fab fragments diluted
1 : 500; Boehringer Mannheim) for 2 h at room temperature.
Nitroblue tetrazolium chloride and 5-bromo-4-chloro-3-indolyl-
phosphate (Boehringer Mannheim) were used as substrates for the
alkaline phosphatase. Controls included use of the sense riboprobe
and omission of the probe.
Tissue culture
Ependymal cells
Tissue culture experiments used cells obtained from mice at 19 days
of gestation (C57BL/J6) (Gabrion et al. 1988; Chauvet et al. 1996).
The brain was removed, the hypothalamic area taken out, and
further dissected until obtaining a region close to the ependymal
layer. The dissection was done with the samples submerged in
10 mM Hepes (pH 7.3) containing 10 mM glucose, 44 mM sacarose,
135 mM NaCl, 5 mM KCl, 0.15 mM Na2HPO4 (340–350 mOsm/L).
Ventricular walls were incubated with 0.1% (w/v) trypsin for 15 min
at 37�C. Trypsinized tissue was transferred to a 15-mL culture tube
containing 10 mL of minimal essential medium (MEM, Gibco Co.,
Rockville, NY, USA) with 10% (v/v) of fetal bovine serum (FBS) to
stop trypsin action. The tissue was dissociated by trituration through
a siliconized Pasteur pipette, until a single cell suspension was
obtained. Cells were centrifuged for 5 min at 200 g, the supernatant
was aspirated off and the cells were resuspended in MEM and
seeded in culture dishes at a concentration of 0.5–1.0 · 106 cells by
dish. Culture medium was supplemented with 10% FBS, 4 mM
L-glutamine, 100 U/mL penicillin and 100 lg/mL streptomycin
(Nalgene, Rochester, NY, USA). Cells were cultured in the same
dish for 5 weeks and fed every 3 days. The dishes with the highest
concentration of epithelial confluent cells were expanded and used
for the uptake experiments.
Astrocytes
Cerebral hemispheres were removed and the meninges were excised
carefully and discarded. Cells were incubated in 0.1% trypsin for
15 min at 37�C and mechanically dissociated. The trypsinized
tissue was transferred to a 15-mL culture tube containing 10 mL of
MEM with 10% of FBS to stop trypsin action. Cells were
centrifuged for 5 min at 200 g, the supernatant aspirated off and the
cells resuspended in MEM and seeded in culture dishes at a
concentration of 0.5–1.0 · 106 cells/dish. The culture medium was
supplemented with 10% FBS, 2 mM L-glutamine, 100 U/mL
penicillin and 100 lg/mL streptomycin.
For immunocytochemistry, cells were grown on 8-well Laborat-
ory-Tek chamber microscopy slides (Nunc, Neperville, IL, USA),
fixed with 4% p-formaldehyde in PBS for 30 min at 4�C, washedwith PBS and incubated in PBS containing 1% bovine serum
albumin (BSA) and 0.2% Triton X-100 for 5 min at room
temperature. Cells were incubated with the different antibodies
overnight at room temperature: Anti-GLUT1-5 (1 : 250, Alpha
Diagnostic), anti-Kir6.1 (1 : 100, Santa Cruz Biotechnology, Santa
Cruz, CA, USA), anti-GFAP (1 : 200, Dako), anti-vimentin mono-
clonal antibody (1 : 10, Boehringer Mannheim), anti-cytokeratin of
high molecular weight (CK-HMW) monoclonal antibody (1 : 500,
Dako), anti-brain-S100a polyclonal antibody (1 : 400, Dako), anti-
myelin basic protein (MBP) monoclonal antibody (1 : 300, Boeh-
ringer Mannheim), anti-neurotrophic growth factor receptor
(p75NGFr) polyclonal antibody (1 : 5000; Chemicon, Temecula,
CA, USA), anti-transthyretin (TTR) polyclonal antibody (1 : 300,
Dako), anti-Tau monoclonal antibody (5 mg/mL, Boehringer
Mannheim), anti-MAP-2 monoclonal antibody (5 mg/mL, Boehrin-
ger Mannheim), anti-von Willebrand factor (HVWF) polyclonal
antibody (1 : 300, Sigma, St Louis, MO, USA), anti-endothelial cell
antigen (CD31) monoclonal antibody (1 : 50, Dako), anti–blood
GLUT2 expression in hypothalamic glial tanycytes 711
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
brain barrier monoclonal antibody (HT-7, 1 : 200, Sigma), and anti-
nitric oxide synthase-brain (bNOS) monoclonal antibody (1 : 250,
Sigma). Cells were then incubated with fluorescein isothiocyanate
conjugated with goat anti-rabbit IgG or rabbit anti-mouse IgG
(1 : 25, Boehringer Mannheim) for 2 h, mounted, and analyzed by
fluorescence microscopy. Similar analyses were done with cultured
astrocytes.
Immunoblotting
For immunoblot analysis, mouse tanycytes, liver and whole brain cell
membraneproteinswereobtainedbyhomogenizing thecells in0.3 mM
sucrose, 3 mM DTT, 1 mM EDTA, 100 lg/mL PMSF, 1 lg/mL
Peptatin A and 2 lg/mL Aprotinin. Total membranes were collected
by high-speed centrifugation. Membrane protein (30 lg) was loadedin each lane, separated by polyacrylamide gel electrophoresis in the
presence of sodium dodecylsulfate, transferred to nitrocellulose
membranes, and probed with anti-GLUT, anti-Kir6.1 or pre-
absorbed antibodies (1 : 500–1 : 1500) (Zamora-Leon et al.
1996). The secondary antibodies were goat anti-rabbit IgG coupled
to peroxidase (1 : 5000) or rabbit anti-goat IgG coupled to
peroxidase (1 : 5000). The reaction was developed with enhanced
chemiluminiscence using the ECL western blotting analysis system
(Amersham Corporation, Arlington Heights, IL, USA).
Uptake analysis
For uptake assays, cells were grown in 6-well plates to a density of
2 · 105 cells per well. Cultures were carefully selected under the
microscope to ensure that only plates showing uniformly growing
cells were used. In each experiment, cells from six wells incubated
with buffer were removed and used to quantify the number of cells
present in each well. We did not observe a significant variation in
cell numbers between the wells after buffer incubation (Vera et al.
1993; Spielholz et al. 1997; Nualart et al. 2003). Cells were washed
with incubation buffer (15 mM HEPES, 135 mM NaCl, 5 mM KCl,
1.8 mM CaCl2, 0.8 mM MgCl2) and incubated in the same medium
for 30 min at room temperature. Uptake assays were performed in
1 mL of incubation buffer containing 0.2 mM deoxy-glucose and
3 lCi of 2-deoxy-D-[1,2-(N)3H]glucose (30.6 Ci/mmol; DuPont–
NEN, Boston, MA, USA). Uptake was stopped by washing the cells
with ice-cold PBS. Cells were lyzed in 0.5 mL of lysis buffer
(10 mM Tris-HCl, pH 8.0, 0.2% SDS), and the incorporated
radioactivity was assayed by liquid scintillation counting. Fructose
uptake assays were performed in incubation buffer containing 1 mM
fructose and 0.8 lCi of D-[U-14C]fructose/mL (285 mCi/mmol;
Amersham) (Zamora-Leon et al. 1996). Samples were processed as
indicated for deoxyglucose uptake. Where appropriate, competitors
and inhibitors were added to the uptake assays or pre-incubated with
the cells. Data represent means ± SD of three experiments done in
duplicate.
Results
Astrocytes and tanycytes localization in the
periventricular zone of the mouse hypothalamus
Astrocytes are stellated cells with multiple fine processes,
some of which are in close contact to capillary walls. Other
astrocyte processes are wrapped around synaptic contacts
which possess receptors for a variety of neurotransmitters.
