Hindawi Publishing Corporation e Scientific World Journal Volume 2013, Article ID 670621, 7 pages http://dx.doi.org/10.1155/2013/670621 Research Article Genetic Variants of Neurotransmitter-Related Genes and miRNAs in Egyptian Autistic Patients Ahmed M. Salem, 1 Samira Ismail, 2 Waheba A. Zarouk, 3 Olwya Abdul Baky, 4 Ahmed A. Sayed, 1 Sawsan Abd El-Hamid, 3 and Sohair Salem 3 1 Department of Biochemistry, Ain Shams University, Cairo, Egypt 2 Department of Clinical Genetics, National Research Centre, Giza, Egypt 3 Department of Molecular Genetics, National Research Centre, Giza, Egypt 4 Department of Child Psychiatry, Ain Shams University, Cairo, Egypt Correspondence should be addressed to Sohair Salem; ssalem [email protected] Received 28 August 2013; Accepted 1 October 2013 Academic Editors: K. Csiszar, S. Mastana, and J. L. Vilotte Copyright © 2013 Ahmed M. Salem et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Autism is a neurodevelopmental disorder with indisputable evidence for a genetic component. is work studied the association of autism with genetic variations in neurotransmitter-related genes, including MAOA uVNTR, MAOB rs1799836, and DRD2 TaqI A in 53 autistic patients and 30 healthy individuals. e study also analyzed sequence variations of miR-431 and miR-21. MAOA uVNTR was genotyped by PCR, MAOB and DRD2 polymorphisms were analyzed by PCR-based RFLP, and miR-431 and miR-21 were sequenced. Low expressing allele of MAOA uVNTR was frequently higher in female patients compared to that in controls (OR = 2.25). MAOB G allele frequency was more significantly increased in autistic patients than in controls ( < 0.001 for both males and females). DRD2 A1+ genotype increased autism risk (OR = 5.1). Severity of autism tends to be slightly affected by MAOA/B genotype. Plasma MAOB activity was significantly reduced in G than in A allele carrying males. ere was no significant difference in patients and maternal plasma MAOA/B activity compared to controls. Neither mutations nor SNPs in miR-431 and miR-21 were found among studied patients. is study threw light on some neurotransmitter-related genes suggesting their potential role in Autism pathogenesis that warrants further studies and much consideration. 1. Introduction Autism is a neurodevelopmental disorder characterized by disturbances in social interactions, language, and communi- cation, as well as by the presence of stereotyped behaviors and restricted interests. e population prevalence of autism is approximately 15–20 in 10,000 [1]. e assessment of candi- date genes for the identification of susceptibility loci in autism is a common molecular strategy. Autism susceptibility loci have been identified on almost every chromosome but no single chromosomal location, however, has been found to be highly significant [2]. e current study investigated genetic variations of two classes of genes, neurotransmission related genes (MAOA/B and DRD2) and miRNAs (miR-431 and miR-21). MAOA/B were chosen due to their role in enzymatic degradation. MAO has been considered a candidate gene for autism susceptibility based not only on its function, but also on its location on the X chromosome; this is due to the skewed sex ratio in autism (4 males : 1 female) [3]. An association between patients and maternal MAOA uVNTR and more severe symptoms of autism spectrum disorders (ASD) was reported [4, 5]. DRD2 gene is related to receptors in postsynaptic component. Functional polymorphisms which affect receptor availability, either post- or presynaptically, may contribute to the impair- ments found in individuals with autism [6]. e role of the DRD2 gene in autism susceptibility was suggested by the fact that antipsychotic medications, which prevent dopamine D2 receptor activation, improve the core symptoms of ASDs [7]. Monoamine oxidase A (MAOA) gene contains a 30 bp variable number of tandem repeats (VNTR) in the promoter (1.2 kb upstream) region, termed uVNTR MAOA. Although

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Hindawi Publishing CorporationThe Scientific World JournalVolume 2013 Article ID 670621 7 pageshttpdxdoiorg1011552013670621
Research ArticleGenetic Variants of Neurotransmitter-Related Genes andmiRNAs in Egyptian Autistic Patients
Ahmed M Salem1 Samira Ismail2 Waheba A Zarouk3 Olwya Abdul Baky4
Ahmed A Sayed1 Sawsan Abd El-Hamid3 and Sohair Salem3
1 Department of Biochemistry Ain Shams University Cairo Egypt2 Department of Clinical Genetics National Research Centre Giza Egypt3 Department of Molecular Genetics National Research Centre Giza Egypt4Department of Child Psychiatry Ain Shams University Cairo Egypt
Correspondence should be addressed to Sohair Salem ssalem nrchotmailcom
Received 28 August 2013 Accepted 1 October 2013
Academic Editors K Csiszar S Mastana and J L Vilotte
Copyright copy 2013 Ahmed M Salem et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Autism is a neurodevelopmental disorder with indisputable evidence for a genetic component This work studied the associationof autism with genetic variations in neurotransmitter-related genes includingMAOA uVNTR MAOB rs1799836 and DRD2 TaqIA in 53 autistic patients and 30 healthy individuals The study also analyzed sequence variations of miR-431 and miR-21 MAOAuVNTR was genotyped by PCR MAOB and DRD2 polymorphisms were analyzed by PCR-based RFLP and miR-431 and miR-21were sequenced Low expressing allele ofMAOA uVNTRwas frequently higher in female patients compared to that in controls (OR= 225)MAOB G allele frequency was more significantly increased in autistic patients than in controls (119875 lt 0001 for both malesand females) DRD2 A1+ genotype increased autism risk (OR = 51) Severity of autism tends to be slightly affected by MAOABgenotype PlasmaMAOB activity was significantly reduced in G than in A allele carrying malesThere was no significant differencein patients and maternal plasmaMAOAB activity compared to controls Neither mutations nor SNPs inmiR-431 andmiR-21 werefound among studied patients This study threw light on some neurotransmitter-related genes suggesting their potential role inAutism pathogenesis that warrants further studies and much consideration
1 Introduction
Autism is a neurodevelopmental disorder characterized bydisturbances in social interactions language and communi-cation as well as by the presence of stereotyped behaviors andrestricted interests The population prevalence of autism isapproximately 15ndash20 in 10000 [1] The assessment of candi-date genes for the identification of susceptibility loci in autismis a common molecular strategy Autism susceptibility locihave been identified on almost every chromosome but nosingle chromosomal location however has been found to behighly significant [2]
The current study investigated genetic variations of twoclasses of genes neurotransmission related genes (MAOABand DRD2) and miRNAs (miR-431 and miR-21) MAOABwere chosen due to their role in enzymatic degradationMAO
has been considered a candidate gene for autism susceptibilitybased not only on its function but also on its location on theX chromosome this is due to the skewed sex ratio in autism(4 males 1 female) [3] An association between patientsand maternal MAOA uVNTR and more severe symptomsof autism spectrum disorders (ASD) was reported [4 5]DRD2 gene is related to receptors in postsynaptic componentFunctional polymorphisms which affect receptor availabilityeither post- or presynaptically may contribute to the impair-ments found in individuals with autism [6] The role of theDRD2 gene in autism susceptibility was suggested by the factthat antipsychotic medications which prevent dopamine D2receptor activation improve the core symptoms of ASDs [7]
Monoamine oxidase A (MAOA) gene contains a 30 bpvariable number of tandem repeats (VNTR) in the promoter(12 kb upstream) region termed uVNTR MAOA Although
2 The Scientific World Journal
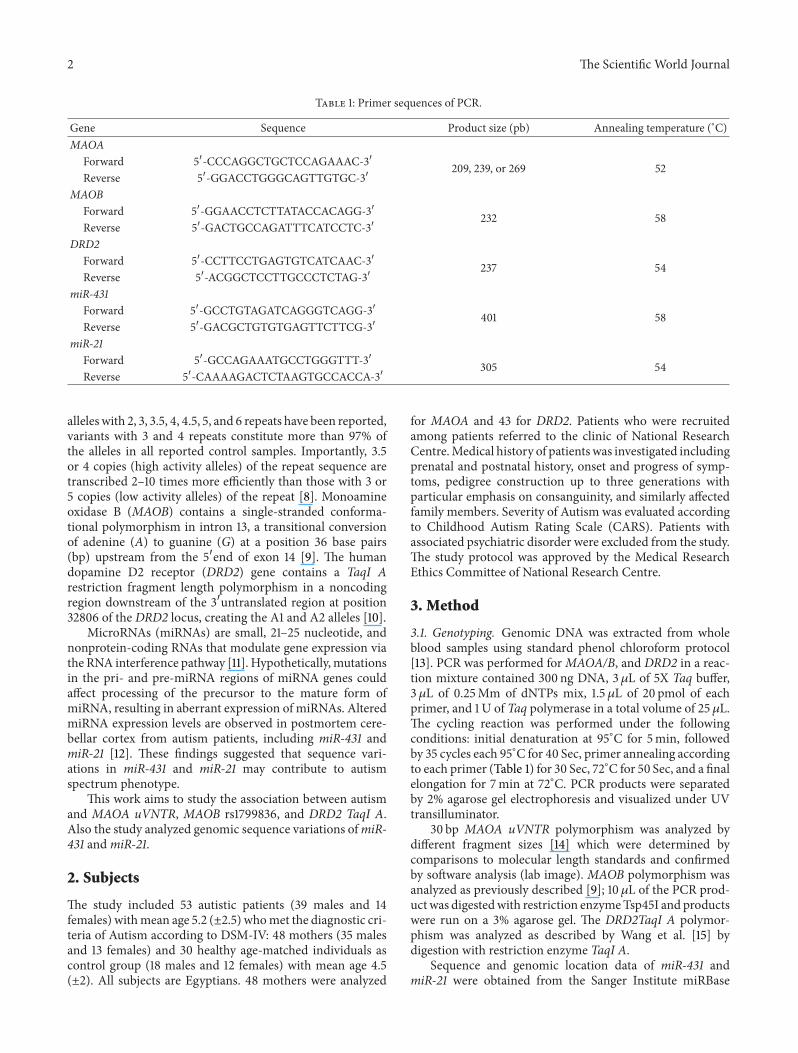
Table 1 Primer sequences of PCR
Gene Sequence Product size (pb) Annealing temperature (∘C)MAOA
Forward 51015840-CCCAGGCTGCTCCAGAAAC-31015840 209 239 or 269 52Reverse 51015840-GGACCTGGGCAGTTGTGC-31015840
MAOBForward 51015840-GGAACCTCTTATACCACAGG-31015840 232 58Reverse 51015840-GACTGCCAGATTTCATCCTC-31015840
DRD2Forward 51015840-CCTTCCTGAGTGTCATCAAC-31015840 237 54Reverse 51015840-ACGGCTCCTTGCCCTCTAG-31015840
miR-431Forward 51015840-GCCTGTAGATCAGGGTCAGG-31015840 401 58Reverse 51015840-GACGCTGTGTGAGTTCTTCG-31015840
miR-21Forward 51015840-GCCAGAAATGCCTGGGTTT-31015840 305 54Reverse 51015840-CAAAAGACTCTAAGTGCCACCA-31015840
alleles with 2 3 35 4 45 5 and 6 repeats have been reportedvariants with 3 and 4 repeats constitute more than 97 ofthe alleles in all reported control samples Importantly 35or 4 copies (high activity alleles) of the repeat sequence aretranscribed 2ndash10 times more efficiently than those with 3 or5 copies (low activity alleles) of the repeat [8] Monoamineoxidase B (MAOB) contains a single-stranded conforma-tional polymorphism in intron 13 a transitional conversionof adenine (A) to guanine (G) at a position 36 base pairs(bp) upstream from the 51015840end of exon 14 [9] The humandopamine D2 receptor (DRD2) gene contains a TaqI Arestriction fragment length polymorphism in a noncodingregion downstream of the 31015840untranslated region at position32806 of the DRD2 locus creating the A1 and A2 alleles [10]
