Histochemical Journal 23, 273--280 (1991) Histochemical profiles of motoneurons innervating muscle fibres with different activity patterns in the zebrafish, Brachydanio rerio F. DE GRAAF, W. VAN RAAMSDONK*, E. VAN ASSELT and P. C. DIEGENBACH Department of Experimental Zoology, University of Amsterdam, Kruislaan 320, I098 SM Amsterdam, the Netherlands Received 9th October 1990 and in revised form 20th January 1991 Sumlnary Enzyme histochemical profiles of spinal motoneurons in the zebrafish were determined. Five enzymes of glucose metabolism were chosen: glucose-6-phosphate dehydrogenase (G6PDH), hexokinase (HK), phosphofructokinase (PFK), succinate dehydrogenase (SDH) and NADH tetrazolium reductase (NADH-TR). Motoneurons were traced with Fluorogold and classified as those that innervate white muscle fibres (W-MNs) and those that innervate red and intermediate muscle fibres (R/ I-MNs). The average enzyme activities per volume of tissue in the somata of both populations differed at most by 25%. Both the average soma volume and the average number of muscle fibres innervated are three times larger for the W-MNs than for the WI-MNs. This suggests that the total amount of enzyme activity within a neuron soma matches target size. In the WI-MNs, the activities of SDH and NADH-TR were closely correlated (correlation coefficient, r = 0.99; p < 0.05) and HK activity correlated well with G6PDH activity (r = 0.94; p < 0.05), but not with PFK (r = 0.64; p > 0.05). In the W-MNs, there was no correlation between SDH and NADH-TR (r = - 0.59; p > 0.05) or between HK and G6PDH (r = 0.50; p > 0.05) and the correlation coefficient between HK and PFK activity was close to zero (r = 0.04; p > 0.05). It was concluded that in the WI-MNs, which are continuously active, firing activity is fuelled by oxidative metabolism. We suggest that in the W-MNs glucose is stored in the form of glycogen and that, despite high levels of NADH-TR present, the energy for intermittent firing activity is provided by glycolysis. Introduction Motoneurons that innervate the axial musculature in the zebrafish, Brachydanio rerio are a good model in which to investigate the relationship between metabo- lic characteristics and neuronal function. Moto- neurons innervate different numbers and types of muscle fibres and their firing activity patterns vary (Burke, 1981). In the zebrafish two populations can be recognized. One population, the white motoneurons, W-MNs, innervates the white muscle fibres which are active at high speeds that are only sustained for short periods. The other population, the red and intermedi- ate motoneurons, WI-MNs, is composed of moto- neurons that innervate the intermediate or red muscle fibres, which are both active at cruising speeds. The two populations differ in somatal positions. The W- MNs are located close to the central canal and the WI- MNs in the ventro-lateral part of the motor column (de Graaf et al., 1990). It has been shown that motoneurons that innervate different muscle fibre types differ in several ways. They differ in oxidative metabolic capacity (Sickles & *To whom all correspondence should be addressed. Oblack, 1984; Ishihara et al., 1988), in physiological characteristics, e.g. duration of the after- hyperpolarization and axonal conduction velocity (Fleshman et al., 1981; Kernell, 1986) and in soma size and morphology of axons and dendrites (Cullheim & Kellerth, 1978; Ulfhake & Kellerth, 1982). Wong-Riley and co-workers have shown that the terminal enzyme of the respiratory chain, cytochrome c oxidase (COX, E.C. 1.9.3.1) is an important marker for neuronal activity (reviewed in Wong-Riley, 1989). In a previous study of the zebrafish spinal moto- neurons one of us showed that COX activity is highest in the W-MNs, whereas sucdnate dehydrogenase activity (SDH, E.C. 1.3.99.1), which is commonly used as a marker of activity in muscle fibres, is highest in the WI-MNs (van Raamsdonk et al., 1987). Several energy and metabolite consuming processes take place in the neuron soma, for instance membrane repolarization and synthesis of proteins and membra- nes. The activity of an enzyme can be of importance for more than one of these processes. The histochemical analysis of a single enzyme will then provide limited information only. We therefore composed histochemi- 0018-2214/91 $03.00 +.12 1991 Chapman and Hall Ltd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Histochemical Journal 23, 273--280 (1991)
Histochemical profiles of motoneurons innervating muscle fibres with different activity patterns in the zebrafish, Brachydanio rerio
F. D E G R A A F , W. V A N R A A M S D O N K * , E. V A N A S S E L T a n d P. C. D I E G E N B A C H
Department of Experimental Zoology, University of Amsterdam, Kruislaan 320, I098 SM Amsterdam, the Netherlands
Received 9th October 1990 and in revised form 20th January 1991
Sumlnary Enzyme histochemical profiles of spinal motoneurons in the zebrafish were determined. Five enzymes of glucose metabolism were chosen: glucose-6-phosphate dehydrogenase (G6PDH), hexokinase (HK), phosphofructokinase (PFK), succinate dehydrogenase (SDH) and NADH tetrazolium reductase (NADH-TR). Motoneurons were traced with Fluorogold and classified as those that innervate white muscle fibres (W-MNs) and those that innervate red and intermediate muscle fibres (R/ I-MNs). The average enzyme activities per volume of tissue in the somata of both populations differed at most by 25%. Both the average soma volume and the average number of muscle fibres innervated are three times larger for the W-MNs than for the WI-MNs. This suggests that the total amount of enzyme activity within a neuron soma matches target size.
