Cryptic diversity among Western Palearctic tree frogs: Postglacial range expansion, range limits, and secondary contacts of three European tree frog lineages (Hyla arborea group) Matthias Stöck a,b , Christophe Dufresnes a , Spartak N. Litvinchuk c , Petros Lymberakis d , Sébastien Biollay a , Matthieu Berroneau e , Amaël Borzée a , Karim Ghali a,f , Maria Ogielska g , Nicolas Perrin a,⇑ a University of Lausanne, Department of Ecology and Evolution (DEE), Biophore, CH-1015 Lausanne, Switzerland b Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), Müggelseedamm 301, D-12587 Berlin, Germany c Institute of Cytology, Russian Academy of Sciences, Tikhoretsky pr. 4, 194064 St. Petersburg, Russia d Natural History Museum of Crete, University of Crete, Knosos Av., P.O. Box 2208, 71409 Irakleio, Crete, Greece e Association Cistude Nature, Chemin du Moulinat, F-33185 Le Haillan, France f Vivarium Lausanne, Chemin de Boissonnet 82, CH-1010 Lausanne, Switzerland g Department of Evolutionary Biology and Conservation of Vertebrates, Wroclaw University, Sienkiewicza 21, 50-335 Wroclaw, Poland article info Article history: Received 24 February 2012 Revised 9 May 2012 Accepted 14 May 2012 Available online 29 May 2012 Keywords: Contact zone Divergence time Diversity Phylogeography Range expansion abstract We characterize divergence times, intraspecific diversity and distributions for recently recognized lin- eages within the Hyla arborea species group, based on mitochondrial and nuclear sequences from 160 localities spanning its whole distribution. Lineages of H. arborea, H. orientalis, H. molleri have at least Pli- ocene age, supporting species level divergence. The genetically uniform Iberian H. molleri, although lar- gely isolated by the Pyrenees, is parapatric to H. arborea, with evidence for successful hybridization in a small Aquitanian corridor (southwestern France), where the distribution also overlaps with H. meridio- nalis. The genetically uniform H. arborea, spread from Crete to Brittany, exhibits molecular signatures of a postglacial range expansion. It meets different mtDNA clades of H. orientalis in NE-Greece, along the Car- pathians, and in Poland along the Vistula River (there including hybridization). The East-European H. ori- entalis is strongly structured genetically. Five geographic mitochondrial clades are recognized, with a molecular signature of postglacial range expansions for the clade that reached the most northern lati- tudes. Hybridization with H. savignyi is suggested in southwestern Turkey. Thus, cryptic diversity in these Pliocene Hyla lineages covers three extremes: a genetically poor, quasi-Iberian endemic (H. molleri), a more uniform species distributed from the Balkans to Western Europe (H. arborea), and a well-structured Asia Minor-Eastern European species (H. orientalis). Ó 2012 Elsevier Inc. All rights reserved. 1. Introduction For many European vertebrate species including amphibians, phylogeographic hypotheses have been established in the last dec- ade (for review: Hewitt, 2011). While morphological or behavioral traits mark the boundaries of some species (e.g. Fijarczyk et al., 2011), the situation is less clear for sibling species and cryptic lin- eages, which are revealed only by the recent application of molec- ular markers (e.g. Stöck et al., 2006; Teacher et al., 2009; Hauswaldt et al., 2011; Recuero et al., 2012; Bisconti et al., 2011; Garcia-Porta et al., 2012). Western Palearctic tree frogs of the Hyla arborea group provide a good example (Faivovich et al., 2005; Smith et al., 2005; Wiens et al., 2005, 2010). Until recently, most European populations were considered to belong to a single spe- cies, H. arborea (e.g. Schneider and Grosse, 2009; http://www. iucnredlist.org/apps/redlist/details/10351/0), except for the Apen- nine Peninsula (plus Sardinia and Corsica), where H. intermedia (resp. H. sarda) had been assigned species status, confirmed by the lack of introgression at a contact zone with H. arborea (Verardi et al., 2009). A phylogenetic analysis based on 3200 bp of mito- chondrial and 860 bp of coding nuclear DNA (Stöck et al., 2008a) revealed this former, wide-ranging H. arborea to comprise three highly diverged lineages: H. arborea, occurring from Greece to northwestern France including Central Europe with the restricted type locality (Zurich; Dubois, 1996); H. molleri (previously consid- ered a subspecies of H. arborea), known from the Iberian Peninsula; and H. orientalis, ranging from Asia Minor to northeastern Europe, and not previously distinguished from H. arborea. Phylogenies based on mtDNA show that H. molleri and H. orientalis are as much diverged from H. arborea as is the recognized species H. intermedia, hence supporting a similar taxonomic status. 1055-7903/$ - see front matter Ó 2012 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.ympev.2012.05.014 ⇑ Corresponding author. Fax: +41 21 692 41 65. E-mail address: [email protected] (N. Perrin). Molecular Phylogenetics and Evolution 65 (2012) 1–9 Contents lists available at SciVerse ScienceDirect Molecular Phylogenetics and Evolution journal homepage: www.elsevier.com/locate/ympev

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular Phylogenetics and Evolution 65 (2012) 1–9
Contents lists available at SciVerse ScienceDirect
Molecular Phylogenetics and Evolution
journal homepage: www.elsevier .com/ locate /ympev
Cryptic diversity among Western Palearctic tree frogs: Postglacial rangeexpansion, range limits, and secondary contacts of three European tree froglineages (Hyla arborea group)
Matthias Stöck a,b, Christophe Dufresnes a, Spartak N. Litvinchuk c, Petros Lymberakis d, Sébastien Biollay a,Matthieu Berroneau e, Amaël Borzée a, Karim Ghali a,f, Maria Ogielska g, Nicolas Perrin a,⇑a University of Lausanne, Department of Ecology and Evolution (DEE), Biophore, CH-1015 Lausanne, Switzerlandb Leibniz-Institute of Freshwater Ecology and Inland Fisheries (IGB), Müggelseedamm 301, D-12587 Berlin, Germanyc Institute of Cytology, Russian Academy of Sciences, Tikhoretsky pr. 4, 194064 St. Petersburg, Russiad Natural History Museum of Crete, University of Crete, Knosos Av., P.O. Box 2208, 71409 Irakleio, Crete, Greecee Association Cistude Nature, Chemin du Moulinat, F-33185 Le Haillan, Francef Vivarium Lausanne, Chemin de Boissonnet 82, CH-1010 Lausanne, Switzerlandg Department of Evolutionary Biology and Conservation of Vertebrates, Wroclaw University, Sienkiewicza 21, 50-335 Wroclaw, Poland
a r t i c l e i n f o a b s t r a c t
Article history:Received 24 February 2012Revised 9 May 2012Accepted 14 May 2012Available online 29 May 2012
Keywords:Contact zoneDivergence timeDiversityPhylogeographyRange expansion
1055-7903/$ - see front matter � 2012 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.ympev.2012.05.014
⇑ Corresponding author. Fax: +41 21 692 41 65.E-mail address: [email protected] (N. Perrin).
We characterize divergence times, intraspecific diversity and distributions for recently recognized lin-eages within the Hyla arborea species group, based on mitochondrial and nuclear sequences from 160localities spanning its whole distribution. Lineages of H. arborea, H. orientalis, H. molleri have at least Pli-ocene age, supporting species level divergence. The genetically uniform Iberian H. molleri, although lar-gely isolated by the Pyrenees, is parapatric to H. arborea, with evidence for successful hybridization ina small Aquitanian corridor (southwestern France), where the distribution also overlaps with H. meridio-nalis. The genetically uniform H. arborea, spread from Crete to Brittany, exhibits molecular signatures of apostglacial range expansion. It meets different mtDNA clades of H. orientalis in NE-Greece, along the Car-pathians, and in Poland along the Vistula River (there including hybridization). The East-European H. ori-entalis is strongly structured genetically. Five geographic mitochondrial clades are recognized, with amolecular signature of postglacial range expansions for the clade that reached the most northern lati-tudes. Hybridization with H. savignyi is suggested in southwestern Turkey. Thus, cryptic diversity in thesePliocene Hyla lineages covers three extremes: a genetically poor, quasi-Iberian endemic (H. molleri), amore uniform species distributed from the Balkans to Western Europe (H. arborea), and a well-structuredAsia Minor-Eastern European species (H. orientalis).
� 2012 Elsevier Inc. All rights reserved.
1. Introduction
For many European vertebrate species including amphibians,phylogeographic hypotheses have been established in the last dec-ade (for review: Hewitt, 2011). While morphological or behavioraltraits mark the boundaries of some species (e.g. Fijarczyk et al.,2011), the situation is less clear for sibling species and cryptic lin-eages, which are revealed only by the recent application of molec-ular markers (e.g. Stöck et al., 2006; Teacher et al., 2009;Hauswaldt et al., 2011; Recuero et al., 2012; Bisconti et al., 2011;Garcia-Porta et al., 2012). Western Palearctic tree frogs of the Hylaarborea group provide a good example (Faivovich et al., 2005;Smith et al., 2005; Wiens et al., 2005, 2010). Until recently, mostEuropean populations were considered to belong to a single spe-
ll rights reserved.
cies, H. arborea (e.g. Schneider and Grosse, 2009; http://www.iucnredlist.org/apps/redlist/details/10351/0), except for the Apen-nine Peninsula (plus Sardinia and Corsica), where H. intermedia(resp. H. sarda) had been assigned species status, confirmed bythe lack of introgression at a contact zone with H. arborea (Verardiet al., 2009). A phylogenetic analysis based on 3200 bp of mito-chondrial and 860 bp of coding nuclear DNA (Stöck et al., 2008a)revealed this former, wide-ranging H. arborea to comprise threehighly diverged lineages: H. arborea, occurring from Greece tonorthwestern France including Central Europe with the restrictedtype locality (Zurich; Dubois, 1996); H. molleri (previously consid-ered a subspecies of H. arborea), known from the Iberian Peninsula;and H. orientalis, ranging from Asia Minor to northeastern Europe,and not previously distinguished from H. arborea. Phylogeniesbased on mtDNA show that H. molleri and H. orientalis are as muchdiverged from H. arborea as is the recognized species H. intermedia,hence supporting a similar taxonomic status.

2 M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9
Based on phylogeographic patterns of European anurans (Stöcket al., 2006; Hofman et al., 2007), and given the constraintsimposed by high altitudes and latitudes on the relatively thermo-philic European tree frogs (e.g. Schneider and Grosse, 2009), wehypothesized that the Carpathians, Alps and Pyrenees play majorroles in maintaining allopatry (leaving potential for secondarycontacts and hybridization in surrounding lowlands). The age ofhybridizing lineages experiencing secondary contact after Quater-nary separation may vary from Late Pleistocene to the LateMiocene (cf. Hewitt, 2011). In the context of our previous workon tree frogs (Stöck et al., 2008a), we further expect to find varyingamounts of geographic genetic structuring within these lineages,with distinctly lower genetic diversity in the northern regionsand higher endemism in the southern ones, which have had arelatively more stable climate since the Last Glacial Maximum(Sandel et al., 2011).
Given that the whole cytochrome b has been shown to be themost informative of several tested mitochondrial markers (Stöcket al., 2008a; Gvozdik et al., 2010), we used this mitochondrialand one nuclear intronic marker to delineate the ranges of thesethree species lineages, evaluate intraspecific diversity, estimatedivergence times, examine signatures of postglacial range expan-sions, and localize secondary contacts.
2. Methods
2.1. Amplification, cloning, and alignment of sequences
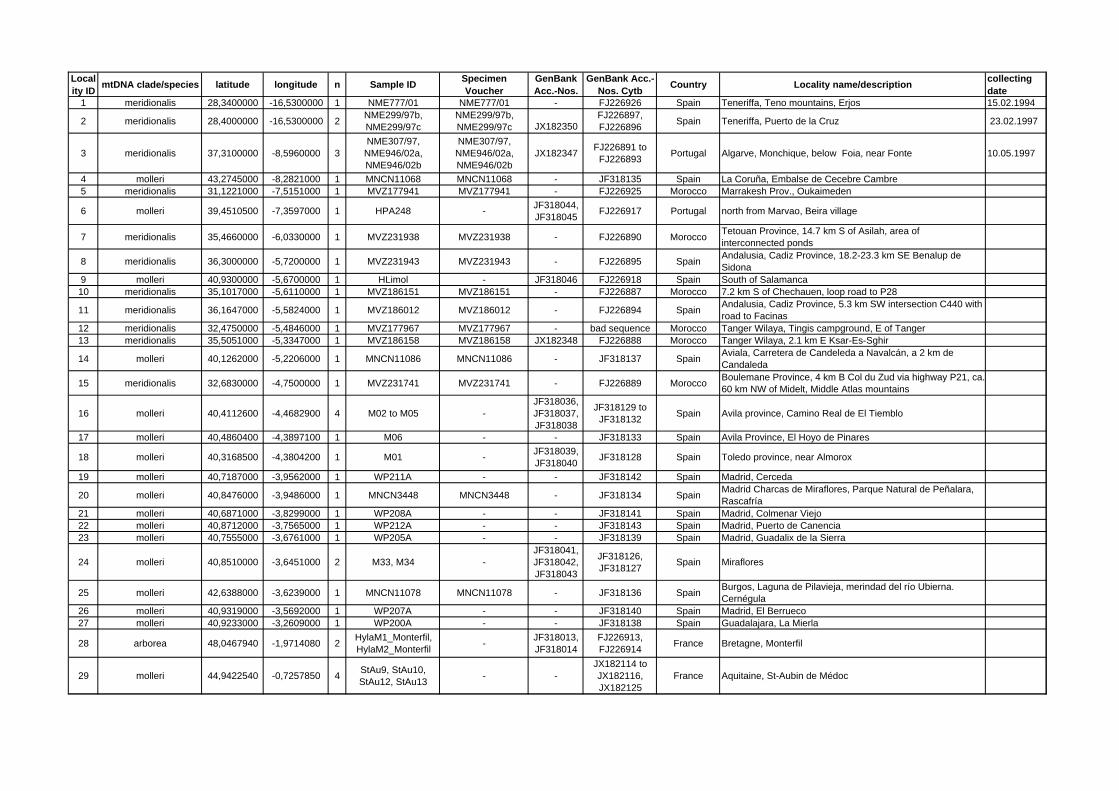
Samples of 462 frogs covering the whole European Hyla distri-bution (Fig. 1) were collected from living adults (buccal swabs),tadpoles (tail tips), or tissues from adult voucher individuals storedin scientific collections (Appendix S1). Buccal swabs were stored at�20 �C, tissue samples in 100% ethanol. DNA was extracted withQiagen DNeasy Tissue Kit or the BioSprint robotic workstation(Qiagen), eluted in a 200 ll Qiagen Buffer AE and stored at�18 �C. The mitochondrial cytochrome b (ca. 1 kb) was amplifiedwith primers L0 and H1046 as described (Stöck et al., 2008a). Toamplify ca. 545 bp of intron 1 of Fibrinogen A, alpha-polypeptide,
Fig. 1. Map with approximate range limits of Western Palearctic tree frogs (range limwww.iucnredlist.org/initiatives/amphibians) with sampling localities (see Map IDs in App‘‘Hyla meridionalis s.l.’’; H. intermedia and ‘‘new taxon 2’’ (Stöck et al., 2008a) as ‘‘Hyla iranges or deficiency of knowledge.
we used two primers (MVZ47: 59_AGTGAAAGATACAGTCACAGTGCTAGG_39; MVZ48: 59_GGAGGATATC-AGCACAGTCT-AAAAAG_39) and the following protocol: PCRs were performed in 12.5 llreactions containing 7.55 ll H2O, 1.25 ll of PCR buffer including1.5 mM MgCl2, 0.1 ll of dNTPs, 0.1 ll Taq QIAGEN, 0.75 ll of eachprimer having a concentration of 10 mM, and 2 ll of genomic DNAwith a concentration of 20 ng/ll. For subsequent cloning, two suchreactions from each individual were pooled to increase volume.The PCR protocol followed a ‘‘touch-up’’ approach with 10 cyclesof annealing temperatures (55–60 �C) increasing by 0.5� each cycle(with 30 s at 95 �C, 30 s at annealing temperature, and 45 s at72 �C), followed by 25 cycles with 30 s at 94 �C, 30 s at 56 �C, and45 s at 72 �C, and a final extension of 7 min at 72 �C. All PCR-prod-ucts (each clone of Fibrinogen; direct sequencing of PCR products ofcytochrome b) were sequenced in both directions, visualized on anABI 3730 sequencer, and aligned with Sequencher 4.9, followed bythe algorithms as implemented in Seaview (Gouy et al., 2010).