MicroRNAs (miRNAs) are small 21ndash25 nucleotide andnonprotein-coding RNAs that modulate gene expression viathe RNA interference pathway [11] Hypothetically mutationsin the pri- and pre-miRNA regions of miRNA genes couldaffect processing of the precursor to the mature form ofmiRNA resulting in aberrant expression of miRNAs AlteredmiRNA expression levels are observed in postmortem cere-bellar cortex from autism patients including miR-431 andmiR-21 [12] These findings suggested that sequence vari-ations in miR-431 and miR-21 may contribute to autismspectrum phenotype
This work aims to study the association between autismand MAOA uVNTR MAOB rs1799836 and DRD2 TaqI AAlso the study analyzed genomic sequence variations ofmiR-431 andmiR-21
2 Subjects
The study included 53 autistic patients (39 males and 14females) withmean age 52 (plusmn25) whomet the diagnostic cri-teria of Autism according to DSM-IV 48 mothers (35 malesand 13 females) and 30 healthy age-matched individuals ascontrol group (18 males and 12 females) with mean age 45(plusmn2) All subjects are Egyptians 48 mothers were analyzed
for MAOA and 43 for DRD2 Patients who were recruitedamong patients referred to the clinic of National ResearchCentreMedical history of patients was investigated includingprenatal and postnatal history onset and progress of symp-toms pedigree construction up to three generations withparticular emphasis on consanguinity and similarly affectedfamily members Severity of Autism was evaluated accordingto Childhood Autism Rating Scale (CARS) Patients withassociated psychiatric disorder were excluded from the studyThe study protocol was approved by the Medical ResearchEthics Committee of National Research Centre
3 Method
31 Genotyping Genomic DNA was extracted from wholeblood samples using standard phenol chloroform protocol[13] PCR was performed forMAOAB and DRD2 in a reac-tion mixture contained 300 ng DNA 3 120583L of 5X Taq buffer3 120583L of 025Mm of dNTPs mix 15 120583L of 20 pmol of eachprimer and 1U of Taq polymerase in a total volume of 25 120583LThe cycling reaction was performed under the followingconditions initial denaturation at 95∘C for 5min followedby 35 cycles each 95∘C for 40 Sec primer annealing accordingto each primer (Table 1) for 30 Sec 72∘C for 50 Sec and a finalelongation for 7min at 72∘C PCR products were separatedby 2 agarose gel electrophoresis and visualized under UVtransilluminator
30 bp MAOA uVNTR polymorphism was analyzed bydifferent fragment sizes [14] which were determined bycomparisons to molecular length standards and confirmedby software analysis (lab image) MAOB polymorphism wasanalyzed as previously described [9] 10 120583L of the PCR prod-uctwas digestedwith restriction enzymeTsp45I and productswere run on a 3 agarose gel The DRD2TaqI A polymor-phism was analyzed as described by Wang et al [15] bydigestion with restriction enzyme TaqI A
Sequence and genomic location data of miR-431 andmiR-21 were obtained from the Sanger Institute miRBase
The Scientific World Journal 3
M 1 2 3
234194
271 + 181
(a)
4M 321 5
232bp146bp
(b)
4M 321
118
237
126111
234194
281 + 271
(c)
Figure 1 Genotyping of MAOAB and DRD2 (a) Different VNTRs of MAOA M indicates DNA marker (120593x174) Lane 1 indicate 5 tandemrepeats at 269 bp Lane 2 indicates 3 and 5 tandem repeats at 209 and 269 bp Lane 3 indicates 4 tandem repeats at 239 (b) PCR-based RFLPofMAOB M is DNAmarker (120593x174) Lanes 1 and 2 indicate AG genotype at 232 and 146 bp (86 bp not shown) Lane 3 indicates AA genotypeat 146 bp Lanes 4 and 5 indicate GG genotype at 232 bp (c) PCR-based RFLP ofDRD2 Lanes 1 and 4 indicate A1A2 genotype at 237 126 and111 bp Lane 2 indicates A2A2 genotype at 126 and 111 Lane 3 indicates A1A1 genotype at 237 bp
(httpmicrornasangeracuksequencesindexshtml) andthe flanking regions of miRNA genes were obtained by BLATanalysis (httpgenomeucsceducgi-binhgBlatcommand=start) Primers used were designed using primer3 (httpbio-toolsumassmededubioappsprimer3 wwwcgi) PCR wasperformed and then sequencing was carried out to screenSNPs or mutations
32 Plasma MAO Activity MAO AB activity in plasma wasmeasured byMAO-Glo assay from promega according to themanufacturer
33 Statistical Analysis Since MAOA and MAOB are Xlinked data were analyzed separately for each gender Mater-nal genotyping was performed only for MAOA and DRD2SNPstats online software was used to analyze genotype andallele frequencies odd ratio (OR) and Hardy Weinbergequilibrium MAOA uVNTR alleles were classified as lowexpressing allele (3 and 5 repeats) and high expressing allele(4 repeats) Autism severity was assessed by CARS as mildmoderate and sever SPSS statistical software used 119905-test tocompare plasmaMAOA andMAOB activity between autisticpatients and controls One way ANOVA test was used tocompare the enzyme activity between different genotypesMales are hemizygous for MAOAB while females may behomo- or heterozygous so genotype frequencywas calculatedonly for females and enzyme activity was analysed for eachgenotype
4 Results
Male-to-female ratio in our study was sim3 1 males (736males and 264 females) Parental consanguinity repre-sented 34 and about 19 of patients had similarly affectedfamily members No significant association was foundbetween severity of Autism and either gender or consan-guinity (119875 = 0622 and 0248 resp) Significant associationwas detected between severity and the presence of similaraffected familymembers as 14mild cases (100) had negativefamily history of the disease (119875 = 0035)
41 MAOA uVNTR The 30 bp-repeat polymorphismshowed three alleles 3 repeats 209 bp 4-repeat 239 bp and5-repeat 269 bp (Figure 1) MAOA uVNTR was classified inmales as low expressing allele (3 repeats) or high expressingallele (4 repeats) the low and high alleles are equallydistributed between male cases and controls (OR = 1)Female genotypes were classified into lowlow (33 35)highhigh (44) and lowhigh (34) distributed as follows214 357 and 429 respectivelyThe lowhigh genotypewas frequently higher in female cases than in controls (OR =375 and 119875 = 014) Low expressing allele was presented inhigh frequency in female cases than in controls (OR = 225119875 = 017) (Table 2)
Maternal low expressing allele was presented in highfrequency than controls (OR = 129 and 159 for male andfemale mothers resp) (Table 2) The genotype lowhigh was
4 The Scientific World Journal
Table 2 Allele frequenciesMAOA uVNTR MAOB rs1799836 and DRD2 Taq1A
Males FemalesCases 119899 () Controls 119899 () OR (95 CI) 119875 value Cases 119899 () Controls 119899 () OR (95 CI) 119875 value
MAOA uVNTRLow expressing allele 13 (333) 6 (333) 1 (031ndash327) mdash 12 (43) 6 (25) 225 (069ndash739) 017High expressing allele 26 (667) 12 (66 7) 16 (57) 18 (75) 0044 (014ndash146)
MaternalMAOA uVNTRLow expressing allele 21 (30) 6 (25) 129 (045ndash37) 064 9 (35) 6 (25) 159 (047ndash542) 046High expressing allele 49 (70) 18 (75) 078 (027ndash224) 17 (65) 18 (75) 063 (018ndash215)
MAOB rs1799836G allele 26 (667) 1 (56) 34 (406ndash284347) lt0001 20 (71) 4 (17) 125 (324ndash4826) lt0001A allele 13 (333) 17 (944) 003 (000ndash025) 8 (29) 20 (83) 008 (002ndash031)
Patients MothersCases 119899 () Controls 119899 () OR (95 CI) P value Cases 119899 () Controls 119899 () OR (95 CI) P value
DRD2 Taq1AA1 allele 64 (60) 29 (48) 163 (086ndash309) 013 25 (29) 29 (48) 044 (022ndash087) 0018A2 allele 42 (40) 31 (52) 061 (032ndash116) 61 (71) 31 (52) 228 (115ndash454)119875 lt 005 is significant OR odd ratio 95 CI 95 confidence interval
Table 3 Relation betweenMAOAB polymorphisms and severity of autistic patients
Males FemalesSevere 119899 () Mild and moderate 119899 () OR 119875 value Severe 119899 () Mild and moderate 119899 () OR 119875 value
MAOA uVNTRLow expressing allele 7 (438) 6 (261) 224 025 5 (50) 7 (39) 157 057High expressing allele 9 (563) 17 (739) 045 5 (50) 11 (61) 064
MAOB rs1799836G allele 11 (688) 15 (562) 1173 082 8 (80) 12 (667) 200 045A allele 5 (312) 8 (348 085 2 (20) 6 (333) 050
more common in mothers than controls (OR = 295 and 58formale and femalemothers resp)The genotype frequenciesofMAOA uVNTR for females mothers and controls were allin Hardy-Weinberg equilibrium (119875 = 062 1 and 009 resp)Patients carrying low expressing allele tend to be severelyautistic rather than mild to moderate (OR = 224 and 157 formale and female patients resp) (Table 3)
42 MAOB Polymorphism Allele A showed two bands of146 and 86 bp (86 bp band not shown) while allele G wasdetected as uncleaved 232 bp band (Figure 1) G allele wassignificantly higher in cases than controls (OR = 34 and125 for male and female cases resp 119875 lt 0001) (Table 2)AA genotype was absent in female cases GG genotype wasabsent in controls and represents 429 of females casesAG genotype was frequently higher in female cases than incontrols (OR = 267 119875 = 022) Genotypes distribution infemale cases and controls were in accordance with HardyWeinberg equilibrium (119875 = 051 and 1 resp) Cases withG allele slightly tend to be severely autistic rather than mildto moderate (OR = 1173 200 for males and females resp)(Table 3)
43 DRD2TaqI A Polymorphism A1 allele was detected bythe presence of uncleaved 237 bp band while A2 allele was
detected by cleavage of 237 bp band into 111 and 126 pb frag-ments (Figure 1) A1 allele was presented in higher frequencyin autistic patients than in controls (OR = 163 119875 = 013)Unlike cases A2 wasmore common inmothers than controls(OR = 228 119875 = 0018) (Table 2) By classifying the casesinto A1+ (A1A1 A1A2) and A1minus (A2A2) the A1+ genotypewas significantly higher in cases than in controls (OR = 51119875 = 004) A1A1 genotype was more common in cases thancontrols (OR = 211 119875 = 021) while A2A2 was significantlyincreased in mothers (OR = 4348 119875 = 0007) Mothersand controls genotypes were in accordance with HardyWeinberg equilibrium (119875 = 046 and 0065 resp) in contrastcases genotype was deviated from Hardy Weinberg equilib-rium (119875 = 00004)