In the periventricular area of the brain, the astrocytes are
located in the subventricular zone close to the ependymal
cells. Consistent with this characteristic, we found a normal
stellated astrocytes distribution in the dorsal ventricular wall
of the third ventricle using the classical astrocyte and
propidium iodine staining (Figs 2a and b, arrows). The
ependymal cells and neurons were negative for anti-GFAP
marker, indicating very-low-to-absent expression of GFAP
(Figs 2a and b). However, in the lower lateral walls of the
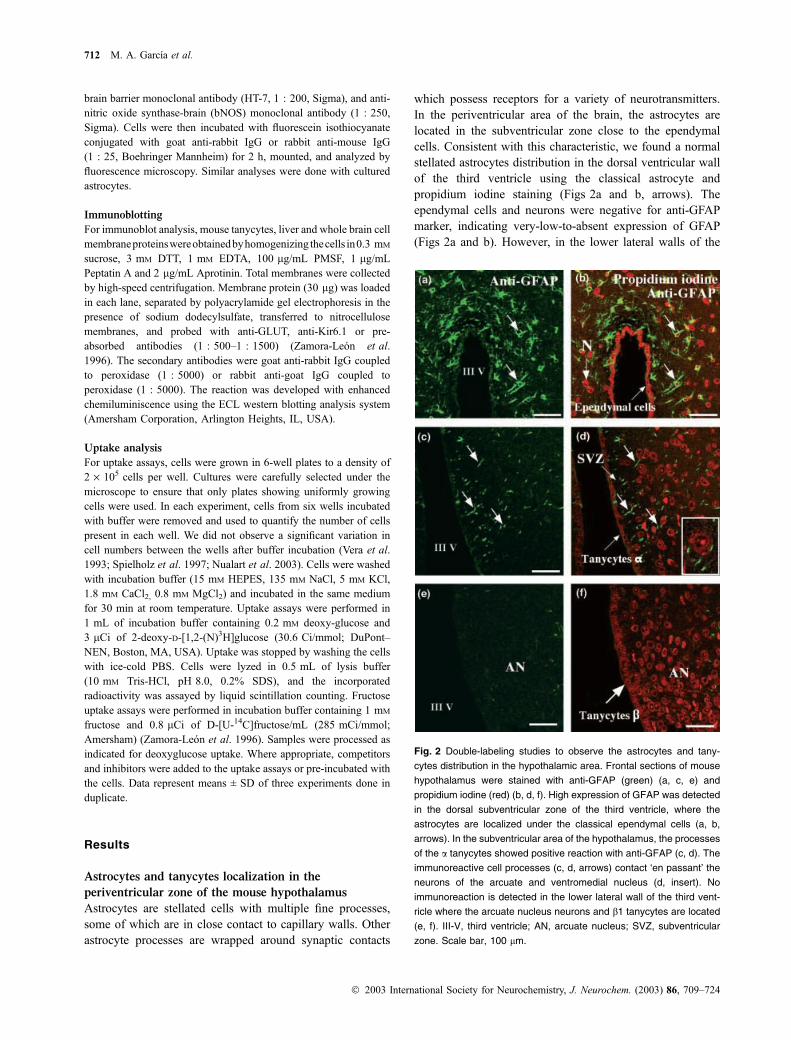
Fig. 2 Double-labeling studies to observe the astrocytes and tany-
cytes distribution in the hypothalamic area. Frontal sections of mouse
hypothalamus were stained with anti-GFAP (green) (a, c, e) and
propidium iodine (red) (b, d, f). High expression of GFAP was detected
in the dorsal subventricular zone of the third ventricle, where the
astrocytes are localized under the classical ependymal cells (a, b,
arrows). In the subventricular area of the hypothalamus, the processes
of the a tanycytes showed positive reaction with anti-GFAP (c, d). The
immunoreactive cell processes (c, d, arrows) contact ‘en passant’ the
neurons of the arcuate and ventromedial nucleus (d, insert). No
immunoreaction is detected in the lower lateral wall of the third vent-
ricle where the arcuate nucleus neurons and b1 tanycytes are located
(e, f). III-V, third ventricle; AN, arcuate nucleus; SVZ, subventricular
zone. Scale bar, 100 lm.
712 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

hypothalamus, stellated and GFAP positive cells were not
detected in the subventricular zone (SVZ) and the reaction
was observed in a tanycytes which presented long immuno-
reactive processes (Figs 2c and d, arrows). These processes
may come into contact, en passant, with the ventromedial
and arcuate nucleus neurons and also the blood vessels
(Fig. 2d, insert and data not shown). In the infundibular walls
of the hypothalamus, where b1 tanycytes contact the neurons
of the arcuate nucleus, we did not find positive reaction with
anti-GFAP (Figs 2e and f), suggesting that astrocytes were
totally replaced by tanycytes in this hypothalamic region.
Our results confirm that tanycytes are the main glial cells in
contact with the endocrine neurons localized in the arcuate
nucleus. This is one of the most important areas responsible
for the generations of signals involved in glucose sensing.
GLUT1 and GLUT2 are expressed in the hypothalamic
ependymal cells
Several studies suggest the expression of the facilitative
glucose transporters, GLUT1 and GLUT2, in ependymal
cells and neurons of the hypothalamic region (Leloup et al.
1994; Maekawa et al. 2000; Ngarmukos et al. 2001). To
confirm the expression of GLUT1 and to establish the precise
localization of GLUT2 in the hypothalamus of adult mice,
we used immunocytochemical detection with anti-GLUTs
polyclonal antibodies and confocal microscopy analysis.
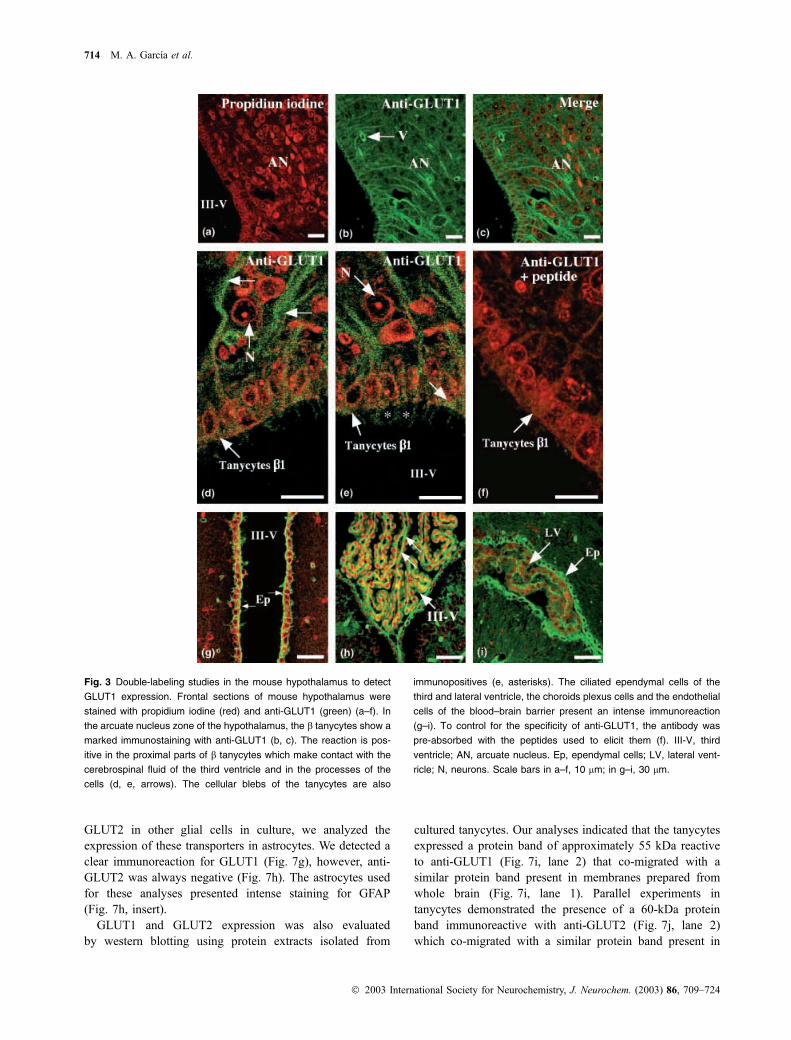
Intense anti-GLUT1 immunoreactivity was observed in the
hypothalamic area, and the immunoreactive material was
associated with endothelial cells, neurophils and tanycytes
(Figs 3a–c). High-magnification analysis showed that
GLUT1 was localized mainly in the processes of the
tanycytes (Fig. 3d, arrows). The apical cytoplasm and the
blebs of the cells were also positives (Fig. 3e, arrow and
asterisks). The reaction was negative when the antibody was
pre-absorbed with the blocking peptide (Fig. 3f). As positive
controls, we observed the intense immunoreaction detected
in the ependymal cells of the third and lateral ventricle
(Figs 3g–i) and in the basolateral membrane of the choroids
plexus cells (Fig. 3h, arrows).
