Extremophiles (1997) 1:2–13 © Springer-Verlag 1997 REVIEW Starch-degrading enzymes Starch is, after cellulose, one of the most abundant polysac- charides produced by plants and is composed of amylose (15%–25%) and amylopectin (75%–85%). Amylose is a linear molecule consisting of 1,4-linked α-d-glucopyranose residues. Amylopectin is a branched polymer and contains α-1,6 glycosidic linkages in addition to the α-1,4 bonds. The hydrolysis of starch requires the coordinate action of sev- eral enzymes, such as α-amylase, β-amylase, glucoamylase, debranching enzymes (pullulanases), and α-glucosidase. The biodegradation of starch is widespread among Bac- teria, fungi, and yeast (Antranikian 1992). Several extreme thermophilic and hyperthermophilic Bacteria and Archaea have been found to grow at high temperatures (>70°C) on starch and produce extremely thermostable starch- degrading enzymes (Leuschner and Antranikian 1995). Table 1 summarizes some properties of selected thermo- stable starch-hydrolyzing enzymes. α-Amylase (1,4-α-d-glucan, 4-glucanhydrolase, EC 3.2.1.1) This endoacting enzyme produces oligosaccharides and glu- cose as end-products by randomly hydrolyzing the α-1,4- glycosidic linkages of starch. The most thermoactive α-amylases that have been purified and characterized are those from the archaeon Pyrococcus woesei (Koch et al. 1991) and P. furiosus (Brown et al. 1990; Koch et al. 1990; Ladermann et al. 1993a,b). The extracellular α-amylase of P. woesei is a monomeric protein with a molecular mass of 68 kDa. The enzyme is active between 40°C–130°C, with an optimum at 100°C and pH 5.5. The intracellular enzyme from P. furiosus has been characterized as a homodimer with a molecular mass of 129 kDa [66 kDa/subunit (SU)]. The en- zyme is active in the range 40°C–120°C with an optimum at 100°C and pH 6.5–7.5; the half-life at 110°C is 4 h. To inac- tivate the α-amylase of P. woesei completely, autoclaving at 120°C for 6 h is necessary. An additional characteristic of Introduction Extreme thermophilic and hyperthermophilic microorgan- isms are classified as those which are adapted to grow opti- mally at temperatures ranging from 70°C to 110°C. A number of such microorganisms have been found to pro- duce thermostable enzymes which are capable of hydrolyz- ing natural polymers such as starch, cellulose, and xylan (Leuschner and Antranikian 1995; Antranikian et al. 1995). Glycosyl hydrolases are a group of enzymes which share the ability to hydrolyze the glycosidic bond between two or more carbohydrates or between a carbohydrate and a non- carbohydrate moiety. Recently, glycosyl hydrolases have been classified into more than 45 families according to amino acid sequence similarities (Henrissat 1991; Henrissat and Bairoch 1993). In starch the glucose residues are linked via α-1,4- and α-1,6-glycosidic bonds, whereas in xylan and cellu- lose the constituent sugars are linked via β-1,4-glycosidic bonds. Microorganisms capable of producing enzymes that hydrolyze these glycosidic linkages play the most important role in the biodegradation of natural polysaccharides. We briefly review glycosyl hydrolases involved in the hydrolysis of natural polysaccharides. In this review we focus on α- and β-glycosyl hydrolases from Bacteria and Archaea that grow optimally above 70°C. Glycosyl hydro- lases from microorganisms that grow below 70°C have been described elsewhere (Rüdiger et al. 1995b). Anwar Sunna · Marco Moracci · Mose Rossi Garabed Antranikian Glycosyl hydrolases from hyperthermophiles Received: September 10, 1996 / Accepted: October 12, 1996 Communicated by K. Horikoshi A. Sunna · G. Antranikian (*) Technical University Hamburg-Harburg, Institute of Biotechnology, Department of Technical Microbiology, Denickestrasse. 15, D-21071 Hamburg, Germany Tel. + 49-40-7718-3117; Fax + 49-40-7718-2909 e-mail: [email protected] M. Moracci Institute of Protein Biochemistry and Enzymology, National Research Council (CNR), Via Marconi 10, I-80125 Naples, Italy M. Rossi Dipartimento di Chimica Organica e Biologica, Universita di Napoli, Via Mezzocannone 16, I-80134 Naples, Italy

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

2 N. Matsuda et al.: EGF receptor and osteoblastic differentiationExtremophiles (1997) 1:2–13 © Springer-Verlag 1997
REVIEW
Starch-degrading enzymes
Starch is, after cellulose, one of the most abundant polysac-charides produced by plants and is composed of amylose(15%–25%) and amylopectin (75%–85%). Amylose is alinear molecule consisting of 1,4-linked α-d-glucopyranoseresidues. Amylopectin is a branched polymer and containsα-1,6 glycosidic linkages in addition to the α-1,4 bonds. Thehydrolysis of starch requires the coordinate action of sev-eral enzymes, such as α-amylase, β-amylase, glucoamylase,debranching enzymes (pullulanases), and α-glucosidase.
The biodegradation of starch is widespread among Bac-teria, fungi, and yeast (Antranikian 1992). Several extremethermophilic and hyperthermophilic Bacteria and Archaeahave been found to grow at high temperatures (>70°C)on starch and produce extremely thermostable starch-degrading enzymes (Leuschner and Antranikian 1995).Table 1 summarizes some properties of selected thermo-stable starch-hydrolyzing enzymes.
α-Amylase (1,4-α-d-glucan,4-glucanhydrolase, EC 3.2.1.1)
This endoacting enzyme produces oligosaccharides and glu-cose as end-products by randomly hydrolyzing the α-1,4-glycosidic linkages of starch.
The most thermoactive α-amylases that have beenpurified and characterized are those from the archaeonPyrococcus woesei (Koch et al. 1991) and P. furiosus(Brown et al. 1990; Koch et al. 1990; Ladermann et al.1993a,b). The extracellular α-amylase of P. woesei is amonomeric protein with a molecular mass of 68kDa. Theenzyme is active between 40°C–130°C, with an optimumat 100°C and pH 5.5. The intracellular enzyme from P.furiosus has been characterized as a homodimer with amolecular mass of 129kDa [66kDa/subunit (SU)]. The en-zyme is active in the range 40°C–120°C with an optimum at100°C and pH 6.5–7.5; the half-life at 110°C is 4h. To inac-tivate the α-amylase of P. woesei completely, autoclaving at120°C for 6h is necessary. An additional characteristic of
Introduction
Extreme thermophilic and hyperthermophilic microorgan-isms are classified as those which are adapted to grow opti-mally at temperatures ranging from 70°C to 110°C. Anumber of such microorganisms have been found to pro-duce thermostable enzymes which are capable of hydrolyz-ing natural polymers such as starch, cellulose, and xylan(Leuschner and Antranikian 1995; Antranikian et al. 1995).
Glycosyl hydrolases are a group of enzymes which sharethe ability to hydrolyze the glycosidic bond between twoor more carbohydrates or between a carbohydrate and a non-carbohydrate moiety. Recently, glycosyl hydrolases havebeen classified into more than 45 families according to aminoacid sequence similarities (Henrissat 1991; Henrissat andBairoch 1993). In starch the glucose residues are linked viaα-1,4- and α-1,6-glycosidic bonds, whereas in xylan and cellu-lose the constituent sugars are linked via β-1,4-glycosidicbonds. Microorganisms capable of producing enzymes thathydrolyze these glycosidic linkages play the most importantrole in the biodegradation of natural polysaccharides.
We briefly review glycosyl hydrolases involved in thehydrolysis of natural polysaccharides. In this review wefocus on α- and β-glycosyl hydrolases from Bacteria andArchaea that grow optimally above 70°C. Glycosyl hydro-lases from microorganisms that grow below 70°C have beendescribed elsewhere (Rüdiger et al. 1995b).
Anwar Sunna · Marco Moracci · Mose RossiGarabed Antranikian
Glycosyl hydrolases from hyperthermophiles
Received: September 10, 1996 / Accepted: October 12, 1996
Communicated by K. Horikoshi
A. Sunna · G. Antranikian (*)Technical University Hamburg-Harburg, Institute of Biotechnology,Department of Technical Microbiology, Denickestrasse. 15, D-21071Hamburg, GermanyTel. +49-40-7718-3117; Fax +49-40-7718-2909e-mail: [email protected]
M. MoracciInstitute of Protein Biochemistry and Enzymology, NationalResearch Council (CNR), Via Marconi 10, I-80125 Naples, Italy
M. RossiDipartimento di Chimica Organica e Biologica, Universita di Napoli,Via Mezzocannone 16, I-80134 Naples, Italy

B. Jochimsen et al.: Stetteria hydrogenophila 3Table 1. Physicochemical properties of selected α-glycosyl hydrolases from extremely thermophilic Bacteria and hyperthermophilic Archaea
Organism Growth Enzymes Samplea Optimal Optimal Mrb Remarks Reference
temperature temperature pH (kDa)(°C) (°C)
Archaea:Desulfurococcus 85 α-Amylase Crude 100 5.5 Canganella et al.mucosus extract 1994
Pullulanase Crude 100 5.0 Canganella et al.type II extract 1994
Pyrococcus 100 α-Amylase Purified 100 6.5–7.5 66/subunit Intracellular/ Ladermannfuriosus (129) cloned et al. 1993a,b
α-Amylase Purified/ 100 7.0 68 Extracellular/ Jorgensen et al.clone cloned 1996
Pullulanase Purified 98 5.5 110 Extracellular/ Brown andtype II glycoprotein Kelly 1993
α-Glucosidase Purified 115 5.5 125 Intracellular Costantino et al.1990
Pyrococcus 100 α-Amylase Purified 100 5.5 68 Extracellular Koch et al. 1991woesei Pullulanase Purified/ 100 6.0 90 Cell-associated/ Rüdiger et al.