44 Plasma MAO Activity in Autistic Patients No differenceof MAOAB plasma activity in autistic patients and theirmothers compared to controls (119875 = 0927 0958 04 and 07for males MAOA MAOB maternal MAOA and maternalMAOB respectively 119875 = 0439 0449 0907 and 06 forfemales MAOA MAOB maternal MAOA and maternalMAOB resp) MAOA activity did not differ between maleslow and high expressing alleles (119875 = 096) nor betweenfemales genotypes (119875 = 0408) MAOB activity was sig-nificantly reduced in G than in A allele carrying males
The Scientific World Journal 5
(a) (b)
180 190 230 240
(e)
(c) (d)
T
T T
T T T T T T TC C C C C C190 200A A A A
A A A A
A A A AAG
G G G G G G G G G G G G G
C A A A A A A AT T T T T T
TTTTTTTTTT
80 90 100 310 320G G G G GG G G G G G G G G G G G G GGC C C C C C
CCCC CCCCCC
CC
GndashA SNP is absent CndashT and GndashTC are absent
GndashA SNP is absentCndashT SNP is absent
AndashG mutation is absent
Figure 2 Sequencing chromatogram ofmiR-21 andmiR-431 (a) it shows the absence of AndashGmutation downstream the pre-miR-21 ((b) (c)(d) and (e)) they show the absence of rs12883709 GndashA upstream the pre-miR-431 rs76090066 CndashT and rs12884005 GndashA in the sequence ofpre-miR-431 rs61993318 CndashT and rs35695758 GndashTC downstream the pre-miR-431
(119875 = 0027) while females showed no difference of MAOBactivity between AG and GG genotypes (119875 = 082)
45 miR-431 and miR-21 Variations rs12883709 GAupstream the pre-miR-431 rs12884005 GA and rs76090066CT in the sequence of the pre-miR-431 rs 61993318 CTand rs35695758 GTC downstream the pre-miR-431 wereall absent Also AndashG mutation at 29-nt downstream ofpre-miR-21 was absent No variations were observed in theamplified sequences (Figure 2)
5 Discussion
Twin and family studies provide indisputable evidence fora genetic component in autism The average concordancefor identical twins versus fraternal twins was frequentlystudied and reported to be 36 versus 3 [16] 64 versus9 [17] and 60 versus 0 [18] However a concordancerate for monozygotic twins of less than 100 indicates thatnongenetic factors also play a causal role The current studyshowed that about 19 of patients had similar affected familymembers Elevated rates of clinical psychiatric disordersdistinct from autism have been reported among the relativesof individuals with autism including schizophrenia anxietydepression and social phobias [19] Parental consanguinityrepresented 34 of cases in our study it is higher than thatof Saltık and Basgul (214) [20] However Datta et al [21]found that possible parental consanguinity increases thelikelihood of autism and behavioral disturbances manyreports did not find a link between consanguinity and autismin Egyptian [22] Saudi [23] and Iranian population [24]
Deletion of MAOAB was associated with severe mentalretardation and unusual stereotypical behaviors of hand
wringing and lip smacking in males [25] Bortolato et al[26] found that both MAOAB knockout mice displayedneuropathological alterations reminiscent of typical ASDfeatures Previous study found consistent association betweenthe ldquolow activityrdquo allele of MAOA and larger brain volumesfor regions of the cortex in children with autism but notin controls In contrast the data did not find association ofthe MAOA promoter polymorphism with autism itself [3]Tassone et al [27] suggested that functionalMAOA promoteralleles play a potential role in the male child the motheror both in ASD In this study no difference in allele fre-quency ofMAOA betweenmale autistic patients and controlsHowever low expressing allele and lowhigh genotype weremore common in female cases and mothers Severity ofautism slightly tends to increase in low expressing allele forboth cases and mothers Previous results found that autisticboys with the low expressing 3-repeat MAOA allele hadmore severe sensory behaviors arousal regulation problemsaggression and worse social communication skills thanmales with the high activity allele and that problems withaggression as well as with fears and rituals were modifiedby the mothersrsquo genotype [5]
The AG (A644G) noncoding SNP (rs1799836) ofMAOBis responsible for altered enzyme activity with tissue speci-ficity [28 29] This MAOB SNP was found to be associatedwith emotional regulation [30 31] and Parkinsonrsquos disease[32] Also it was implicated as risk factor for schizophrenia ina Spanish population [33] and in Han Chinese [34] So far noavailable information was found regarding relation ofMAOBpolymorphism and autism however this study highlightedthe importance of G allele in both male and female autisticpatients G allele was significantly higher in autistic casesthan in controls Interestingly GG genotype was absent in
6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009
2 The Scientific World Journal
Table 1 Primer sequences of PCR
Gene Sequence Product size (pb) Annealing temperature (∘C)MAOA
Forward 51015840-CCCAGGCTGCTCCAGAAAC-31015840 209 239 or 269 52Reverse 51015840-GGACCTGGGCAGTTGTGC-31015840
MAOBForward 51015840-GGAACCTCTTATACCACAGG-31015840 232 58Reverse 51015840-GACTGCCAGATTTCATCCTC-31015840
DRD2Forward 51015840-CCTTCCTGAGTGTCATCAAC-31015840 237 54Reverse 51015840-ACGGCTCCTTGCCCTCTAG-31015840
miR-431Forward 51015840-GCCTGTAGATCAGGGTCAGG-31015840 401 58Reverse 51015840-GACGCTGTGTGAGTTCTTCG-31015840
miR-21Forward 51015840-GCCAGAAATGCCTGGGTTT-31015840 305 54Reverse 51015840-CAAAAGACTCTAAGTGCCACCA-31015840
alleles with 2 3 35 4 45 5 and 6 repeats have been reportedvariants with 3 and 4 repeats constitute more than 97 ofthe alleles in all reported control samples Importantly 35or 4 copies (high activity alleles) of the repeat sequence aretranscribed 2ndash10 times more efficiently than those with 3 or5 copies (low activity alleles) of the repeat [8] Monoamineoxidase B (MAOB) contains a single-stranded conforma-tional polymorphism in intron 13 a transitional conversionof adenine (A) to guanine (G) at a position 36 base pairs(bp) upstream from the 51015840end of exon 14 [9] The humandopamine D2 receptor (DRD2) gene contains a TaqI Arestriction fragment length polymorphism in a noncodingregion downstream of the 31015840untranslated region at position32806 of the DRD2 locus creating the A1 and A2 alleles [10]
MicroRNAs (miRNAs) are small 21ndash25 nucleotide andnonprotein-coding RNAs that modulate gene expression viathe RNA interference pathway [11] Hypothetically mutationsin the pri- and pre-miRNA regions of miRNA genes couldaffect processing of the precursor to the mature form ofmiRNA resulting in aberrant expression of miRNAs AlteredmiRNA expression levels are observed in postmortem cere-bellar cortex from autism patients including miR-431 andmiR-21 [12] These findings suggested that sequence vari-ations in miR-431 and miR-21 may contribute to autismspectrum phenotype
This work aims to study the association between autismand MAOA uVNTR MAOB rs1799836 and DRD2 TaqI AAlso the study analyzed genomic sequence variations ofmiR-431 andmiR-21
2 Subjects
The study included 53 autistic patients (39 males and 14females) withmean age 52 (plusmn25) whomet the diagnostic cri-teria of Autism according to DSM-IV 48 mothers (35 malesand 13 females) and 30 healthy age-matched individuals ascontrol group (18 males and 12 females) with mean age 45(plusmn2) All subjects are Egyptians 48 mothers were analyzed
for MAOA and 43 for DRD2 Patients who were recruitedamong patients referred to the clinic of National ResearchCentreMedical history of patients was investigated includingprenatal and postnatal history onset and progress of symp-toms pedigree construction up to three generations withparticular emphasis on consanguinity and similarly affectedfamily members Severity of Autism was evaluated accordingto Childhood Autism Rating Scale (CARS) Patients withassociated psychiatric disorder were excluded from the studyThe study protocol was approved by the Medical ResearchEthics Committee of National Research Centre
3 Method
31 Genotyping Genomic DNA was extracted from wholeblood samples using standard phenol chloroform protocol[13] PCR was performed forMAOAB and DRD2 in a reac-tion mixture contained 300 ng DNA 3 120583L of 5X Taq buffer3 120583L of 025Mm of dNTPs mix 15 120583L of 20 pmol of eachprimer and 1U of Taq polymerase in a total volume of 25 120583LThe cycling reaction was performed under the followingconditions initial denaturation at 95∘C for 5min followedby 35 cycles each 95∘C for 40 Sec primer annealing accordingto each primer (Table 1) for 30 Sec 72∘C for 50 Sec and a finalelongation for 7min at 72∘C PCR products were separatedby 2 agarose gel electrophoresis and visualized under UVtransilluminator
30 bp MAOA uVNTR polymorphism was analyzed bydifferent fragment sizes [14] which were determined bycomparisons to molecular length standards and confirmedby software analysis (lab image) MAOB polymorphism wasanalyzed as previously described [9] 10 120583L of the PCR prod-uctwas digestedwith restriction enzymeTsp45I and productswere run on a 3 agarose gel The DRD2TaqI A polymor-phism was analyzed as described by Wang et al [15] bydigestion with restriction enzyme TaqI A
Sequence and genomic location data of miR-431 andmiR-21 were obtained from the Sanger Institute miRBase
The Scientific World Journal 3
M 1 2 3
234194
271 + 181
(a)
4M 321 5
232bp146bp
(b)
4M 321
118
237
126111
234194
281 + 271
(c)
