Short Notes Genetic differentiation in two European tree frog (Hyla arborea) metapopulations in contrasted landscapes of western Switzerland Sylvain Dubey 1,2, ∗ , Sylvain Ursenbacher 1,3 , Jérôme Pellet 1,4 , Luca Fumagalli 1 Abstract. The survival of threatened species as the European tree frog (Hyla arborea) is strongly dependent on the genetic variability within populations, as well as gene flow between them. In Switzerland, only two sectors in its western part still harbour metapopulations. The first is characterised by a very heterogeneous and urbanized landscape, while the second is characterised by a uninterrupted array of suitable habitats. In this study, six microsatellite loci were used to establish levels of genetic differentiation among the populations from the two different locations. The results show that the metapopulations have: (i) weak levels of genetic differentiation (F ST within metapopulation ≈ 0.04), (ii) no difference in levels of genetic structuring between them, (iii) significant (p = 0.019) differences in terms of genetic diversity (Hs) and observed heterozygozity (Ho), the metapopulation located in a disturbed landscape showing lower values. Our results suggest that even if the dispersal of H. arborea among contiguous ponds seems to be efficient in areas of heterogeneous landscape, a loss of genetic diversity can occur. Keywords: amphibians, conservation, fragmentation, Hylidae, microsatellite markers. As a result of growing urbanization and sub- sequent habitat degradation and fragmentation, numerous animal species currently face extinc- tion or have declined drastically during the last century. These disturbances commonly result in population size reduction and decrease the possibility of migration and subsequent gene flow between populations (e.g. Frankham et al., 2002; Cushman, 2006). The genetic conse- quences of population fragmentation are com- plex and critically depend upon levels of gene 1 - Department of Ecology and Evolution, Laboratory for Conservation Biology, Biophore, CH-1015 Lausanne, Switzerland 2 - Heydon-Laurence Bld, A08, Science Road, School of Biological Sciences, University of Sydney, Sydney, NSW 2006, Australia 3 - Department of Environmental Sciences, Section of Con- servation Biology, University of Basel, St. Johanns- Vorstadt 10, CH-4056 Basel, Switzerland 4 - A. Maibach Sàrl, CP99, Ch. de la Poya 10, 1610 CH- Oron-la-Ville, Switzerland ∗ Corresponding author; e-mail: [email protected] flow between fragments. When it is restricted, population fragmentation is expected to reduce within-population genetic polymorphism and increase genetic differentiation among popula- tions, typically leading to a loss of genetic diver- sity within fragments (e.g. Hitchings and Bee- bee, 1997). Consequently, understanding the ef- fects of population fragmentation is crucial in conservation biology, since extinction risks are more elevated in fragmented populations with low levels of genetic variability (Frankham et al., 2002; Frankham, 2005). The studied species, The European tree frog (Hyla arborea), is a pond-breeding species which possess a distribution area extending from Portugal to southern Sweden, and from the Balkans to west Asia (Gasc et al., 1997). As in several other amphibian species (Wake, 1991), a strong decline across its entire dis- tribution range has been recorded during the last decades (Gasc et al., 1997). In Switzerland, once widespread across this country, the species has been reduced to less than one dozen sec- © Koninklijke Brill NV, Leiden, 2009. Amphibia-Reptilia 30 (2009): 127-133 Also available online - www.brill.nl/amre

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Short Notes
Genetic differentiation in two European tree frog (Hyla arborea)metapopulations in contrasted landscapes of western Switzerland
Sylvain Dubey12lowast Sylvain Ursenbacher13 Jeacuterocircme Pellet14 Luca Fumagalli1
Abstract The survival of threatened species as the European tree frog (Hyla arborea) is strongly dependent on thegenetic variability within populations as well as gene flow between them In Switzerland only two sectors in its westernpart still harbour metapopulations The first is characterised by a very heterogeneous and urbanized landscape while thesecond is characterised by a uninterrupted array of suitable habitats In this study six microsatellite loci were used toestablish levels of genetic differentiation among the populations from the two different locations The results show thatthe metapopulations have (i) weak levels of genetic differentiation (FST within metapopulation asymp 004) (ii) no differencein levels of genetic structuring between them (iii) significant (p = 0019) differences in terms of genetic diversity (Hs) andobserved heterozygozity (Ho) the metapopulation located in a disturbed landscape showing lower values Our results suggestthat even if the dispersal of H arborea among contiguous ponds seems to be efficient in areas of heterogeneous landscape aloss of genetic diversity can occur
Keywords amphibians conservation fragmentation Hylidae microsatellite markers
As a result of growing urbanization and sub-sequent habitat degradation and fragmentationnumerous animal species currently face extinc-tion or have declined drastically during the lastcentury These disturbances commonly resultin population size reduction and decrease thepossibility of migration and subsequent geneflow between populations (eg Frankham etal 2002 Cushman 2006) The genetic conse-quences of population fragmentation are com-plex and critically depend upon levels of gene
1 - Department of Ecology and Evolution Laboratory forConservation Biology Biophore CH-1015 LausanneSwitzerland
2 - Heydon-Laurence Bld A08 Science Road School ofBiological Sciences University of Sydney SydneyNSW 2006 Australia
3 - Department of Environmental Sciences Section of Con-servation Biology University of Basel St Johanns-Vorstadt 10 CH-4056 Basel Switzerland
4 - A Maibach Sagraverl CP99 Ch de la Poya 10 1610 CH-Oron-la-Ville SwitzerlandlowastCorresponding author e-mailsylvaindubeybiousydeduau
flow between fragments When it is restrictedpopulation fragmentation is expected to reducewithin-population genetic polymorphism andincrease genetic differentiation among popula-tions typically leading to a loss of genetic diver-sity within fragments (eg Hitchings and Bee-bee 1997) Consequently understanding the ef-fects of population fragmentation is crucial inconservation biology since extinction risks aremore elevated in fragmented populations withlow levels of genetic variability (Frankham etal 2002 Frankham 2005)
The studied species The European tree frog(Hyla arborea) is a pond-breeding specieswhich possess a distribution area extendingfrom Portugal to southern Sweden and fromthe Balkans to west Asia (Gasc et al 1997)As in several other amphibian species (Wake1991) a strong decline across its entire dis-tribution range has been recorded during thelast decades (Gasc et al 1997) In Switzerlandonce widespread across this country the specieshas been reduced to less than one dozen sec-
copy Koninklijke Brill NV Leiden 2009 Amphibia-Reptilia 30 (2009) 127-133Also available online - wwwbrillnlamre
128 Short Notes
tors which undergo local extinctions (Grossen-bacher 1988 1994) This severe decline isthought to be mainly due to anthropogenic ac-tivities Studies carried out on the Europeantree frog in Sweden (Carlson and Edenhamn2000) as well as in Switzerland (Pellet et al2006) highlighted regular extinction and re-colonisation events characteristic of a metapop-ulation dynamics (Hanski and Gilpin 1997)Extensive theory has been developed to modelgenetic processes within metapopulation struc-tures Due to frequent extinctions and bottle-necks during recolonisations metapopulationsare likely to suffer more rapidly from inbreed-ing and fitness reduction than single large popu-
lations with the same total size (Gilpin 1991Hanski and Gilpin 1997)
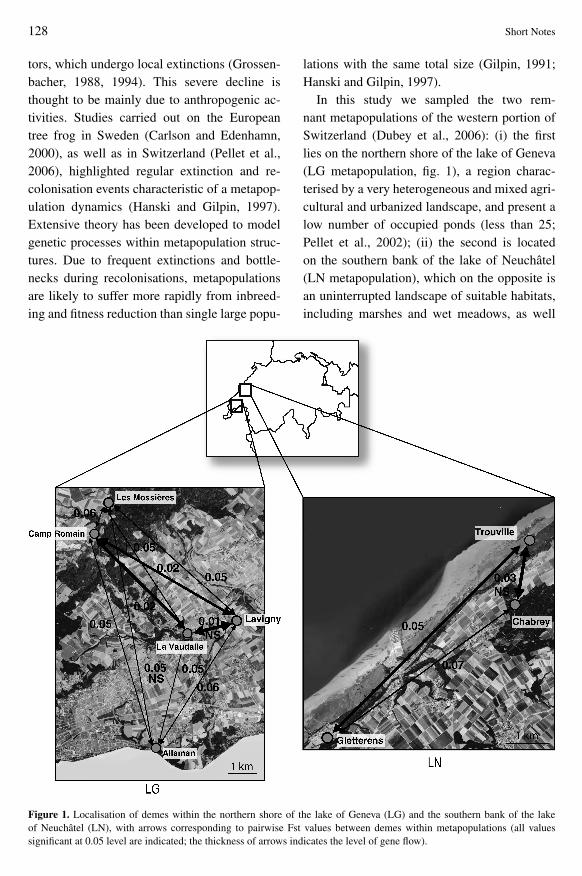
In this study we sampled the two rem-nant metapopulations of the western portion ofSwitzerland (Dubey et al 2006) (i) the firstlies on the northern shore of the lake of Geneva(LG metapopulation fig 1) a region charac-terised by a very heterogeneous and mixed agri-cultural and urbanized landscape and present alow number of occupied ponds (less than 25Pellet et al 2002) (ii) the second is locatedon the southern bank of the lake of Neuchacirctel(LN metapopulation) which on the opposite isan uninterrupted landscape of suitable habitatsincluding marshes and wet meadows as well