In the WI-MNs, the activities of SDH and NADH-TR were closely correlated (correlation coefficient, r = 0.99; p < 0.05) and HK activity correlated well with G6PDH activity (r = 0.94; p < 0.05), but not with PFK (r = 0.64; p > 0.05). In the W-MNs, there was no correlation between SDH and NADH-TR (r = - 0.59; p > 0.05) or between HK and G6PDH (r = 0.50; p > 0.05) and the correlation coefficient between HK and PFK activity was close to zero (r = 0.04; p > 0.05).
It was concluded that in the WI-MNs, which are continuously active, firing activity is fuelled by oxidative metabolism. We suggest that in the W-MNs glucose is stored in the form of glycogen and that, despite high levels of NADH-TR present, the energy for intermittent firing activity is provided by glycolysis.
In t roduc t ion
Motoneurons that innervate the axial muscula ture in the zebrafish, Brachydanio rerio are a good model in which to investigate the relat ionship be tween metabo- lic characteristics and neurona l function. Moto- neurons innervate different numbers and types of muscle fibres and their firing activity pat terns vary (Burke, 1981). In the zebrafish two popula t ions can be recognized. One populat ion, the white motoneurons , W-MNs, innervates the white muscle fibres which are active at h igh speeds that are only sustained for short periods. The other populat ion, the red and intermedi- ate motoneurons , WI-MNs, is composed of moto- neurons that innervate the intermediate or red muscle fibres, which are bo th active at cruising speeds. The two popula t ions differ in somatal positions. The W- MNs are located close to the central canal and the WI- MNs in the ventro-lateral par t of the motor co lumn (de Graaf et al., 1990).
It has been shown that mo toneu rons that innervate different muscle fibre types differ in several ways. They differ in oxidative metabolic capacity (Sickles &
*To whom all correspondence should be addressed.
Oblack, 1984; Ishihara et al., 1988), in physiological characteristics, e.g. durat ion of the after- hyperpolar iza t ion and axonal conduct ion velocity (Fleshman et al., 1981; Kernell, 1986) and in soma size and morpho logy of axons and dendri tes (Cullheim & Kellerth, 1978; Ulfhake & Kellerth, 1982).
Wong-Riley and co-workers have sh o w n that the terminal en zy m e of the respira tory chain, cy tochrome c oxidase (COX, E.C. 1.9.3.1) is an impor tan t marker for neurona l activity (reviewed in Wong-Riley, 1989). In a previous s tudy of the zebrafish spinal moto- neurons one of us showed that COX activity is highest in the W-MNs, whereas sucdna te dehydrogenase activity (SDH, E.C. 1.3.99.1), which is commonly used as a marker of activity in muscle fibres, is highest in the WI-MNs (van Raamsdonk et al., 1987).
Several energy and metabolite consuming processes take place in the neu ron soma, for instance m embrane repolarizat ion and synthesis of proteins and membra- nes. The activity of an en zy m e can be of impor tance for more than one of these processes. The histochemical analysis of a single enzyme will then provide limited informat ion only. We therefore composed histochemi-
0018-2214/91 $03.00 +.12 �9 1991 Chapman and Hall Ltd

274 DE GRAAF, VAN RAAMSDONK, VAN ASSELT and DIEGENBACH
cal profiles f rom the activities of several enzymes , each representa t ive of one of the ma in metabol ic routes .
Under normal nutr i t ional condit ions, glucose is the source of ene rgy for ne rvous tissue (Sokoloff, 1974). The his tochemical profiles therefore included: hexoki- nase (HK, E.C. 2.7.1.1), the first and rate-l imiting e n z y m e of glucose utilization; g lucose-6-phospha te d e h y d r o g e n a s e (G6PDH, E.C. 1.1.1.49), key e n z y m e of the ox ida t ive pen tose p h o s p h a t e p a t h w a y and indicative of the util ization of glucose in biosynthesis ; phosphof ruc tok inase (PFK, E.C. 2.7.1.11), key e n z y m e of glycolysis; SDH, one of the Krebs cycle enzymes ; N A D H te t razol ium reductase (NADH-TR, E.C. 1.6.99.3), the first s tep in the electron t r anspor t chain. NADH-TR, like COX, m a y be indicative of overall neurona l activity as all o ther metabol ic pa th- ways converge into the electron t ranspor t sys tem.
The p resen t s tudy reveals that W-MNs a n d WI-MNs have different h is tochemical profiles. O n the basis of correlat ions b e t w e e n e n z y m e activities we sugges t that the cont inuous firing activity in the WI-MNs is fuelled by oxidative metabol i sm, whe rea s the inter- mi t ten t firing activity of the W - M N s is fuelled b y glycolysis.
and stored under liquid nitrogen until used. Sections containing labelled somata were photographed prior to incubation, while they were still dry, because tracer diffused from the sections in aqueous media. A Zeiss fluorescence microscope with an excitation filter of 440 to 500 nm and a barrier filter of 515 nm was used.
Labelled somata in the medio-dorsal part of the motor column were classified as W-MNs and in the ventro-lateraI part as WI-MNs. Further subdivision into R-MNs and I-MNs could not be made because their locations largely overlap (de Graaf et al., 1990).
