INGESTIVE BEHAVIOR AND OBESITY Hypothalamic Dopamine and Serotonin in the Regulation of Food Intake Michael M. Meguid, MD, PhD, Serguei O. Fetissov, MD, PhD, Madhu Varma, MD, MS, Tomoi Sato, MD, Lihua Zhang, MD, PhD, Alessandro Laviano, MD, and Filippo Rossi-Fanelli, MD From the Neuroscience Program, Surgical Metabolism and Nutrition Laboratory, Department of Surgery, University Hospital, SUNY Upstate Medical University, Syracuse, New York, USA; and the Obesity Unit, Department of Clinical Medicine, University of “La Sapienza,” Rome, Italy Because daily food intake is the product of the size of a meal and the frequency of meals ingested, the characteristic of meal size to meal number during a 24-h light– dark cycle constitutes an identifiable pattern specific to normal states and obesity and that occurs during early cancer anorexia. An under- standing of simultaneous changes in meal size and meal number (constituting a change in feeding patterns) as opposed to an understanding of only food intake provides a more insightful dynamic picture reflecting integrated behavior. We have correlated this to simultaneous changes in dopamine and serotonin concentrations and to their postsynaptic receptors, focusing simultaneously on two discrete hypothalamic food-intake–related nuclei, in response to the ingestion of food. The relation between concentrations of dopamine and serotonin limited to the lateral hypothalamic area (LHA) and the ventromedial nucleus (VMN) as they relate to the influence of meal size and meal number during the hyperphagia of obesity and anorexia of cancer as measured in our experiments are discussed. Based on these data, conceptual models are proposed concerning: 1) an “afferent– efferent neurotransmitter unit,” with facilitatory or inhibitory neuropeptide properties to generate an appropriate neuroendocrine and neuronal response that ultimately modifies food intake; 2) initiation and termination of a meal, thereby determining the number and size of a meal under normal conditions; and 3) a schema integrating the onset mechanism of cancer anorexia. Nicotine is used as a tool to further explore the relation of meal size to meal number, with a focus on simultaneous changes in dopamine and serotonin concentrations in the LHA and VMN with the onset of acute anorexia of nicotine infusion and acute hyperphagia of nicotine cessation. Data concerning the role of sex-related hormones on dopamine and serotonin with regard to the LHA and VMN in relation to the modulation of food intake are also presented. Nutrition 2000;16:843– 857. ©Elsevier Science Inc. 2000 Key words: cancer anorexia, dopamine, serotonin, lateral hypothalamic area, ventromedial nucleus, cancer, obesity, nicotine INTRODUCTION Based on a body mass index (BMI) criterion of 25.0 to 29.9 kg/m 2 , approximately 25% to 33% of the U.S. population is overweight and another 30% is obese. 1,2 Growing evidence links an increase in BMI with an increased incidence of coronary heart disease, ele- vated blood pressure, and diabetes. 3,4 The higher prevalence of these conditions with obesity has focused public-health concern on the risk of being overweight. 5 There is increasing medical aware- ness of the physical, social, and economic costs of obesity in the industrialized world 6,7 that continues to be a major health problem with no effective treatment in sight. As a last resort, the treatment of morbid obesity has been relegated to the surgeon to perform a variety of surgical treatment manipulations (e.g., gastric stapling), but surgical treatment has a 20% long-term complication rate and a significant mortality risk. These procedures constitute a “fix” and do not presume to be based on an understanding of the cause of obesity. At the other end of the spectrum, a fall in BMI to less than 20 kg/m 2 in association with ongoing weight loss commonly occurs with the anorexia of chronic wasting diseases such as cancer, inflammatory bowel disease, or human immunodeficiency virus. Here, too, the risk of adverse effects on health and general well- being increases. People with a BMI of less than 18.5 kg/m 2 have a smaller muscle mass and their physical work capacity is re- duced. 8,9 This has serious implications that are even more signif- icant especially during illness because both myocardial and respi- ratory function are compromised. 10,11 Furthermore, with a BMI of less than 18.5 kg/m 2 , a deterioration in psychologic function is manifest by apathy, lassitude, lack of self-help motivation, depres- sion, and anxiety. 12 There is also evidence that the malnutrition secondary to hypophagia and anorexia of disease compromises the immune system, leading to impairment of cell-mediated immunity, This work was supported in part by NIH grants DK43796 and CA70239, American Institute for Cancer Research grant 85A95, Syracuse University Hendricks Award, NATO grant CRG940731, and Smokeless Tobacco Research Council Inc. grant 211-2506A. Dr. Laviano was supported by grant AI93.00131.04 from the Italian National Research Council (CNR). Correspondence to: Michael M. Meguid, MD, PhD, Department of Sur- gery, University Hospital, Upstate Medical University, 750 East Adams Street, Syracuse, NY 13210, USA. E-mail: [email protected] Date accepted: July 5, 2000. Nutrition 16:843– 857, 2000 0899-9007/00/$20.00 ©Elsevier Science Inc., 2000. Printed in the United States. All rights reserved. PII S0899-9007(00)00449-4

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

INGESTIVE BEHAVIOR AND OBESITY
Hypothalamic Dopamine and Serotonin in theRegulation of Food Intake
Michael M. Meguid, MD, PhD, Serguei O. Fetissov, MD, PhD, Madhu Varma, MD, MS,Tomoi Sato, MD, Lihua Zhang, MD, PhD, Alessandro Laviano, MD, and
Filippo Rossi-Fanelli, MDFrom the Neuroscience Program, Surgical Metabolism and Nutrition Laboratory,
Department of Surgery, University Hospital, SUNY Upstate Medical University, Syracuse,New York, USA; and the Obesity Unit, Department of Clinical Medicine, University of
“La Sapienza,” Rome, Italy
Because daily food intake is the product of the size of a meal and the frequency of meals ingested, thecharacteristic of meal size to meal number during a 24-h light–dark cycle constitutes an identifiablepattern specific to normal states and obesity and that occurs during early cancer anorexia. An under-standing of simultaneous changes in meal size and meal number (constituting a change in feedingpatterns) as opposed to an understanding of only food intake provides a more insightful dynamic picturereflecting integrated behavior. We have correlated this to simultaneous changes in dopamine and serotoninconcentrations and to their postsynaptic receptors, focusing simultaneously on two discrete hypothalamicfood-intake–related nuclei, in response to the ingestion of food. The relation between concentrations ofdopamine and serotonin limited to the lateral hypothalamic area (LHA) and the ventromedial nucleus(VMN) as they relate to the influence of meal size and meal number during the hyperphagia of obesityand anorexia of cancer as measured in our experiments are discussed. Based on these data, conceptualmodels are proposed concerning: 1) an “afferent–efferent neurotransmitter unit,” with facilitatory orinhibitory neuropeptide properties to generate an appropriate neuroendocrine and neuronal response thatultimately modifies food intake; 2) initiation and termination of a meal, thereby determining the numberand size of a meal under normal conditions; and 3) a schema integrating the onset mechanism of canceranorexia. Nicotine is used as a tool to further explore the relation of meal size to meal number, with afocus on simultaneous changes in dopamine and serotonin concentrations in the LHA and VMN with theonset of acute anorexia of nicotine infusion and acute hyperphagia of nicotine cessation. Data concerningthe role of sex-related hormones on dopamine and serotonin with regard to the LHA and VMN in relationto the modulation of food intake are also presented.Nutrition 2000;16:843–857. ©Elsevier Science Inc.2000
Key words: cancer anorexia, dopamine, serotonin, lateral hypothalamic area, ventromedial nucleus,cancer, obesity, nicotine
INTRODUCTION
Based on a body mass index (BMI) criterion of 25.0 to 29.9 kg/m2,approximately 25% to 33% of the U.S. population is overweightand another 30% is obese.1,2 Growing evidence links an increase inBMI with an increased incidence of coronary heart disease, ele-vated blood pressure, and diabetes.3,4 The higher prevalence ofthese conditions with obesity has focused public-health concern onthe risk of being overweight.5 There is increasing medical aware-ness of the physical, social, and economic costs of obesity in the
industrialized world6,7 that continues to be a major health problemwith no effective treatment in sight. As a last resort, the treatmentof morbid obesity has been relegated to the surgeon to perform avariety of surgical treatment manipulations (e.g., gastric stapling),but surgical treatment has a 20% long-term complication rate anda significant mortality risk. These procedures constitute a “fix” anddo not presume to be based on an understanding of the cause ofobesity.
At the other end of the spectrum, a fall in BMI to less than 20kg/m2 in association with ongoing weight loss commonly occurswith the anorexia of chronic wasting diseases such as cancer,inflammatory bowel disease, or human immunodeficiency virus.Here, too, the risk of adverse effects on health and general well-being increases. People with a BMI of less than 18.5 kg/m2 havea smaller muscle mass and their physical work capacity is re-duced.8,9 This has serious implications that are even more signif-icant especially during illness because both myocardial and respi-ratory function are compromised.10,11Furthermore, with a BMI ofless than 18.5 kg/m2, a deterioration in psychologic function ismanifest by apathy, lassitude, lack of self-help motivation, depres-sion, and anxiety.12 There is also evidence that the malnutritionsecondary to hypophagia and anorexia of disease compromises theimmune system, leading to impairment of cell-mediated immunity,
This work was supported in part by NIH grants DK43796 and CA70239,American Institute for Cancer Research grant 85A95, Syracuse UniversityHendricks Award, NATO grant CRG940731, and Smokeless TobaccoResearch Council Inc. grant 211-2506A. Dr. Laviano was supported bygrant AI93.00131.04 from the Italian National Research Council (CNR).
Correspondence to: Michael M. Meguid, MD, PhD, Department of Sur-gery, University Hospital, Upstate Medical University, 750 East AdamsStreet, Syracuse, NY 13210, USA. E-mail: [email protected]
Date accepted: July 5, 2000.
Nutrition 16:843–857, 2000 0899-9007/00/$20.00©Elsevier Science Inc., 2000. Printed in the United States. All rights reserved. PII S0899-9007(00)00449-4

phagocytosis, and antibody response.13 These changes in hostresistance increase susceptibility to infection.14 Consequently, pa-tients spend more time in the hospital,15 cannot tolerate full treat-ment regimens, experience a 40% to 60% greater frequency ofcomplications from surgical and medical treatment, take longer torecover from surgery, and have a two- to threefold increase indeath.16 Thus, the deviation beyond normal BMI range to the twoextremes causes major health problems associated with increasedphysical, social, and economic costs to the individual and tosociety.
Ultimately, an understanding of the neurobiochemical changesassociated with the hyperphagia or hypergluttony of obesity andthe hypophagia or anorexia of disease, i.e., an understanding of themechanisms that influence and control food intake and appetite, isessential to rationally treat these life-threatening conditions. Suchinformation will also advance our understanding of the normalcontrol of appetite and food intake, not only on earth but also in themicrogravity environment of space, where anorexia prevailsamong the astronauts.17,18
The long-term objectives of this laboratory continue to be theelucidation of the central and peripheral mechanisms that regulatefood intake and influence circadian rhythm in food intake. Westarted by focusing on the lateral hypothalamic area (LHA) andventromedial nucleus (VMN) and studying changes in dopamine(DA) and serotonin as measured by in vivo microdialysis. Thesemeasurements were related to simultaneous changes in meal sizeand meal number versus only food intake because this paradigmprovides a more meaningful approach to the understanding ofmechanism(s) in relation to food-intake control. From a pragmaticview point, we have approached the problem based on the thesisthat food intake (FI) is a function of the product of meal size (MZ)and meal number (MN), i.e.,FI 5 MZ 3 MN.
