Heterologous expression, purification, and properties of human cytochrome P450 27C1 Zhong-Liu Wu, Cheryl J. Bartleson, Amy-Joan L. Ham, F. Peter Guengerich * Department of Biochemistry and Center in Molecular Toxicology, Vanderbilt University School of Medicine, Nashville, TN 37232 0146, USA Received 27 October 2005, and in revised form 4 November 2005 Available online 28 November 2005 Abstract Cytochrome P450 (P450) 27C1 is one of the ‘‘orphan’’ P450 enzymes without a known biological function. A human P450 27C1 cDNA with a nucleotide sequence modified for Escherichia coli usage was prepared and modified at the N-terminus, based on the expect- ed mitochondrial localization. A derivative with residues 3–60 deleted was expressed at a level of 1350 nmol/L E. coli culture and had the characteristic P450 spectra. The identity of the expressed protein was confirmed by mass spectrometry of proteolytic fragments. The puri- fied P450 was in the low-spin iron state, and the spin equilibrium was not perturbed by any of the potential substrates vitamin D 3 ,1a- or 25-hydroxy vitamin D 3 , or cholesterol. P450s 27A1 and 27B1 are known to catalyze the 25-hydroxylation of vitamin D 3 and the 1a-hy- droxylation of 25-hydroxy vitamin D 3 , respectively. In the presence of recombinant human adrenodoxin and adrenodoxin reductase, recombinant P450 27C1 did not catalyze the oxidation of vitamin D 3 ,1a- or 25-hydroxy vitamin D 3 , or cholesterol at detectable rates. P450 27C1 mRNA was determined to be expressed in liver, kidney, pancreas, and several other human tissues. Ó 2005 Elsevier Inc. All rights reserved. Keywords: Cytochrome P450; P450 27C1; Adrenodoxin; Cholesterol; Vitamin D; Expression in Escherichia coli Cytochrome P450 1 enzymes are involved in the oxidation of a large variety of xenobiotic chemicals including drugs, carcinogens, pesticides, etc. [2]. However, more than one-half of the human P450s have characterized roles in the metabo- lism of endogenous chemicals or vitamins [3]. Approximate- ly, 13 of 57 human P450 genes do not have any defined functions in the oxidation of endogenous or xenobiotic chem- icals, and these can accordingly be termed ‘‘orphans’’ [4]. One approach to the characterization of the P450 orphans is the heterologous expression of large amounts of the proteins to facilitate studies on involvement in reactions, using approaches focused on candidate sub- strates inferred from knowledge about related P450s and also broader searches [4]. One of the orphan P450s is 27C1, which has no reported information at this time concerning its site of expression, or even if it is expressed [4]. On the basis of the sequence similarity with P450s 27A1 and 27B1, this might be expected to be a mito- chondrial protein with possible catalytic activity toward Vit D 3 or its hydroxylated forms [3,5–10]. We developed conditions for the high-level expression of a modified version of human P450 27C1 in Escherichia coli and its purification. The distribution of P450 27C1 mRNA in human tissues was analyzed, and the possibility that P450 27C1 is involved in Vit D 3 or cholesterol metabolism was examined. Materials and methods Chemicals Oligonucleotides for cDNA synthesis and N-terminal modifications were purchased from Operon (Huntsville, www.elsevier.com/locate/yabbi Archives of Biochemistry and Biophysics 445 (2006) 138–146 ABB 0003-9861/$ - see front matter Ó 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.abb.2005.11.002 * Corresponding author. Fax: +1 615 322 3141. E-mail address: [email protected] (F.P. Guengerich). 1 Abbreviations used: Adr, NADPH-adrenodoxin reductase; Adx, adre- nodoxin; LB, Luria–Bertani medium; NTA, nitrilotriacetate; OH, hydro- xy; (OH) 2 , dihydroxy; P450, cytochrome P450 (also termed ‘‘heme-thiolate protein P450’’ [1]); PAGE, polyacrylamide gel electrophoresis; PCA, polymerase cycling assembly; SDS, sodium dodecyl sulfate; TB, Terrific broth (medium); Vit D 3 , cholecalciferol (vitamin D 3 ).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/yabbi
Archives of Biochemistry and Biophysics 445 (2006) 138–146
ABB
Heterologous expression, purification, and propertiesof human cytochrome P450 27C1
Zhong-Liu Wu, Cheryl J. Bartleson, Amy-Joan L. Ham, F. Peter Guengerich *
Department of Biochemistry and Center in Molecular Toxicology, Vanderbilt University School of Medicine, Nashville, TN 37232 0146, USA
Received 27 October 2005, and in revised form 4 November 2005Available online 28 November 2005
Abstract
Cytochrome P450 (P450) 27C1 is one of the ‘‘orphan’’ P450 enzymes without a known biological function. A human P450 27C1cDNA with a nucleotide sequence modified for Escherichia coli usage was prepared and modified at the N-terminus, based on the expect-ed mitochondrial localization. A derivative with residues 3–60 deleted was expressed at a level of 1350 nmol/L E. coli culture and had thecharacteristic P450 spectra. The identity of the expressed protein was confirmed by mass spectrometry of proteolytic fragments. The puri-fied P450 was in the low-spin iron state, and the spin equilibrium was not perturbed by any of the potential substrates vitamin D3, 1a- or25-hydroxy vitamin D3, or cholesterol. P450s 27A1 and 27B1 are known to catalyze the 25-hydroxylation of vitamin D3 and the 1a-hy-droxylation of 25-hydroxy vitamin D3, respectively. In the presence of recombinant human adrenodoxin and adrenodoxin reductase,recombinant P450 27C1 did not catalyze the oxidation of vitamin D3, 1a- or 25-hydroxy vitamin D3, or cholesterol at detectable rates.P450 27C1 mRNA was determined to be expressed in liver, kidney, pancreas, and several other human tissues.� 2005 Elsevier Inc. All rights reserved.
Keywords: Cytochrome P450; P450 27C1; Adrenodoxin; Cholesterol; Vitamin D; Expression in Escherichia coli
Cytochrome P4501 enzymes are involved in the oxidationof a large variety of xenobiotic chemicals including drugs,carcinogens, pesticides, etc. [2].However,more thanone-halfof the human P450s have characterized roles in the metabo-lism of endogenous chemicals or vitamins [3]. Approximate-ly, 13 of 57 human P450 genes do not have any definedfunctions in theoxidationofendogenousorxenobiotic chem-icals, and these can accordingly be termed ‘‘orphans’’ [4].