Labelling percentages should not exceed 100%. These percentages were calculated from counts in 90 transverse serial sections of 10 ~m thickness of the spinal cords. Neuron somata that occurred in more than one section were counted only once. One body segment is supplied with innervation by a stretch of spinal cord that measures approximately 700~m and contains on average 130 motoneurons (van Raamsdonk et al., 1983).
One fish was used for the demonstration of acetylcholi- nesterase activity (ACHE, E.C. 3.1.1.7) 5.5 weeks after injection with FLG (Fig. 1). Absence of AChE activity in FLG positive neurons would imply labelling of other neurons besides motoneurons. AChE activity was demonstrated according to Tsuji (1974), but 50 mM MES-KOH buffer, pH 5.5, was used instead of acetate.
Materials and methods
Experimental animals Adult zebrafish (Brachydanio rerio) ranging in length from 35 to 48 mm were used. They were fed a commercial flake food once a day, kept on a day-night cycle of 16 h light and 8 h dark, and at a water temperature of 25 ~ C.
Substrates and reagents Paraformaldehyde (PFA), glutaraldehyde (GA) and ATP were obtained from Merck. Fluorogold (FLG) was obtained from Fluorochrome Inc., Colorado, USA. Phenazine metho- sulphate (PMS), MES, HEPES, NAD +, NADP +, NADH, nitro-blue tetrazolium (NBT), tetranitro-blue tetrazolium (TNBT) and glyceraldehyde-3-phosphate dehydrogenase (Ga3PDH, E.C. 1.2.1;9) were obtained from Sigma. Succin- ate (di-Na salt), acetylthiocholine-iodide, dimethylforma- mide (DMF) and polyvhlyl alcohol (PVA, M.W. 14,000) were obtained from BDH. Aldolase (E.C. 4.1.2.7), G6PDH, glucose-6-phosphate (G6P) and fructose-6-phosphate (F6P) were obtained from Boehringer. All other reagents used were of analytical grade.
Identification of motoneurons Motoneurons were identified by retrograde tracing with FLG (Schmued & Fallon, 1986). On two consecutive days 1 ~1 of a 4% solution was injected in the axial muscle just caudal to the anus. The fish were killed by decapitation on the 8th day following the first injection. The spinal cord was removed and embedded in 15% (w/v) gelatin in phosphate buffered saline (PBS) and frozen in liquid nitrogen. Trans- verse cryostat sections (10 ~m) were cut at - 30 ~ C, collected on gelatin-chrome-alum coated slides and air-dried at room temperature. The slides were wrapped in aluminiurn foil
Histochemical procedures NADH-TR activity was demonstrated as described by Lojda et al. (1976).
SDH activity was demonstrated according to methods as described by Van Noorden et al. (1983).
G6PDH activity was demonstrated according to van Noorden et al. (1984).
HK activity was demonstrated in a medium containing 10ram glucose, 10m~ ATP, 1ram NADP +, 10mM MgC12, 5mM NAN3, 1.25U m1-1 G6PDH, 0.33ram PMS, in 0.1M HEPES-KOH buffer pH 7.9, 18% PVA and 5.8mM TNBT, dissolved by mild heating in a mixture of ethanol and DMF. The final concentration of DMF was 4% (v/v) and of ethanol 2% (v/v). For a discussion of the use of PVA see van Noorden and Vogels (1989).
The method for the demonstration of PFK activity was optimized for this project and will be described in detail elsewhere. It was performed in a medium containing 12.5mM F6P, 2.5ram ATP, 5ram NAD +, 5ram AMP, 5ram MgClv 5 m ~ NAN3, 1.4 U m1-1 aldolase, 10 U m1-1 Ga3PDH, 0.33mM PMS in 0.1M phosphate buffer pH 7.4, 18% PVA and 5.8 mM TNBT dissolved as described above.
PVA media were removed from the sections with tap water at 60 ~ C. Sections were postfixed for 20 min in 4% PFA, 2% CaC12 at room temperature. All sections were enclosed in glycerin gelatin.
Sections for G6PDH were incubated at 25~ for 15rain. Sections for HK, PFK, SDH and NADH-TR at 37 ~ C for 15, 15, 30 and 10 rain respectively. These conditions were within the range in which we observed linearity between time and reaction product formation.
Control incubations were performed by omitting subs- trates from the media.
To evaluate the effect of FLG on demonstrable enzyme activity large unlabelled somata in the medio-dorsal part of

Histochemical profiles of m o t o n e u r o n s wi th different activity pa t t e rns 275
the motor column were measured. These somata are almost certainly W-MNs (de Graaf et al., 1990). The nearest labelled W-MN was chosen for comparison with each unl~belled soma (Fig. 2).
Quantification End point measurements were made to determine the enzyme activity in the motoneuron somata. Circular fields with a diameter of 2 ~tm were measured on a Zeiss cyto- spectrophotometer with a x40 objective at a wavelength of 550nm. For each soma the mean absorbance of three separate measurements was determined.
Statistics For statistical analysis the package 'Statworks' was used on an Apple Macintosh microcomputer. Linear regression analysis and ANOVA tests were used; samples were considered to be from different populations if p < 0.05.