Additionally, we used immunofluorescent and confocal
microscopy to analyze, with higher sensitivity, the expression
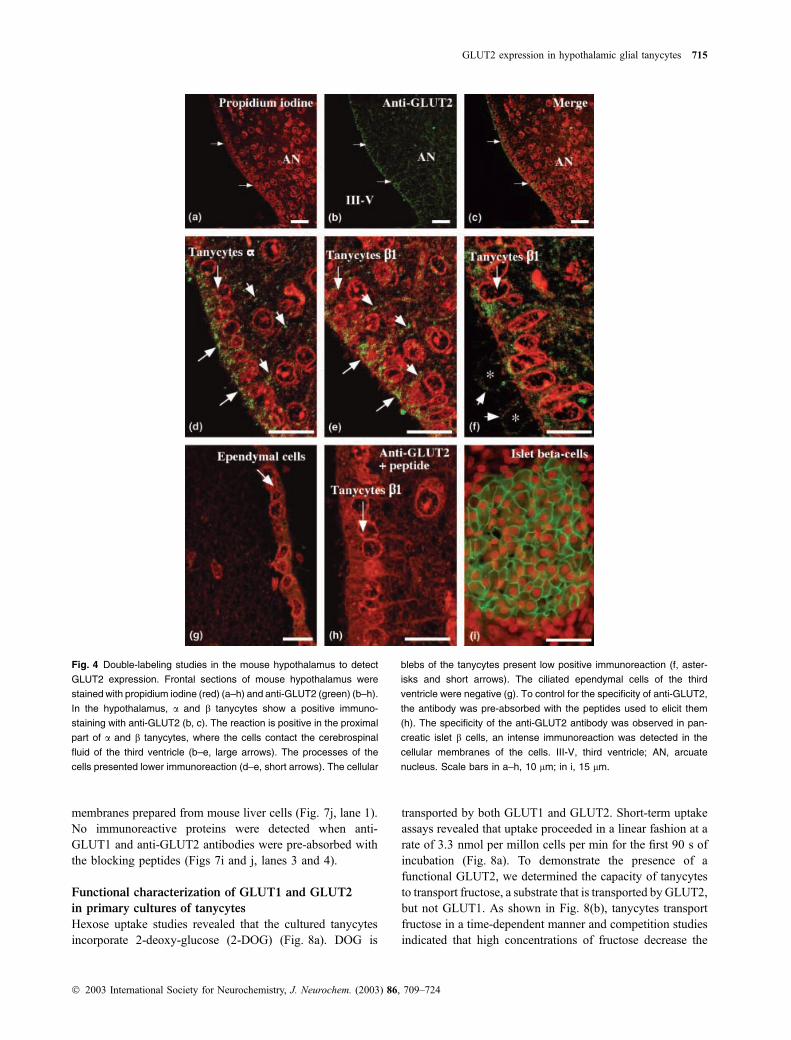
of GLUT2 in the hypothalamic area. Our results showed
GLUT2 staining in a and b tanycytes (Figs 4a–c, arrows)
mainly localized in the apical cellular membranes and the
ventricular cytoplasmic regions of the cells (Figs 4d and e,
large arrows). The cell processes of both a and b tanycytes
presented a weak immunoreactivity (Figs 4d and e, short
arrows). In the lower infundibular region of the third
ventricle the b tanycytes presented a low positive reaction
in the blebs of the cells, similar to the reaction observed with
anti-GLUT1 (Fig. 4f, asterisks and short arrows). No immu-
nostaining was observed in the ependymal cells (Fig. 4g). An
intense immunoreaction was observed in the cellular mem-
brane of the islet b cells (Fig. 4i), demonstrating the high
specificity of the anti-GLUT2 antibody. The reaction was
always negative when we used the blocking peptide to
absorb the antibody (Fig. 4h).
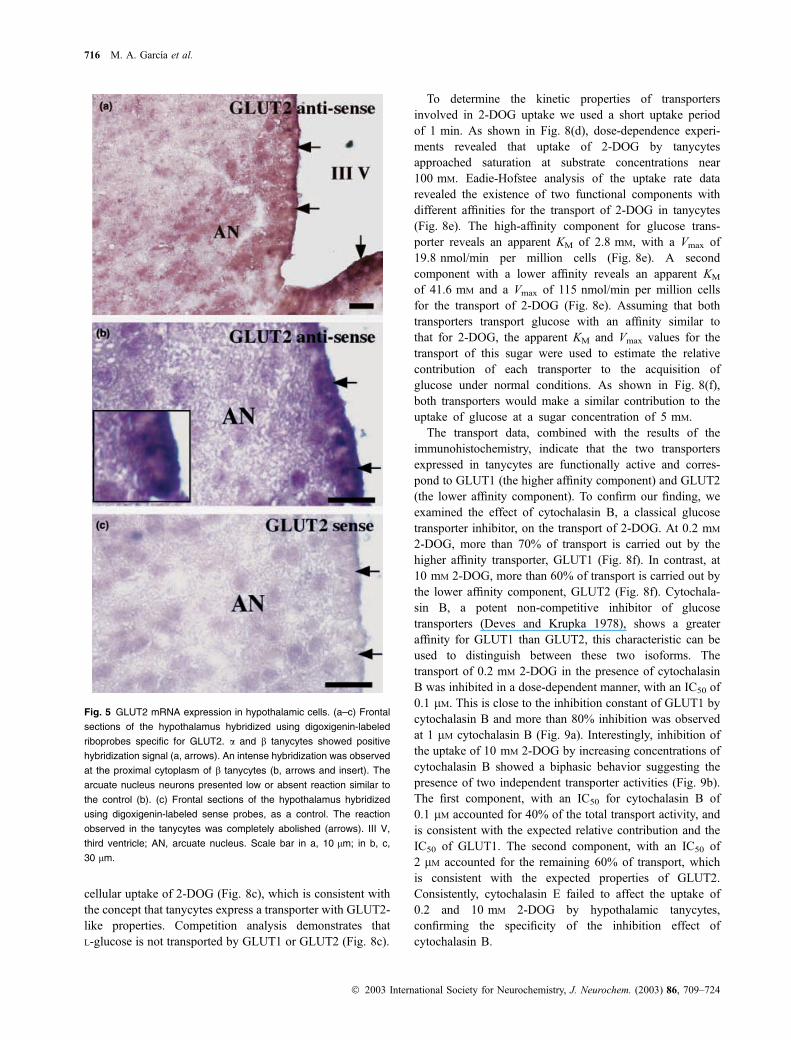
Isotopic in situ hybridization analyses have suggested the
expression of GLUT2 mRNA in hypothalamic neurons and
ependymal cells, but the low resolution of this technique has
prevented a clear identification of the specific cell types
expressing GLUT2. We analyzed the gene expression of
GLUT2 at the mRNA level by in situ hybridization using
digoxigenin-labeled cRNA probes specific for GLUT2. Both
hypothalamic tanycytes, a and b, showed a positive hybrid-
ization signal (Figs 5a and b, arrows) concentrated in the
region facing the third ventricle (Fig. 5b, insert). The neurons
showed a very low reaction similar to the staining observed
with the sense riboprobe (Figs 5b and c), however, the
reaction detected in tanycytes was completely abolished
when the sense probe was used (Fig. 5c, arrows). In
conclusion, these experiments confirmed GLUT1 expression
in hypothalamic tanycytes and demonstrated that these cells
also express the low affinity transporter GLUT2.
GLUT2 is highly expressed in primary tanycyte cultures
We seeded our cultures with cells obtained by thoroughly
dissecting the pre-natal mouse hypothalamic area. After
5 weeks in culture without passage, we selected flasks with
confluent cell growth having an elongated, epithelial aspect
(Fig. 6a). Most cells showed a polarized morphology that
consisted of a wide proximal cytoplasmic region containing
the nucleus and a long basal process (Fig. 6b). Immuno-
histochemical analysis revealed an intense positive reaction
with anti-vimentin and anti-p75 NGFr (Table 1, Figs 6c and
f, and 7a), and a low anti-S100a immunoreaction (Table 1,
Fig. 6e) in cultured tanycytes. Anti-GFAP, anti-CK-HMW
and anti-TTR produced a negative immunoreaction (Table 1,
Fig. 6d). The cells showed negative immunoreactivity for
antibodies against neurons (anti-Tau and anti-MAP2), oligo-
dendroglia (anti-MBP) and endothelial cell markers (anti-
HVWF, anti-BBB, anti-bNOS and anti-endothelial cells)
(Table 1). Astrocytes showed an intense immunostaining
with anti-GFAP (Table 1, Fig. 6g), however, they were
negative for anti-vimentin (Table 1 and Fig. 6h), anti-S100a,
anti-MBP and antip75 NGFr (Table 1).