Figure 1 Genotyping of MAOAB and DRD2 (a) Different VNTRs of MAOA M indicates DNA marker (120593x174) Lane 1 indicate 5 tandemrepeats at 269 bp Lane 2 indicates 3 and 5 tandem repeats at 209 and 269 bp Lane 3 indicates 4 tandem repeats at 239 (b) PCR-based RFLPofMAOB M is DNAmarker (120593x174) Lanes 1 and 2 indicate AG genotype at 232 and 146 bp (86 bp not shown) Lane 3 indicates AA genotypeat 146 bp Lanes 4 and 5 indicate GG genotype at 232 bp (c) PCR-based RFLP ofDRD2 Lanes 1 and 4 indicate A1A2 genotype at 237 126 and111 bp Lane 2 indicates A2A2 genotype at 126 and 111 Lane 3 indicates A1A1 genotype at 237 bp
(httpmicrornasangeracuksequencesindexshtml) andthe flanking regions of miRNA genes were obtained by BLATanalysis (httpgenomeucsceducgi-binhgBlatcommand=start) Primers used were designed using primer3 (httpbio-toolsumassmededubioappsprimer3 wwwcgi) PCR wasperformed and then sequencing was carried out to screenSNPs or mutations
32 Plasma MAO Activity MAO AB activity in plasma wasmeasured byMAO-Glo assay from promega according to themanufacturer
33 Statistical Analysis Since MAOA and MAOB are Xlinked data were analyzed separately for each gender Mater-nal genotyping was performed only for MAOA and DRD2SNPstats online software was used to analyze genotype andallele frequencies odd ratio (OR) and Hardy Weinbergequilibrium MAOA uVNTR alleles were classified as lowexpressing allele (3 and 5 repeats) and high expressing allele(4 repeats) Autism severity was assessed by CARS as mildmoderate and sever SPSS statistical software used 119905-test tocompare plasmaMAOA andMAOB activity between autisticpatients and controls One way ANOVA test was used tocompare the enzyme activity between different genotypesMales are hemizygous for MAOAB while females may behomo- or heterozygous so genotype frequencywas calculatedonly for females and enzyme activity was analysed for eachgenotype
4 Results
Male-to-female ratio in our study was sim3 1 males (736males and 264 females) Parental consanguinity repre-sented 34 and about 19 of patients had similarly affectedfamily members No significant association was foundbetween severity of Autism and either gender or consan-guinity (119875 = 0622 and 0248 resp) Significant associationwas detected between severity and the presence of similaraffected familymembers as 14mild cases (100) had negativefamily history of the disease (119875 = 0035)
41 MAOA uVNTR The 30 bp-repeat polymorphismshowed three alleles 3 repeats 209 bp 4-repeat 239 bp and5-repeat 269 bp (Figure 1) MAOA uVNTR was classified inmales as low expressing allele (3 repeats) or high expressingallele (4 repeats) the low and high alleles are equallydistributed between male cases and controls (OR = 1)Female genotypes were classified into lowlow (33 35)highhigh (44) and lowhigh (34) distributed as follows214 357 and 429 respectivelyThe lowhigh genotypewas frequently higher in female cases than in controls (OR =375 and 119875 = 014) Low expressing allele was presented inhigh frequency in female cases than in controls (OR = 225119875 = 017) (Table 2)
Maternal low expressing allele was presented in highfrequency than controls (OR = 129 and 159 for male andfemale mothers resp) (Table 2) The genotype lowhigh was
4 The Scientific World Journal
Table 2 Allele frequenciesMAOA uVNTR MAOB rs1799836 and DRD2 Taq1A
Males FemalesCases 119899 () Controls 119899 () OR (95 CI) 119875 value Cases 119899 () Controls 119899 () OR (95 CI) 119875 value
MAOA uVNTRLow expressing allele 13 (333) 6 (333) 1 (031ndash327) mdash 12 (43) 6 (25) 225 (069ndash739) 017High expressing allele 26 (667) 12 (66 7) 16 (57) 18 (75) 0044 (014ndash146)
MaternalMAOA uVNTRLow expressing allele 21 (30) 6 (25) 129 (045ndash37) 064 9 (35) 6 (25) 159 (047ndash542) 046High expressing allele 49 (70) 18 (75) 078 (027ndash224) 17 (65) 18 (75) 063 (018ndash215)
MAOB rs1799836G allele 26 (667) 1 (56) 34 (406ndash284347) lt0001 20 (71) 4 (17) 125 (324ndash4826) lt0001A allele 13 (333) 17 (944) 003 (000ndash025) 8 (29) 20 (83) 008 (002ndash031)
Patients MothersCases 119899 () Controls 119899 () OR (95 CI) P value Cases 119899 () Controls 119899 () OR (95 CI) P value
DRD2 Taq1AA1 allele 64 (60) 29 (48) 163 (086ndash309) 013 25 (29) 29 (48) 044 (022ndash087) 0018A2 allele 42 (40) 31 (52) 061 (032ndash116) 61 (71) 31 (52) 228 (115ndash454)119875 lt 005 is significant OR odd ratio 95 CI 95 confidence interval
Table 3 Relation betweenMAOAB polymorphisms and severity of autistic patients
Males FemalesSevere 119899 () Mild and moderate 119899 () OR 119875 value Severe 119899 () Mild and moderate 119899 () OR 119875 value
MAOA uVNTRLow expressing allele 7 (438) 6 (261) 224 025 5 (50) 7 (39) 157 057High expressing allele 9 (563) 17 (739) 045 5 (50) 11 (61) 064
MAOB rs1799836G allele 11 (688) 15 (562) 1173 082 8 (80) 12 (667) 200 045A allele 5 (312) 8 (348 085 2 (20) 6 (333) 050
more common in mothers than controls (OR = 295 and 58formale and femalemothers resp)The genotype frequenciesofMAOA uVNTR for females mothers and controls were allin Hardy-Weinberg equilibrium (119875 = 062 1 and 009 resp)Patients carrying low expressing allele tend to be severelyautistic rather than mild to moderate (OR = 224 and 157 formale and female patients resp) (Table 3)
42 MAOB Polymorphism Allele A showed two bands of146 and 86 bp (86 bp band not shown) while allele G wasdetected as uncleaved 232 bp band (Figure 1) G allele wassignificantly higher in cases than controls (OR = 34 and125 for male and female cases resp 119875 lt 0001) (Table 2)AA genotype was absent in female cases GG genotype wasabsent in controls and represents 429 of females casesAG genotype was frequently higher in female cases than incontrols (OR = 267 119875 = 022) Genotypes distribution infemale cases and controls were in accordance with HardyWeinberg equilibrium (119875 = 051 and 1 resp) Cases withG allele slightly tend to be severely autistic rather than mildto moderate (OR = 1173 200 for males and females resp)(Table 3)
43 DRD2TaqI A Polymorphism A1 allele was detected bythe presence of uncleaved 237 bp band while A2 allele was
detected by cleavage of 237 bp band into 111 and 126 pb frag-ments (Figure 1) A1 allele was presented in higher frequencyin autistic patients than in controls (OR = 163 119875 = 013)Unlike cases A2 wasmore common inmothers than controls(OR = 228 119875 = 0018) (Table 2) By classifying the casesinto A1+ (A1A1 A1A2) and A1minus (A2A2) the A1+ genotypewas significantly higher in cases than in controls (OR = 51119875 = 004) A1A1 genotype was more common in cases thancontrols (OR = 211 119875 = 021) while A2A2 was significantlyincreased in mothers (OR = 4348 119875 = 0007) Mothersand controls genotypes were in accordance with HardyWeinberg equilibrium (119875 = 046 and 0065 resp) in contrastcases genotype was deviated from Hardy Weinberg equilib-rium (119875 = 00004)
44 Plasma MAO Activity in Autistic Patients No differenceof MAOAB plasma activity in autistic patients and theirmothers compared to controls (119875 = 0927 0958 04 and 07for males MAOA MAOB maternal MAOA and maternalMAOB respectively 119875 = 0439 0449 0907 and 06 forfemales MAOA MAOB maternal MAOA and maternalMAOB resp) MAOA activity did not differ between maleslow and high expressing alleles (119875 = 096) nor betweenfemales genotypes (119875 = 0408) MAOB activity was sig-nificantly reduced in G than in A allele carrying males
The Scientific World Journal 5
(a) (b)
180 190 230 240
(e)
(c) (d)
T
T T
T T T T T T TC C C C C C190 200A A A A
A A A A
A A A AAG
G G G G G G G G G G G G G
C A A A A A A AT T T T T T
TTTTTTTTTT
80 90 100 310 320G G G G GG G G G G G G G G G G G G GGC C C C C C
CCCC CCCCCC
CC
GndashA SNP is absent CndashT and GndashTC are absent
GndashA SNP is absentCndashT SNP is absent
AndashG mutation is absent
Figure 2 Sequencing chromatogram ofmiR-21 andmiR-431 (a) it shows the absence of AndashGmutation downstream the pre-miR-21 ((b) (c)(d) and (e)) they show the absence of rs12883709 GndashA upstream the pre-miR-431 rs76090066 CndashT and rs12884005 GndashA in the sequence ofpre-miR-431 rs61993318 CndashT and rs35695758 GndashTC downstream the pre-miR-431
(119875 = 0027) while females showed no difference of MAOBactivity between AG and GG genotypes (119875 = 082)
45 miR-431 and miR-21 Variations rs12883709 GAupstream the pre-miR-431 rs12884005 GA and rs76090066CT in the sequence of the pre-miR-431 rs 61993318 CTand rs35695758 GTC downstream the pre-miR-431 wereall absent Also AndashG mutation at 29-nt downstream ofpre-miR-21 was absent No variations were observed in theamplified sequences (Figure 2)
5 Discussion
Twin and family studies provide indisputable evidence fora genetic component in autism The average concordancefor identical twins versus fraternal twins was frequentlystudied and reported to be 36 versus 3 [16] 64 versus9 [17] and 60 versus 0 [18] However a concordancerate for monozygotic twins of less than 100 indicates thatnongenetic factors also play a causal role The current studyshowed that about 19 of patients had similar affected familymembers Elevated rates of clinical psychiatric disordersdistinct from autism have been reported among the relativesof individuals with autism including schizophrenia anxietydepression and social phobias [19] Parental consanguinityrepresented 34 of cases in our study it is higher than thatof Saltık and Basgul (214) [20] However Datta et al [21]found that possible parental consanguinity increases thelikelihood of autism and behavioral disturbances manyreports did not find a link between consanguinity and autismin Egyptian [22] Saudi [23] and Iranian population [24]
Deletion of MAOAB was associated with severe mentalretardation and unusual stereotypical behaviors of hand