This simple but useful equation was initially proposed byothers.19,20 The concepts suggested by this equation are attractivebecause it provides several potential insights. 1) Meal size andmeal number are functionally interrelated, but to be separatelycontrolled they must be independently regulated, probably byseparate neuroanatomic sites. Under normal physiologic condi-tions in rats with ad libitum feeding, a change in one brings abouta compensatory change in the other, so as to maintain constancy ofdaily food intake.21 2) During human illness or in rat models ofdisease states such as hyperphagia of obesity22,23 or anorexia of
cancer,24,25 this reciprocal relation is disassociated. 3) Meal sizeand meal number are influenced by circadian rhythm: in normalrats, 70% is consumed during the dark cycle, whereas in hyperpha-gia of obesity an increase in light-phase food intake occurs,22 andin the anorexia of cancer a significant decrease in food intakeoccurs in the dark phase.25 4) Sex influences the relation of mealsize to meal number.26
Models Used to Study Hypothalamic Dopamine and Serotonin
We have used a variety of models that reflect dopamine andserotonin and leptin perturbations of food-intake controls. We haveused models of nicotine infusion to study acute anorexia and itscessation to gain insight into acute hyperphagia.27 To study theneurobiology of chronic anorexia we have used a rat cancermodel24,25; the study of the obese Zucker rat has provided insightinto the neurophysiology of chronic hyperphagia of obesity.28,29
Further insight was gained by studying the existence of sex dif-ferences.30,31 The integrated approach to investigating appetitecontrols involves the correlation and combination of several fac-tors, as outlined in Figure 1. These include 1) behavioral studies(measuring food intake and its constituents: meal size and mealnumber by a computerized rat eater meter developed in our labo-ratory32), 2) biochemical correlation to hypothalamic dopamineand serotonin via in vivo microdialysis, and 3) changes in hypo-thalamic receptor mRNA of dopamine and serotonin via reversetranscription–polymerase chain reaction, in situ hybridization, andimmunochemistry reflecting the coexpression with food-intake–related peptidergic neuromodulators. These studies have been fa-cilitated by using hypothalamically injected receptor subtype-specific agonists and antagonists to dopamine and serotonin.
Since the experiments by Hetherington and Ranson on bilaterallesion of the mediobasal hypothalamus including the VMN and thearcuate nuclei,33 the involvement of this area in food intake andbody-weight control has been generally accepted. Recently, itsneurochemical basis has gained even further support by the dis-covery that leptin brings about its action of reducing food intakeand body weight mostly through the neurons in the arcuate nucleusand the VMN.34 In addition to central anatomic areas, peripheralsites influencing the gastrointestinal tract and fat storage contributeto the regulation of food intake by changes in circulating nutrient
FIG. 1. Schema showing an afferent–efferent neurotransmitter unit, with a focus on dopamine and serotonin in relation to monoaminergic/peptidergic controlpoints and potential sites of their investigation in the study of food-intake regulation.
844 Meguid et al. Nutrition Volume 16, Number 10, 2000

concentrations and by related changes in neuroendocrinemediators.
Afferent vagal fibers from the gastrointestinal tract convergeonto the nucleus of tractus solitarius and from there project ontothe LHA. Because the study by Arees and Mayer35 demonstratedneuronal connections between the lateral and medial hypothala-mus, much additional anatomic, electrophysiologic, functional,neurochemical, and pathophysiologic data have consolidated theirrole in the reciprocal function in the regulation of food intake (seereferences in Table I). In this context, the LHA as the parasym-pathetic nervous system center and the ventromedial hypothalamusas the neuroendocrine sympathetic nervous system center are func-tionally integrated with other hypothalamic areas and nuclei arealso recognized in the control of food intake. These structures
include the paraventricular nucleus (PVN),36,37 the arcuate nucle-us,38,39 the dorsomedial hypothalamic nucleus,40 the VMN of thehypothalamus and the LHA,41–43, among others. Nevertheless,insufficient data exist to link together all these neuronal areas, sothat a comprehensive integrative functional picture of their inter-action in the regulation of food intake can emerge. However, theclose relation between two such areas, the LHA and VMN, hasbeen documented by data providing a relatively greater under-standing of not only their role in food-intake control but also theirinfluence on meal size and meal number as they relate to normaland diseased states, as summarized in Table I. This paradigmunderpins our present research in relation to the function of dopa-mine and serotonin.
LHA and VMN are involved reciprocally (Table I), beingcritical to food intake and energy-metabolism regulation.41–43En-hanced release of both norepinephrine and dopamine in the LHAis associated with inhibition of food intake44,45 but in the medialhypothalamic nuclei is associated with stimulation of food intake.Enhanced release of serotonin in the medial hypothalamic nuclei isassociated with inhibition of food intake.46 Thus, an effect ofmonoamine release seems to exist, which is specifically coordi-nated between lateral and medial hypothalamic areas in relation tofood intake, meal size, and meal number.
Rats, like humans, have a circadian rhythm in food intake. Inrats more than 70% of their food is consumed during the darkphase. The mechanism of this diurnal rhythm is not clear, but theretinohypothalamic projection to the suprachiasmatic nucleus(SCN), a pathway by which light information reaches the SCN, isinvolved in the generation of the circadian rhythm.47,48 The SCNsends monosynaptic efferents to the VMN. Electrophysiologicstudies have shown that the SCN exerts mainly an excitatoryinfluence on glucoreceptor neurons in the VMN. The SCN hasmultisynaptic inhibitory influence on the glucose-sensitive neuronsin the LHA. Thus, with light-on stimulus the SCN stimulates VMNand inhibits LHA, resulting in suppression of feeding. Further, thelight-on stimulus is a major regulatory factor of melatonin synthe-sis and secretion in the pineal gland. VMN neuronal activity isinfluenced by the pineal gland, which in turn is affected by theSCN.49 The SCN is involved in the entrainment and rhythmgeneration; the light–dark cycle being the primary synchronizingagent. Bilateral SCN lesions alter the circadian rhythms of feeding,whereas continuous light exposure suppresses total daily food andwater intake. In addition to the SCN light-entrained endogenouscircadian oscillator, the VMN50 and the LHA51 are entrained byfood intake as a second circadian oscillator. Glucose (i.e., carbo-hydrate) versus fat primarily affects the food-entrained ossilator.52
Neurons in VMN contain insulin, leptin, and estrogen receptorsand thus receive hormonal signals concerning small daily gains orlosses in calorie storage in peripheral fat deposits53 regulatingenergy metabolism.54,55 Electrolytic lesion of the VMN attenuatethe daily expression of several rhythms including feeding behaviorand corticosterone levels.56 Thus, the food-intake circadian systemconsists of at least two major pace-making systems, one entrainedby light (SCN) and the other entrained by food (the anatomicallyinterconnected and functionally interrelated VMN and LHA).
Broad Concepts of Food-Intake Regulation
The ingestion of food is a complex process that integrates sensoryinformation related to sight, smell, and taste of food, to previousingestion experience, and to satiety signals elicited by ingestionand hormonal signals related to energy balance. Conceptually,information concerning body energy and nutritional status (fromgut, liver, and fat stores) is continuously transmitted to the brain by1) the afferent autonomic neurons, 2) changes in concentrations ofcirculating nutrients and hormones, and 3) gastrointestinal andcentral neuroendocrine mediators. Relevant areas of the forebrainand midbrain further modulate this information. This “afferent
TABLE I.
RELATIONSHIP BETWEEN LHA AND VMN
Factor Comments
Anatomic Dendrites from both LHA and VMN are in closeproximity, enabling passage of LHA andVNM fiber trajectories interact with each other
Electrophysiologic VMN neuron frequency increase correlates witha decrease in LHA neuron frequency
Highly significant inverse relation of SUDSbetween VNM and LHA
Activity of LHA neurons affects gastricdistention
Physiologic LHA neuron stimulation activates feeding,whereas VMN neuron stimulation inhibitsfeeding
LHA DA increases in response to food intakeafter food deprivation
VMN DA decreases in response to food intakeafter food deprivation
LHA DA correlates with meal sizeVMN DA concentrations control the intermeal
interval and thus meal numberLeptin acts in both LHA and VMN
Functional–behavioral
Food stimulant (odor/sight) LHA EGG changedfrom high-amplitude, slow waves to low-amplitude fast waves; EGG of VMN showedthe reverse trend
Electrical stimulation of VMN and LHA causesreciprocal effects on glucose and glycogenlevels
LHA lesions increase intestinal transitNeurochemical Shows how neuropeptide CART acts on VMN to
cause anorexiaDA stimulation has opposite effects on the LHA
and VMNOrexin’s effect on LHA causes increase in food
intakeLHA controls short-term satiety and VMN
controls long-term satiationPathophysiologic VMN lesion reduces energy expenditure
Anorexia or cancer and IBD mediated by IL-1on VMN reduce meal number
LHA DA concentrations and meal size arebigger in obese than in lean Zucker rats
CART, cocaine and amphetamine-regulated transcript; DA, dopamine;EEG, electroencephalogram; IBD, inflammatory bowel disease; IL-1,interleukin-1; LHA, lateral hypothalamic area; SUDS, sudden unex-plained death syndrome; VMN, ventromedial nucleus.
Nutrition Volume 16, Number 10, 2000 845Dopamine and Serotonin in the Regulation of Food Intake

sensory” information is ultimately integrated into the hypotha-lamic food-intake–related nuclei by an “afferent–efferent neuro-transmitter unit” with facilitatory or inhibitory neuropeptide prop-erties to generate an adequate neuroendocrine and neural responsethat is manifested by changes in meal size and meal number (Fig.1).
We postulate that at this “neurotransmitter unit” the afferentinformation, which includes current metabolic status, is conveyedto the presynaptic monoaminergic system of the hypothalamus,where it is stored at the end of the presynaptic afferent neuron asprepackaged synaptosomes of neurotransmitters dopamine andserotonin. The presynaptic cell influences postsynaptic cells byrelease of 103 to 105 molecules of dopamine and serotonin bindingto postsynaptic receptors. Dopamine and serotonin may eitherfacilitate or inhibit second-messenger intracellular activity basedon the function of their receptor subtype to which they bind. Asoutlined schematically, this conveys information concerning mealsize, meal number, and food intake by activation of postsynapticneurons of the stimulatory food-intake neuropeptides (e.g.,melanin-concentrating hormone (MCH), neuropeptide Y (NPY),galanin, and orexin A/B) or by the inhibitory food-intake neu-ropeptides (e.g., melanocyte-stimulating hormone (MSH), cocaineand amphetamine-regulated transcript (CART), and corticotropin-releasing factor (CRF), among others), depending on the prevail-ing metabolic conditions, to effector organs to modulate foodintake. Leptin, insulin, and other peripheral peptides and steroidhormones modulate the synthesis and release of dopamine andserotonin. The concept of the “afferent–efferent neurotransmitterunit” (which “ reads” information from periphery and initiatescentrally mediated efferent response) provides a “cytomolecularworking diagram” to facilitate the investigation of normal anddisease-related food intake processes. Our laboratory has used theconcept shown in Figure 1 to formulate hypotheses to investigatekey control points upon which the data in this review are based.
The Neurotransmitters Dopamine and Serotonin
Dopamine and serotonin are two critical food-intake–related neu-rotransmitters. Among the monoamines, dopamine and serotoninare relevant brain messengers in food-intake regulation.44–46,57–62
Among other appetite-related brain messengers, dopamine is es-sential for feeding. The absence of dopamine production in theknockout mouse, which does not express tyrosine hydroxylase,44,63
does not inhibit motor capabilities to eat but causes an inability toinitiate feeding; this ability can be restored by gene delivery oftyrosine hydroxylase into the striatum.64 Dopamine release seemsto have a site-specific action in food-intake regulation, such as inthe nucleus accumbens, and it is associated with the reinforcementeffect in feeding.65 In the hypothalamus dopamine release is as-sociated with the duration of meal consumption, which is a factorin determining feeding pattern. Hence, dopamine is required toinitiate each meal and is thus associated with meal number andduration, i.e., meal size.41,42 Increased dopamine in the VMN, thearea of neuroendocrine and autonomic regulation of metabolism,accompanies food intake.66 Medial hypothalamic lesions produceboth hyperphagia and morbid obesity, suggesting that VMN do-pamine is involved in regulating both food intake and the body-weight setpoint.