type II clone cloned 1995a
α-Glucosidase Purified 110 5.0–5.5 Intracellular Linke et al. 1992
Pyrodictium 98 α-Amylase Crude 90 5.0 Andrade et al.abysii extract 1996
Pullulanase Crude 100 9.0 Andrade et al.type II extract 1996
Staphylothermus 90 α-Amylase Crude 100 5.0 Canganella et al.marinus extract 1994
Sulfolobus 88 Glucoamylase Partially 70 5.5 Cytoplasmic Lama et al. 1991solfataricus MT-4 purified fraction
Sulfolobus 80 α-Amylase Purified 120/subunit Extracellular Haseltine et al.solfataricus 98/2 (240) 1996
α-Glucosidase Purified 105 4.5 80/subunit Intracellular Rolfsmeier and(400) Blum 1995
Thermococcus 85 α-Amylase Crude 90 5.5 Canganella et al.celer extract 1994
Pullulanase Crude 90 5.5 Canganella et al.type II extract 1994
Thermococcus 90 Pullulanase Purified 98 5.5 119 Extracellular/ Brown andlitoralis type II glycoprotein Kelly 1993
Thermococcus 80 α-Amylase Purified 80 5.5–6.0 42 Extracellular Chung et al.profundus 1995DT5432
Thermococcus 75 α-Glucosidase Purified 75c 7.0 60 Extracellular Piller et al. 1996strain AN1
Thermococcus 85 α-Amylase Crude 100 5.5 Canganella et al.strain TY extract 1994
Pullulanase Crude 100 6.5 Canganella et al.type II extract 1994
Thermococcus 85 α-Amylase Crude 100 6.5 Canganella et al.strain TYS extract 1994
Pullulanase Crude 100 6.5 Canganella et al.type II extract 1994
Bacteria:Dictyoglomus 78 α-Amylase Purified/ 90 5.5 75 Cytoplasmic Fukusumi et al.thermophilum clone fraction 1988H-6-12
Thermotoga 80 Glucoamylase/ Partially 90 6.0 60 Toga-associated Schumann et al.maritima MSB8 α-amylase purified 1991
Thermus 75 Pullulanase Purified 75 5.5 65 Cell-associated Kim et al. 1996caldophilus type IGK-24
a Pure enzymes or crude extracts were used for enzyme characterization.b Values in brackets represent the size of the native enzyme.c Assay temperature.

4 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
these two enzymes is that, unlike mesophilic α-amylases,they do not require Ca2+ for activity or thermostability.
The P. woesei α-amylase is capable of hydrolyzing ran-domly the α-1,4-glycosidic linkages in various substratessuch as soluble and native starch, glycogen, amylopectin,and amylose. The results of hydrolysis are various oligosac-charides, but unlike other α-amylases, glucose is not formedas an end-product. Maltoheptaose is the smallest substratethat can be hydrolyzed by this enzyme. The intracellular α-amylase of P. furiosus hydrolyzes starch to glucose andmaltose in addition to a mixture of oligosaccharides.
The gene encoding for a distinct thermostable extracel-lular α-amylase from P. furiosus has been cloned, se-quenced, and expressed in E. coli and B. subtilis (Jorgensenet al. 1996 unpublished work). This recombinant α-amylaseshows sequence homology to the α-amylases belonging tofamily 13 of glycosyl hydrolases. This enzyme, however, isnot homologous to the intracellular α-amylase of the samearchaeon (Ladermann et al. 1993a,b). The gene encodingfor an intracellular α-amylase from P. furiosus has also beencloned and expressed in E. coli (Ladermann et al. 1993a).The recombinant α-amylase has no leader sequence and itsamino acid sequence is homologous to that of the α-amylasefrom Dictyoglomus thermophilum (Fukusumi et al. 1988).
The hyperthermophilic archaeon, Thermococcusprofundus DT5432, has been reported to produce extracel-lular thermostable amylases. One of these enzymes, namelyamylase S, has been purified and characterized (Chung et al.1995). This enzyme consists of a single subunit with a mo-lecular mass of 42kDa. The enzyme exhibits catalytic activ-ity between 40°C and 100°C with an optimum at 80°C andpH 5.5–6.0. The half-life of the enzyme at 90°C is 15min. Atthe same temperature in the presence of Ca2+, however, thehalf-life is increased to 4h.
The amylase S of T. profundus DT 5432 hydrolyzessoluble starch, glycogen, amylose, and amylopectin produc-ing mainly maltose and maltotriose as major end-products.Maltose is not hydrolyzed and, like the α-amylases from P.furiosus, amylase S possesses transferase activity.
Although only few archaeal amylases have been purifiedand characterized, the production of amylases seems to bewidely distributed among Archaea. Canganella et al. (1994)and Andrade et al. (1996) reported on the presence of ther-mostable amylases in the crude extracts of two archaeabelonging to the order Thermoproteales (Desulfurococcusmucosus, Staphylothermus marinus), four archaea belong-ing to the order Thermococcales (Thermococcus celer, T.litoralis, Thermococcus strain TYS, and Thermococcusstrain TY), and one archaeon belonging to the orderPyrodictiales (Pyrodictium abyssi). The enzymes are opti-mally active between 90°C and 100°C and at pH values of5.0–6.0. All the enzymes exhibit remarkable thermostabil-ity in the absence of substrate and calcium ions.
The extreme thermoacidophilic archaeon Sulfolobussolfataricus is able to grow on several sugars, includingstarch and maltose, as carbon and energy sources (Grogan1989). A thermostable amylolytic enzyme from the cyto-plasmic fraction of S. solfataricus strain MT-4 was partiallypurified and characterized (Lama et al. 1991). The enzyme
is optimally active at 70°C and pH 5.5 and shows no calciumrequirement for thermostability. The main productsresulting from the hydrolysis of soluble starch, amylose,and amylopectin were glucose and trehalose, indicating thatthe amylolytic activity is of the glucoamylase type. Re-cently, an α-amylase from the culture supernatant ofSulfolobus solfataricus 98/2 grown on starch was purified(Haseltine et al. 1996). The enzyme is a homodimer with asubunit size of 120kDa. The enzyme is an endoactingamylase capable of hydrolyzing starch, dextrin, and α-cyclodextrin with similar efficiencies. The α-amylase is pro-duced constitutively at low levels and the addition of starchinduces the synthesis of higher levels of enzyme. Glucose,on the other hand, represses the synthesis of α-amylase bycatabolite repression.
The anaerobic thermophilic bacterium Dictyoglomusthermophilum produces a heat-stable amylase complexwhen grown on starch (Kobayashi et al. 1988). The prelimi-nary characterization of the crude extract revealed an opti-mal temperature and pH for activity at 90°C and 5.0,respectively. In the absence of substrate the amylolytic ac-tivity was stabilized in the presence of calcium. Chromato-graphic fractionation of the crude extract showed thepresence of at least four fractions with different amylolyticactivities. One amylase gene (amyA) from Dictyoglomusthermophilum has been cloned and expressed in E. coli(Fukusumi et al. 1988). The recombinant enzyme did notpossess an NH2-secretion signal peptide and was found tobe localized in the cytoplasmic fraction of E. coli. The puri-fied recombinant amylase shows a molecular mass of 75kDaand is optimally active at 90°C and pH 5.5. The amylasehydrolyzes starch to produce mainly maltotriose, maltose,and glucose.
The extreme thermophilic anaerobic bacteriumThermotoga maritima is able to grow on starch as a carbonsource without releasing amylase into the culture medium(Schumann et al. 1991; Jaenicke 1993). Two distinct amy-lolytic fractions were found to be associated with the “toga”(the outer sheath covering the cells). Fraction I seems topossess β-amylase that is optimally active at 95°C and pH5.0. Fraction II, on the other hand, seems to contain α-amylase and glucoamylase activities with an optimum at90°C and pH 6.0.
Pullulanase type I (α-dextrin6-glucanohydrolase, EC 3.2.1.41)
Pullulanase type I hydrolyzes exclusively the α-1,6-glycosidic linkages in branched oligosaccharides, polysac-charides, and pullulan to form linear oligosaccharides asend-products. Most of the thermoactive pullulanases stud-ied so far belong to the type II pullulanases, and attack bothα-1,4- and α-1,6-glycosidic linkages in branched substrates.
The aerobic thermophilic bacterium Thermuscaldophilus GK-24 grows optimally at 75°C and producesthermostable pullulanase of type I when grown on starch(Kim et al. 1996). The enzyme has a molecular mass of65kDa and is optimally active at 75°C and pH 5.5. The

B. Jochimsen et al.: Stetteria hydrogenophila 5
pullulanase is thermostable up to 90°C and Ca2+ ions do nothave any effect on the enzyme activity. The pullulanaseattacks the α-1,6-linkages of pullulan and producesmaltotriose. The α-1,6-linkages of α,β-limit dextrins, amy-lopectin, and glycogen are hydrolyzed less efficiently. Adebranching enzyme with similar properties was purifiedfrom a newly isolated anaerobic bacterium which was iden-tified as Fervidobacterium pennavorans strain Ven 5. Theenzyme, with a molecular mass of 240kDa (77kDa/SU), isoptimally active at 85°C and pH 5.0–7.0 (Leuschner andAntranikian 1995).