wringing and lip smacking in males [25] Bortolato et al[26] found that both MAOAB knockout mice displayedneuropathological alterations reminiscent of typical ASDfeatures Previous study found consistent association betweenthe ldquolow activityrdquo allele of MAOA and larger brain volumesfor regions of the cortex in children with autism but notin controls In contrast the data did not find association ofthe MAOA promoter polymorphism with autism itself [3]Tassone et al [27] suggested that functionalMAOA promoteralleles play a potential role in the male child the motheror both in ASD In this study no difference in allele fre-quency ofMAOA betweenmale autistic patients and controlsHowever low expressing allele and lowhigh genotype weremore common in female cases and mothers Severity ofautism slightly tends to increase in low expressing allele forboth cases and mothers Previous results found that autisticboys with the low expressing 3-repeat MAOA allele hadmore severe sensory behaviors arousal regulation problemsaggression and worse social communication skills thanmales with the high activity allele and that problems withaggression as well as with fears and rituals were modifiedby the mothersrsquo genotype [5]
The AG (A644G) noncoding SNP (rs1799836) ofMAOBis responsible for altered enzyme activity with tissue speci-ficity [28 29] This MAOB SNP was found to be associatedwith emotional regulation [30 31] and Parkinsonrsquos disease[32] Also it was implicated as risk factor for schizophrenia ina Spanish population [33] and in Han Chinese [34] So far noavailable information was found regarding relation ofMAOBpolymorphism and autism however this study highlightedthe importance of G allele in both male and female autisticpatients G allele was significantly higher in autistic casesthan in controls Interestingly GG genotype was absent in
6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009

The Scientific World Journal 3
M 1 2 3
234194
271 + 181
(a)
4M 321 5
232bp146bp
(b)
4M 321
118
237
126111
234194
281 + 271
(c)
Figure 1 Genotyping of MAOAB and DRD2 (a) Different VNTRs of MAOA M indicates DNA marker (120593x174) Lane 1 indicate 5 tandemrepeats at 269 bp Lane 2 indicates 3 and 5 tandem repeats at 209 and 269 bp Lane 3 indicates 4 tandem repeats at 239 (b) PCR-based RFLPofMAOB M is DNAmarker (120593x174) Lanes 1 and 2 indicate AG genotype at 232 and 146 bp (86 bp not shown) Lane 3 indicates AA genotypeat 146 bp Lanes 4 and 5 indicate GG genotype at 232 bp (c) PCR-based RFLP ofDRD2 Lanes 1 and 4 indicate A1A2 genotype at 237 126 and111 bp Lane 2 indicates A2A2 genotype at 126 and 111 Lane 3 indicates A1A1 genotype at 237 bp
(httpmicrornasangeracuksequencesindexshtml) andthe flanking regions of miRNA genes were obtained by BLATanalysis (httpgenomeucsceducgi-binhgBlatcommand=start) Primers used were designed using primer3 (httpbio-toolsumassmededubioappsprimer3 wwwcgi) PCR wasperformed and then sequencing was carried out to screenSNPs or mutations
32 Plasma MAO Activity MAO AB activity in plasma wasmeasured byMAO-Glo assay from promega according to themanufacturer
33 Statistical Analysis Since MAOA and MAOB are Xlinked data were analyzed separately for each gender Mater-nal genotyping was performed only for MAOA and DRD2SNPstats online software was used to analyze genotype andallele frequencies odd ratio (OR) and Hardy Weinbergequilibrium MAOA uVNTR alleles were classified as lowexpressing allele (3 and 5 repeats) and high expressing allele(4 repeats) Autism severity was assessed by CARS as mildmoderate and sever SPSS statistical software used 119905-test tocompare plasmaMAOA andMAOB activity between autisticpatients and controls One way ANOVA test was used tocompare the enzyme activity between different genotypesMales are hemizygous for MAOAB while females may behomo- or heterozygous so genotype frequencywas calculatedonly for females and enzyme activity was analysed for eachgenotype
4 Results
Male-to-female ratio in our study was sim3 1 males (736males and 264 females) Parental consanguinity repre-sented 34 and about 19 of patients had similarly affectedfamily members No significant association was foundbetween severity of Autism and either gender or consan-guinity (119875 = 0622 and 0248 resp) Significant associationwas detected between severity and the presence of similaraffected familymembers as 14mild cases (100) had negativefamily history of the disease (119875 = 0035)
41 MAOA uVNTR The 30 bp-repeat polymorphismshowed three alleles 3 repeats 209 bp 4-repeat 239 bp and5-repeat 269 bp (Figure 1) MAOA uVNTR was classified inmales as low expressing allele (3 repeats) or high expressingallele (4 repeats) the low and high alleles are equallydistributed between male cases and controls (OR = 1)Female genotypes were classified into lowlow (33 35)highhigh (44) and lowhigh (34) distributed as follows214 357 and 429 respectivelyThe lowhigh genotypewas frequently higher in female cases than in controls (OR =375 and 119875 = 014) Low expressing allele was presented inhigh frequency in female cases than in controls (OR = 225119875 = 017) (Table 2)
Maternal low expressing allele was presented in highfrequency than controls (OR = 129 and 159 for male andfemale mothers resp) (Table 2) The genotype lowhigh was
4 The Scientific World Journal
Table 2 Allele frequenciesMAOA uVNTR MAOB rs1799836 and DRD2 Taq1A
Males FemalesCases 119899 () Controls 119899 () OR (95 CI) 119875 value Cases 119899 () Controls 119899 () OR (95 CI) 119875 value
MAOA uVNTRLow expressing allele 13 (333) 6 (333) 1 (031ndash327) mdash 12 (43) 6 (25) 225 (069ndash739) 017High expressing allele 26 (667) 12 (66 7) 16 (57) 18 (75) 0044 (014ndash146)
MaternalMAOA uVNTRLow expressing allele 21 (30) 6 (25) 129 (045ndash37) 064 9 (35) 6 (25) 159 (047ndash542) 046High expressing allele 49 (70) 18 (75) 078 (027ndash224) 17 (65) 18 (75) 063 (018ndash215)
MAOB rs1799836G allele 26 (667) 1 (56) 34 (406ndash284347) lt0001 20 (71) 4 (17) 125 (324ndash4826) lt0001A allele 13 (333) 17 (944) 003 (000ndash025) 8 (29) 20 (83) 008 (002ndash031)
Patients MothersCases 119899 () Controls 119899 () OR (95 CI) P value Cases 119899 () Controls 119899 () OR (95 CI) P value
DRD2 Taq1AA1 allele 64 (60) 29 (48) 163 (086ndash309) 013 25 (29) 29 (48) 044 (022ndash087) 0018A2 allele 42 (40) 31 (52) 061 (032ndash116) 61 (71) 31 (52) 228 (115ndash454)119875 lt 005 is significant OR odd ratio 95 CI 95 confidence interval
Table 3 Relation betweenMAOAB polymorphisms and severity of autistic patients
Males FemalesSevere 119899 () Mild and moderate 119899 () OR 119875 value Severe 119899 () Mild and moderate 119899 () OR 119875 value
MAOA uVNTRLow expressing allele 7 (438) 6 (261) 224 025 5 (50) 7 (39) 157 057High expressing allele 9 (563) 17 (739) 045 5 (50) 11 (61) 064
MAOB rs1799836G allele 11 (688) 15 (562) 1173 082 8 (80) 12 (667) 200 045A allele 5 (312) 8 (348 085 2 (20) 6 (333) 050
more common in mothers than controls (OR = 295 and 58formale and femalemothers resp)The genotype frequenciesofMAOA uVNTR for females mothers and controls were allin Hardy-Weinberg equilibrium (119875 = 062 1 and 009 resp)Patients carrying low expressing allele tend to be severelyautistic rather than mild to moderate (OR = 224 and 157 formale and female patients resp) (Table 3)
42 MAOB Polymorphism Allele A showed two bands of146 and 86 bp (86 bp band not shown) while allele G wasdetected as uncleaved 232 bp band (Figure 1) G allele wassignificantly higher in cases than controls (OR = 34 and125 for male and female cases resp 119875 lt 0001) (Table 2)AA genotype was absent in female cases GG genotype wasabsent in controls and represents 429 of females casesAG genotype was frequently higher in female cases than incontrols (OR = 267 119875 = 022) Genotypes distribution infemale cases and controls were in accordance with HardyWeinberg equilibrium (119875 = 051 and 1 resp) Cases withG allele slightly tend to be severely autistic rather than mildto moderate (OR = 1173 200 for males and females resp)(Table 3)
43 DRD2TaqI A Polymorphism A1 allele was detected bythe presence of uncleaved 237 bp band while A2 allele was
detected by cleavage of 237 bp band into 111 and 126 pb frag-ments (Figure 1) A1 allele was presented in higher frequencyin autistic patients than in controls (OR = 163 119875 = 013)Unlike cases A2 wasmore common inmothers than controls(OR = 228 119875 = 0018) (Table 2) By classifying the casesinto A1+ (A1A1 A1A2) and A1minus (A2A2) the A1+ genotypewas significantly higher in cases than in controls (OR = 51119875 = 004) A1A1 genotype was more common in cases thancontrols (OR = 211 119875 = 021) while A2A2 was significantlyincreased in mothers (OR = 4348 119875 = 0007) Mothersand controls genotypes were in accordance with HardyWeinberg equilibrium (119875 = 046 and 0065 resp) in contrastcases genotype was deviated from Hardy Weinberg equilib-rium (119875 = 00004)
44 Plasma MAO Activity in Autistic Patients No differenceof MAOAB plasma activity in autistic patients and theirmothers compared to controls (119875 = 0927 0958 04 and 07for males MAOA MAOB maternal MAOA and maternalMAOB respectively 119875 = 0439 0449 0907 and 06 forfemales MAOA MAOB maternal MAOA and maternalMAOB resp) MAOA activity did not differ between maleslow and high expressing alleles (119875 = 096) nor betweenfemales genotypes (119875 = 0408) MAOB activity was sig-nificantly reduced in G than in A allele carrying males
The Scientific World Journal 5
(a) (b)
180 190 230 240
(e)
(c) (d)
T
T T
T T T T T T TC C C C C C190 200A A A A
A A A A
A A A AAG
G G G G G G G G G G G G G
C A A A A A A AT T T T T T
TTTTTTTTTT
