http://het.sagepub.com/ Human & Experimental Toxicology http://het.sagepub.com/content/early/2012/01/09/0960327111432500 The online version of this article can be found at: DOI: 10.1177/0960327111432500 published online 12 January 2012 Hum Exp Toxicol Pant and S. Jahan M A. Siddiqui, V. Kumar, M. P. Kashyap, M. Agarwal, A. K. Singh, V. K. Khanna, A. A. Al-Khedhairy, J. Musarrat, A. B. Short-term exposure of 4-hydroxynonenal induces mitochondria-mediated apoptosis in PC12 cells Published by: http://www.sagepublications.com can be found at: Human & Experimental Toxicology Additional services and information for http://het.sagepub.com/cgi/alerts Email Alerts: http://het.sagepub.com/subscriptions Subscriptions: http://www.sagepub.com/journalsReprints.nav Reprints: http://www.sagepub.com/journalsPermissions.nav Permissions: What is This? - Jan 12, 2012 Proof >> at UNIV OF LOUISVILLE on January 14, 2012 het.sagepub.com Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

http://het.sagepub.com/Human & Experimental Toxicology
http://het.sagepub.com/content/early/2012/01/09/0960327111432500The online version of this article can be found at:
DOI: 10.1177/0960327111432500
published online 12 January 2012Hum Exp ToxicolPant and S. Jahan
M A. Siddiqui, V. Kumar, M. P. Kashyap, M. Agarwal, A. K. Singh, V. K. Khanna, A. A. Al-Khedhairy, J. Musarrat, A. B.Short-term exposure of 4-hydroxynonenal induces mitochondria-mediated apoptosis in PC12 cells
Published by:
http://www.sagepublications.com
can be found at:Human & Experimental ToxicologyAdditional services and information for
http://het.sagepub.com/cgi/alertsEmail Alerts:
http://het.sagepub.com/subscriptionsSubscriptions:
http://www.sagepub.com/journalsReprints.navReprints:
http://www.sagepub.com/journalsPermissions.navPermissions:
What is This?
- Jan 12, 2012Proof >>
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

Original Paper
Short-term exposure of4-hydroxynonenal inducesmitochondria-mediated apoptosisin PC12 cells
MA Siddiqui1, V Kumar2, MP Kashyap2, M Agarwal2,AK Singh2, VK Khanna2, AA Al-Khedhairy1, J Musarrat1,AB Pant2 and S Jahan2
Abstract4-Hydroxynonenal (4-HNE) is one of the most reactive aldehydic by-products of lipid peroxidation. Therole of 4-HNE in the etiology of various neurodegenerative disorders including cerebral ischemia/reper-fusion, Alzheimer’s disease, Parkinson’s disease, etc. has been documented. We and others have reportedthat long-term toxic insults of 4-HNE triggers apoptotic signals and oxidative stress in various cells.However, the status of apoptosis following short-term exposure and underlying mechanisms has not beenexplored so far. We studied the apoptotic changes in PC12 cells receiving short-term exposure of4-HNE. A significant dose-dependent induction in reactive oxygen species (ROS) and early responsemarkers (c-Fos, c-Jun, and GAP-43) were observed in cells exposed to 4-HNE (10, 25, and 50 mM) for1h. Following the exposure of PC12 cells to 4-HNE, the levels of protein and messenger RNA expres-sions of P53, Bax, and caspase 3 were significantly upregulated, whereas the levels of Bcl2 was downre-gulated. We could record the apoptotic signals and ROS generation in PC12 cells receiving 4-HNEexposure for such a short period of time. Induction in the expression and activity of caspase 3 has alsoindicated the mitochondrial mediation in the apoptosis induction.
KeywordsPC12 cells, 4-hydroxynonenal, reactive oxygen species (ROS), apoptosis
Introduction
Aldehydic products of lipid peroxidation of biological
membranes have been reported as important etiologi-
cal factors in numerous neurodegenerative disor-
ders.1–3 Of these, 4-hydroxynonenal (4-HNE), a
long-chain a, b-unsaturated aldehyde, is known to
be most toxic.4,5 Higher concentrations of 4-HNE are
cytotoxic and causes oxidative stress-mediated cell
death in a variety of cell types including PC12
cells.6–9 The low-level exposure of 4-HNE also
modulates intracellular signaling by activating the
mitogen activated protein kinases (MAPK), stress-
activated protein kinase and c-Jun N-terminal protein
kinase cascades, and inhibiting the nuclear factor k-B
activity.7,10,11 We and others have shown the associa-
tion of 4-HNE-induced oxidative stress-mediated
cytotoxicity/genotoxicity with dopamine (DA-D2),
cholinergic (muscarinic), benzodiazepine, and seroto-
nin (5-HT)-2A receptors in cultured cells of human
and animal origins.8,12 The expression of GSTP1-1
and increased levels of intracellular Caþþ are also
associated with 4-HNE exposure in cells.9
1 Department of Zoology, College of Science, King SaudUniversity, Riyadh, Saudi Arabia2 In Vitro Toxicology Laboratory, Indian Institute of ToxicologyResearch, Lucknow, India
Corresponding author:AB Pant, In Vitro Toxicology Laboratory, Indian Institute ofToxicology Research, PO Box 80, MG Marg, Lucknow 226001,Uttar Pradesh, IndiaEmail: [email protected]; [email protected]
Human and Experimental Toxicology1–10
ª The Author(s) 2011Reprints and permission:
sagepub.co.uk/journalsPermissions.navDOI: 10.1177/0960327111432500
het.sagepub.com
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

In our earlier studies, a concentration-dependent
metabolism of 4-HNE in PC12 cells was observed in
the form of glutathionyl conjugates and 4-Hydroxy-
trans-2-nonenoic acid (HNA) and 1,4-Dihydroxy-2-
nonene (DHN) metabolites within 1 h of exposure.13
Although, 4-HNE has been reported to induce apopto-
sis14–16 and necrosis,17–19 the underlying mechanisms
following short-term exposure of 4-HNE is not known.
Thus, the present study was carried out to test whether
the expression of selected apoptosis markers in PC12
cells, receiving short-term exposure of 4-HNE is
altered. Our results show significant dose-dependent
alterations in reactive oxygen species (ROS), and early
responses to specific apoptotic markers in cells exposed
to 4-HNE. Furthermore, enhanced expression and activ-
ity of caspase 3 following 4-HNE exposure to cells indi-
cates the mitochondrial mediation in the apoptosis
induction. Together, the results from these studies may
provide a mechanism through which 4-HNE induces
apoptosis in neuronal cells.