Serotonin neurons of the midbrain and brainstem raphe nucleiare among the earliest neuronal phenotype to become differenti-ated during development of the central nervous system, and sero-tonin is thought to act as a regulatory and developmentalagent.67,68The more rostral nuclei (primarily the dorsal and medialraphe) form ascending projections, densely innervating such fore-brain areas as the hypothalamus, hippocampus, and cortex. Thus,the serotonergic system is involved in the regulation of suchdiverse functions as reproduction, mood, sleep, and cognition.Mice lacking serotonergic receptors display food-intake and body-
weight–related abnormalities.69 Serotonin release in the hypothal-amus is enhanced during feeding,70 is thought to promote satiation,and reflects carbohydrate ingestion.71
Numerous studies have characterized serotonin and the sero-toninergic system in the inhibitory role of food intake. The sug-gestion that serotonin acts to inhibit food intake is based onfindings that a variety of different directly and indirectly actingagonists at central serotonin synapses cause dramatic reduction infood intake.72 A body of evidence exists showing that dopaminerelease is regulated in part by serotonin-release.45,73,74
To function as compensatory mechanisms, it is likely that mealsize and meal number are independently regulated in different yetconnected anatomic sites of the brain. Using the automated com-puterized rat eater meter32 and the in vivo brain microdialysistechnique,75 we measured simultaneously the intra-VMN andintra-LHA concentrations of dopamine and serotonin as they re-lated to the control of meal size and meal number. Our data andthose of others indicate that, under normal conditions, the interac-tion between dopamine and serotonin within the LHA influencesmeal size42,70,76–78and that intra-VMN dopamine and serotonininfluence meal number.41,71,79
ROLE OF DOPAMINE AND SEROTONIN IN THEREGULATION OF FOOD INTAKE IN OBESITY
The feeding pattern of the obese Zucker rat versus its lean coun-terpart demonstrates a consistently larger meal size throughout the24-h light–dark cycle; thus, the obese Zucker rat consistentlyconsumes more food than its lean counterpart. The importantcontribution of dopamine to the regulation of food intake andfeeding pattern has been shown in the medial hypothalamus by thestudy of both pre- and postsynaptic dopaminergic systems, asmeasured by in vivo microdialysis. Dopamine release is rapidlybut transiently increased in the VMN during spontaneous eating inlean and obese rats, and this increase is more pronounced in obeserats, which consume larger meals. When rats are deprived of foodfor 24 h, food intake brings about an immediate decrease ofdopamine release to 65% of preeating levels in both lean and obeserats.29 These two findings shows that release of dopamine isassociated with both short-term (individual meals) and long-term(hunger) regulation of food intake. These data also suggest thatduring obesity the presynaptic dopaminergic system adequatelyresponds to the change of feeding status.
Extracellular concentration of VMN dopamine can be raisedphysiologically by grafting embryonic dopaminergic cells, as doc-umented by microdialysis. This grafting resulted in the increase offood intake by meal number and then by meal size.22 The VMNdopaminergic graft also resulted in changing the diurnal distribu-tion of food intake, so that food intake in the dark phase increasedthe change in the feeding pattern in obese rats.23 By measuringdopamine release every 20 min by microdialysis in the VMN for24 h, we found that the release has a diurnal rhythm influenced byfood deprivation.51 This change in feeding pattern that occurs inthe obese rat suggests an involvement of the diurnal rhythm inVMN dopamine release, resulting in abnormally high eating ac-tivity of obese rats during the light phase. A food-intake stimula-tory function for VMN dopamine was confirmed by a singlepharmacologic injection (10 nM) of dopamine into the VMN offreely moving lean and obese rats, which caused an increase ofmeal size with a compensatory decrease in meal number (unpub-lished observations).
Dopamine acts on its receptors postsynaptically to modulate theactivity of target neurons by stimulation or inhibition of adenylylcyclase activity by D1 and D5 or D2 to D4 subtypes of receptors,respectively. The finding that hypothalamic neurons express bothsubtypes of dopaminergic receptors provides evidence that dopa-mine may activate or inhibit its target neurons based on the 1)
846 Meguid et al. Nutrition Volume 16, Number 10, 2000

function of the subtype and 2) level of the dopaminergic receptorexpressed for a given metabolic condition under normal or patho-logic conditions. Using reverse transcription–polymerase chainreaction, we found that, under ad libitum feeding conditions, thelevel of expression of D1- and D2-receptor mRNA measured whenthe feeding activity of rat is minimal (between 10 and 11AM) wasdifferent in the VMN of lean and obese rats (unpublished obser-vations). D1 receptor, which normally is not abundantly expressedin the hypothalamus, was found highly expressed in the VMN ofobese rats, whereas D2 receptor was detected in the lean rat but atan undetectable level in the VMN of the obese Zucker rat. Thedifferent levels of dopaminergic receptor expression in the twophenotypes may be related to the relatively low concentration ofligand (dopamine) in obese as opposed to lean rats,80 or it maydirectly depend on altered leptin signaling in postsynaptic neurons.
We tested the relevance of different levels of dopaminergicreceptor expression between obese and lean rats on food intake andfeeding pattern by using a single pharmacologic injection (10 nM)of specific receptor antagonists. A different response to food intakeand feeding pattern (relation of meal size to meal number) wasfound in the obese than in the lean rat. This finding allows us toconclude that the level of expression of different subtypes ofdopaminergic receptors in the VMN is directly involved in theregulation of meal size and meal number.
Model of Hypothalamic Dopaminergic and SerotonergicRegulation in Obesity
Based on our results,22,23,28,29,41,42,51,72,76–82we constructed amodel of dopaminergic and serotonergic regulation of food intake,as shown in Figure 2. This model is based on the followingpostulates. 1) Dopamine may act on three populations of neurons:neurons that influence meal number by prolonging intermeal in-tervals by the release of food-intake inhibitory neuropeptides suchasa-MSH and CART; neurons influencing meal size by startingand maintaining the meal by the release of food-intake stimulatory
neuropeptides such as NPY and Agouti-related protein (AgRP);and neurons influencing meal size by stopping the meal by therelease of food-intake inhibitory neurohormones and neuropep-tides such as vasopressin, oxytocine (OT), or corticotropin-releasing factor (CRF). 2) All three populations of neurons mayexpress both D1 and D2 receptors, which can be activated orinhibited by dopamine. 3) Dopamine first activates the D2 and thenthe D1 receptor, which have different thresholds of activation. Inthis way, dopamine can change its effect from inhibition to acti-vation according to its concentration at a specific subtype receptor.
According to our model (Fig. 2), the circadian or sensory-stimulated release of dopamine will inhibit food-intake inhibitoryneurons (2FI) and then activate food-intake stimulatory neurons(1FI) to start the next meal. The ingestion of food also stimulatesdopamine release through its oral and gastric components, whichmaintain the meal through1FI neurons until2FI neurosecretoryneurons stop the meal. Le Magnen and Devos’s postulate83 that ahighly significant correlation exists between meal size and elapsedtime after the onset of a new meal could be a reflection ofdopamine’s action. This action would stimulate inhibitory neu-ropeptide synthesis in quantities proportional to meal size.
This model is an example of how one neurotransmitter could beinvolved in the regulation of food intake. Other monoaminergicand peptidergic neurotransmitters exist that probably activate orinhibit either adenylyl cyclase or a different second messenger thatwould function synergistically or antagonistically with dopa-mine.74,84 However, having a simple model facilitates our under-standing of the involvement of dopamine in the complex regula-tion of food intake.
An example of the interaction of dopamine with another mono-amine is the release of serotonin in the hypothalamus. We inves-tigated this possibility by inserting double cannulas, one placedinto the ipsilateral LHA and the other into the contralateral VMN.Microdialysis was performed every 20 min simultaneously inawake, freely moving active rats during a complete 24-h light–dark cycle. Two paradigms were studied: the fed state and the
FIG. 2. Proposed model of hypothalamic dopamine and serotonin and neuropeptide interaction in the regulation of food intake, meal size, and meal numberbased on data derived from studies in the lateral and ventromedial hypothalamus.
Nutrition Volume 16, Number 10, 2000 847Dopamine and Serotonin in the Regulation of Food Intake
fasted state. A well-defined rise in dopamine occurred in the LHAat late midday before the initiation of feeding. Dopamine rose to apeak with food intake. In contrast, in the fasted state, the initialdopamine rise was blunted in the absence of food. By applyingtime-series analysis to the data of simultaneous serotonin anddopamine release as measured by microdialysis every 20 min for24 h, we found that concurrent release of dopamine and serotoninwithin the VMN and the LHA is synchronized in the fed stateduring intermeal intervals.51 It was not synchronized during eatingor during fasting; food intake desynchronized dopamine and sero-tonin release within and between the ventromedial hypothalamusand the LHA.
These data point to a tight relation between dopaminergic andserotonergic systems, which is influenced by the feeding state of ananimal and which may be involved in maintaining the balancewithin and between sympathetic and parasympathetic nervoussystem centers. The data show that food intake is coupled unequiv-ocally to the release of dopamine and serotonin in the VMN andLHA, suggesting it as a mechanism of activation of postsynapticneurons associated with prevailing metabolic conditions.
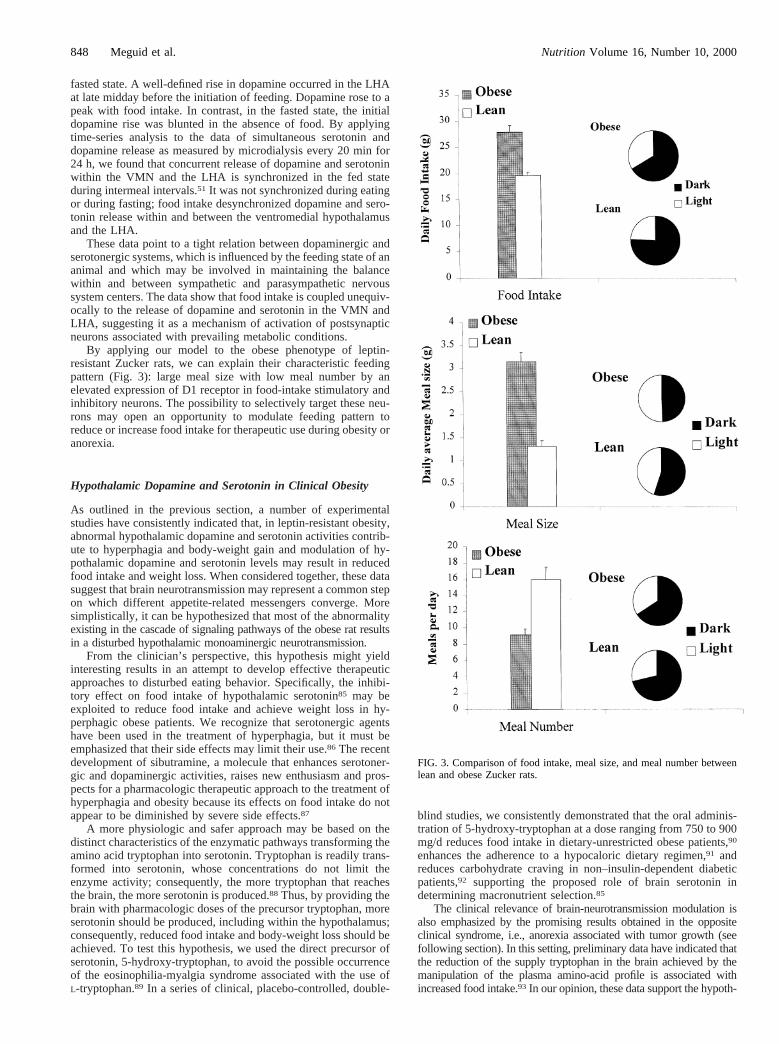
By applying our model to the obese phenotype of leptin-resistant Zucker rats, we can explain their characteristic feedingpattern (Fig. 3): large meal size with low meal number by anelevated expression of D1 receptor in food-intake stimulatory andinhibitory neurons. The possibility to selectively target these neu-rons may open an opportunity to modulate feeding pattern toreduce or increase food intake for therapeutic use during obesity oranorexia.
Hypothalamic Dopamine and Serotonin in Clinical Obesity
As outlined in the previous section, a number of experimentalstudies have consistently indicated that, in leptin-resistant obesity,abnormal hypothalamic dopamine and serotonin activities contrib-ute to hyperphagia and body-weight gain and modulation of hy-pothalamic dopamine and serotonin levels may result in reducedfood intake and weight loss. When considered together, these datasuggest that brain neurotransmission may represent a common stepon which different appetite-related messengers converge. Moresimplistically, it can be hypothesized that most of the abnormalityexisting in the cascade of signaling pathways of the obese rat resultsin a disturbed hypothalamic monoaminergic neurotransmission.