Figure 1 Localisation of demes within the northern shore of the lake of Geneva (LG) and the southern bank of the lakeof Neuchacirctel (LN) with arrows corresponding to pairwise Fst values between demes within metapopulations (all valuessignificant at 005 level are indicated the thickness of arrows indicates the level of gene flow)
Short Notes 129
as woodland bordering the lake in a continuousway In the year 2000 40 calling ponds weredetected totalling several thousand frogs (Pel-let and Neet 2001) This area is considered asone of the largest metapopulation in Switzer-land (Grossenbacher 1988)
Information concerning the extent of popu-lation fragmentation is critical to determinewhether a species requires proactive manage-ment plans to reduce extinction risks associ-ated with genetic stochasticity Hence to es-timate the effect of habitat fragmentation onthe genetic structure and variability of tree frogpopulations we investigated the two isolatedmetapopulations in western Switzerland withsix microsatellite loci We hypothesized that inmetapopulations embedded in continuous habi-tats genetic structure would be lower while ge-netic variability and gene flow higher comparedto metapopulations characterized by lower den-sities and where heterogeneous habitat may actas a barrier to gene flow
During spring 2002 and 2003 a total of 235 samplesof Hyla arborea (tadpoles and eggs) were collected in 8ponds in the two studied areas (fig 1) Five ponds weresampled within the LG metapopulation (Allaman CampRomain Lavigny Mossiegraveres Vaudalle) and three pondsin the LN metapopulation (Chabrey Gletterens Trouville)When eggs were collected only one egg per clutch was usedfor genetic analyses
DNA extraction from tissues and egg samples was car-ried out using the QIAamp DNA Mini Kit (QIAGEN) orwith a CTAB protocol (Milligan 1992) Six microsatelliteloci isolated and characterized for Hyla arborea (Wha1-9 Wha1-20 Wha1-25 Wha1-103 Wha1-104 Wha1-140Arens et al 2000) were amplified and scored (see Arens etal (2000) for the specific PCR profiles) Amplified productswere genotyped with an ABI PRISM 377 DNA Sequencerusing genescan analysis 21 software (Applied Biosystems)
Gene diversities comprising observed (Ho) expected(Hs) within-deme (the populations will be treated as demesin the different sections of the manuscript) and expectedoverall heterozygosities (Ht) were estimated following (Neiand Chesser 1983) Genotypic disequilibrium betweenloci in each sample and deviations from Hardy-Weinbergequilibrium (HWE) within samples were tested based on2400 permutations and 10 000 randomizations respectivelyWrightrsquos fixation indices for within-deme deviation fromrandom mating (FIS) as well as pairwise deme differen-tiation (FST ) were estimated following Weir and Cocker-ham (1984) Deviation from random mating within demes(FIS) per locus and sample were computed with a boot-strap procedure (2000 randomizations) Statistical support
for pairwise deme differentiation was obtained through ex-act G-tests on allelic frequencies as described by Goudet etal (1996) with 2000 randomizations All summary statisticsand tests mentioned above have been computed using FSTAT
Version 2932 (Goudet 1995) Permutation tests were car-ried out in order to detect significant differences in allelicrichness expected (Hs) and observed (Ho) heterozygositiesand FST indices among the two studied metapopulations
Genetic isolation by distance at the metapopulationallevel and overall was tested by using a partial Mantel test(Mantel 1967) p-value were given after 10 000 randomiza-tions
A Bayesian model-based clustering method (Pritchardet al 2000) for inferring population structure and assign-ing individuals to populations was used as implemented instructure version 21 (Falush et al 2003) Based on allelefrequencies individuals are assigned through the use of aMarkov chain Monte Carlo (MCMC) simulation a member-ship coefficient for each of K populations We performed 10runs of 6 middot105 iterations (the first 105 considered as burn-in)for K = 1 to K = 10 (i) including all the populations and(ii) within the two metapopulations The number of popu-lations best fitting our data set was defined as described inEvanno et al (2005) The latter statistics compares the rateof change in the log probability of data between successiveK and the corresponding variance of log probabilities
We used the software migrate 206 (Beerli and Felsen-stein 2001 Beerli 2004) to estimate the scaled migrationrate (M) between demes within metapopulations This soft-ware is based on a coalescence model with mutation andmigration and estimates a measure of effective populationsize θ defined as 4Neμ where μ denotes mutation rateand migration M defined as mμ where m denotes migra-tion rate We assumed a stepwise mutation model and basedestimates on 15 short [104 Markov Chain Monte Carlo(MCMC) steps] and five long (105 MCMC steps) chainsTo ensure convergence we used the lsquoadaptive heatingrsquo op-tion with one lsquocoldrsquo and three lsquohotrsquo chains
Tests for HWE indicated that all loci testedwere at HWE and in genotypic equilibrium Forthe six microsatellite loci the number of allelesper locus ranged from 7 to 17 (average = 983)with a total of 59 alleles across 6 loci The al-lelic richness within deme ranges from 441 to529 with an overall mean of 625 (table 1) Ex-pected heterozygosities per locus within demes(Hs) ranged from 016 to 090 with an av-erage of 058 whereas expected overall het-erozygosity (Ht) averaged 062 (range per lo-cus 035-087) Observed heterozygosity (Ho)values varied from 044 to 068 with an aver-age of 051 (see table 1) There was a signifi-cant deviation from random mating in the ana-lyzed demes (overall FIS = 012 p gt 0001
130 Short Notes
LN FIS = 006 LG FIS = 016) suggest-ing the occurence of a within-sample sub-structure The genetic differentiation betweendemes (pairwise FST ) within each metapopu-lation was low ranging from 001 to 007 allvalues being significant except for the pairs ofdemes Mossiegravere-Allaman Vaudalle-Lavignyand Trouville-Chabrey (p gt 005) Compara-ble overall FST values were found in the LN andLG metapopulations (respectively 0046 95CI 0017-005 and 0039 95 CI 0025-0079
Table 1 Genetic diversities for eight Hyla arborea demesin two distinct metapopulations based on 6 microsatelliteloci N = maximum number of singers in the population in2002 (Pellet et al 2002) n = sample size Ho = observedheterozygosity Hs = expected heterozygosity AR = allelicrichness
Site N n Ho Hs AR
1 Allaman 20 20 045 054 4622 Camp romain 27 24 045 059 5303 Lavigny 149 24 044 049 5124 Mossiegraveres 25 12 047 051 5225 Vaudalle 11 20 044 055 524
Mean LG (1 to 5) 232 100 045 054 510
6 Chabrey 26 21 058 066 5297 Gletterens 11 19 063 060 4418 Trouville 99 22 068 063 496
Mean LN (6 to 8) 136 62 060 064 489
TotalTotal Mean (1 to 8) 368 162 051 058 625
p = 086 see fig 1) The pairwise FST val-ues among demes from separated metapopula-tions varies from 0086 to 0142 (mean = 011)for geographical distances ranging from 592 to697 km
No isolation by distance within the LGmetapopulation was detected (p = 040 r2 =009) indicating the absence of a pattern of ge-netic isolation with geographical distance (nottested on the three populations of LN) Signifi-cant differences between metapopulations wereobserved for Ho (LN Ho = 060 LG Ho =045 p = 0019) and Hs (LN Hs = 064 LGHs = 054 p = 0019) whereas allelic rich-ness (AR) FST and FIS were not significantlydifferent (respectively p = 043 p = 086 andp = 011 see table 1)
The analyses performed with structure re-vealed that the number of populations best fit-ting our data set is K = 2 No substructure wasrevealed within the metapopulations
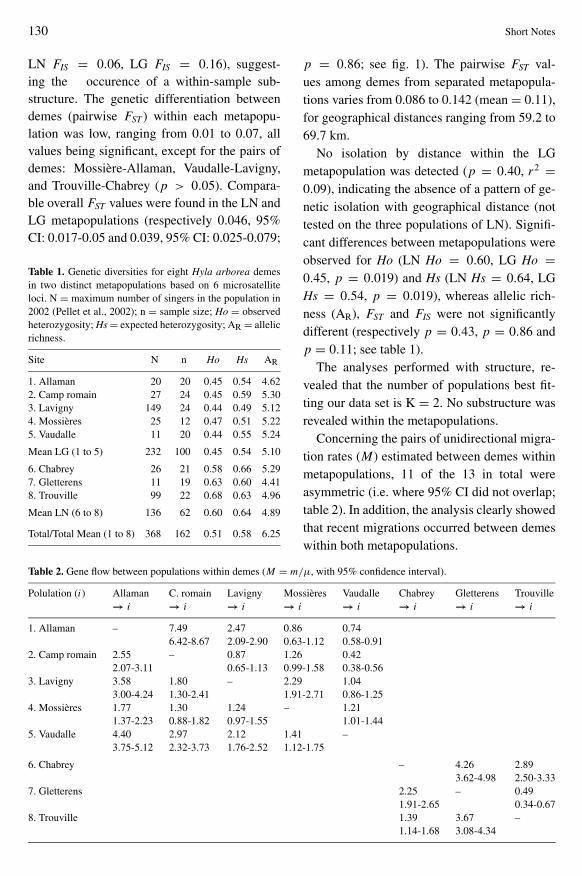
Concerning the pairs of unidirectional migra-tion rates (M) estimated between demes withinmetapopulations 11 of the 13 in total wereasymmetric (ie where 95 CI did not overlaptable 2) In addition the analysis clearly showedthat recent migrations occurred between demeswithin both metapopulations
