Animal Reproduction Science 112 (2009) 251–260 GnRH or eCG treatment fails to restore reproductive function in GnRH immunized ewes ¨ O. G ¨ okdal a,∗ , O. Atay a , H. ¨ Ulker b , E. Yaralı a , ˙ I.B. Helva a , D.M. DeAvila c , J.J. Reeves c a Adnan Menderes University, C ¸ine Vocational High School, 09500 C ¸ ine, Aydın, Turkey b Y¨ uz¨ unc¨ u Yıl University, Faculty of Agriculture, Department of Animal Science, Van, Turkey c Department of Animal Sciences, Washington State University, Pullman, WA, USA Received 22 February 2008; received in revised form 2 April 2008; accepted 23 April 2008 Available online 2 May 2008 Abstract This study was designed to evaluate the potential of using eCG or GnRH in restoring reproductive functions in GnRH immunized ewes. Thirty-three multiparous Kıvırcık ewes were randomly assigned into either control group (n = 11) or immunization group (n = 22). Ewes were immunized against GnRH by injecting with a cocktail of ovalbumin-LHRH-7 (ovalbumin-GnRH-7) and thioredoxin-LHRH-7 (thioredoxin-GnRH- 7) fusion proteins generated by recombinant DNA technology in April. 500 IU eCG or 0.008 mg GnRH analogue was used to induce ovulations. Serum GnRH antibodies were present in animals of the immunized group beginning the second week after the first immunization and maintained throughout the study (14 months). Immunization caused anestrus in immunized ewes. eCG or GnRH analogue administration given after 14 days progestagen (20 mg fluorogestone acetate, FGA) treatment during breeding season (mid July) did not induce ovulation in these ewes. Two more attempts with single or multiple eCG injections failed to induce ovulation in this group as well. It appears that the gonadotropin stimulation was not of adequate time since neither eCG nor GnRH administration was able to restore reproductive function in immunized animals. The immunization effect lasted more than a year. These results suggest that GnRH immunization exerts its effect via the hypothalamo-pituitary axis and that more than such stimulation is required to overcome the reproductive suppression. © 2008 Elsevier B.V. All rights reserved. Keywords: GnRH; Fusion proteins; Immunization; Ewes ∗ Corresponding author. Tel.: +90 256 7117052; fax: +90 256 7117054. E-mail address: [email protected] ( ¨ O. G ¨ okdal). 0378-4320/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.anireprosci.2008.04.023

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Animal Reproduction Science 112 (2009) 251–260
GnRH or eCG treatment fails to restore reproductivefunction in GnRH immunized ewes
O. Gokdal a,∗, O. Atay a, H. Ulker b, E. Yaralı a, I.B. Helva a,D.M. DeAvila c, J.J. Reeves c
a Adnan Menderes University, Cine Vocational High School, 09500 Cine, Aydın, Turkeyb Yuzuncu Yıl University, Faculty of Agriculture, Department of Animal Science, Van, Turkey
c Department of Animal Sciences, Washington State University, Pullman, WA, USA
Received 22 February 2008; received in revised form 2 April 2008; accepted 23 April 2008Available online 2 May 2008
Abstract
This study was designed to evaluate the potential of using eCG or GnRH in restoring reproductive functionsin GnRH immunized ewes. Thirty-three multiparous Kıvırcık ewes were randomly assigned into eithercontrol group (n = 11) or immunization group (n = 22). Ewes were immunized against GnRH by injectingwith a cocktail of ovalbumin-LHRH-7 (ovalbumin-GnRH-7) and thioredoxin-LHRH-7 (thioredoxin-GnRH-7) fusion proteins generated by recombinant DNA technology in April. 500 IU eCG or 0.008 mg GnRHanalogue was used to induce ovulations. Serum GnRH antibodies were present in animals of the immunizedgroup beginning the second week after the first immunization and maintained throughout the study (14months). Immunization caused anestrus in immunized ewes. eCG or GnRH analogue administration givenafter 14 days progestagen (20 mg fluorogestone acetate, FGA) treatment during breeding season (mid July)did not induce ovulation in these ewes. Two more attempts with single or multiple eCG injections failed toinduce ovulation in this group as well. It appears that the gonadotropin stimulation was not of adequate timesince neither eCG nor GnRH administration was able to restore reproductive function in immunized animals.The immunization effect lasted more than a year. These results suggest that GnRH immunization exerts itseffect via the hypothalamo-pituitary axis and that more than such stimulation is required to overcome thereproductive suppression.© 2008 Elsevier B.V. All rights reserved.