2.2. Phylogenetic analyses
In a first step, we reduced the total number of mtDNA se-quences to the number of haplotypes found at each locality. Max-imum likelihood (ML) phylogenies were generated with PhyML 3.0(Guindon et al., 2010) using the GTR model for cytochrome b andHKY model for the Fibrinogen alpha nuclear marker. For each case,we chose a BioNJ tree as a starting tree and used the combined sub-tree pruning and regrafting (SPR) plus nearest neighbor inter-change (NNI) options for tree improvement. All other parameterswere set as default (http://atgc.lirmm.fr/phyml/). Bootstrap valueswere based on 1000 (mtDNA) or 100 (nuDNA) resampled datasets.Bayesian phylogenetic analysis using the reported marker-specificsubstitution models was performed in MrBayes v3.1.0 (Ronquistand Huelsenbeck, 2003), with the default heating values for threeout of four chains, running 20 � 106 generations separately forthe mtDNA and nDNA datasets, with tree sampling every 1000generations. The ‘‘burnin’’-value was selected by visualizing thelog likelihoods associated with the posterior distribution of treesin the program Tracer (http://tree.bio.ed.ac.uk/software/tracer/).
its according to maps available through the Global Amphibian Assessment http://endix S1). Hyla meridionalis and ‘‘new taxon 1’’ (acc. Stöck et al., 2008a) are united as
ntermedia s.l.’’. Approximated range limits in interleaved colors indicate parapatric

H. intermedia s.l.
H. orientalis
H. savignyi
H. felixarabica
H. sarda
H. meridionalis s.l.
H. molleri
H. arborea
92/100
95/100
-/72 85/
100
98/100 100/100
99/100
100/100
100/100
100/100
100/100 99/99
87/100
91/100
94/100
94/100
100/100
99/100
-/75
M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9 3
All trees generated before the log likelihood curve flattened werediscarded.
2.3. Demographic analyses and estimates of divergence time
We used DnaSP v.5 (Librado and Rozas, 2009) to calculate andvisualize the distributions of observed and expected pairwisenucleotide site differences (‘mismatch distributions’), between allindividuals within the mtDNA clades of Hyla arborea, H. molleri,and subclades within H. orientalis, as well as the respective ex-pected values for growing populations (Librado and Rozas, 2009).We included only cytochrome b markers for which >904 bp 100%readable sequences were available (H. arborea: 86%, H. orientalis:94% H. molleri: 100%, total: 92%).
Divergence times to the most recent common ancestors wereestimated from the cytochrome b and Fibrinogen alpha markersindependently, assuming an uncorrelated exponential relaxedmolecular clock (BEAST v. 1.6; Drummond et al., 2006; http://beast.bio.ed.ac.uk/Main_Page). In the absence of appropriate fos-sils, we based our prior on results from previous work (Smithet al., 2005; Stöck et al., 2008a), assuming a normal distributionfor the divergence time between H. meridionalis and other treefrogs, with a mean of 10 millions of years ago (Mya) and standarddeviation of 1 My (thus effectively spanning a large range from 7.5to 12.5 Mya).
We applied the marker specific models of sequence evolution asdescribed for PhyML, and a Yule tree prior (constant speciation rateper lineage) as most appropriate for species-level divergences(Drummond et al., 2007). DNA cytochrome b data were analyzedboth with and without codon partition, with different partitionsfor codons 1 + 2 and 3.
Outgroup(H. japonica)
0.04 subst./site100/100
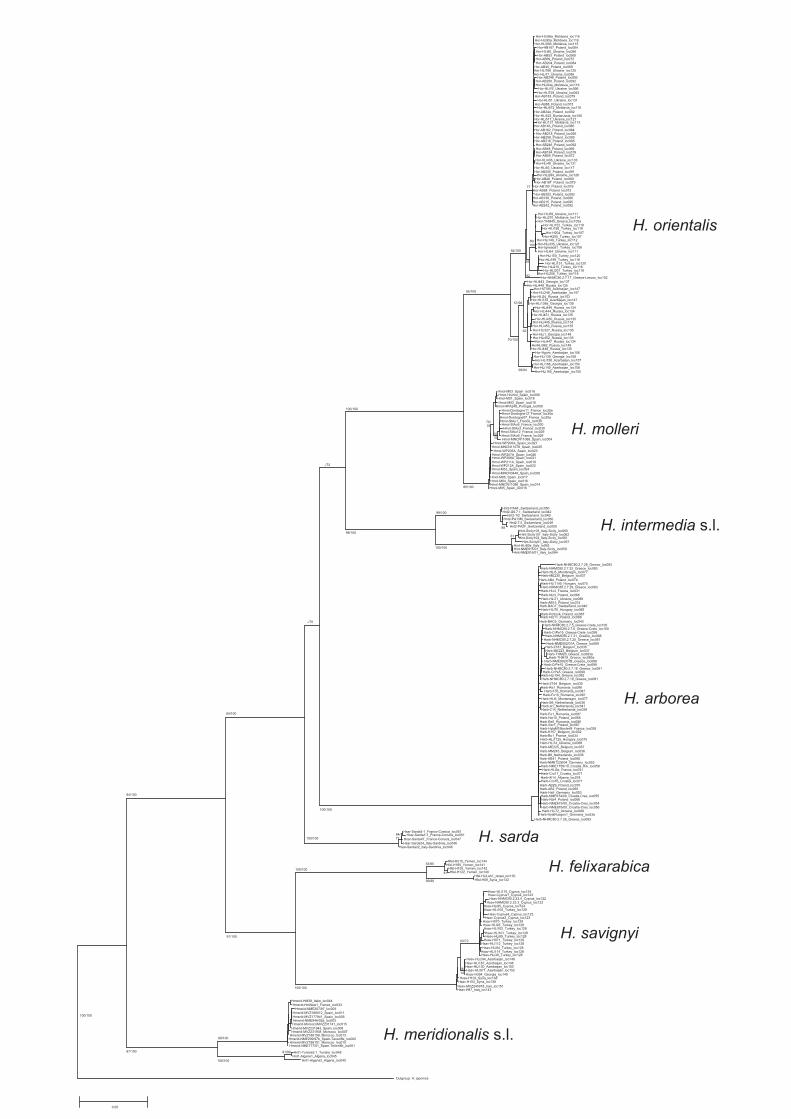
Fig. 2. Schematic maximum likelihood tree obtained with the program PhyMLbased on ca. 1 kb of mtDNA cytochrome b (a detailed version with all individuallabels is shown in Fig. S1) with bootstrap support values obtained from 1000resampled data sets (major nodes below 50% remained unlabeled; before ‘‘/’’),followed by Bayesian posterior support values (%) for major respective nodes (afterthe ‘‘/’’) from analysis using Mr. Bayes v3.1.0. Color codes of clades correspond tothose of localities in Fig. 1. (For interpretation of the references to color in this figurelegend, the reader is referred to the web version of this article.)
3. Results
Maximum likelihood and Bayesian phylogenetic analysesyielded mtDNA trees with congruent topologies (Figs. 2 and S1).The same clades were recovered for the nuDNA tree, but withmarkedly lower support (Fig. 3). Results turned out to be very ro-bust regarding partitioning. In the following, we focus on each ofthe three species lineages of the Hyla arborea group.
3.1. European tree frog (Hyla arborea)
This species ranges from the Western Balkan Peninsula acrossCentral into mainland Western Europe (Fig. 1), showing almostno genetic structure on the mitochondrial or the nuclear level(Figs. 2 and 3). Specifically, it occurs on Crete (locs. 94, 97–101),the Peloponnesus and mainland Greece (locs. 81–83, 89), alongthe eastern Adriatic coast (locs. 54–56, 58, 71, 77, 78), andthroughout the Eastern Pannonian Basin (Hungary, NE-Romania,W-Ukraine: locs. 75, 85–87), where it is separated from the easterntree frog (H. orientalis) by the Carpathian Arc. Hyla arborea is theonly tree-frog taxon occurring from central Poland (west of theVistula River: locs. 65–67, 70) throughout central (locs. 52 and53) to northwestern (locs. 34–41, 43) and western Europe (locs.28, 31, 32). The mtDNA mismatch distribution (Fig. 4a) showssignificantly high matching of simulated and observed curves(Table 1), pointing to a recent and rapid expansion.
3.2. Eastern tree frog (Hyla orientalis)
Our data show that this lineage, whose old name was resur-rected when molecular evidence showed its mitochondrial andnuclear divergence from H. arborea (Stöck et al., 2008a; Gvozdiket al., 2010), in fact represents a genetically very diverse and
well-differentiated group of lineages based on both mtDNA(Figs. 2 and 5a) and nuclear DNA (Fig. 3). Using mtDNA, wefound five well-supported subclades (Fig. 5): one in the TalyshRange (locs. 154–158), and a second well-structured one in theCaucasus and adjacent areas (locs. 134–139, 147–149, 153); forboth, mismatch distribution analyses (Figs. 5b and c) failed toreach significance (Table 1). Another well-structured mtDNA-group with two subclades inhabits western Asia Minor (locs.107, 118–120) and the western coast of the Black Sea (locs.108, 111, 112, 114, 121), without signs of recent demographicchanges (Table 1). Finally, a well-supported, widespread haplo-type clade with almost no substructure inhabits the Crimea,the northwestern coast of the Black Sea and the entire northeast-ern European region including Ukraine, Belarus, Russia, and Po-land, with the Vistula River as its approximate western border.For this latter group, demographic analyses revealed an almostperfect match of simulated to empirical data (Fig. 5e and Table 1),also pointing to a recent expansion.
The two groups of subclades based on mtDNA (i: Caucasus andTalysh vs. ii: Asia Minor and Black Sea, Eastern Europe and Crimea;Fig. 5a) are not entirely recovered based on nuDNA, where twoweakly supported subclusters (Fig. 3) unite Eastern European andTalysh with western Asia Minor frogs.
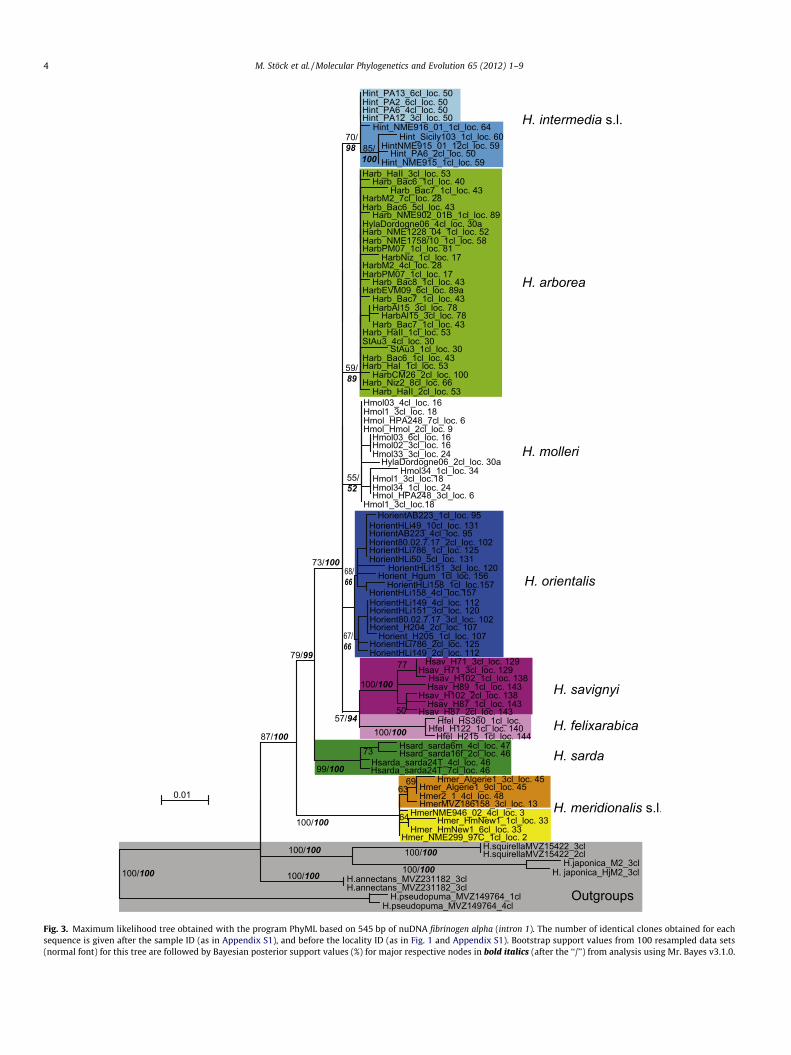
Fig. 3. Maximum likelihood tree obtained with the program PhyML based on 545 bp of nuDNA fibrinogen alpha (intron 1). The number of identical clones obtained for eachsequence is given after the sample ID (as in Appendix S1), and before the locality ID (as in Fig. 1 and Appendix S1). Bootstrap support values from 100 resampled data sets(normal font) for this tree are followed by Bayesian posterior support values (%) for major respective nodes in bold italics (after the ‘‘/’’) from analysis using Mr. Bayes v3.1.0.
4 M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9
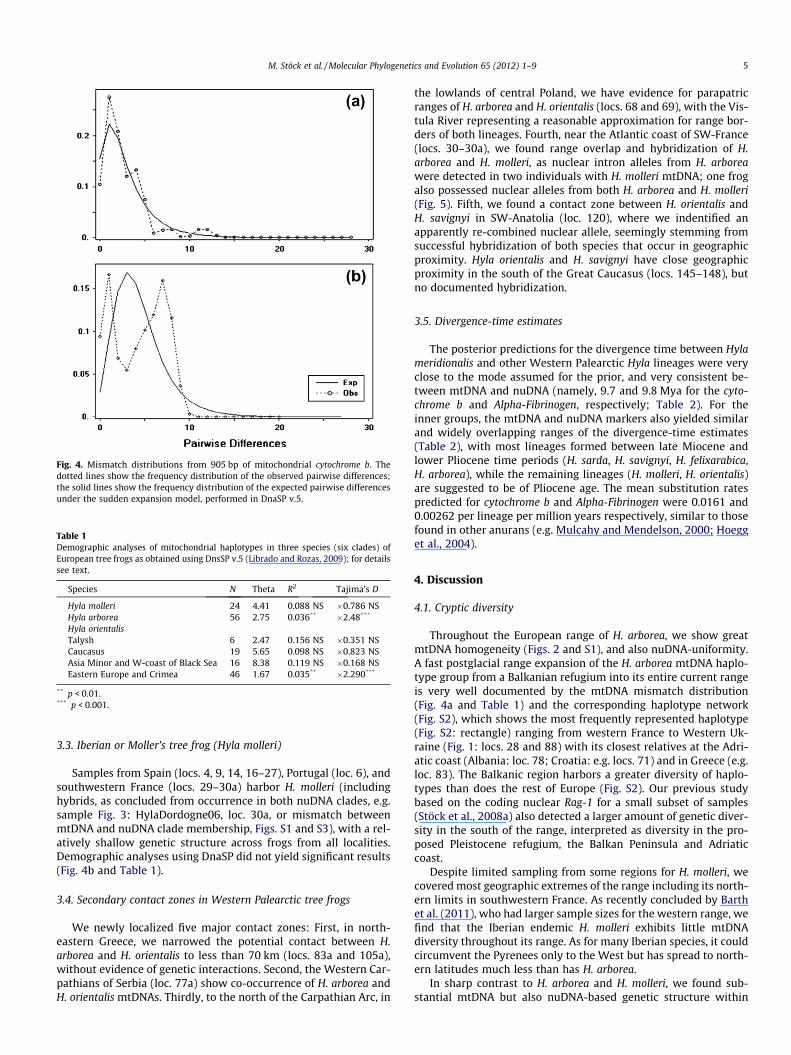
Fig. 4. Mismatch distributions from 905 bp of mitochondrial cytochrome b. Thedotted lines show the frequency distribution of the observed pairwise differences;the solid lines show the frequency distribution of the expected pairwise differencesunder the sudden expansion model, performed in DnaSP v.5.