Table 2 Gene flow between populations within demes (M = mμ with 95 confidence interval)
Polulation (i) Allaman C romain Lavigny Mossiegraveres Vaudalle Chabrey Gletterens Trouvillerarr i rarr i rarr i rarr i rarr i rarr i rarr i rarr i
1 Allaman ndash 749 247 086 074642-867 209-290 063-112 058-091
2 Camp romain 255 ndash 087 126 042207-311 065-113 099-158 038-056
3 Lavigny 358 180 ndash 229 104300-424 130-241 191-271 086-125
4 Mossiegraveres 177 130 124 ndash 121137-223 088-182 097-155 101-144
5 Vaudalle 440 297 212 141 ndash375-512 232-373 176-252 112-175
6 Chabrey ndash 426 289362-498 250-333
7 Gletterens 225 ndash 049191-265 034-067
8 Trouville 139 367 ndash114-168 308-434
Short Notes 131
The main result stemming from this studyis the absence of substantial differences interms of genetic differentiation between the twometapopulations despite an important contrastin habitat fragmentation and demes densities asshown by (i) the low global structure observedwithin metapopulations (mean FST asymp 004)with pair-wise FST ranging from 001 to 007(ii) the absence of substructure within metapop-ulations as revealed by clustering analyses and(iii) the unidirectional indices of migration (M)observed between demes within metapopula-tions with some very high values observed (Mmax for LN 426 and for LG 749) Thereforeour data suggest a high overall rate of disper-sal within both metapopulations Though thepairs of unidirectional indices of migration (M)were mostly asymmetric within both metapop-ulations (11 of 13 pairs) with values varyingfrom 042 to 749 revealing that the demes con-tributed differently to the low structure observedbetween them Therefore the genetic diversityof demes could be reduced by the effect ofasymmetric gene flow
In contrast with the low structure observedwithin both areas the analyses revealed signif-icant (p = 0019) lower gene diversity (Hs)and observed heterozygosity (Ho) within LGmetapopulation as well as a higher but not sig-nificantly different substructure (FIS)
Consequently the results pointed out that thelandscape of LG is less suitable for the Euro-pean tree frog than that of LN and as a resultdispersal is less effective to maintain gene flowamong local demes and as consequence geneticdiversity within demes This pattern is illus-trated within LG by a strong variability of pair-wise FST values between spatially closer demesdistant of 15 and 18 km with FST varyingfrom 001 to 006 respectively Thus it con-firmed that other independent factors than dis-tance alone must be taken into account in thedispersion of the tree frog such as roads urbanareas as well as natural obstacles and historicalfactors (Pellet et al 2004)
Overall and despite significant differencesobserved in term of genetic diversity betweenour two metapopulations these values are com-parable to genetic variation at microsatellite locifound in other amphibian species eg fromArens et al (2007) on Rana arvalis or fromScribner et al (2001) on Bufo bufo In additionour estimates of genetic variability Hs vary-ing from 049 to 066 are slightly in contrastwith the data from Andersen et al (2004 035-053) and Arens et al (2006 039-059) on Harborea metapopulations in Denmark and theNetherlands respectively or eg from Hitch-ings and Beebee (1997) on Rana temporariaand Newman and Squire (2001) on Rana syl-vatica where heterozygosity levels are lower
Mostly based on the same microsatellitemarkers genetic differentiation is in opposi-tion substantially higher within the Danish andDutch metapopulations (Andersen et al 2004Arens et al 2006) characterized by FST val-ues ranging from 003 to 033 (overall FST =022) and from 0 to 035 (overall = 019) re-spectively Several mutually non-exclusive hy-potheses could explain the substantial diffe-rence in genetic differentiation between our re-sults and the two available studies on H ar-borea (i) in contrast with the area studied byAndersen et al (2004) and Arens et al (2006)some natural structuring elements such as ripar-ian ecosystem exist in the LG metapopulationlandscape and such complementary terrestrialhabitats could act as potential dispersal corri-dors for H arborea (ii) geographical scale issmaller in our study (groups of ponds in Ander-sen et al (2004) are for instance separated byseveral km) (iii) the Danish and Dutch stud-ied areas being situated in the northern mar-gins of the distribution of the species phenom-ena as local drift and local extinction could bemore significant leading to an increase of ge-netic structure (iv) when sampling tadpoles po-tential sampling of siblings (individuals fromthe same clutches) may upwardly bias FST es-timates Although the occurrence of closely re-lated tadpoles could not be excluded our sam-
132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008
128 Short Notes
tors which undergo local extinctions (Grossen-bacher 1988 1994) This severe decline isthought to be mainly due to anthropogenic ac-tivities Studies carried out on the Europeantree frog in Sweden (Carlson and Edenhamn2000) as well as in Switzerland (Pellet et al2006) highlighted regular extinction and re-colonisation events characteristic of a metapop-ulation dynamics (Hanski and Gilpin 1997)Extensive theory has been developed to modelgenetic processes within metapopulation struc-tures Due to frequent extinctions and bottle-necks during recolonisations metapopulationsare likely to suffer more rapidly from inbreed-ing and fitness reduction than single large popu-
lations with the same total size (Gilpin 1991Hanski and Gilpin 1997)
In this study we sampled the two rem-nant metapopulations of the western portion ofSwitzerland (Dubey et al 2006) (i) the firstlies on the northern shore of the lake of Geneva(LG metapopulation fig 1) a region charac-terised by a very heterogeneous and mixed agri-cultural and urbanized landscape and present alow number of occupied ponds (less than 25Pellet et al 2002) (ii) the second is locatedon the southern bank of the lake of Neuchacirctel(LN metapopulation) which on the opposite isan uninterrupted landscape of suitable habitatsincluding marshes and wet meadows as well
Figure 1 Localisation of demes within the northern shore of the lake of Geneva (LG) and the southern bank of the lakeof Neuchacirctel (LN) with arrows corresponding to pairwise Fst values between demes within metapopulations (all valuessignificant at 005 level are indicated the thickness of arrows indicates the level of gene flow)
Short Notes 129
as woodland bordering the lake in a continuousway In the year 2000 40 calling ponds weredetected totalling several thousand frogs (Pel-let and Neet 2001) This area is considered asone of the largest metapopulation in Switzer-land (Grossenbacher 1988)
Information concerning the extent of popu-lation fragmentation is critical to determinewhether a species requires proactive manage-ment plans to reduce extinction risks associ-ated with genetic stochasticity Hence to es-timate the effect of habitat fragmentation onthe genetic structure and variability of tree frogpopulations we investigated the two isolatedmetapopulations in western Switzerland withsix microsatellite loci We hypothesized that inmetapopulations embedded in continuous habi-tats genetic structure would be lower while ge-netic variability and gene flow higher comparedto metapopulations characterized by lower den-sities and where heterogeneous habitat may actas a barrier to gene flow
During spring 2002 and 2003 a total of 235 samplesof Hyla arborea (tadpoles and eggs) were collected in 8ponds in the two studied areas (fig 1) Five ponds weresampled within the LG metapopulation (Allaman CampRomain Lavigny Mossiegraveres Vaudalle) and three pondsin the LN metapopulation (Chabrey Gletterens Trouville)When eggs were collected only one egg per clutch was usedfor genetic analyses
DNA extraction from tissues and egg samples was car-ried out using the QIAamp DNA Mini Kit (QIAGEN) orwith a CTAB protocol (Milligan 1992) Six microsatelliteloci isolated and characterized for Hyla arborea (Wha1-9 Wha1-20 Wha1-25 Wha1-103 Wha1-104 Wha1-140Arens et al 2000) were amplified and scored (see Arens etal (2000) for the specific PCR profiles) Amplified productswere genotyped with an ABI PRISM 377 DNA Sequencerusing genescan analysis 21 software (Applied Biosystems)