Pullulanase type II (amylopullulanase)
This class of enzyme is capable of randomly hydrolyzingα-1,4-linkages in addition to the branching points (α-1,6-linkages) in polysaccharides and limit dextrins. The action ofpullulanase type II on poly- and oligosaccharides results inthe formation of glucose, maltose, and maltotriose as majorproducts (Antranikian 1992). Unlike pullulanase type I,pullulanase type II is widely distributed among extremethermophilic Bacteria and hyperthermophilic Archaea.
The most thermostable and thermoactive pullulanasetype II reported to date are derived from thehyperthermophilic archaeon Pyrococcus woesei (Linke etal. 1992; Rüdiger et al. 1995a) and P. furiosus (Brown et al.1990; Brown and Kelly 1993). Unlike the pyrococcal α-amylase, the pullulanase was found to be cell-associated.The gene encoding the pullulanase of P. woesei was clonedand expressed in E. coli. This is the first archaealpullulanase that has been cloned and expressed in amesophilic host (Linke et al. 1992; Rüdiger et al. 1995a).The purification of the recombinant pullulanase wasachieved by thermal denaturation of the host proteinand recovery of the thermostable pullulanase in thesupernatant. The enzyme possesses a signal sequence and iscomposed of a single polypeptide chain with a molecularmass of 90kDa. The pullulanase is optimally active at 100°Cand pH 6.0. The half-life of the enzyme at 110°C in thepresence of maltodextrin (0.25%) or Ca2+ (1mM) is raisedfrom 7 to 20min. Enzyme activation (up to 370%) isachieved in the presence of Ca2+ (0.3mM) and reducingagents such as β-mercaptoethanol (10mM) anddithiothreitol (10mM).
The pullulanases of type II which are produced byThermococcus litoralis and P. furiosus are glycoproteinswith a molecular mass of 119 and 110kDa, respectively(Brown and Kelly 1993). Both enzymes are optimally activeat around 100°C and pH 5.5. The presence of Ca2+ (5mM)had a positive effect on the activity of both enzymes at hightemperatures. At 120°C in the presence of substrate(maltoheptaose) and Ca2+, the pullulanases of T. litoralisand P. furiosus have half-lives of 5 and 12min, respectively.In addition to thermal stability, calcium stimulates thepullulan-hydrolyzing activity of these enzymes while inhib-iting the starch-hydrolyzing activity. At high temperaturesboth enzymes show higher specificity towards pullulan thantowards starch or glycogen.
Thermoactive pullulanases of type II also have been re-ported from Thermococcus celer, Thermococcus strainTYS, Thermococcus strain TY, Desulfurococcus mucosus,and Staphylothermus marinus (Canganella et al. 1994).Only recently, the pullulanase from the hyperthermophilePyrodictium abyssi was characterized (Andrade et al. 1996).The enzyme is optimally active at 100°C and at alkaline pH(pH 9.0).
α-Glucosidase (α-d-glucosideglucohydrolase, EC 3.2.1.20)
α-Glucosidases hydrolyze terminal, nonreducing 1,4- and,to a lesser extent, 1,6-linked α-glucose residues of disaccha-rides, oligosaccharides, and aryl-glucosides. They are in-volved in the last step of the degradation of starch, formingglucose as an end-product. Organisms growing on starchproduce intracellular α-glucosidases together with extracel-lular amylolytic enzymes; these activities constitute thestarch-degrading system. Among Archaea, α-glucosidaseshave been characterized from two species of the genusPyrococcus: P. furiosus and P. woesei (Costantino et al.1990; Leuschner and Antranikian 1995).
The α-glucosidase activity in P. furiosus is mostlyintracellular, and its expression, as for other starch-degrading enzymes from this source, is induced by carbohy-drates containing α-1,4- linkages (Costantino et al. 1990).However, the mechanism of induction remains unknown.This α-glucosidase has highest activity toward maltose andisomaltose, but significant activity against aryl-glucosidesis also displayed. The enzyme is a monomer with amolecular mass of 125kDa, it has optimal activity overa broad temperature range, about 105°C–115°C, and itexhibits remarkable thermal stability (2 days at 98°C).The observed inactivation in the presence of EDTA sug-gests that metal ions may be important for activity and/orstability.
Recently, a study by subtractive hybridization of theregulation of the genes for maltose fermentation in P.furiosus was reported (Robinson et al. 1994). Althoughsome of the genes isolated with this technique are requiredfor maltose utilization (i.e., pyruvate dikinase), being typi-cal of the pyroglycolysis pathway, no α-glycohydrolase-encoding genes were isolated.
An intracellular α-glucosidase activity has been purifiedto homogeneity in S. solfataricus strain 98/2 (Rolfsmeierand Blum 1995). The enzyme is made of four to fiveidentical subunits of 80kDa; it displays maximal activity onmaltose, and is active to a lesser extent on aryl-glucosides,but not on starch. Maltosaccharides of up to seven glucoseunits are substrates of the enzyme that is also moderatelyactive on α-1,6 disaccharides. Interestingly, the enzymeis optimally active at 105°C, more than 20°C above theoptimal growth temperature of this S. solfataricus strain,and has an half-life of 3 h at 95°C. These data suggestthat the enzyme is structurally different from P. furiosusenzymes although they show similar specificities. The iden-tification of maltose as preferred substrate would suggest

6 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
that the S. solfataricus α-glucosidase is a proper maltasethat allows the use of this sugar as a carbon source invivo.
The purification and characterization of an extracellularα-glucosidase from the archaeon Thermococcus strainAN1 has been recently reported (Piller et al. 1996). Theenzyme is a monomer with a molecular mass of 60kDaand a half-life of 35min at 98°C; the substrate preference isaryl-α-d-glucoside > nigerose > panose > palatinose >isomaltose > maltose > turanose. These authors reportedthat the enzyme reduces its thermostability on purification;however, its half-life is increased in the presence ofseveral additives such as sorbitol, dithiothreitol, or bovineserum albumin (BSA) and by co-cross-linking with BSA.The biochemical data reported show a certain varietyamong the α-glucosidases from hyperthermophilic Archaea:α-glucosidases from P. furiosus, P. woesei, and S.solfataricus are intracellular and oligomeric, whereas theenzyme from Thermococcus is smaller, monomeric, andmainly extracellular. However, in the absence of the genescoding for these enzymes, it is not possible to analysein detail the structural similarities among archaeal α-glucosidases.
Xylan-degrading enzymes
Xylan is the major component of plant hemicelluloses.Plant xylans are heterogeneous polysaccharides consistingof a main chain of β-1,4-linked d-xylopyranosyl residues.The backbone structure contains, depending on the origin,acetyl, arabinosyl, and glucuronosyl substituents. Thedepolymerization action of the endoxylanase resultsin the conversion of the polymeric substrate intoxylooligosaccharides, which can be further hydrolyzed toxylose by the action of the β-xylosidase. The side-chainsubstituents are removed by a set of enzymes whichinclude α-arabinofuranosidase, acetylxylan esterases, andα-glucuronidase. In Table 2 some properties of thermo-stable xylan-hydrolyzing enzymes are presented.
Endoxylanase (1,4-β-d-xylan xylanohydrolase,EC 3.2.1.8)
β-1,4-Endoxylanases catalyze the hydrolysis of the internalglycosidic linkages of a heteroxylan backbone resulting in adecreased degree of polymerization of the substrate. Themost thermostable endoxylanases that have been describedso far are those purified from the anaerobic thermophilicspecies of the genus Thermotoga, namely Thermotoga sp.strain FjSS3-B.1 (Simpson et al. 1991), T. maritima(Winterhalter and Liebl 1995), T. neapolitana (Bok et al.1994), and T. thermarum (Sunna et al. 1996). Like the α-amylase produced by T. maritima, all the thermostablexylanases produced by these microorganisms are mainlycell-associated and most probably localized within the toga(Schumann et al. 1991; Ruttersmith et al. 1992; Winterhalterand Liebl 1995; Sunna and Antranikian 1996).
Thermotoga sp. strain FjSS3-B.1 produces a thermo-stable endoxylanase with a molecular mass of 31kDa(Simpson et al. 1991). The endoxylanase is optimally activeat 105°C and pH 5.4. The enzyme is thermostable at 95°Cand exhibits a half-life of 8min at 100°C. At 105°C the half-life of the enzyme is less than 2min; however, in the pres-ence of 0.25% xylan alone or 0.25% xylan plus 0.2% BSA,the half-life of the enzyme increases to 9 and 11min, respec-tively. The endoxylanase hydrolyzes oat spelts xylan tomainly xylobiose and xylotriose.