Table 1Demographic analyses of mitochondrial haplotypes in three species (six clades) ofEuropean tree frogs as obtained using DnsSP v.5 (Librado and Rozas, 2009); for detailssee text.
Species N Theta R2 Tajima’s D
Hyla molleri 24 4.41 0.088 NS �0.786 NSHyla arborea 56 2.75 0.036** �2.48***
Hyla orientalisTalysh 6 2.47 0.156 NS �0.351 NSCaucasus 19 5.65 0.098 NS �0.823 NSAsia Minor and W-coast of Black Sea 16 8.38 0.119 NS �0.168 NSEastern Europe and Crimea 46 1.67 0.035** �2.290***
** p < 0.01.*** p < 0.001.
M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9 5
3.3. Iberian or Moller’s tree frog (Hyla molleri)
Samples from Spain (locs. 4, 9, 14, 16–27), Portugal (loc. 6), andsouthwestern France (locs. 29–30a) harbor H. molleri (includinghybrids, as concluded from occurrence in both nuDNA clades, e.g.sample Fig. 3: HylaDordogne06, loc. 30a, or mismatch betweenmtDNA and nuDNA clade membership, Figs. S1 and S3), with a rel-atively shallow genetic structure across frogs from all localities.Demographic analyses using DnaSP did not yield significant results(Fig. 4b and Table 1).
3.4. Secondary contact zones in Western Palearctic tree frogs
We newly localized five major contact zones: First, in north-eastern Greece, we narrowed the potential contact between H.arborea and H. orientalis to less than 70 km (locs. 83a and 105a),without evidence of genetic interactions. Second, the Western Car-pathians of Serbia (loc. 77a) show co-occurrence of H. arborea andH. orientalis mtDNAs. Thirdly, to the north of the Carpathian Arc, in
the lowlands of central Poland, we have evidence for parapatricranges of H. arborea and H. orientalis (locs. 68 and 69), with the Vis-tula River representing a reasonable approximation for range bor-ders of both lineages. Fourth, near the Atlantic coast of SW-France(locs. 30–30a), we found range overlap and hybridization of H.arborea and H. molleri, as nuclear intron alleles from H. arboreawere detected in two individuals with H. molleri mtDNA; one frogalso possessed nuclear alleles from both H. arborea and H. molleri(Fig. 5). Fifth, we found a contact zone between H. orientalis andH. savignyi in SW-Anatolia (loc. 120), where we indentified anapparently re-combined nuclear allele, seemingly stemming fromsuccessful hybridization of both species that occur in geographicproximity. Hyla orientalis and H. savignyi have close geographicproximity in the south of the Great Caucasus (locs. 145–148), butno documented hybridization.
3.5. Divergence-time estimates
The posterior predictions for the divergence time between Hylameridionalis and other Western Palearctic Hyla lineages were veryclose to the mode assumed for the prior, and very consistent be-tween mtDNA and nuDNA (namely, 9.7 and 9.8 Mya for the cyto-chrome b and Alpha-Fibrinogen, respectively; Table 2). For theinner groups, the mtDNA and nuDNA markers also yielded similarand widely overlapping ranges of the divergence-time estimates(Table 2), with most lineages formed between late Miocene andlower Pliocene time periods (H. sarda, H. savignyi, H. felixarabica,H. arborea), while the remaining lineages (H. molleri, H. orientalis)are suggested to be of Pliocene age. The mean substitution ratespredicted for cytochrome b and Alpha-Fibrinogen were 0.0161 and0.00262 per lineage per million years respectively, similar to thosefound in other anurans (e.g. Mulcahy and Mendelson, 2000; Hoegget al., 2004).
4. Discussion
4.1. Cryptic diversity
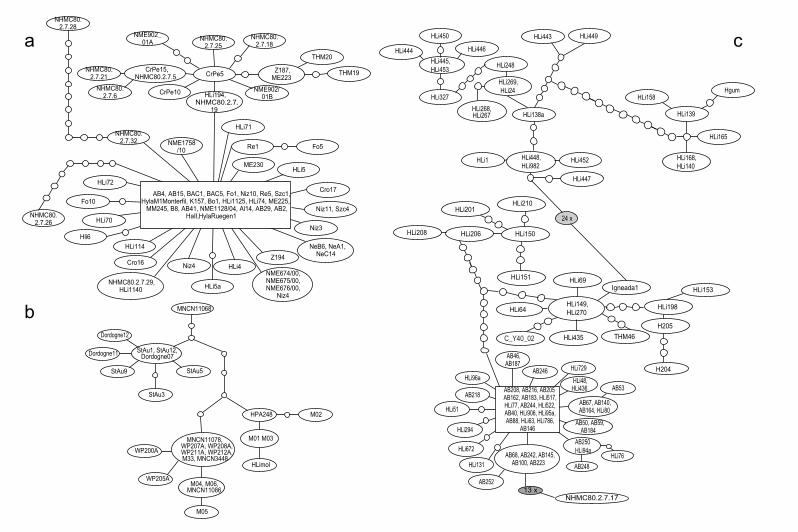
Throughout the European range of H. arborea, we show greatmtDNA homogeneity (Figs. 2 and S1), and also nuDNA-uniformity.A fast postglacial range expansion of the H. arborea mtDNA haplo-type group from a Balkanian refugium into its entire current rangeis very well documented by the mtDNA mismatch distribution(Fig. 4a and Table 1) and the corresponding haplotype network(Fig. S2), which shows the most frequently represented haplotype(Fig. S2: rectangle) ranging from western France to Western Uk-raine (Fig. 1: locs. 28 and 88) with its closest relatives at the Adri-atic coast (Albania: loc. 78; Croatia: e.g. locs. 71) and in Greece (e.g.loc. 83). The Balkanic region harbors a greater diversity of haplo-types than does the rest of Europe (Fig. S2). Our previous studybased on the coding nuclear Rag-1 for a small subset of samples(Stöck et al., 2008a) also detected a larger amount of genetic diver-sity in the south of the range, interpreted as diversity in the pro-posed Pleistocene refugium, the Balkan Peninsula and Adriaticcoast.
Despite limited sampling from some regions for H. molleri, wecovered most geographic extremes of the range including its north-ern limits in southwestern France. As recently concluded by Barthet al. (2011), who had larger sample sizes for the western range, wefind that the Iberian endemic H. molleri exhibits little mtDNAdiversity throughout its range. As for many Iberian species, it couldcircumvent the Pyrenees only to the West but has spread to north-ern latitudes much less than has H. arborea.
In sharp contrast to H. arborea and H. molleri, we found sub-stantial mtDNA but also nuDNA-based genetic structure within
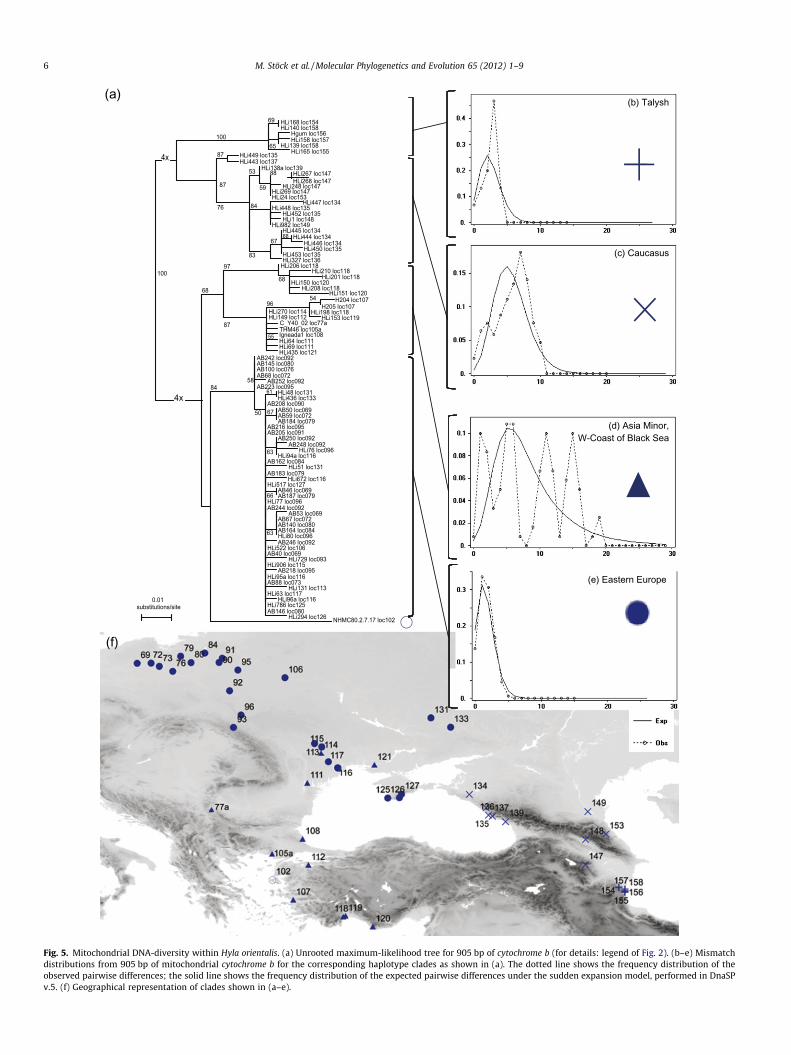
Fig. 5. Mitochondrial DNA-diversity within Hyla orientalis. (a) Unrooted maximum-likelihood tree for 905 bp of cytochrome b (for details: legend of Fig. 2). (b–e) Mismatchdistributions from 905 bp of mitochondrial cytochrome b for the corresponding haplotype clades as shown in (a). The dotted line shows the frequency distribution of theobserved pairwise differences; the solid line shows the frequency distribution of the expected pairwise differences under the sudden expansion model, performed in DnaSPv.5. (f) Geographical representation of clades shown in (a–e).
6 M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9

Table 2Divergence time (millions of years, My) to the most recent common ancestor (including stem), estimates based on the program BEAST using mitochondrial (cytochrome b; meanrate: 0.0161 per lineage per million years) and nuclear (Fibrinogen alpha; mean rate 0.00262 per lineage per million years) DNA sequences.
Species H. meridionalis H. sarda H. savignyi H. felixarabica H. arborea H. intermedia H. molleri H. orientalis
Marker mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA mtDNA nuDNA
My 9.7(7.7–11.6)
9.8(7.6–12.0)
6.1(4.3–8.3)
7.7(4.5–11.1)
6.2(4.3–8.4)
5.2(2.9–7.9)
6.2(4.3–8.4)
3.9(2.0–6.1)
6.1(4.3–8.1)
4.5(2.3–7.0)
6.1(4.5–8.2)
3.6(1.7–5.8)
3.7(2.6–5.0)
3.5(1.7–5.4)
3.7(2.6–5-5)
4.6(2.4–6.9)
M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9 7
the recently recognized Eastern tree frog H. orientalis (Figs. 3 and5). Much of H. orientalis’ diversity occurs in Asia Minor and sug-gests circum-Black Sea Pleistocene refugia. The clade thatpost-glacially colonized the northern latitudes shows highmtDNA-uniformity and significant signs of recent range expan-sion (Fig. 5e), similar to the signature across all of H. arborea’smtDNA (Fig. 4a). The reason that Gvozdik et al. (2010) foundall their ‘‘H. orientalis samples (to) form a compact cluster withsubstantial genetic variation, although without any deep diver-gences’’ appears to result from sampling only some Asia Minorand Caucasian regions.
4.2. Divergence times
The posterior predictions for the divergence time with H. merid-ionalis are extremely close to the mode assumed for the prior andvery consistent between cytochrome b and Alpha-Fibrinogen. Fur-thermore, the associated mean substitution rates are similar tothose found in other anurans (e.g. Mulcahy and Mendelson,2000; Hoegg et al., 2004), providing support for our calibration ofthe phylogenies with Hyla meridionalis
Despite previous estimates of divergence time for some Wes-tern Palearctic tree frog species (Canestrelli et al., 2007: H. interme-dia; Recuero et al., 2007: H. meridionalis), Gvozdik et al., 2010: H.orientalis, H. savignyi, H. felixarabica, H. meridionalis), our study isthe first that includes all extant species, and that uses mitochon-drial and nuclear sequence markers. As far as comparable (diver-gence of H. orientalis vs. H. savignyi + H. felixarabica) ourestimates are compatible, given the highest posterior densityinterval spanning ‘‘the period from the Early Pliocene throughthe Miocene, between 4.9 and 23.0 My’’ (Gvozdik et al., 2010).
Some of the discrepancies between our mtDNA and nuDNA-based estimates (Table 2) may be explicable by fewer data on thenuclear than on the mtNDA level for several clades. We confirmconsiderable divergences between two subclades of both ‘‘H.meridionalis s.l.’’ and ‘‘H. intermedia s.l.’’ (Fig. 2), as previouslyshown by other authors and markers (Recuero et al., 2007;Canestrelli et al., 2007), and temporarily called ‘‘new taxa1 and2’’ (Stöck et al., 2008a). More work is needed to understand poten-tial taxonomic implications for these lineages but is beyond thescope of this paper.
4.3. Contact and hybrid zones
The Eastern Mediterranean contains several major Pleistocenerefugia, with the territory of Greece representing a meeting zoneof faunal elements of Asia Minor and of Balkan Peninsular (plusAfrican) origin (Lymberakis and Poulakakis, 2010). WesternGreece, Crete and some western Aegean islands are colonized bythe mitochondrial lineage that also occurs on the western BalkanPeninsula and stretches into Central and even Western Europe(H. arborea), while the eastern Greek provinces of Macedonia,Thrace and the eastern Aegean islands are phylogenetically closeto the clade of Asia Minor origin (H. orientalis). Although we nar-rowed the potential contact to ca. 70 km (locs. 83a and 105a),our data are not sufficient yet to reveal potential contacts of
tree-frog lineages in northeastern Greece and the Aegean islands.To the north of Greece, the Carpathians represent a major barrierfor tree frogs. West of this mountain range occurs the H. arboreahaplotype group, and to the east of the Carpathian Arc that of H.orientalis, which also inhabits the entire rest of the Eastern Euro-pean Hyla range. Large Carpathian river valleys provide rare oppor-tunities for secondary contacts, with so far one locality of co-occurrence of both mtDNAs (loc. 77a). To the north of the Carpa-thian Arc, secondary contact and hybridization between H. arboreaand H. orientalis are documented by mtDNA and microsatellite datafrom the lowlands of Poland (Borzée, 2010, in prep.). Interestingly,the mtDNA subclade of H. orientalis that meets the uniform H. arbo-rea in Poland (Fig. 1 and 5) differs from the subclade (Figs. 1 and4d: triangles) that is in potential contact in Serbia and northeasternGreece. This offers interesting comparative research opportunitieson secondary contacts of differently, but quite closely relatedpopulations.