Gene diversities comprising observed (Ho) expected(Hs) within-deme (the populations will be treated as demesin the different sections of the manuscript) and expectedoverall heterozygosities (Ht) were estimated following (Neiand Chesser 1983) Genotypic disequilibrium betweenloci in each sample and deviations from Hardy-Weinbergequilibrium (HWE) within samples were tested based on2400 permutations and 10 000 randomizations respectivelyWrightrsquos fixation indices for within-deme deviation fromrandom mating (FIS) as well as pairwise deme differen-tiation (FST ) were estimated following Weir and Cocker-ham (1984) Deviation from random mating within demes(FIS) per locus and sample were computed with a boot-strap procedure (2000 randomizations) Statistical support
for pairwise deme differentiation was obtained through ex-act G-tests on allelic frequencies as described by Goudet etal (1996) with 2000 randomizations All summary statisticsand tests mentioned above have been computed using FSTAT
Version 2932 (Goudet 1995) Permutation tests were car-ried out in order to detect significant differences in allelicrichness expected (Hs) and observed (Ho) heterozygositiesand FST indices among the two studied metapopulations
Genetic isolation by distance at the metapopulationallevel and overall was tested by using a partial Mantel test(Mantel 1967) p-value were given after 10 000 randomiza-tions
A Bayesian model-based clustering method (Pritchardet al 2000) for inferring population structure and assign-ing individuals to populations was used as implemented instructure version 21 (Falush et al 2003) Based on allelefrequencies individuals are assigned through the use of aMarkov chain Monte Carlo (MCMC) simulation a member-ship coefficient for each of K populations We performed 10runs of 6 middot105 iterations (the first 105 considered as burn-in)for K = 1 to K = 10 (i) including all the populations and(ii) within the two metapopulations The number of popu-lations best fitting our data set was defined as described inEvanno et al (2005) The latter statistics compares the rateof change in the log probability of data between successiveK and the corresponding variance of log probabilities
We used the software migrate 206 (Beerli and Felsen-stein 2001 Beerli 2004) to estimate the scaled migrationrate (M) between demes within metapopulations This soft-ware is based on a coalescence model with mutation andmigration and estimates a measure of effective populationsize θ defined as 4Neμ where μ denotes mutation rateand migration M defined as mμ where m denotes migra-tion rate We assumed a stepwise mutation model and basedestimates on 15 short [104 Markov Chain Monte Carlo(MCMC) steps] and five long (105 MCMC steps) chainsTo ensure convergence we used the lsquoadaptive heatingrsquo op-tion with one lsquocoldrsquo and three lsquohotrsquo chains
Tests for HWE indicated that all loci testedwere at HWE and in genotypic equilibrium Forthe six microsatellite loci the number of allelesper locus ranged from 7 to 17 (average = 983)with a total of 59 alleles across 6 loci The al-lelic richness within deme ranges from 441 to529 with an overall mean of 625 (table 1) Ex-pected heterozygosities per locus within demes(Hs) ranged from 016 to 090 with an av-erage of 058 whereas expected overall het-erozygosity (Ht) averaged 062 (range per lo-cus 035-087) Observed heterozygosity (Ho)values varied from 044 to 068 with an aver-age of 051 (see table 1) There was a signifi-cant deviation from random mating in the ana-lyzed demes (overall FIS = 012 p gt 0001
130 Short Notes
LN FIS = 006 LG FIS = 016) suggest-ing the occurence of a within-sample sub-structure The genetic differentiation betweendemes (pairwise FST ) within each metapopu-lation was low ranging from 001 to 007 allvalues being significant except for the pairs ofdemes Mossiegravere-Allaman Vaudalle-Lavignyand Trouville-Chabrey (p gt 005) Compara-ble overall FST values were found in the LN andLG metapopulations (respectively 0046 95CI 0017-005 and 0039 95 CI 0025-0079
Table 1 Genetic diversities for eight Hyla arborea demesin two distinct metapopulations based on 6 microsatelliteloci N = maximum number of singers in the population in2002 (Pellet et al 2002) n = sample size Ho = observedheterozygosity Hs = expected heterozygosity AR = allelicrichness
Site N n Ho Hs AR
1 Allaman 20 20 045 054 4622 Camp romain 27 24 045 059 5303 Lavigny 149 24 044 049 5124 Mossiegraveres 25 12 047 051 5225 Vaudalle 11 20 044 055 524
Mean LG (1 to 5) 232 100 045 054 510
6 Chabrey 26 21 058 066 5297 Gletterens 11 19 063 060 4418 Trouville 99 22 068 063 496
Mean LN (6 to 8) 136 62 060 064 489
TotalTotal Mean (1 to 8) 368 162 051 058 625
p = 086 see fig 1) The pairwise FST val-ues among demes from separated metapopula-tions varies from 0086 to 0142 (mean = 011)for geographical distances ranging from 592 to697 km
No isolation by distance within the LGmetapopulation was detected (p = 040 r2 =009) indicating the absence of a pattern of ge-netic isolation with geographical distance (nottested on the three populations of LN) Signifi-cant differences between metapopulations wereobserved for Ho (LN Ho = 060 LG Ho =045 p = 0019) and Hs (LN Hs = 064 LGHs = 054 p = 0019) whereas allelic rich-ness (AR) FST and FIS were not significantlydifferent (respectively p = 043 p = 086 andp = 011 see table 1)
The analyses performed with structure re-vealed that the number of populations best fit-ting our data set is K = 2 No substructure wasrevealed within the metapopulations
Concerning the pairs of unidirectional migra-tion rates (M) estimated between demes withinmetapopulations 11 of the 13 in total wereasymmetric (ie where 95 CI did not overlaptable 2) In addition the analysis clearly showedthat recent migrations occurred between demeswithin both metapopulations
Table 2 Gene flow between populations within demes (M = mμ with 95 confidence interval)
Polulation (i) Allaman C romain Lavigny Mossiegraveres Vaudalle Chabrey Gletterens Trouvillerarr i rarr i rarr i rarr i rarr i rarr i rarr i rarr i
1 Allaman ndash 749 247 086 074642-867 209-290 063-112 058-091
2 Camp romain 255 ndash 087 126 042207-311 065-113 099-158 038-056
3 Lavigny 358 180 ndash 229 104300-424 130-241 191-271 086-125
4 Mossiegraveres 177 130 124 ndash 121137-223 088-182 097-155 101-144
5 Vaudalle 440 297 212 141 ndash375-512 232-373 176-252 112-175
6 Chabrey ndash 426 289362-498 250-333
7 Gletterens 225 ndash 049191-265 034-067
8 Trouville 139 367 ndash114-168 308-434
Short Notes 131
The main result stemming from this studyis the absence of substantial differences interms of genetic differentiation between the twometapopulations despite an important contrastin habitat fragmentation and demes densities asshown by (i) the low global structure observedwithin metapopulations (mean FST asymp 004)with pair-wise FST ranging from 001 to 007(ii) the absence of substructure within metapop-ulations as revealed by clustering analyses and(iii) the unidirectional indices of migration (M)observed between demes within metapopula-tions with some very high values observed (Mmax for LN 426 and for LG 749) Thereforeour data suggest a high overall rate of disper-sal within both metapopulations Though thepairs of unidirectional indices of migration (M)were mostly asymmetric within both metapop-ulations (11 of 13 pairs) with values varyingfrom 042 to 749 revealing that the demes con-tributed differently to the low structure observedbetween them Therefore the genetic diversityof demes could be reduced by the effect ofasymmetric gene flow
In contrast with the low structure observedwithin both areas the analyses revealed signif-icant (p = 0019) lower gene diversity (Hs)and observed heterozygosity (Ho) within LGmetapopulation as well as a higher but not sig-nificantly different substructure (FIS)
Consequently the results pointed out that thelandscape of LG is less suitable for the Euro-pean tree frog than that of LN and as a resultdispersal is less effective to maintain gene flowamong local demes and as consequence geneticdiversity within demes This pattern is illus-trated within LG by a strong variability of pair-wise FST values between spatially closer demesdistant of 15 and 18 km with FST varyingfrom 001 to 006 respectively Thus it con-firmed that other independent factors than dis-tance alone must be taken into account in thedispersion of the tree frog such as roads urbanareas as well as natural obstacles and historicalfactors (Pellet et al 2004)