Keywords: GnRH; Fusion proteins; Immunization; Ewes
∗ Corresponding author. Tel.: +90 256 7117052; fax: +90 256 7117054.E-mail address: [email protected] (O. Gokdal).
0378-4320/$ – see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.anireprosci.2008.04.023

252 O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260
1. Introduction
Immunization against gonadotropin releasing hormone (GnRH) has been described as oneof the methods to reduce reproductive functions in farm animals and a possible alternativeto surgical castration (Reeves et al., 1989; Bonneau and Enright, 1995; Thompson, 2000).Gonadotropin hormone concentrations, testicular development and sexual activities are sup-pressed in GnRH immunized animals (Robertson et al., 1982; Hoskinson et al., 1990; Adamsand Adams, 1992). Two recombinant fusion proteins, ovalbumin-LHRH-7 (ovalbumin-GnRH-7)(OL) and thioredoxin-LHRH-7 (thioredoxin-GnRH-7) (TL), were developed to be used as a steril-ization vaccine (Zhang et al., 1999; Quesnell et al., 2000). The effectiveness of these recombinantproteins in suppressing reproductive functions was demonstrated in heifers (Sosa et al., 2000),bulls (Aissat et al., 2002) and ram lambs (Ulker et al., 2001; Ulker et al., 2005). The effective-ness of these recombinant proteins in suppressing reproductive functions in ewes has not beenstudied.
Many researchers report that active immunization against GnRH induces only a temporarysuppression of reproductive functions after which animals return to normal fertility (Reeves et al.,1989; D’Occhio, 1993; Bonneau and Enright, 1995; Thompson, 2000). Alternatively, reproductivefunctions in GnRH immunized animals could be restored by using GnRH agonist that did notcross-react with the antibodies (Adams and Adams, 1986; Herman and Adams, 1990; Sakuraiet al., 1992), multiple injections of FSH, LH, hCG (Mariana et al., 1998) or eCG (Oatley etal., 2005). Nevertheless, single injection of GnRH to restore reproductive functions after activeimmunization done in early life (Brown et al., 1994, 1995; Clarke et al., 1998) or adult age inewes (Jeffcoate et al., 1978) failed to do so. Immunizing farm animals against GnRH allowsanimal owners to keep the male and female animals together as long as the immunization effectlasts. Restoring reproductive functions in immunized animals without waiting for the decline incirculating anti-GnRH antibodies below a threshold required to neutralize GnRH could be a usefulmanagement tool. Nevertheless, no such studies related to restoring reproductive function afteractive immunization against GnRH at adult age have been shown. In sheep immunized againstGnRH at prepubertal or peripubertal age, plasma luteinizing hormone (LH) concentrations werenot restored after GnRH injection at a time when anti-GnRH antibodies are low (Brown et al.,1994, 1995) or not detectable (Clarke et al., 1998). These findings have led to the suggestionthat in the young ewe the basal hypothalamus-median eminence is a target site for anti-GnRHantibodies which cause lesions disrupting the integrity of hypothalamus and, consequently, longterm reproductive suppression. Findings in swine support this suggestion (Molenaar et al., 1993).D’Occhio et al. (2001) suggest the presence of a long-term permanent effect in bulls immunizedat adult age. Conversely, Mariana et al. (1998) were able to induce preovulatory follicles in ewesimmunized against GnRH early in life by multiple injections of FSH, LH or hCG. Similarly,spermatogenesis has been restored in GnRH immunized bulls using eCG administered IM every2 week for 80 days (Oatley et al., 2005). It is not known whether reproductive functions couldbe restored by eCG injection in ewes actively immunized against GnRH at an adult age usingrecombinant fusion proteins.
Suppression of LH, which is a result of GnRH immunization, has been suggested to causeearly disruption of pregnancy through inadequate pituitary support to corpus luteum (CL) (Tastet al., 2000). So, even though reproductive functions are restored in immunized animals usinggonadotropin hormones, i.e. eCG, there is a possibility that pregnancy might be ended becauseof circulating anti-GnRH antibodies neutralizing GnRH and consequently low levels of LH.However, considering the longer half life and LH like effect (Murphy and Martinuk, 1991), eCG

O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260 253
could provide adequate LH support during the early days of pregnancy. If ovarian functionsof GnRH immunized animals can be induced by eCG administration, then, valuable informationcould be obtained on establishment and maintenance of pregnancy under high anti-LHRH antibodyconcentrations.
The purposes of this study were (1) to determine the effectiveness of recombinant LHRH fusionproteins in suppressing reproductive functions in mature, pluriparous ewes and (2) to investigatethe possibilities of using eCG or LHRH in restoring reproductive functions in LHRH immunizedewes.