Antibodies specific for facilitative glucose transporters
revealed expression of GLUT1 and GLUT2 in cultured
tanycytes (Table 1, Figs 7b–f). The anti-GLUT1 immuno-
reactivity showed some heterogeneity (Fig. 7b); in compar-
ison, anti-GLUT2 staining was consistently more intense and
was evenly distributed throughout the cell population
(Figs 7c and d). Moreover, the cell processes showed a
homogeneous and particularly intense reaction with anti-
GLUT2 (Fig. 7e, arrows). To control for the specificity of the
anti-GLUT1 and anti-GLUT2 immunoreactivity, we used
primary antibodies pre-absorbed with the blocking peptides
(Fig. 7f). To control for the expression of GLUT1 and
GLUT2 expression in hypothalamic glial tanycytes 713
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
GLUT2 in other glial cells in culture, we analyzed the
expression of these transporters in astrocytes. We detected a
clear immunoreaction for GLUT1 (Fig. 7g), however, anti-
GLUT2 was always negative (Fig. 7h). The astrocytes used
for these analyses presented intense staining for GFAP
(Fig. 7h, insert).
GLUT1 and GLUT2 expression was also evaluated
by western blotting using protein extracts isolated from
cultured tanycytes. Our analyses indicated that the tanycytes
expressed a protein band of approximately 55 kDa reactive
to anti-GLUT1 (Fig. 7i, lane 2) that co-migrated with a
similar protein band present in membranes prepared from
whole brain (Fig. 7i, lane 1). Parallel experiments in
tanycytes demonstrated the presence of a 60-kDa protein
band immunoreactive with anti-GLUT2 (Fig. 7j, lane 2)
which co-migrated with a similar protein band present in
Fig. 3 Double-labeling studies in the mouse hypothalamus to detect
GLUT1 expression. Frontal sections of mouse hypothalamus were
stained with propidium iodine (red) and anti-GLUT1 (green) (a–f). In
the arcuate nucleus zone of the hypothalamus, the b tanycytes show a
marked immunostaining with anti-GLUT1 (b, c). The reaction is pos-
itive in the proximal parts of b tanycytes which make contact with the
cerebrospinal fluid of the third ventricle and in the processes of the
cells (d, e, arrows). The cellular blebs of the tanycytes are also
immunopositives (e, asterisks). The ciliated ependymal cells of the
third and lateral ventricle, the choroids plexus cells and the endothelial
cells of the blood–brain barrier present an intense immunoreaction
(g–i). To control for the specificity of anti-GLUT1, the antibody was
pre-absorbed with the peptides used to elicit them (f). III-V, third
ventricle; AN, arcuate nucleus. Ep, ependymal cells; LV, lateral vent-
ricle; N, neurons. Scale bars in a–f, 10 lm; in g–i, 30 lm.
714 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
membranes prepared from mouse liver cells (Fig. 7j, lane 1).
No immunoreactive proteins were detected when anti-
GLUT1 and anti-GLUT2 antibodies were pre-absorbed with
the blocking peptides (Figs 7i and j, lanes 3 and 4).
Functional characterization of GLUT1 and GLUT2
in primary cultures of tanycytes
Hexose uptake studies revealed that the cultured tanycytes
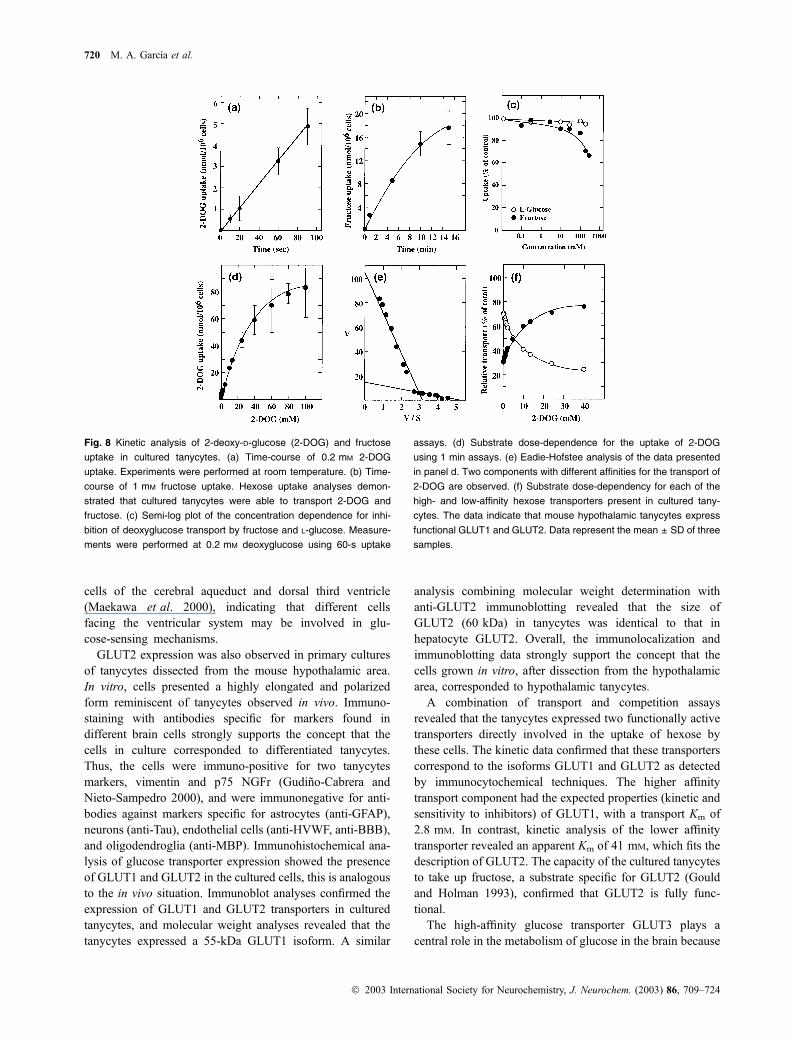
incorporate 2-deoxy-glucose (2-DOG) (Fig. 8a). DOG is
transported by both GLUT1 and GLUT2. Short-term uptake
assays revealed that uptake proceeded in a linear fashion at a
rate of 3.3 nmol per millon cells per min for the first 90 s of
incubation (Fig. 8a). To demonstrate the presence of a
functional GLUT2, we determined the capacity of tanycytes
to transport fructose, a substrate that is transported by GLUT2,
but not GLUT1. As shown in Fig. 8(b), tanycytes transport
fructose in a time-dependent manner and competition studies
indicated that high concentrations of fructose decrease the
Fig. 4 Double-labeling studies in the mouse hypothalamus to detect
GLUT2 expression. Frontal sections of mouse hypothalamus were
stained with propidium iodine (red) (a–h) and anti-GLUT2 (green) (b–h).
In the hypothalamus, a and b tanycytes show a positive immuno-
staining with anti-GLUT2 (b, c). The reaction is positive in the proximal
part of a and b tanycytes, where the cells contact the cerebrospinal
fluid of the third ventricle (b–e, large arrows). The processes of the
cells presented lower immunoreaction (d–e, short arrows). The cellular
blebs of the tanycytes present low positive immunoreaction (f, aster-
isks and short arrows). The ciliated ependymal cells of the third
ventricle were negative (g). To control for the specificity of anti-GLUT2,
the antibody was pre-absorbed with the peptides used to elicit them
(h). The specificity of the anti-GLUT2 antibody was observed in pan-
creatic islet b cells, an intense immunoreaction was detected in the
cellular membranes of the cells. III-V, third ventricle; AN, arcuate
nucleus. Scale bars in a–h, 10 lm; in i, 15 lm.
GLUT2 expression in hypothalamic glial tanycytes 715
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
cellular uptake of 2-DOG (Fig. 8c), which is consistent with
the concept that tanycytes express a transporter with GLUT2-
like properties. Competition analysis demonstrates that
L-glucose is not transported by GLUT1 or GLUT2 (Fig. 8c).
To determine the kinetic properties of transporters
involved in 2-DOG uptake we used a short uptake period
of 1 min. As shown in Fig. 8(d), dose-dependence experi-
ments revealed that uptake of 2-DOG by tanycytes
approached saturation at substrate concentrations near
100 mM. Eadie-Hofstee analysis of the uptake rate data
revealed the existence of two functional components with
different affinities for the transport of 2-DOG in tanycytes
(Fig. 8e). The high-affinity component for glucose trans-
porter reveals an apparent KM of 2.8 mM, with a Vmax of
19.8 nmol/min per million cells (Fig. 8e). A second
component with a lower affinity reveals an apparent KM
of 41.6 mM and a Vmax of 115 nmol/min per million cells
for the transport of 2-DOG (Fig. 8e). Assuming that both
transporters transport glucose with an affinity similar to
that for 2-DOG, the apparent KM and Vmax values for the
transport of this sugar were used to estimate the relative
contribution of each transporter to the acquisition of
glucose under normal conditions. As shown in Fig. 8(f),
both transporters would make a similar contribution to the
uptake of glucose at a sugar concentration of 5 mM.