Materials and methods
Cell culture
PC12 cells used in the study were originally procured
from National Centre for Cell Sciences, Pune, India,
and grown in Nutrient Mixture/F-12 (Hams) supple-
mented with 2.5% fetal bovine serum (FBS), 15% horse
serum, 0.2% sodium bicarbonate, and antibiotic/anti-
mycotic solution (100�, 1 ml/100 ml of medium;
Invitrogen, Life technologies, Staley Road, Grand
Island, NY 14072, USA). The cells were maintained
in 5% CO2–95% atmosphere under high humidity at
37�C. Cells were assessed for cell viability by trypan
blue dye exclusion assay as described earlier by Pant
et al.20 and batches showing viability of more than
95% were used in the experiments.
Reagents and consumables
All the specified chemicals, reagents, and diagnostic
kits were procured from Sigma Chemical Company
Pvt. Ltd. (St. Louis, Missouri, USA), unless otherwise
stated. Nutrient mixture F-12 Hams, antibiotics/anti-
mycotics solution (100�), FBS (Staley Road, Grand
Island, NY, USA), and horse serum were purchased
from Gibco BRL (Kamstrupvej 90, Denmark-4000,
Roskilde, USA). Culture wares and other plastic
consumables used in the study were procured from
Nunc (Denmark). Milli-Q water was used in all the
experiments. 4-HNE, an unsaturated aldehyde, was
generously gifted by Dr Sanjay Srivastava, Depart-
ment of Cardiology, University of Louisville (Ken-
tucky, USA).
Experimental design
PC12 cells were exposed to various concentrations
(1–50 mM) of 4-HNE for 1 h. Cells receiving 4-
HNE insult were then analyzed for ROS generation
and to determine the changes in the expression of
early response proteins (c-Fos, c-Jun, and GAP-43)
as well as selected marker genes associated with
mitochondria-mediated apoptosis using real time-
polymerase chain reaction (RT-PCR). The possible
associations of these proteins in 4-HNE-induced
apoptosis were also assessed using immunoblot–
Western blot analysis.
ROS generation
ROS generation was assessed using 2,7-
dichlorodihydrofluorescein diacetate (DCFH-DA;
Sigma Aldrich (St. Louis, Missouri, USA), USA) dye
as a fluorescence agent following the protocol earlier
described by us.21 In brief, cells (5 � 104 per well)
were allowed to adhere to poly-L-lysine-coated
eight-well chamber slide flasks. Following the expo-
sure of 4-HNE for 1 h, cells were washed with
phosphate-buffered saline (PBS) and incubated for
the next 30 min in DCFH-DA (20 mM) containing
incomplete culture medium in the dark. Then, the
slides were analyzed for intracellular fluorescence
using an upright fluorescence microscope (Nikon
Eclipse 80i (Yokohama, Japan) equipped with Nikon
DS-Ri1 12.7 megapixel camera). Image analysis soft-
ware Leica Qwin 500 (Leica Microsystem, Wetzlar,
Germany) was used to quantify the change in the
fluorescence and data were expressed as percentage
change in the fluorescence area.
Transcriptional changes
Alterations in the messenger RNA (mRNA) expres-
sion of apoptosis marker genes (P53, Bax, Bcl2, and
caspase 3) were studied following the protocol as
described earlier by us.21,22 In brief, cells (1 � 106)
were allowed to grow in six-well culture plates unless
reached to 80–85% confluence. Following the expo-
sure of cells to 4-HNE for 1 h, total RNA was isolated
from both experimental and control sets using GeneE-
lute mammalian total RNA Miniprep Kit (catalog no.
RTN-70, Sigma). The purity and yield of RNA was
2 Human and Experimental Toxicology
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

assessed by Nanodrop ND-1000 Spectrophotometer
V3.3 (Nanodrop Technologies Inc., Wilmington, Dela-
ware, USA). RNA quality in terms of integrity was also
checked by running RNA onto 2% denaturing agarose
gel. Total RNA (2 mg) was reverse-transcribed into
cDNA by SuperScript III first-strand cDNA synthesis
Kit (catalog no. 18080-051, Invitrogen Life Science,
USA). qRT-PCR was performed by SYBR Green dye
(ABI, Applied Biosystem, 850 Lincoln Centre Drive
Foster City California, USA) using ABI PRISM1
7900HT Sequence Detection System (Applied Biosys-
tems, USA). Real-time reactions were carried out in tri-
plicate. Specificity of primer sets and genomic DNA
contamination were assessed for all the samples by ana-
lyzing the melting curve and no template control
(NTCs), respectively. GAPDH was used as internal con-
trol to normalize the data. 4-HNE-induced alterations in
mRNA expression were expressed in terms of relative
quantity.
Protein expression (Western blotting)
Alterations in the expression of selected marker pro-
teins (P53, Bax, Bcl2, and activated caspase 3) associ-
ated with apoptosis were carried out by Western
immunoblotting as described earlier.16 In brief, after
respective exposure, cells were washed twice with
ice-cold PBS (pH 7.4) and centrifuged at 1000 rpm for
10 min. Cell pellets were lysed using cell lysis reagent
(catalog no. C2978, Sigma) in the presence of 1� pro-
tein inhibitor cocktail (catalog no. P8340; Sigma). An
equal amount (50 mg/well) of proteins was loaded in
10% Tricine–sodium dodecyl sulfate gel and electro-
phoresis was carried out. Following electrophoresis,
protein bands from the gel were transferred on polyvi-
nylidene fluoride membrane (Millipore, Bedford,
Massachusetts, USA) in an electrophoresis transfer
apparatus (BioRad, Hercules, California, USA). The
membrane was blocked overnight in Tris-Buffered
Saline Tween-20 (TBST) (30 mM Tris, pH 7.5,
150 mM NaCl, and 0.1% Tween 20) containing 5%nonfat milk. Membranes were then probed with pri-
mary antibodies specific for Bax (1:500, Santa Cruz,
USA), Bcl2, activated caspase 3 (1:1000, Cell Signal-
ing Technology (CST), USA), and b-actin (1:2000,
Santa Cruz). After several washings with TBST, mem-
brane was incubated with horseradish peroxidase-
conjugated secondary antibody (goat anti-rabbit for
Bax and cleaved-caspase 3, goat anti-mouse for all
others) for 2 h at room temperature. Then, the blots
were developed using 3,30,5,50-tetramethylbenzidine
(TMB)–hydrogen peroxide (H2O2; Sigma Weltevreden
Park, Johannesburg, 1715, S.A.). Pictures of specific
band were taken by gel documentation system
(Alpha Innotech, USA) and densitometric analysis
was done by AlphaEase� FC StandAlone V. 4.0.0
software.