2. Materials and methods
2.1. Animals and treatments
Thirty-three multiparous Kıvırcık ewes, a multi-purpose (meat, milk and wool) thin-tailedlocal sheep breed of Turkey, at 3–8 years of age and weighing average 39 kg were randomlyassigned into control (n = 11) and immunization (n = 22) groups. All ewes were subjected toestrous synchronization program consisting of 14 days progestagen (20 mg FGA) impregnatedsponge plus 500 IU eCG injection at sponge withdrawal at least once previously. Breeding seasonstarts in June and majority of the ewes are mated during July in Cine district (latitude 37◦61′E,longitude 28◦06′N) of Aydın region, Turkey. Therefore, the immunization schedule was designedto obtain high antibody concentrations during the mating period. Thus, ewes in the immunizationgroup were immunized against GnRH using GnRH fusion proteins on April 11, which was 111days prior to proposed mating time. Since all ewes were being suckled during that time in orderto eliminate the possibility that suckling may suppress ovarian activity, ewes were subjected toa ‘21-day decreasing suckling program’ beginning 1 week after immunization to get them dry.Booster immunization was done 1 month after the first immunization.
2.2. Preparation of antigens and immunizations
OL and TL proteins were produced from previously constructed ovalbumin-GnRH-7 andthioredoxin-GnRH-7 genes generated by recombinant DNA techniques as described by Zhanget al. (1999) and Quesnell et al. (2000), respectively. Recombinant genes were over expressed inEscherichia Coli. His-bind affinity chromatography using a Ni2+ column allowed for purificationof the proteins. Equimolar amounts of each GnRH fusion protein (10 nM) totalling 0.75 mg of pro-tein were suspended in 6 M urea and emulsified in 0.5 ml of modified complete Freund’s adjuvant(Sigma, St. Louis, MO, USA) for the primary immunization and incomplete Freund’s adjuvantwas used for the booster injection. Immunizations were distributed over four subcutaneous siteson the inside surface of the legs.
2.3. Data collection
Two weeks after the booster a harnessed teaser ram was introduced to stimulate sexual func-tions. Behavioral estrus was observed. Ninety-five days after the first immunization all animalswere subjected to a typical estrus synchronization program: Progestagen (20 mg fluorogestoneacetate, FGA) impregnated sponges (Chronogest®, Intervet International, Boxmeer, The Nether-lands) were inserted for 14 days. At sponge withdrawal control animals and 11 animals inimmunization group were administered with 500 IU eCG, with i.m. injecion. The other 11 ani-

254 O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260
Fig. 1. Diagram of experimental design (synchronization of estrus and first attempt to restore reproductive functions inGnRH immunized ewes).
mals in immunization group were administered i.m. with 0.9 ml/animal GnRH analogue (0.008 mgBuserilin [Receptal]; Hoechst UK Ltd., Milton Keynes, UK). Two days after eCG or GnRH ana-logue injections rams were introduced. One ram was assigned per 4–5 ewes in separate pens. Heatand mating activities were observed (Fig. 1).
Since no estrus activity was observed in the immunized ewes 10 days after sponge removal allanimals in this group were injected i.m. with 400 IU eCG to induce ovulation (second attempt).Rams introduced after sponge withdrawal remained with ewes until pregnancy determination(Fig. 2).
All animals we examined for pregnancy using 6 MHz linear rectal probe (Pie medical, 100Falco vet.) 35 days after sponge withdrawal. Since no animals were pregnant in immunizationgroup a ‘long term ovarian stimulating program, multiple eCG injections’ was initiated (thirdattempt). For this purpose 14 animals in immunization group were injected i.m. with 400 IU eCGonce and then 280 IU eCG i.m. three times at 6 days intervals and once 3 days after forth one.
Fig. 2. Diagram of second (single eCG injection) and third (multiple eCG injections) attempts to restore reproductivefunctions in GnRH immunized ewes.
O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260 255
Progestagen containing sponges were inserted at the third eCG injection and removed at the lastinjection. At sponge removal fertile rams were introduced and estrus behaviors were observed.The rams remained with ewes all time (Fig. 2). All control and 14 animals from immunized groupwere kept with fertile rams during the subsequent year’s mating season.
Beginning from the first immunization all animals were bled every 3–4 days to monitor levelsof ovarian hormones. Also all animals were bled every 2 weeks to determine anti-GnRH antibodyconcentrations. For the last 7 months of the study, only monthly blood collection was performed.Blood samples were refrigerated overnight at +4 ◦C, and then centrifuged at 3000 rpm for 15 min.Serum was harvested and stored at −20 ◦C until subsequent analyses.