From the clinician’s perspective, this hypothesis might yieldinteresting results in an attempt to develop effective therapeuticapproaches to disturbed eating behavior. Specifically, the inhibi-tory effect on food intake of hypothalamic serotonin85 may beexploited to reduce food intake and achieve weight loss in hy-perphagic obese patients. We recognize that serotonergic agentshave been used in the treatment of hyperphagia, but it must beemphasized that their side effects may limit their use.86 The recentdevelopment of sibutramine, a molecule that enhances serotoner-gic and dopaminergic activities, raises new enthusiasm and pros-pects for a pharmacologic therapeutic approach to the treatment ofhyperphagia and obesity because its effects on food intake do notappear to be diminished by severe side effects.87
A more physiologic and safer approach may be based on thedistinct characteristics of the enzymatic pathways transforming theamino acid tryptophan into serotonin. Tryptophan is readily trans-formed into serotonin, whose concentrations do not limit theenzyme activity; consequently, the more tryptophan that reachesthe brain, the more serotonin is produced.88 Thus, by providing thebrain with pharmacologic doses of the precursor tryptophan, moreserotonin should be produced, including within the hypothalamus;consequently, reduced food intake and body-weight loss should beachieved. To test this hypothesis, we used the direct precursor ofserotonin, 5-hydroxy-tryptophan, to avoid the possible occurrenceof the eosinophilia-myalgia syndrome associated with the use ofL-tryptophan.89 In a series of clinical, placebo-controlled, double-
blind studies, we consistently demonstrated that the oral adminis-tration of 5-hydroxy-tryptophan at a dose ranging from 750 to 900mg/d reduces food intake in dietary-unrestricted obese patients,90
enhances the adherence to a hypocaloric dietary regimen,91 andreduces carbohydrate craving in non–insulin-dependent diabeticpatients,92 supporting the proposed role of brain serotonin indetermining macronutrient selection.85
The clinical relevance of brain-neurotransmission modulation isalso emphasized by the promising results obtained in the oppositeclinical syndrome, i.e., anorexia associated with tumor growth (seefollowing section). In this setting, preliminary data have indicated thatthe reduction of the supply tryptophan in the brain achieved by themanipulation of the plasma amino-acid profile is associated withincreased food intake.93 In our opinion, these data support the hypoth-
FIG. 3. Comparison of food intake, meal size, and meal number betweenlean and obese Zucker rats.
848 Meguid et al. Nutrition Volume 16, Number 10, 2000

esis that brain neurotransmission may represent a common step onwhich different appetite-related messengers converge.
ROLE OF SEROTONIN AND DOPAMINE IN THEREGULATION OF FOOD INTAKE IN ANOREXIA OFCANCER
Sex differences in feeding patterns exist in normal rats.26 They alsooccur in tumor-bearing (TB) rats.94 As outlined in the schema (Fig.4), the changes in the feeding patterns as observed in canceranorexia are a manifestation of a chain of biochemical eventspreceding and leading to abnormal hypothalamic monoaminergicactivity.
In male anorectic Fischer 344 rats with methyl cholanthrene(MCA) sarcoma, anorexia is observed to occur with tumor growth.As shown in Figure 5, in the early phase meal number progres-sively decreases, but food intake is maintained relatively constantthrough an apparently compensatory increase in meal size. Ap-proximately 1 to 2 d later, meal size significantly decreases andanorexia becomes profound and apparent.24,25,95
Anorexia also develops in female Fischer-344 rats. It is pre-dominantly due to a decrease in meal number; meal size remainsrelatively constant. Differences in feeding patterns between normaland TB female and male rats is dependent on the presence offemale sex hormone.94 This hypothesis was tested using ovariec-tomized female TB rats. The pattern of development of anorexia inovariectomized TB rats was the same as that in male rats: an initialdecrease in meal number followed by a decrease in meal size. Insham-operated TB female rats, meal size remained unchanged.The degree of anorexia in ovariectomized TB rats was significantlygreater, with a consequent lower food intake. This observationsuggests that female sex hormones, predominantly estradiol, playa significant role in feeding behavior and in immune functioncomparable to that observed in female septic patients.96
A similar pattern, i.e., a decrease in food intake through aninitial decrease in meal number followed by a decrease in mealsize, can be reproduced with systemic injection of interleukin(IL)–1a and/or tumor necrosis factor (TNF)–a.97,98Because at theonset of anorexia IL-1a in the cerebrospinal fluid is elevated in TB
rats99 and IL-1 receptors exist in the VMN,100 the role of cytokinesin cancer anorexia was determined by injecting IL-1 receptorantagonist in the VMN of anorectic TB rats.101 The results showedthat food intake increased only through an increase in meal num-ber. Furthermore, the systemic infusion of the TNF-receptor con-struct into anorectic TB rats increased food intake by both in-creases in both meal size and meal number.102 Based on the sumof these data, IL-1 and TNF-a play a pivotal role in the mechanismof cancer anorexia. Such cytokines act directly103and indirectly onthe hypothalamus by changes in monoamine release of dopamineand serotonin.104 Abnormalities of monoaminergic105,106and pep-tidergic107,108systems occur during cancer anorexia.
The concentration of NPY decreased in the PVN but increasedin the arcuate nucleus,108 both of which play a significant role inthe regulation of food intake. These data suggest that peptidergicdysfunction in the hypothalamic food-intake–related nuclei con-tributes to the mechanism(s) of cancer anorexia. Such mechanismsare well summarized elsewhere109; in this report, we focus ourinterest on hypothalamic dopaminergic and serotonergic dysfunc-tion in cancer anorexia.
The hypothalamic serotonergic system contributes to canceranorexia.105,110 Serotonin is a known satiating factor and sup-presses food intake when injected into the VMN of normal rats,111
where it plays a critical role in the cancer anorexia.95 Serotoninconcentrations are increased in the VMN in anorectic TB rats, asmeasured by in vivo microdialysis, and normalize after tumorresection.106We were also able to increase serotonin concentrationin the VMN by injection of IL-1a into the VMN.104
The synthesis of brain serotonin directly depends on the plasmaconcentration of its precursor, tryptophan.88,112In anorectic cancerpatients and TB rats, we measured an increase of tryptophanconcentration in plasma and cerebrospinal fluid24,113,114that pos-itively correlated with the degree of anorexia. Plasma-free trypto-phan is carried competitively across the blood–brain barrier by ashared transport system with other large neutral amino acids.115
Hence, the molar ratio of plasma-free tryptophan to large neutralamino acid is an important factor determining the brain availabilityof tryptophan and thus brain serotonin synthesis and activity, aspreviously reported.113 In addition, endotoxin or cytokines (IL-1)also facilitate plasma-free tryptophan synthesis through the sym-
FIG. 4. Schema indicating interaction of cytokines, monoamines, and neuropeptides with changes in meal size and meal number in cancer anorexia.
Nutrition Volume 16, Number 10, 2000 849Dopamine and Serotonin in the Regulation of Food Intake

pathetic nervous system,116 indicating a link between cancer, cy-tokine, and serotonin (tryptophan). Furthermore, oral administra-tion of branched-chain amino acid, to decrease the ratio oftryptophan to large neutral amino acids, significantly increasedfood intake in cancer patients93; in patients with cancer plasma-free tryptophan decreased after curative tumor resection.117
In TB states hypothalamic dopaminergic system also appears tobe abnormal, which may be a consequence of serotoninergicsystem dysfunction, because of their interaction.73,118We reportedthat IL-1a facilitated VMN dopamine release in normal rats.104
However, in anorectic TB rats, we found low dopamine concen-trations in the VMN and a high turnover rate.106
We recently observed that dopaminergic receptor mRNA ex-pression also changed in anorectic TB rats. We measured foodintake, meal size, and meal number and dopaminergic D1- andD2-receptor mRNA expression in the hypothalamus in anorecticTB and non-TB normal rats. When anorexia developed, mani-fested by the characteristic feeding-pattern changes, D2 and D1receptors were highly expressed. D2 is postulated to influencesatiety119; D1 receptor is not abundantly expressed under normalconditions.120
To determine the function of the changes in the expression ofD1 and D2 receptor, we injected specific dopaminergic receptorantagonists into the VMN in anorectic TB and non-TB normal rats.The results showed that sulpiride (D2-receptor antagonist) signif-icantly increased food intake by an increase in meal number onlyin anorectic TB rats but did not significantly increase food intakein non-TB normal rats. SCH23390 (D1-receptor antagonist) de-creased food intake by a decrease in meal size in anorectic TB ratsbut failed to change food intake in non-TB normal rats. ObeseZucker rats have high D1- and low D2-receptor expression asso-ciated with huge meal size and increased food intake. Based onthese two sets of data, we deduce that, in anorectic TB rats, the
highly expressed D1 receptor in the hypothalamus facilitates foodintake and meal size, whereas the highly expressed D2 receptor inthe hypothalamus contributes to anorexia. Consequently, thesechanges in dopaminergic receptor expression in the hypothalamuscontribute to the feeding pattern that characteristically occurs incancer.
Leptin has anorectic properties when injected centrally andperipherally. Because cytokines such as TNF and IL-1 stimulateplasma leptin and leptin mRNA expression in the adipose tissue,121
determining role of leptin in cancer anorexia is of interest. Thus,we measured plasma leptin concentration at an early and a latephase of anorexia of cancer. We found that plasma leptin concen-tration at the early phase was not different between TB andnon-TB pair-fed rats. However, higher leptin concentrations wereobserved at the late phase of anorexia in TB as opposed to non-TBpair-fed rats,122 although the concentrations in both groups weresignificantly lower compared with those at the early phase, afinding that is in agreement with that reported by others.123,124
From this study, we conclude that leptin does not initiate anorexiaof cancer but is involved in the development and the maintenanceof cancer anorexia.
NPY-deficient mice have normal feeding behavior,125 whereasdopamine-deficient mice are aphagic,63 indicating that peptidergicsystem dysfunction can be compensated for by the monoaminergicsystem to initiate feeding. Consequently, manipulating the mono-aminergic system in cancer to compensate for peptidergic dysfunc-tion results in amelioration of cancer anorexia.
We injected specific monoaminergic receptor antagonists into thehypothalamus. Mianserin (serotonin 2C–receptor antagonist126) andsulpiride (D2-receptor antagonist) injected into the VMN or LHAincreased food intake in anorectic TB rats. However, enhancingendogenous hypothalamic dopamine production by implanting dopa-
FIG. 5. Changes in food intake with onset of anorexia. (a) Daily light- and dark-phase food intake, (b) meal number, and (c) meal size in anorectictumor-bearing rats. Day 0 is the day of onset of anorexia defined as a decrease in the rat’s food intake per 100 g of body weight, which is 1 g lower thanthe average in each rat before tumor inoculation for 3 consecutive d. Data from day23, the last day of normal food intake, to day12. Coefficient correlationswere applied to changes in food intake, meal number, and meal size for the period from day23 to day 0 to statistically describe the changes in the relationof meal number to meal size to determine the development of cancer anorexia. The mean linear correlation coefficient between food intake and meal numberwas 0.95 and that between food intake and meal size was 0.49, suggesting that the reduction of food intake at the onset of anorexia is due mainly to thedecrease in meal number. On day12, meal size decreased significantly.
850 Meguid et al. Nutrition Volume 16, Number 10, 2000

minergic cells into the hypothalamus to facilitate physiologic dopa-mine release23 changed the feeding pattern but failed to changetemporally and quantitatively the development of anorexia in TB rats.