One approach to the characterization of the P450orphans is the heterologous expression of large amountsof the proteins to facilitate studies on involvement inreactions, using approaches focused on candidate sub-
0003-9861/$ - see front matter � 2005 Elsevier Inc. All rights reserved.
doi:10.1016/j.abb.2005.11.002
* Corresponding author. Fax: +1 615 322 3141.E-mail address: [email protected] (F.P. Guengerich).
1 Abbreviations used: Adr, NADPH-adrenodoxin reductase; Adx, adre-nodoxin; LB, Luria–Bertani medium; NTA, nitrilotriacetate; OH, hydro-xy; (OH)2, dihydroxy; P450, cytochrome P450 (also termed ‘‘heme-thiolateprotein P450’’ [1]); PAGE, polyacrylamide gel electrophoresis; PCA,polymerase cycling assembly; SDS, sodium dodecyl sulfate; TB, Terrificbroth (medium); Vit D3, cholecalciferol (vitamin D3).
strates inferred from knowledge about related P450sand also broader searches [4]. One of the orphan P450sis 27C1, which has no reported information at this timeconcerning its site of expression, or even if it is expressed[4]. On the basis of the sequence similarity with P450s27A1 and 27B1, this might be expected to be a mito-chondrial protein with possible catalytic activity towardVit D3 or its hydroxylated forms [3,5–10].
We developed conditions for the high-level expression ofa modified version of human P450 27C1 in Escherichia coli
and its purification. The distribution of P450 27C1 mRNAin human tissues was analyzed, and the possibility thatP450 27C1 is involved in Vit D3 or cholesterol metabolismwas examined.
Materials and methods
Chemicals
Oligonucleotides for cDNA synthesis and N-terminalmodifications were purchased from Operon (Huntsville,

Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146 139
AL, USA) in either 96-well format, at 10-nmol scale in wetform (normalized to 100 lM each), or tube format, at50 nmol scale. All oligonucleotides were of salt-free qualityand used directly without further purification. Vit D3, 25-OH Vit D3, 1a, 25-(OH)2 Vit D3, and cholesterol were pur-chased from Sigma-Aldrich (St. Louis, MO, USA), and 1a-OH Vit D3 was obtained from Calbiochem-EMD Biosci-ences (La Jolla, CA, USA). [4-14C]Cholesterol (53 mCi/mmol) was purchased from Perkin-Elmer (Boston, MA,USA). All other reagents and solvents were obtained fromgeneral commercial suppliers and used without furtherpurification.
Optimization of the P450 27C1 nucleotide sequence forheterologous expression and oligonucleotide design for the
synthesis of P450 27C1 cDNA
Automatic codon optimization and oligonucleotidedesign for polymerase chain reaction (PCR)-based genesynthesis were performed using DNAWorks 2.4 [11], devel-oped by the Center for Information Technology, NationalInstitutes of Health, Department of Health and HumanServices (http://molbio.info.nih.gov/dnaworks).
The amino acid sequence and the native cDNAsequence information for human P450 27C1 were obtainedfrom D. R. Nelson�s P450 homepage (http://drnelson.utmem.edu/human.P450.seqs.html) and assembled fromGenBank sequence AC027142. An EcoRI restriction sitewas selected from the native cDNA sequence and used tomanually separate the full-length gene into two synthonsencoding amino acids 6–288 (Synthon 1) and 289–542(Synthon 2). In addition, a (His)5 tag was added to theC-terminal to facilitate purification. A series of overlap-ping 40- to 50-mers were prepared for each synthon (seeSupplementary Material), based on the DNAWorks results(vide supra).
The codons were optimized automatically to suit thecodon preference bias of E. coli. The codon frequencythreshold was set to be 20% in the program. Overlap melt-ing temperatures were designed to be 65 ± 3 �C. The 5 0-and 3 0-flanking sequences for Synthon 1 were 5 0-TAGGAGGTCATATGGCTTTATTAGCACGTATT-3 0 (sense)and 5 0-GAATTCTGC-3 0 (sense) respectively, which con-tained an NdeI restriction site and AT enrichment for thecodons of the first seven amino acids on the 5 0 end andan EcoRI restriction site on the 3 0 end. The flankingsequences for Synthon 2 were 5 0-CGCGAATTCTGC-3 0
(sense) and 5 0-TAATCTAGAGGTA-3 0 (sense), whichcontained an EcoRI restriction site on the 5 0 end and anXbaI restriction site on the 3 0 end. Other than that, allDNA sequences were designed to lack the NdeI, XbaI,and EcoRI sites needed for subsequent manipulation.
PCR-based gene synthesis
Ten microliters of each of the oligonucleotide compo-nents (normalized to 100 lM) of Synthons 1 and 2 were
added into two separate pools and diluted to 2 lMeach.
Polymerase cycling assembly was carried out in a 50-ll reaction mixture: 2.5 U PfuUltra High-Fidelity DNApolymerase (Stratagene, La Jolla, CA, USA), 0.2 mMeach dNTP, and 5.0 ll of 10· PfuUltra buffer. Thermalcycling began with a 5-min denaturing step at 95 �Cand was followed by 25 cycles at 95 �C for 30 s, 60 �Cfor 30 s, and 72 �C for 90 s, with a final extension timeof 10 min.
Polymerase chain reaction amplification mixtures(50 ll) contained 2.5 U of PfuUltra High-Fidelity DNApolymerase, 0.2 mM each dNTP, 5.0 ll of 10· PfuUltra
buffer, 1 ll of each PCA product, and 0.4 lM of eachof the outermost oligonucleotides. The thermal cyclingparameters were essentially the same as for PCA, exceptthat the number of cycles was raised to 30.
The 866- and 796-bp amplified PCR fragments of Syn-thons 1 and 2 were purified by agarose gel electrophoresisand double digested with NdeI and EcoRI and with XbaIand EcoRI, respectively. Both fragments with cohesiveends were ligated together in a ‘‘monocistronic’’ pCW vec-tor (not coding for NADPH-P450 reductase [12]) anddigested with NdeI and XbaI. The ligation mixture wastransformed into E. coli DH5a competent cells and selectedon LB plates with 50 lg/ml ampicillin. The plasmids werepurified using a QIAprep Miniprep kit (Qiagen, Valencia,CA, USA) and screened by restriction digest analysis withNdeI and XbaI. The plasmids (construct 1) that yielded1.6 kb fragments were sequenced in both the sense andthe antisense strands in the Vanderbilt facility with anApplied Biosystems Model 3700 fluorescence sequencingunit using a Taq dye terminator kit (PE Applied Biosys-tems, Foster City, CA, USA).