Since the splitting of H. molleri from H. arborea by Stöck et al.(2008a), occurrence and range limits at the Atlantic coast of SW-France, in the Aquitaine region, have been ambiguous with respectto species (see also Barth et al., 2011). Our new data (locs. 29–30a)not only revealed the only Western Palearctic region with three co-occurring tree frog taxa but also (at least) F1-hybridization be-tween H. molleri and H. arborea. As in the overlapping distributionsof H. meridionalis and H. molleri, in the Spanish Sistema CentralMountains, few hybridization events have been reported (Oliveiraet al., 1991; Barbadillo and Lapena, 2003); even genetic interac-tions between three species appear possible, but more research isrequired.
In addition to the three newly localized contact zones of H.arborea with H. orientalis (NE-Greece, Poland), and with H. molleri(SW-France), a well-known contact zone with H. intermedia existsin NE-Italy (Verardi et al., 2009), where neither hybrids nor back-crosses were identified, indicating a lack of current gene exchangebetween the two species. However, introgressed alleles appearedin both species, indicating past introgressive hybridization. Usingbioacoustic inference, pending genetic confirmation, Schneider(2001) narrowed the contact between H. orientalis (as ‘‘H. arborea’’)and H. savigny to less than 10 km in the Anamur plain of south-west Anatolia. Parapatry with one documented locality of hybrid-ization (Karkom, Israel) has been shown between H. savignyi andH. felixarabica (Gvozdik et al., 2010).
4.4. Comparisons with phylogeographic patterns of other terrestrialgroups
Our data contribute to knowledge of the evolutionary historyof Western Palearctic tree frogs as well as the comparative phy-logeography of Europe, and should improve conservation mea-sures. As recently noted by Rissler and Smith (2010) for NorthAmerica: ‘‘Identifying congruence in the geographical positionof lineage breaks and species range limits across multiple taxais a focus (. . .) of comparative phylogeography. These regionsare biogeographical hotspots for investigations into the pro-cesses driving divergence at multiple phylogenetic levels’’. In-deed, the postglacial colonization routes and resulting

8 M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9
secondary hybrid zones of tree frogs in the Western Palearcticscoincide with several of those known from other terrestrial spe-cies. Namely, the postglacial colonization route of H. arborearesembles that of the grasshopper Chorthippus parallelus, andthe advance of beech (Fagus sylvatica) and black alder (Alnusglutinosa) from their Balkanian refugia (King and Ferris, 1998;Hewitt, 1999, 2004; Magri, 2008), with the broad-leaf forestproviding direct summer habitats for tree frogs, suggesting par-tial co-colonization. As do H. arborea and H. molleri, these threespecies meet Iberian counterparts in the Pyrenees (Hewitt,1999) and form hybrid zones in their vicinity. Postglacial colo-nization of northeastern Europe to the east of the Carpathiansby H. orientalis resembles that by the green toad Bufo variabilis(Stöck et al., 2006, 2008b).
4.5. Implications for conservation of European tree frogs
Amphibians are undergoing a massive and extensive crisis(Wake and Vredenburg, 2008; Hoffmann et al., 2010), with com-plex causes that include land-use changes (Hof et al., 2011). Theremaining amphibian biodiversity should thus be especiallyassessed and protected in regions with industrial agriculture andintense land use and fragmentation (such as Western Europe) orcurrently facing major land-use changes due to political andeconomic transformations (such as Eastern Europe). While mostHyla species are still common in parts of their Western Palearcticrange, habitats are fragmented, and these frogs are in significantdecline over much of their Western European distribution(http://www.iucnredlist.org/apps/redlist/details/10351/0), mainlyby ‘‘loss of breeding habitats, habitat isolation, fragmentation,and pollution’’. Tree frogs are considered less threatened in EasternEurope (www.amhibiaweb.org, incl. refs.). However, land-usechanges caused by ongoing political and economic transformationpose upcoming threats also for the latter regions. Our data there-fore support conservation efforts by fine-tuning measured loca-tions of refugia harboring great genetic diversity (e.g. Moritz,2002), which are ‘‘essential refuges for Earth’s many small-rangedspecies’’ (Sandel et al., 2011). The localized areas of secondary con-tact should be considered ‘‘natural arenas to investigate processesdriving speciation’’ (Rissler and Smith, 2010), which require specialconservation efforts.
Acknowledgments
This work was supported by the Swiss National Science Foun-dation Grant 31003A-129894 to NP, the Fondation Agassiz andFondation du 450e from the University of Lausanne (Grant26076864 to CD), and a travel grant from the Swiss Academyof Sciences (SCNAT+) to AB and MSt; work of SNL was in partfunded by Grants RFBR.12-04-01277, NSh-3299.2010.4 andMCB-N22n. We thank A. Askanderov, O.S. Bezman-Moseyko, L.J.Borkin, G. Delaunay, G. Dzukic, W.-R. Grosse, J. Jaquiéry, V.I.Kazakov, O.V. Kukushkin, G.A. Lada, K.Y. Lotiev, I. Martinez-Solan-o, L.F. Mazanaeva, D.A. Melnikov, K.D. Milto, C. Muster, O. Pearl-son, I. Sas, U. Scheidt, Isa Schön, D.V. Shabanov, S. Shaytan, D.V.Skorinov, B. Wielstra, O.I. Zinenko, and R. Zollinger for providingsamples, Z. Dumas and R. Savary for help in the laboratory, H.Jourdan for range map information, and A. Ogrodowczyk for helpduring the fieldwork.
Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.ympev.2012.05.014.
References
Barbadillo, L.J., Lapena, M., 2003. Hibridacion natural de Hyla arborea Linnaeus,1758. e Hyla meridionalis Boettger, 1874) en la Peninsula Ibérica. Munibe 16,140–145.
Barth, A., Galan, P., Donaire, D., Gonzalez de la Vega, J.P., Pabijan, M., Vences, V.,2011. Mitochondrial uniformity in populations of the treefrog Hyla molleriacross the Iberian Peninsula. Amph. Rept. 32, 557–564.
Bisconti, R., Canestrelli, R., Colangelo, P., Nascetti, G., 2011. Multiple lines ofevidence for demographic and range expansion of a temperate species (Hylasarda) during the last glaciation. Mol. Ecol. 20, 5313–5327.
Borzée, A., 2010. Secondary Contacts of Two Tree Frog Taxa from the Hyla arboreagroup. Unpubl. Master Thesis, Behav., Evol., Conserv., School of Biol., Univ. ofLausanne.
Canestrelli, D., Cimmurata, R., Nascetti, G., 2007. Phylogeography and historicaldemography of the Italian treefrog, Hyla intermedia, reveals multiple refugia,population expansions and secondary contacts within peninsular Italy. Mol.Ecol. 16, 4808–4821.
Clement, M., Posada, D., Crandall, K., 2000. TCS: a computer program to estimategene genealogies. Mol. Ecol. 9, 1657–1660.
Drummond, A.J., Ho, S.Y.W., Phillips, M.J., Rambaut, A., 2006. Relaxed phylogeneticsand dating with confidence. PLoS Biol. 4, 699–710.
Drummond, A.J., Ho, S.Y.W., Rawlence, N., Rambaut, A., 2007. A Rough Guide toBEAST 1.4. <http://code.google.com/p/beast-mcmc/downloads/list>.
Dubois, A., 1996. The valid scientific name of the Italian treefrog, withcomments on the status of some early scientific names of Amphibia,Anura, and on some articles of the Code concerning secondary homonyms.Dumerilia 2, 55–71.
Faivovich, J., Haddad, C.F.B., Garcia, P.C.A., Frost, D.R., Campbell, J.A., Wheeler, W.C.,2005. Systematic review of the frog family Hylidae, with special reference toHylinae: phylogenetic analysis and taxonomic revision. Bull. Am. Mus. Nat. Hist.294, 1–240.
Fijarczyk, A., Nadachowska, K., Hofman, S., Litvinchuk, S.N., Babik, W., Stuglik, M.,Gollmann, G., Choleva, L.S., Cogalniceanu, D., Vukov, T., Dzukic, G., Szymura,J.M., 2011. Nuclear and mitochondrial phylogeography of the European fire-bellied toads Bombina bombina and Bombina variegata supports theirindependent histories. Mol. Ecol. 20, 3381–3398.
Garcia-Porta, J., Litvinchuk, S.N., Crochet, P.A., Romano, A., Geniez, P.H., Lo-Valvo, M.,Lymberakis, P., Carranza, S., 2012. Molecular phylogenetics and historicalbiogeography of the west-palearctic common toads (Bufo bufo speciescomplex). Mol. Phylogen. Evol. 63, 113–130.
Gouy, M., Guindon, S., Gascuel, O., 2010. SeaView version 4.0: a multiplatformgraphical user interface for sequence alignment and phylogenetic tree building.Mol. Phylogenet. Evol. 27, 221–224.
Guindon, S., Dufayard, J.F., Lefort, V., Anisimova, M., Hordijk, W., Gascuel, O., 2010.New algorithms and methods to estimate maximum-likelihood phylogenies:assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321.
Gvozdik, V., Moravec, J., Klütsch, C., Kotlik, P., 2010. Phylogeography of the MiddleEastern tree frogs Hyla, Hylidae, Amphibia) as inferred from nuclear andmitochondrial DNA variation, with a description of a new species. Mol.Phylogenet. Evol. 55, 1146–1166.
Hauswaldt, J.S., Angelini, C., Pollok, A., Steinfartz, S., 2011. Hybridization of twoancient salamander lineages: molecular evidence for endemic spectacledsalamanders on the Apennine Peninsula. J. Zool. 284, 248–256.
Hewitt, G.M., 1999. Post-glacial recolonization of European Biota. Biol. J. Linn. Soc.68, 87–112.
Hewitt, G.M., 2004. The structure of biodiversity – insights from molecularphylogeography. Front. Zool. 1, 4.
Hewitt, G.F., 2011. Quaternary phylogeography: the roots of hybrid zones. Genetica139, 617–638.
Hoegg, S., Vences, M., Brinkmann, H., Meyer, A., 2004. Phylogeny and comparativesubstitution rates of frogs inferred from sequences of three nuclear genes. Mol.Biol. Evol. 21, 1188–1200.
Hof, C., Arujo, M.B., Jetz, W., Rahbeck, C., 2011. Additive threats from pathogens,climate and land-use change for global amphibian diversity. Nature 480, 516–521.
Hoffmann, M., Hilton-Taylor, C., Angulo, A., Böhm, M., Brooks, T.M., et al., 2010. Theimpact of conservation on the status of the world’s vertebrates. Science 330,1503–1509.
Hofman, S., Spolsky, C., Uzzell, T., Cogalniceanu, D., Babik, W., Szymura, J.M., 2007.Phylogeography of the fire-bellied toads Bombina: independent Pleistocenehistories inferred from mitochondrial genomes. Mol. Ecol. 16, 2301–2316.
King, R.A., Ferris, C., 1998. Chloroplast DNA phylogeography of Alnus glutinosa (L.)Gaertn. Mol. Ecol. 7, 1151–1161.
Librado, P., Rozas, J., 2009. DnaSP v5: a software for comprehensive analysis of DNApolymorphism data. Bioinformatics 25, 1451–1452.
Lymberakis, P., Poulakakis, N., 2010. Three continents claiming an Archipelago: theevolution of Aegean’s herpetofaunal diversity. Diversity 2, 233–255.
Magri, D., 2008. Patterns of post-glacial spread and the extent of glacial refugia ofEuropean beech (Fagus sylvatica). J. Biogeogr. 35, 450–463.
Moritz, C., 2002. Strategies to protect biological diversity and the evolutionaryprocesses that sustain it. Syst. Biol. 51, 238–254.
Mulcahy, D.G., Mendelson, J.R., 2000. Phylogeography and speciation of themorphologically variable, widespread species Bufo valliceps, based onmolecular evidence from mtDNA. Mol. Phylogenet. Evol. 17, 173–189.

M. Stöck et al. / Molecular Phylogenetics and Evolution 65 (2012) 1–9 9
Oliveira, M.E., Paillette, M., Rosa, H.D., Crespo, H.D., 1991. A natural hybrid betweenHyla arborea and Hyla meridionalis detected by mating calls. [note that the taxonnamed ‘‘arborea’’ here is now H. molleri]. Amph. Rept. 12, 15–20.
Recuero, E., Iraola, A., Rubio, X., Machordom, A., Garcia-Paris, M., 2007.Mitochondrial differentiation and biogeography of Hyla meridionalis (Anura:Hylidae): an unusual phylogeographical pattern. J. Biogeogr. 34, 1207–1219.
Recuero, E., Canestrelli, D., Vörös, J., Szabo, J.K., Poyarkov, N.K., Arntzen, J.W.,Crnobrnja-Isailovic, J., Kidov, A.A., Cogalniceanu, D., Caputo, F.P., Nascetti, G.,Martinez-Solano, I., 2012. Multilocus species tree analyses resolve the radiationof the widespread Bufo bufo species group (Anura, Bufonidae). Mol. Phylogen.Evol. 62, 71–86.
Rissler, L., Smith, W.H., 2010. Mapping amphibian contact zones andphylogeographical break hotspots across the United States. Mol. Ecol. 19,5404–5416.
Ronquist, F., Huelsenbeck, J.P., 2003. MrBayes 3: Bayesian phylogenetic inferenceunder mixed models. Bioinformatics 19, 1572–1574.
Sandel, B., Arge, L., Dalsgaard, B., Davies, B.G., Gaston, K.J., Sutherland, W.J.,Svenning, J.-C., 2011. The influence of late Quaternary climate-change velocityon species endemism. Science 334, 660–664.
Schneider, H., 2001. The distribution of Hyla arborea and H. savignyi on the southcoast of Turkey. Zool. Midd. East 23, 61–69.
Schneider, H., Grosse, W.-R., 2009. Hyla arborea (Linnaeus, 1758) – EuropäischerLaubfrosch. In: Grossenbacher, K. (Ed.), Handbuch der Amphibien undReptilien Europas, Froschlurche II (Handbook of the Amphibians and Reptilesof Europe. vol. 5 Anura II), vol. 5. AULA-Verlag Wiesbaden, pp. 413–498 (inGerman).
Smith, S.A., Stephens, P.R., Wiens, J.J., 2005. Replicate patterns of species richness,historical biogeography and phylogeny in Holarctic tree frogs. Evolution 59,2433–2450.
Stöck, M., Moritz, C., Hickerson, M., Frynta, D., Dujsebayeva, T., Eremchenko, V.,Macey, J.R., Papenfuss, T.J., Wake, D.B., 2006. Evolution of mitochondrialrelationships and biogeography of Palearctic green toads (Bufo viridissubgroup) with insights in their genomic plasticity. Mol. Phylogenet. Evol. 41,663–689.
Stöck, M., Dubey, S., Klütsch, C., Litvinchuk, S.N., Scheidt, U., Perrin, N., 2008a.Mitochondrial and nuclear phylogeny of circum-Mediterranean tree frogs fromthe Hyla arborea group. Mol. Phylogenet. Evol. 49, 1019–1024.