Three xylanase genes (xynA, xynB, and xynC) havebeen found in Thermotoga sp. strain FjSS3-B.1 (Saul et al.1995). One gene, namely xynA, has been isolated and over-expressed in E. coli. The xynA gene encodes a thermostablexylanase with a molecular mass of 40.5kDa. This enzymebelongs to the glycosyl hydrolase family 10 and is closelyrelated to XynZ from Clostridium thermocellum (Grépinetet al. 1988) and XynA from the thermophilic anaerobeCaldicellulosiruptor Rt8B.4 (Dwivedi et al. 1996). However,the xynA gene from Thermotoga sp. strain FjSS3-B.1 showsonly low homology to the T. maritima xynA gene reportedby Winterhalter et al. (1995). The pH optimum and thermo-stability of the recombinant XynA differ from those re-ported previously for the purified endoxylanase fromThermotoga sp. strain FjSS3-B.1. Thus, the recombinant
Table 2. Physicochemical properties of selected β-glycosyl hydrolases from extremely thermophilic Bacteria and hyperthermophilic Archaea
Organism Growth Enzyme Samplea Optimal Optimal Mrb Remarks Reference
temperature temperature pH (kDa)(°C) (°C)
Archaea:Pyrococcus 100 β-Glucosidase Purified 102–105 5.0 58/subunit Intracellular/ Kengen et al.furiosus (230) cloned 1993
Pyrodictium 98 Endoxylanase Crude 110 5.5 Andrade et al.abysii extract 1996
Sulfolobus 88 β-Glycosidase Purified >90 6.5 56/subunit Intracellular/ Nucci et al. 1993solfataricus (240) cloned/broadMT-4 specificity
Sulfolobus 80 β-Glycosidase Purified >77c 5.0 60–65/ Intracellular/ Grogan 1991solfataricus P2 subunit broad specificity
(>200)
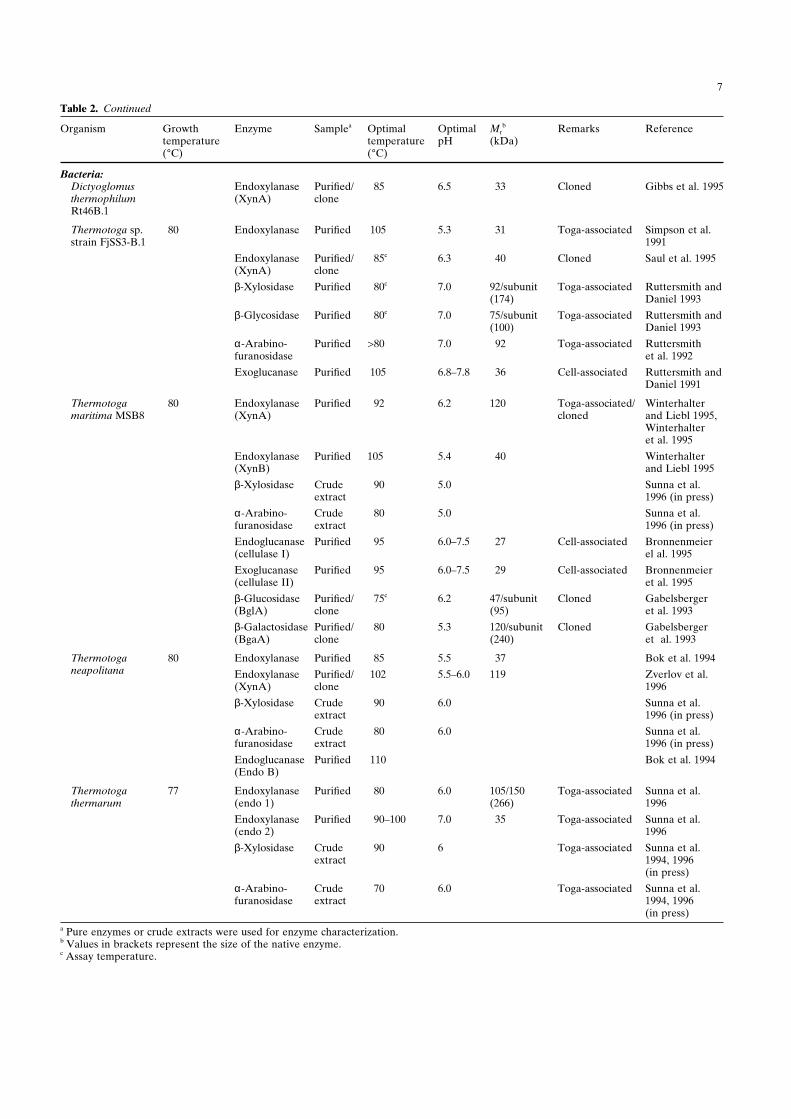
B. Jochimsen et al.: Stetteria hydrogenophila 7
Table 2. Continued
Organism Growth Enzyme Samplea Optimal Optimal Mrb Remarks Reference
temperature temperature pH (kDa)(°C) (°C)
Bacteria:Dictyoglomus Endoxylanase Purified/ >85 6.5 33 Cloned Gibbs et al. 1995thermophilum (XynA) cloneRt46B.1
Thermotoga sp. 80 Endoxylanase Purified 105 5.3 31 Toga-associated Simpson et al.strain FjSS3-B.1 1991
Endoxylanase Purified/ >85c 6.3 40 Cloned Saul et al. 1995(XynA) clone
β-Xylosidase Purified >80c 7.0 92/subunit Toga-associated Ruttersmith and(174) Daniel 1993
β-Glycosidase Purified >80c 7.0 75/subunit Toga-associated Ruttersmith and(100) Daniel 1993
α-Arabino- Purified >80 7.0 92 Toga-associated Ruttersmithfuranosidase et al. 1992
Exoglucanase Purified 105 6.8–7.8 36 Cell-associated Ruttersmith andDaniel 1991
Thermotoga 80 Endoxylanase Purified >92 6.2 120 Toga-associated/ Winterhaltermaritima MSB8 (XynA) cloned and Liebl 1995,
Winterhalteret al. 1995
Endoxylanase Purified 105 5.4 40 Winterhalter(XynB) and Liebl 1995
β-Xylosidase Crude >90 5.0 Sunna et al.extract 1996 (in press)
α-Arabino- Crude >80 5.0 Sunna et al.furanosidase extract 1996 (in press)
Endoglucanase Purified >95 6.0–7.5 27 Cell-associated Bronnenmeier(cellulase I) el al. 1995
Exoglucanase Purified >95 6.0–7.5 29 Cell-associated Bronnenmeier(cellulase II) et al. 1995
β-Glucosidase Purified/ >75c 6.2 47/subunit Cloned Gabelsberger(BglA) clone (95) et al. 1993
β-Galactosidase Purified/ >80 5.3 120/subunit Cloned Gabelsberger(BgaA) clone (240) et al. 1993
Thermotoga 80 Endoxylanase Purified >85 5.5 37 Bok et al. 1994neapolitana Endoxylanase Purified/ 102 5.5–6.0 119 Zverlov et al.
(XynA) clone 1996
β-Xylosidase Crude >90 6.0 Sunna et al.extract 1996 (in press)
α-Arabino- Crude >80 6.0 Sunna et al.furanosidase extract 1996 (in press)
Endoglucanase Purified 110 Bok et al. 1994(Endo B)
Thermotoga 77 Endoxylanase Purified >80 6.0 105/150 Toga-associated Sunna et al.thermarum (endo 1) (266) 1996
Endoxylanase Purified >90–100 7.0 35 Toga-associated Sunna et al.(endo 2) 1996
β-Xylosidase Crude >90 6 Toga-associated Sunna et al.extract 1994, 1996
(in press)
α-Arabino- Crude >70 6.0 Toga-associated Sunna et al.furanosidase extract 1994, 1996
(in press)a Pure enzymes or crude extracts were used for enzyme characterization.b Values in brackets represent the size of the native enzyme.c Assay temperature.

8 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
XynA seem to be similar but not identical to the enzymeoriginally purified by Simpson et al. (1991).
During growth with xylan or xylose as the sole carbonsource, Thermotoga maritima MSB8 produces two distinctthermostable endoxylanases referred to as XynA and XynB(Winterhalter and Liebl 1995). XynA is a membrane-bound, multidomain enzyme with a molecular mass of120kDa and optimal activity at 92°C and pH 6.2. XynB onthe other hand, has a molecular mass of 40kDa and isoptimally active at 105°C and pH 5.4. Both enzymes pro-duce mainly xylotriose, xylobiose, and low amounts of xy-lose upon hydrolysis of oat spelts xylan. Unlike XynA,XynB activity is stimulated twofold by the addition of500mM NaCl. XynA is able to adsorb to microcrystallinecellulose, indicating the presence of a cellulose-binding do-main. Both enzymes show high substrate specificity towardsdifferent xylans and none of the enzymes was able to hydro-lyze cellulose, mannan, arabinogalactan, pullulan, amylose,or starch. However, in contrast to XynA, XynB was able tocleave aryl β-d-xylosides.
XynA and XynB are encoded by two different genes.The xynA gene encoding for the thermostableendoxylanase XynA has been cloned and expressed in E.coli (Winterhalter et al. 1995). XynA is a modular enzymecomposed of five domains. The XynA of T. maritimaMSB8 is most closely related to the xylanase ofThermoanaerobacterium saccharolyticum B6A-RI (Lee etal. 1993). The catalytic domain of XynA shares aminoacid sequence similarity with xylanases of the glycosylhydrolase family 10. Cellulose-binding studies indicatethat the C-terminal domains C1 and C2 promote cellulose-binding. These two domains represent cellulose-bindingdomains of a novel type that do not share amino acidsequence similarity to any other known cellulose-bindingdomain.