Hypothalamic Dopamine and Serotonin in Clinical CancerAnorexia
The sum of the evidence highlights the fact that, when the anorexiaof cancer is present, abnormal hypothalamic dopaminergic andserotonergic neurotransmission is contemporaneously detectable.Cytokines are intimately involved in the process of cancer an-orexia through different mechanisms including interference withmonoaminergic neurotransmission. Hence, pharmacologic manip-ulation to inhibit their synthesis usingv-3 fatty-acid–based phar-maconutrition or to suppress their function using pentoxifylline,for example, may provide a beneficial effect to counteract anorexiaand its detrimental metabolic sequelae by increasing food intake.In keeping with this reasoning, interesting and promising resultshave been obtained in anorectic TB rats receiving a dimeric,pegylated TNF receptor construct to block TNF activity.102 In thisstudy, we found that by blocking TNF activity a significant im-provement of food intake can be achieved. A similar drug isalready available on the market (Infliximab), but no report hasbeen published testing its effects on food intake in anorexia. Thus,it is extremely difficult to speculate whether this therapeutic strat-egy will result in clinically relevant benefits. Also, because themechanism(s) of cancer anorexia is multifactorial, it seems un-likely that blocking one of many pathways will have a significantand consistent effect.
In 1979, Krause et al. first demonstrated in an animal modelthat the decrease in food intake after tumor implantation is asso-ciated with a significant increase in brain tryptophan and serotonincontent.105 From these data, it was hypothesized that increasedbrain serotonin activity secondary to increased brain availability oftryptophan contributes to anorexia in cancer. Supporting this hy-pothesis, we confirmed these data in an experimental model ofanorexia,24 but we were also able to show that the increase of braintryptophan availability and content are common features in cancerpatients.113,114 Strengthening the relation between tumor, trypto-phan, and anorexia, we also showed that plasma tryptophan levelsnormalize after curative tumor resection in patients, resulting inimproved food intake.117Such data lend credence to the hypothesisthat serotonin is a critical anorectic agent.
In keeping with our hypothesis, the hypothalamic monoamin-ergic system may represent a major target on which differentappetite-related messengers converge; the strategy of interferingwith serotonin systems may produce encouraging results. Becausethe brain availability of tryptophan depends on the molar ratio ofplasma-free tryptophan to large neutral amino acid, which includethe branched-chain amino acids, we hypothesized that changes inplasma amino-acid concentration induced by oral administration ofbranched-chain amino acids, to decrease the ratio in cancer pa-tients, would decrease brain tryptophan availability and thus sero-tonin synthesis, leading to increased appetite and food intake. Inline with this reasoning, administration of branched-chain aminoacids to anorectic cancer patients resulted in the normalization ofthe ratio of plasma-free tryptophan to large neutral amino acid,decreased the incidence of anorexia in the study group, and sig-nificantly improved food intake.93
From the sum of these experimental and clinical data, wededuce that monoamines play a major role in the neural modula-tion of feeding behavior. Their manipulation appears to compen-sate for other abnormalities of food-intake–related systems. Fur-thermore, from the clinician’s perspective, the administration ofbranched-chain amino acids to modulate brain serotonin concen-tration appears to be safe and likely provides effective therapy toincrease food intake. Unfortunately, no available data currentlyexist indicating whether increased food intake in anorectic cancer
patients results in improved long-term endpoints, i.e., morbidityand mortality rates. However, while waiting for clinical reports toanswer these questions, it must be emphasized that restoring foodintake to normal or near normal significantly improves the qualityof life of the patient with anorexia, thus positively affecting psy-chological status, which is of great help in withstanding the det-rimental effects of aggressive antineoplastic regimens often usedin the treatment of cancer patients.
ROLE OF NEUROTRANSMITTERS AND NPY IN THEREGULATION OF FOOD INTAKE DURING NICOTINECONSUMPTION
Despite the fact that most people are aware of the health risksassociated with using tobacco products, approximately one-fourthof adults use them.127 Obviously, there must be some compellingreasons for its continued use. One frequently cited reason is thebelief that the use of tobacco products can suppress appetite andcontrol body-weight gain. There are also studies documentingweight gain after the cessation of smoking.128
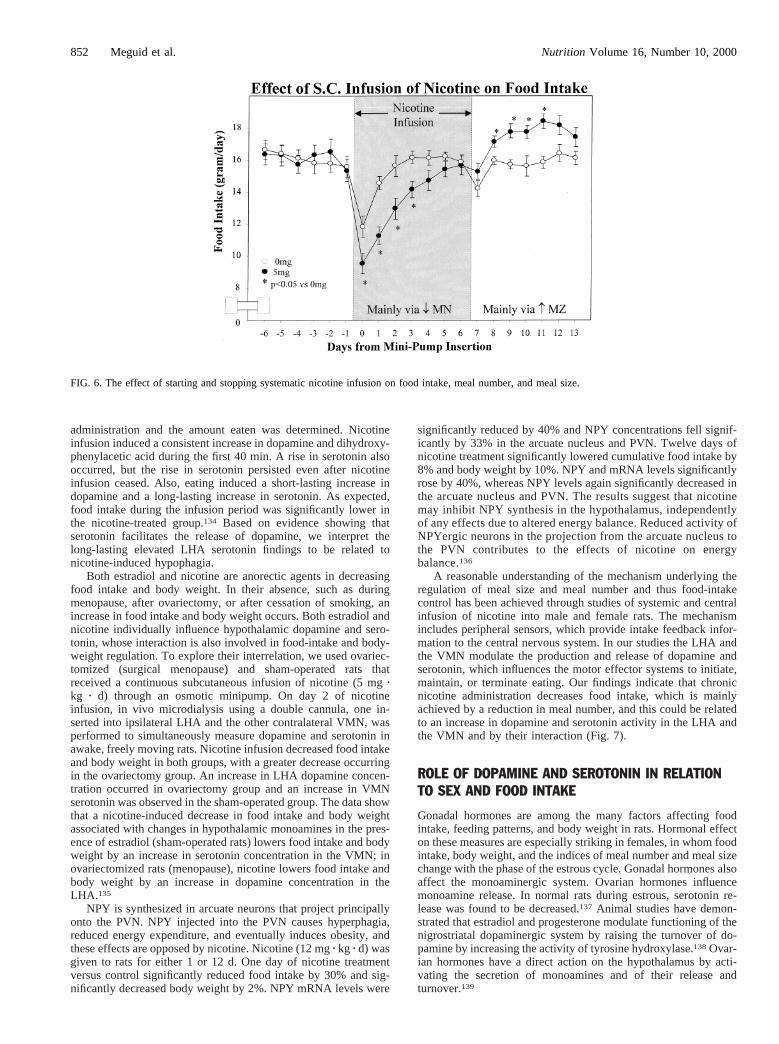
Nicotine is the most pharmacologically active ingredient intobacco and is believed to be responsible for the tobacco-inducedinhibitory effect on eating, in addition to its psychostimulantproperties. Studies in rats have demonstrated that the chronicinfusion of nicotine decreases both food intake and body-weightgain and that cessation of nicotine reverses the process.129–131In asystemic infusion study, we administered nicotine subcutaneouslythrough mini-osmotic pump to male rats at 5 mg/kg of body weightper day for 7 d and then stopped the infusion. Control rats receivedsaline. As shown in Figure 6, nicotine decreased food intakepredominantly through a decrease in meal number, with an inhib-itory effect on meal size. The hyperphagia of nicotine cessation ledto increased food intake predominantly through an increase inmeal size only.
Because nicotine alters food-intake and feeding patterns, nico-tine seems to influence the VMN and LHA because these nucleiaffect meal number and meal size, respectively. Both food intake(by an increase in meal size only) and daily body weight returnedto normal several days after cessation of the nicotine administra-tion.27 When the same study was conducted in female rats, wherethe fluctuation of daily food intake corresponds to the estrouscycle, nicotine decreased food intake through meal number, pre-serving meal size. A concomitant reduction in daily body weightalso occurred. From these data we conclude that nicotine has aprimary effect on the VMN in the female.
To further assess the affect of nicotine on LHA dopamine andserotonin, microdialysis of the LHA was performed after 2 d ofnicotine infusion and 2 d after the nicotine infusion was stopped.During the nicotine infusion, both dopamine and serotonin con-centrations were significantly elevated relative to their concentra-tions in the saline-infusion control group. After cessation of thenicotine infusion, dopamine concentration fell below that of con-trol levels. The infusion of nicotine into the male rat did not raiseplasma leptin concentrations.132 However, changes in vagal affer-ent stimuli were recorded when nicotine was injected into theportal vein. Below 1 ng, a decrease in vagal afferent stimulioccurred; above 10 ng, an increase was recorded. These datasuggest that some of the anorectic effects of nicotine are mediatedthrough peripheral activity.133
In contrast to that long-term study, short-term experimentswere conducted in which overnight food-deprived male ratsunderwent nicotine administration directly into the LHA for 60min by using a microdialysis (push and pull) technique. LHAdopamine, dihydroxyphenylacetic acid, serotonin, and 5-hydroxy-indoleacetic acid were measured by microdialysis before, during,and after LHA nicotine administration. Food was made availablefor 40 min beginning immediately after cessation of nicotine
Nutrition Volume 16, Number 10, 2000 851Dopamine and Serotonin in the Regulation of Food Intake
administration and the amount eaten was determined. Nicotineinfusion induced a consistent increase in dopamine and dihydroxy-phenylacetic acid during the first 40 min. A rise in serotonin alsooccurred, but the rise in serotonin persisted even after nicotineinfusion ceased. Also, eating induced a short-lasting increase indopamine and a long-lasting increase in serotonin. As expected,food intake during the infusion period was significantly lower inthe nicotine-treated group.134 Based on evidence showing thatserotonin facilitates the release of dopamine, we interpret thelong-lasting elevated LHA serotonin findings to be related tonicotine-induced hypophagia.
Both estradiol and nicotine are anorectic agents in decreasingfood intake and body weight. In their absence, such as duringmenopause, after ovariectomy, or after cessation of smoking, anincrease in food intake and body weight occurs. Both estradiol andnicotine individually influence hypothalamic dopamine and sero-tonin, whose interaction is also involved in food-intake and body-weight regulation. To explore their interrelation, we used ovariec-tomized (surgical menopause) and sham-operated rats thatreceived a continuous subcutaneous infusion of nicotine (5 mgzkg z d) through an osmotic minipump. On day 2 of nicotineinfusion, in vivo microdialysis using a double cannula, one in-serted into ipsilateral LHA and the other contralateral VMN, wasperformed to simultaneously measure dopamine and serotonin inawake, freely moving rats. Nicotine infusion decreased food intakeand body weight in both groups, with a greater decrease occurringin the ovariectomy group. An increase in LHA dopamine concen-tration occurred in ovariectomy group and an increase in VMNserotonin was observed in the sham-operated group. The data showthat a nicotine-induced decrease in food intake and body weightassociated with changes in hypothalamic monoamines in the pres-ence of estradiol (sham-operated rats) lowers food intake and bodyweight by an increase in serotonin concentration in the VMN; inovariectomized rats (menopause), nicotine lowers food intake andbody weight by an increase in dopamine concentration in theLHA.135
NPY is synthesized in arcuate neurons that project principallyonto the PVN. NPY injected into the PVN causes hyperphagia,reduced energy expenditure, and eventually induces obesity, andthese effects are opposed by nicotine. Nicotine (12 mgz kg z d) wasgiven to rats for either 1 or 12 d. One day of nicotine treatmentversus control significantly reduced food intake by 30% and sig-nificantly decreased body weight by 2%. NPY mRNA levels were
significantly reduced by 40% and NPY concentrations fell signif-icantly by 33% in the arcuate nucleus and PVN. Twelve days ofnicotine treatment significantly lowered cumulative food intake by8% and body weight by 10%. NPY and mRNA levels significantlyrose by 40%, whereas NPY levels again significantly decreased inthe arcuate nucleus and PVN. The results suggest that nicotinemay inhibit NPY synthesis in the hypothalamus, independentlyof any effects due to altered energy balance. Reduced activity ofNPYergic neurons in the projection from the arcuate nucleus tothe PVN contributes to the effects of nicotine on energybalance.136
A reasonable understanding of the mechanism underlying theregulation of meal size and meal number and thus food-intakecontrol has been achieved through studies of systemic and centralinfusion of nicotine into male and female rats. The mechanismincludes peripheral sensors, which provide intake feedback infor-mation to the central nervous system. In our studies the LHA andthe VMN modulate the production and release of dopamine andserotonin, which influences the motor effector systems to initiate,maintain, or terminate eating. Our findings indicate that chronicnicotine administration decreases food intake, which is mainlyachieved by a reduction in meal number, and this could be relatedto an increase in dopamine and serotonin activity in the LHA andthe VMN and by their interaction (Fig. 7).