Protein expression (Immunocytochemistry)
Immunocytochemical localization for early response
gene proteins (c-Fos, c-Jun, and GAP-43) was carried
out using specific antibodies following the protocol.22
In brief, 4-HNE exposed cells were fixed in 4%paraformaldehyde for 10 min. Cells were washed
with PBS thrice and incubated with 0.5% H2O2
(w/v) in methanol to quench endogenous peroxidase.
Nonspecific binding sites were blocked by incubating
cells with 0.5% bovine serum albumin and
0.1% Triton X-100 in phosphate buffer saline (PBS)
for 2 h. Cells were then incubated in primary mono-
clonal antibodies specific for c-Fos, c-Jun, and Gap-
43 proteins (1:200, Sigma Chemicals Company Pvt.
Ltd.) for 1 h. Following the washing with PBS, cells
were again reincubated with goat anti-rabbit horse-
radish peroxidase conjugate secondary antibody
(1:500) for 2 h. Cells were washed with PBS to
remove any unbound secondary antibody and incu-
bated with diaminobenzidine hydrochloride for 5–
15 min to develop the color. Cells were visualized and
images were captured using an upright microscope
(Nikon Eclipse 80i equipped with Nikon DS-Ri1
12.7 megapixel camera). Image analysis software
Leica Qwin 500 was used to quantify the percentage
change in the area of expression of protein early
response proteins. Similar experiments in cells with-
out any exposure were run under parallel and served
as controls.
Caspase 3 activity
4-HNE-induced alterations in the activity of caspase 3
were monitored using kits (Biovision, catalog no.
K106, USA). Following 4-HNE exposure for 1 h,
cells were pelleted, resuspended in prechilled extrac-
tion buffer (50 ml), and incubated for 10 min on ice.
Then, the samples were centrifuged for 5 min at 500g
and the clear supernatant (50 ml per well) transferred
to 96-well culture plates. Assay buffer (50 ml) and
substrate conjugate (5 ml) were added and mixed
well. Immediately after the completion of 2 h incu-
bation at 37�C in dark, the contents were mixed
thoroughly and read for absorbance at 400 nm. The
Siddiqui MA et al. 3
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from
values of exposed groups were compared with
unexposed control sets and the data expressed as
a percentage of control.
Statistical analysis
Results are expressed as mean + standard error
(SE) of the values obtained from three independent
experiments, and in each experiment, a minimum
of four replicates were used. Statistical analysis
was performed using one-way analysis of variance,
and post hoc Dunnett’s test was applied to compare
values between control and treated groups. The
values of p < 0.05 were considered as statistically
significant.
Results
ROS generation
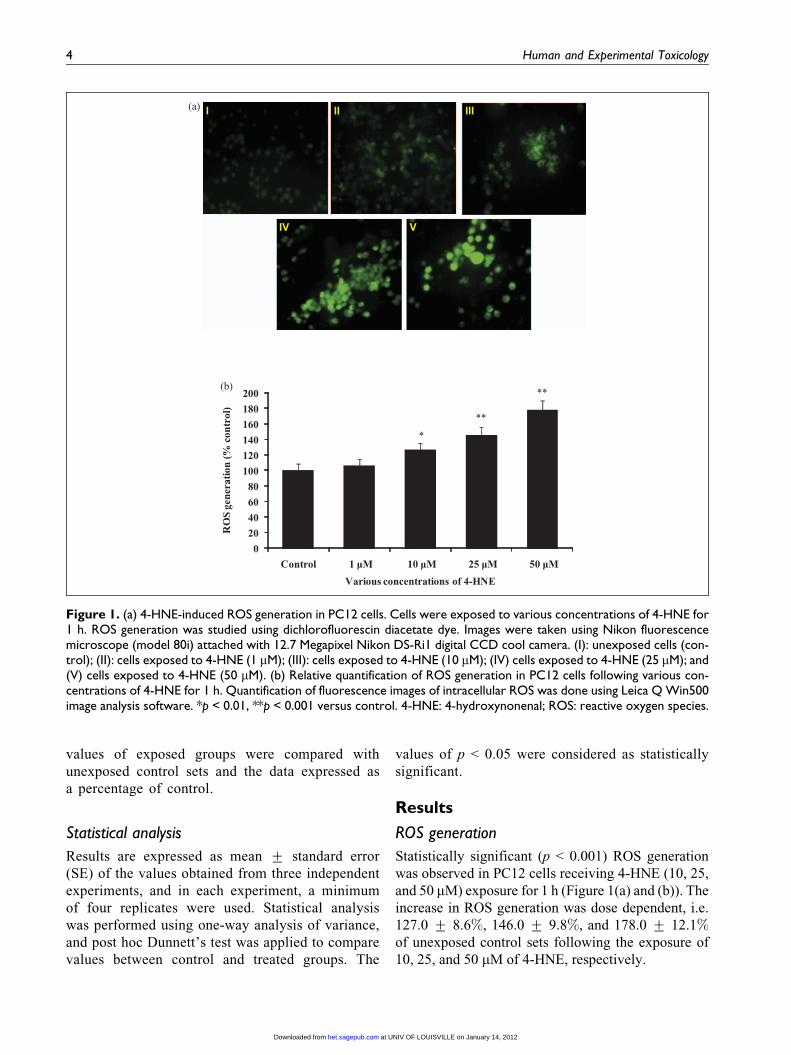
Statistically significant (p < 0.001) ROS generation
was observed in PC12 cells receiving 4-HNE (10, 25,
and 50 mM) exposure for 1 h (Figure 1(a) and (b)). The
increase in ROS generation was dose dependent, i.e.
127.0 + 8.6%, 146.0 + 9.8%, and 178.0 + 12.1%of unexposed control sets following the exposure of
10, 25, and 50 mM of 4-HNE, respectively.
0
20
40
60
80
100
120
140
160
180
200
Control 1 µM 10 µM 25 µM 50 µM
RO
S ge
nera
tion
(% c
ontr
ol)
Various concentrations of 4-HNE
*
**
**
I II III
VIV
(b)
(a)
Figure 1. (a) 4-HNE-induced ROS generation in PC12 cells. Cells were exposed to various concentrations of 4-HNE for1 h. ROS generation was studied using dichlorofluorescin diacetate dye. Images were taken using Nikon fluorescencemicroscope (model 80i) attached with 12.7 Megapixel Nikon DS-Ri1 digital CCD cool camera. (I): unexposed cells (con-trol); (II): cells exposed to 4-HNE (1 mM); (III): cells exposed to 4-HNE (10 mM); (IV) cells exposed to 4-HNE (25 mM); and(V) cells exposed to 4-HNE (50 mM). (b) Relative quantification of ROS generation in PC12 cells following various con-centrations of 4-HNE for 1 h. Quantification of fluorescence images of intracellular ROS was done using Leica Q Win500image analysis software. *p < 0.01, **p < 0.001 versus control. 4-HNE: 4-hydroxynonenal; ROS: reactive oxygen species.