A radioactive binding assay was used to evaluate the percentage of 125I-GnRH that would bindto the anti-GnRH antibodies present in the serum at a 1:1000 dilution. Mouse anti-sheep gammaglobulin was used as the second antibody at 1:20 dilution. Iodination of GnRH was performedusing GnRH (2.5 �g/25 �l H2O) with 0.5 mCi (5 �l) 125I, 30 �l of 0.5 M PBS (pH 7.5) and 10 �lChloramine-T (600 ng/10 �l of 0.5 M PBS, pH 7.5). I-GnRH was separated on a QAE-Sephadexcolumn using column buffer (10 mM Tris, 1 mM CaCl2; 0.1% BSA, pH 7.2). The percentage of125I-GnRH bound to serum diluted 1:1000 was used as the index for GnRH antibody titers.
Serum progesterone concentrations were determined by RIA (DSL-3400, Diagnostic SystemsLaboratories, Inc. Webster, TX). All procedures were conducted according to the manufacturer’sinstructions.
Data analysis was performed using GLM procedure of SAS (SAS Inst. Inc., Cary, NC) forrepeated measures to determine main effects of treatment, time and treatment × time for each ofresponse variables (serum anti-GnRH antibody percentage bound, progesterone concentration).Data are presented as means ± S.E.M.
3. Results
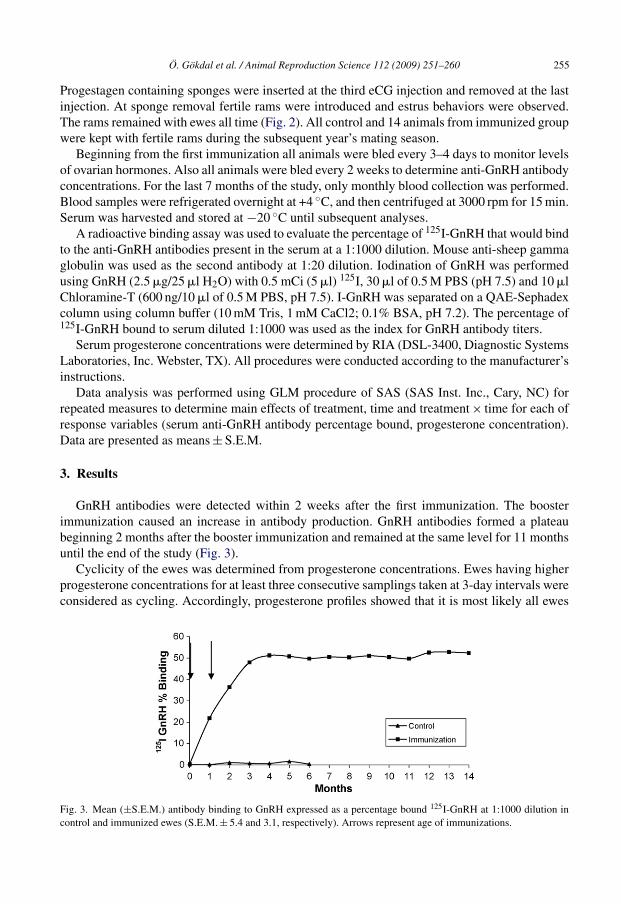
GnRH antibodies were detected within 2 weeks after the first immunization. The boosterimmunization caused an increase in antibody production. GnRH antibodies formed a plateaubeginning 2 months after the booster immunization and remained at the same level for 11 monthsuntil the end of the study (Fig. 3).
Cyclicity of the ewes was determined from progesterone concentrations. Ewes having higherprogesterone concentrations for at least three consecutive samplings taken at 3-day intervals wereconsidered as cycling. Accordingly, progesterone profiles showed that it is most likely all ewes
Fig. 3. Mean (±S.E.M.) antibody binding to GnRH expressed as a percentage bound 125I-GnRH at 1:1000 dilution incontrol and immunized ewes (S.E.M. ± 5.4 and 3.1, respectively). Arrows represent age of immunizations.