ROLE OF DOPAMINE AND SEROTONIN IN RELATIONTO SEX AND FOOD INTAKE
Gonadal hormones are among the many factors affecting foodintake, feeding patterns, and body weight in rats. Hormonal effecton these measures are especially striking in females, in whom foodintake, body weight, and the indices of meal number and meal sizechange with the phase of the estrous cycle. Gonadal hormones alsoaffect the monoaminergic system. Ovarian hormones influencemonoamine release. In normal rats during estrous, serotonin re-lease was found to be decreased.137 Animal studies have demon-strated that estradiol and progesterone modulate functioning of thenigrostriatal dopaminergic system by raising the turnover of do-pamine by increasing the activity of tyrosine hydroxylase.138Ovar-ian hormones have a direct action on the hypothalamus by acti-vating the secretion of monoamines and of their release andturnover.139
FIG. 6. The effect of starting and stopping systematic nicotine infusion on food intake, meal number, and meal size.
852 Meguid et al. Nutrition Volume 16, Number 10, 2000

Food intake in the mature male rat is relatively stable.26,30,140
Temporally there is an increase in meal size and a decrease in mealnumber. In the female rat, meal number and meal size changecyclically and reciprocally during the estrus cycle.26 Specifically,during the proestrous phase of the estrous cycle, when serumestradiol titers are rising and are approaching their peak concen-tration, meal number peaks, with a decrease in the correspondingmeal size. During the metestrus/diestrus phase, when serum estra-diol titers are lowest, meal number decreases with a correspondingincrease in meal size. Although food intake per 100 g of bodyweight was similar in males and females, weight gain in males wasapproximately seven times greater.26
After orchiectomy (surgical removal of the testes), food intakein the orchiectomized rat decreased significantly in dark-phasemeal number and was not compensated for by increases in dark-phase meal size.30 There was a shift in the circadian distribution offood intake such that the rat ate more during the light phase. Afterhormone replacement these changes were reversed. After ovariec-tomy the estrous cycling disappeared, the relation of meal numberto meal size was altered, and there was an increase in daily foodintake.31 After ovariectomy daily food intake increased signifi-cantly in light-phase food intake, achieved by significant increasesin meal size. Daily hormone replacement (not given cyclically) inthe ovariectomized rat, with estradiol alone or in combination withprogesterone, normalized food intake. Mean body weight rapidlyincreased after ovariectomy and decreased after orchiectomy.
These studies show that gonadal hormones influence foodintake and that there are clear sex differences in food intake andfeeding pattern. Absence of gonadal hormones attenuates sexdifferences in circadian distribution in food intake. Alterations ofthe internal hormonal milieu affect the circadian rhythm of feedingand disrupt the feeding patterns. Because both male and femalerats showed an increase in light-phase feeding and in meal size anda decrease in meal number after castration, similar mechanism(s)are operative in both sexes. The exact mechanisms by whichgonadal hormones affect the neuroendocrine and biochemicalprocesses involved in food regulation are not clear, but bothcentral and peripheral systems are affected. Testosterone ismetabolized to estradiol and dihydrotestosterone (DHT) at var-ious sites including the VMN, which is believed to be a neuralsite regulating food intake. It is believed that aromatization oftestosterone to estrogens may be responsible for the centraleffects of testosterone on food intake.140
Because specific changes were observed in meal number andmeal size, whose product equals food intake, after removing thegonads in both sexes and because these individual indices appearto be differentially regulated by different hypothalamic sites,namely the VMN and LHA, we examined the role that VMN playsin the relation of meal number to meal size by analyzing feedingpatterns in both sexes after specific VMN block. The VMN wasblocked in age-matched young adult male and female rats byinjecting colchicine intrahypothalamically. After the VMNblock141 with colchicine, food intake significantly increased by60% in both dark and light phases. This was achieved as a resultof a significant 70% increase in meal number in both dark and lightphases, with no significant change in meal size. The female rat alsoincreased its food intake in both dark and light phases, with themagnitude of light-phase increase being much more than that ofthe dark phase (84% versus 26%). This was achieved by anincrease in the meal size in both the dark and light phases.
These results show a sexual dimorphism between male andfemale rats after VMN inhibition. After castration or in the absenceor impairment of the neural target site (VMN block), sex differ-ences in food intake are attenuated, but differences in the feedingindex by which the male and female rat increase total food intake,i.e., meal size in the female rat, meal number in the male rat, isprobably sex related and indicates that the significance of gonadalhormones is not only the regulation of food intake but also themechanisms by which regulation is achieved. Neuroanatomic dif-ferences in the morphology in the rat central nervous system exist1) in the preoptic area, the amygdala and VMN, neuronal nuclearand nucleolar size, 2) in the arcuate nucleus in synaptic vesiclesand terminals, 3) in the preoptic area in synaptic organization inthe suprachiasmatic nucleus, 4) in dendritic branching patterns inthe preoptic area, and 5) in the lumbar spinal cord (fifth and sixthsegments) in gross nuclear volume. Thus, neurons sensitive togonadal steroids seem to occupy prominent positions within neuralcircuits. Steroid-sensitive neurons can increase neurotransmitterbiosynthesis, decrease uptake or degradation, or enhance receptorsensitivity to neurotransmitters.142
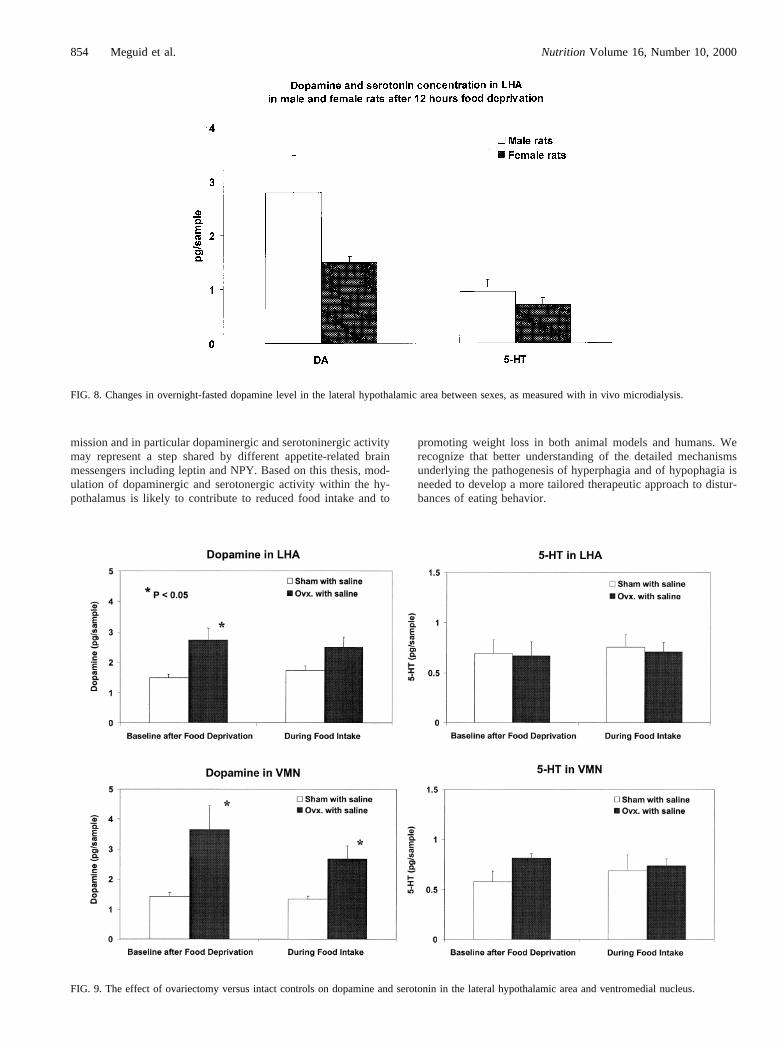
Using in vivo microdialysis, the intra-VMN and intra-LHAlevels of dopamine and serotonin were measured as they relate tothe control of meal size and meal number in normal male andfemale rats to examine their sex differences. After 12 h of fooddeprivation, normal intact male rats appeared to show higher LHAdopamine and serotonin levels than their normal intact femalecounterparts (Fig. 8). After ovariectomy, female rat VMN andLHA dopamine levels increased (Fig. 9) and the level of serotoninwas unchanged, although an increase was seen in the VMN.Whether sex differences exist in mRNA expression of the genesfor dopaminergic receptors in the VMN between male and femaleFischer rats in the normal state was also examined. In our intactnormal male rat, only the D2 receptor was expressed; in theproestrus female, both D1 and D2 receptors were expressed. Ourstudies show that there are sex differences in observed behaviorssuch as food intake because of sex differences in gene expression.In addition to morphologic differences in the hypothalamus, weshowed a functional difference, as documented by changes inhypothalamic dopamine and in their receptor expression. Thesechanges, influenced by gonadal hormones, translate into functionaldiversities characterized by different behaviors in each sex.
CONCLUSION
The control of food intake and energy expenditure is extremelycomplex and involves a number of different messengers.143 How-ever, different lines of investigations suggest that disturbances ofeating behavior are associated with abnormal hypothalamic neu-rotransmission, regardless of the signaling pathways involved intheir pathogenesis. This indicates that hypothalamic neurotrans-
FIG. 7. Schema showing interrelationship of the LHA and VMN, withnicotine and estrogen postulated to influence meal number and meal size.
Nutrition Volume 16, Number 10, 2000 853Dopamine and Serotonin in the Regulation of Food Intake
mission and in particular dopaminergic and serotoninergic activitymay represent a step shared by different appetite-related brainmessengers including leptin and NPY. Based on this thesis, mod-ulation of dopaminergic and serotonergic activity within the hy-pothalamus is likely to contribute to reduced food intake and to
promoting weight loss in both animal models and humans. Werecognize that better understanding of the detailed mechanismsunderlying the pathogenesis of hyperphagia and of hypophagia isneeded to develop a more tailored therapeutic approach to distur-bances of eating behavior.
FIG. 8. Changes in overnight-fasted dopamine level in the lateral hypothalamic area between sexes, as measured with in vivo microdialysis.
FIG. 9. The effect of ovariectomy versus intact controls on dopamine and serotonin in the lateral hypothalamic area and ventromedial nucleus.
854 Meguid et al. Nutrition Volume 16, Number 10, 2000

ACKNOWLEDGMENTS
Ms. Dawn Jensen-Nobile is thanked for her editorial assistance.