Overall and despite significant differencesobserved in term of genetic diversity betweenour two metapopulations these values are com-parable to genetic variation at microsatellite locifound in other amphibian species eg fromArens et al (2007) on Rana arvalis or fromScribner et al (2001) on Bufo bufo In additionour estimates of genetic variability Hs vary-ing from 049 to 066 are slightly in contrastwith the data from Andersen et al (2004 035-053) and Arens et al (2006 039-059) on Harborea metapopulations in Denmark and theNetherlands respectively or eg from Hitch-ings and Beebee (1997) on Rana temporariaand Newman and Squire (2001) on Rana syl-vatica where heterozygosity levels are lower
Mostly based on the same microsatellitemarkers genetic differentiation is in opposi-tion substantially higher within the Danish andDutch metapopulations (Andersen et al 2004Arens et al 2006) characterized by FST val-ues ranging from 003 to 033 (overall FST =022) and from 0 to 035 (overall = 019) re-spectively Several mutually non-exclusive hy-potheses could explain the substantial diffe-rence in genetic differentiation between our re-sults and the two available studies on H ar-borea (i) in contrast with the area studied byAndersen et al (2004) and Arens et al (2006)some natural structuring elements such as ripar-ian ecosystem exist in the LG metapopulationlandscape and such complementary terrestrialhabitats could act as potential dispersal corri-dors for H arborea (ii) geographical scale issmaller in our study (groups of ponds in Ander-sen et al (2004) are for instance separated byseveral km) (iii) the Danish and Dutch stud-ied areas being situated in the northern mar-gins of the distribution of the species phenom-ena as local drift and local extinction could bemore significant leading to an increase of ge-netic structure (iv) when sampling tadpoles po-tential sampling of siblings (individuals fromthe same clutches) may upwardly bias FST es-timates Although the occurrence of closely re-lated tadpoles could not be excluded our sam-
132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008

Short Notes 129
as woodland bordering the lake in a continuousway In the year 2000 40 calling ponds weredetected totalling several thousand frogs (Pel-let and Neet 2001) This area is considered asone of the largest metapopulation in Switzer-land (Grossenbacher 1988)
Information concerning the extent of popu-lation fragmentation is critical to determinewhether a species requires proactive manage-ment plans to reduce extinction risks associ-ated with genetic stochasticity Hence to es-timate the effect of habitat fragmentation onthe genetic structure and variability of tree frogpopulations we investigated the two isolatedmetapopulations in western Switzerland withsix microsatellite loci We hypothesized that inmetapopulations embedded in continuous habi-tats genetic structure would be lower while ge-netic variability and gene flow higher comparedto metapopulations characterized by lower den-sities and where heterogeneous habitat may actas a barrier to gene flow
During spring 2002 and 2003 a total of 235 samplesof Hyla arborea (tadpoles and eggs) were collected in 8ponds in the two studied areas (fig 1) Five ponds weresampled within the LG metapopulation (Allaman CampRomain Lavigny Mossiegraveres Vaudalle) and three pondsin the LN metapopulation (Chabrey Gletterens Trouville)When eggs were collected only one egg per clutch was usedfor genetic analyses
DNA extraction from tissues and egg samples was car-ried out using the QIAamp DNA Mini Kit (QIAGEN) orwith a CTAB protocol (Milligan 1992) Six microsatelliteloci isolated and characterized for Hyla arborea (Wha1-9 Wha1-20 Wha1-25 Wha1-103 Wha1-104 Wha1-140Arens et al 2000) were amplified and scored (see Arens etal (2000) for the specific PCR profiles) Amplified productswere genotyped with an ABI PRISM 377 DNA Sequencerusing genescan analysis 21 software (Applied Biosystems)
Gene diversities comprising observed (Ho) expected(Hs) within-deme (the populations will be treated as demesin the different sections of the manuscript) and expectedoverall heterozygosities (Ht) were estimated following (Neiand Chesser 1983) Genotypic disequilibrium betweenloci in each sample and deviations from Hardy-Weinbergequilibrium (HWE) within samples were tested based on2400 permutations and 10 000 randomizations respectivelyWrightrsquos fixation indices for within-deme deviation fromrandom mating (FIS) as well as pairwise deme differen-tiation (FST ) were estimated following Weir and Cocker-ham (1984) Deviation from random mating within demes(FIS) per locus and sample were computed with a boot-strap procedure (2000 randomizations) Statistical support
for pairwise deme differentiation was obtained through ex-act G-tests on allelic frequencies as described by Goudet etal (1996) with 2000 randomizations All summary statisticsand tests mentioned above have been computed using FSTAT
Version 2932 (Goudet 1995) Permutation tests were car-ried out in order to detect significant differences in allelicrichness expected (Hs) and observed (Ho) heterozygositiesand FST indices among the two studied metapopulations
Genetic isolation by distance at the metapopulationallevel and overall was tested by using a partial Mantel test(Mantel 1967) p-value were given after 10 000 randomiza-tions
A Bayesian model-based clustering method (Pritchardet al 2000) for inferring population structure and assign-ing individuals to populations was used as implemented instructure version 21 (Falush et al 2003) Based on allelefrequencies individuals are assigned through the use of aMarkov chain Monte Carlo (MCMC) simulation a member-ship coefficient for each of K populations We performed 10runs of 6 middot105 iterations (the first 105 considered as burn-in)for K = 1 to K = 10 (i) including all the populations and(ii) within the two metapopulations The number of popu-lations best fitting our data set was defined as described inEvanno et al (2005) The latter statistics compares the rateof change in the log probability of data between successiveK and the corresponding variance of log probabilities
We used the software migrate 206 (Beerli and Felsen-stein 2001 Beerli 2004) to estimate the scaled migrationrate (M) between demes within metapopulations This soft-ware is based on a coalescence model with mutation andmigration and estimates a measure of effective populationsize θ defined as 4Neμ where μ denotes mutation rateand migration M defined as mμ where m denotes migra-tion rate We assumed a stepwise mutation model and basedestimates on 15 short [104 Markov Chain Monte Carlo(MCMC) steps] and five long (105 MCMC steps) chainsTo ensure convergence we used the lsquoadaptive heatingrsquo op-tion with one lsquocoldrsquo and three lsquohotrsquo chains
Tests for HWE indicated that all loci testedwere at HWE and in genotypic equilibrium Forthe six microsatellite loci the number of allelesper locus ranged from 7 to 17 (average = 983)with a total of 59 alleles across 6 loci The al-lelic richness within deme ranges from 441 to529 with an overall mean of 625 (table 1) Ex-pected heterozygosities per locus within demes(Hs) ranged from 016 to 090 with an av-erage of 058 whereas expected overall het-erozygosity (Ht) averaged 062 (range per lo-cus 035-087) Observed heterozygosity (Ho)values varied from 044 to 068 with an aver-age of 051 (see table 1) There was a signifi-cant deviation from random mating in the ana-lyzed demes (overall FIS = 012 p gt 0001
130 Short Notes
LN FIS = 006 LG FIS = 016) suggest-ing the occurence of a within-sample sub-structure The genetic differentiation betweendemes (pairwise FST ) within each metapopu-lation was low ranging from 001 to 007 allvalues being significant except for the pairs ofdemes Mossiegravere-Allaman Vaudalle-Lavignyand Trouville-Chabrey (p gt 005) Compara-ble overall FST values were found in the LN andLG metapopulations (respectively 0046 95CI 0017-005 and 0039 95 CI 0025-0079
Table 1 Genetic diversities for eight Hyla arborea demesin two distinct metapopulations based on 6 microsatelliteloci N = maximum number of singers in the population in2002 (Pellet et al 2002) n = sample size Ho = observedheterozygosity Hs = expected heterozygosity AR = allelicrichness
