Genregulation Literature - Alberts/Lehninger - Kim Sneppen & G. Zocchi: Physics in Molecular Biology - E. Klipp et al. : Systems Biology in Practice Systems biophysics 2010/05/11 Physics of transcription control and expression analysis

Genregulation
Dec 31, 2015
Genregulation. Physics of transcription control and expression analysis. Systems biophysics 2010/05/11. Literature Alberts/Lehninger Kim Sneppen & G. Zocchi: Physics in Molecular Biology E. Klipp et al. : Systems Biology in Practice. From genetic approach to sytemic approach. - PowerPoint PPT Presentation
Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Genregulation
Literature
- Alberts/Lehninger- Kim Sneppen & G. Zocchi: Physics in Molecular Biology- E. Klipp et al. : Systems Biology in Practice
Systems biophysics 2010/05/11
Physics of transcription control and expression analysis

From genetic approach to sytemic approach
genregulation
mRNA regulation
DNA mutations / evolution
protein functions
spatiotemporal structure formationMorphogenesis
signal transduction=> Topics of systems biophysics

Biological Pattern formation and Morphogenesis11.05.2010
Zur Anzeige wird der QuickTime™ Dekompressor „TIFF (LZW)“
benötigt.
E S k1 ES k2 E P
k 1 Enzymatic Reactions
Michaelis-Menton-KineticsInhibation, Regulation
Reaction-Diffusion-Model of Morphogenesis
Biochemical Network

E.coli as model system
E.coli has a single DNA molecule which is 4.6 106 basepairs long. It encodes 4226 proteins and a couple of RNA molecules. The information content of the genome is is bigger than the structural information of the encoded Proteins-> regulatory mechanisms are encoded
Genregulation allows adaption to changing environmental conditions, and regulation of metabolism

Content of this lecture:
Basics: Monod Model, Lac Operon
Statistical Physics of DNA-binding Proteins
Modelling of genregulatory Networks
(ODE & Boolian Networks)
Dynamics of Protein-DNA binding
DNA looping
Analysis of gene expression data
Synthetic Networks

Operon-Modell
operon
Operon: Genetic subunit, that consists of regulated genes with similar functionality.It includes- Promotor: Binding site for RNA polymerase - Operator: controls access of the RNA-Polymerase structural gene - Structural genes: Polypeptide encoding genes
Francois Jacob und Jaques Monod, 1961

The Trp Operator as a switch:• Within the promotor lies a short DNA region as binding site for a
repressor. A bound repressor prevents the Polymerase from binding.

The OUTSIDE of proteins can be recognized by proteins
Distinct basepairs can be recognized by their marginsDNA binding motivs
Small channel
Large channel

Binding of Tryptophane to the Tryptophane-Repressorproteine changes the conformation of the repressor, Repressor can bind to the repressor binding site

Identification of promotor sequences

Transcription-Activation proteins switch on genes

Gen-Regulation with Feedback:lac-Operon
LacI
IPTG, TMG

Campbell, N.A., Biology
A cis-regulatory element or cis-element is a region of DNA or RNA that regulates the expression of genes located on that same strand. This term is constructed from the Latin word cis, which means "on the same side as". These cis-regulatory elements are often binding sites of one or more trans-acting factors.
IPTG (Isopropyl β-D-1-thiogalactopyranoside)This compound is used as a molecular mimic of allolactose, a lactose metabolite that triggers transcription of the lac operon. Unlike allolactose, the sulfur (S) atom creates a chemical bond which is non-hydrolyzable by the cell, preventing the cell from "eating up" or degrading the inductant. IPTG induces activity of beta-galactosidase, an enzyme that promotes lactose utilization, by binding and inhibiting the lac repressor. In cloning experiments, the lacZ gene is replaced with the gene of interest and IPTG is then used to induce gene expression.
Non-metabolizable inducer are used to induce gene expression

Variation of Protein-Concentration with IPTG
Northern Blot: measurement of the messenger RNA (mRNA) concentration
External and internal Inductor-concentration is equal in equilibriumThe mRNA concentration increases linear with the concentration of inductor, saturation over 60%
The operon enables a variation of Protein concentration. What is missing to make a switch?
60
40
20
0
[mR
NA
]
0.100.00[IPTG Induktor]Long, C et al, J.Bacteriol. 2001

Transkription und Translation in E.coliTypical times and rates
1 Molecule / cell = 1nMComplete mass2.5 106 Da
TRANSKRIPTIONrate 1/s - 1/18sTranskriptionsrate: 30bps-90bps
TRANSLATION10.000-15.000 RibosomesTranslation rate 6-22 codons/s(40 Proteine/mRNA)