REFERENCES
1. NIH Consensus Statement. Methods for voluntary weight loss and control.Nutrition 1996;12:663
2. Flegal KM, Carrol MD, Kuczmarski RJ, Johnson CL. Overweight and obesityin the United States: prevalence and trends. 1960–1994. Int J Obes Relat MetabDisord 1998:22:39
3. Manson JE, Colditz GA, Stampfer MJ, et al. A prospective study of obesity andrisk of coronary heart disease in women. N Engl J Med 1990;322:882
4. Staessen J, Fagard R, Lijnen P, Amery A. Body weight, sodium intake andblood pressure. J Hypertens 1989;7(suppl):S19
5. Shaper AG, Wanamethee SG, Walker M. Body weight: implications for theprevention of coronary heart disease, stroke, and diabetes mellitus in a cohortstudy of middle aged men. Br Med J 1998;514:1311
6. Wolf AM, Colditz GA. Social and economic effects of body weight in theUnited States. Am J Clin Nutr 1996;63(suppl 3):466S
7. Seidell JC. Societal and personal costs of obesity. Exp Clin Endocrinol Diabetes1998;106(suppl 2):7
8. Spurr GB. Physical activity, nutritional status and physical work capacity inrelation to agricultural production. In: Pollitt E, Amante P, eds.Energy intakeand activity.New York: Alan R. Liss, 1984:207
9. Kennedy E, Garcia M. Body mass index and economic productivity. Eur J ClinNutr 1994;48(suppl 3):S45
10. Arora NS, Rochester DF. Respiratory muscle strength and maximal voluntaryventilation in undernourished patients. Am Rev Respir Dis 1982;126:5
11. Heymsfield SB, Bethel RA, Ansley JD, et al. Cardiac abnormalities in cachecticpatients before and during nutritional repletion. Am Heart J 1978;95:584
12. Landel AM, Lo C-C, Meguid MM. Observations on predicted brain influx ratesof neurotransmitter precursors: effects of tumor, operative stress with tumorremoval, and postoperative TPN of varying amino acid compositions. Cancer1987;59:1192
13. Chandra RK. The relation between immunology, nutrition and diseases inelderly people. Age Ageing 1990;19:S25
14. Watson RR, ed.Nutrition, disease resistance and immune function. New York:Marcel Dekker, 1984
15. Robinson C. Goldstein N, Levine GM. Impact of nutritional status on DRGlength of stay. JPEN 1987;11:49
16. Hill GD. Body composition research: implications for the practice of clinicalnutrition. JPEN 1998;16:197
17. Varma M, Sato T, Zhang L, Meguid MM. Space flight related anorexia. Lancet2000;356(9230):681
18. Stein TP, Leskiw JM, Schuluter MD, et al. Energy expenditure and balanceduring space flight on the Space Shuttle. Am J Physiol 1999;276:R1739
19. Levitsky DA, Collier G. Effects of diet and deprivation on meal eating behaviorin rats. Physiol Behav 1968;3:137
20. Becker EE, Kissileff HR. Inhibitory controls of feeding by the ventromedialhypothalamus. Am J Physiol 1974;226:383
21. Meguid M, Laviano A, Rossi-Fanelli F. Food intake equals meal size times mealnumber. Appetite 1998;31:404
22. Fettisov S, Meguid M, Shafiroff M, Miyata G, Torelli G. Dopamine in the VMNof the hypothalamus is important for diurnal distribution of eating in obese maleZucker rats. Nutrition 2000;16:65
23. Fetissov SO, Meguid MM, Miyata G, Torelli GF, Shafiroff M. VMN dopami-nergic graft and feeding pattern in obese Zucker rats. Int J Obes 2000;24:376
24. Meguid MM, Muscaritoli M, Beverly JL, et al. The early cancer anorexiaparadigm: changes in plasma free tryptophan and feeding indexes. JPEN 1992;16:56S
25. Meguid MM, Sato T, Torelli GF, Laviano A, Rossi-Fanelli F. An analysis oftemporal changes in meal number and meal size at onset of anorexia in maletumor-bearing rats. Nutrition 2000;16:305
26. Laviano A, Meguid MM, Gleason JR, Yang Z-J, Renvyle T. Comparison oflong-term feeding pattern between male and female Fischer 344 rats: influenceof estrous cycle. Am J Physiol 1996;270:R413
27. Miyata G, Meguid MM, Torelli GF, Fetissov S. Nicotine’s effect on hypotha-lamic neurotransmitters and appetite regulation. Surgery 1999;126:255
28. Meguid MM, Fetissov S, Miyata GF, Torelli G. Feeding pattern in obese Zuckerrats after dopaminergic and serotonergic LHA grafts. NeuroReport 1999;10:1049
29. Meguid MM, Fetissov SO, Blaha V, Yang ZJ. Dopamine and serotonin VMNrelease is related to feeding status in obese and lean Zucker rats. NeuroReport2000;11:2069
30. Chai J-K, Blaha V, Meguid MM, et al. Use of orchiectomy and testosteronereplacement to explore meal number to meal size relationship in male rats. Am JPhysiol 1999;45:R1366
31. Varma M, Chai J-K, Meguid MM, et al. Effect of estradiol and progresterone ondaily rhythm in food intake and feeding patterns in Fischer rats. Physiol Behav.1999;68:99
32. Meguid MM, Kawashima Y, Campos ACL, et al. Automated computerized rateater meter: description and application. Physiol Behav 1990;48:759
33. Hetherington AW, Ranson SW. Hypothalmic lesions and adiposity in the rat.Anat Rec 1940;78:149
34. Baskin DG, Figlewicz Lattemann D, Seeley RJ, et al. Insulin and leptin: dualadiposity signals to the brain for the regulation of food intake and body weight.Brain Res 1999;848:114
35. Arees EA, Mayer J. Anatomical connections between medial and lateral regionsof the hypothalamus concerned with food intake. Science 1967;157:1574
36. Morien A, Wellman J, Fojt J. Diurnal rhythms of paraventricular hypothalamicnorepinephrine and food intake in rats. Pharmacol Biochem Behav 1995;52:169
37. Lin L, York DA. Enterostatin actions in the amygdala and PVN to suppressfeeding in the rat. Peptides 1997;18:1341
38. Satoh N, Ogawa Y, Katsuura G, et al. The arcuate nucleus as a primary site ofsatiety effect of leptin in rats. Neurosci Lett 1997;224:149
39. Burlet A, Grouzmann E, Musse N, et al. The immunological impairment ofarcuate neuropeptide Y neurons by ricin A chain produces persistent decrease offood intake and body weight. Neuroscience 1995;66:151
40. Bellinger LL, Mendel VE, Bernardis LL, Castongua TW. Meal patterns of ratswith dorsomedial hypothalamic nuclei lesions or sham operations. PhysiolBehav 1986;36:693
41. Meguid MM, Yang Z-J, Laviano A. Meal size and number: relationship todopamine levels in the ventromedial hypothalamic nucleus. Am J Physiol1997;272:R1925
42. Meguid MM, Yang Z-J, Koseki M. Eating induced rise in LHA-dopaminecorrelates with meal size in normal and bulbectomized rats. Brain Res Bull1995;36:487
43. Bernardis LL, Bellinger LL. The lateral hypothalamic area revisited: ingestivebehavior. Neurosci Biobehav Rev 1996;20:189
44. Szczypka MS, Rainey MA, Palmiter RD. Dopamine is required for hyperphagiain Lep(ob/ob) mice. Nat Genet 2000;25:102
45. Giannakopoulos G, Galanopoulou P, Daifotis Z, Couvaris C. Effects of metu-lergine treatment on diet selection, brain serotonin (5-HT) and dopamine (DA)turnover in free feeding rats. Prog Neuropsychopharmacol Biol Psychiatry1998;22:803
46. Smith BK, York DA, Bray GA. Activation of hypothalamic serotonin receptorsreduced intake of dietary fat and protein but not carbohydrate. Am J Physiol1999;277(3, Pt 2):R802
47. Moore RY, Lenn NJ. A retinohypothalamic projection in the rat. J Comp Neurol1972;146:1
48. Moore RY. Organization and function of a central nervous system circadianoscillator: the suprachiasmatic hypothalamic nucleus. Fed Proc 1983;42:2783
49. Peschke D, Teichmann J, Peschke E. Pineal influence on annular nuclearvolume in ventromedial hypothalamic nucleus neurons of the male wistar rat.Chronobiol Int 2000;17:15
50. Choi SJ, Wong LS, Yamat C, Dallman MF. Hypothalmic ventromedial nucleusamplify circadian rhythms; do they contain a food entrained endogenous oscil-lator. J Neurosci 1988;18:3843
51. Fetissov SO, Meguid MM, Chen C, Miyata G. Synchronized release of dopa-mine and serotonin in the medial and lateral hypothalamus of rats. Neuroscience2000(in press)
52. Stephan FK, Davidson AJ. Glucose but not fat, phase shifts the feeding en-trained circadian clock. Physiol Behav 1998;65:277
53. Levin BE, Routh VH. Role of brain in energy balance and obesity. Am J Physiol1996;271:R491
54. Bray GA. hypothalamic and genetic obesity in experimental animals. An auto-nomic and endocrine hypothesis. Physiol Rev 1979;59:719
55. Bellinger LL, Bernadis LL, Mendel VE. Effect of ventromedial and dorsomedialhypothalamic lesions on circadian corticosterone rhythms. Neuroendocrinology1976;22:216
56. Egawa M, Inoue S, Satoh S, et al. Acute and chronic effects of VMH lesions oncircadian rhythms in food intake and metabolites. Brain Res Bull 1993;31:293
57. Grignaschi G, Fanelli E, Scagnol I, Samanin R. Studies on the role of serotoninreceptor subtypes in the effect of sibutramine in various feeding paradigms inrats. Br J Pharmacol 1999;127:1190
58. Leibowitz SF, Rossakis C. Mapping study of brain dopamine and epinephrine-sensitive sites which cause feeding suppression in the rats. Brain Res 1979;172:101
59. Leibowitz SF. Brain monoamines and peptides: role in the control of eatingbehavior. Fed Proc 1986;45:1396
Nutrition Volume 16, Number 10, 2000 855Dopamine and Serotonin in the Regulation of Food Intake

60. Leibowitz SF, Alexander JT, Cheung WK, Weiss GF. Effects of 5HT and 5HTblocker metergoline on meal patterns & macronutrient selection. PharmacolBiochem Behav 1993;45:185
61. Mori RC, Guimaraes RB, Nascimento CM, Ribeiro EB. Lateral hypothalamicserotonergic responsiveness to food intake in rat obesity as measured bymicrodialysis. Can J Physiol Pharmacol 1999;77:286
62. Rouch C, Nicolaidis S, Orosco M. Determination, using microdialysis, ofhypothalamic serotonin variations in response to different macronutrients.Physiol Behav 1999;65:653
63. Zhou Q-Y, Palmiter RD. Dopamine-deficient mice are severely hypoactive,adipsic and aphagic. Cell 1995;83:1197
64. Szczypka MS, Mandel RJ, Donahue BA, et al. Viral gene delivery selectivelyrestores feeding and prevents lethality of dopamine-deficient mice. Neuron1999;22:167
65. Salamone JD, Cousins MS, Snyder BJ. Behavioral functions of nucleus accum-bens dopamine: empirical and conceptual problems with the anhedonia hypoth-esis. Neurosci Biobehav Rev 1997;21:341
66. Orosco M, Rouch C, Meile MJ, Nicolaidis S. Spontaneous feeding-relatedmonoamine changes in rostromedial hypothalamus of the obese Zucker rat: amicrodialysis study. Physiol Behav 1995;57:1103
67. Lauder JM. Ontogeny of the serotonergic system in the rat. Serotonin as adevelopmental signal. Ann NY Acad Sci 1990;600:297
68. Whitaker-Azmitia PM. Role of serotonin and other neurotransmitter receptors inbrain development basis for developmental pharmacology. Pharmacol Rev1990;43:553
69. Nonogaki K, Strack AM, Dallman MF, Tecott LH. Leptin-independent hy-perphagia and type 2 diabetes in mice with a mutated serotonin 5-HT2C receptorgene. Nat Med 1998;4:1152
70. Schwartz DH, McClane S, Hernandez L, Hoebel BG. Feeding increases extra-cellular serotonin in the lateral hypothalamus of the rat as measured by micro-dialysis. Brain Res 1989;479:349
71. Hoebel BG, Hernandez L, Schwartz DH, et al. Microdialysis studies of the brainnorepinephrine, serotonin and dopamine release during ingestive behavior:theoretical and clinical implications. Ann NY Acad Sci 1989;575:171
72. Fetissov S, Meguid MM, Zhang LH, Sato T. Catabolic effect of 5HT in themedial hypothalamus is diminished in leptin resistant obese rats. Brain Res Bull2000(in press)