N-Terminal modifications
N-Terminal mutations were introduced into native con-struct (pCW 27C1) by PCR-based mutagenesis. The N-ter-minal segment of 27C1 was amplified between the NdeI andthe EcoRI sites using 5 0 PCR primers containing thedesired mutations and a complementary 3 0 PCR primer(5 0-GCAGAATTCGCGCCACGGCTTTGGGATAAATGGACGTAAC-3 0), which was also the 3 0 outmost oligo-nucleotide of Synthon 1. The mutagenic primers for con-structs 2 and 3 were 5 0-GTCTGCCACATATGGCTCGTGCAGAAGATAAAGGTGCAGGCCGTCCAGGTAG-3 0 and 5 0-GTG GTGGTCATATGGCTGGTCCACGTAGTCTGGCGCATGCCAGGTCCAC-3 0, respectively.PfuUltra High-Fidelity DNA polymerase was used forthe PCR amplification at an annealing temperature of60 �C. The products were purified by agarose gel electro-phoresis, double digested with NdeI and EcoRI and ligatedwith the 5.8 kb fragment derived from the digestion of con-struct 1 with the same enzymes. The modifications wereconfirmed by sequencing the open reading frame regionsof the new constructs.

140 Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146
Expression of P450 27C1 and groEL/ES in E. coli DH5a
Both plasmids pGro12 ES/EL [13] and each of the threeconstructs of pCW 27C1 were transformed into E. coli
DH5a competent cells and selected on LB plates contain-ing 50 lg/ml ampicillin and 20 lg/ml kanamycin. Singlecolonies were grown overnight in LB media fortified with100 lg/ml ampicillin and 50 lg/ml kanamycin, at 37 �Cwith gyrorotary shaking at 250 rpm; 0.5 ml of each over-night culture was then inoculated into 50 ml of TB brothcontaining 100 lg/ml ampicillin, 50 lg/ml kanamycin,and 0.025% (v/v) of a mixture of trace elements [14] in a250 ml flask. The cultures were incubated at 37 �C withgyrorotary shaking at 220 rpm for 4 h; induction of P45027C1 and groEL/ES transcription was then initiated withthe addition of 1.0 mM IPTG and 1 mg/ml arabinose,respectively. 5-Aminolevulinic acid was also supplementedsimultaneously, at a final concentration of 0.5 mM. Theincubation continued at 27 �C with gyrorotary shaking at200 rpm for another 48 h, and the expression level ofP450 27C1 was monitored at 20, 24, 40, 42, and 46 h.
Large scale expression for construct 3 was performed in2.8-L Fernbach flasks with 500 ml media, essentially in thesame manner as that for small scale trials, except that afterinduction the cultures were incubated at 27 �C with gyroro-tary shaking at 190 rpm for 40 h in a New Brunswick Inno-va 4300 shaker (New Brunswick Scientific, Edison, NJ,USA).
Purification of recombinant P450 27C1
All purification steps were performed at 4 �C. E. colimembranes were prepared as described elsewhere [15,16].The membrane fractions (from a 1-L culture) were dilut-ed in buffer A, which consisted of 20 mM potassiumphosphate (pH 7.4), 20% glycerol (v/v), 0.625% sodiumcholate (w/v), 0.625% Tergitol NP-10 (w/v), 0.1 mMdithiothreitol, and 3 mM magnesium acetate. The mix-ture was stirred for 4 h and centrifuged at 105· g for30 min. The resulting supernatant, with KCl added toa final concentration of 0.5 M, was loaded onto aNi2+–NTA agarose column (Qiagen; 5 ml packed-bedvolume), equilibrated with 10 column volumes of bufferA plus 0.5 M KCl and 5 mM imidazole (buffer B). Afterwashing of the column with 10 volumes of buffer B,P450 27C1 was eluted at a flow rate of 0.5 ml/min withbuffer B containing 100 mM imidazole.
To remove the detergents, the eluted reddish fractionswere applied to a Biogel HTP hydroxylapatite column(10 ml packed-bed volume; Bio-Rad, Richmond, CA,USA) equilibrated with 5 column volumes of 20 mM potas-sium phosphate buffer (pH 7.7) containing 20% glycerol (v/v) (buffer C) and washed with buffer C until the A280 of theeffluent was <0.02. P450 27C1 was then eluted by raisingthe potassium phosphate concentration to 0.7 M, withsodium cholate present at a final concentration of 0.2%.The eluted P450 27C1 was dialyzed overnight against 40
volumes of 100 mM potassium phosphate (pH 7.4) con-taining 20% glycerol (v/v) and 0.1 mM EDTA to removecholate.
Analysis of amino acid sequence by mass spectrometry
The purified P450 27C1 protein was exchanged fromstorage buffer to digestion buffer using a 30-kDa UltrafreeMC regenerated cellulose filter (Millipore), and the pro-teins were subsequently digested directly off the filter aspreviously described [17]. The proteins were subjected todigestion with either trypsin (37 �C for 30 min, 2 h, or18 h) or chymotrypsin (4 h, room temperature). Liquidchromatography–mass spectrometry (LC–MS) analysis ofthe resulting peptides was performed using a ThermoFinn-igan LTQ ion trap mass spectrometer equipped with aThermo MicroAS autosampler and Thermo SurveyorHPLC pump, Nanospray source, and Xcalibur 1.4 instru-ment control (Thermo Electron, San Jose, CA, USA).The peptides were separated on a packed capillary tip,100 lm · 11 cm, with C18 resin (Monitor C18, 5 lm,100 A; Column Engineering, Ontario, CA, USA) usingan in-line solid phase extraction column (100 lm · 6 cm)packed with the same C18 resin (using a frit generated fromliquid silicate Kasil 1 [18] similar to that previouslydescribed [19], except that the flow from the HPLC pumpwas split prior to the injection valve. The flow rate duringthe solid-phase extraction of the gradient was 1 ll/min andduring the separation phase was 700 nl/min. Mobile phaseA was 0.1% aqueous HCO2H (v/v) and mobile phase Bconsisted of CH3CN with 0.1% HCO2H (v/v). A 95-mingradient was performed with a 15-min washing period(100% A for the first 10 min followed by a gradient to98% A, v/v, at 15 min) to allow for solid-phase extractionand removal on any residual salts. After the initial washingperiod, a 60-min gradient was performed where the first35 min was a slow, linear gradient from 98% A to 75%A, v/v, followed by a faster gradient to 10% A at 65 minand an isocratic phase at 10% A (v/v) to 75 min. TheMS/MS spectral analysis of the peptides was performedusing data-dependent scanning in which one full MS spec-tra, using a full mass range of 400–2000 amu, was followedby three MS/MS spectra. Peptides were identified using theSEQUEST algorithm [20] and the SEQUEST Browsersoftware in the Bioworks 3.1 software package (ThermoElectron), using the human subset of the Uniref100 data-base to which the P450 27C1 protein sequence was addedand all redundant sequences to this protein were removed.