The arabinose system1
Uptake
Reporter
Regulator
Break down
pBAD24 2
~55 copies/cell
[1] R. Schleif. Trends in Genetics, 16(12):559–565, 2000[2] L. M. Guzman, D. Belin, M. J. Carson, and J. Beckwith. J.Bacteriol., 177(14):4121–4130, 1995[3] D. A. Siegele and J. C. Hu. Proc. Natl. Acad. Sci. USA, 94(15):8168–8172, 1997

automated data aquisition
define ROIs
measure total intensity
DICtn
N
DICt0
Fluorescencet0
t1
tn
background correction
calibration and conversion into molecular units
Time-lapse Fluorescence Microscopy and Quantitative Image Processing


8x105
6
4
2
0
To
tal F
luo
resc
en
ce [
a.u
.]
806040200
Time [min]
0.2% arabinose
Single cell expression kinetics
30min 40min 60min50min 70min
5min 15min 35min 45min25min
Saturating induction
Subsaturating induction
Image series correspond to blue curves
Fluorescence measurement• Cell outlines are determined using bright field images• The signal is integrated within the outline in each fluorescence image
8x105
6
4
2
0
To
tal F
luo
resc
en
ce [
a.u
.]
806040200
Time [min]
0.01% arabinose

Gene expression model
Deterministic rate model with time delay d
8x105
6
4
2
0
Z(
) [a
.u.]
806040200
[min]
Reporter module Uptake module
Induction: t=0min

Curve Fitting
Fixed Parameters
Saturating induction
Subsaturating induction
Fit Parameters
Fit expression function
Time delay
Protein synthesisrate
Literature
Measured
8x105
6
4
2
0
To
tal F
luo
resc
en
ce [
a.u
.]
806040200
Time [min]
0.01% arabinose
8x105
6
4
2
0
To
tal F
luo
resc
en
ce [
a.u
.]
806040200
Time [min]
0.2% arabinose

Ohter example: Quorum Sensing

Squid with floodlamp
Phänomena:Squid (Euprymna scolopes) emmits light due the night Squid isn´t recognized as prey in the moonlight
Explanation:
Light organ of the squid collects luminescent bacteria (Vibrio fischerei)
Question:
Why does V. fischerei emmit light within the lightorgan of the squid, but not in open sea?

Quorum sensing
Bakterien detect their own cell density Density regulates the expression of luminescent genes
K. Nelson, Cell-Cell Signalling in Bacteria
Bacteria increase exponentialOD: optical density

Molekular picture of QS
• Bakteria export oligopeptides (Pheromones)• Oligopeptides accumulate with increasing cell density• Oligopeptide diffuse into cell membrane and regulates the expression of luminescent genes

Searching the binding site

Searching the binding site: timescales
D kT
6R
P(r, t) 1
4tDexp
r2
4Dt
tdiffusion d2
2D
Stokes Einstein equation(z.B. DGFP=3-7µm2/s)
Probability distribution
1µm
Typical timescale for a proteine to find an arbitrary point in an E.coli: tD 0.1s

Diffusion to a target site (binding disc)
J D4r2 dC
dr
dC
dt
1
r2D
d
dr4r2 dC
dr
C(r) J
D4r C()
C() N V
C() 0
J 4DN
V
on V
4DN20s N

Residence times for transcription factors
1
off
1
on
V exp G kT 4D M exp G kT
(from on=20s/N follows, that 1 molecule in 1µm3 occupies half an Operator)
for specific bindings (operon) with 1M-1=1.6nm3 and Gspez=-12.6kcal/mol, =1 follows
off 20s
for unspecific binding sites with Guspez=-10-4 kcal/mol, follows
off 10 4 s

Search of the binding sites on a DNA strand
DayssD
L2000.200
2 1
2
Unspecific binding events of TFs is a problem, since the time to find a binding site is increased. For a infinite staytime, a 1D- random walk over the strand would last:
(L=1.5mm und D1≈D)

Accelerated search: jumps between strands decrease time to find a binding site.
l2
D1
L
l
Ll
D1
Mit L=1.5mm, l=150nm follows
50s

Boolian Networks, or what cells and computers have in common.