4 Human and Experimental Toxicology
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from
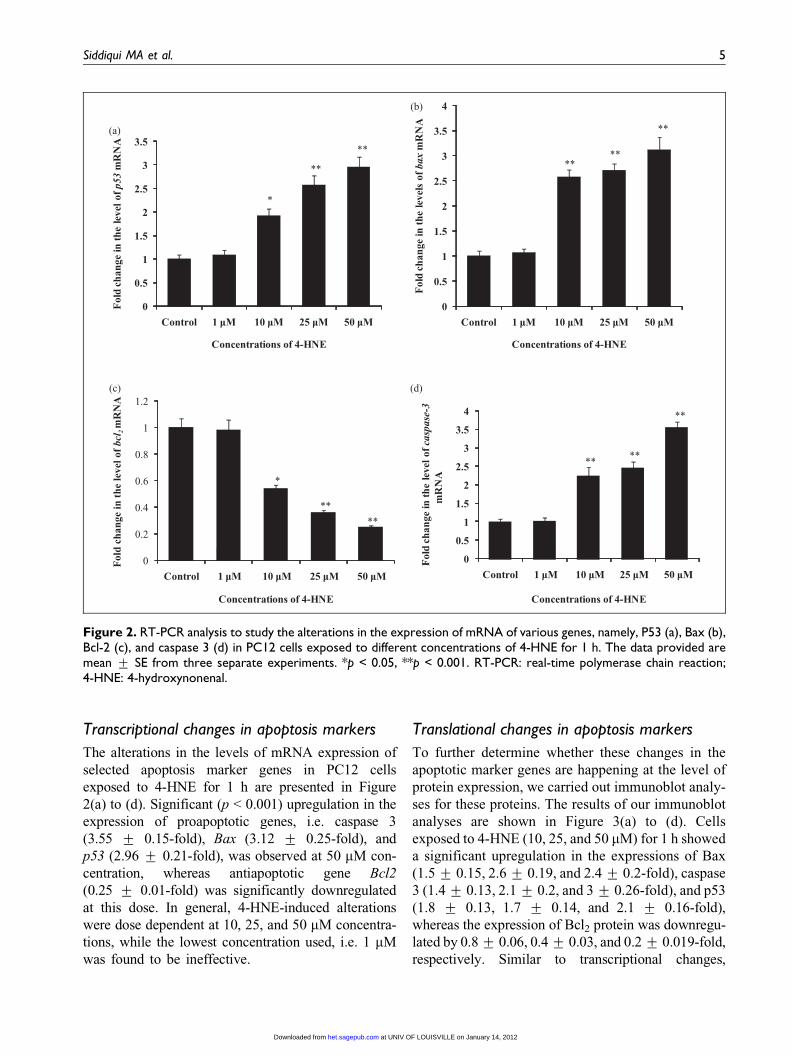
Transcriptional changes in apoptosis markers
The alterations in the levels of mRNA expression of
selected apoptosis marker genes in PC12 cells
exposed to 4-HNE for 1 h are presented in Figure
2(a) to (d). Significant (p < 0.001) upregulation in the
expression of proapoptotic genes, i.e. caspase 3
(3.55 + 0.15-fold), Bax (3.12 + 0.25-fold), and
p53 (2.96 + 0.21-fold), was observed at 50 mM con-
centration, whereas antiapoptotic gene Bcl2
(0.25 + 0.01-fold) was significantly downregulated
at this dose. In general, 4-HNE-induced alterations
were dose dependent at 10, 25, and 50 mM concentra-
tions, while the lowest concentration used, i.e. 1 mM
was found to be ineffective.
Translational changes in apoptosis markers
To further determine whether these changes in the
apoptotic marker genes are happening at the level of
protein expression, we carried out immunoblot analy-
ses for these proteins. The results of our immunoblot
analyses are shown in Figure 3(a) to (d). Cells
exposed to 4-HNE (10, 25, and 50 mM) for 1 h showed
a significant upregulation in the expressions of Bax
(1.5 + 0.15, 2.6 + 0.19, and 2.4 + 0.2-fold), caspase
3 (1.4 + 0.13, 2.1 + 0.2, and 3 + 0.26-fold), and p53
(1.8 + 0.13, 1.7 + 0.14, and 2.1 + 0.16-fold),
whereas the expression of Bcl2 protein was downregu-
lated by 0.8 + 0.06, 0.4 + 0.03, and 0.2 + 0.019-fold,
respectively. Similar to transcriptional changes,
0
0.5
1
1.5
2
2.5
3
3.5
Control 1 µM 10 µM 25 µM 50 µM
Fol
d ch
ange
in th
e le
vel o
f p53
mR
NA
Concentrations of 4-HNE
**
**
*
0
0.5
1
1.5
2
2.5
3
3.5
4
Control 1 µM 10 µM 25 µM 50 µM
Fol
d ch
ange
in th
e le
vels
of bax
mR
NA
Concentrations of 4-HNE
**
**
**
0
0.2
0.4
0.6
0.8
1
1.2
Control 1 µM 10 µM 25 µM 50 µM
Fol
d ch
ange
in t
he le
vel o
f bc
l 2 m
RN
A
Concentrations of 4-HNE
**
**
*
0
0.5
1
1.5
2
2.5
3
3.5
4
Control 1 µM 10 µM 25 µM 50 µM
Fol
d ch
ange
in th
e le
vel o
f cas
pase
-3m
RN
A
Concentrations of 4-HNE
**
**
**
(a)
(c)
(b)
(d)
Figure 2. RT-PCR analysis to study the alterations in the expression of mRNA of various genes, namely, P53 (a), Bax (b),Bcl-2 (c), and caspase 3 (d) in PC12 cells exposed to different concentrations of 4-HNE for 1 h. The data provided aremean + SE from three separate experiments. *p < 0.05, **p < 0.001. RT-PCR: real-time polymerase chain reaction;4-HNE: 4-hydroxynonenal.