Analysis of mRNA expression
The amino acid sequences of all 57 human P450s werealigned to determine the best region for probe design usingMacVector 8.0 (Accelrys, Burlington, MA, USA). On thebasis of this alignment, nucleotide probes were designedfor P450s 27A1, 27B1, and 27C1 (Table 1). All three oligo-nucleotides were synthesized and purified using PAGE

Table 1Sequences of oligonucleotides used in hybridization analysis
27A1: 5 0-TGACTTTATGACTCGACTGGACCAGCTGCGGGCAGAGAGTGCTTCGGGGAAC-3 0
27B1: 50-TCGGGACGTGGCGGGGGAATTTTACAAGTTCGGACTGGAAGGCATCGCCGCG-3 0
27C1: 50-TGGGTGCCTTGCCAGGAGGTACACAGTCCAAGACAAGGTGAAGGACGTCGTG-3 0
Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146 141
(Operon). A control nitrocellulose BA-S83 sheet (Schlei-cher & Schuell, Keene, NH, USA) was manually spottedwith the 27A1, 27B1, and 27C1 complementary oligonucle-otides. The oligonucleotides were fixed to the nitrocelluloseas described elsewhere [21]. The 32P-labeled P450 27C1probe was generated using a RediPrime II random prime-labeling system (Amersham Biosciences, Piscataway, NJ,USA). The control nitrocellulose and the human MultipleTissue Expression array (BD Biosciences-Clontech, PaloAlto, CA, USA) were hybridized with the 32P-labeledP450 27C1 probe (Table 1) according to the manufacturer�sinstructions. Briefly, the blot/array was blocked withExpressHyb solution containing 1.5 mg of sheared salmontestis DNA for 30 min at 60 �C. Fresh ExpressHyb solutioncontaining newly prepared 32P-labeled P450 27C1 probereplaced the blocking solution, and the blot/array wasincubated overnight at 60 �C. The next day the blot/arraywas washed four times at room temperature with 2· SSC(30 mM sodium citrate buffer, pH 7.0, plus 300 mM NaCl)containing 1% SDS (w/v). The final wash was conducted at60 �C. The blot and array were developed using a Molecu-lar Imager FX phosphorimager (Bio-Rad).
Enzyme assays
Assays of cholesterol oxidation were performed using ageneral procedure described elsewhere [22,23]. PurifiedP450 27C1 (0.1–0.3 lM) or P450 27A1 (0.1–0.25 lM) wasmixed with E. coli recombinant mouse Adx (2.0–5.0 lM)and E. coli recombinant bovine Adr (0.2–0.5 lM) (bothwere generous gifts of I. A. Pikuleva, University of Texas,Galveston, TX) in 1.0 ml of 50 mM potassium phosphatebuffer (pH 7.4) containing 1 mM of EDTA and either50 lM cholesterol together with 3 ll [4-14C]cholesterol(53 mCi/mmol, stock 40 lCi/ml in C2H5OH, 2.66 · 105
dpm total) or Vit D3, 1a-OH Vit D3, or 25-OH Vit D3.Reactions (at 37 �C) were initiated by the addition of anNADPH-generating system composed of (final concentra-tions of) 0.5 mM NADPH, 10 mM glucose 6-phosphate,and 1.0 IU yeast glucose 6-phosphate dehydrogenase permilliliter [24]. Reactions were quenched after 30 min bythe addition of 1.0 ml CH2Cl2 and mixing with a vortexdevice. The layers were separated by centrifugation(3 · 103· g) and the lower organic layer was removed.The CH2Cl2 extraction step was repeated. The combinedorganic extracts were dried under an N2 stream, dissolvedin CH3OH and subjected to high-performance liquidchromatography (HPLC). HPLC was performed on a6.2 mm · 80 mm Zorbax C8 column (3 lm; Agilent Tech-nologies, Palo Alto, CA, USA) with a CH3OH/H2O mobilephase at a flow rate of 1 ml/min with a 90–100% CH3OH
linear gradient over 10 min (in H2O, v/v) and 100%CH3OH for 10 min. Detection was at 265 nm with a Hew-lett-Packard 1040A diode array detector (Hewlett-Packard,Palo Alto, CA, USA) for Vit D3, 1a-OH Vit D3, and 25-OH Vit D3 products, or with a 2B b-RAM flow counter(IN/US Systems, Tampa, FL, USA) for [4-14C]cholesterolproducts.
Other assays and methods
P450 concentrations were estimated as described byOmura and Sato [25] using an OLIS/Aminco DW2a spec-trophotometer (OLIS, Bogart, GA, USA). Interactions ofpotential ligands with P450 27C1 were examined by addingcompounds dissolved in CH3OH (Vit D3 and 25-OH VitD3) or in a 45% aqueous solution of 2-hydroxypropyl-b-cy-clodextrin (for cholesterol) to the enzyme [22,23] andobserving spectral changes in the region of 350–500 nm[26].
Protein concentrations were estimated using a bicinch-oninic acid method following the manufacturer�s instruc-tions (Pierce Chemical, Rockford, IL, USA).
Sodium dodecyl sulfate–PAGE was performed usingReady Gel Tris–HCl gels (4–20% gradient, w/v; Bio-Rad)and stained with Bio-Safe Coomassie stain (Bio-Rad) fol-lowing the manufacturer�s instructions.
Results and discussion
Synthesis of codon-optimized P450 27C1 cDNA
At the start of this project, no P450 27C1 cDNA wasavailable and no information existed about sites of expres-sion. cDNA was prepared by PCA of overlapping oligonu-cleotides [27]. Because of the plan to express the cDNA inE. coli, the nucleotide sequence was optimized for codonusage in the process [11,28,29]. To improve the score gen-erated by DNAWorks 2.4 (judged by various factorsincluding possible formation of hairpins, mispriming, andrepeating regions, which can affect the error rate of genesynthesis especially for longer cDNAs), the whole cDNAsequence of P450 27C1 was manually divided into two syn-thons at an EcoRI restriction site. This EcoRI site is locat-ed in the middle of the P450 27C1 cDNA (862–867) andallows the formation of two synthons with sizes <1 kb thatcan be joined together. The amino acids encoded by thisregion (GAATTC) are Glu and Phe, with respective codonfrequencies of 67 and 42% in E. coli (http://molbio.info.nih.gov/dnaworks/dnaworks_CFT.html), which, withoutbeing subjected to codon optimization, presumably shouldnot have a major effect on the whole-codon optimization

ig. 2. Agarose gel electrophoresis of the gene synthesis products ofynthons 1 and 2. Lanes 1 and 4, markers; lane 2, Synthon 1; lane 3,ynthon 2.