Thermotoga neapolitana is capable of growth on xylan atup to 90°C and produces several thermostable xylanases(Sunna et al. 1992). One endoxylanase from T. neapolitanahas been purified and characterized (Bok et al. 1994). Theenzyme is a monomeric protein with a molecular mass of37kDa. The temperature and pH optima are 85°C and 5.5,respectively. The xylanase is thermostable and exhibits at82°C a half-life of 130h. The main products resulting fromthe hydrolysis of xylan are xylooligosaccharides rangingfrom xylobiose to xyloheptaose.
The genes encoding for several carbohydrate-degradingenzymes of T. neapolitana have been cloned and expressedin E. coli (Dakhova et al. 1993). Recently, the nucleotidesequence of the xynA gene, encoding for the thermostableextracellular xylanase (XynA) of T. neapolitana, has beendetermined and the recombinant enzyme has been charac-terized (Zverlov et al. 1996). The molecular mass of theenzyme is 119kDa and the optimal temperature and pH foractivity are 102°C and 5.5–6.0, respectively. The enzymeexhibits remarkable thermal stability at 100°C with a half-life of 120min. The endoxylanase displays low β-1,3- and β-1,4-glucanase activity as well as cellobiohydrolase andxylosidase activities. The XynA from T. neapolitana is com-posed of three different domains, namely A, B, and C, with
domain C being responsible for cellulose binding. The en-zyme displays almost 89% identity to the multidomainXynA of T. maritima (Winterhalter et al. 1995). Further-more, high similarity was found between the XynA from T.neapolitana and the N-terminal catalytic domain of CelB ofCaldocellum saccharolyticum (Saul et al. 1990; renamed toCaldicellulosiruptor saccharolyticus).
Cultivation of the nonmarine bacterium T. thermarumon xylan or xylose is accompanied by the induction of high-molecular-mass (HMM) and low-molecular-mass (LMM)thermostable endoxylanases. In the absence of these sub-strates, however, only the LMM xylanase was present in lowamounts (Sunna and Antranikian 1996). From both frac-tions, two thermostable enzymes from T. thermarum werepurified and identified as endoxylanase 1 and endoxylanase2 (Sunna et al. 1996). Endoxylanase 1 is a dimeric enzymewith a native molecular mass of 266 kDa (105 and 150 kDa/SU). The enzyme displays maximal activity at 80°C andpH 6.0. Endoxylanase 2, on the other hand, is composedof a single subunit with a molecular mass of 35 kDa.Endoxylanase 2 is optimally active between 90°C and 100°Cand at pH 7.0. Both enzymes do not require metal ionsfor activity and were not active against carboxymethyl(CM)-cellulose. Unlike endoxylanase 1, endoxylanase 2 wasactive towards galactomannan indicating the presence of amannanase activity. Both enzymes hydrolyze partiallysoluble and insoluble xylans to produce mainly xylobiose asend-product.
The xylanases from the anaerobic thermophilicbacterium Dictyoglomus thermophilum and those fromseveral Dictyoglomus strains have been characterized byMathrani and Ahring (1992). All these enzymes have anoptimum temperature for activity at around 80°C and areactive between pH 5.5 and 9.0. Recently, a xylanase gene(xynA) from D. thermophilum Rt46B.1 was cloned and ex-pressed in E. coli (Gibbs et al. 1995). The recombinantenzyme is a single-domain xylanase which belongs to family10 of glycosyl hydrolases and shows the highest homologywith other thermophilic xylanases of the same family.The thermostable xylanase, Xyn A, is a single proteinwith a molecular mass of 33kDa. The enzyme is optimallyactive at 85°C with an optimum pH at 6.5. The XynA wasthermostable in the absence of substrate at 85°C. At thistemperature and at pH 6.8 and 9.3 the enzyme displayedhalf-lives of 24 and 6.5h, respectively. Substrate hydrolysisexperiments with xylan indicate that XynA is anendoxylanase producing xylobiose and xylotriose as mainproducts.
Archaea growing at high temperatures seem to beunable to utilize xylan as a carbon source or to producethermostable xylanases (Sunna and Antranikian [in press]).Recently, however, it was shown that the hyperther-mophilic archaeon Pyrodictium abysii is capable of produc-ing a unique thermostable endoxylanase when grown in thepresence of xylan, xylose, or arabinose (Andrade et al.1996). The enzyme, which is inducible, displays optimalactivity at 110°C and pH 5.5. The main products resultingfrom the hydrolysis of xylan are xylose, xylobiose, andxylotriose.

B. Jochimsen et al.: Stetteria hydrogenophila 9
β-Xylosidase (β-d-xyloside xylohydrolase, EC 3.2.1.37)
β-Xylosidases can be classified according to their relativeaffinities towards xylobiose and larger oligosaccharides;they generate d-xylose from short oligosaccharides andxylobiose in a concerted action with xylan-degrading en-zymes. In this respect, they play an important role in de-grading xylobiose, an inhibitor of endoxylanases.
β-Xylosidase activities from hyperthermophilic Archaeahave been found so far to be one of the enzymatic activitiesof β-glycosidases with broad specificity (see later). Bycontrast, true β-xylosidases have been purified and charac-terized from the thermophilic bacterium Thermotoga sp.strain FjSS3-B.1 (Ruttersmith and Daniel 1993). The β-xylosidase from this microorganism is associated with thetoga, and it is a homodimer (92kDa/SU), showing remark-able stability to heat (half-life 2 hours at 80°C) and anoptimum of activity at pH 7.0. This enzyme shows highactivity with xylobiose, aryl-α-l-arabinofuranoside andaryl-α-l-arabinopyranoside, but it is inhibited by aryl-β-d-xylopyranoside. Interestingly, the β-glucosidase fromThermotoga sp. strain FjSS3-B.1 is active on β-d-xylopyranoside, but not on xylobiose, and it differs from theβ-xylosidase in its structural properties (Ruttersmith andDaniel 1993).
Other β-xylosidases that are active between 50°C and100°C and at pH values from 4.0 to 9.0 have been detectedin the crude extract of T. maritima, T. neapolitana, and T.thermarum (Sunna et al. [in press]).
α-l-Arabinofuranosidase (EC 3.2.1.55)
α-Arabinofuranosidases are active against branchedarabinoxylans, arabinose-substituted xylooligosaccharides,and p-nitrophenyl-α-l-arabinofuranoside. The hydrolysisof arabinoxylans with α-arabinofuranosidase results in therelease of arabinose residues. An α-arabinofuranosidasefrom Thermotoga sp. strain FjSS3-B.1 has been purified tohomogeneity and characterized (Ruttersmith et al. 1992).The enzyme has a molecular mass of 92kDa and is thermo-stable at 98°C. The enzyme displayed high activity towardsxylobiose and is also classified as a β-xylosidase.
Also, thermostable α-arabinofuranosidase activitieshave been detected in the crude extracts of T. maritima, T.neapolitana, and T. thermarum when grown on xylan as thecarbon source (Sunna et al. 1994; Sunna et al. [in press]).The enzymes from T. maritima and T. neapolitana areoptimally active at temperatures between 80°C and 90°Cand pH values between 5.0 and 6.0. On the other hand,the α-arabinofuranosidase from T. thermarum is lessthermoactive, showing optimal activity at 70°C and pH 6.0.
Cellulose-degrading enzymes
Cellulose is the most abundant organic biopolymer in na-ture. Cellulose consists of up to 14000 glucose units, linkedtogether by β-1,4-glycosidic bonds. The enzymatic hydroly-
sis of cellulose to its monomer glucose requires the synergis-tic action of at least three different enzymes, namelyendoglucanase, exoglucanase (cellobiohydrolase), and β-glucosidase. Cellulases are widespread in fungi and Bacteriabut interestingly, until now, they have not been found inhyperthermophilic Archaea. Table 2 summarizes someproperties of selected thermostable cellulose-hydrolyzingenzymes.
Endoglucanase (1,4-β-d-glucan glucanohydrolase,EC 3.2.1.4)
This enzyme hydrolyzes cellulose randomly, producing oli-gosaccharides, cellobiose, and glucose as end-products.Recently, a thermostable cellulase (cellulase I) from T.maritima MSB8 has been characterized (Bronnenmeier etal. 1995). The enzyme was purified to homogeneity and hasa molecular mass of 27kDa. The enzyme is optimally activeat 95°C and at pH values between 6.0 and 7.5. When incu-bated without substrate at 95°C, cellulase I exhibits a half-life of about 120min. The purified cellulase I is activeagainst CM-cellulose and is accordingly classified as anendoglucanase.
Thermotoga neapolitana produces two endoglucanases,Endo A and Endo B, when grown in a medium containingcellobiose as the carbon source (Bok et al. 1994). The puri-fied Endo B exhibits a temperature optimum for activity at110°C. The enzyme has a remarkable thermostability, withhalf-lives at 100°C and 106°C of 8 and 2.2h, respectively.Both enzymes show high specificity towards CM-cellulose.