The transport data, combined with the results of the
immunohistochemistry, indicate that the two transporters
expressed in tanycytes are functionally active and corres-
pond to GLUT1 (the higher affinity component) and GLUT2
(the lower affinity component). To confirm our finding, we
examined the effect of cytochalasin B, a classical glucose
transporter inhibitor, on the transport of 2-DOG. At 0.2 mM
2-DOG, more than 70% of transport is carried out by the
higher affinity transporter, GLUT1 (Fig. 8f). In contrast, at
10 mM 2-DOG, more than 60% of transport is carried out by
the lower affinity component, GLUT2 (Fig. 8f). Cytochala-
sin B, a potent non-competitive inhibitor of glucose
transporters (Deves and Krupka 1978), shows a greater
affinity for GLUT1 than GLUT2, this characteristic can be
used to distinguish between these two isoforms. The
transport of 0.2 mM 2-DOG in the presence of cytochalasin
B was inhibited in a dose-dependent manner, with an IC50 of
0.1 lM. This is close to the inhibition constant of GLUT1 by
cytochalasin B and more than 80% inhibition was observed
at 1 lM cytochalasin B (Fig. 9a). Interestingly, inhibition of
the uptake of 10 mM 2-DOG by increasing concentrations of
cytochalasin B showed a biphasic behavior suggesting the
presence of two independent transporter activities (Fig. 9b).
The first component, with an IC50 for cytochalasin B of
0.1 lM accounted for 40% of the total transport activity, and
is consistent with the expected relative contribution and the
IC50 of GLUT1. The second component, with an IC50 of
2 lM accounted for the remaining 60% of transport, which
is consistent with the expected properties of GLUT2.
Consistently, cytochalasin E failed to affect the uptake of
0.2 and 10 mM 2-DOG by hypothalamic tanycytes,
confirming the specificity of the inhibition effect of
cytochalasin B.
Fig. 5 GLUT2 mRNA expression in hypothalamic cells. (a–c) Frontal
sections of the hypothalamus hybridized using digoxigenin-labeled
riboprobes specific for GLUT2. a and b tanycytes showed positive
hybridization signal (a, arrows). An intense hybridization was observed
at the proximal cytoplasm of b tanycytes (b, arrows and insert). The
arcuate nucleus neurons presented low or absent reaction similar to
the control (b). (c) Frontal sections of the hypothalamus hybridized
using digoxigenin-labeled sense probes, as a control. The reaction
observed in the tanycytes was completely abolished (arrows). III V,
third ventricle; AN, arcuate nucleus. Scale bar in a, 10 lm; in b, c,
30 lm.
716 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

Tanycytes in culture express the K-ATP channels
pore-forming subunit Kir6.1
Another component of the glucose-sensing mechanism is
the K-ATP channel. There is immunohistochemical data
showing the expression of the K-ATP channel pore-
forming subunit Kir6.1 in hypothalamic tanycytes in situ
(Thomzig et al. 2001). As the expression of this channel is
crucial to argument that tanycytes are potentially involved
in glucose sensing, we studied the expression of Kir6.1
subunit in cultured tanycytes using immunofluoresce and
immunoblot analysis.
Immunocytochemical analysis using specific antibodies
for Kir6.1 subunit indicates that Kir6.1 subunit is
expressed in tanycyte primary cultures (Table 1, Fig. 10a).
Positive immunoreactivity is detected in the cytoplasm and
processes of the tanycytes. Propidium iodine staining after
RNAase treatment confirms that most of the cells in
culture expressed Kir6.1 immunoreactivity (Fig. 10b).
Fig. 6 Immunocytochemistry analyses of cultured tanycytes and ast-
rocytes. The cells were obtained from mouse hypothalamus at
19 days of gestation. (a) Tanycytes after 5 weeks in culture. The cells
are organized in monolayers and show an elongated form. (b) High
magnification of a single cell using Nomarsky optics. The cell shows a
polarized aspect with an apical expanded area and a long single
process (labeled P). The position of the nucleus is also indicated
(labeled N). (c–f) Tanycytes immunostained with anti-vimentin (c),
anti-GFAP (d), anti-brain S100a (e) and anti-p75 NGFr (f). The tany-
cytes were anti-GFAP negative, however, the cells presented a strong
immunoreaction with anti-vimentin and anti-p75 NGFr, both markers
for tanycytes. (g, h) Astrocytes at 2 weeks in culture immunostained
with anti-GFAP (g) and anti-vimentin (h). Insert in h, Hoechst staining
indicating the cells present in the observed field. Scale bars in a,
100 lm; in b, 5 lm. Scale bars in c–h, 30 lm.
GLUT2 expression in hypothalamic glial tanycytes 717
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
Anti-Kir6.1 primary antibodies pre-absorbed with the
blocking peptide indicate that the immunoreaction was
specific (data not shown). Western immunoblotting analy-
sis using anti-Kir6.1 antibodies revealed a 47-kDa protein
band (Fig. 10d, lane 3). A similar protein band was found
in extract prepared from hypothalamus and liver (Fig. 10,
lanes 1 and 2).
Discussion
Initial biochemical studies indicate that the relative expres-
sion of GLUT2 in the brain is low (Brant et al. 1993; Leloup
et al. 1994). However, low relative levels of GLUT2 may be
due to restricted expression in a few cell types. Our detailed
immunohistochemical and functional analysis of the low
affinity glucose transporter, both in situ and in vitro,
consistently indicates that GLUT2 is primarily expressed in
a and b tanycytes of the mouse hypothalamus. These
experiments were performed using an anti-GLUT2 antibody
that gave a positive immunoblot reaction in membranes
isolated from mouse hepatocytes expressing high levels of
GLUT2 and in pancreatic b cells in situ. Interestingly, the
preferential GLUT2 expression observed in hypothalamic aand b tanycytes was localized in the proximal part of the cell
body, which corresponds to the region of the cell that is in
contact with the cerebrospinal fluid. Other cells of the
hypothalamus, such as neurons and endothelial cells, were
consistently negative for GLUT2 in our immunohistochem-
ical analysis.
Immunohistochemical data have indicated GLUT2 local-
ization in numerous punctate structures localized in the
hypothalamic area between the arcuate nucleus and ventro-
medial hypothalamus (Leloup et al. 1994). Further, immu-
nodetection using electron-microscopy suggests GLUT2
localization in astrocytes processes in close relationship with
nerve terminals or neuronal cell bodies (Leloup et al. 1994).
However, the antibodies used in this study were unable to
detect GLUT2 proteins by western blot analysis of samples
isolated from the hypothalamic arcuate nucleus. Ngarmukos
et al. (2001) were unable to detect GLUT2 expression in
ventromedial hypothalamic cells; however, GLUT2 immu-
noreactivity was detected in the ependymal cells of the dorsal
third ventricle and in scattered cells in the arcuate and
periventricular nuclei. Further, Maekawa et al. (2000) detec-
ted a clear expression of GLUT2 in ependymal cells of the
dorsal third ventricle and cerebral aqueduct, but the
hypothalamic glial cells were negative. In detail, GLUT2
was localized in ciliated ependymal cells of the cerebral
aqueduct, specifically in the cell membrane of the cilia
(Maekawa et al. 2000). Thus, the localized expression of
GLUT2 in a few ventricular ependymal cells may explain the
low relative expression of this transporter when the analysis
includes the whole brain. In situ hybridization analyses have
revealed GLUT2 expression in a region of the hypothalamus
containing neurons and ependymal cells (Navarro et al.
1996). However, the lower resolution capacity of autoradi-
ographic detection of the GLUT2 probe prevented the
identification of the specific cell types expressing GLUT2.
Our in situ non-isotopic hybridization clearly identifies the
a- and b-tanycytes as the hypothalamic cells expressing
GLUT2. We observed a reduced hybridization in neuronal
soma, however, the negative controls using the sense probe
showed a similar reaction, indicating low-to-absent expres-
sion of GLUT2 mRNA in arcuate nucleus neurons and
astrocytes-like cells.