80 90 100 310 320G G G G GG G G G G G G G G G G G G GGC C C C C C
CCCC CCCCCC
CC
GndashA SNP is absent CndashT and GndashTC are absent
GndashA SNP is absentCndashT SNP is absent
AndashG mutation is absent
Figure 2 Sequencing chromatogram ofmiR-21 andmiR-431 (a) it shows the absence of AndashGmutation downstream the pre-miR-21 ((b) (c)(d) and (e)) they show the absence of rs12883709 GndashA upstream the pre-miR-431 rs76090066 CndashT and rs12884005 GndashA in the sequence ofpre-miR-431 rs61993318 CndashT and rs35695758 GndashTC downstream the pre-miR-431
(119875 = 0027) while females showed no difference of MAOBactivity between AG and GG genotypes (119875 = 082)
45 miR-431 and miR-21 Variations rs12883709 GAupstream the pre-miR-431 rs12884005 GA and rs76090066CT in the sequence of the pre-miR-431 rs 61993318 CTand rs35695758 GTC downstream the pre-miR-431 wereall absent Also AndashG mutation at 29-nt downstream ofpre-miR-21 was absent No variations were observed in theamplified sequences (Figure 2)
5 Discussion
Twin and family studies provide indisputable evidence fora genetic component in autism The average concordancefor identical twins versus fraternal twins was frequentlystudied and reported to be 36 versus 3 [16] 64 versus9 [17] and 60 versus 0 [18] However a concordancerate for monozygotic twins of less than 100 indicates thatnongenetic factors also play a causal role The current studyshowed that about 19 of patients had similar affected familymembers Elevated rates of clinical psychiatric disordersdistinct from autism have been reported among the relativesof individuals with autism including schizophrenia anxietydepression and social phobias [19] Parental consanguinityrepresented 34 of cases in our study it is higher than thatof Saltık and Basgul (214) [20] However Datta et al [21]found that possible parental consanguinity increases thelikelihood of autism and behavioral disturbances manyreports did not find a link between consanguinity and autismin Egyptian [22] Saudi [23] and Iranian population [24]
Deletion of MAOAB was associated with severe mentalretardation and unusual stereotypical behaviors of hand
wringing and lip smacking in males [25] Bortolato et al[26] found that both MAOAB knockout mice displayedneuropathological alterations reminiscent of typical ASDfeatures Previous study found consistent association betweenthe ldquolow activityrdquo allele of MAOA and larger brain volumesfor regions of the cortex in children with autism but notin controls In contrast the data did not find association ofthe MAOA promoter polymorphism with autism itself [3]Tassone et al [27] suggested that functionalMAOA promoteralleles play a potential role in the male child the motheror both in ASD In this study no difference in allele fre-quency ofMAOA betweenmale autistic patients and controlsHowever low expressing allele and lowhigh genotype weremore common in female cases and mothers Severity ofautism slightly tends to increase in low expressing allele forboth cases and mothers Previous results found that autisticboys with the low expressing 3-repeat MAOA allele hadmore severe sensory behaviors arousal regulation problemsaggression and worse social communication skills thanmales with the high activity allele and that problems withaggression as well as with fears and rituals were modifiedby the mothersrsquo genotype [5]
The AG (A644G) noncoding SNP (rs1799836) ofMAOBis responsible for altered enzyme activity with tissue speci-ficity [28 29] This MAOB SNP was found to be associatedwith emotional regulation [30 31] and Parkinsonrsquos disease[32] Also it was implicated as risk factor for schizophrenia ina Spanish population [33] and in Han Chinese [34] So far noavailable information was found regarding relation ofMAOBpolymorphism and autism however this study highlightedthe importance of G allele in both male and female autisticpatients G allele was significantly higher in autistic casesthan in controls Interestingly GG genotype was absent in
6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009
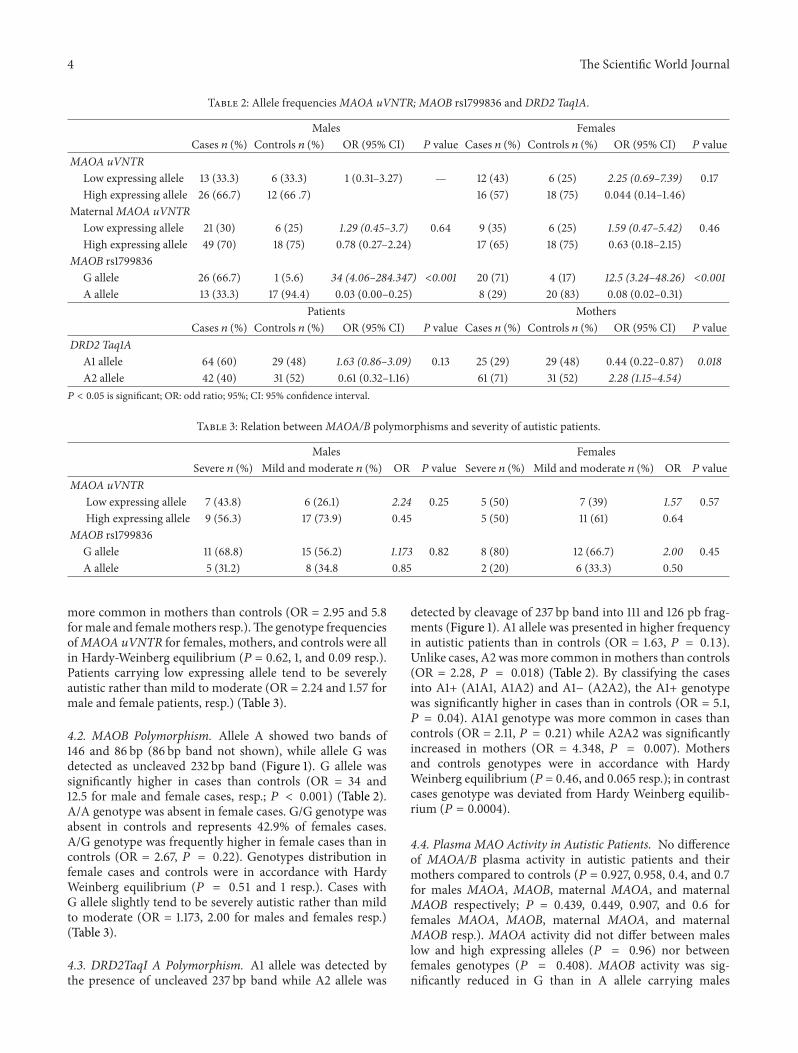
4 The Scientific World Journal
Table 2 Allele frequenciesMAOA uVNTR MAOB rs1799836 and DRD2 Taq1A
Males FemalesCases 119899 () Controls 119899 () OR (95 CI) 119875 value Cases 119899 () Controls 119899 () OR (95 CI) 119875 value
MAOA uVNTRLow expressing allele 13 (333) 6 (333) 1 (031ndash327) mdash 12 (43) 6 (25) 225 (069ndash739) 017High expressing allele 26 (667) 12 (66 7) 16 (57) 18 (75) 0044 (014ndash146)
MaternalMAOA uVNTRLow expressing allele 21 (30) 6 (25) 129 (045ndash37) 064 9 (35) 6 (25) 159 (047ndash542) 046High expressing allele 49 (70) 18 (75) 078 (027ndash224) 17 (65) 18 (75) 063 (018ndash215)
MAOB rs1799836G allele 26 (667) 1 (56) 34 (406ndash284347) lt0001 20 (71) 4 (17) 125 (324ndash4826) lt0001A allele 13 (333) 17 (944) 003 (000ndash025) 8 (29) 20 (83) 008 (002ndash031)
Patients MothersCases 119899 () Controls 119899 () OR (95 CI) P value Cases 119899 () Controls 119899 () OR (95 CI) P value
DRD2 Taq1AA1 allele 64 (60) 29 (48) 163 (086ndash309) 013 25 (29) 29 (48) 044 (022ndash087) 0018A2 allele 42 (40) 31 (52) 061 (032ndash116) 61 (71) 31 (52) 228 (115ndash454)119875 lt 005 is significant OR odd ratio 95 CI 95 confidence interval
Table 3 Relation betweenMAOAB polymorphisms and severity of autistic patients
Males FemalesSevere 119899 () Mild and moderate 119899 () OR 119875 value Severe 119899 () Mild and moderate 119899 () OR 119875 value
MAOA uVNTRLow expressing allele 7 (438) 6 (261) 224 025 5 (50) 7 (39) 157 057High expressing allele 9 (563) 17 (739) 045 5 (50) 11 (61) 064
MAOB rs1799836G allele 11 (688) 15 (562) 1173 082 8 (80) 12 (667) 200 045A allele 5 (312) 8 (348 085 2 (20) 6 (333) 050
more common in mothers than controls (OR = 295 and 58formale and femalemothers resp)The genotype frequenciesofMAOA uVNTR for females mothers and controls were allin Hardy-Weinberg equilibrium (119875 = 062 1 and 009 resp)Patients carrying low expressing allele tend to be severelyautistic rather than mild to moderate (OR = 224 and 157 formale and female patients resp) (Table 3)
42 MAOB Polymorphism Allele A showed two bands of146 and 86 bp (86 bp band not shown) while allele G wasdetected as uncleaved 232 bp band (Figure 1) G allele wassignificantly higher in cases than controls (OR = 34 and125 for male and female cases resp 119875 lt 0001) (Table 2)AA genotype was absent in female cases GG genotype wasabsent in controls and represents 429 of females casesAG genotype was frequently higher in female cases than incontrols (OR = 267 119875 = 022) Genotypes distribution infemale cases and controls were in accordance with HardyWeinberg equilibrium (119875 = 051 and 1 resp) Cases withG allele slightly tend to be severely autistic rather than mildto moderate (OR = 1173 200 for males and females resp)(Table 3)
43 DRD2TaqI A Polymorphism A1 allele was detected bythe presence of uncleaved 237 bp band while A2 allele was
detected by cleavage of 237 bp band into 111 and 126 pb frag-ments (Figure 1) A1 allele was presented in higher frequencyin autistic patients than in controls (OR = 163 119875 = 013)Unlike cases A2 wasmore common inmothers than controls(OR = 228 119875 = 0018) (Table 2) By classifying the casesinto A1+ (A1A1 A1A2) and A1minus (A2A2) the A1+ genotypewas significantly higher in cases than in controls (OR = 51119875 = 004) A1A1 genotype was more common in cases thancontrols (OR = 211 119875 = 021) while A2A2 was significantlyincreased in mothers (OR = 4348 119875 = 0007) Mothersand controls genotypes were in accordance with HardyWeinberg equilibrium (119875 = 046 and 0065 resp) in contrastcases genotype was deviated from Hardy Weinberg equilib-rium (119875 = 00004)
44 Plasma MAO Activity in Autistic Patients No differenceof MAOAB plasma activity in autistic patients and theirmothers compared to controls (119875 = 0927 0958 04 and 07for males MAOA MAOB maternal MAOA and maternalMAOB respectively 119875 = 0439 0449 0907 and 06 forfemales MAOA MAOB maternal MAOA and maternalMAOB resp) MAOA activity did not differ between maleslow and high expressing alleles (119875 = 096) nor betweenfemales genotypes (119875 = 0408) MAOB activity was sig-nificantly reduced in G than in A allele carrying males
The Scientific World Journal 5
(a) (b)
180 190 230 240
(e)
(c) (d)
T
T T
T T T T T T TC C C C C C190 200A A A A
A A A A
A A A AAG
G G G G G G G G G G G G G
C A A A A A A AT T T T T T
TTTTTTTTTT
80 90 100 310 320G G G G GG G G G G G G G G G G G G GGC C C C C C
CCCC CCCCCC
CC
GndashA SNP is absent CndashT and GndashTC are absent
GndashA SNP is absentCndashT SNP is absent
AndashG mutation is absent
Figure 2 Sequencing chromatogram ofmiR-21 andmiR-431 (a) it shows the absence of AndashGmutation downstream the pre-miR-21 ((b) (c)(d) and (e)) they show the absence of rs12883709 GndashA upstream the pre-miR-431 rs76090066 CndashT and rs12884005 GndashA in the sequence ofpre-miR-431 rs61993318 CndashT and rs35695758 GndashTC downstream the pre-miR-431
(119875 = 0027) while females showed no difference of MAOBactivity between AG and GG genotypes (119875 = 082)
45 miR-431 and miR-21 Variations rs12883709 GAupstream the pre-miR-431 rs12884005 GA and rs76090066CT in the sequence of the pre-miR-431 rs 61993318 CTand rs35695758 GTC downstream the pre-miR-431 wereall absent Also AndashG mutation at 29-nt downstream ofpre-miR-21 was absent No variations were observed in theamplified sequences (Figure 2)
5 Discussion