73. West AR, Galloway MP. Regulation of serotonin-facilitated dopamine release invivo: the role of protein kinase A activating transduction mechanisms. Synapse1996;23:20
74. Brunetti L, Michelotto B, Orlando G, Vacca M. Leptin inhibits norepinephrineand dopamine release from rat hypothalamic neuronal endings. Eur J Pharmacol1999;372:237
75. Miyata G, Meguid MM, Torelli GF, Fetisov S. A complication of long-termbrain microdialysis. Nutrition 1999;15:723
76. Yang Z-J, Koseki M, Meguid MM, Laviano A. Eating-related increase ofdopamine concentration in the LHA with oronasal stimulation. Am J Physiol1996;270:R315
77. Yang Z-J, Meguid MM, Koseki M, et al. Increased food intake and body weightgain after lateral hypothalamic dopaminergic cell implantation. NeuroReport1996;7:449
78. Yang Z-J, Meguid MM. LHA-dopaminergic activity in obese and lean Zuckerrats. NeuroReport 1995;6:1191
79. Yang Z-J, Meguid MM, Oler A. Eating-associated VMN-dopamine levels ofrats: comparison of oral and intragastric feeding. NeuroReport 1997;8:1543
80. Orosco M, Trouvin JH, Cohen Y, Jocquot C. Ontogeny of brain monoamines inlean and obese female Zucker rats. Physiol Behav 1986;36:853
81. Yang Z-J, Meguid MM, Chai J-K, Chen C, Oler A. Bilateral hypothalamicdopamine infusion in male Zucker rat suppresses feeding due to reduced mealsize. Pharmacol Biochem Behav. 1997;58:631
82. Fetissov S, Torelli GF, Meguid MM, Miyata G, Varma M. Effects of implan-tation of dopaminergic neurons into the ventromedial nucleus of the hypothal-amus. Surg Forum 1998;49:496
83. Le Magnen J, Devos M. Metabolic correlates of the meal onset in the free foodintake of rats. Physiol Behav 1970;5:805
84. Rada P, Mark GP, Hoebel BG. Galanin in the hypothalamus raises dopamineand lowers acetylcholine release in the nucleus accumbens: a possible mecha-nism for hypothalamic initiation of feeding behavior. Brain Res 1998;798:1
85. Leibowitz SF. The role of serotonin in eating disorders. Drugs 1990;39(suppl3):33
86. Jung RT. Drug treatment of obesity: general principles and current therapies. In:Kopelman PG, Stock MJ, eds.Clinical obesity. London: Blackwell Science,1998:469
87. Bray GA, Tartaglia LA. Medicinal strategies in the treatment of obesity. Nature2000;404:672
88. Tagliamonte A, Biggio G, Vargiu L, Gessa GL. Free tryptophan in serumcontrols brain tryptophan levels and serotonin synthesis. Life Sci 1973;12:277
89. Duffy J. The lessons of eosinophilia-myalgia syndrome. Hosp Pract 1992;30:6590. Ceci F, Cangiano C, Cairella M, et al. The effects of oral 5-hydroxytryptophan
administration on feeding behavior in obese adult female subjects. J NeuralTransm 1989;76:109
91. Cangiano C, Ceci F, Cascino A, et al. Eating behavior and adherence to dietaryprescriptions in obese adult subjects treated with 5-hydroxytryptophan. Am JClin Nutr 1992;56:86
92. Cangiano C, Laviano A, Del Ben M, et al. Effects of oral 5-hdroxy-tryptophanon energy intake and macronutrient selection in non-insulin dependent diabeticpatients. Int J Obes Relat Metab Disord 1998;22:648
93. Cangiano C, Laviano A, Meguid MM, et al. Effects of administration of oralbranched-chain amino acids onanorexia and caloric intake in cancer patients.J Natl Cancer Inst 1996;88:550
94. Chai J-K, Varma M, Meguid MM, Torelli GF, Yang Z-J. Estradiol modulatestumor-induced anorexia and feeding pattern. Surg Forum 1998;49:479
95. Varma M, Torelli GF, Meguid MM, et al. Potential strategies for amelioratingearly cancer anorexia. J Surg Res 1999;81:69
96. Schro¨der J, Kahlke V, Staubach KH, Zabel P, Stuber F. Gender differences inhuman sepsis. Arch Surg 1998;133:1200
97. Yang ZJ, Koseki M, Meguid MM, Gleason JR, Debonis D. Synergistic effect ofrhTNF-a and rhIL-1a in inducing anorexia in rats. Am J Physiol 1994;267:R1056
98. Debonis D, Meguid MM, Laviano A, Yang ZJ, Gleason JR. Temporal changesin meal number and meal size relationship in response to rHu IL-1a. Neuro-Report 1995;6:1752
99. Opara EI, Laviano A, Meguid MM, Yang ZJ. Correlation between food intakeand CSF IL-1a in anorectic tumor bearing rats. NeuroReport 1995;6:750
100. Gayle D, Ilyin SE, Plata-Salaman CR. Central nervous system IL-1 beta systemand neuropeptide Y mRNAs during IL-1 induced anorexia in rats. Brain ResBull 1997;44:311
101. Laviano A, Renvyle T, Meguid MM, et al. Relationship between interleukin-1and cancer anorexia. Nutrition 1995;11:680
102. Torelli GF, Meguid MM, Moldawer LL, et al. Use of recombinant humansoluble TNF receptor in anorectic tumor-bearing rats. Am J Physiol 1999;277:R850
103. Plata-Salaman CR, Oomura Y, Kai Y. Tumor necrosis factor and interleukin 1beta: suppression of food intake by direct action in the central nervous system.Brain Res 1988;448:106
104. Yang ZJ, Blaha V, Meguid MM, Oler A, Zadak Z. Interleukin-1 injection intoventromedial hypothalamic nucleus depress food intake and increases release ofdopamine and 5-HT. Pharmacol Biochem Behav 1999;62:61
105. Krause R, James JH, Ziparo V, Fischer JE. Brain tryptophan and the neoplasticanorexia-cachexia syndrome. Cancer 1979;44:1003
106. Blaha V, Yang ZJ, Meguid MM, et al. Ventromedial nucleus of hypothalamusis related to the development of cancer induced anorexia: in vivo microdialysisstudy. Acta Med (Hradec Kralove) 1998;41:3
107. Chance WT, Balasubramaniam A, Dayal R, Brown J, Fischer JE. Hypothalamicconcentration and release of neuropeptide Y into microdialysates is reduced inanorectic tumor-bearing rats. Life Sci 1994;54:1869
108. McCarthy HD, McKibbin PE, Perkins AV, Linton EA, Williams G. Alterationsin hypothalamic NPY and CRF in anorexic tumor-bearing rats. Am J Physiol1993;264:E638
109. Inui A. Cancer anorexia-cachexia syndrome. Are neuropeptides the key? CancerRes 1999;59:4493
110. Rossi-Fanelli F, Cangiano C, Ceci F, et al. Plasma tryptophan and anorexia inhuman cancer. Eur J Clin Oncol 1986;22:89
111. Leibowitz SF, Weiss GF, Suh JS. Medial hypothalamic nuclei mediate seroto-nin’s inhibitory effect on feeding behavior. Pharmacol Biochem Behav 1990;37:735
112. Venero JL, Herrera AJ, Machado A, Cano J. Changes in neurotransmitter levelsassociated with the deficiency of some essential amino acids in the diet. Br JNutr 1992;68:409
113. Muscaritoli M, Meguid MM, Beverly JL, et al. Mechanism of early tumoranorexia. J Surg Res 1996;60:389
114. Cangiano C, Cascio A, Ceci F, et al. Plasma and CSF tryptophan and anorexiain cancer anorexia. J Neural Transm 1990;81:225
115. Pe´rez-Cruet J, Chase TN, Murphy DL. Dietary regulation of brain tryptophanmetabolism by plasma ratio of free tryptophan and neutral amino acids inhumans. Nature 1974;248:693
116. Dunn AJ. Endotoxin-induced activation of cerebral catecholamine and serotoninmetabolism: comparison with interleukin-1. J Pharmacol Exp Ther 1990;261:964
856 Meguid et al. Nutrition Volume 16, Number 10, 2000

117. Cangiano C, Testa U, Muscaritoli M, et al. Cytokines, tryptophan and anorexiain cancer patients before and after surgical tumor ablation. Anticancer Res1994;14:1451
118. Thorre´ K, Sarre S, Smolders I, Ebinger G, Michotte Y. Dopaminergic regulationof serotonin release in the substantia nigra of the freely moving rat usingmicrodialysis. Brain Res 1998;796:107
119. Leibowitz SF, Rossakis C. Pharmacological characterization of perifornicalhypothalamic dopamine receptors mediating feeding inhibition in the rat. BrainRes 1979;172:115
120. Weiner DM, Levey AI, Sunahara RK, et al. D1 and D2 dopamine receptormRNA in rat brain. Proc Natl Acad Sci USA 1991;88:1859
121. Grunfeld C, Zhao C, Fuller J, et al. Endotoxin and cytokines induce expressionof leptin, the ob gene product, in hamsters. J Clin Invest 1996;97:2152
122. Sato T, Meguid MM, Fetissov S, et al. Leptin contributes to progression oftumor-induced anorexia in rats. Surg Forum 1999;50:354
123. Chance WT, Sheriff S, Moore J, Peng F, Balasubramaniam A. Reciprocalchanges in hypothalamic receptor binding and circulating leptin in anorectictumor-bearing rats. Brain Res 1998;803:27
124. Simons JP, Schols AM, Campfield LA, Wouters EF, Saris WH. Plasma con-centration of total leptin and human lung-cancer-associated cachexia. Clin Sci1997;93:272
125. Erickson JC, Clegg KE, Palmiter RD. Sensitivity to leptin and susceptibility toseizures of mice lacking neuropeptide Y. Nature 1996;381:415
126. Laviano A, Gleason JR, Meguid MM, et al. Effects of Intra-VMN mianserin andIL-1ra on meal number in anorectic tumor-bearing rats. J Invest Med 2000;48:40
127. Centers for Disease Control.The health consequences of smoking: nicotineaddiction—a report of the Surgeon General. DHHS Publication no. 8808406.Rockville, MD: U.S. Department of Health and Human Services, Public HealthService, CDC, 1988
128. Wack JT, Rodin J. Smoking and its effects on body weight and the systems ofcaloric regulation. Am J Clin Nutr 1982;35:366
129. Grunberg NE, Bowen DJ, Winders SE. Effects of nicotine on body weight andfood consumption in female rats. Psychopharmacology 1986;90:101
130. Levin ED, Morgan MM, Galvez C, Ellison GD. Chronic nicotine and with-
drawal effects on body weight and food and water consumption in female rats.Physiol Behav 1987;39:441
131. Morgan MM, Ellison GD. Different effects of chronic nicotine treatment regi-mens on body weight and tolerance in the rat. Psychopharmacology 1987;91:236
132. Miyata G, Meguid MM. Is leptin involved in the acute anorectic effect ofnicotine? Nutrition 2000;16:141
133. Miyata G, Niijima A, Meguid MM, Yang ZJ. Hepatic involvement wherebynicotine induces its anorectic effect. Surg Forum 1999;50:262
134. Yang ZJ, Blaha V, Meguid MM, et al. Infusion of nicotine into the LHAenhances dopamine and 5-HT release and suppresses food intake. PharmacolBiochem Behav 1999;64:155
135. Zhang L, Varma M, Meguid MM. Changes in hypothalamic monoamines withnicotine in menopausal females: implications in food intake and body weightregulation. Surg Forum 2000;L1:545
136. Frankish HM, Dryden S, Wang Q, et al. Nicotine administration reducesneuropeptide Y and neuropeptide Y mRNA concentrations in the rat hypothal-amus: NPY may mediate nicotine’s effects on energy balance. Brain Res1995;694:139
137. Gundlah C, Simon L, Auerbach S. Differences in hypothalamic serotoninbetween estrous phases and in vivo microdialysis study. Brain Res 1998;785:91
138. Bedard PJ, di Paolo T, Langelier P, Poyet P, Labrie F. Behavioral and biochem-ical evidence of an effect of estradiol on striatal dopamine receptors. In: Fuxe K,Gustafsson J-A, Welterberg L, eds.Steroid hormone regulation of the brain.Oxford: Pergamon Press, 1981:331
139. Salmanca S, Uphouse L. Estradiol modulation of the rat hyperphagia induced bythe 5 HT1A agonist, 8 OH-DPAT. Pharmacol Biochem Behav1992;43:953
140. Nunez AA, Siegel LI, Wade GN. Central effects of testosterone on food intakein male rats. Physiol Behav 1980;24:469
141. Varma M, Laviano A, Meguid MM, et al. Comparison of early feeding patterndynamics in female and male Fischer rats after reversible VMN Block. J InvestMed 2000(in press)
142. McEwen S. Neural gonadal steroid action. Science 1981;211:1301143. Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG. Central nervous
system control of food intake. Nature 2000;404:661
Nutrition Volume 16, Number 10, 2000 857Dopamine and Serotonin in the Regulation of Food Intake
Related Documents