The expression of GLUT2 in a- and b-tanycytes suggest
that tanycytes contacting ventromedial hypothalamic neu-
rons (a) and arcuate nucleus neurons (b) are involved in
glucose uptake using the same low-affinity transporter. In
both types of cells, the localization of GLUT2 is observed
in the proximal pole of the tanycyte which contains the
cerebrospinal fluid, and thus tanycytes may be primarily
involved in detecting glucose concentration in the cere-
brospinal fluid of the ventricular system. Similar expres-
sion of GLUT2 transporter has been detected in ependymal
Table 1 Comparative immunohistochemical analysis of cultured
tanycytes and astrocytes
Tanycytes Astrocytes
Antibodies Reaction
Glial and ependymal cell markers
Anti-vimentin +++ –
Anti-b-S100a + –
Anti-GFAP – +++
Anti-CK-HMW – nd
Anti-TTR – nd
Anti-MBP – –
Anti-p75 NGFr +++ –
Neuronal markers
Anti-Tau – nd
Anti-MAP-2 – nd
Endothelial cell markers
Anti-HVWF – nd
Anti-endothelial cells – nd
Anti-BBB – nd
Anti-bNOS – nd
Glucose transporters markers
Anti-GLUT1 +++ +++
Anti-GLUT2 +++ –
Anti-GLUT3 – –
Anti-GLUT4 – –
Anti-GLUT5 – –
K-ATP channels subunit
Anti-Kir6.1 +++ nd
nd, not determined; –, negative; +, weak reaction; +++, strong reac-
tion.
718 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

Fig. 7 Cultured tanycytes express GLUT1 and GLUT2. Immunocyto-
chemical and immunoblot analysis. (a–f) Tanycyctes at 5 weeks in
culture analyzed with different antibodies. (a) Anti-vimentin. (b) Anti-
GLUT1. (c) and (e, f) anti-GLUT2. GLUT2 is detected in the proximal
part of the tanycytes (c) and in the long processes of the cells (e). (d)
Hoechst staining indicating the total cells present in the observed field.
The numbers 1–4 represent the same cells shown in c with anti-
GLUT2. (f) Control of the immunostaining using anti-GLUT2 pre-
absorbed with the peptide. (g, h) Cultured astrocytes analysed with
anti-GLUT1 (g) and anti-GLUT2 (h). The astrocytes were positives to
anti-GFAP (h, insert). (i) Immunoblotting of total cellular membranes of
mouse brain (lane 1) and tanycytes (lane 2) incubated with anti-
GLUT1. The tanycytes showed a 55-kDa band (lane 2) similar to the
band located in cellular membranes isolated from whole mouse brains
(lane 1). (j) Immunoblotting of total cellular membranes of mouse liver
(1) and tanycytes (2) incubated with anti-GLUT2. A 60-kDa band that
corresponds to GLUT2 was detected in both samples. A total of 30 lg
of membrane protein was loaded in each lane. Controls were per-
formed incubating the membranes with antibodies pre-absorbed with
the peptides (i, j, lanes 3–4). Scale bars in a–h, 30 lm.
GLUT2 expression in hypothalamic glial tanycytes 719
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
cells of the cerebral aqueduct and dorsal third ventricle
(Maekawa et al. 2000), indicating that different cells
facing the ventricular system may be involved in glu-
cose-sensing mechanisms.
GLUT2 expression was also observed in primary cultures
of tanycytes dissected from the mouse hypothalamic area.
In vitro, cells presented a highly elongated and polarized
form reminiscent of tanycytes observed in vivo. Immuno-
staining with antibodies specific for markers found in
different brain cells strongly supports the concept that the
cells in culture corresponded to differentiated tanycytes.
Thus, the cells were immuno-positive for two tanycytes
markers, vimentin and p75 NGFr (Gudino-Cabrera and
Nieto-Sampedro 2000), and were immunonegative for anti-
bodies against markers specific for astrocytes (anti-GFAP),
neurons (anti-Tau), endothelial cells (anti-HVWF, anti-BBB),
and oligodendroglia (anti-MBP). Immunohistochemical ana-
lysis of glucose transporter expression showed the presence
of GLUT1 and GLUT2 in the cultured cells, this is analogous
to the in vivo situation. Immunoblot analyses confirmed the
expression of GLUT1 and GLUT2 transporters in cultured
tanycytes, and molecular weight analyses revealed that the
tanycytes expressed a 55-kDa GLUT1 isoform. A similar
analysis combining molecular weight determination with
anti-GLUT2 immunoblotting revealed that the size of
GLUT2 (60 kDa) in tanycytes was identical to that in
hepatocyte GLUT2. Overall, the immunolocalization and
immunoblotting data strongly support the concept that the
cells grown in vitro, after dissection from the hypothalamic
area, corresponded to hypothalamic tanycytes.
A combination of transport and competition assays
revealed that the tanycytes expressed two functionally active
transporters directly involved in the uptake of hexose by
these cells. The kinetic data confirmed that these transporters
correspond to the isoforms GLUT1 and GLUT2 as detected
by immunocytochemical techniques. The higher affinity
transport component had the expected properties (kinetic and
sensitivity to inhibitors) of GLUT1, with a transport Km of
2.8 mM. In contrast, kinetic analysis of the lower affinity
transporter revealed an apparent Km of 41 mM, which fits the
description of GLUT2. The capacity of the cultured tanycytes
to take up fructose, a substrate specific for GLUT2 (Gould
and Holman 1993), confirmed that GLUT2 is fully func-
tional.
The high-affinity glucose transporter GLUT3 plays a
central role in the metabolism of glucose in the brain because
Fig. 8 Kinetic analysis of 2-deoxy-D-glucose (2-DOG) and fructose
uptake in cultured tanycytes. (a) Time-course of 0.2 mM 2-DOG
uptake. Experiments were performed at room temperature. (b) Time-
course of 1 mM fructose uptake. Hexose uptake analyses demon-
strated that cultured tanycytes were able to transport 2-DOG and
fructose. (c) Semi-log plot of the concentration dependence for inhi-
bition of deoxyglucose transport by fructose and L-glucose. Measure-
ments were performed at 0.2 mM deoxyglucose using 60-s uptake
assays. (d) Substrate dose-dependence for the uptake of 2-DOG
using 1 min assays. (e) Eadie-Hofstee analysis of the data presented
in panel d. Two components with different affinities for the transport of
2-DOG are observed. (f) Substrate dose-dependency for each of the
high- and low-affinity hexose transporters present in cultured tany-
cytes. The data indicate that mouse hypothalamic tanycytes express
functional GLUT1 and GLUT2. Data represent the mean ± SD of three
samples.
720 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

it is the main glucose transporter expressed by neurons
(Vannucci 1994). The results of our functional studies
indicated absence of expression of a GLUT3-like high
affinity glucose transporter (Km < 1 mM) in the cultured
tanycytes. We confirmed the absence of expression of
GLUT3 in the tanycytes in vitro and in situ by immunocyto-
chemistry and immunobloting with anti-GLUT3 antibodies
(Table 1 and data not shown). Thus, the data indicate that
GLUT3 plays no role in glucose uptake by the tanycytes.
The expression of GLUT2 in hypothalamic tanycytes may
have important physiological consequences. An elevated Km
for glucose transport implies that tanycytes expressing
GLUT2 will increase their glucose uptake rate in direct
proportion to extracellular changes in glucose concentration.
This property of GLUT2 determines its participation in the
glucose-sensing mechanism of the pancreatic b cell (Guillam
et al. 1997; Yang et al. 1999; Guillam et al. 2000; Schuit
et al. 2001). Therefore, it is possible that GLUT2 might play
a similar role in the brain (Wan et al. 1998), which opens up
the possibility for the involvement of tanycytes in a glucose-
sensing mechanism in the hypothalamus. In this context,
there is data indicating that proteins of the type involved in
the b cell glucose-sensing mechanism are expressed in the
hypothalamus (Alvarez et al. 1996; Navarro et al. 1996;
Miki et al. 2001) (Fig. 1).
In order to obtain more evidence to reinforce our theory
that tanycytes are involved in glucose sensing, we analyzed
the expression of ATP-sensitive potassium channels in
cultured tanycytes. These channels are essential to couple
the energy state of a cell to its excitability, participating in
glucose-sensing mechanism in pancreatic islet b cells (Schuit
et al. 2001). Recently, the presence of functional K-ATP
channels in glial cells has been suggested. This study
detected the presence of the Kir6.1 subunit, the main pore-
forming protein detected in hippocampal, cortical and
cerebellar astrocytes, tanycytes and Bergmann glial cells
(Thomzig et al. 2001). We confirmed the expression of the
K-ATP channel subunit Kir6.1 in cultured tanycytes, indica-
ting that these cells maintain in vitro some of the molecular
and functional properties observed in situ.