Cellobiohydrolase (1,4-β-d-glucancellobiohydrolase, EC 3.2.1.91)
Cellobiohydrolases are exoglucanases that attack thenonreducing end of cellulose to produce cellobiose. Themost thermostable cellobiohydrolase that has been re-ported so far is the one purified from the thermophilicbacterium Thermotoga sp. strain FjSS3-B.1 (Ruttersmithand Daniel 1991). The enzyme has a molecular mass of36kDa and shows maximal activity at 105°C and pH valuesbetween 6.8 and 7.8. At 108°C and 113°C the cello-biohydrolase exhibits half-lives of 70 and 7min, respec-tively. Thermostability is not enhanced by the addition ofglycerol, sorbitol, CM-cellulose, or CaCl2. However, in thepresence of NaCl (0.8M) at temperatures of 108°C and113°C the half-lives of the enzyme increased almost two-fold. The cellobiohydrolase from Thermotoga sp. strainFjSS3-B.1 is active against amorphous cellulose andCM-cellulose, with the main hydrolysis products beingcellobiose.
Bronnenmeier et al. (1995) reported the purification of athermostable cellobiohydrolase from T. maritima MSB8.The enzyme has a molecular mass of 29kDa and an optimalactivity at 95°C and pH 6.0–7.5. In the absence of substrate,at 95°C, cellobiohydrolase retains 50% of its initial activityafter 120min. The enzyme is able to hydrolyze Avicel, CM-cellulose, β-glucan, and p-nitrophenyl-β-d-cellobioside. The

10 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
main products resulting from the hydrolysis of Avicel arecellobiose and cellotriose.
β-Glucosidase (β-d-glucosideglucohydrolase, EC 3.2.1.21)
β-Glycosidases catalyse the hydrolysis of O-glycosidicbonds with β-anomeric configuration in disaccharides orbetween a saccharide and an aglycon group. Differentclasses of enzymes are able to cleave β-glycosidic bonds:(1) phospho-β-glucosidases and -galactosidases, specific forphosphorylated substrates; (2) typical β-galactosidases,relatively specific for β-galactosides; and (3) β-glucosidases.The latter group contains β-glycosidases, mostly microbial,with rather broad specificity toward β-glucosides, galacto-sides, fucosides, and xylosides; and cellobiases, the enzymeshydrolyzing cellobiose to glucose in the last step of cellulosedegradation. Cellobiose-degrading enzymes are often de-scribed as the least-stable member of the cellulolyticenzyme system and this is one of the reasons for the notice-ably widespread expectation that these enzymes fromhyperthermophiles will find important applications.
High levels of β-glucosidase activity (5% of total cellprotein) were found in P. furiosus cells grown on cellobiose(Kengen et al. 1993). Interestingly, two enzymes contribute,to different extents, to the total activity found in the cellextract. The major β-glucosidase activity isolated in P.furiosus (Pfuβ-gly) is optimally active at 102°C–105°C andis very thermostable, with half-lives of 3.5 days and 13 hoursat 100°C and 110°C, respectively. Pfuβ-gly is a tetramer of230kDa (58kDa/SU); it shows broad substrate specificity,hydrolyzing aryl-galactoside, -xyloside, and -mannoside sub-strates; and it does not require metal ions for activity. Theenzyme is unable to produce glucose from cellulose or CM-cellulose, but remarkable activity is found on disaccharidessuch as cellobiose and salicin. The enzyme is intracellularand, since the organism is unable to grow on glucose, cello-biose is expected to be transported into the cell. The genecoding for Pfuβ-gly has recently been isolated and ex-pressed in E. coli (Voorhorst et al. 1995); its transcriptionand translation initiation sites have been determined. Theorganization of the gene indicates that it is transcribed in asingle unit and no genes coding for hypothetical sugar trans-porters could be found.
β-Glucosidase has been detected in differentSulfolobales species such as S. acidocaldarius DSM 639; S.solfataricus MT4 and P2, and S. shibatae B12 (Grogan 1991)and a gene for a putative β-galactosidase from S.solfataricus DSM 1616 has been cloned by directed expres-sion in E. coli (Little et al. 1989). Interestingly, furtherstudies indicated that the true source of this gene was a S.acidocaldarius strain of unknown origin (Grogan 1991).Among these enzymes, the only ones that were purified tohomogeneity were those from S. solfataricus strains. The β-glycosidase from S. solfataricus MT4 strain (Ssβ-gly) wasoriginally classified as a β-galactosidase (Pisani et al. 1990),but further biochemical characterization demonstrated abroad substrate specificity (Nucci et al. 1993). This is in
agreement with the results previously found for the strainP2 (Grogan 1991): the β-galactosidase and the β-glucosi-dase activities reside on the same enzyme. Hence, MT4 andP2 strains are closely related, and the corresponding en-zymes show similar characteristics.
The native molecular mass of Ssβ-gly is 240kDa, beinga tetramer of 56kDa identical subunits, and it does notneed any cofactors for activity. Aryl-glycosides are the pre-ferred substrates: the enzyme hydrolyzes aryl-glucosides,-fucosides, -galactosides, and -xylosides. Noteworthy, itscatalytic efficiency is β-1,3 > β-1,4 > β-1,6, and cellotetraose> cellotriose > cellobiose, for glucose dimers and oligomers,respectively. Cellopentaose, the longest oligosaccharidetested, is hydrolyzed less efficiently. Ssβ-gly, like other en-zymes from hyperthermophiles, shows noticeable resistanceto heat and other common protein denaturants (alcohols,detergents, etc.), and enhanced activity at temperatures upto 85°C (Pisani et al. 1990). The gene coding for Ssβ-gly(lacS) has been cloned and overexpressed in Saccharomy-ces cerevisiae and E. coli, and its organization in the S.solfataricus genome was determined (Cubellis et al. 1990;Moracci et al. 1992, 1995; Prisco et al. 1994). A gene encod-ing a putative membrane protein homologous to the majorfacilitator superfamily was found with the same orientationupstream from lacS (Prisco et al. 1994). Each gene has itsown archaeal consensus promoter, but the terminator se-quences between them are absent. Northern blot analysis ofRNA prepared from S. solfataricus reveals transcripts of thedimension expected for the two genes; however, they showdifferent relative abundances, and no larger transcripts areevident. Hence, the putative permease-encoding gene istranscribed in vivo, but it is still not clear if these two genesare cotranscribed and/or regulated in a proper operon.
The physiological role of these β-glycohydrolases inArchaea is still obscure: growth on cellulose has never beenreported. Furthermore, typical cellulolytic enzymes, such asendoglucanases and cellobiohydrolases, are unknown inArchaea; presumably, other cellulolytic organisms shouldprovide cellobiose in vivo. Sulfolobus solfataricus P2 andMT4 strains can grow on cellobiose as a source of carbonand energy, but this does not significantly induce the en-zyme (Grogan 1991). In P. furiosus, the mechanism used bycells growing on cellobiose to regulate the expression ofthe enzyme is still unknown. In future, the availabilityof well-established molecular genetic techniques inhyperthermophilic Archaea will certainly improve thesestudies in vivo.
Among the enzymatic activities typical of the cellulolyticsystem that have been reported from Thermotogales, thereare several β-glucosidases. In T. maritima strain MSB8,one β-galactosidase and two β-glucosidases were found(Gabelsberger et al. 1993). Unlike the specific β-glucosidase(BglB), the β-glucosidase (BglA) shows a broad substratespecificity. β-Galactosidase (BgaA) and β glucosidase(BglA) were studied by direct expression in E. coli, andbiochemically characterized. BglA and BgaA are bothdimers of 95 and 120kDa, respectively; the BglA enzymehydrolyzes aryl-glucoside, -galactoside, and -fucoside sub-strates and several disaccharides such as cellobiose, lactose,

B. Jochimsen et al.: Stetteria hydrogenophila 11
arbutin, and salicin, while BgaA is active on aryl-galactosides and lactose substrates. These enzymes showsimilar optimal temperatures for activity, but differentoptimal pHs: 6.2 and 5.3 for BglA and BgaA, respectively.Recently, the coding sequence of BglA was reported (Lieblet al. 1994; Moore et al. 1994).
The β-glucosidase from Thermotoga strain FjSS3-B.1(Ruttersmith and Daniel 1993) shows a broad specificity byhydrolysing aryl-glucosides, -fucosides, and -xylosides, andβ-linkages in the order β-1,2 > β-1,6 > β-1,4 in disaccharides.This enzyme resembles BglA from strain MSB8, but themolecular cloning of the gene would allow a better struc-tural comparison of these enzymes. Interestingly, in FjSS3-B.1 strain, a second β-glucosidase activity was detected.
Several enzymatic activities involved in carbohydratedegradation have been identified in T. neapolitana, strainZ2706-MC24, by directly cloning and expressing their genesin E. coli (Dakhova et al. 1993). Along with amylolytic,endo- and exo-glucanase, laminarase, and xylanase activi-ties, two classes of clones expressing β-glucosidase activitieswere isolated: one more active on cellobiose, and one onaryl-glucosides. However, no further biochemical informa-tion is available on these enzymes.