Site N n Ho Hs AR
1 Allaman 20 20 045 054 4622 Camp romain 27 24 045 059 5303 Lavigny 149 24 044 049 5124 Mossiegraveres 25 12 047 051 5225 Vaudalle 11 20 044 055 524
Mean LG (1 to 5) 232 100 045 054 510
6 Chabrey 26 21 058 066 5297 Gletterens 11 19 063 060 4418 Trouville 99 22 068 063 496
Mean LN (6 to 8) 136 62 060 064 489
TotalTotal Mean (1 to 8) 368 162 051 058 625
p = 086 see fig 1) The pairwise FST val-ues among demes from separated metapopula-tions varies from 0086 to 0142 (mean = 011)for geographical distances ranging from 592 to697 km
No isolation by distance within the LGmetapopulation was detected (p = 040 r2 =009) indicating the absence of a pattern of ge-netic isolation with geographical distance (nottested on the three populations of LN) Signifi-cant differences between metapopulations wereobserved for Ho (LN Ho = 060 LG Ho =045 p = 0019) and Hs (LN Hs = 064 LGHs = 054 p = 0019) whereas allelic rich-ness (AR) FST and FIS were not significantlydifferent (respectively p = 043 p = 086 andp = 011 see table 1)
The analyses performed with structure re-vealed that the number of populations best fit-ting our data set is K = 2 No substructure wasrevealed within the metapopulations
Concerning the pairs of unidirectional migra-tion rates (M) estimated between demes withinmetapopulations 11 of the 13 in total wereasymmetric (ie where 95 CI did not overlaptable 2) In addition the analysis clearly showedthat recent migrations occurred between demeswithin both metapopulations
Table 2 Gene flow between populations within demes (M = mμ with 95 confidence interval)
Polulation (i) Allaman C romain Lavigny Mossiegraveres Vaudalle Chabrey Gletterens Trouvillerarr i rarr i rarr i rarr i rarr i rarr i rarr i rarr i
1 Allaman ndash 749 247 086 074642-867 209-290 063-112 058-091
2 Camp romain 255 ndash 087 126 042207-311 065-113 099-158 038-056
3 Lavigny 358 180 ndash 229 104300-424 130-241 191-271 086-125
4 Mossiegraveres 177 130 124 ndash 121137-223 088-182 097-155 101-144
5 Vaudalle 440 297 212 141 ndash375-512 232-373 176-252 112-175
6 Chabrey ndash 426 289362-498 250-333
7 Gletterens 225 ndash 049191-265 034-067
8 Trouville 139 367 ndash114-168 308-434
Short Notes 131
The main result stemming from this studyis the absence of substantial differences interms of genetic differentiation between the twometapopulations despite an important contrastin habitat fragmentation and demes densities asshown by (i) the low global structure observedwithin metapopulations (mean FST asymp 004)with pair-wise FST ranging from 001 to 007(ii) the absence of substructure within metapop-ulations as revealed by clustering analyses and(iii) the unidirectional indices of migration (M)observed between demes within metapopula-tions with some very high values observed (Mmax for LN 426 and for LG 749) Thereforeour data suggest a high overall rate of disper-sal within both metapopulations Though thepairs of unidirectional indices of migration (M)were mostly asymmetric within both metapop-ulations (11 of 13 pairs) with values varyingfrom 042 to 749 revealing that the demes con-tributed differently to the low structure observedbetween them Therefore the genetic diversityof demes could be reduced by the effect ofasymmetric gene flow
In contrast with the low structure observedwithin both areas the analyses revealed signif-icant (p = 0019) lower gene diversity (Hs)and observed heterozygosity (Ho) within LGmetapopulation as well as a higher but not sig-nificantly different substructure (FIS)
Consequently the results pointed out that thelandscape of LG is less suitable for the Euro-pean tree frog than that of LN and as a resultdispersal is less effective to maintain gene flowamong local demes and as consequence geneticdiversity within demes This pattern is illus-trated within LG by a strong variability of pair-wise FST values between spatially closer demesdistant of 15 and 18 km with FST varyingfrom 001 to 006 respectively Thus it con-firmed that other independent factors than dis-tance alone must be taken into account in thedispersion of the tree frog such as roads urbanareas as well as natural obstacles and historicalfactors (Pellet et al 2004)
Overall and despite significant differencesobserved in term of genetic diversity betweenour two metapopulations these values are com-parable to genetic variation at microsatellite locifound in other amphibian species eg fromArens et al (2007) on Rana arvalis or fromScribner et al (2001) on Bufo bufo In additionour estimates of genetic variability Hs vary-ing from 049 to 066 are slightly in contrastwith the data from Andersen et al (2004 035-053) and Arens et al (2006 039-059) on Harborea metapopulations in Denmark and theNetherlands respectively or eg from Hitch-ings and Beebee (1997) on Rana temporariaand Newman and Squire (2001) on Rana syl-vatica where heterozygosity levels are lower
Mostly based on the same microsatellitemarkers genetic differentiation is in opposi-tion substantially higher within the Danish andDutch metapopulations (Andersen et al 2004Arens et al 2006) characterized by FST val-ues ranging from 003 to 033 (overall FST =022) and from 0 to 035 (overall = 019) re-spectively Several mutually non-exclusive hy-potheses could explain the substantial diffe-rence in genetic differentiation between our re-sults and the two available studies on H ar-borea (i) in contrast with the area studied byAndersen et al (2004) and Arens et al (2006)some natural structuring elements such as ripar-ian ecosystem exist in the LG metapopulationlandscape and such complementary terrestrialhabitats could act as potential dispersal corri-dors for H arborea (ii) geographical scale issmaller in our study (groups of ponds in Ander-sen et al (2004) are for instance separated byseveral km) (iii) the Danish and Dutch stud-ied areas being situated in the northern mar-gins of the distribution of the species phenom-ena as local drift and local extinction could bemore significant leading to an increase of ge-netic structure (iv) when sampling tadpoles po-tential sampling of siblings (individuals fromthe same clutches) may upwardly bias FST es-timates Although the occurrence of closely re-lated tadpoles could not be excluded our sam-
132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008
130 Short Notes
LN FIS = 006 LG FIS = 016) suggest-ing the occurence of a within-sample sub-structure The genetic differentiation betweendemes (pairwise FST ) within each metapopu-lation was low ranging from 001 to 007 allvalues being significant except for the pairs ofdemes Mossiegravere-Allaman Vaudalle-Lavignyand Trouville-Chabrey (p gt 005) Compara-ble overall FST values were found in the LN andLG metapopulations (respectively 0046 95CI 0017-005 and 0039 95 CI 0025-0079
Table 1 Genetic diversities for eight Hyla arborea demesin two distinct metapopulations based on 6 microsatelliteloci N = maximum number of singers in the population in2002 (Pellet et al 2002) n = sample size Ho = observedheterozygosity Hs = expected heterozygosity AR = allelicrichness
Site N n Ho Hs AR
1 Allaman 20 20 045 054 4622 Camp romain 27 24 045 059 5303 Lavigny 149 24 044 049 5124 Mossiegraveres 25 12 047 051 5225 Vaudalle 11 20 044 055 524
Mean LG (1 to 5) 232 100 045 054 510
6 Chabrey 26 21 058 066 5297 Gletterens 11 19 063 060 4418 Trouville 99 22 068 063 496
Mean LN (6 to 8) 136 62 060 064 489
TotalTotal Mean (1 to 8) 368 162 051 058 625
p = 086 see fig 1) The pairwise FST val-ues among demes from separated metapopula-tions varies from 0086 to 0142 (mean = 011)for geographical distances ranging from 592 to697 km
No isolation by distance within the LGmetapopulation was detected (p = 040 r2 =009) indicating the absence of a pattern of ge-netic isolation with geographical distance (nottested on the three populations of LN) Signifi-cant differences between metapopulations wereobserved for Ho (LN Ho = 060 LG Ho =045 p = 0019) and Hs (LN Hs = 064 LGHs = 054 p = 0019) whereas allelic rich-ness (AR) FST and FIS were not significantlydifferent (respectively p = 043 p = 086 andp = 011 see table 1)
The analyses performed with structure re-vealed that the number of populations best fit-ting our data set is K = 2 No substructure wasrevealed within the metapopulations