256 O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260
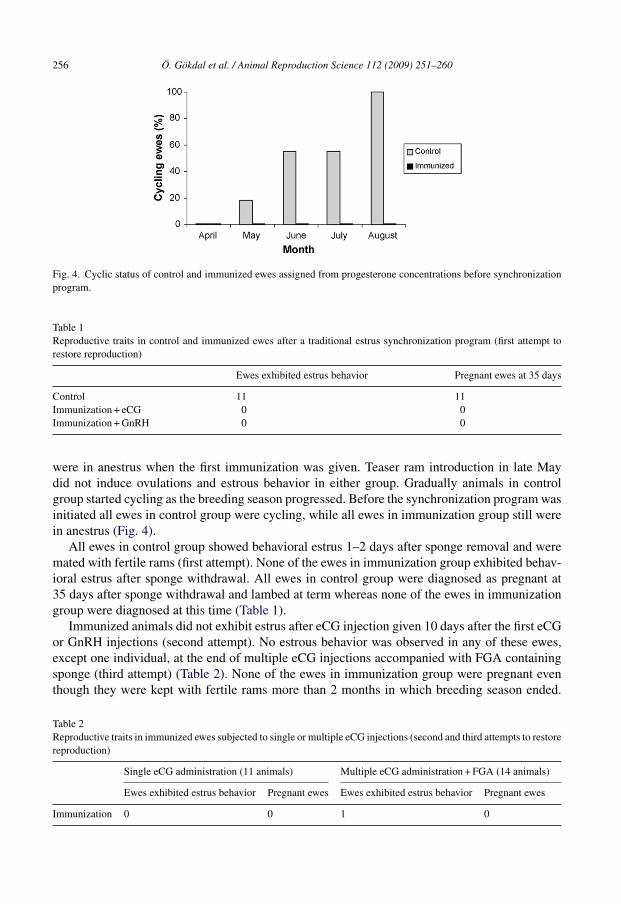
Fig. 4. Cyclic status of control and immunized ewes assigned from progesterone concentrations before synchronizationprogram.
Table 1Reproductive traits in control and immunized ewes after a traditional estrus synchronization program (first attempt torestore reproduction)
Ewes exhibited estrus behavior Pregnant ewes at 35 days
Control 11 11Immunization + eCG 0 0Immunization + GnRH 0 0
were in anestrus when the first immunization was given. Teaser ram introduction in late Maydid not induce ovulations and estrous behavior in either group. Gradually animals in controlgroup started cycling as the breeding season progressed. Before the synchronization program wasinitiated all ewes in control group were cycling, while all ewes in immunization group still werein anestrus (Fig. 4).
All ewes in control group showed behavioral estrus 1–2 days after sponge removal and weremated with fertile rams (first attempt). None of the ewes in immunization group exhibited behav-ioral estrus after sponge withdrawal. All ewes in control group were diagnosed as pregnant at35 days after sponge withdrawal and lambed at term whereas none of the ewes in immunizationgroup were diagnosed at this time (Table 1).
Immunized animals did not exhibit estrus after eCG injection given 10 days after the first eCGor GnRH injections (second attempt). No estrous behavior was observed in any of these ewes,except one individual, at the end of multiple eCG injections accompanied with FGA containingsponge (third attempt) (Table 2). None of the ewes in immunization group were pregnant eventhough they were kept with fertile rams more than 2 months in which breeding season ended.
Table 2Reproductive traits in immunized ewes subjected to single or multiple eCG injections (second and third attempts to restorereproduction)
Single eCG administration (11 animals) Multiple eCG administration + FGA (14 animals)
Ewes exhibited estrus behavior Pregnant ewes Ewes exhibited estrus behavior Pregnant ewes
Immunization 0 0 1 0

O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260 257
Table 3Pregnancy results in control and immunized ewes for 2 years.
First year Second year
Lambed ewes Lamb numbers per ewe Lambed ewes Lamb numbers per ewe
Control 11 1.4 11 1.4Immunization 0 0 0 0
Furthermore, none of these immunized animals got pregnant in the second year’s breeding season,while all control animals got pregnant and lambed at term (Table 3).
4. Discussion
Theoretically, immunocastration is reversible phenomenon. The reversibility of immunocastra-tion in natural process, i.e., with gradual decrease in GnRH antibody concentrations over the time,has been noted in several studies (Keeling and Crighton, 1984; Grieger et al., 1990; D’Occhioet al., 2001). However, several studies related to restoration of pituitary and ovarian functionsusing hormone treatment after immunocastration have been reported. Serum concentrations ofLH in actively or passively immunized ewes were restored by circhoral (hourly) administration ofa GnRH agonist that did not cross-react with the antibodies (Adams and Adams, 1986; Hermanand Adams, 1990; Sakurai et al., 1992). Also, restoration of spermatogenesis using eCG in GnRHimmunized bulls has been achieved (Oatley et al., 2005). The use of eCG in estrous synchronizationprotocols in sheep is well established. Fourteen days progestagen impregnated sponges insertionaccompanied with a single eCG injection at sponge withdrawal increases ovarian response andpercentage of multiple births from induced ovulation (Pearce and Robinson, 1985). Additionally,GnRH treatment is known to induce ovulation in anestrus sheep using traditional synchronizationprograms (Bartlewski et al., 2001, 2004). In the present study neither eCG nor GnRH analogueinjections combined with FGA (progestagen) treatment were able to induce estrous in immunizedewes.