Siddiqui MA et al. 5
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from
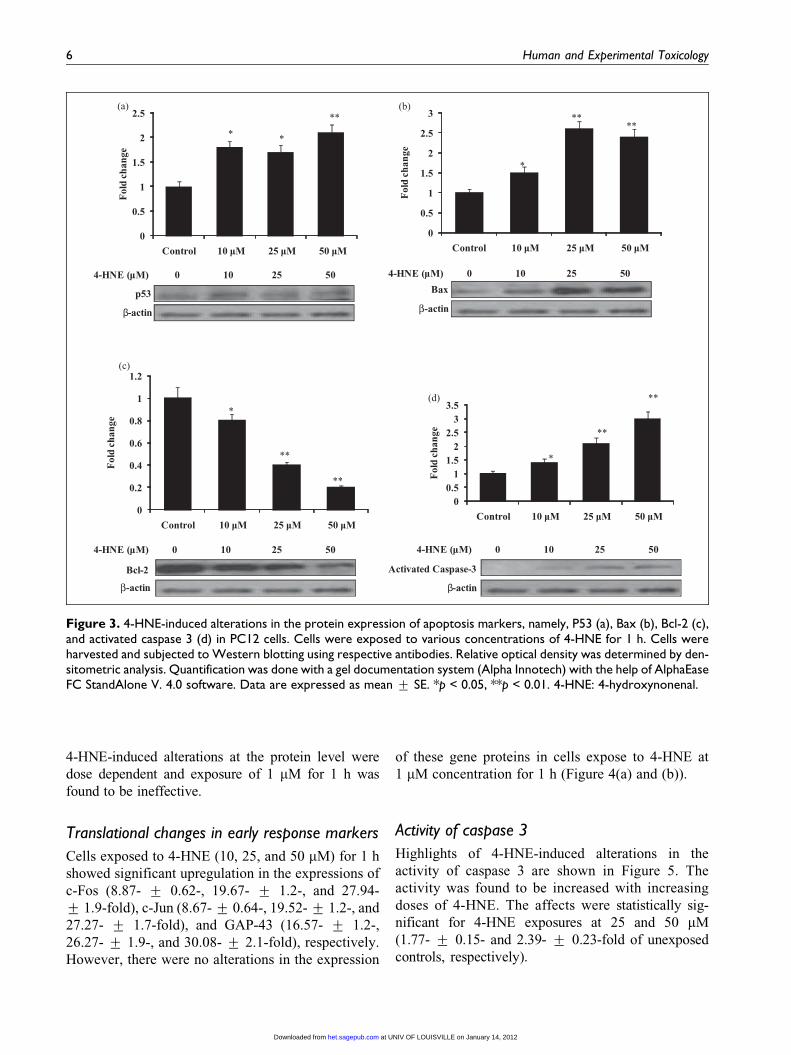
4-HNE-induced alterations at the protein level were
dose dependent and exposure of 1 mM for 1 h was
found to be ineffective.
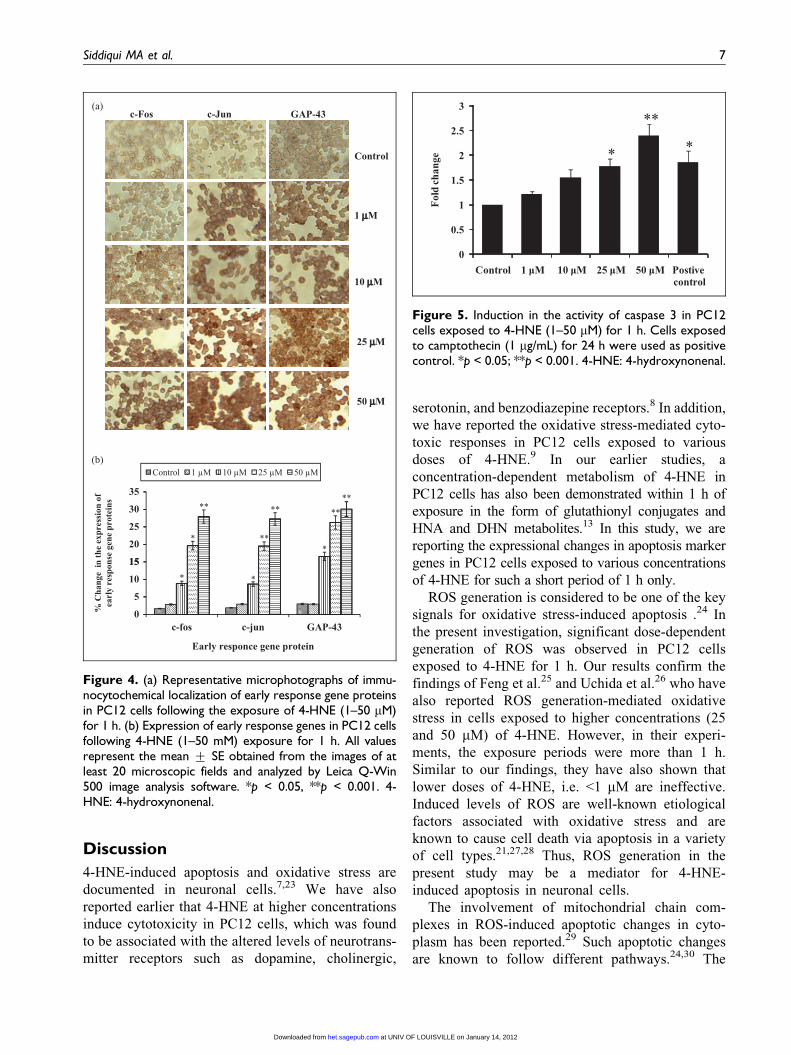
Translational changes in early response markers
Cells exposed to 4-HNE (10, 25, and 50 mM) for 1 h
showed significant upregulation in the expressions of
c-Fos (8.87- + 0.62-, 19.67- + 1.2-, and 27.94-
+ 1.9-fold), c-Jun (8.67- + 0.64-, 19.52- + 1.2-, and
27.27- + 1.7-fold), and GAP-43 (16.57- + 1.2-,
26.27- + 1.9-, and 30.08- + 2.1-fold), respectively.
However, there were no alterations in the expression
of these gene proteins in cells expose to 4-HNE at
1 mM concentration for 1 h (Figure 4(a) and (b)).