Stöck M., Roth P., Podloucky R., Grossenbacher, K., 2008b. Wechselkröten – unterBerücksichtigung von Bufo viridis virdis Laurenti, 1768; Bufo variabilis Pallas,1769; Bufo boulengeri Lataste, 1879; Bufo balearicus Böttger, 1880 und Bufosiculus Stöck, Sicilia, Belfiore, Lo Brutto, Lo Valvo und Arculeo, 2008. In:Grossenbacher, K., (Ed.), Handbuch der Amphibien und Reptilien Europas.Froschlurche II (Handbook of the Amphibians and Reptiles of Europe. vol. 5Anura II), vol. 5. AULA-Verlag Wiesbaden, pp. 413–498 (in German).
Teacher, A.G.F., Garner, T.W.J., Nichols, R.A., 2009. European phylogeography of thecommon frog (Rana temporaria): routes of postglacial colonization into theBritish Isles, and evidence for an Irish glacial refugium. Heredity 102, 490–496.
Verardi, A., Canestrelli, D., Nascetti, G., 2009. Nuclear and mitochondrial patterns ofintrogression between the parapatric European treefrogs Hyla arborea and H.intermedia. Ann. Zool. Fenn. 46, 247–258.
Wake, D.B., Vredenburg, V., 2008. Are we in the midst of the sixth mass extinction?A view from the world of amphibians. Proc. Nat. Acad. Sci. USA 105, 11466–11473.
Wiens, J.J., Fetzner Jr., J.W., Parkinson, C.L., Reeder, T.W., 2005. Hylid frog phylogenyand sampling strategies for speciose clades. Syst. Biol. 54, 719–748.
Wiens, J.J., Kuczynski, C.A., Hua, X., Moen, D.S., 2010. An expanded phylogeny oftreefrogs (Hylidae) based on nuclear and mitochondrial sequence data. Mol.Phylogenet. Evol. 55, 871–882.
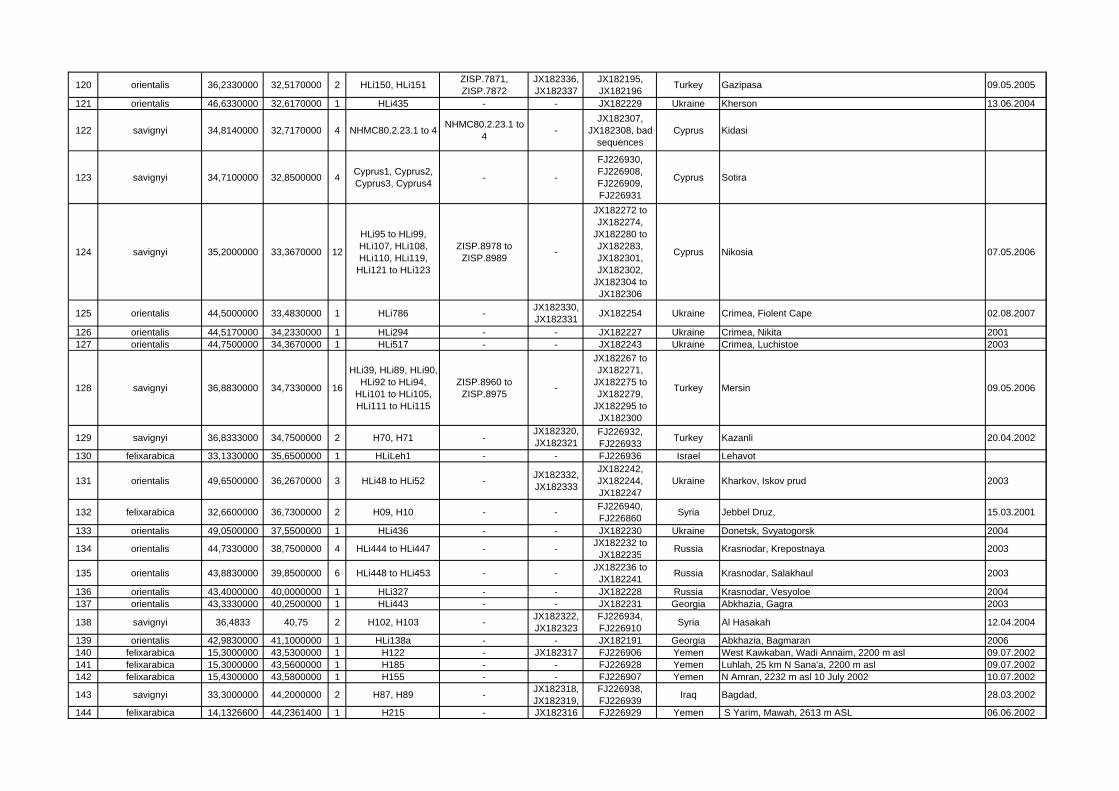
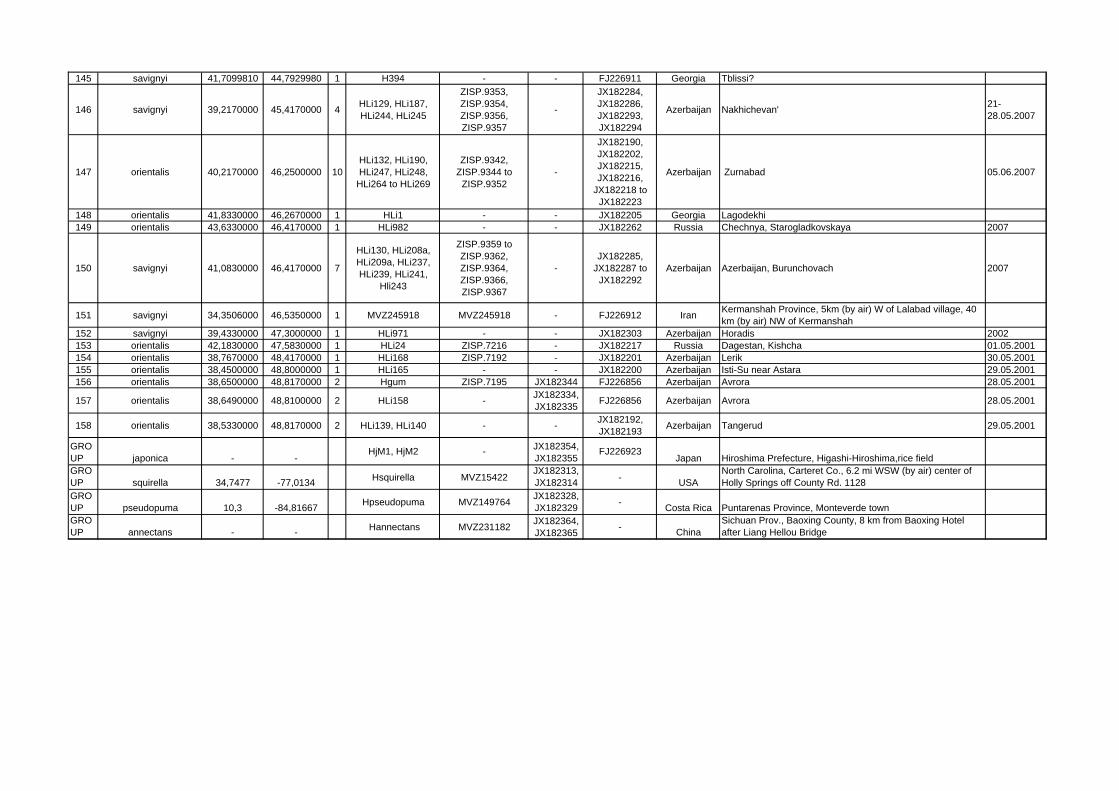
Locality ID mtDNA clade/species latitude longitude n Sample ID Specimen
VoucherGenBank Acc.-Nos.
GenBank Acc.-Nos. Cytb Country Locality name/description collecting
date1 meridionalis 28,3400000 -16,5300000 1 NME777/01 NME777/01 - FJ226926 Spain Teneriffa, Teno mountains, Erjos 15.02.1994
2 meridionalis 28,4000000 -16,5300000 2 NME299/97b, NME299/97c
NME299/97b, NME299/97c JX182350
FJ226897, FJ226896 Spain Teneriffa, Puerto de la Cruz 23.02.1997
3 meridionalis 37,3100000 -8,5960000 3NME307/97,
NME946/02a, NME946/02b
NME307/97, NME946/02a, NME946/02b
JX182347 FJ226891 to FJ226893 Portugal Algarve, Monchique, below Foia, near Fonte 10.05.1997
4 molleri 43,2745000 -8,2821000 1 MNCN11068 MNCN11068 - JF318135 Spain La Coruña, Embalse de Cecebre Cambre5 meridionalis 31,1221000 -7,5151000 1 MVZ177941 MVZ177941 - FJ226925 Morocco Marrakesh Prov., Oukaimeden
6 molleri 39,4510500 -7,3597000 1 HPA248 - JF318044, JF318045 FJ226917 Portugal north from Marvao, Beira village
7 meridionalis 35,4660000 -6,0330000 1 MVZ231938 MVZ231938 - FJ226890 Morocco Tetouan Province, 14.7 km S of Asilah, area of interconnected ponds
8 meridionalis 36,3000000 -5,7200000 1 MVZ231943 MVZ231943 - FJ226895 Spain Andalusia, Cadiz Province, 18.2-23.3 km SE Benalup de Sidona
9 molleri 40,9300000 -5,6700000 1 HLimol - JF318046 FJ226918 Spain South of Salamanca10 meridionalis 35,1017000 -5,6110000 1 MVZ186151 MVZ186151 - FJ226887 Morocco 7.2 km S of Chechauen, loop road to P28
11 meridionalis 36,1647000 -5,5824000 1 MVZ186012 MVZ186012 - FJ226894 Spain Andalusia, Cadiz Province, 5.3 km SW intersection C440 with road to Facinas
12 meridionalis 32,4750000 -5,4846000 1 MVZ177967 MVZ177967 - bad sequence Morocco Tanger Wilaya, Tingis campground, E of Tanger13 meridionalis 35,5051000 -5,3347000 1 MVZ186158 MVZ186158 JX182348 FJ226888 Morocco Tanger Wilaya, 2.1 km E Ksar-Es-Sghir
14 molleri 40,1262000 -5,2206000 1 MNCN11086 MNCN11086 - JF318137 Spain Aviala, Carretera de Candeleda a Navalcán, a 2 km de Candaleda
15 meridionalis 32,6830000 -4,7500000 1 MVZ231741 MVZ231741 - FJ226889 Morocco Boulemane Province, 4 km B Col du Zud via highway P21, ca.60 km NW of Midelt, Middle Atlas mountains
16 molleri 40,4112600 -4,4682900 4 M02 to M05 -JF318036, JF318037, JF318038
JF318129 to JF318132 Spain Avila province, Camino Real de El Tiemblo
17 molleri 40,4860400 -4,3897100 1 M06 - - JF318133 Spain Avila Province, El Hoyo de Pinares
18 molleri 40,3168500 -4,3804200 1 M01 - JF318039, JF318040 JF318128 Spain Toledo province, near Almorox
19 molleri 40,7187000 -3,9562000 1 WP211A - - JF318142 Spain Madrid, Cerceda
20 molleri 40,8476000 -3,9486000 1 MNCN3448 MNCN3448 - JF318134 Spain Madrid Charcas de Miraflores, Parque Natural de Peñalara, Rascafría
21 molleri 40,6871000 -3,8299000 1 WP208A - - JF318141 Spain Madrid, Colmenar Viejo22 molleri 40,8712000 -3,7565000 1 WP212A - - JF318143 Spain Madrid, Puerto de Canencia23 molleri 40,7555000 -3,6761000 1 WP205A - - JF318139 Spain Madrid, Guadalix de la Sierra
24 molleri 40,8510000 -3,6451000 2 M33, M34 -JF318041, JF318042, JF318043
JF318126, JF318127 Spain Miraflores
25 molleri 42,6388000 -3,6239000 1 MNCN11078 MNCN11078 - JF318136 Spain Burgos, Laguna de Pilavieja, merindad del río Ubierna. Cernégula
26 molleri 40,9319000 -3,5692000 1 WP207A - - JF318140 Spain Madrid, El Berrueco27 molleri 40,9233000 -3,2609000 1 WP200A - - JF318138 Spain Guadalajara, La Mierla
28 arborea 48,0467940 -1,9714080 2 HylaM1_Monterfil, HylaM2_Monterfil - JF318013,
JF318014FJ226913, FJ226914 France Bretagne, Monterfil
29 molleri 44,9422540 -0,7257850 4 StAu9, StAu10, StAu12, StAu13 - -
JX182114 to JX182116, JX182125
France Aquitaine, St-Aubin de Médoc
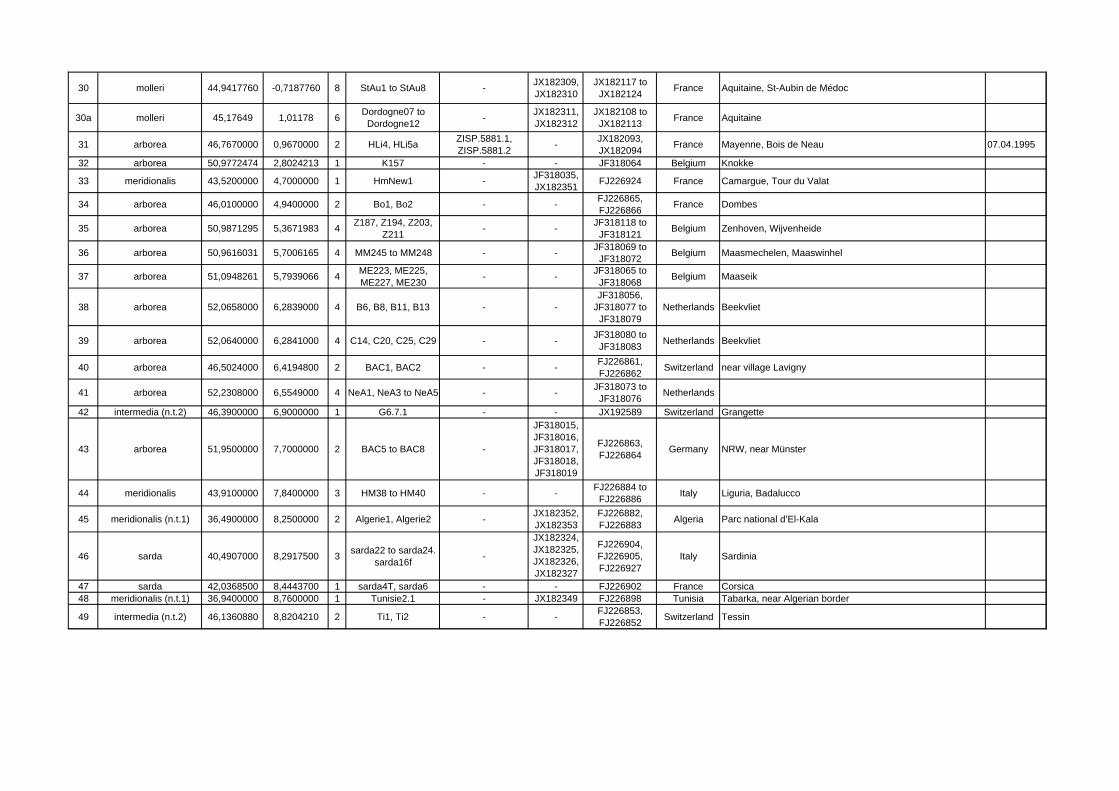
30 molleri 44,9417760 -0,7187760 8 StAu1 to StAu8 - JX182309, JX182310
JX182117 to JX182124 France Aquitaine, St-Aubin de Médoc
30a molleri 45,17649 1,01178 6 Dordogne07 to Dordogne12 - JX182311,
JX182312JX182108 to
JX182113 France Aquitaine
31 arborea 46,7670000 0,9670000 2 HLi4, HLi5a ZISP.5881.1, ZISP.5881.2 - JX182093,
JX182094 France Mayenne, Bois de Neau 07.04.1995
32 arborea 50,9772474 2,8024213 1 K157 - - JF318064 Belgium Knokke
33 meridionalis 43,5200000 4,7000000 1 HmNew1 - JF318035, JX182351 FJ226924 France Camargue, Tour du Valat
34 arborea 46,0100000 4,9400000 2 Bo1, Bo2 - - FJ226865, FJ226866 France Dombes
35 arborea 50,9871295 5,3671983 4 Z187, Z194, Z203, Z211 - - JF318118 to
JF318121 Belgium Zenhoven, Wijvenheide
36 arborea 50,9616031 5,7006165 4 MM245 to MM248 - - JF318069 to JF318072 Belgium Maasmechelen, Maaswinhel
37 arborea 51,0948261 5,7939066 4 ME223, ME225, ME227, ME230 - - JF318065 to
JF318068 Belgium Maaseik
38 arborea 52,0658000 6,2839000 4 B6, B8, B11, B13 - -JF318056,
JF318077 to JF318079
Netherlands Beekvliet
39 arborea 52,0640000 6,2841000 4 C14, C20, C25, C29 - - JF318080 to JF318083 Netherlands Beekvliet
40 arborea 46,5024000 6,4194800 2 BAC1, BAC2 - - FJ226861, FJ226862 Switzerland near village Lavigny