Concerning the pairs of unidirectional migra-tion rates (M) estimated between demes withinmetapopulations 11 of the 13 in total wereasymmetric (ie where 95 CI did not overlaptable 2) In addition the analysis clearly showedthat recent migrations occurred between demeswithin both metapopulations
Table 2 Gene flow between populations within demes (M = mμ with 95 confidence interval)
Polulation (i) Allaman C romain Lavigny Mossiegraveres Vaudalle Chabrey Gletterens Trouvillerarr i rarr i rarr i rarr i rarr i rarr i rarr i rarr i
1 Allaman ndash 749 247 086 074642-867 209-290 063-112 058-091
2 Camp romain 255 ndash 087 126 042207-311 065-113 099-158 038-056
3 Lavigny 358 180 ndash 229 104300-424 130-241 191-271 086-125
4 Mossiegraveres 177 130 124 ndash 121137-223 088-182 097-155 101-144
5 Vaudalle 440 297 212 141 ndash375-512 232-373 176-252 112-175
6 Chabrey ndash 426 289362-498 250-333
7 Gletterens 225 ndash 049191-265 034-067
8 Trouville 139 367 ndash114-168 308-434
Short Notes 131
The main result stemming from this studyis the absence of substantial differences interms of genetic differentiation between the twometapopulations despite an important contrastin habitat fragmentation and demes densities asshown by (i) the low global structure observedwithin metapopulations (mean FST asymp 004)with pair-wise FST ranging from 001 to 007(ii) the absence of substructure within metapop-ulations as revealed by clustering analyses and(iii) the unidirectional indices of migration (M)observed between demes within metapopula-tions with some very high values observed (Mmax for LN 426 and for LG 749) Thereforeour data suggest a high overall rate of disper-sal within both metapopulations Though thepairs of unidirectional indices of migration (M)were mostly asymmetric within both metapop-ulations (11 of 13 pairs) with values varyingfrom 042 to 749 revealing that the demes con-tributed differently to the low structure observedbetween them Therefore the genetic diversityof demes could be reduced by the effect ofasymmetric gene flow
In contrast with the low structure observedwithin both areas the analyses revealed signif-icant (p = 0019) lower gene diversity (Hs)and observed heterozygosity (Ho) within LGmetapopulation as well as a higher but not sig-nificantly different substructure (FIS)
Consequently the results pointed out that thelandscape of LG is less suitable for the Euro-pean tree frog than that of LN and as a resultdispersal is less effective to maintain gene flowamong local demes and as consequence geneticdiversity within demes This pattern is illus-trated within LG by a strong variability of pair-wise FST values between spatially closer demesdistant of 15 and 18 km with FST varyingfrom 001 to 006 respectively Thus it con-firmed that other independent factors than dis-tance alone must be taken into account in thedispersion of the tree frog such as roads urbanareas as well as natural obstacles and historicalfactors (Pellet et al 2004)
Overall and despite significant differencesobserved in term of genetic diversity betweenour two metapopulations these values are com-parable to genetic variation at microsatellite locifound in other amphibian species eg fromArens et al (2007) on Rana arvalis or fromScribner et al (2001) on Bufo bufo In additionour estimates of genetic variability Hs vary-ing from 049 to 066 are slightly in contrastwith the data from Andersen et al (2004 035-053) and Arens et al (2006 039-059) on Harborea metapopulations in Denmark and theNetherlands respectively or eg from Hitch-ings and Beebee (1997) on Rana temporariaand Newman and Squire (2001) on Rana syl-vatica where heterozygosity levels are lower
Mostly based on the same microsatellitemarkers genetic differentiation is in opposi-tion substantially higher within the Danish andDutch metapopulations (Andersen et al 2004Arens et al 2006) characterized by FST val-ues ranging from 003 to 033 (overall FST =022) and from 0 to 035 (overall = 019) re-spectively Several mutually non-exclusive hy-potheses could explain the substantial diffe-rence in genetic differentiation between our re-sults and the two available studies on H ar-borea (i) in contrast with the area studied byAndersen et al (2004) and Arens et al (2006)some natural structuring elements such as ripar-ian ecosystem exist in the LG metapopulationlandscape and such complementary terrestrialhabitats could act as potential dispersal corri-dors for H arborea (ii) geographical scale issmaller in our study (groups of ponds in Ander-sen et al (2004) are for instance separated byseveral km) (iii) the Danish and Dutch stud-ied areas being situated in the northern mar-gins of the distribution of the species phenom-ena as local drift and local extinction could bemore significant leading to an increase of ge-netic structure (iv) when sampling tadpoles po-tential sampling of siblings (individuals fromthe same clutches) may upwardly bias FST es-timates Although the occurrence of closely re-lated tadpoles could not be excluded our sam-
132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008

Short Notes 131
The main result stemming from this studyis the absence of substantial differences interms of genetic differentiation between the twometapopulations despite an important contrastin habitat fragmentation and demes densities asshown by (i) the low global structure observedwithin metapopulations (mean FST asymp 004)with pair-wise FST ranging from 001 to 007(ii) the absence of substructure within metapop-ulations as revealed by clustering analyses and(iii) the unidirectional indices of migration (M)observed between demes within metapopula-tions with some very high values observed (Mmax for LN 426 and for LG 749) Thereforeour data suggest a high overall rate of disper-sal within both metapopulations Though thepairs of unidirectional indices of migration (M)were mostly asymmetric within both metapop-ulations (11 of 13 pairs) with values varyingfrom 042 to 749 revealing that the demes con-tributed differently to the low structure observedbetween them Therefore the genetic diversityof demes could be reduced by the effect ofasymmetric gene flow
In contrast with the low structure observedwithin both areas the analyses revealed signif-icant (p = 0019) lower gene diversity (Hs)and observed heterozygosity (Ho) within LGmetapopulation as well as a higher but not sig-nificantly different substructure (FIS)
Consequently the results pointed out that thelandscape of LG is less suitable for the Euro-pean tree frog than that of LN and as a resultdispersal is less effective to maintain gene flowamong local demes and as consequence geneticdiversity within demes This pattern is illus-trated within LG by a strong variability of pair-wise FST values between spatially closer demesdistant of 15 and 18 km with FST varyingfrom 001 to 006 respectively Thus it con-firmed that other independent factors than dis-tance alone must be taken into account in thedispersion of the tree frog such as roads urbanareas as well as natural obstacles and historicalfactors (Pellet et al 2004)
Overall and despite significant differencesobserved in term of genetic diversity betweenour two metapopulations these values are com-parable to genetic variation at microsatellite locifound in other amphibian species eg fromArens et al (2007) on Rana arvalis or fromScribner et al (2001) on Bufo bufo In additionour estimates of genetic variability Hs vary-ing from 049 to 066 are slightly in contrastwith the data from Andersen et al (2004 035-053) and Arens et al (2006 039-059) on Harborea metapopulations in Denmark and theNetherlands respectively or eg from Hitch-ings and Beebee (1997) on Rana temporariaand Newman and Squire (2001) on Rana syl-vatica where heterozygosity levels are lower