In addition to GLUT2 and K-ATP channels, cells
involved in glucose sensing express a high Km glucokin-
ase, an enzyme that shows low affinity for glucose and is
not inhibited by glucose-6-phosphate. Glucokinase is
expressed in the hypothalamus, specifically in ependymal
cells (tanycytes) and some neurons (Roncero et al. 2000).
The high Km glucokinase has also been detected in the
lower brain stem (Lynch et al. 2000; Maekawa et al.
2000). Other areas within the brain, specifically the area
post rema, the medulla oblongata and the tractus solitarius
nucleus have also been postulated to have glucose sensor
mechanisms that modulate feeding and reproduction
(Leloup et al. 1994; Schwartz et al. 2000). Additionally,
a sensor mechanism triggered by low glucose concentra-
tions in the hind brain controls the secretion of GnRH that
inhibits the pulsatile LH secretion in rats (Murahashi et al.
1996). It might be possible that the effects associated with
a glucose-sensing mechanism in the lower brain stem may
be functionally co-ordinated with a hypothalamic glucose-
sensing mechanism (Grill and Kaplan 2002). GnRH is
secreted in the basal and lateral part of the median
eminence, an area in which the axonal terminals of the
neurons that release GnRH are in contact with the end feet
of the tanycyte processes (Meister et al. 1988). The end
feet of the tanycyte processes wrap around and support the
portal blood vessels of the median eminence, thereby
blocking the nerve endings, containing hypothalamic
factors, from reaching the peri-capillary spaces in the
Fig. 9 Effect of cytochalasin B on 2-deoxy-D-glucose uptake by cul-
tured tanycytes. (a) Dose-dependent effect of cytochalasin B and cy-
tochalasin E using a concentration of 0.2 mM 2-DOG to test the higher
affinity transporter. (b) Dose-dependent effect of cytochalasin B and
cytochalasin E using a concentration of 10 mM 2-DOG to test the lower
affinity transporter. Data represent the mean ± SD of three samples.
DOG, deoxyglucose; Cyt, cytochalasin.
GLUT2 expression in hypothalamic glial tanycytes 721
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

median eminence (Kozlowski and Coates 1985; Hokfelt
et al. 1988). It has been proposed that dopamine regulates
the contact between the tanycyte ending and the blood
vessels (Bergland and Page 1979; Hokfelt et al. 1988).
When dopamine is released, it induces the retraction of the
tanycyte endings, allowing the access of the neuropeptide-
containing nerve terminal to the portal blood vessels. This
property of tanycytes is regulated by dopamine, but its
function may also be induced by intracellular signals
generated directly in the tanycytes. Tanycytes are in
simultaneous contact with the cerebrospinal fluid and the
blood vessels, and it is reasonable to think that they may
be able to detect changes in glucose concentration
occurring in both fluids. Thus, variations in glucose
concentration may induce the first changes in tanycytes,
and dopamine (whose release may be regulated by axons
afferent from the lower brain stem) could modulate the
structural change. These various systems provide a selec-
tion of routes by which glucose may reach the brain and
affect phenomena as diverse as sleep, orgasm, reproduction
cycle and feeding behavior (Bergland and Page 1979).
The level of GLUT2 expression in tanycytes may be
similar to the expression of GLUT3 and GLUT1 in
neurons and astrocytes. Thus, these transporters need high-
sensitivity immunohistochemical methods (Leloup et al.
1994). Additionally, an up-regulation in the expression of
GLUT1 and GLUT3 has been observed in cultured cells
(Gould and Holdman 1993). A similar up-regulation of
GLUT2 occurred in cultured tanycytes, allowing us to
detect the transporter with conventional immunofluores-
cence. We did not detect changes in GLUT2 expression in
cells after prolonged culture, suggesting that the regulation
of GLUT2 expression in tanycytes is different to that in
hepatocytes, where cells lost the expression of GLUT2 in
short time cultures with a simultaneous induction of
GLUT1 expression (Zheng et al. 1995).
Acknowledgements
The authors thank Ximena Campos, Universidad de Concepcion for
her technical support and Dr Simon Watkins, Department of Cell
Biology and Physiology, University of Pittsburgh, for his support in
confocal microscopy. We also thank Dr Rodolfo Medina for the
critical reading of the manuscript. This work was in part supported
by grants 1010843 from FONDECYT, Chile, DIUC-GIA
201.034.006–1.4, DIUC 202.031.089–1 and DIUC 201.035.002–
1.0, Universidad de Concepcion, Chile.
Fig. 10 Tanycytes in vitro express the K-ATP channel subunit Kir6.1.
The cells were obtained from mouse hypothalamus at 19 days of
gestation and used to perform immunocytochemical (a, b) or immu-
noblot (c) analysis. (a, b) Tanycytes after 5 weeks in culture stained
with anti-Kir6.1 subunit antibody (a) and propidium iodine (b). The
merge image shows that most of the cells in culture present Kir6.1
immunoreaction (b). (c) Immunoblotting of total cellular membranes of
liver (lane 1), hypothalamus (lane 2) and cultured tanycytes (lane 3)
incubated with anti-Kir6.1. The tanycytes show a 47-kDa band similar
to the band located in cellular membranes isolated from liver and
hypothalamus. Scale bar in c, 15 lm.
722 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

References
Akmayev I. G. and Fidelina O. V. (1974) Morphological aspects of the
hypothalamic-hypophyseal system. V. The tanycytes: their relation
to the hypophyseal adrenocorticotrophic function. An enzyme-
histochemical study. Cell Tissue Res. 152, 403–410.
Alvarez E., Roncero I., Chowen J. A., Thorens B. and Blazquez E.
(1996) Expression of the glucagon-like peptide-1 receptor gene in
rat brain. J. Neurochem. 66, 920–927.
Bergland R. M. and Page R. B. (1979) Pituitary-brain vascular relations:
a new paradigm. Science 204, 18–24.
Brant A. M., Jess T. J., Milligan G., Brown C. M. and Gould G. W.
(1993) Immunological analysis of glucose transporters expressed
in different regions of the rat brain and central nervous system.
Biochem. Biophys. Res. Commun. 192, 1297–1302.
Chauvet N., Parmentier M. L. and Alonso G. (1995) Transected axons of
adult hypothalamo-neurohypophysial neurons regenerate along
tanycytic processes. J. Neurosci. Res. 41, 129–144.
Chauvet N., Privat A. and Alonso G. (1996) Aged median eminence
glial cell cultures promote survival and neurite outgrowth of
cocultured neurons. Glia 18, 211–223.
Deves R. and Krupka R. M. (1978) Cytochalasin B and the kinetics of
inhibition of biological transport: a case of asymmetric binding to
the glucose carrier. Biochim. Biophys. Acta 510, 339–348.
Flament-Durand J. and Brion J. (1985) Tanycytes: morphology and
functions: a review. Int. Rev. Cytol. 96, 121–155.
Gabrion J., Peraldi S., Faivre-Bauman A., Klotz C., Ghandour M. S.,
Paulin D., Assenmacher I. and Tixier-Vidal A. (1988) Characteri-
zation of ependymal cells in hypothalamic and choroidal primary
cultures. Neuroscience 24, 993–1007.
Garcıa M. A., Carrasco M., Godoy A., Reinicke K., Montecinos V. P.,
Aguayo L. G., Tapia J. C., Vera J. C. and Nualart F. (2001)
Elevated expression of glucose transporter-1 in hypothalamic
ependymal cells not involved in the formation of the brain–
cerebrospinal fluid barrier. J. Cell Biochem. 80, 491–503.
Gerhart D. Z., Leino R. L., Borson N. D., Taylor W. E., Gronlund K. M.,
McCall A. L. and Drewes L. R. (1995) Localization of glucose
transporter GLUT3 in brain: comparison of rodent and dog using
species-specific carboxy-terminal antisera. Neuroscience 66,
237–246.
Gould G. W. and Holdman G. D. (1993) The glucose transporter family:
structures, function and tissue-specific expression. Biochem. J. 295,
329–341.
Grill H. and Kaplan J. (2002) The neuroanatomical axis for control of
energy balance. Front. Neuroendocrinol. 23, 2–40.
Gudino-Cabrera G. and Nieto-Sampedro M. (2000) Schwann-like
macroglia in adult rat brain. Glia 30, 49–63.
Guillam M. T., Hummler E., Schaerer E., Yeh J. Y., Birnbaum M. J.,
Beermann F., Schmidt A., Deriaz N. and Thorens B. (1997) Early
diabetes and abnormal pancreatic islet development in mice lack-
ing GLUT2. Nat. Genet. 17, 327–330.