GnRH injections to the non-cyclic immunized ewes in early ages (Brown et al., 1995) or adultages (Jeffcoate et al., 1978) did not cause a marked increase in LH concentrations. It was concludedthat immunization may have resulted in either reduced sensitivity of the pituitary gland to GnRHstimulation, a diminished capacity to produce and secrete LH and FSH or depleted pituitary poolof gonadotropins. In 35% of sexually mature bulls immunized against LHRH the testes continuedto decrease in size for 4 months and did not show any re-initiation of growth for 1 year afterimmunization (D’Occhio et al., 2001). It was concluded, therefore, that active immunizationagainst GnRH can induce a long-term, possibly permanent, suppression of reproductive functionin bulls. Thus, a permanent castration like effect could be speculated for protein combination usedin the present study as well.
The mechanism for sustained suppression of reproductive functions after immunizationagainst GnRH is poorly understood. Preliminary findings in sheep and swine suggest that basalhypothalamus-median eminence is a primary target site for anti-GnRH antibodies, and immuniza-tion may lead to a disruption of the integrity of basal hypothalamus-median eminence (Molenaaret al., 1993; Clarke et al., 1998). Hernandez et al. (2005) reported vacuolized large basophilcells known as ‘castrate cells’ in histological examination in castrated rats’ pituitaries but not inthe immunocastrated ones. Similarly, histological evaluation of anterior pituitaries of castrated

258 O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260
and GnRH immunized bulls and ram lambs did not indicate any pathological or morphologi-cal differences (unpublished results). It appears that there is little information to indicate thatGnRH immunization can generate a permanent effect on the pituitary by causing pathologicalchanges.
There are some reports that eCG induces some immune response and therefore eCG antibodiesare generated (Bodin et al., 1997). In some estrous synchronization programs lower fertility rateswere attributed to this fact. Animals in this study had been treated with eCG previously duringthe preceding years. So, in order to eliminate this possibility half of the animals in immunizationgroup were treated with GnRH because GnRH treatment is known to induce ovulation in anestroussheep (Bartlewski et al., 2001, 2004). Nevertheless, this approach did not contribute to restorationof reproductive functions.
The response of ovaries to eCG injection in immunized animals remains to be evaluated. eCGexerts its gonadotropic effect via FSH and LH receptors on the ovary (Murphy and Martinuk,1991). Neither in the first attempt in eCG treated immunized group nor in the second and thirdattempts could induce ovarian functions in immunized animals. Mariana et al. (1998) were able toinduce preovulatory follicles in ewes immunized against GnRH early in life by multiple injectionsof FSH, LH or hCG. Similarly, spermatogenesis has been restored in GnRH immunized bullsusing eCG administered i.m., every 2 weeks for 80 days (Oatley et al., 2005). In the presentstudy, immunization, possibly, caused the ovaries of ewes to regress to a pre-pubertal stage andthe time and total dose of eCG applied in the present study were too short to recover from thisstage. Alternatively, eCG could induce follicular developments on the ovary but preovulatorysurge could not occur because of high level of circulating GnRH antibodies. This aspect requiresfurther investiagation.
Immunized animals did not get pregnant in the second year’s breeding season. GnRH antibodyconcentrations were still high during this time (Fig. 3). Prolonged immunocastration might be theresult of high antibody concentrations.
There has been no dose response trial for GnRH fusion proteins in the ewes. In order to achievea reversible or transient immunocastration it is important that immunization against GnRH shouldachieve a short-term direct immunoneutralization of GnRH by GnRH antibodies. If immuniza-tion disrupts the tissues in the median eminence associated with the presence of high levels ofGnRH antibodies a long-term immunocastration is achieved, but this immunocastration will notbe reversible.
5. Conclusion
There is a possibility that lower doses of these fusion proteins may be able to induce onlyshort-term immunoneutralization of GnRH by GnRH antibodies and therefore allows restorationof reproductive functions upon request.
These results suggest that GnRH immunization exerts its effect via the hypothalamo-pituitaryaxis and that more than such stimulation is required to overcome the reproductive suppression.Determining the way of response of ovaries to eCG injection in immunized animals requiresfurther investigation.