Mostly based on the same microsatellitemarkers genetic differentiation is in opposi-tion substantially higher within the Danish andDutch metapopulations (Andersen et al 2004Arens et al 2006) characterized by FST val-ues ranging from 003 to 033 (overall FST =022) and from 0 to 035 (overall = 019) re-spectively Several mutually non-exclusive hy-potheses could explain the substantial diffe-rence in genetic differentiation between our re-sults and the two available studies on H ar-borea (i) in contrast with the area studied byAndersen et al (2004) and Arens et al (2006)some natural structuring elements such as ripar-ian ecosystem exist in the LG metapopulationlandscape and such complementary terrestrialhabitats could act as potential dispersal corri-dors for H arborea (ii) geographical scale issmaller in our study (groups of ponds in Ander-sen et al (2004) are for instance separated byseveral km) (iii) the Danish and Dutch stud-ied areas being situated in the northern mar-gins of the distribution of the species phenom-ena as local drift and local extinction could bemore significant leading to an increase of ge-netic structure (iv) when sampling tadpoles po-tential sampling of siblings (individuals fromthe same clutches) may upwardly bias FST es-timates Although the occurrence of closely re-lated tadpoles could not be excluded our sam-
132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008

132 Short Notes
pling was performed in order to minimize theirpresence On the contrary the Danish samplingpossibly included related individuals as fourdifferent tadpoles per clutch were analyzed (noinformation was given concerning the Dutchsampling strategy)
Overall our results suggest that even ifthe dispersal of H arborea among contiguousponds is high in areas of heterogeneous land-scape as shown in LG a loss of genetic diver-sity can occur Although no signs of inbreed-ing depression have been highlighted withinthe fragmented LG metapopulation of H ar-borea the small size of the demes coupledwith a lower genetic diversity compared to LNmetapopulation might increase the risk of de-mographic and genetic stochasticity Thus it islikely that the size and the level of connectiv-ity among demes is a crucial factor for the sur-vival of the metapopulations of Hyla arboreaConsequently in order to protect the Europeantree frog populations in an efficient way prior-ity should be given to conserving areas of suit-able habitat to promote population connectivityand maintain genetic diversity and evolutionarypotential In particular conservation manage-ment actions should be focused on (i) increas-ing the metapopulation sizes and the number ofdemes (ii) encourage the connectivity withinthe metapopulations In this context the rela-tively high migration potential of the Europeantree frog should permit a rapid natural coloni-sation of newly created ponds Although thetwo metapopulations have been separated foronly 20 years they are genetically well differ-entiated (mean FST between the two metapop-ulations 011) Consequently environment im-provements between them might be undertakenin order to reconnect each other and to recre-ate gene flow between both regions Since theEuropean tree frog is a mobile species the con-nection between the two metapopulations couldprobably be realised with the creation of onlya limited group of ponds with favourable sur-rounding habitats
Acknowledgements This work was partly funded by theOFEV (Office Feacutedeacuteral de lrsquoEnvironnement E Kohli) Weacknowledge N Duvoisin and N di Marco for help in thelaboratory The samples were collected with the autorisa-tion of the ldquoConservation de la Faune du Canton de Vaudrdquo(C Neet) and the KARCH (Centre de Coordination pourla protection des Amphibiens et des Reptiles de SuisseS Zumbach and J-C Monney) We also thank J-M Fi-vat J Goudet S Trouveacute and two anonymous referees forhelpful comments
References
Andersen LW Fog K Damgaard C (2004) Habitatfragmentation causes bottlenecks and inbreeding in theEuropean tree frog (Hyla arborea) P Roy Soc B-BiolSci 271 1293-1302
Arens P Bugter R vanrsquot Westende W Zollinger RStronks J Vos CC Smulders MJM (2006) Mi-crosatellite variation and population structure of a re-covering Tree frog (Hyla arborea L) metapopulationConserv Genet 7 825-834
Arens P van der Sluis T vanrsquot Westende WPC Vos-man B Vos CC Smulders MJM (2007) Geneticpopulation differentiation and connectivity among frag-mented Moor frog (Rana arvalis) populations in TheNetherlands Landscape Ecol 22 1489-1500
Arens P vanrsquot Westende W Bugter R SmuldersMJM Vosman B (2000) Microsatellite markers forthe European tree frog Hyla arborea Mol Ecol 91944-1946
Beerli P (2004) Migrate documentation and programpart of LAMARC Version 20 httpevolutiongswashingtonedulamarchtml
Beerli P Felsenstein J (2001) Maximum likelihood esti-mation of a migration matrix and effective populationssizes in n subspopulations by using a coalescent ap-proach Proc Natl Acad Sci USA 98 4563-4568
Carlson A Edenhamn P (2000) Extinction dynamics andthe regional persistence of a tree frog metapopulationP Roy Soc B-Biol Sci 267 1311-1313
Cushman SA (2006) Effects of habitat loss and fragmen-tation on amphibians a review and prospectus BiolCons 128 231-240
Dubey S Ursenbacher S Fumagalli L (2006) Origin oftree frog (Hyla spp) populations in western SwitzerlandRev Suisse Zool 113 879-887
Evanno G Regnaut S Goudet J (2005) Detecting thenumber of clusters of individuals using the softwarestructure a simulation study Mol Ecol 14 2611-2620
Falush D Stephens M Pritchard JK (2003) Inferenceof population structure using multilocus genotype datalinked loci and correlated allele frequencies Genetics164 1567-1587
Frankham R (2005) Genetics and extinction Biol Con-serv 126 131-140
Frankham R Ballou JD Briscoe DA (2002) Introduc-tion to Conservation Genetics Cambridge UK Cam-bridge University Press
Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008

Short Notes 133
Gasc J-P Cabela A Crnobrnja-Isailovic J Dolmen DGrossenbacher K Haffner P Lescure J Martens HMartiacutenez Rica JP Maurin H Oliveira ME Sofian-idou TS Veith M Zuiderwijk K (1997) Atlas ofAmphibians and Reptiles in Europe Paris Societas Eu-ropaea Herpetologica Museacuteum National drsquoHistoire Na-turelle
Gilpin ME (1991) The genetic effective size of ametapopulation In Metapopulation Dynamics Empir-ical and Theoretical Investigations Gilpin ME Han-ski I Eds London Academic Press
Goudet J (1995) FSTAT (Version 12) A computer pro-gram to calculate F-statistics J Hered 86 485-486
Goudet J Raymond M deMeeus T Rousset F (1996)Testing differentiation in diploid populations Genetics144 1933-1940
Grossenbacher K (1988) Verbreitungsatlas der Amphibiender Schweiz
Grossenbacher K (1994) Liste rouge des amphibiens men-aceacutes de Suisse In Liste rouge des espegraveces animalesmenaceacutees de Suisse p 97 Duelli P Ed Bern OFEFP
Hanski IA Gilpin ME (1997) Metapopulation BiologyEcology Genetics and Evolution London UK Acad-emic Press
Hitchings SP Beebee TJC (1997) Genetic substructur-ing as a result of barriers to gene flow in urban Ranatemporaria (common frog) populations implications forbiodiversity conservation Heredity 79 117-127
Mantel N (1967) Detection of disease clustering and ageneralized regression approach Cancer Res 27 209
Milligan B (1992) Plant DNA isolation In MolecularGenetic Analysis of Populations A Practical Approachp 59-88 Hoezel AR Ed IRL Press Oxford
Nei M Chesser RK (1983) Estimation of fixation in-dexes and gene diversities Ann Hum Genet 47 253-259
Newman RA Squire T (2001) Microsatellite variationand fine-scale population structure in the wood frog(Rana sylvatica) Mol Ecol 10 1087-1100
Pellet J Dubey S Hoehn S (2002) The amphibiansof the watershed of the Aubonne river (Switzerland)distribution and habitat analysis Bull Soc Vaud SciNat 88 41-57
Pellet J Guisan A Perrin N (2004) A concentric analy-sis of the impact of urbanization on the threatened Euro-pean tree frog in an agricultural landscape Cons Biol18 1599-1606
Pellet J Maze G Perrin N (2006) The contributionof patch topology and demographic parameters to PVApredictions the case of the European tree frog PopulEcol 48 353-361
Pellet J Neet C (2001) The European tree frog (Hylaarborea Anura) in the canton de Vaud present state ofknowledge Bull Soc Vaud Sci Nat 87 287-303
Pritchard JK Stephens M Donnelly P (2000) Inferenceof population structure using multilocus genotype dataGenetics 155 945-959
Scribner KT Arntzen JW Cruddace N Oldham RS(2001) Environmental correlates of toad abundance andpopulation genetic diversity Biol Cons 98 201-210
Wake W (1991) Species in danger Nature Canada 20 51Weir BS Cockerham CC (1984) Estimating F -statistics
for the analysis of population structure Evolution 381358-1370
Received January 20 2008 Accepted July 16 2008
Related Documents