Activity of caspase 3
Highlights of 4-HNE-induced alterations in the
activity of caspase 3 are shown in Figure 5. The
activity was found to be increased with increasing
doses of 4-HNE. The affects were statistically sig-
nificant for 4-HNE exposures at 25 and 50 mM
(1.77- + 0.15- and 2.39- + 0.23-fold of unexposed
controls, respectively).
p53
β-actin
0
0.5
1
1.5
2
2.5
Control 10 µM 25 µM 50 µM
Fol
d ch
ange
**
**
4-HNE (µM) 0 10 25 50
Bax
β-actin
4-HNE (µM) 0 10 25 50
0
0.5
1
1.5
2
2.5
3
Control 10 µM 25 µM 50 µM
Fol
d ch
ange
****
*
0
0.2
0.4
0.6
0.8
1
1.2
Control 10 µM 25 µM 50 µM
Fol
d ch
ange
Bcl-2
β-actin
**
**
*
4-HNE (µM) 0 10 25 50
00.5
11.5
22.5
33.5
Control 10 µM 25 µM 50 µM
Fol
d ch
ange
Activated Caspase-3
β-actin
**
**
*
4-HNE (µM) 0 10 25 50
(a)
(c)
(d)
(b)
Figure 3. 4-HNE-induced alterations in the protein expression of apoptosis markers, namely, P53 (a), Bax (b), Bcl-2 (c),and activated caspase 3 (d) in PC12 cells. Cells were exposed to various concentrations of 4-HNE for 1 h. Cells wereharvested and subjected to Western blotting using respective antibodies. Relative optical density was determined by den-sitometric analysis. Quantification was done with a gel documentation system (Alpha Innotech) with the help of AlphaEaseFC StandAlone V. 4.0 software. Data are expressed as mean + SE. *p < 0.05, **p < 0.01. 4-HNE: 4-hydroxynonenal.
6 Human and Experimental Toxicology
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from
Discussion
4-HNE-induced apoptosis and oxidative stress are
documented in neuronal cells.7,23 We have also
reported earlier that 4-HNE at higher concentrations
induce cytotoxicity in PC12 cells, which was found
to be associated with the altered levels of neurotrans-
mitter receptors such as dopamine, cholinergic,
serotonin, and benzodiazepine receptors.8 In addition,
we have reported the oxidative stress-mediated cyto-
toxic responses in PC12 cells exposed to various
doses of 4-HNE.9 In our earlier studies, a
concentration-dependent metabolism of 4-HNE in
PC12 cells has also been demonstrated within 1 h of
exposure in the form of glutathionyl conjugates and
HNA and DHN metabolites.13 In this study, we are
reporting the expressional changes in apoptosis marker
genes in PC12 cells exposed to various concentrations
of 4-HNE for such a short period of 1 h only.
ROS generation is considered to be one of the key
signals for oxidative stress-induced apoptosis .24 In
the present investigation, significant dose-dependent
generation of ROS was observed in PC12 cells
exposed to 4-HNE for 1 h. Our results confirm the
findings of Feng et al.25 and Uchida et al.26 who have
also reported ROS generation-mediated oxidative
stress in cells exposed to higher concentrations (25
and 50 mM) of 4-HNE. However, in their experi-
ments, the exposure periods were more than 1 h.
Similar to our findings, they have also shown that
lower doses of 4-HNE, i.e. <1 mM are ineffective.
Induced levels of ROS are well-known etiological
factors associated with oxidative stress and are
known to cause cell death via apoptosis in a variety
of cell types.21,27,28 Thus, ROS generation in the
present study may be a mediator for 4-HNE-
induced apoptosis in neuronal cells.
The involvement of mitochondrial chain com-
plexes in ROS-induced apoptotic changes in cyto-
plasm has been reported.29 Such apoptotic changes
are known to follow different pathways.24,30 The
Control
1 μμM
10 μμM
25 μμM
50 μμM
c-Fos GAP-43c-Jun
35
Control 1 µM 10 µM 25 µM 50 µM
****
15
20
25
30 ****
***
*
0
5
10
15
% C
hang
e in
the
expr
essi
on o
fea
rly
resp
onse
gen
e pr
otei
ns
**
c-fos
Early responce gene protein
GAP-43c-jun
(b)
(a)
Figure 4. (a) Representative microphotographs of immu-nocytochemical localization of early response gene proteinsin PC12 cells following the exposure of 4-HNE (1–50 mM)for 1 h. (b) Expression of early response genes in PC12 cellsfollowing 4-HNE (1–50 mM) exposure for 1 h. All valuesrepresent the mean + SE obtained from the images of atleast 20 microscopic fields and analyzed by Leica Q-Win500 image analysis software. *p < 0.05, **p < 0.001. 4-HNE: 4-hydroxynonenal.
0
0.5
1
1.5
2
2.5
3
Control 1 µM 10 µM 25 µM 50 µM Postive control
*
**
*
Fol
d ch
ange
Figure 5. Induction in the activity of caspase 3 in PC12cells exposed to 4-HNE (1–50 mM) for 1 h. Cells exposedto camptothecin (1 mg/mL) for 24 h were used as positivecontrol. *p < 0.05; **p < 0.001. 4-HNE: 4-hydroxynonenal.
Siddiqui MA et al. 7
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

mitochondrial-dependent apoptotic pathway is also
known to be involved in 4-HNE-induced cytotoxicity
in various cells31,32 including PC12 cells.16,23 We
observed that 4-HNE significantly upregulates the
expression (mRNA and protein) of P53, Bax, caspase
3, and downregulated Bcl2 in a dose-dependent man-
ner. In general, transcriptional changes were well
coordinated with translational changes and with phy-
siological activity of caspase 3. The upregulation of
nuclear P53 protein is known to play an important role
in minimizing DNA damage by inducing transcrip-
tional reprogramming, which eventually leads to
controlled cell death.33–35 However, higher level of
cytoplasmic P53 protein interacts with mitochondria,
thereby promoting mitochondrial membrane permea-
bilization36 and plays an important role in the regula-
tion of apoptosis.37 This cytoplasmic P53 protein has
been suggested to induce proapoptotic members of
the Bcl2 family such as Bax and Bak, and their displa-
cement with antiapoptotic Bcl2 proteins.38 Thus, the
alterations in the expression profile of marker genes
in this study indicates that P53 triggers the mitochon-
drial apoptotic cascade in PC12 cells exposed to
4-HNE. Similar to our findings, 4-HNE-induced
apoptosis in other cell types has also been suggested
through the P53-dependent intrinsic pathway.11,39 The
induction of P53 by 4-HNE followed by the activation
of caspase 3 and the onset of apoptosis has also been
reported by Li et al.40 Association of 4-HNE-induced
apoptosis with P53 accumulation suggests that the
process of apoptosis elicited by 4-HNE may be due
to the changes in the availability or sensitivity of
specific regulatory pathways that are activated by the
4-HNE.