Results
Identification of motoneurons The distr ibution of FLG labelled somata in the spinal cord co r r e sponded to the distr ibution prev ious ly obse rved fol lowing tracing with horse rad ish perox- idase (de Graaf et al., 1990). AChE activity could be d e m o n s t r a t e d in all somata labelled by FLG injection 5.5 weeks before (Fig. 1) and the labelling percen tage ave raged 75%. It is therefore unlikely that o ther neu rons bes ides m o t o n e u r o n s were labelled.
Control reactions Control incubat ions resul ted in negligible s ta ining (less than 10% of normal intensity). This is in accordance wi th p rev ious results in zebrafish (van R a a m s d o n k et al., 1987) and in weak ly electric fish (Straatsburg et al., 1989).
Fig. 1. (A) Fluorescence photograph of dry cryostat section with FLG labelled neurons. Survival time in this experiment was 5.5 weeks. Labelling of glial cells neighbouring the central canal (CC) can easily be discerned from neuronal labelling (arrow heads). (B) Same section as in A after staining for ACHE. Arrow heads at AChE-positive neurons. Scale bar = 50 ~m.
Fig. 2. (A) Fluorescence photograph of dry cryostat section containing FLG labelled neurons. (B) Same section as in A after staining for PFK. Note the equal staining intensity of the labelled neurons (arrow heads) compared to the contralateral large diameter neuron (arrow), that contains no label. CC = central canal. Scale bar = 50 ~m.
276 DE GRAAF, VAN RAAMSDONK, VAN ASSELT and DIEGENBACH
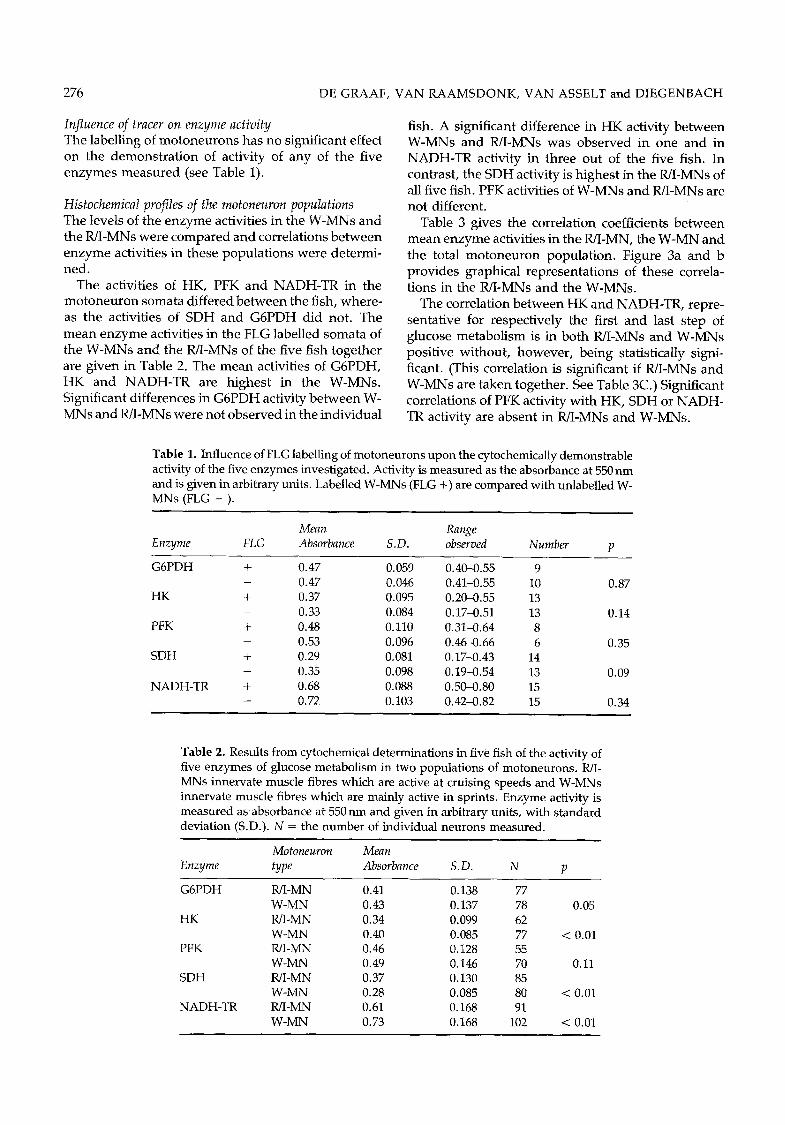
Influence of tracer on enzyme activity The labelling of m o t o n e u r o n s has no significant effect on the demons t r a t ion of activity of any of the five e n z y m e s m e a s u r e d (see Table 1).
HistochemicaI profiles of the motoneuron populations The levels of the e n z y m e activities in the W-MNs and the R/I-MNs were c o m p a r e d and correlat ions b e t w e e n e n z y m e activities in these popula t ions were determi- ned.
The activities of HK, PFK and N A D H - T R in the m o t o n e u r o n somata differed b e t w e e n the fish, where - as the activities of SDH and G 6 P D H did not. The m e a n e n z y m e activities in the FLG labelled soma ta of the W-MNs and the WI-MNs of the five fish toge ther are g iven in Table 2. The m e a n activities of G6PDH, H K and N A D H - T R are h ighes t in the W-MNs. Significant differences in G 6 P D H activity b e t w e e n W- M N s and R/I-MNs were not obse rved in the individual
fish. A significant difference in H K activity be tw een W-MNs and WI-MNs was obse rved in one and in N A D H - T R activity in three out of the five fish. In contrast , the SDH activity is h ighes t in the R/I-MNs of all five fish. PFK activities of W - M N s and WI-MNs are not different.