Molecular cloning of the genes encoding hyper-thermophilic archaeal and bacterial β-glucosidases allowedtheir classification into glycosyl hydrolase families based onamino acid sequence similarities. Bacterial β-glucosidase(T. maritima MSB8) and archaeal β-glucosidase (P.furiosus), β-glycosidase (S. solfataricus MT4), and β-galactosidase (S. solfataricus DSM 1616, putative S.acidocaldarius) were placed in family 1 along with bacterialβ-glycosidases, 6-phospho-β-glucosidases and -galactosi-dases, cyanogenic β-glycosidases, plant myrosinases, andmammalian gut lactases. Archaeal β-glycosidases show thehighest similarity scores (approximately 80%), and muchlower scores with enzymes from other domains (Moracci etal. 1994; Voorhorst et al. 1995). Interestingly, lower similar-ity is found also between archaeal enzymes and T. maritimaBglA, suggesting that the adaptation to high temperaturesfollowed different strategies in different domains. Sequencemotifs surrounding the active site found in these enzymesare common to glycosyl hydrolase families 2, 5, 10, and 17(Jenkins et al. 1995). Furthermore, by hydrophobic clusteranalysis, an even wider structural conservation amongglycosyl hydrolases was observed (Henrissat et al. 1995).Family 1 enzymes catalyse the hydrolysis of substratesthrough a covalent intermediate and without the help ofcofactors, whereas family 2 enzymes require Mg2+ ions forfull activity: this confirms the biochemical data collectedso far. Recently, the three-dimensional structures of bothfamily 1 and family 2 mesophilic enzymes have becameavailable (Jacobson et al. 1994; Barret et al. 1995;Wiesmann et al. 1995), and S. solfataricus MT4 β-glycosidase has been crystallised (Pearl et al. 1993). Whenthe structures of hyperthermophilic glycosyl hydrolases be-come available, their detailed inspection and comparisonwith those of mesophilic homologous enzymes would cer-tainly throw some light on the origins of thermal stabilityand activity.
Acknowledgments The work performed at the authors’ laboratorieswas supported by the European Commission (Biotech Generic Project“Biotechnology of Extremophiles,” contract BIO 2-CT-93-0274).
References
Andrade CM, Morana A, de Rosa M, Antranikian G (1996) Produc-tion and characterization of amylolytic and xylanolytic enzymesfrom the hyperthermophilic archaeon Pyrodictium abyssi. First In-ternational Congress on Extremophiles, Estoril, Portugal, 2–6 June1996, Book of Abstracts: 98
Antranikian G (1992) Microbial degradation of starch. In:Winkelmann G (ed) Microbial degradation of natural products.VCH, Weinheim, pp 27–56
Antranikian G, Rüdiger A, Canganella F, Klingeberg M, Sunna A(1995) Biodegradation of polymers at temperatures up to 130°C. J MS – Pure Appl Chem A32:661–669
Barret T, Suresh CG, Tolley SP, Dodson EJ, Hughes MA (1995) Thecrystal structure of a cyanogenic β-glucosidase from white clover, afamily 1 glycosyl hydrolase. Structure 3:951–960
Bok JD, Goers SK, Eveleigh DE (1994) Cellulase and xylanase systemsof Thermotoga neapolitana. ACS Symp Ser 566:54–65
Bronnenmeier K, Kern A, Liebl W, Staudenbauer WL (1995)Purification of Thermotoga maritima enzymes for the degradationof cellulosic materials. Appl Environ Microbiol 61:1399–1407
Brown SH, Kelly RM (1993) Characterization of amylolytic enzymes,having both α-1,4 and α-1,6 hydrolytic activity from the thermophilicarchaea Pyrococcus furiosus and Thermococcus litoralis. ApplEnviron Microbiol 59:2614–2621
Brown SH, Costantino HR, Kelly RM (1990) Characterization of amy-lolytic enzyme activities associated with the hyperthermophilicarchaebacterium Pyrococcus furiosus. Appl Environ Microbiol56:1985–1991
Canganella F, Andrade CM, Antranikian G (1994) Characterization ofamylolytic and pullulytic enzymes from thermophilic archaea andfrom a new Fervidobacterium species. Appl Microbiol Biotechnol42:239–245
Chung YC, Kobayashi T, Kanai H, Akiba T, Kudo T (1995) Purifi-cation and properties of an extracellular amylase from thehyperthermophilic archaeon Thermococcus profundus DT5432.Appl Environ Microbiol 61:1502–1506
Costantino HR, Brown SH, Kelly RM (1990) Purification and charac-terization of an α-glucosidase from hyperthermophilicArchaebacterium, Pyrococcus furiosus, exhibiting a temperatureoptimum of 105°C to 115°C. J Bacteriol 172:3654–3660
Cubellis MV, Rozzo C, Montecucchi P, Rossi M (1990) Isolation andsequencing of a new β-galactosidase-encoding archaebacterial gene.Gene 94:89–94
Dakhova ON, Kurepina NE, Zverlov VV, Svetlichnyi VA,Velikodvorskaya GA (1993) Cloning and expression in Escherichiacoli of Thermotoga neapolitana genes coding for enzymes of carbo-hydrate substrate degradation. Biochem Biophys Res Commun194:1359–1364
Dwivedi PP, Gibbs MD, Saul DJ, Bergquist PL (1996) Cloning, se-quencing and overexpression in Escherichia coli of a xylanase gene,xynA, from the themophilic bacterium Caldicellulosiruptor Rt8B.4.Appl Microbiol Biotechnol 45:86–93
Fukusumi S, Kamizono A, Horinouchi S, Beppu T (1988) Cloning andnucleotide sequence of a heat-stable amylase gene from an anaero-bic thermophile, Dictyoglomus thermophilum. Eur J Biochem174:15–21
Gabelsberger J, Liebl W, Schleifer KH (1993) Cloning and character-ization of β-galactoside and β-glucoside hydrolysing enzymes ofThermotoga maritima. FEMS Microbiol Lett 109:131–138
Gibbs MD, Reeves RA, Bergquist PL (1995) Cloning, sequencing, andexpression of a xylanase gene from the extreme thermophileDictyoglomus thermophilum Rt46B.1 and activity of the enzymeon fiber-bound substrate. Appl Environ Microbiol 61:4403–4408
Grépinet O, Chebrou MC, Béguin P (1988) Nucleotide sequence anddeletion analysis of the xylanase gene (xynZ) of Clostridiumthermocellum. J Bacteriol 170:4582–4588

12 N. Matsuda et al.: EGF receptor and osteoblastic differentiation
Grogan DW (1989) Phenotypic characterization of the archaebacterialgenus Sulfolobus: comparison of five wild-type strains. J Bacteriol171:6710–6719
Grogan DW (1991) Evidence that β-galactosidase of Sulfolobussolfataricus is only one of several activities of a thermostable β-d-glycosidase. Appl Environ Microbiol 57:1644–1649
Haseltine C, Rolfsmeier M, Blum P (1996) The glucose effect andregulation of α-amylase synthesis in the hyperthermophilic archaeonSulfolobus solfataricus. J Bacteriol 178:945–950
Henrissat B (1991) A classification of glycosyl hydrolases based onamino acid sequence similarity. Biochem J 280:309–316
Henrissat B, Bairoch A (1993) New families in the classification ofglycosyl hydrolases based on amino acid sequence similarities.Biochem J 293:781–788
Henrissat B, Callebaut I, Fabrega S, Lehn P, Mornon JP, Davies G(1995) Conserved catalytic machinery and the prediction of a com-mon fold for several families of glycosyl hydrolases. Proc Natl AcadSci USA 92:7090–7094
Jacobson RH, Zhang X-J, DuBose RF, Matthews BW (1994) Three-dimensional structure of β-galactosidase from E. coli. Nature369:761–766
Jaenicke R (1993) Structure–function relationship of hyper-thermophilic enzymes. ACS Symp Ser 516:53–67
Jenkins J, Lo Leggio L, Harris G, Pickersgill R (1995) β-Glucosidase,β-galactosidase, family A cellulases, family F xylanases and twobarley glycanases form a superfamily of enzymes with 8-fold β/αarchitecture and with two conserved glutamates near the carboxy-terminal ends of β-strands four and seven. FEBS Lett 362:281–285
Kengen SWM, Luesink EJ, Stams AJM, Zehnder AJB (1993) Purifica-tion and characterization of an extremely thermostable β-glucosidase from the hyperthermophilic archaeon Pyrococcusfuriosus. Eur J Biochem 213:305–312
Kim CH, Nashiru O, Ko JH (1996) Purification and biochemical char-acterization of pullulanase type I from Thermus caldophilus GK-24.FEMS Microbiol Lett 138:147–152
Kobayashi Y, Motoike M, Fukusumi S, Ohshima T, Saiki T, Beppu T(1988) Heat-stable amylase complex produced by a strictly anaero-bic and extremely thermophilic bacterium, Dictyoglomusthermophilum. Agric Biol Chem 52:615–616
Koch R, Zablowski P, Sprinat A, Antranikian G (1990) Extremelythermostable amylolytic enzyme from the archaebacteriumPyrococcus furiosus. FEMS Microbiol Lett 71:21–26
Koch R, Spreinat A, Lemke K, Antranikian G (1991) Purificationand properties of a hyperthermoactive α-amylase from thearchaebacterium Pyrococcus woesei. Arch Microbiol 155:572–578