Initially, P53 is activated in response to DNA dam-
age by 4-HNE, and these upregulated levels of P53
promote induction in the expression of proapoptotic
Bax, while downregulating the expression of antia-
poptotic Bcl2 protein. This imbalance in the ratio of
Bax/Bcl-2 could lead to the dissipation in mitochon-
drial membrane potential. Finally, dysfunctional
mitochondria could release cytochrome-c in cytosol,
which could activate caspase 3 via the activation of
procaspase 9. In the case of 4-HNE-induced apopto-
sis, an imbalance in Bax/Bcl-2 protein may be an
upstream event followed by mitochondrial-mediated
activation of caspase 3, a final executer caspase,
which leads to cell apoptosis. Activation of P53 is
associated with a rapid increase in its levels and with
an increased ability to bind DNA and mediate tran-
scriptional activation of apoptotic signals.21,41
Induction in the levels of early response gene pro-
teins such as c-Fos, c-Jun, and GAP-43, which are
integral components of transcription factor AP-1, has
been discussed as a regulator of cell death, survival,
and regeneration.42 In our present study, exposure of
4-HNE-induced expression of c-Fos, c-Jun, and
GAP-43 shows the effects of 4-HNE as a toxicant and
could be well correlated with previously reported
induced expression of these genes in the appearance
of neurotoxicity.22,43 Oxidation products of 4-HNE
thereby may have a role in regulating cell death/
survival as in the case of neurodegeneration.
Together, our data provide insights to the induction
of apoptotic damages in neuronal cells following
short-term exposure (1 h) of 4-HNE, which may help
understand the possible cellular and molecular
mechanisms involved in 4-HNE-induced neuronal
damage.
Funding
Financial support was provided by Council of
Scientific and Industrial Research, New Delhi, India
(SIP-08).
Acknowledgements
The authors thank the Director, Indian Institute of
Toxicology Research, Lucknow, India, for his keen interest
in the present work and technical laboratory assistance of
Mr Rajesh Misra.
Declaration of Conflict of Interest
The authors declared no conflicts of interest.
References
1. Lu Z, Li CM, Qiao Y, Yan Y and Yang X. Effect of
inhaled formaldehyde on learning and memory of
mice. Indoor Air 2008; 18: 77–83.
2. Cai J, Chen J, He H, Yin Z, Zhu Z and Yin D. Carbonyl
stress: malondialdehyde induces damage on rat hippo-
campal neurons by disturbance of Caþ homeostasis.
Cell Biol Toxicol 2009; 25: 435–445.
3. Butterfield DA, Bader Lange ML and Sultana R.
Involvements of the lipid peroxidation product,
HNE, in the pathogenesis and progression of
Alzheimer’s disease. Biochim Biophys Acta 2010;
1801: 924–929.
4. Sakai T, Shimizu Y and Kawahara S. Effect of
NaCl on the lipid peroxidationderived aldehyde,
4-hydroxy-2-nonenal, formation in boiled pork. Biosci
Biotechnol Biochem 2006; 70: 815–820.
8 Human and Experimental Toxicology
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

5. Lopachin RM, Geohagen BC and Gavin T. Synaptoso-
mal toxicity and nucleophilic targets of 4-hydroxy-2-
nonenal. Toxicol Sci 2009; 107: 171–181.
6. Arakawa M, Ishimura A, Arai Y, Kawabe K, Suzuki S,
Ishige K, et al. NAcetylcysteine and ebselen but not
nifedipine protected cerebellar granule neurons against
4-hydroxynonenal-induced neuronal death. Neurosci
Res 2007; 57: 220–229.
7. Abdul HM and Butterfield DA. Involvement of PI3K/
PKG/ERK1/2 signaling pathways in cortical neurons
to trigger protection by co-treatment of acetyl-L-carni-
tine and alpha-lipoic acid against HNE-mediated oxi-
dative stress and neurotoxicity implications for
Alzheimer’s disease. Free Radic Biol Med 2007; 42:
371–384.
8. Siddiqui MA, Singh G, Kashyap MP, Khanna VK,
Yadav S, Chandra D, et al. Influence of cytotoxic doses
of 4-hydroxynonenal on selected neurotransmitter
receptors in PC-12 cells. Toxicol In Vitro 2008; 22:
1681–1688.
9. Siddiqui MA, Kashyap MP, Khanna VK, Yadav S and
Pant AB. NGF induced differentiated PC12 cells as in
vitro tool to study 4-hydroxynonenal induced cellular
damage. Toxicol In Vitro 2010; 24: 1681–1688.
10. Srivastava S, Ramana KV, Tammali R, Srivastava SK
and Bhatnagar A. Contribution of aldose reductase to
diabetic hyperproliferation of vascular smooth muscle
cells. Diabetes. 2006; 55: 901–910.
11. Sharma R, Sharma A, Dwivedi S, Zimniak P,
Awasthi S and Awasthi YC. 4-Hydroxynonenal
self-limits fas-mediated DISC-independent apoptosis
by promoting export of Daxx from the nucleus to
the cytosol and its binding to Fas. Biochemistry
2008; 47: 143–156.
12. Yadav UC, Ramana KV, Awasthi YC and Srivastava
SK. Glutathione level regulates HNE-induced geno-
toxicity in human erythroleukemia cells. Toxicol Appl
Pharmacol 2008; 227: 257–264.
13. Siddiqui MA, Kashyap MP, Khanna VK, Gupta VK,
Tripathi VK, Srivastava S, et al. Metabolism of
4-hydroxy trans 2-nonenal (HNE) in cultured PC12
cells. Ann Neurosci 2008b; 15: 60–68.
14. Zhang H, Court N and Forman HJ. Submicromolar
concentrations of 4-hydroxynonenal induce glutamate
cysteine ligase expression in HBE1 cells. Redox Rep
2007; 12: 101–106.
15. Jang YJ, Kim JE, Kang NJ, Lee KW and Lee HJ.
Piceatannol attenuates 4-hydroxynonenal-induced
apoptosis of PC12 cells by blocking activation of
c-Jun N-terminal kinase. Ann N Y Acad Sci 2009;
1171: 176–182.
16. Siddiqui MA, Kashyap MP, Kumar V, Al-Khedhairy
AA, Musarrat J and Pant AB. Protective potential of
trans-resveratrol against 4-hydroxynonenal induced
damage in PC12 cells. Toxicol In Vitro 2010b; 24:
1592–1598.
17. Gallagher EP, Huisden CM and Gardner JL.
Transfection of HepG2 cells with hGSTA4 provides
protection against 4-hydroxynonenal-mediated oxida-
tive injury. Toxicol In Vitro 2007; 21: 1365–1372.