Table 3 gives the correlat ion coefficients b e t w e e n m e a n e n z y m e activities in the WI-MN, the W - M N and the total m o t o n e u r o n popula t ion . Figure 3a and b p rov ides graphical represen ta t ions of these correla- t ions in the WI-MNs and the W-MNs.
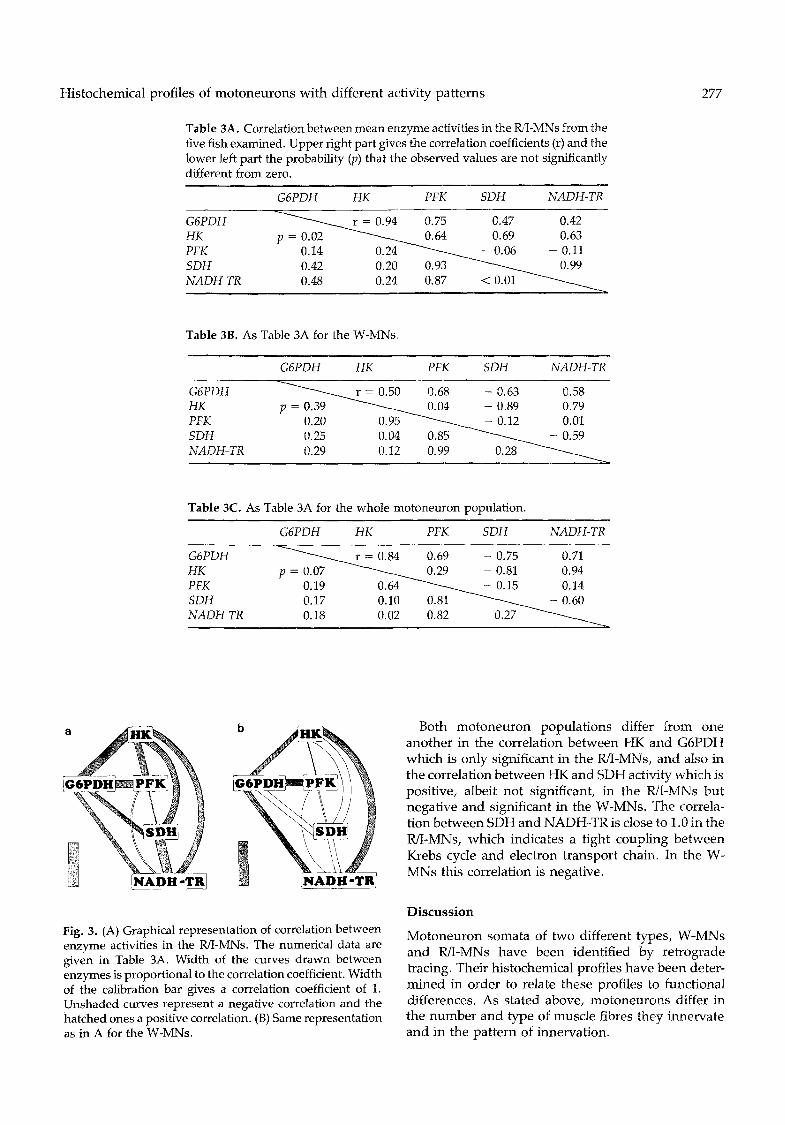
The correlat ion b e t w e e n H K and NADH-TR, repre- sentat ive for respect ively the first and last s tep of glucose metabo l i sm is in bo th R/I-MNs and W-MNs posi t ive wi thout , however , be ing statistically signi- ficant. (This correlat ion is significant if R/I-MNs and W-MNs are t aken together . See Table 3C.) Significant correlations of PFK activity wi th HK, SDH or N A D H - TR activity' are absent in WI-MNs and W-MNs.
Table 1. Influence of FLG labelling of motoneurons upon the cytochemically demonstrable activity of the five enzymes investigated. Activity is measured as the absorbance at 550 nm and is given in arbitrary units. Labelled W-MNs (FLG +) are compared with unlabelled W- MNs (FLG - ).
Mean Range Enzyme FLG Absorbance S.D. observed Number p
G6PDH + 0.47 0.059 0.40-0.55 9 - 0.47 0.046 0.41-0.55 10
HK + 0.37 0.095 0.20-0.55 13 - 0.33 0.084 0.17--0.51 13
PFK + 0.48 0.110 0.31--0.64 8 - 0.53 0.096 0.46-0.66 6
SDH + 0.29 0.081 0.17-0.43 14 - 0.35 0.098 0.19--0.54 13
NADH-TR + 0.68 0.088 0.50-0.80 15 - 0.72 0.103 0.42--0.82 15
0.87
0.14
0.35
0.09
0.34
Table 2. Results from cytochemical determinations in five fish of the activity of five enzymes of glucose metabolism in two populations of motoneurons. R/I- MNs innervate muscle fibres which are active at cruising speeds and W-MNs innervate muscle fibres which are mainly active in sprints. Enzyme activity is measured as absorbance at 550 nm and given in arbitrary units, with standard deviation (S.D.). N = the number of individual neurons measured.