142 Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146
process. The alignment of the final optimized cDNAsequence is compared with the native sequence of P45027C1 (Fig. 1).
The desired oligonucleotide size was set in 40- to 50-merrange. The designs obtained with the best overall score ofzero and the shortest lengths of oligonucleotides were cho-sen for both Synthons 1 and 2 (see Supplementary Materialfor oligonucleotide sequences). The oligonucleotides foreach synthon were assembled by PCA and amplified bysubsequent PCR. A dominant single band of the correctsize was achieved for both synthons when the oligonucleo-tide mixture was used at final concentration of 0.4 and0.2 lM for Synthons 1 and 2, respectively, as shown bygel electrophoresis (Fig. 2).
The two synthetic cDNA fragments were intergratedtogether into the expression vector pCW�ori and subjectedto sequencing. Of the four plasmids sequenced, three had asingle point mutation and one had a perfect sequence,which gave an overall error rate of 0.46 errors/kb for thefour, comparable to previously reported results for PCR-based gene synthesis [11,30–33]. The plasmid with the per-fect P450 27C1 sequence was used directly in subsequentexperiments.
An attempt to combine PCA and PCR together in onestep by including 0.4 lM of each of the outermost oligonu-cleotides in the PCA reaction mixture was successful and
Fig. 1. Modifications introduced into P450 27C1 cDNA to optimize codonnucleotide sequence predicted from genomic DNA; lower line, nucleotide sequ
FSS
yielded one distinct band of the desired size, but the errorrate was not examined.
Expression of N-terminal variants
Previous work on the expression of mammalian micro-somal or mitochondrial P450s in E. coli has shown thatin most situations the N-terminus of the native protein
usage in E. coli. Top line, predicted amino acid sequence; middle line,ence optimized for E. coli expression.
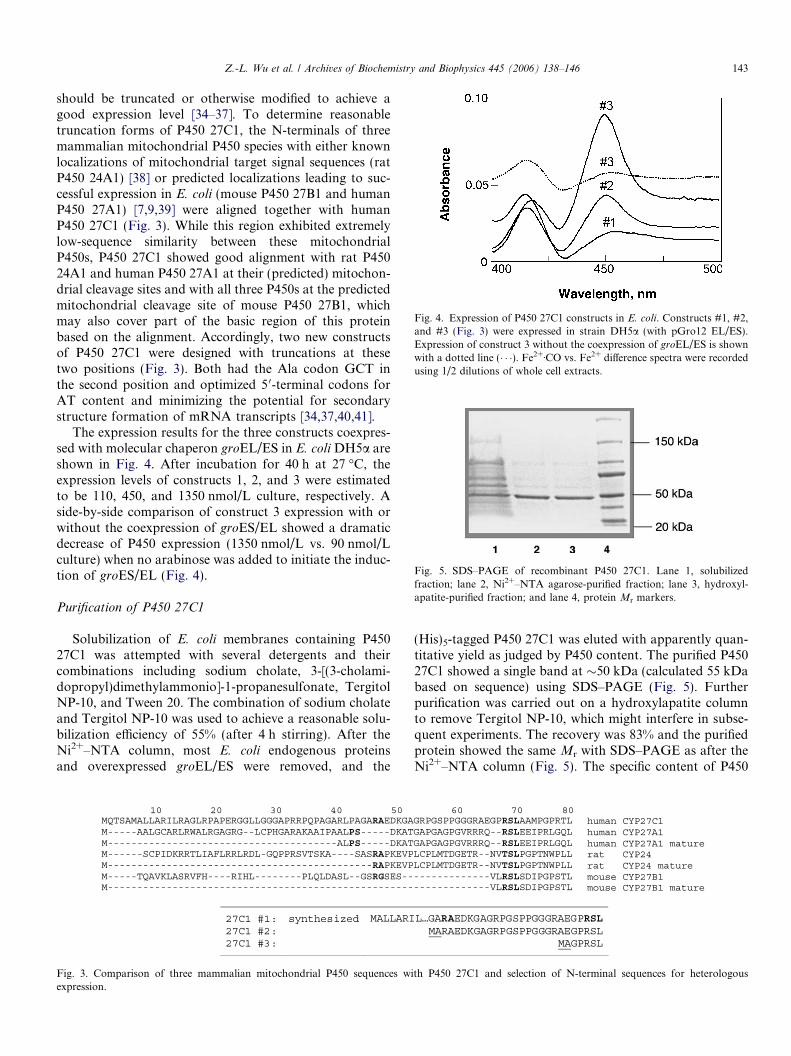
Fig. 4. Expression of P450 27C1 constructs in E. coli. Constructs #1, #2,and #3 (Fig. 3) were expressed in strain DH5a (with pGro12 EL/ES).Expression of construct 3 without the coexpression of groEL/ES is shownwith a dotted line (� � �). Fe2+ÆCO vs. Fe2+ difference spectra were recordedusing 1/2 dilutions of whole cell extracts.
Fig. 5. SDS–PAGE of recombinant P450 27C1. Lane 1, solubilizedfraction; lane 2, Ni2+–NTA agarose-purified fraction; lane 3, hydroxyl-apatite-purified fraction; and lane 4, protein Mr markers.
Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146 143
should be truncated or otherwise modified to achieve agood expression level [34–37]. To determine reasonabletruncation forms of P450 27C1, the N-terminals of threemammalian mitochondrial P450 species with either knownlocalizations of mitochondrial target signal sequences (ratP450 24A1) [38] or predicted localizations leading to suc-cessful expression in E. coli (mouse P450 27B1 and humanP450 27A1) [7,9,39] were aligned together with humanP450 27C1 (Fig. 3). While this region exhibited extremelylow-sequence similarity between these mitochondrialP450s, P450 27C1 showed good alignment with rat P45024A1 and human P450 27A1 at their (predicted) mitochon-drial cleavage sites and with all three P450s at the predictedmitochondrial cleavage site of mouse P450 27B1, whichmay also cover part of the basic region of this proteinbased on the alignment. Accordingly, two new constructsof P450 27C1 were designed with truncations at thesetwo positions (Fig. 3). Both had the Ala codon GCT inthe second position and optimized 5 0-terminal codons forAT content and minimizing the potential for secondarystructure formation of mRNA transcripts [34,37,40,41].