Twin and family studies provide indisputable evidence fora genetic component in autism The average concordancefor identical twins versus fraternal twins was frequentlystudied and reported to be 36 versus 3 [16] 64 versus9 [17] and 60 versus 0 [18] However a concordancerate for monozygotic twins of less than 100 indicates thatnongenetic factors also play a causal role The current studyshowed that about 19 of patients had similar affected familymembers Elevated rates of clinical psychiatric disordersdistinct from autism have been reported among the relativesof individuals with autism including schizophrenia anxietydepression and social phobias [19] Parental consanguinityrepresented 34 of cases in our study it is higher than thatof Saltık and Basgul (214) [20] However Datta et al [21]found that possible parental consanguinity increases thelikelihood of autism and behavioral disturbances manyreports did not find a link between consanguinity and autismin Egyptian [22] Saudi [23] and Iranian population [24]
Deletion of MAOAB was associated with severe mentalretardation and unusual stereotypical behaviors of hand
wringing and lip smacking in males [25] Bortolato et al[26] found that both MAOAB knockout mice displayedneuropathological alterations reminiscent of typical ASDfeatures Previous study found consistent association betweenthe ldquolow activityrdquo allele of MAOA and larger brain volumesfor regions of the cortex in children with autism but notin controls In contrast the data did not find association ofthe MAOA promoter polymorphism with autism itself [3]Tassone et al [27] suggested that functionalMAOA promoteralleles play a potential role in the male child the motheror both in ASD In this study no difference in allele fre-quency ofMAOA betweenmale autistic patients and controlsHowever low expressing allele and lowhigh genotype weremore common in female cases and mothers Severity ofautism slightly tends to increase in low expressing allele forboth cases and mothers Previous results found that autisticboys with the low expressing 3-repeat MAOA allele hadmore severe sensory behaviors arousal regulation problemsaggression and worse social communication skills thanmales with the high activity allele and that problems withaggression as well as with fears and rituals were modifiedby the mothersrsquo genotype [5]
The AG (A644G) noncoding SNP (rs1799836) ofMAOBis responsible for altered enzyme activity with tissue speci-ficity [28 29] This MAOB SNP was found to be associatedwith emotional regulation [30 31] and Parkinsonrsquos disease[32] Also it was implicated as risk factor for schizophrenia ina Spanish population [33] and in Han Chinese [34] So far noavailable information was found regarding relation ofMAOBpolymorphism and autism however this study highlightedthe importance of G allele in both male and female autisticpatients G allele was significantly higher in autistic casesthan in controls Interestingly GG genotype was absent in
6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009
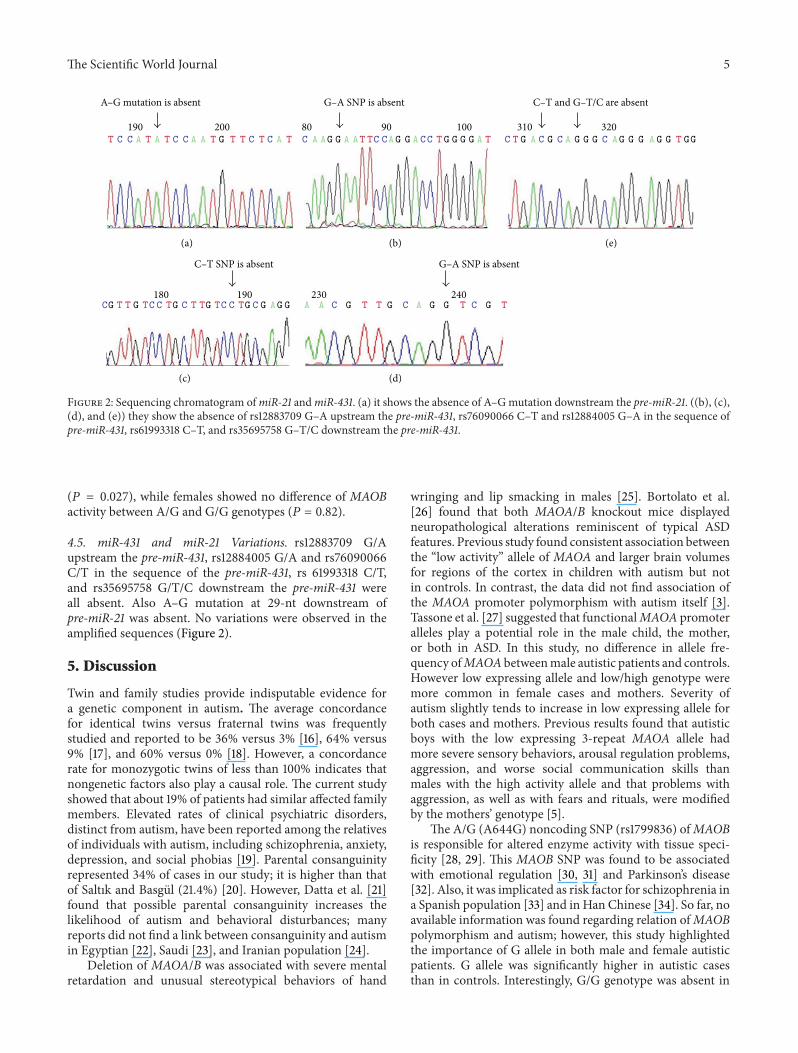
The Scientific World Journal 5
(a) (b)
180 190 230 240
(e)
(c) (d)
T
T T
T T T T T T TC C C C C C190 200A A A A
A A A A
A A A AAG
G G G G G G G G G G G G G
C A A A A A A AT T T T T T
TTTTTTTTTT
80 90 100 310 320G G G G GG G G G G G G G G G G G G GGC C C C C C
CCCC CCCCCC
CC
GndashA SNP is absent CndashT and GndashTC are absent
GndashA SNP is absentCndashT SNP is absent
AndashG mutation is absent
Figure 2 Sequencing chromatogram ofmiR-21 andmiR-431 (a) it shows the absence of AndashGmutation downstream the pre-miR-21 ((b) (c)(d) and (e)) they show the absence of rs12883709 GndashA upstream the pre-miR-431 rs76090066 CndashT and rs12884005 GndashA in the sequence ofpre-miR-431 rs61993318 CndashT and rs35695758 GndashTC downstream the pre-miR-431
(119875 = 0027) while females showed no difference of MAOBactivity between AG and GG genotypes (119875 = 082)
45 miR-431 and miR-21 Variations rs12883709 GAupstream the pre-miR-431 rs12884005 GA and rs76090066CT in the sequence of the pre-miR-431 rs 61993318 CTand rs35695758 GTC downstream the pre-miR-431 wereall absent Also AndashG mutation at 29-nt downstream ofpre-miR-21 was absent No variations were observed in theamplified sequences (Figure 2)
5 Discussion
Twin and family studies provide indisputable evidence fora genetic component in autism The average concordancefor identical twins versus fraternal twins was frequentlystudied and reported to be 36 versus 3 [16] 64 versus9 [17] and 60 versus 0 [18] However a concordancerate for monozygotic twins of less than 100 indicates thatnongenetic factors also play a causal role The current studyshowed that about 19 of patients had similar affected familymembers Elevated rates of clinical psychiatric disordersdistinct from autism have been reported among the relativesof individuals with autism including schizophrenia anxietydepression and social phobias [19] Parental consanguinityrepresented 34 of cases in our study it is higher than thatof Saltık and Basgul (214) [20] However Datta et al [21]found that possible parental consanguinity increases thelikelihood of autism and behavioral disturbances manyreports did not find a link between consanguinity and autismin Egyptian [22] Saudi [23] and Iranian population [24]
Deletion of MAOAB was associated with severe mentalretardation and unusual stereotypical behaviors of hand
wringing and lip smacking in males [25] Bortolato et al[26] found that both MAOAB knockout mice displayedneuropathological alterations reminiscent of typical ASDfeatures Previous study found consistent association betweenthe ldquolow activityrdquo allele of MAOA and larger brain volumesfor regions of the cortex in children with autism but notin controls In contrast the data did not find association ofthe MAOA promoter polymorphism with autism itself [3]Tassone et al [27] suggested that functionalMAOA promoteralleles play a potential role in the male child the motheror both in ASD In this study no difference in allele fre-quency ofMAOA betweenmale autistic patients and controlsHowever low expressing allele and lowhigh genotype weremore common in female cases and mothers Severity ofautism slightly tends to increase in low expressing allele forboth cases and mothers Previous results found that autisticboys with the low expressing 3-repeat MAOA allele hadmore severe sensory behaviors arousal regulation problemsaggression and worse social communication skills thanmales with the high activity allele and that problems withaggression as well as with fears and rituals were modifiedby the mothersrsquo genotype [5]
The AG (A644G) noncoding SNP (rs1799836) ofMAOBis responsible for altered enzyme activity with tissue speci-ficity [28 29] This MAOB SNP was found to be associatedwith emotional regulation [30 31] and Parkinsonrsquos disease[32] Also it was implicated as risk factor for schizophrenia ina Spanish population [33] and in Han Chinese [34] So far noavailable information was found regarding relation ofMAOBpolymorphism and autism however this study highlightedthe importance of G allele in both male and female autisticpatients G allele was significantly higher in autistic casesthan in controls Interestingly GG genotype was absent in
6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009

6 The Scientific World Journal
controls whileAAgenotypewas absent in female cases Suchfindings may suggest its potential role in the impairmentsfound in individuals with autism
No correspondence was found between the low and highMAOA genotype and brain MAOA activity in healthy males[35] and in postmortem samples [36]This study showed thatneitherMAOA norMAOB activity had significant differencein plasma of autistic patients and mothers compared tocontrols Partially results of this study contradict a study onOmani autistic childrenwhich showed a significant reductionin the level of plasma MAOA activity with nonsignificantchange in the level ofMAOB [37] No association was foundbetween MAOA genotype and plasma enzyme activity Theprevious results suggesting that genotype does not have adirect effect on brain enzyme activity [35] MAOB activitywas significantly reduced in G allele than in A allele carryingmales (119875 = 0027) The G allele had been associated withlower MAOB activity in human brain [38] In contrastGarpenstrand et al [39] found that individuals with the ldquoA-allelerdquo displayed significantly lower platelets enzyme activitythan individuals with the ldquoG-allelerdquo However Pivac et al [29]found that plateletMAOB activity did not differ betweenmensubjects subdivided into those with A allele or G allele
Postsynaptic D2 receptors and presynaptic D2 autorecep-tors are involved in the dopaminergic (DAergic) modulationof cognitive and emotional processes that are impaired inindividuals with autism [40] Previous results showed that thesubjects with one or two A1 alleles had lower DRD2 densitythan those without this allele Therefore the DRD2 TaqI Apolymorphism may be one of the important markers for theDRD2 density and function [41] Minor allele frequency forthe Taq1 A ranges from 20 in Caucasians to 44 in otherethnic groups [42] In this study A1 allele was predominatingin autistic patients than in controls Unlike cases A2 wasmore common in mothers than controls