Ladermann KA, Asada K, Uemori T, Mukai H, Taguchi Y, Kato I,Anfinsen CB (1993a) α-Amylase from the hyperthermophilicarchaebacterium Pyrococcus furiosus. J Biol Chem 268:24402–24407
Ladermann KA, Davis BR, Krutzsch HC, Lewis MS, Griko YV,Privalov PL, Anfinsen CB (1993b) The purification and characteri-zation of an extremely thermostable α-amylase from thehyperthermophilic archaebacterium Pyrococcus furiosus. J BiolChem 268:24394–24401
Lama L, Nicolaus B, Trincone A, Morzillo P, Calandrelli V,Gambacorta A (1991) Thermostable amylolytic activity fromSulfolobus solfataricus. Biotech Forum Europe 8:201–203
Lee YE, Lowe SE, Henrissat B, Zeikus JG (1993) Characterization ofthe active site and thermostability regions of endoxylanase fromThermoanaerobacterium saccharolyticum B6A-RI. J Bacteriol175:5890–5898
Leuschner C, Antranikian G (1995) Heat-stable enzymes from ex-tremely thermophilic and hyperthermophilic microorganisms.World J Microbiol Biotechnol 11:95–114
Liebl W, Gabelsberger J, Schleifer KH (1994) Comparative amino acidsequence analysis of Thermotoga maritima β-glucosidase (BglA)deduced from the nucleotide sequence of the gene indicates distantrelationship between β-glucosidases of the BGA family and otherfamilies of β-1,4-glycosyl hydrolases. Mol Gen Genet 242:111–115
Linke B, Rüdiger A, Wittenberg G, Jorgensen PL, Antranikian G(1992) Production of heat-stable pullulanase and α-glucosidase fromthe extreme thermophilic archaeon Pyrococcus woesei. DECHEMABiotech Conf 5:161–163
Little S, Cartwright P, Campbell C, Prenneta A, McChesney J, Moun-tain A, Robinson M (1989) Nucleotide sequence of a thermostableβ-galactosidase from Sulfolobus solfataricus. Nucleic Acids Res17:7980
Mathrani IM, Ahring BK (1992) Thermophilic and alkalophilicxylanases from several Dictyoglomus isolates. Appl MicrobiolBiotechnol 38:23–27
Moore JB, Markiewicz P, Miller JH (1994) Identification and sequenc-ing of the Thermotoga maritima lacZ gene, part of a divergentlytranscribed operon. Gene 147:101–106
Moracci M, La Volpe A, Pulitzer JF, Rossi M, Ciaramella M (1992)Expression of the thermostable β-galactosidase gene from thearchaebacterium Sulfolobus solfataricus in Saccharomyces cerevisiaeand characterization of a new inducible promoter for heterologousexpression. J Bacteriol 174:873–882
Moracci M, Ciaramella M, Nucci R, Pearl LH, Sanderson I, TrinconeA, Rossi M (1994) Thermostable β-glycosidases from Sulfolobussolfataricus. Biocatalysis 11:89–103
Moracci M, Nucci R, Febbraio F, Vaccaro C, Vespa N, La Cara F,Rossi M (1995) Expression and extensive characterisation of aβ-glycosidase from the extreme thermoacidophilic archaeonSulfolobus solfataricus in Escherichia coli: authenticity of the recom-binant enzyme. Enzyme Microb Technol 17:992–997
Nucci R, Moracci M, Vaccaro C, Vespa N, Rossi M (1993) Exo-glucosidase activity and substrate specificity of the β-glycosidaseisolated from the extreme thermophile Sulfolobus solfataricus. Bio-technol Appl Biochem 17:239–250
Pearl LH, Hemmings AM, Nucci R, Rossi M (1993) Crystallisation andpreliminary X-ray analysis of the β-galactosidase from the extremethermophilic archaebacterium Sulfolobus solfataricus. J Mol Biol229:558–560
Piller K, Daniel RM, Petach HH (1996) Properties and stabilization ofan extracellular α-glucosidase from the extremely thermophilicarchaebacteria Thermococcus strain AN1: enzyme activity at 130°C.Biochim Biophys Acta 1292:197–205
Pisani FM, Rella R, Raia CA, Rozzo C, Nucci R, Gambacorta A, DeRosa M, Rossi M (1990) Thermostable β-galactosidase from thearchaebacterium Sulfolobus solfataricus – purification and proper-ties. Eur J Biochem 187:321–328
Prisco A, Moracci M, Rossi M, Ciaramella M (1994) A gene encodinga putative membrane protein homologous to the major facilitatorsuperfamily of transporters maps upstream of the β-glycosidase genein the archaeon Sulfolobus solfataricus. J Bacteriol 177:1614–1619
Robinson KA, Robb FT, Schreiner HJ (1994) Isolation of maltose-regulated genes from the hyperthermophilic archaeum, Pyrococcusfuriosus, by subtractive hybridization. Gene 148:137–141
Rolfsmeier M, Blum P (1995) Purification and characterization of amaltase from the extremely thermophilic crenarchaeote Sulfolobussolfataricus. J Bacteriol 177:482–485
Rüdiger A, Jorgensen PL, Antranikian G (1995a) Isolation and charac-terization of a heat-stable pullulanase from the hyperthermophilicarchaeon Pyrococcus woesei after cloning and expression of its genein Escherichia coli. Appl Environ Microbiol 61:567–575
Rüdiger A, Sunna A, Antranikian G (1995b) Polysaccharides-degrading enzymes. In: Drauz H, Waldmann H (eds) Enzyme cataly-sis in organic synthesis. VCH, Weinheim, pp 316–339
Ruttersmith LD, Daniel RM (1991) Thermostable cellobiohydrolasefrom the thermophilic eubacterium Thermotoga sp. strain FjSS3-B.1.Biochem J 277:887–890
Ruttersmith LD, Daniel RM (1993) Thermostable β-glucosidase and β-xylosidase from Thermotoga sp. strain FjSS3-B.1. Biochim BiophysActa 1156:167–172
Ruttersmith LD, Daniel RM, Simpson HD (1992) Cellulolytic andhemicellulolytic enzymes functional above 100°C. Ann NY Acad Sci672:137–141
Saul DJ, Williams LC, Grayling RA, Chamley LW, Love DR,Bergquist PL (1990) celB, a gene coding for a bifunctional cellulasefrom the extreme thermophile “Caldocellum saccharolyticum.” ApplEnviron Microbiol 56:3117–3124
Saul DJ, Williams LC, Reeves RA, Gibbs MD, Bergquist PL (1995)Sequence and expression of a xylanase gene from thehyperthermophile Thermotoga sp. strain FjSS3–B.1 and character-ization of the recombinant enzyme and its activity on kraft pulp.Appl Environ Microbiol 61:4110–4113

B. Jochimsen et al.: Stetteria hydrogenophila 13
Schumann J, Wrba A, Jaenicke R, Stetter KO (1991) Topographicaland enzymatic characterization of amylase from the extremely ther-mophilic eubacterium Thermotoga maritima. FEBS Lett 282:122–126
Simpson HD, Haufler UR, Daniel RM (1991) An extremely thermo-stable xylanase from the thermophilic eubacterium Thermotoga.Biochem J 277:413–417
Sunna A, Antranikian G (1996) Growth and production of xylanoly-tic enzymes by the extreme thermophilic anaerobic bacteriumThermotoga thermarum. Appl Microbiol Biotechnol 45:671–676
Sunna A, Antranikian G (in press) Xylanolytic enzymes from fungiand bacteria. CRC Crit Rev Biotechnol
Sunna A, Stoffregen T, Antranikian G (1992) Xylan-degrading en-zymes from thermophilic bacteria. DECHEMA Biotech Conf 5:97–99
Sunna A, Puls J, Antranikian G (1994) Characterization of thexylanolytic enzyme system of the thermophilic eubacteriumThermotoga thermarum. In: Alberghino L, Frontali L, Sensi P (eds)Proceedings of the 6th European Congress on Biotechnology.Elsevier, Amsterdam, pp 267–270
Sunna A, Puls J, Antranikian G (1996) Purification and characteriza-tion of two thermostable endo-1,4-β-d-xylanases from Thermotogathermarum. Biotechnol Appl Biochem 24:177–185
Sunna A, Puls J, Antranikian G (in press) Characterization of thexylanolytic enzyme system of the extreme thermophilic anaerobicbacteria Thermotoga maritima, T. neapolitana, and T. thermarum.Comp Biochem Physiol
Voorhorst WGB, Eggen RIL, Luesink EJ, de Vos WM (1995) Charac-terization of the celB gene coding for β-glucosidase from thehyperthermophilic archaeon Pyrococcus furiosus and its expressionand site-directed mutation in Escherichia coli. J Bacteriol 177:7105–7111
Wiesmann C, Beste G, Hengstenberg W, Schulz GE (1995) Thethree- dimensional structure of 6-phospho-β-galactosidase fromLactococcus lactis. Structure 3:961–968
Winterhalter C, Liebl W (1995) Two extremely thermostable xylanasesof the hyperthermophilic bacterium Thermotoga maritima MSB8.Appl Environ Microbiol 61:1810–1815
Winterhalter C, Heinrich P, Candussio A, Wich G, Liebl W(1995) Identification of a novel cellulose-binding domain withinthe multidomain 120-kDa xylanase XynA of the hyperther-mophilic bacterium Thermotoga maritima. Mol Microbiol 15:431–444
Zverlov V, Piotukh K, Dakhova O, Velikodvorskaya G, Borris R(1996) The multidomain xylanase A of the hyperthermophilic bacte-rium Thermotoga neapolitana is extremely thermoresistant. ApplMicrobiol Biotechnol 45:245–247
Related Documents