Acknowledgements
This project was supported by Adnan Menderes University Scientific Research ProjectsCommission (Project no: CMYO-03001) and National Research Initiative Competitive Grant

O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260 259
no: 2003-25203-13514 from the USDA Cooperative State Research, Education, and ExtensionService.
References
Adams, T.E., Adams, B.M., 1986. Gonadotrope function in ovariectomized ewes actively immunized against gonadotropin-releasing hormone (GnRH). Biol. Reprod. 35 (2), 360–367.
Adams, T.E., Adams, B.M., 1992. Feedlot performance of steers and bulls actively immunised against GnRH. J. Anim.Sci. 70, 1691–1698.
Aissat, D., Sosa, J.M., de Avila, D.M., Bertrand, K.P., Reeves, J.J., 2002. Endocrine, growth, and carcass characteristicsof bulls immunized against luteinizing hormone-releasing hormone fusion proteins. J. Anim. Sci. 80, 2209–2213.
Bartlewski, P.M., Beard, A.P., Chapman, C.L., Nelson, M.L., Palmer, B., Aravindakshan, J., Cook, S.J., Rawlings,N.C., 2001. Ovarian responses in gonadotrophin-releasing hormone-treated anoestrous ewes: follicular and endocrinecorrelates with luteal outcome. Reprod. Fertil. Dev. 13 (2–3), 133–142.
Bartlewski, P.M., Aravindakshan, J., Beard, A.P., Nelson, M.L., Batista-Arteaga, M., Cook, S.J., Rawlings, N.C., 2004.Effects of medroxyprogesterone acetate (MAP) on ovarian antral follicle development, gonadotrophin secretion andresponse to ovulation induction with gonadotrophin-releasing hormone (GnRH) in seasonally anoestrous ewes. Anim.Reprod. Sci. 81 (1–2), 63–71.
Bodin, L., Drion, P.V., Remy, B., Brice, G., Cognie, Y., Beckers, J.F., 1997. Anti-PMSG antibody levels in sheep subjectedannually to oestrus synchronisation. Reprod. Nutr. Dev. 37 (6), 651–660.
Bonneau, M., Enright, W.J., 1995. Immunocastration in cattle and pig. Livestock Prod. Sci. 42, 193–200.Brown, B.W., Mattner, P.E., Carroll, P.A., Holland, E.J., Paull, D.R., Hoskinson, R.M., Rigby, R.D.G., 1994. Immunization
of sheep against GnRH early in life: effects on reproductive function and hormones in rams. J. Reprod. Fertil. 101,15–21.
Brown, B.W., Mattner, P.E., Carrol, P.A., Hoskinson, R.M., Rigby, R.D.G., 1995. Immunization of sheep against GnRHearly in life: effects of reproduction functions and hormones in ewes. J. Reprod. Fertil. 103, 131–135.
Clarke, I.J., Brown, B.W., Tran, V.V., Scott, C.J., Fry, R., Millar, R.P., Rao, A., 1998. Neonatal immunization againstGonadotropin-releasing hormone (GnRH) results in diminished GnRH secretion in adulthood. Endocrinology 139(4), 2007–2014.
D’Occhio, M.J., 1993. Immunological suppression of reproductive functions in male and female mammals. Anim. Reprod.Sci. 33 (1–4), 345–372.
D’Occhio, M.J., Aspden, W.J., Trigg, T.E., 2001. Sustained testicular atrophy in bulls actively immunized against GnRH:potential to control carcase characteristics. Anim. Reprod. Sci. 66, 47–58.
Grieger, D.M., Scarborough, R., De Avila, D.M., Johnson, H.E., Reeves, J.J., 1990. Active immunization of heifers againstluteinizing hormone. III. Evaluation of dose and longevity. J. Anim. Sci. 68, 3755–3764.
Herman, M.E., Adams, T.E., 1990. Gonadotropin secretion in ovariectomized ewes: effect of passive immunization againstgonadotropin-releasing hormone (GnRH) and infusion of a GnRH agonist and estradiol. Biol. Reprod. 42 (2), 273–280.
Hernandez, J.A., Sosa Manzo, J.M., Reeves, J.J., McLean, D.J., 2005. Changes in pituitary gene expression betweensurgically castrated and LHRH immunocastrated male rats. Proc. Western Sect. Am. Soc. Anim. Sci. 56, 343–346.
Hoskinson, M., Rigby, R.D.G., Mattner, P.E., Huynh, V.L., D’Occhio, M.J., Neish, A., Trigg, T.E., Moss, B.A., Lindsey,M.J., Coleman, G.D., Schwartzkoff, C.L., 1990. Waxtrate: an anti-reproductive vaccine for cattle. Austr. J. Biotech.4, 166–170.