Motoneuron Mean Enzyme type Absorbance S.D. N p
G6PDH WI-MN 0.41 0.138 77 W-MN 0.43 0.137 78
HK WI-MN 0.34 0.099 62 W-MN 0.40 0.085 77
PFK WI-MN 0.46 0.128 55 W-MN 0.49 0.146 70
SDH R/I-MN 0.37 0.130 85 W-MN 0.28 0.085 80
NADH-TR R/I-MN 0.61 0.168 9I W-MN 0.73 0.168 102
0.05
< 0.01
0.11
< 0,01
< 0,01
Histochemical profiles of m o t o n e u r o n s wi th different activity pa t te rns 277
Table 3A. Correlation between mean enzyme activities in the R//-MNs from the five fish examined. Upper right part gives the correlation coefficients (r) and the lower left part the probability (p) that the observed values are no~ significantly different from zero.
G6PDH HK PFK SDH NADH-TR
G6PDH HK PFK SDH NADH-TR
~ ~ = 0.94 0.75 0.47 0.42 p = 0.02 ~ 0.64 0.69 0.63
0.14 0.24 ~ - 0.06 - 0.11 0.42 0.20 0.93 ~ , ~ 0.99 0.48 0.24 0.87 ~
Table 3B. As Table 3A for the W-MNs.
G6PDH HK PFK SDH NADH-TR
G6PDH = 0.50 0.68 - 0.63 0.58 HK p = 0.39 ~ 0.04 - 0.89 0.79 PFK 0.20 0.95 ~ - 0.12 0.01 SDH 0.25 0.04 0.85 ~ ~ - 0.59 NADH-TR 0.29 0.12 0.99 0.28
Table 3C. As Table 3A for the whole motoneuron population.
G6PDH HK PFK SDH NADH-TR
G6PDH ~ = 0.84 0.69 - 0.75 0.71 HK p = 0.07 ~ 0.29 - 0.81 0.94 PFK 0.19 0.64 ~ - 0.15 0.14 SDH 0.17 0.10 0.81 ~ - 0 . 6 0
027 NADH TR 0.18 0.02 0.82
Fig. 3. (A) Graphical representation of correlation between enzyme activities in the R/I-MNs. The numerical data are given in Table 3A. Width of the curves drawn between enzymes is proportional to the correlation coefficient. Width of the calibration bar gives a correlation coefficient of 1. Unshaded curves represent a negative correlation and the hatched ones a positive correlation. (B) Same representation as in A for the W-MNs.
Both m o t o n e u r o n popula t ions differ f rom one ano the r in the correlat ion b e t w e e n H K and G6PDH which is only significant in the WI-MNs, and also in the correlation b e t w e e n H K and SDH activity which is posit ive, albeit not significant, in the WI-MNs but negat ive and significant in the W-MNs. The correla- t ion b e t w e e n SDH and N A D H - T R is close to 1.0 in the WI-MNs, which indicates a t ight coupl ing be tw een Krebs cycle and electron t ranspor t chain. In the W- MNs this correlat ion is negat ive.
Discussion
M o t o n e u r o n somata of two different types, W - M N s and WI-MNs have been identified by re t rograde tracing. Their h is tochemical profiles have been deter- m ined in order to relate these profiles to functional differences. As s ta ted above, m o t o n e u r o n s differ in the n u m b e r and type of muscle fibres they innerva te and in the pa t t e rn of innervat ion.

278 DE GRAAF, VAN RAAMSDONK, VAN ASSELT and DIEGENBACH
Target size and metabolic capacity From counts of muscle fibres and ventral root axons (van Raamsdonk et al., 1983) it appears that the average number of muscle fibres innervated by W- MNs is approximately three times larger than that of the WI-MNs.
Biosynthetic processes such as protein synthesis are almost confined to the neuron soma and the products are transported along the axons. The total amount of a biosynthesis-linked enzyme present within a neuron soma may therefore match the target size. An estimate for this enzyme capacity of a neuron soma is obtained by multiplying the enzyme activity, which is mea- sured per unit volume, by the soma volume.
Previous measurements of the cross sectional area of HRP-traced motoneuron somata (de Graaf et al., 1990) show that the average soma volume of the W- MNs is approximately three times larger than that of the R/I-MNs. Average enzyme activities per unit volume differ between both populations by at most 25% (SDH, Table 2). The average capacity for each of the five enzymes is thus about three times larger in the W-MNs than in the WI-MNs. These differences corres- pond to differences in the average target size between W-MNs and WI-MNs.
[nnervation pattern and metabolic specialization The red muscle fibres are, like the intermediate muscle fibres, involved in continuous swimming at low speeds. The white fibres are activated at high speeds and for short periods (Johnston et al., 1977). The motoneurons in fish fire bursts of action potentials followed by silent periods. This enables the alternate activation of the left and right muscle. Differences among motoneurons in impulse frequency, that prob- ably relate to unit type have been observed (Roberts & Williamson, 1983). It is assumed that the energy requirement for a single burst will be higher in the W- MNs. First, they have a larger average soma size and therefore need more energy for membrane repolariza- tion. Second, it has been shown in mammals that the motoneurons of fast twitch units fire at higher frequencies than the motoneurons of slow twitch units (Kernell, 1979, 1986; Henneman & Mendell, 1981).