41 arborea 52,2308000 6,5549000 4 NeA1, NeA3 to NeA5 - - JF318073 to JF318076 Netherlands
42 intermedia (n.t.2) 46,3900000 6,9000000 1 G6.7.1 - - JX192589 Switzerland Grangette
43 arborea 51,9500000 7,7000000 2 BAC5 to BAC8 -
JF318015, JF318016, JF318017, JF318018, JF318019
FJ226863, FJ226864 Germany NRW, near Münster
44 meridionalis 43,9100000 7,8400000 3 HM38 to HM40 - - FJ226884 to FJ226886 Italy Liguria, Badalucco
45 meridionalis (n.t.1) 36,4900000 8,2500000 2 Algerie1, Algerie2 - JX182352, JX182353
FJ226882, FJ226883 Algeria Parc national d’El-Kala
46 sarda 40,4907000 8,2917500 3 sarda22 to sarda24. sarda16f -
JX182324, JX182325, JX182326, JX182327
FJ226904, FJ226905, FJ226927
Italy Sardinia
47 sarda 42,0368500 8,4443700 1 sarda4T, sarda6 - - FJ226902 France Corsica48 meridionalis (n.t.1) 36,9400000 8,7600000 1 Tunisie2.1 - JX182349 FJ226898 Tunisia Tabarka, near Algerian border
49 intermedia (n.t.2) 46,1360880 8,8204210 2 Ti1, Ti2 - - FJ226853, FJ226852 Switzerland Tessin
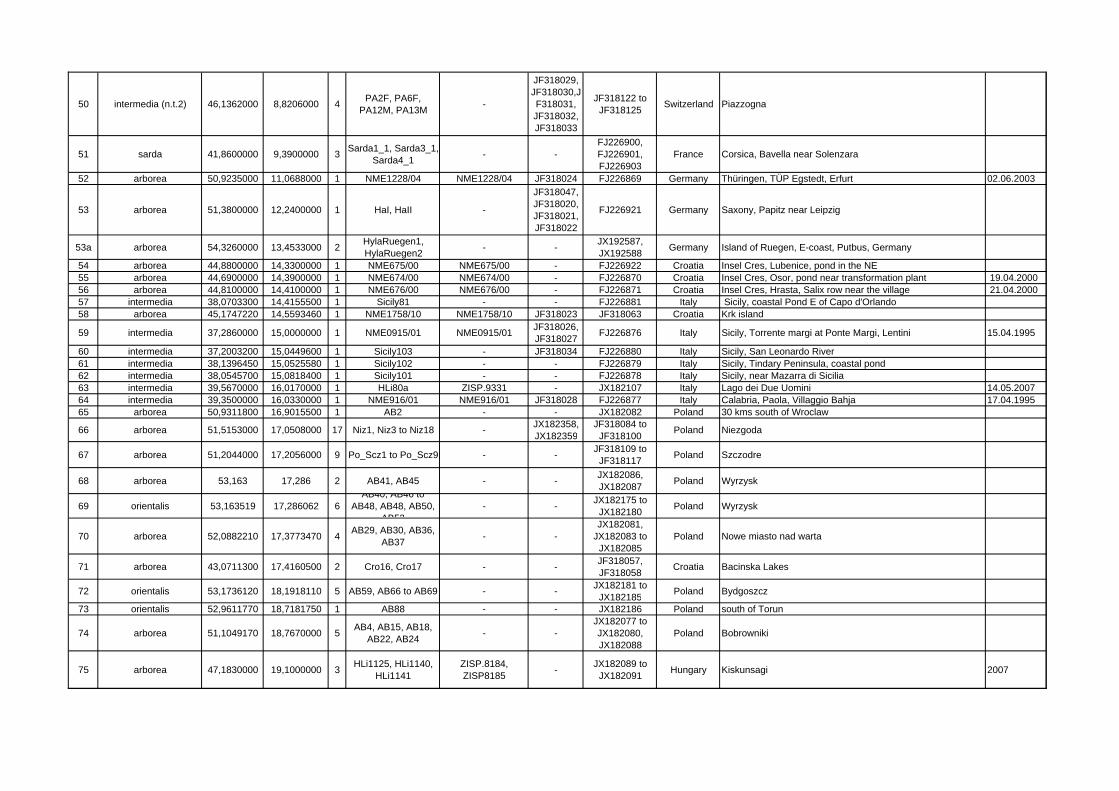
50 intermedia (n.t.2) 46,1362000 8,8206000 4 PA2F, PA6F, PA12M, PA13M -
JF318029, JF318030,JF318031, JF318032, JF318033
JF318122 to JF318125 Switzerland Piazzogna
51 sarda 41,8600000 9,3900000 3 Sarda1_1, Sarda3_1, Sarda4_1 - -
FJ226900, FJ226901, FJ226903
France Corsica, Bavella near Solenzara
52 arborea 50,9235000 11,0688000 1 NME1228/04 NME1228/04 JF318024 FJ226869 Germany Thüringen, TÜP Egstedt, Erfurt 02.06.2003
53 arborea 51,3800000 12,2400000 1 HaI, HaII -
JF318047, JF318020, JF318021, JF318022
FJ226921 Germany Saxony, Papitz near Leipzig
53a arborea 54,3260000 13,4533000 2 HylaRuegen1, HylaRuegen2 - - JX192587,
JX192588 Germany Island of Ruegen, E-coast, Putbus, Germany
54 arborea 44,8800000 14,3300000 1 NME675/00 NME675/00 - FJ226922 Croatia Insel Cres, Lubenice, pond in the NE55 arborea 44,6900000 14,3900000 1 NME674/00 NME674/00 - FJ226870 Croatia Insel Cres, Osor, pond near transformation plant 19.04.200056 arborea 44,8100000 14,4100000 1 NME676/00 NME676/00 - FJ226871 Croatia Insel Cres, Hrasta, Salix row near the village 21.04.200057 intermedia 38,0703300 14,4155500 1 Sicily81 - - FJ226881 Italy Sicily, coastal Pond E of Capo d'Orlando58 arborea 45,1747220 14,5593460 1 NME1758/10 NME1758/10 JF318023 JF318063 Croatia Krk island
59 intermedia 37,2860000 15,0000000 1 NME0915/01 NME0915/01 JF318026, JF318027 FJ226876 Italy Sicily, Torrente margi at Ponte Margi, Lentini 15.04.1995
60 intermedia 37,2003200 15,0449600 1 Sicily103 - JF318034 FJ226880 Italy Sicily, San Leonardo River61 intermedia 38,1396450 15,0525580 1 Sicily102 - - FJ226879 Italy Sicily, Tindary Peninsula, coastal pond62 intermedia 38,0545700 15,0818400 1 Sicily101 - - FJ226878 Italy Sicily, near Mazarra di Sicilia63 intermedia 39,5670000 16,0170000 1 HLi80a ZISP.9331 - JX182107 Italy Lago dei Due Uomini 14.05.200764 intermedia 39,3500000 16,0330000 1 NME916/01 NME916/01 JF318028 FJ226877 Italy Calabria, Paola, Villaggio Bahja 17.04.199565 arborea 50,9311800 16,9015500 1 AB2 - - JX182082 Poland 30 kms south of Wroclaw
66 arborea 51,5153000 17,0508000 17 Niz1, Niz3 to Niz18 - JX182358, JX182359
JF318084 to JF318100 Poland Niezgoda
67 arborea 51,2044000 17,2056000 9 Po_Scz1 to Po_Scz9 - - JF318109 to JF318117 Poland Szczodre
68 arborea 53,163 17,286 2 AB41, AB45 - - JX182086, JX182087 Poland Wyrzysk
69 orientalis 53,163519 17,286062 6AB40, AB46 to
AB48, AB48, AB50, AB53
- - JX182175 to JX182180 Poland Wyrzysk
70 arborea 52,0882210 17,3773470 4 AB29, AB30, AB36, AB37 - -
JX182081, JX182083 to
JX182085Poland Nowe miasto nad warta
71 arborea 43,0711300 17,4160500 2 Cro16, Cro17 - - JF318057, JF318058 Croatia Bacinska Lakes
72 orientalis 53,1736120 18,1918110 5 AB59, AB66 to AB69 - - JX182181 to JX182185 Poland Bydgoszcz
73 orientalis 52,9611770 18,7181750 1 AB88 - - JX182186 Poland south of Torun
74 arborea 51,1049170 18,7670000 5 AB4, AB15, AB18, AB22, AB24 - -
JX182077 to JX182080, JX182088
Poland Bobrowniki
75 arborea 47,1830000 19,1000000 3 HLi1125, HLi1140, HLi1141
ZISP.8184, ZISP8185 - JX182089 to
JX182091 Hungary Kiskunsagi 2007
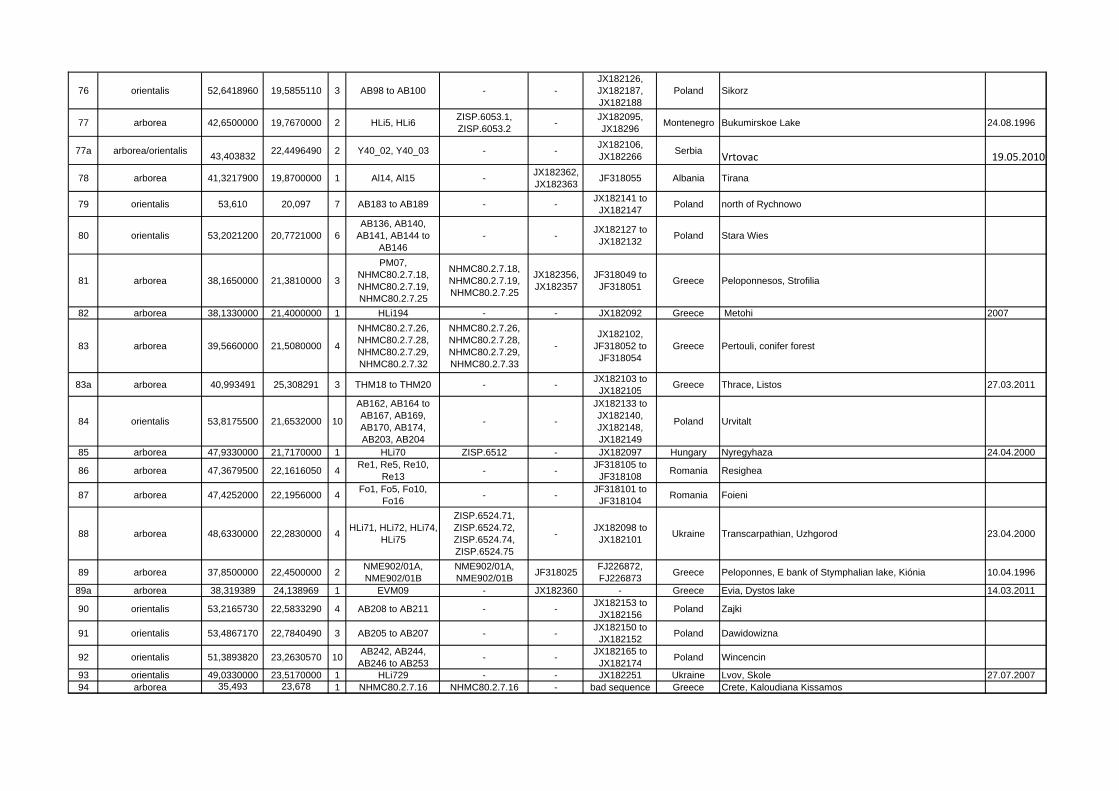
76 orientalis 52,6418960 19,5855110 3 AB98 to AB100 - -JX182126, JX182187, JX182188
Poland Sikorz
77 arborea 42,6500000 19,7670000 2 HLi5, HLi6 ZISP.6053.1, ZISP.6053.2 - JX182095,
JX18296 Montenegro Bukumirskoe Lake 24.08.1996
77a arborea/orientalis 43,403832 22,4496490 2 Y40_02, Y40_03 - - JX182106, JX182266 Serbia
Vrtovac 19.05.2010
78 arborea 41,3217900 19,8700000 1 Al14, Al15 - JX182362, JX182363 JF318055 Albania Tirana
79 orientalis 53,610 20,097 7 AB183 to AB189 - - JX182141 to JX182147 Poland north of Rychnowo
80 orientalis 53,2021200 20,7721000 6AB136, AB140,
AB141, AB144 to AB146
- - JX182127 to JX182132 Poland Stara Wies
81 arborea 38,1650000 21,3810000 3
PM07, NHMC80.2.7.18, NHMC80.2.7.19, NHMC80.2.7.25
NHMC80.2.7.18, NHMC80.2.7.19, NHMC80.2.7.25
JX182356, JX182357
JF318049 to JF318051 Greece Peloponnesos, Strofilia
82 arborea 38,1330000 21,4000000 1 HLi194 - - JX182092 Greece Metohi 2007
83 arborea 39,5660000 21,5080000 4
NHMC80.2.7.26, NHMC80.2.7.28, NHMC80.2.7.29, NHMC80.2.7.32
NHMC80.2.7.26, NHMC80.2.7.28, NHMC80.2.7.29, NHMC80.2.7.33
-JX182102,
JF318052 to JF318054
Greece Pertouli, conifer forest
83a arborea 40,993491 25,308291 3 THM18 to THM20 - - JX182103 to JX182105 Greece Thrace, Listos 27.03.2011
84 orientalis 53,8175500 21,6532000 10
AB162, AB164 to AB167, AB169, AB170, AB174, AB203, AB204
- -
JX182133 to JX182140, JX182148, JX182149
Poland Urvitalt
85 arborea 47,9330000 21,7170000 1 HLi70 ZISP.6512 - JX182097 Hungary Nyregyhaza 24.04.2000
86 arborea 47,3679500 22,1616050 4 Re1, Re5, Re10, Re13 - - JF318105 to
JF318108 Romania Resighea
87 arborea 47,4252000 22,1956000 4 Fo1, Fo5, Fo10, Fo16 - - JF318101 to
JF318104 Romania Foieni
88 arborea 48,6330000 22,2830000 4 HLi71, HLi72, HLi74, HLi75
ZISP.6524.71, ZISP.6524.72, ZISP.6524.74, ZISP.6524.75
- JX182098 to JX182101 Ukraine Transcarpathian, Uzhgorod 23.04.2000
89 arborea 37,8500000 22,4500000 2 NME902/01A, NME902/01B
NME902/01A, NME902/01B JF318025 FJ226872,
FJ226873 Greece Peloponnes, E bank of Stymphalian lake, Kiónia 10.04.1996
89a arborea 38,319389 24,138969 1 EVM09 - JX182360 - Greece Evia, Dystos lake 14.03.2011
90 orientalis 53,2165730 22,5833290 4 AB208 to AB211 - - JX182153 to JX182156 Poland Zajki
91 orientalis 53,4867170 22,7840490 3 AB205 to AB207 - - JX182150 to JX182152 Poland Dawidowizna
92 orientalis 51,3893820 23,2630570 10 AB242, AB244, AB246 to AB253 - - JX182165 to