18. Borovic S, Cipak A, Meinitzer A, Kejla Z, Perovic D,
Waeg G and Zarkovic N. Differential sensitivity to
4-hydroxynonenal for normal and malignant mesench-
ymal cells. Redox Rep 2007; 12: 50–54.
19. Miranda CL, Reed RL, Kuiper HC, Alber S and Ste-
vens JF. Ascorbic acid promotes detoxification and
elimination of 4-hydroxy-2(E)-nonenal in human
monocytic THP-1 cells. Chem Res Toxicol 2009; 22:
863–874.
20. Pant AB, Agarwal AK, Sharma VP and Seth PK. In
vitro cytotoxicity evaluation of plastic biomedical
devices. Hum Exp Toxicol 2001; 20: 412–417.
21. Kashyap MP, Singh AK, Siddiqui MA, Kumar V, Tri-
pathi VK, Khanna VK, et al. Caspase cascade regu-
lated mitochondria mediated apoptosis in
monocrotophos exposed PC12 cells. Chem Res Toxicol
2010; 23: 1663–1672.
22. Kashyap MP, Singh AK, Kumar V, Tripathi VK,
Srivastava VK, Agarwal M, et al. Monocrotophos
induced apoptosis in PC12 cells: role of xenobiotic
metabolizing cytochrome P450s. PLoS ONE 2011;
6: e17757.
23. Raza H and John A. 4-hydroxynonenal induces mito-
chondrial oxidative stress, apoptosis and expression
of glutathione S-transferase A4-4 and cytochrome
P450 2E1 in PC12 cells. Toxicol Appl Pharmacol
2006; 216: 309–318.
24. Kroemer G, Galluzzi L and Brenner C. Mitochondrial
membrane permeabilization in cell death. Physiol Rev
2007; 87: 99–163.
25. Feng Q, Kumagai T, Torii Y, Nakamura Y, Osawa T
and Uchida K. Anticarcinogenic antioxidants as inhibi-
tors against intracellular oxidative stress. Free Radic
Res 2001; 35: 779–788.
26. Uchida Y, Ohba K, Yoshioka T, Irie K, Muraki T
and Maru Y. Cellular carbonyl stress enhances the
expression of plasminogen activator inhibitor-1 in rat
white adipocytes via reactive oxygen
species-dependent pathway. J Biol Chem 2004; 279:
4075–4083.
27. Fortunato JJ, Agostinho FR, Reus GZ, Petronilho FC,
Dal-Pizzol F and Quevedo J. Lipid peroxidative
Siddiqui MA et al. 9
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from

damage on malathion exposure in rats. Neurotox Res
2006; 9: 23–28.
28. Sompol P, Ittarat W, Tangpong J, Chen Y, Doubins-
kaia I, Batinic-Haberle I, et al. A neuronal model of
Alzheimer’s disease: an insight into the mechanisms
of oxidative stress-mediated mitochondrial injury.
Neuroscience 2008; 153: 120–130.
29. Galluzzi L, Morselli E, Kepp O, Tajeddine N and
Kroemer G. Targeting p53 to mitochondria for cancer
therapy. Cell Cycle 2008; 7: 1949–1955.
30. Segui B and Legembre P. Redistribution of CD95 into
the lipid rafts to treat cancer cells? Recent Pat Antican-
cer Drug Discov 2010; 5: 22–28.
31. Jin Xu, Ji L-D and Xu L-H. Lead-induced apoptosis in
PC 12 cells: involvement of p53, Bcl-2 family and cas-
pase-3. Toxicol Lett 2006; 166: 160–167.
32. Vaillancourt F, Fahmi H, Shi Q, Lavigne P, Ranger P,
Fernandes JC and Benderdour M. 4-Hydroxynonenal
induces apoptosis in human osteoarthritic chondrocytes:
the protective role of glutathione-S-transferase. Arthritis
Res Ther 2008; 10: R107.
33. Bunz F, Dutriaux A, Lengauer C, Waldman T, Zhou S,
Brown JP, et al. Requirement for p53 and p21 to sus-
tain G2 arrest after DNA damage. Science 1998; 282:
1497–1501.
34. Bargonetti J and Manfredi JJ. Multiple roles of the tumor
suppressor p53. Curr Opin Oncol 2002; 14: 86–91.
35. Cui Q, Yu JH, Wu JN, Tashiro S, Onodera S, Minami
M and Ikejima T. P53-mediated cell cycle arrest and
apoptosis through a caspase-3- independent, but
caspase-9-dependent pathway in oridonin-treated
MCF-7 human breast cancer cells. Acta Pharmacol Sin
2007; 8: 1057–1066.
36. Moll UM, Wolff S, Speidel D and Deppert W. Tran-
scription independent pro-apoptotic functions of p53.
Curr Opin Cell Biol 2005; 17: 631–636.
37. Yu J and Zhang L. The transcriptional targets of p53 in
apoptosis control. Biochem Biophys Res Commun
2005; 331: 851–858.
38. Galluzzi L, Blomgren K and Kroemer G. Mitochon-
drial membrane permeabilization in neuronal injury.
Nat Rev Neurosci 2009; 10: 481–494.
39. Awasthi YC, Sharma R, Sharma A, Yadav S, Singhal
SS, Chaudary P, et al. Self-regulatory role of
4-hydroxynonenal in signaling for stress-induced pro-
grammed cell death. Free Radic Biol Med 2008; 45:
111–118.
40. Li D, Hinshelwood A, Gardner R, McGarvie G and
Ellis EM. Mouse aldo-keto reductase AKR7A5
protects V79 cells against 4-hydroxynonenal-induced
apoptosis. Toxicology 2006; 226: 172–180.
41. Lakin ND and Jackson SP. Regulation of p53 in
response to DNA damage. Oncogene 1999; 18,
7644–7655.
42. Vollgraf U, Wegner M and Richter-Landsberg C.
Activation of AP-1 and nuclear factor-kappaB tran-
scription factors is involved in hydrogen
peroxide-induced apoptotic cell death of oligodendro-
cytes. J Neurochem 1999; 73: 2501–2509.
43. Vaudano E, Rosenbald C and Bjorklund A. Injury
induced c-Jun expression and phosphorylation in the
dopaminergic nigral neurons of the rat: correlation
with neuronal death and modulation by
glial-cell-line derived neurotropic factor. Eur J Neu-
rosci 2001; 13: 1–14.
10 Human and Experimental Toxicology
at UNIV OF LOUISVILLE on January 14, 2012het.sagepub.comDownloaded from
Related Documents