The major part of a neuron's energy intake is consumed in firing activity (Sokoloff, 1974). For reasons outlined above, it is assumed that the energy requirement for firing activity in the R/I-MNs is continuous and at a low level and in the W-MNs incidental but at a high level. It is unknown which of the two motoneuron types consumes the most energy for firing activity measured over larger periods of time (e.g. hours) under normal aquarium conditions. We will consider whether the differences in the pattern of impulse activity is reflected in the histochemical profiles. In doing so we regard enzymes with signi- ficantly correlating activities as functionally related. In
short this is the constant proportion hypothesis as put forward by Pette and Hoffer (1980).
R/I-MNs. There is a significant eorrela tion between HK and G6PDH activity in WI-MNs and no significant correlation between HK and PFK activity. Glucose is thus primarily of importance for biosynthetic proces- ses. The SDH activity is high (Table 2) and correlates closely with NADH-TR activity (Table 3a and Fig. 3a). This indicates a tight coupling of the Krebs cycle and electron transport chain. We conclude that in the WI- MNs, firing activity is fuelled by oxidative metabolism or, more accurately, by a joint activity of Krebs cycle electron transport chain.
W-MNs. The W-MNs have a low activity of SDH (Table 2), which is not positively correlated with NADH-TR activity. A coupling between Krebs cycle and electron transport chain is thus absent. We conclude that the firing activity in the W-MNs is not fuelled by a joint activity of Krebs cycle and electron transport chain. The correlation between HK and G6PDH activity is not significant. The metabolic flux from HK to G6PDH will therefore be smaller than in the WI-MNs. The correla- t-ion coefficient between HK and PFK activity is close to zero. The flux from HK directly to PFK is therefore low or insignificant. It is likely that glucose is temporarily stored in the form of glycogen. The physiological implications of this will be outlined below. In short, we suggest that the W-MNs rely on glycolysis starting from glycogen to sustain the firing activity; a high phosphorylase (E. C. 2.4.1.1) activity is thus expected.
Campa and Engel (1970, 1971) qualitatively determi- ned phosphorylase and SDH in spinal neurons in segment L7 of the cat. They reported a high phos- phorylase activity in the large neurons and a high SDH and low phosphorylase activity in the smaller and medium sized neurons. Sickles and McLendon (1983) determined phosphorylase quantitatively in rat lum- bosacral a-motoneurons and also reported the highest activity in the largest neurons.
Physiological implications One ATP molecule is lost for each molecule of glucose stored as glycogen (Stryer, 1980). Initiating glycolysis from glycogen however, enables, in the short time of a single burst, the generation of larger amounts of ATP; first, because glycolysis starts with G6P instead of glucose, second because, contrary to HK, phosphorylase is not influenced by product inhibition of G6P. A higher substrate concentration for PFK can therefore be built up, which results in a higher glycolytic rate (Shulman, 1988). Glycolysis has a cytosolic location. Hence it can be located in closer proximity to the plasma membrane than mitochond- rial enzymes. This may be advantageous for a local- ized, high energy release in a short interval. The high

Histochemical profiles of m o t o n e u r o n s wi th different activity pa t te rns 279
ene rgy r equ i remen t for repolar izat ion in the immed i - ate vicinity of the m e m b r a n e m a y have implicat ions for the distr ibution of e n z y m e s within the n eu ron soma. This will be the subject of future research.
In conclusion, a low a n d con t inuous ene rgy require- m e n t in the R/I-MNs is m e t b y oxidative me tabo l i sm or a joint activity of Krebs cycle and electron t r anspor t chain. We sugges t that a h igh and inf requent firing activity of the W-MNs is m e t by t e m p o r a r y s torage of glucose w h e r e b y a h igher rate of glycolysis can be achieved.
Acknowledgements
We thank Prof Dr B. L. Roberts a n d Dr C. J. F. van N o o r d e n for critical c o m m e n t s on the manuscr ip t .
References
BURKE, R. E (1981) Motor units: anatomy, physiology and functional organization. In Handbook of physiology, Sec- tion I. The Nervous System, Vot II. Motor systems (edited by BROOKS, V. B.) pp. 345--422. Washington: American Physiological Society,
C A M P A , I . F. & E N G E L , W. K. (1970) Histochemistry of motor neurons and interneurons in the cat lumbar spinal cord. Neurology 20, 559--68.
C A M P A , J. F. & E N G E L , W. K. (1971) Histochemical and functional correlations in anterior horn neurons of the cat spinal cord. Science 171, 198-9.
C U L L H E I M , S. & K E L L E R T H , J . - O . (1978) A morphological study of the axons and recurrent collaterals of cat a- motoneurones supplying different functional types of muscle unit. J. Physiol. 281, 301-13.
DE G R A A F , F . , V A N R A A M S D O N K , W . , V A N A S S E L T r E. &
OIEGENBACH, P. C. (1990) Identification of moto- neurons in the spinal cord of the zebrafish (Brachydanio rerio), with special reference to motoneurons that innervate intermediate muscle fibers. Anat. EmbryoI. 182, 93-102.
F L E S H M A N , J. W . , M U N S O N / J. B. , SYPERT, G. W. &
rRIEOMAN, W. A. (1981) Rheobase, input resistance, and motor-unit type in medial gastrocnemius moto- neurons in the cat. J. Neurophysiol. 46, 1326-38.