The expression results for the three constructs coexpres-sed with molecular chaperon groEL/ES in E. coli DH5a areshown in Fig. 4. After incubation for 40 h at 27 �C, theexpression levels of constructs 1, 2, and 3 were estimatedto be 110, 450, and 1350 nmol/L culture, respectively. Aside-by-side comparison of construct 3 expression with orwithout the coexpression of groES/EL showed a dramaticdecrease of P450 expression (1350 nmol/L vs. 90 nmol/Lculture) when no arabinose was added to initiate the induc-tion of groES/EL (Fig. 4).
Purification of P450 27C1
Solubilization of E. coli membranes containing P45027C1 was attempted with several detergents and theircombinations including sodium cholate, 3-[(3-cholami-dopropyl)dimethylammonio]-1-propanesulfonate, TergitolNP-10, and Tween 20. The combination of sodium cholateand Tergitol NP-10 was used to achieve a reasonable solu-bilization efficiency of 55% (after 4 h stirring). After theNi2+–NTA column, most E. coli endogenous proteinsand overexpressed groEL/ES were removed, and the
Fig. 3. Comparison of three mammalian mitochondrial P450 sequences wiexpression.
(His)5-tagged P450 27C1 was eluted with apparently quan-titative yield as judged by P450 content. The purified P45027C1 showed a single band at �50 kDa (calculated 55 kDabased on sequence) using SDS–PAGE (Fig. 5). Furtherpurification was carried out on a hydroxylapatite columnto remove Tergitol NP-10, which might interfere in subse-quent experiments. The recovery was 83% and the purifiedprotein showed the same Mr with SDS–PAGE as after theNi2+–NTA column (Fig. 5). The specific content of P450
th P450 27C1 and selection of N-terminal sequences for heterologous

Fig. 7. Spectra of purified P450 27C1. (A) Absorbance spectra of P45027C1 (‘‘absolute spectra’’). The P450 concentration was 2.0 lM. Thespectra are offset for clarity. (B) Second derivative of absorbance spectrum(ferric state). (C) Fe2+ÆCO vs. Fe2+ difference spectrum. The P450concentration was 2.4 lM (kmax = 449 nm).
144 Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146
27C1 was estimated to be 14.7 nmol/mg protein at the finalstep, based on the colorimetric protein estimation method.
Purified P450 27C1 was digested with trypsin and withchymotrypsin (individual digests), and the products wereanalyzed by LC–MS. Of the expected petptides, 90% couldbe identified (Fig. 6). The N-terminus was not detected inthe trypsin digest, but the peptides AGPRSLAAMPGPR-TLANLAEFFR and AGPRSLAAM*PGPRTL (whereM* is Met sulfoxide) were identified in the chymotrypsindigest. (We presume that the Met sulfoxide is an artifact ofthe processing.) These peptides correspond to the expectedN-terminusMAGPRSL. . . (Fig. 3) with loss of the N-termi-nal Met due to aminopeptidase activity, which is expectedwhen Ala follows (N-formyl)Met in bacteria [42,43].
Spectral properties of P450 27C1
The purified enzyme showed an absorbance spectrumwith wavelength maxima at 417, 532, and 566 nm for theferric form (Fig. 7). Second-derivative analysis of the Soretpeak, using the OLIS software, indicated that �90% of theiron is in the low-spin state (Fig. 6B), as judged by the rel-ative intensities of the bands at 390 and 418 nm [44,45].
The ferrous enzyme had a Soret maximum at 414 nmand the ferrous-CO complex at 448 nm (Fig. 6A). The kmax
in the typical ferrous-CO vs. ferrous spectrum was at 449nm (Fig. 6C).
The effects of potential substrates on the spectrum weremeasured, in light of the possibility that a low- to high-spiniron transition might be observed and associated with sub-strate binding [26]. Because of the literature indicating thatVit D and cholesterol are substrates for P450s 27A1 and27B1 [3,6], studies were carried out with varying concentra-tions (maximum indicated in parentheses) of cholesterol(30 lM, added in 45% aqueous 2-hydroxypropyl-b-cyclo-dextrin), Vit D3 (cholecalciferol, 100 lM), 25-OH Vit D3
(50 lM), or 1a-OH Vit D3 (50 lM), but no changes wereobserved (other than dilution) (results not shown).
Attempts to identify catalytic activities of P450 27C1
The coding sequence of P450 27C1 and comparisonswith P450s 27A1 and 27B1 indicate that this should havea mitochondrial localization. Therefore, the expectedelectron transfer partner should be Adx. Recombinant
MAGPRSLAAMPGPRTLANLAEFFCRDGFSRIHE
AQVLRAEGAAPQRANMESWREYRDLRGRATGL
EVNQVIADLIKRIYLLRSQAEDGETVTNVNDLFFK
ALELMFSMFKTSMYAGAIPRWLRPFIPKPWREF
RVSGGLLTYLFLSQALTLQEIYANVTEMLLAGVD
GERHVPTAADVPKVPLVRALLKETLRLFPVLPGN
QDENFPRAKEFRPERWLRKGDLDRVDNFGSIPF
SSQTNAVHAKTHGLLTPGGPIHVRFVNRKHHHH
Fig. 6. MS analysis of peptide digests of P450 27C1. Peptides identified areexperiments with the trypsin and chymotrypsin digests are combined here.
Adr and Adx supported the oxidation of cholesterol andof 1-OH Vit D3 by recombinant P450 27A1 (all proteinswere kindly provided by I. A. Pikuleva, University of Tex-as) (Supplementary Material, Fig. S1). (The products werenot identified but are presumed to be 27-OH cholesteroland 1a,25-(OH)2 Vit D3, respectively [3,6,7].) No detectableoxidation of cholesterol or 1-OH Vit D3 was observed withpurified P450 27C1 under identical conditions (Supplemen-tary Material, Fig. S1), with a limit of detection of0.01 nmol product formed/min/nmol P450.