JX182174 Poland Wincencin
93 orientalis 49,0330000 23,5170000 1 HLi729 - - JX182251 Ukraine Lvov, Skole 27.07.200794 arborea 35,493 23,678 1 NHMC80.2.7.16 NHMC80.2.7.16 - bad sequence Greece Crete, Kaloudiana Kissamos
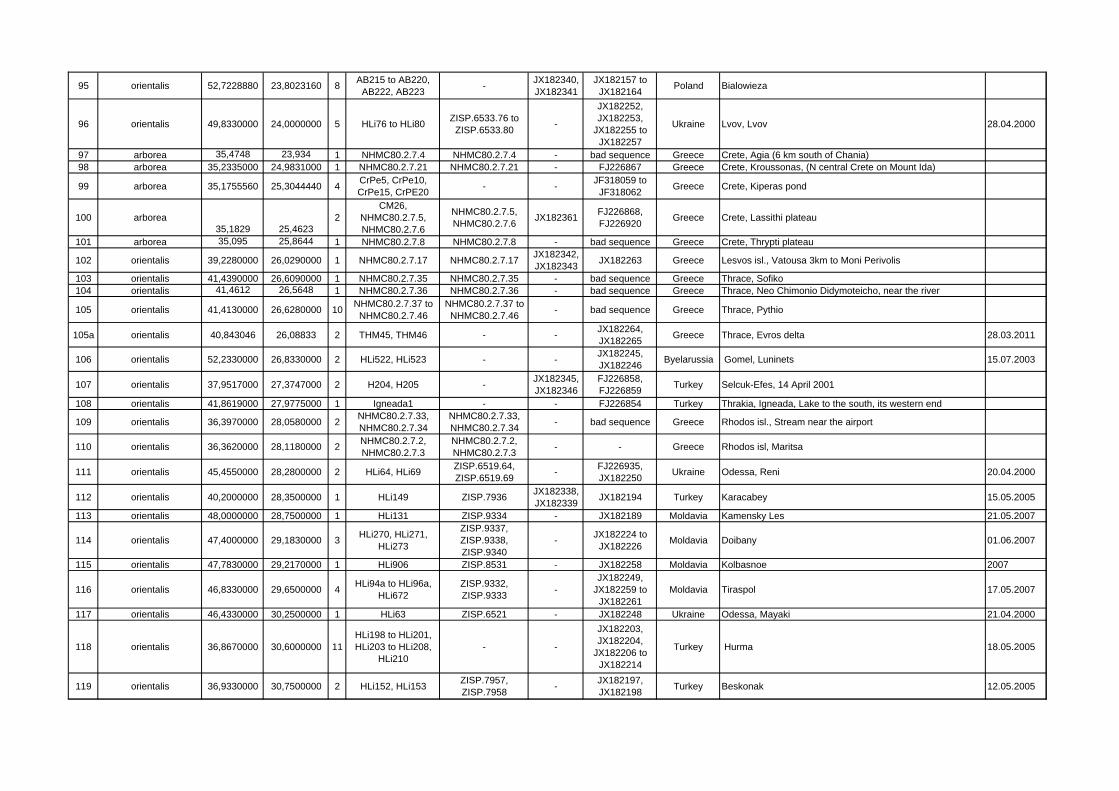
95 orientalis 52,7228880 23,8023160 8 AB215 to AB220, AB222, AB223 - JX182340,
JX182341JX182157 to
JX182164 Poland Bialowieza
96 orientalis 49,8330000 24,0000000 5 HLi76 to HLi80 ZISP.6533.76 to ZISP.6533.80 -
JX182252, JX182253,
JX182255 to JX182257
Ukraine Lvov, Lvov 28.04.2000
97 arborea 35,4748 23,934 1 NHMC80.2.7.4 NHMC80.2.7.4 - bad sequence Greece Crete, Agia (6 km south of Chania)98 arborea 35,2335000 24,9831000 1 NHMC80.2.7.21 NHMC80.2.7.21 - FJ226867 Greece Crete, Kroussonas, (N central Crete on Mount Ida)
99 arborea 35,1755560 25,3044440 4 CrPe5, CrPe10, CrPe15, CrPE20 - - JF318059 to
JF318062 Greece Crete, Kiperas pond
100 arborea35,1829 25,4623
2CM26,
NHMC80.2.7.5, NHMC80.2.7.6
NHMC80.2.7.5, NHMC80.2.7.6 JX182361 FJ226868,
FJ226920 Greece Crete, Lassithi plateau
101 arborea 35,095 25,8644 1 NHMC80.2.7.8 NHMC80.2.7.8 - bad sequence Greece Crete, Thrypti plateau
102 orientalis 39,2280000 26,0290000 1 NHMC80.2.7.17 NHMC80.2.7.17 JX182342, JX182343 JX182263 Greece Lesvos isl., Vatousa 3km to Moni Perivolis
103 orientalis 41,4390000 26,6090000 1 NHMC80.2.7.35 NHMC80.2.7.35 - bad sequence Greece Thrace, Sofiko104 orientalis 41,4612 26,5648 1 NHMC80.2.7.36 NHMC80.2.7.36 - bad sequence Greece Thrace, Neo Chimonio Didymoteicho, near the river
105 orientalis 41,4130000 26,6280000 10 NHMC80.2.7.37 to NHMC80.2.7.46
NHMC80.2.7.37 to NHMC80.2.7.46 - bad sequence Greece Thrace, Pythio
105a orientalis 40,843046 26,08833 2 THM45, THM46 - - JX182264, JX182265 Greece Thrace, Evros delta 28.03.2011
106 orientalis 52,2330000 26,8330000 2 HLi522, HLi523 - - JX182245, JX182246 Byelarussia Gomel, Luninets 15.07.2003
107 orientalis 37,9517000 27,3747000 2 H204, H205 - JX182345, JX182346
FJ226858, FJ226859 Turkey Selcuk-Efes, 14 April 2001
108 orientalis 41,8619000 27,9775000 1 Igneada1 - - FJ226854 Turkey Thrakia, Igneada, Lake to the south, its western end
109 orientalis 36,3970000 28,0580000 2 NHMC80.2.7.33, NHMC80.2.7.34
NHMC80.2.7.33, NHMC80.2.7.34 - bad sequence Greece Rhodos isl., Stream near the airport
110 orientalis 36,3620000 28,1180000 2 NHMC80.2.7.2, NHMC80.2.7.3
NHMC80.2.7.2, NHMC80.2.7.3 - - Greece Rhodos isl, Maritsa
111 orientalis 45,4550000 28,2800000 2 HLi64, HLi69 ZISP.6519.64, ZISP.6519.69 - FJ226935,
JX182250 Ukraine Odessa, Reni 20.04.2000
112 orientalis 40,2000000 28,3500000 1 HLi149 ZISP.7936 JX182338, JX182339 JX182194 Turkey Karacabey 15.05.2005
113 orientalis 48,0000000 28,7500000 1 HLi131 ZISP.9334 - JX182189 Moldavia Kamensky Les 21.05.2007
114 orientalis 47,4000000 29,1830000 3 HLi270, HLi271, HLi273
ZISP.9337, ZISP.9338, ZISP.9340
- JX182224 to JX182226 Moldavia Doibany 01.06.2007
115 orientalis 47,7830000 29,2170000 1 HLi906 ZISP.8531 - JX182258 Moldavia Kolbasnoe 2007
116 orientalis 46,8330000 29,6500000 4 HLi94a to HLi96a, HLi672
ZISP.9332, ZISP.9333 -
JX182249, JX182259 to
JX182261Moldavia Tiraspol 17.05.2007
117 orientalis 46,4330000 30,2500000 1 HLi63 ZISP.6521 - JX182248 Ukraine Odessa, Mayaki 21.04.2000
118 orientalis 36,8670000 30,6000000 11HLi198 to HLi201, HLi203 to HLi208,
HLi210- -
JX182203, JX182204,
JX182206 to JX182214
Turkey Hurma 18.05.2005
119 orientalis 36,9330000 30,7500000 2 HLi152, HLi153 ZISP.7957, ZISP.7958 - JX182197,
JX182198 Turkey Beskonak 12.05.2005
120 orientalis 36,2330000 32,5170000 2 HLi150, HLi151 ZISP.7871, ZISP.7872
JX182336, JX182337
JX182195, JX182196 Turkey Gazipasa 09.05.2005
121 orientalis 46,6330000 32,6170000 1 HLi435 - - JX182229 Ukraine Kherson 13.06.2004
122 savignyi 34,8140000 32,7170000 4 NHMC80.2.23.1 to 4 NHMC80.2.23.1 to 4 -
JX182307, JX182308, bad
sequencesCyprus Kidasi
123 savignyi 34,7100000 32,8500000 4 Cyprus1, Cyprus2, Cyprus3, Cyprus4 - -
FJ226930, FJ226908, FJ226909, FJ226931
Cyprus Sotira
124 savignyi 35,2000000 33,3670000 12
HLi95 to HLi99, HLi107, HLi108, HLi110, HLi119,
HLi121 to HLi123
ZISP.8978 to ZISP.8989 -
JX182272 to JX182274,
JX182280 to JX182283, JX182301, JX182302,
JX182304 to JX182306
Cyprus Nikosia 07.05.2006
125 orientalis 44,5000000 33,4830000 1 HLi786 - JX182330, JX182331 JX182254 Ukraine Crimea, Fiolent Cape 02.08.2007
126 orientalis 44,5170000 34,2330000 1 HLi294 - - JX182227 Ukraine Crimea, Nikita 2001127 orientalis 44,7500000 34,3670000 1 HLi517 - - JX182243 Ukraine Crimea, Luchistoe 2003
128 savignyi 36,8830000 34,7330000 16
HLi39, HLi89, HLi90, HLi92 to HLi94,
HLi101 to HLi105, HLi111 to HLi115
ZISP.8960 to ZISP.8975 -
JX182267 to JX182271,
JX182275 to JX182279,
JX182295 to JX182300
Turkey Mersin 09.05.2006
129 savignyi 36,8333000 34,7500000 2 H70, H71 -JX182320, JX182321
FJ226932, FJ226933 Turkey Kazanli 20.04.2002
130 felixarabica 33,1330000 35,6500000 1 HLiLeh1 - - FJ226936 Israel Lehavot
131 orientalis 49,6500000 36,2670000 3 HLi48 to HLi52 - JX182332, JX182333
JX182242, JX182244, JX182247
Ukraine Kharkov, Iskov prud 2003
132 felixarabica 32,6600000 36,7300000 2 H09, H10 - - FJ226940, FJ226860 Syria Jebbel Druz, 15.03.2001
133 orientalis 49,0500000 37,5500000 1 HLi436 - - JX182230 Ukraine Donetsk, Svyatogorsk 2004
134 orientalis 44,7330000 38,7500000 4 HLi444 to HLi447 - - JX182232 to JX182235 Russia Krasnodar, Krepostnaya 2003
135 orientalis 43,8830000 39,8500000 6 HLi448 to HLi453 - - JX182236 to JX182241 Russia Krasnodar, Salakhaul 2003
136 orientalis 43,4000000 40,0000000 1 HLi327 - - JX182228 Russia Krasnodar, Vesyoloe 2004137 orientalis 43,3330000 40,2500000 1 HLi443 - - JX182231 Georgia Abkhazia, Gagra 2003
138 savignyi 36,4833 40,75 2 H102, H103 - JX182322, JX182323
FJ226934, FJ226910 Syria Al Hasakah 12.04.2004
139 orientalis 42,9830000 41,1000000 1 HLi138a - - JX182191 Georgia Abkhazia, Bagmaran 2006140 felixarabica 15,3000000 43,5300000 1 H122 - JX182317 FJ226906 Yemen West Kawkaban, Wadi Annaim, 2200 m asl 09.07.2002141 felixarabica 15,3000000 43,5600000 1 H185 - - FJ226928 Yemen Luhlah, 25 km N Sana'a, 2200 m asl 09.07.2002142 felixarabica 15,4300000 43,5800000 1 H155 - - FJ226907 Yemen N Amran, 2232 m asl 10 July 2002 10.07.2002
143 savignyi 33,3000000 44,2000000 2 H87, H89 - JX182318, JX182319,
FJ226938, FJ226939 Iraq Bagdad, 28.03.2002
144 felixarabica 14,1326600 44,2361400 1 H215 - JX182316 FJ226929 Yemen S Yarim, Mawah, 2613 m ASL 06.06.2002
145 savignyi 41,7099810 44,7929980 1 H394 - - FJ226911 Georgia Tblissi?