Jeffcoate, I.A., Foster, J.P., Crighton, D.B., 1978. Effect of active immunization of ewes against synthetic luteinisinghormone releasing hormone. Theriogenology 10 (4), 323–335.
Keeling, B.J., Crighton, D.B., 1984. Reversibility of the effects of active immunization against LHRH. In: Crighton, D.B.(Ed.), Immunological Aspects of Reproduction in Mammals. Butterworths, London, pp. 379–398.
Mariana, J.C., Monniaux, D., Caraty, A., Pisselet, C., Fontaine, J., Solari, A., 1998. Immunization of sheep against GnRHearly in life: effects on gonadotropins, follicular growth and responsiveness of granulosa cells to FSH and IGF-I in twobreeds of sheep with different prolificacy (Romanov and Ile-de-France). Domest. Anim. Endocrinol. 15 (4), 195–207.
Molenaar, G.J., Lugard-Kok, C., Meloen, R.H., Oonk, R.B., de Koning, J., Wensing, C.J., 1993. Lesions in the hypotha-lamus after active immunisation against GnRH in the pig. J. Neuroimmunol. 48 (1), 1–11.
Murphy, B.D., Martinuk, S.D., 1991. Equine chorionic gonadotropin. Endocr. Rev. 12 (1), 27–44.Oatley, J.M., Tibary, A., de Avila, D.M., Wheaton, J.E., McLean, D.J., Reeves, J.J., 2005. Changes in spermatogenesis and
endocrine function in the ram testis due to irradiation and active immunization against luteinizing hormone-releasinghormone. J. Anim. Sci. 83, 604–612.

260 O. Gokdal et al. / Animal Reproduction Science 112 (2009) 251–260
Pearce, D.T., Robinson, T.J., 1985. Plasma progesterone concentrations, ovarian and endocrinological response and spermtransport in ewes with synchronized oestrus. J. Reprod. Fertil. 75 (1), 49–62.
Quesnell, M.M., Zhang, Y., De Avila, D.M., Bertrand, K.P., Reeves, J.J., 2000. Immunization of male mice with luteinizinghormone-releasing hormone fusion proteins reduces testicular and accessory sex gland function. Biol. Reprod. 63,347–353.
Reeves, J.J., Chang, C.F., De Avila, D.M., Grieger, H.E., Johnson, H.E., Roberts, A.J., 1989. Vaccine against endogenoushormones: a possible future tool in animal production. J. Dairy Sci. 72, 3363–3371.
Robertson, I.S., Fraser, H.M., Innes, G.M., Jones, A.S., 1982. Effects of immunocastration on sexual and productioncharacteristics of male cattle. Vet. Rec. 111, 529–531.
Sakurai, H., Adams, B.M., Adams, T.E., 1992. Pattern of gonadotropin-releasing hormone (GnRH)-like stimuli sufficientto induce follicular growth and ovulation in ewes passively immunized against GnRH. Biol. Reprod. 47 (2), 177–184.
Sosa, J.M., Zhang, Y., De Avila, D.M., Bertrand, K.P., Reeves, J.J., 2000. Technical note: recombinant LHRH fusionprotein suppresses estrus in heifers. J. Anim. Sci. 78, 1310–1312.
Tast, A., Love, R.J., Clarke, I.J., Evans, G., 2000. Effects of active and passive gonadotropin-releasing hormone immu-nization on recognition and establishment of pregnancy in pigs. Reprod. Fertil. Dev. 12, 277–282.
Thompson, D.L., 2000. Immunization against GnRH in male species (comperative aspects). Anim. Reprod. Sci. 60–61,459–469.
Ulker, H., Kanter, M., Gokdal, O., Aygun, T., Karakus, F., Sakarya, M.E., De Avila, D.M., Reeves, J.J., 2005. Testiculardevelopment, ultrasonographic and histological appearance of the testis in ram lambs immunized against recombinantLHRH fusion proteins. Anim. Reprod. Sci. 86, 205–219.
Ulker, H., Kanter, M., Gokdal, O., De Avila, D.M., Reeves, J.J., 2001. The effects of recombinant LHRH fusion proteinson testicular development and histology in ram lambs. Deutsche Tierarztliche Wochenschrift 108, 459–464.
Zhang, Y., Rozell, T.G., De Avila, D.M., Bertrand, K.P., Reeves, J.J., 1999. Development of recombinant ovalbumin-luteinizing hormone releasing hormone as a potential sterilization vaccine. Vaccine 17, 2185–2191.
Related Documents