Vit D3 and 25-OH Vit D3 were also examined as poten-tial substrates (at concentrations of 50 lM each), using thesame Adx/Adr system. The P450 27C1 system was tested
IQQKHTREYGKIFKSHFGPQFVVSIADRDMV
ISAEGEQWLKMRSVLRQRILKPKDVAIYSG
YSMEGVATILYESRLGCLENSIPQLTVEYIE
CRSWDGLFKFSQIHVDNKLRDIQYQMDRGR
TTSFTLSWTVYLLARHPEVQQTVYREIVKNL
GRVTQEDLVIGGYLIPKGTQLALCHYATSY
GHGVRSCIGRRIAELEIHLVVIQLLQHFEIKT
H
shown as underlined bold letters. The results of the LC–MS-electrospray

Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146 145
under same conditions as the positive control (P450 27A1)in the previous system. No activity was detected with anyof these substrates, at the limit of �0.01 nmol productformed/min/nmol P450.
Analysis of expression of P450 27C1 mRNA in humantissues
A probe was constructed to differentiate P450 27C1mRNA from P450s 27A1 and 27B1 (see Materials andmethods) and a control experiment established the selectiv-ity (Fig. 8). The 27C1 probe (32P-labeled) was used with acommercial source of mRNA samples from differenthuman tissues (Fig. 8). The results show the presence ofmRNA in liver, kidney, and pancreas, plus less in lung,ovary, adrenal, thyroid, salivary gland, mammary gland,and several fetal tissues, with much less in cancer tissues.
These results for P450 27C1 can be compared with theknown localizations of P450 27A1 mRNA in liver (plusleukocytes, skin fibroblasts, kidney, and arterial wall) andof P450 27B1 in kidney (plus skin, lymph nodes, colon,pancreas, adrenal, brain, placenta, cervix, and parathyroidgland) [3]. Thus, P450 27A1, 27B1, and 27C1 genes appearto be transcribed in many tissues.
Fig. 8. Analysis of sites of P450 27C1 mRNA expression in individualhuman tissues. Both the nitrocellulose blot (Part a) and the mRNA array(Part b) were treated and developed according to the protocol described inMaterials and methods. (a) Stringency conditions were developed tominimize hybridization with the corresponding regions of the P450 27A1and 27B1 cDNAs. A 32P-labeled oligonucleotide (52-mer, Table 1) wasselected for optimal recognition of P450 27C1 mRNA, using thecomplementary sections of the P450 27A1, 27B1, and 27C1 cDNAsshown in Table 1, with (A) 27A1 oligonucleotide (18 ng), (B), 27B1oligonucleotide (23 ng), and (C), 27C1 oligonucleotide (28 ng). (b) Acommercial blot with mRNA samples from individual human tissues wasprobed: A1, kidney, A6, lymph node; B1, lung; B6, ovary; C1, liver; C2,pancreas; C3, adrenal gland; C4, thyroid gland; C5, salivary gland; C6,mammary gland; D1, leukemia HL-60; D2, HeLa S3; D3, leukemia K-562;D4, leukemia MOLT-4; D5, Burkitt�s lymphoma, Raji; D6, Burkitt�slymphoma, Daudi; F2, yeast tRNA; F3, E. coli rRNA; F4, E. coli DNA;F5, poly(r(A)); and F6, human Cot-1 DNA.
Conclusions
In an effort to characterize the functions of humanP450s that do not have known substrates, P450 27C1 waschosen as a candidate and methods were developed tobegin the analysis of this and other ‘‘orphan’’ P450s. Thechoice of the N-terminal sequence and presence of a chap-erone-based expression system affected the expression ofP450 27C1 (Fig. 4), as might be expected [34,37,40,46,47].The expression level of the synthetic cDNA, optimizedfor E. coli codon usage (http://molbio.info.nih.gov/dna-works; see also http://www.kazusa.or.jp/codon/), was veryhigh, but a control study with the unaltered sequence hasnot been carried out for comparison.
On the basis of the sequence of P450 27C1, we assumethat it should have a mitochondrial localization and ana-lyzed this with an adrenodoxin-based electron transfer sys-tem. No activity toward the potential substrates cholesteroland Vit D3 and its major products was observed underthese conditions. Expression of P450 27C1 mRNA wasdetected in human liver, pancreas, kidney, and severalother tissues (Fig. 8).
It is of interest to note that mice express P450 27A1 and27B1, but P450 27C1 is not present in the mouse genome[48]. At this time our results do not provide a clear indica-tion of catalytic activity for P450 27C1, but other possibil-ities cannot be excluded. A literature search at the time ofsubmission/publication of this manuscript provides noreports of human P450 27C1 except for a patent applica-tion (Cytochroma) which is not available.
Acknowledgments
This work was supported by US Public Health ServiceGrants R01 CA090426 and P30 ES000267. We thank I.A. Pikuleva (University of Texas), N. Avadhani (Universi-ty of Pennsylvania, Philadelphia, PA), and M. R. Water-man (Vanderbilt University, Nashville, TN) for providingAdx and Adr, N. Kagawa (Vanderbilt University) for thegroEL/ES expression plasmid, pGro12 (HSP ResearchLaboratory, Kyoto, Japan), and K. Trisler for assistancein preparation of the manuscript.
Appendix A. Supplementary material
Supplementary data associated with this article can befound, in the online version, at doi:10.1016/j.abb.2005.11.002.
References
[1] G. Palmer, J. Reedijk, J. Biol. Chem. 267 (1992) 665–677.[2] P.R. Ortiz de Montellano (Ed.), Cytochrome P450: Structure,
Mechanism, and Biochemistry, third ed., Kluwer Academic/PlenumPublishers, New York, 2005.
[3] F.P. Guengerich, in: P.R. Ortiz de Montellano (Ed.), CytochromeP450: Structure, Mechanism, and Biochemistry, third ed., KluwerAcademic/Plenum Publishers, New York, 2005, pp. 377–531.

146 Z.-L. Wu et al. / Archives of Biochemistry and Biophysics 445 (2006) 138–146
[4] F.P. Guengerich, Z.-L. Wu, C.J. Bartleson, Biochem. Biophys. Res.Commun. 338(1) (2005) 465–469.
[5] N. Sawada, T. Sakaki, M. Ohta, K. Inouye, Biochem. Biophys. Res.Commun. 273 (2000) 977–984.
[6] K. Wikvall, Int. J. Mol. Med. 7 (2001) 201–209.[7] I.A. Pikuleva, I. Bjorkhelm, M.R. Waterman, Arch. Biochem.
Biophys. 343 (1997) 123–130.[8] N. Sawada, T. Sakaki, S. Kitanaka, K. Takeyama, S. Kato, K.
Inouye, Eur. J. Biochem. 265 (1999) 950–956.[9] E. Axen, H. Postlind, H. Sjoberg, K. Wikvall, Proc. Natl. Acad. Sci.