(Nature, Dec 99)

Combinatoric gene regulation: Genetic networks
transcriptiontranscription
translationtranslation
Genregulatoric proteineGenregulatoric proteine

A transcription-activator and a transcription-repressor regulate the lac-Operon

Thermodynamicc model of a combinatoric transcription logics
P : bindingprobability
Gerland et al. PNAS, 2005
Gene regulation follows the mechanics of „Boltzmann-machines“

Statistical physics of protein - DNA binding
CI O CIO
K k
k
CI O CIO
CIO Ototal
CI
K CI
Binding-isothermes:

Cooperativity due to dimer binding
CI D O CIO
KD CI M 2
CI D
CIO Ototal
CI M 2
K KD CI M 2
Cooperative binding
CI M CI M
CI D

The statistical weight of the „on“ state
Pon Z(on)
Z
Pon
Poff
Z(on)
Z(off )
c
exp G kT CI K
The free-energy difference is normalized to 1mol/l . The real change in free energy of the binding event depends on the concentration of TF in solution [Cl] :
G* kT ln Z(on) ln Z(off ) G kT ln CI
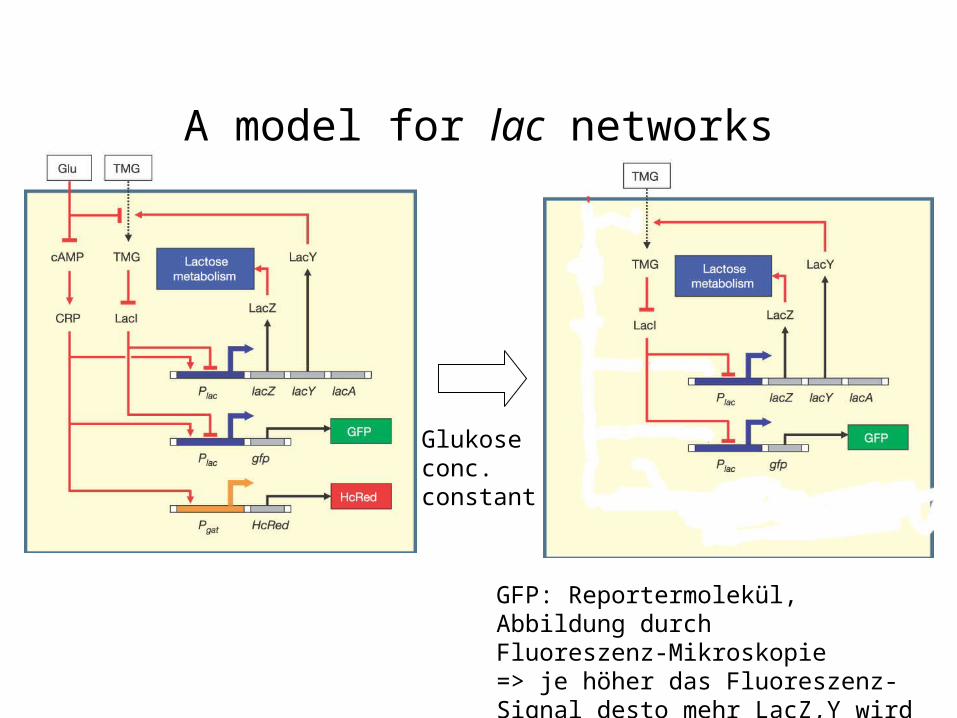
A model for lac networks
Glukoseconc.constant
GFP: Reportermolekül, Abbildung durchFluoreszenz-Mikroskopie=> je höher das Fluoreszenz-Signal desto mehr LacZ,Y wird exprimiert

Experimental proof for a switch
Start: not induced
After induction exist 2 populations:
green: induced bacteria
white, not induced population
Bistable area (grey)
Arrow marks the start state:
on-off state of bacteria depend on the on-off state in the beginning!
switch with hysteresisOzbudak et al, Nature 2004

modelling of genregulatory networks: example

Modelling in mRNA level

Timetrace of mRNA concentrations
Problem: kinetic binding constants are usually not known and hard to measure
Steady state

Simplification of genregulatory networks
transcriptiontranscription
translationtranslation
Genregulatory proteinGenregulatory protein

Abstraction of genetic networks
Gen X
Gen Y
Gen Z
+
-

Boolean networks(Kauffman 1989)

Boolean networkmodel
• N Genes (nodes)
• with 2N different states
• with possible rules
• K is the number of possible inputs per node
22K

Boolean rules for N=2 und K=2

Back to the example:
We learn: if a=0, then follows0101 stationary
if a=1, then follows oscilatory behaviour1000->1001->1111->1010->1000
Related Documents