A mutation or a single nucleotide polymorphism (SNP)at the miRNA gene region might affect the transcription ofpri-miRNA transcripts the processing of miRNA precur-sors to mature miRNAs or miRNA-target interactions [43]It was suggested that dysregulation of miRNA expressioncontributes to the observed alterations in gene expressionand in turn may lead to the pathophysiological conditionsunderlying autism [2] A study by Abu-Elneel et al [12] foundthat altered miRNA expression levels are observed in post-mortem cerebellar cortex from autism patients Among thesedysregulated miRNAs were miR-431 (14q322) and miR-21(17q231) rs12883709 GA rs12884005 GA rs76090066 CTrs61993318 CT and rs35695758 GTC were absent in thecurrent study Zhu et al [43] reported AndashG mutation at 29-nt downstream of pre-miR-21 led to a conformational changeof the secondary structure close to the stem reaching intothe pre-miR-21 and a relative reduction of the mature miR-21 expression in vivo Even though we did not report any ofSNPs or mutation in regions amplified it could be attributedto small number of cases so a larger number of patientsmiRNAs and amplified sequences are recommended
This study threw light on some neurotransmitter-relatedgenes suggesting their potential role in Autism pathogenesisthat warrants further studies and much consideration
Conflict of Interests
The authors declare no conflict of interests
References
[1] S Chakrabarti and E Fombonne ldquoPervasive developmentaldisorders in preschool children confirmation of high preva-lencerdquo American Journal of Psychiatry vol 162 no 6 pp 1133ndash1141 2005
[2] T Sarachana R Zhou G Chen H K Manji and V WHu ldquoInvestigation of post-transcriptional gene regulatory net-works associated with autism spectrumdisorders bymicroRNAexpression profiling of lymphoblastoid cell linesrdquo GenomeMedicine vol 2 no 4 article 23 2010
[3] L K Davis H C Hazlett A L Librant et al ldquoCortical enlarge-ment in autism is associated with a functional VNTR in themonoamine oxidase a generdquo American Journal of MedicalGenetics B vol 147 no 7 pp 1145ndash1151 2008
[4] I L Cohen X Liu C Schutz et al ldquoAssociation of autism sever-ity with a monoamine oxidase A functional polymorphismrdquoClinical Genetics vol 64 no 3 pp 190ndash197 2003
[5] I L Cohen X Liu M E S Lewis et al ldquoAutism severity isassociated with child and maternal MAOA genotypesrdquo ClinicalGenetics vol 79 no 4 pp 355ndash362 2011
[6] J A Hettinger X Liu M L Hudson et al ldquoDRD2 andPPP1R1B (DARPP-32) polymorphisms independently conferincreased risk for autism spectrum disorders and additivelypredict affected status in male-only affected sib-pair familiesrdquoBehavioral and Brain Functions vol 8 article 19 2012
[7] P W Troost B E Lahuis M-P Steenhuis et al ldquoLong-term effects of risperidone in children with autism spectrumdisorders a placebo discontinuation studyrdquo Journal of theAmerican Academy of Child and Adolescent Psychiatry vol 44no 11 pp 1137ndash1144 2005
[8] S Z Sabol S Hu and D Hamer ldquoA functional polymorphismin the monoamine oxidase A gene promoterrdquoHuman Geneticsvol 103 no 3 pp 273ndash279 1998
[9] R M Wu C W Cheng K H Chen et al ldquoThe COMT L allelemodifies the association between MAOB polymorphism andPD in Taiwaneserdquo Neurology vol 56 no 3 pp 375ndash382 2001
[10] A Suzuki KMihara T Kondo et al ldquoThe relationship betweendopamine D2 receptor polymorphism at the Taq1 A locus andtherapeutic response to nemonapride a selective dopamineantagonist in schizophrenic patientsrdquo Pharmacogenetics vol10 no 4 pp 335ndash341 2000
[11] V Ambros ldquoThe functions of animal microRNAsrdquo Nature vol431 no 7006 pp 350ndash355 2004
[12] K Abu-Elneel T Liu F S Gazzaniga et al ldquoHeterogeneousdysregulation of microRNAs across the autism spectrumrdquoNeurogenetics vol 9 no 3 pp 153ndash161 2008
[13] J Sambrook and D Russell Commonly Used Techniques inMolecular Cloning vol 3 ofMolecular Cloning appendix 8 ColdSpringHarbor Laboratory Press Cold SpringHarbor NY USA3rd edition 2001
[14] L G Schmidt T Sander S Kuhn et al ldquoDifferent allele distri-bution of a regulatory MAOA gene promoter polymorphism inantisocial and anxious-depressive alcoholicsrdquo Journal of NeuralTransmission vol 107 no 6 pp 681ndash689 2000
[15] T-J Wang S-Y Huang W-W Lin et al ldquoPossible interactionbetween MAOA and DRD2 genes associated with antisocial
The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009

The Scientific World Journal 7
alcoholism among Han Chinese men in Taiwanrdquo Progress inNeuro-Psychopharmacology and Biological Psychiatry vol 31no 1 pp 108ndash114 2007
[16] M B Lauritsen and H Ewald ldquoThe genetics of autismrdquo ActaPsychiatrica Scandinavica vol 103 no 6 pp 411ndash427 2001
[17] L DeFrancesco ldquoAutism on the riserdquo The Scientist vol 15 pp16ndash19 2001
[18] R Muhle S V Trentacoste and I Rapin ldquoThe genetics ofautismrdquo Pediatrics vol 113 no 5 pp e472ndashe486 2004
[19] S Bolte S Knecht and F Poustka ldquoA case-control study ofpersonality style and psychopathology in parents of subjectswith autismrdquo Journal of Autism and Developmental Disordersvol 37 no 2 pp 243ndash250 2007
[20] S Saltık and S Basgul ldquoNeurological disorders combined withautism in childrenrdquoNobel Medicus Journal vol 8 no 3 pp 113ndash120 2012
[21] A KDatta SMandal and S Bhattacharya ldquoAutism andmentalretardationwith convulsion in tuberous sclerosis a case reportrdquoCases Journal vol 2 no 7 article 7061 2009
[22] M A El Sawy H I Awadalla R R Mohamed O K ZakiandO AMohamed ldquoStudy of some environmental and geneticdeterminants of autism in Egyptian childrenrdquoThe InternationalJournal of Psychology and Counselling vol 3 no 8 pp 130ndash1362011
[23] S M Al-Salehi E H Al-Hifthy andM Ghaziuddin ldquoAutism inSaudi Arabia presentation clinical correlates and comorbidityrdquoTranscultural Psychiatry vol 46 no 2 pp 340ndash347 2009
[24] R Sasanfar S Haddad A Tolouei M Ghadami D Yu and SSantangelo ldquoPaternal age increases the risk for autism in anIranian population samplerdquo Molecular Autism vol 1 no 1article 2 2010
[25] L Rodriguez-Revenga I Madrigal L S Alkhalidi et al ldquoCon-tiguous deletion of the NDP MAOA MAOB and EFHC2 genesin a patientwithNorrie disease severe psychomotor retardationand myoclonic epilepsyrdquo American Journal of Medical GeneticsA vol 143 no 9 pp 916ndash920 2007
[26] M Bortolato S C Godar L Alzghoul et al ldquoMonoamineoxidase A andAB knockoutmice display autistic-like featuresrdquoThe International Journal of Neuropsychopharmacology vol 16no 4 pp 869ndash888 2013
[27] F Tassone L Qi W Zhang R L Hansen I N Pessah and IHertz-Picciotto ldquoMAOA DBH and SLC6A4 variants inCHARGE a case-control study of autism spectrum disordersrdquoAutism Research vol 4 no 4 pp 250ndash261 2011
[28] V Filic A Vladic J Stefulj et al ldquoMonoamine oxidases A andB gene polymorphisms in migraine patientsrdquo Journal of theNeurological Sciences vol 228 no 2 pp 149ndash153 2005
[29] N Pivac J KnezevicMMustapic et al ldquoThe lack of associationbetween monoamine oxidase (MAO) intron 13 polymorphismand platelet MAO-B activity among menrdquo Life Sciences vol 79no 1 pp 45ndash49 2006
[30] A M Dlugos A A Palmer and H deWit ldquoNegative emotion-ality monoamine oxidase B gene variants modulate personalitytraits in healthy humansrdquo Journal of Neural Transmission vol116 no 10 pp 1323ndash1334 2009
[31] M Bortolato and J C Shih ldquoBehavioral outcomes of mono-amine oxidase deficiency preclinical and clinical evidencerdquoInternational Review of Neurobiology vol 100 pp 13ndash42 2011
[32] J H Kurth M C Kurth S E Poduslo and J D SchwankhausldquoAssociation of a monoamine oxidase B allele with Parkinsonrsquosdiseaserdquo Annals of Neurology vol 33 no 4 pp 368ndash372 1993
[33] P Gasso M Bernardo S Mas et al ldquoAssociation of AG poly-morphism in intron 13 of the monoamine oxidase B gene withschizophrenia in a Spanish populationrdquo Neuropsychobiologyvol 58 no 2 pp 65ndash70 2008
[34] Y-LWei C-X Li S-B Li Y Liu and L Hu ldquoAssociation studyof monoamine oxidase AB genes and schizophrenia in HanChineserdquo Behavioral and Brain Functions vol 7 article 42 2011
[35] J S FowlerNAlia-Klein AKriplani et al ldquoEvidence that brainMAO A activity does not correspond to MAO A genotype inhealthy male subjectsrdquo Biological Psychiatry vol 62 no 4 pp355ndash358 2007
[36] L Emilsson U Pettersson E E Jazin L Oreland and J Bal-ciuniene ldquoInvestigation of the functional effect of monoamineoxidase polymorphisms in human brainrdquoHuman Genetics vol110 no 1 pp 1ndash7 2002
[37] M M Essa M M Al-Sharbati Y M Al-Farsi et al ldquoAlteredactivities of monoamine oxidase A in omani autistic childrenmdasha brief reportrdquo International Journal of Biological and MedicalResearch vol 2 no 3 pp 811ndash813 2011
[38] N Alia-Klein R Z Goldstein A Kriplani et al ldquoBrain mono-amine oxidase A activity predicts trait aggressionrdquo Journal ofNeuroscience vol 28 no 19 pp 5099ndash5104 2008
[39] H Garpenstrand J Ekblom K Forslund G Rylander andL Oreland ldquoPlatelet monoamine oxidase activity is related toMAOB intron 13 genotyperdquo Journal of Neural Transmission vol107 no 5 pp 523ndash530 2000
[40] S B Floresco O Magyar S Ghods-Sharifi C Vexelman andM T L Tse ldquoMultiple dopamine receptor subtypes in themedial prefrontal cortex of the rat regulate set-shiftingrdquo Neu-ropsychopharmacology vol 31 no 2 pp 297ndash309 2006
[41] A Suzuki T Kondo K Mihara et al ldquoAssociation betweenTaqI a dopamine D2 receptor polymorphism and therapeu-tic response to bromperidol a preliminary reportrdquo EuropeanArchives of Psychiatry amp Clinical Neuroscience vol 251 no 2pp 57ndash59 2001
[42] J-P Zhang T Lencz and A K Malhotra ldquoD2 receptor geneticvariation and clinical response to antipsychotic drug treatmenta meta-analysisrdquo American Journal of Psychiatry vol 167 no 7pp 763ndash772 2010
[43] Z Zhu W Gao Z Qian and Y Miao ldquoGenetic variation ofmiRNA sequence in pancreatic cancerrdquo Acta Biochimica etBiophysica Sinica vol 41 no 5 Article ID gmp023 pp 407ndash4132009
Related Documents