USA 91 (1994) 10014–10018.[10] T. Monkawa, T. Yoshida, S. Wakino, T. Shinki, H. Anazawa, H.F.
DeLuca, T. Suda, M. Hayashi, T. Saruta, Biochem. Biophys. Res.Commun. 239 (1997) 527–533.
[11] D.M. Hoover, J. Lubkowski, Nucleic Acids Res. 30 (2002) e43.[12] A. Parikh, E.M.J. Gillam, F.P. Guengerich, Nat. Biotechnol. 15
(1997) 784–788.[13] K. Nishihara, M. Kanemori, M. Kitagawa, H. Yanagi, T. Yura,
Appl. Environ. Microbiol. 64 (1998) 1694–1699.[14] P. Sandhu, T. Baba, F.P. Guengerich, Arch. Biochem. Biophys. 306
(1993) 443–450.[15] E.M.J. Gillam, T. Baba, B.-R. Kim, S. Ohmori, F.P. Guengerich,
Arch. Biochem. Biophys. 305 (1993) 123–131.[16] F.P. Guengerich, M.V. Martin, in: I.R. Phillips, E. Shephard (Eds.),
Methods in Molecular Genetics, Cytochrome P450 Protocols, Aca-demic Press, Orlando, FL, 2006, pp. 31–37.
[17] L.L. Manza, S.L. Stamer, A.J. Ham, S.G. Codreanu, D.C. Liebler,Proteomics 5 (2005) 1742–1745.
[18] H.J. Cortes, C.D. Pfeiffer, B.E. Richter, T.S. Stevens, J. High, Resol.Chromatog. Chromatog. Commun. 10 (1987) 446–448.
[19] L.J. Licklider, C.C. Thoreen, J. Peng, S.P. Gygi, Anal. Chem. 74(2002) 3076–3083.
[20] J.R. Yates III, J.K. Eng, A.L. McConnack, D. Schieltz, Anal. Chem.67 (1994) 1426–1436.
[21] F.M. Ausubel, R. Brent, R.E. Kingston, D.D. Moore, J.G. Seidman,J.A. Smith, K. Struhl, Current Protocols in Molecular Biology, JohnWiley, New York, 1995.
[22] I.A. Pikuleva, A. Puchkaev, I. Bjorkhem, Biochemistry 40 (2001)7621–7629.
[23] N. Mast, S.E. Graham, U. Andersson, I. Bjorkhem, C. Hill, J.Peterson, I.A. Pikuleva, Biochemistry 44 (2005) 3259–3271.
[24] F.P. Guengerich, in: A.W. Hayes (Ed.), Principles and Methods ofToxicology, Taylor & Francis, Philadelphia, 2001, pp. 1625–1687.
[25] T. Omura, R. Sato, J. Biol. Chem. 239 (1964) 2370–2378.
[26] J.B. Schenkman, H. Remmer, R.W. Estabrook, Mol. Pharmacol. 3(1967) 113–123.
[27] W.P. Stemmer, A. Crameri, K.D. Ha, T.M. Brennan, H.L. Heyneker,Gene 164 (1995) 49–53.
[28] C. Gustafsson, S. Govindarajan, J. Minshull, Trends Biotechnol. 22(2004) 346–353.
[29] A. Carbone, A. Zinovyev, F. Kepes, Bioinformatics 19 (2003) 2005–2015.
[30] X. Gao, P. Yo, A. Keith, T.J. Ragan, T.K. Harris, Nucleic Acids Res.31 (2003) e143.
[31] A.S. Xiong, Q.H. Yao, R.H. Peng, X. Li, H.Q. Fan, Z.M. Cheng, Y.Li, Nucleic Acids Res. 32 (2004) e98.
[32] S.J. Kodumal, K.G. Patel, R. Reid, H.G. Menzella, M. Welch, D.V.Santi, Proc. Natl. Acad. Sci. USA 101 (2004) 15573–15578.
[33] H.O. Smith, C.A. Hutchison III, C. Pfannkoch, J.C. Venter, Proc.Natl. Acad. Sci. USA 100 (2003) 15440–15445.
[34] H.J. Barnes, M.P. Arlotto, M.R. Waterman, Proc. Natl. Acad. Sci.USA 88 (1991) 5597–5601.
[35] A. Wada, P.A. Mathew, H.J. Barnes, D. Sanders, R.W. Estabrook,M.R. Waterman, Arch. Biochem. Biophys. 290 (1991) 376–380.
[36] T.H. Richardson, F. Jung, K.J. Griffin, M. Wester, J.L. Raucy, B.Kemper, L.M. Bornheim, C. Hassett, C.J. Omiecinski, E.F. Johnson,Arch. Biochem. Biophys. 323 (1995) 87–96.
[37] H.J. Barnes, Methods Enzymol. 272 (1996) 3–14.[38] Y. Ohyama, M. Noshiro, K. Okuda, FEBS Lett. 278 (1991) 195–198.[39] T. Sakaki, N. Sawada, Y. Nonaka, Y. Ohyama, K. Inouye, Eur. J.
Biochem. 262 (1999) 43–48.[40] F.P. Guengerich, E.M.J. Gillam, T. Shimada, CRC Crit. Rev.
Toxicol. 26 (1996) 551–583.[41] N. Kagawa, Q. Cao, Recent Res. Dev. Biophys. Biochem. 1 (2001)
99–107.[42] T. Meinnel, Y. Mechulam, S. Blanquet, Biochimie 75 (1993) 1061–
1075.[43] M.-S. Dong, L.C. Bell, Z. Guo, D.R. Phillips, I.A. Blair, F.P.
Guengerich, Biochemistry 35 (1996) 10031–10040.[44] T.C. O�Haver, G.L. Green, Anal. Chem. 48 (1976) 312–318.[45] F.P. Guengerich, Biochemistry 22 (1983) 2811–2820.[46] E. Uchida, N. Kagawa, T. Sakaki, N. Urushino, N. Sawada, M.
Kamakura, M. Ohta, S. Kato, K. Inouye, Biochem. Biophys. Res.Commun. 323 (2004) 505–511.
[47] K. Kusano, M.R. Waterman, M. Sakaguchi, T. Omura, N. Kagawa,Arch. Biochem. Biophys. 367 (1999) 129–136.
[48] D.R. Nelson, D.C. Zeldin, S.M. Hoffman, L.J. Maltais, H.M. Wain,D.W. Nebert, Pharmacogenetics 14 (2004) 1–18.
Related Documents