146 savignyi 39,2170000 45,4170000 4 HLi129, HLi187, HLi244, HLi245
ZISP.9353, ZISP.9354, ZISP.9356, ZISP.9357
-
JX182284, JX182286, JX182293, JX182294
Azerbaijan Nakhichevan' 21-28.05.2007
147 orientalis 40,2170000 46,2500000 10HLi132, HLi190, HLi247, HLi248,
HLi264 to HLi269
ZISP.9342, ZISP.9344 to
ZISP.9352 -
JX182190, JX182202, JX182215, JX182216,
JX182218 to JX182223
Azerbaijan Zurnabad 05.06.2007
148 orientalis 41,8330000 46,2670000 1 HLi1 - - JX182205 Georgia Lagodekhi149 orientalis 43,6330000 46,4170000 1 HLi982 - - JX182262 Russia Chechnya, Starogladkovskaya 2007
150 savignyi 41,0830000 46,4170000 7
HLi130, HLi208a, HLi209a, HLi237, HLi239, HLi241,
Hli243
ZISP.9359 to ZISP.9362, ZISP.9364, ZISP.9366, ZISP.9367
-JX182285,
JX182287 to JX182292
Azerbaijan Azerbaijan, Burunchovach 2007
151 savignyi 34,3506000 46,5350000 1 MVZ245918 MVZ245918 - FJ226912 Iran Kermanshah Province, 5km (by air) W of Lalabad village, 40 km (by air) NW of Kermanshah
152 savignyi 39,4330000 47,3000000 1 HLi971 - - JX182303 Azerbaijan Horadis 2002153 orientalis 42,1830000 47,5830000 1 HLi24 ZISP.7216 - JX182217 Russia Dagestan, Kishcha 01.05.2001154 orientalis 38,7670000 48,4170000 1 HLi168 ZISP.7192 - JX182201 Azerbaijan Lerik 30.05.2001155 orientalis 38,4500000 48,8000000 1 HLi165 - - JX182200 Azerbaijan Isti-Su near Astara 29.05.2001156 orientalis 38,6500000 48,8170000 2 Hgum ZISP.7195 JX182344 FJ226856 Azerbaijan Avrora 28.05.2001
157 orientalis 38,6490000 48,8100000 2 HLi158 - JX182334, JX182335 FJ226856 Azerbaijan Avrora 28.05.2001
158 orientalis 38,5330000 48,8170000 2 HLi139, HLi140 - - JX182192, JX182193 Azerbaijan Tangerud 29.05.2001
GROUP japonica - -
HjM1, HjM2 - JX182354, JX182355
FJ226923Japan Hiroshima Prefecture, Higashi-Hiroshima,rice field
GROUP squirella 34,7477 -77,0134 Hsquirella MVZ15422
JX182313, JX182314 - USA
North Carolina, Carteret Co., 6.2 mi WSW (by air) center of Holly Springs off County Rd. 1128
GROUP pseudopuma 10,3 -84,81667 Hpseudopuma MVZ149764 JX182328,
JX182329 - Costa Rica Puntarenas Province, Monteverde townGROUP annectans - - Hannectans MVZ231182 JX182364,
JX182365 - ChinaSichuan Prov., Baoxing County, 8 km from Baoxing Hotel after Liang Hellou Bridge
Outgroup: H. japonica
Hnt1-Algerie2_Algeria_loc045Hnt1-Algerie1_Algeria_loc045Hnt1-Tunisie2.1_Tunisia_loc04891/99
Hmerid-NME777/01_Spain-Teneriffe_loc001Hmerid-MVZ186151_Morocco_loc010
Hmerid-NME299/97b_Spain-Teneriffe_loc002Hmerid-MVZ186158_Morocco_loc013Hmerid-MVZ231938_Morocco_loc007Hmerid-MVZ231943_Spain_loc008Hmerid-MoroccoMVZ231741_loc015Hmerid-NME946/02a_loc003Hmerid-MVZ177941_Spain_loc005Hmerid-MVZ186012_Spain_loc011
Hmerid-NME307/97_loc003Hmerid-HmNew1_France_loc033
Hmerid-HM38_Italie_loc044
100/100
99/100
Hsav-H87_Iraq_loc143Hsav-MVZ245918_Iran_loc151Hsav-H103_Syria_loc138Hsav-H102_Syria_loc138
Hsav-H394_Georgia_loc145Hsav-HLi971_Azerbaijan_loc152
Hsav-HLi130_Azerbaijan_loc150Hsav-HLi187_Azerbaijan_loc146Hsav-HLi244_Azerbaijan_loc146
Hsav-HLi39_Turkey_loc128Hsav-HLi114_Turkey_loc128Hsav-HLi94_Turkey_loc128
Hsav-HLi112_Turkey_loc128Hsav-H071_Turkey_loc129Hsav-HLi89_Turkey_loc128Hsav-HLi101_Turkey_loc128
Hsav-HLi102_Turkey_loc128Hsav-HLi93_Turkey_loc128Hsav-H070_Turkey_loc129
Hsav-Cyprus3_Cyprus_loc123Hsav-Cyprus4_Cyprus_loc123
Hsav-HLi105_Turkey_loc128Hsav-HLi95_Cyprus_loc124
Hsav-NHMC80.2.23.3_Cyprus_loc122Hsav-NHMC80.2.23.4_Cyprus_loc122
Hsav-Cyprus1_Cyprus_loc123Hsav-HLi110_Cyprus_loc124
68/100
63/72
Hfel-H09_Syria_loc132Hfel-HLiLeh1_Israel_loc130
Hfel-H122_Yemen_loc140Hfel-H155_Yemen_loc142
Hfel-H185_Yemen_loc141Hfel-H215_Yemen_loc144
81
99/99
53/95
100/100
100/100
Hsar-Sarda22_Italy-Sardinia_loc046Hsar-Sarda24_Italy-Sardinia_loc046Hsar-Sarda4T_France-Corsica_loc047
Hsar-Sarda1-1_France-Corsica_loc051Hsar-Sarda3-1_France-Corsica_loc051
84/71
Harb-NHMC80.2.7.26_Greece_loc083
Harb-HLi72_Ukraine_loc088Harb-NME676/00_Croatia-Cres_loc056Harb-NME675/00_Croatia-Cres_loc054Harb-Niz4_Poland_loc066Harb-NME674/00_Croatia-Cres_loc055Harb-HaII_Germany_loc053Harb-AB2_Poland_loc065Harb-AB29_Poland_loc070Harb-Cro16_Croatia_loc071Harb-Al14_Albania_loc078Harb-Cro17_Croatia_loc071Harb-HLi5a_France_loc031
Harb-NME1758/10_Croatia_Krk_loc058Harb-NME1228/04_Germany_loc052Harb-AB41_Poland_loc068Harb-B8_Netherlands_loc038Harb-MM245_Belgium_loc036Harb-ME225_Belgium_loc037Harb-HLi74_Ukraine_loc088Harb-HLi1125_Hungary_loc075Harb-Bo1_France_loc034Harb-K157_Belgium_loc032Harb-HylaM1Monterfil_France_loc028Harb-Szc1_Poland_loc067Harb-Re5_Romania_loc086Harb-Niz10_Poland_loc066Harb-Fo1_Romania_loc087Harb-C14_Netherlands_loc039Harb-A1_Netherlands_loc041Harb-B6_Netherlands_loc038Harb-HLi6_Montenegro_loc077Harb-Fo10_Romania_loc087Harb-Fo5_Romania_loc087
Harb-Re1_Romania_loc086Harb-Z194_Belgium_loc035Harb-NHMC80.2.7.19_Greece_loc081Harb-HLi194_Greece_loc082Harb-CrPe5_Greece_loc099
Harb-NHMC80.2.7.18_Greece_loc081Harb-CrPe10_Greece-Crete_loc099Harb-NME902/01B_Greece_loc089
Harb-ME223_Belgium_loc037Harb-Z187_Belgium_loc035Harb-NME902/01A_Greece_loc089
Harb-NHMC80.2.7.25_Greece_loc081Harb-NHMC80.2.7.21_Greece_loc098
Harb-CrPe15_Greece-Crete_loc099Harb-NHMC80.2.7.6_Greece-Crete_loc100
Harb-NHMC80.2.7.5_Greece-Crete_loc100Harb-BAC5_Germany_loc043Harb-Niz11_Poland_loc066Harb-PoSzc4_Poland_loc067Harb-HLi70_Hungary_loc085
Harb-BAC1_Switzerland_loc040Harb-AB15_Poland_loc074Harb-HLi71_Ukraine_loc088Harb-Niz3_Poland_loc066Harb-HLi4_France_loc031Harb-NHMC80.2.7.29_Greece_loc083Harb-HLi1140_Hungary_loc075
Harb-AB4_Poland_loc074Harb-ME230_Belgium_loc037Harb-HLi5_Montenegro_loc077Harb-NHMC80.2.7.32_Greece_loc083
Harb-NHMC80.2.7.28_Greece_loc083
Hint-NME916/01_Italy_loc064Hint-NME915/01_Italy-Sicily_loc059Hint-HLi80a_Italy_lo063
Hint-Sicily81_Italy-Sicily_loc057Hint-Sicily102_Italy-Sicily_loc061Hint-Sicily101_Italy-Sicily_loc062
Hint-Sicily103_Italy-Sicily_loc06061
Hnt2-PA2F_Switzerland_loc050Hnt2-Ti1_Switzerland_loc049
Hnt2-PA13M_Switzerland_loc050Hnt2-Ti2_Switzerland_loc049
88
Hnt2-G6.7.1_Switzerland_loc042Hnt2-PA6F_Switzerland_loc050
100/100
99/100
Hmol-M05_Spain_loc016Hmol-MNCN11086_Spain_loc014Hmol-M04_Spain_loc016Hmol-M06_Spain_loc017Hmol-MNCN3448_Spain_loc020Hmol-M33_Spain_loc024Hmol-WP212A_Spain_loc022Hmol-WP211A_Spain_loc019Hmol-WP208A_Spain_loc021Hmol-WP207A_Spain_loc026Hmol-WP205A_Spain_loc023
Hmol-MNCN11078_Spain_loc025Hmol-WP200A_Spain_loc027
Hmol-MNCN11068_Spain_loc004Hmol-StAu9_France_loc029Hmol-StAu12_France_loc029
Hmol-StAu3_France_loc030Hmol-StAu5_France_loc030
Hmol-StAu1_France_loc030
Hmol-HPA248_Portugal_loc006Hmol-M02_Spain_loc016
Hmol-M01_Spain_loc018Hmol-HLimol_Spain_loc009Hmol-M03_Spain_loc016
84/100
70/76
Hor-HLi165_Azerbaijan_loc155Hor-HLi140_Azerbaijan_loc158Hor-HLi168_Azerbaijan_loc154Hor-HLi158_Azerbaijan_loc157
Hor-HLi139_Georgia_loc158Hor-Hgum_Azerbaijan_loc156
Hor-HLi448_Russia_loc135HorHLi982_Russia_loc149
Hor-HLi447_Russia_loc134Hor-HLi452_Russia_loc135Hor-HLi1_Georgia_loc148Hor-HLi327_Russia_loc136Hor-HLi453_Russia_loc135Hor-HLi445_Russia_loc134
Hor-HLi450_Russia_loc135Hor-HLi451_Russia_loc135Hor-HLi444_Russia_loc134Hor-HLi446_Russia_loc134
Hor-HLi138a_Georgia_loc139Hor-HLi132_Azerbaijan_loc147Hor-HLi24_Russia_loc153Hor-HLi248_Azerbaijan_loc147Hor-HLi190_Azerbaijan_loc147
Hor-HLi449_Russia_loc135Hor-HLi443_Georgia_loc137
62
99/94
57/98
Hor-NHMC80.2.7.17_Greece-Lesvos_loc102Hor-HLi206_Turkey_loc118
Hor-HLi201_Turkey_loc118Hor-HLi210_Turkey_loc118
Hor-HLi151_Turkey_loc120Hor-HLi199_Turkey_loc118Hor-HLi150_Turkey_loc120
Hor-HLi64_Ukraine_loc111Hor-Igneada1_Turkey_loc108Hor-HLi435_Ukraine_loc121Hor-HLi149_Turkey_loc112
Hor-H205_Turkey_loc107Hor-H204_Turkey_loc107
Hor-HLi198_Turkey_loc118Hor-HLi152_Turkey_loc119
Hor-HLi270_Moldavia_loc114Hor-HLi69_Ukraine_loc111
92
89/100
Hor-AB242_Poland_loc092Hor-AB215_Poland_loc095Hor-AB136_Poland_loc080Hor-AB252_Poland_loc092
Hor-AB68_Poland_loc072Hor-AB100_Poland_loc076
Hor-AB187_Poland_loc079Hor-AB46_Poland_loc069Hor-HLi294_Ukraine_loc126
Hor-AB205_Poland_loc091Hor-HLi63_Ukraine_loc117Hor-HLi48_Ukraine_loc131Hor-HLi436_Ukraine_loc133Hor-AB59_Poland_loc072Hor-AB184_Poland_loc079Hor-AB48_Poland_loc069Hor-AB246_Poland_loc092
Hor-AB216_Poland_loc095Hor-AB208_Poland_loc090Hor-AB218_Poland_loc095
Hor-AB162_Poland_loc084Hor-AB146_Poland_loc080
Hor-HLi131_Moldavia_loc113Hor-HLi517_Ukraine_loc127Hor-HLi522_Byelarussia_loc106Hor-AB244_Poland_loc092
Hor-HLi672_Moldavia_loc116Hor-AB88_Poland_loc073
Hor-HLi51_Ukraine_loc131Hor-AB183_Poland_loc079
Hor-HLi729_Ukraine_loc093Hor-HLi76_Ukraine_loc096
Hor-HLi94a_Moldavia_loc116Hor-AB250_Poland_loc092Hor-AB248_Poland_loc092
Hor-HLi77_Ukraine_loc096Hor-HLi786_Ukraine_loc125Hor-AB40_Poland_loc069Hor-AB204_Poland_loc084Hor-AB69_Poland_loc072Hor-AB53_Poland_loc069
Hor-HLi80_Ukraine_loc096Hor-AB167_Poland_loc084
Hor-HLi906_Moldavia_loc115Hor-HLi95a_Moldavia_loc116Hor-HLi96a_Moldavia_loc116
69
77
70/100
92/100
85/100
95/100
98/100
100/100
100/100
100/100
91/100
94/100
87/100
94/100
100/100
0.05
Harb-THM20_Greece_loc083aHarb-THM19_Greece_loc083a
Harb-HylaRuegen1_Germany_loc53a
Hor-THM45_Greece_loc105a
Hmol-Dordogne07_France_loc30a
Hmol-Dordogne11_France_loc30aHmol-Dordogne12_France_loc30a
H. intermedia s.l.
H. arborea
H. molleri
H. orientalis
H. savignyi
H. felixarabica
H. sarda
H. meridionalis s.l.
-/72
-/75
HLi72
Fo10
HLi70
Hli6 HLi114
Cro16 Niz4
ME230
HLi5a
HLi4 Z194
NeB6, NeA1, NeC14
Niz3
Niz11, Szc4
Cro17
HLi5
HLi194, NHMC80.2.7.
19
NHMC80.2.7.32
NHMC80.2.7.28
Re1 Fo5
CrPe5 CrPe15, NHMC80.2.7.5
NHMC80.2.7.6
NHMC80.2.7.21
CrPe10
NHMC80.2.7.25
NHMC80.2.7.18
Z187,ME223
THM19
NHMC80.2.7.26
NME1758/10
HLi71
NHMC80.2.7.29, HLi1140
NME674/00, NME675/00, NME676/00,
Niz4
NME902/01B
THM20
NME902_01A
AB4, AB15, BAC1, BAC5, Fo1, Niz10, Re5, Szc1, HylaM1Monterfil, K157, Bo1, HLi1125, HLi74, ME225,
MM245, B8, AB41, NME1128/04, Al14, AB29, AB2, HaII, HylaRuegen1
WP205A
StAu1, StAu12, Dordogne07
StAu5 StAu9
MNCN11068
StAu3
HPA248
MNCN11078, WP207A, WP208A, WP211A, WP212A,M33, MNCN3448
M04, M06, MNCN11086
M05
WP200A
M01 M03
HLimol
M02
Dordogne11
Dordogne12
a c
b
HLi672
HLi51
HLi294
AB248
AB250HLi94a AB68, AB242, AB145,
AB100, AB223
AB252
AB50, AB59, AB184
AB53
AB67, AB140, AB164, HLi80
HLi48, HLi436
HLi729
AB46, AB187
HLi131 HLi76
AB218
HLi96a AB208, AB216, AB205 AB162, AB183, HLi517, HLi77, AB244, HLi522, AB40, HLi906, HLi95a, AB88, HLi63, HLi786,
AB146
HLi206 HLi150HLi208
HLi201
HLi210
HLi149, HLi270
HLi198
HLi153
H205
HLi435
Igneada1 HLi69
HLi64
THM46
HLi447
HLi448,HLi982
HLi452 HLi1
HLi138a
HLi269, HLi24
HLi248
HLi267 HLi268,
HLi327
HLi445,HLi453
HLi444 HLi446
HLi450 HLi449HLi443
HLi165
HLi168, HLi140
HLi139
HLi158 Hgum
NHMC80.2.7.17
24 x
HLi151
H204
HLi201
C_Y40_02
13 x
AB246
Related Documents