H E N N E M A N , E. & M E N D E L L , L. h i . (1981) Functional organization of motoneuron pool and its inputs. In Handbook of physiology, Section I. The Nervous System. VoI II. Motor systems (edited by B R O O K S , V. B.) pp. 423-507. Washington: American Physiological Society.
I S H I H A R A , A.~ N A I T O H , H . , A R A K I , H . & N I S H I H A R A , Y.
(1988) Soma size and oxidative enzyme activity of motoneurons supplying the fast twitch and slow twitch muscles in the rat. Brain Res. 446, 195-8.
J O H N S T O N , I . A . , D A V I S O N , W. & G O L D S P I N K , G. (1977) Energy metabolism of carp swimming muscle. J. Comp. Physiol. 114, 203-16.
KERNELL, D. (1979) Rhythmic properties of motoneurons innervating muscle fibres of different speed in m. gastrocnemius medialis of the cat. Brain Res. 160, 159- 62.
KERNELL, D. (1986) Organization and properties of spinal motoneurons and motor units. Prog. Brain Res. 64, 21- 30.
L O J D A , Z . , G O S S R A U , R. & S C H I E B E R , T. t I . (1976) Enzymhis- tochemische Methoden. Berlin: Springer Verlag.
I'ETTE, D. & HOttER, H. W. (1980) The constant proportion enzyme group concept in the selection of reference enzymes in metabolism. In Trends in Enzyme Histoche- mistry and Cytochemistry (edited by EVERED, D. a O'CONNOR, M.) pp. 231--44. Amsterdam: Excerpta Medica.
R O B E R T S , B. L. & W I L L I A M S O N , R. M. (1983) Motor pattern formation in the dogfish spinal cord. In Neural origin of rhythmic movements (edited by ROBERTS, A. & ROBERTS, B.) pp. 331--50. Cambridge: Society for Experimental Biology.
S C H M U E D , L. C. & F A L L O N , J. H . (1986) Fluoro-Gold: a n e w
fluorescent retrograde axonal tracer with numerous unique properties. Brain Res. 377, 147-54.
SHULMAN, R. G. (1988) High resolution NMR in vivo. Trends Biochem. Sci. 13, 37-9.
S I C K L E S , D. W. & M c L E N D O N , R. E. (1983) Metabolic variation among rat lumbosacral c~-motoneurons. His- tochemistry 79, 205-17.
S I C K L E S , D. W. & O B L A K , T. G. (1984) Metabolic variation among motoneurons innervating different muscle fibre types, I. Oxidative enzyme activity. J. Neurophysiol. 51, 529-37.
SOKOLOFr, L. (1974) Changes in enzyme activities in neural tissues with maturation and development of the ner- vous system. In The Neurosciences: Third Study Program. (edited by SCI-IMITT, V. O. & W O R D E N , F. G . ) p p . 885-98. Cambridge, MA, MIT Press.
S T R A A T S B U R G , I . H . , DE G R A A F , F . , V A N N O O R D E N , C. J . F.
a VAN RAAMSDONK, W. (1989) Enzyme reaction rate studies in electromotor neurons of the weakly electric fish Apteronotus leptorhynchus. Histochem. J. 21, 609-17.
STRYER, L. (1980) Biochemistry. 2nd edn. San Francisco: W. H. Freeman.
TSUJI, S. (1974) On the chemical basis of thiocholine methods for demonstration of acetylcholinesterase acti- vities. Histochemistry 42, "99-110.
U L F H A K E , S. & K E L L E R T H , J.-O. (1982) Does ~ -
motoneurone size correlate with motor unit type in cat triceps surae? Brain Res. 251, 201-9.
VAN NOOROEN, C. J. V. (1984) Histochemistly and cytoche- mistry of glucose-6-phosphate dehydrogenase. Prog. Histochem. Cytochem. 15, 1-85.
VAN NOORDEN, C. J. r. & VOCALS, I. M. C. (1989) Polyvinyl alcohol and other tissue protectants in enzyme histo- chemistry: a consumer's guide. Histochem. J. 21, 373-9.
V A N N O O R D E N , C. J . F . , B H A T T A C H A R Y A ~ R. D. & V O G E L S ,
I. M. C. (1983) Enzyme cytochemical staining of indi- vidual cells with the use of a polyacrylamide carrier. Acta Histochem 73, 71-8.
V A N R A A M S D O N K r W . , M O S t W . , S M I T - O N E L , M. J.~ V A N
DER L A A R S E , W. J . & F E H R E S , R. (1983) The develop- ment of the spinal motor column in relation to the myotomal muscle fibers in the zebrafish (Brachydanio rerio). I. Posthatching development. Anat. Embryol 167, 125-39.

280 DE GRAAF, VAN RAAMSDONK, VAN ASSELT and DIEGENBACH
VAN RAAMSDONK~ W.p SMIT-ONEL~ M.~ DONSELAARr Y. &
OIEC~NBACH, I". (1987) Quantitative cytochemical analysis of cytochrome oxidase and succinate dehydro- genase activity in spinal neurons. Acta Histochem. 81, 129-41.
WONG-RILEY, M. T. T. (1989) Cytochrome oxidase: an endogenous metabolic marker for neuronal activity. Trends Neurosci. 12, 94-101.
Related Documents