Guillam M. T., Dupraz P. and Thorens B. (2000) Glucose uptake,
utilization, and signaling in Glut2-Null islets. Diabetes 49,
1485–1491.
Hokfelt T., Foster G., SchultzbergM., Meister B., SchallingM., Goldstein
M., Hemmings H. C. Jr, Ouimet C. and Greengard P. (1988)
DARPP-32 as a marker for D-1 dopaminoceptive cells in the rat
brain: prenatal development and presence in glial elements (tany-
cytes) in the basal hypothalamus. Adv. Exp. Med. Biol. 235, 65–82.
Joost H. G., Bell G. I., Best J. D., Birnbaum M. J., Charron M. J., Chen
Y. T., Doege H., James D. E., Lodish H. F., Moley J. F., Mueckler
M., Rogers S., Schurmann A., Seino S. and Thorens B. (2002)
Nomenclature of the GLUT/SLC2A family of sugar/polyol trans-
port facilitators. Am. J. Physiol. Endocrinol. Metab. 282, 974–976.
Kalaria R. N., Gravina S. A., Schmidley J. W., Perry G. and Harik S. I.
(1988) The glucose transporter of the human brain and blood–brain
barrier. Ann. Neurol. 24, 757–764.
Kozlowski G. P. and Coates P. W. (1985) Ependymoneuronal speciali-
zations between LHRH fibers and cells of the cerebroventricular
system. Cell Tissue Res. 242, 301–311.
Leloup C., Arluison M., Lepetit N., Cartier N., Marfaing-Jallat P., Ferre
P. and Penicaud L. (1994) Glucose transporter 2 (GLUT2):
expression in specific brain nuclei. Brain Res. 638, 221–226.
Levin B. E., Dunn-Meynell A. A. and Routh V. H. (2001) Brain
glucosensing and the KATP channel. Nat. Neurosci. 4, 459–460.
Lynch R. M., Tompkins L. S., Brooks H. L., Dunn-Meynell A. A. and
Levin B. E. (2000) Localization of glucokinase gene expression in
the rat brain. Diabetes 49, 693–700.
Maekawa F., Toyoda Y., Torii N., Miwa I., Thompson R. C., Foster D.,
Tsukahara S., Tsukamura H. and Maeda K. (2000) Localization of
glucokinase-like immunoreactivity in the rat lower brain stem: for
possible location of brain glucose-sensing mechanisms. Endo-
crinology 141, 375–384.
Meister B., Hokfelt T., Tsuruo Y., Hemmings H., Ouimet C., Greengard
P. and Glodstein M. (1988) DARPP-32, a dopamine-and cyclic
AMP-regulated phosphoprotein in tanycytes of the mediobasal
hypothalamus: distribution and relation to dopamine and lutein-
izing hormone-releasing hormone neurons and other glial elements.
Neuroscience 27, 607–622.
Miki T., Liss B., Minami K., Shiuchi T., Saraya A., Kashima Y., Hor-
iuchi M., Ashcroft F., Minokoshi Y., Roeper J. and Seino S. (2001)
ATP-sensitive K+ channels in the hypothalamus are essential for
the maintenance of glucose homeostasis. Nat. Neurosci. 4, 507–
512.
Murahashi K., Bucholtz D. C., Nagatani S., Tsukahara S., Tsukamura H.,
Foster D. L. and Maeda K. I. (1996) Supression of luteinizing
hormone pulses by restriction of glucose availability is mediated by
sensors in the brain stem. Endocrinology 137, 1171–1176.
Navarro M., Rodrıguez de Fonseca F., Alvarez E., Chowen J. A., Zueco
J. A., Gomez R., Eng J. and Blazquez E. (1996) Colocalization of
glucagon-like peptide-1 (GLP-1) receptors, glucose transporter
GLUT-2, and glucokinase mRNAs in rat hypothalamic cells: evi-
dence for a role of GLP-1 receptor agonists as an inhibitory signal
for food and water intake. J. Neurochem. 67, 1982–1991.
Ngarmukos C., Baur E. and Kumagai A. (2001) Co-localization of
GLUT1 and GLUT4 in the blood–brain barrier of the rat ventro-
medial hypothalamus. Brain Res. 900, 1–8.
Nualart F., Hein S., Rodrıguez E. and Oksche A. (1991) Identification
and partial characterization of the secretory glycoproteins of the
bovine subcommissural organ-Reissner’s fiber complex. Evidence
for the existence of two precursor forms. Brain Res. Mol. Brain.
Res. 11, 227–238.
Nualart F., Godoy A. and Reinicke K. (1999) Expression of the hexose
transporters GLUT1 and GLUT2 during the early development of
the human brain. Brain Res. 824, 97–104.
Nualart F., Rivas C., Montecinos V., Godoy A., Guaiquil V., Golde D.
and Vera J. C. (2003) Recycling of vitamin C by a bystander effect.
J. Biol. Chem. 278, 10128–10133.
Oomura Y., Ono T., Ooyama H. and Wayner M. J. (1969) Glucose and
osmosensitive neurons of the rat hypothalamus. Nature 222, 282–
284.
Peruzzo B., Pastor F., Blazquez J., Schoebitz K., Pelaez B., Amat P. and
Rodrıguez E. (2000) A second look at the barriers of the medial
basal hypothalamus. Exp. Brain Res. 132, 10–26.
Rodrıguez E. M., Pena P., Aguado L. and Schoebitz K. (1985)
Involvement of tanycytes in the control of gonadotropin secretion,
in Proceedings of the International Symposium Comparative
Endocrinology. Current Trends in Comparative Endocrinology
GLUT2 expression in hypothalamic glial tanycytes 723
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724

(Lofts B. and Holmes W. N., eds), pp. 113–116. Hong Kong
University Press, Hong Kong.
Roncero I., Alvarez E., Vasquez P. and Blasquez E. (2000) Functional
glucokinase isoforms are expressed in rat brain. J. Neurochem. 74,
1848–1857.
Schuit F. C., Huypens P., Heimberg H. and Pipeleers D. G. (2001)
Glucose sensing in pancreatic b-cells. A model for study of other
glucose-regulated cells in gut, pancreas, and hypothalamus. Dia-
betes 50, 1–11.
Schwartz M. W., Woods S. C., Porte D. Jr, Seeley R. J. and Beskin D. G.
(2000) Central nervous system control of food intake. Nature 404,
661–671.
Spielholz C., Golde D. W., Houghton A. N., Nualart F. and Vera J. C.
(1997) Increased facilitated transport of dehydroascorbic acid
without changes in sodium-dependent ascorbate transport in human
melanoma cells. Cancer Res. 57, 2529–2537.
Thomzig A., Wenzel M., Karschin Ch, Eaton M., Skatchkov S. N.,
Karschin A. and Veh R. W. (2001) Kir6.1 is the principal pore-
forming subunit of astrocyte but not neuronal plasma membrane
K-ATP channels. Mol. Cell Neurosci. 18, 671–690.
Vannucci S. J. (1994) Developmental expression of GLUT1 and GLUT3
glucose transporters in rat brain. J. Neurochem. 62, 240–246.
Vera J. C., Rivas C., Fischbarg J. and Golde D. (1993) Mammalian
facilitative hexose transporters mediate the transport of dehydro-
ascorbic acid. Nature 364, 79–82.
Wan H. Z., Hulsey M. G. and Martin R. J. (1998) Intracerebroven-
tricular administration of antisense oligodeoxynucleotide against
GLUT2 glucose transporter mRNA reduces food intake, body
weight change and glucoprivic response in rats. J. Nutr. 128,
287–329.
Yang X. J., Kow L. M., Funabashi T. and Mobbs C. V. (1999)
Hypothalamic glucose sensor: similarities to and differences from
pancreatic b-cells mechanisms. Diabetes 48, 1763–1772.
Zamora-Leon S. P., Golde D. W., Concha I. I., Rivas C. I., Delgado-
Lopez F., Baselga J., Nualart F. and Vera J. C. (1996) Expression of
the fructose transporter GLUT5 in human breast cancer. Proc. Natl
Acad. Sci. USA 93, 1847–1852.
Zheng Q., Levitsky L. L., Mink K. and Rhoads D. B. (1995) Glucose
regulation of glucose transporters in cultured adult and fetal
hepatocytes. Metabolism 44, 1553–1558.
724 M. A. Garcıa et al.
� 2003 International Society for Neurochemistry, J. Neurochem. (2003) 86, 709–724
Related Documents











