1912-1918 Nucleic Acids Research, 1995, Vol. 23, No. 11 gar2 is a nucleolar protein from Schizosaccharomyces pombe required for 18S rRNA and 40S ribosomal subunit accumulation Marie-Pierre Gulli, Jean-Philippe Girard+, Dan Zabetakis1, Bruno Lapeyre§, Teri Melesel and Michble Caizergues-Ferrer* Laboratoire de Biologie Moleculaire Eucaryote du CNRS, 118 Route de Narbonne, 31062 Toulouse, France and 1Columbia University, Department of Biological Sciences, New York, NY, USA Received February 23, 1995; Revised and Accepted Aprl 27, 1995 ABSTRACT Several nucleolar proteins, such as nucleolin, NOP1/ fibrillarin, SSB1, NSR1 and GARI share a common glycine and arginine rich structural motif called the GAR domain. To identify novel nucleolar proteins from fission yeast we screened Schizosaccharomyces pombe genomic DNA libraries with a probe en- compassing the GAR structural motif. Here we report the identification and characterization of a S.pombe gene coding for a novel nucleolar protein, designated gar2. The structure of the fission yeast gar2 is reminiscent of that of nucleolin from vertebrates and NSR1 from Saccharomyces cerevislae. In addition, like these proteins, gar2 has a nucleolar localisatlon. The disruption of the gar2+ gene affects normal cell growth, leads to an accumulation of 35S pre-rRNA and a decrease of mature 18S rRNA steady state levels. Moreover, ribosomal profiles of the mutant show an increase of free 60S ribosomal subunits and an absence of free 40S ribosomal subunits. gar2 is able to rescue a S.cerevsiae mutant lacking NSR1, thus establishing gar2 as a functional homolog of NSRI. We propose that gar2 helps the assembly of pre-ribosomal particles containing 18S rRNA. INTRODUCTION In eukaryotes, most steps of ribosome biogenesis occur within a specialized nuclear domain, the nucleolus. The RNA polymerase I transcibes a primary single transcript, the pre-rRNA. In addition to 18S, 5.8S and 25S rRNAs sequences which are maintained in the mature rRNA forms, the pre-rRNA contains spacer regions not found in cytoplasmic ribosomes. Evidence exists that the growing pre-rRNA chain interacts with ribosomal proteins (1), non-ribosomal proteins and small nucleolar RNAs (snoRNAs) (2), leading to the formation of pre-ribosomal particles. In these RNP complexes, the pre-rRNA undergoes base modification, EMBL accession no. Z48166 methylation and a series of endo- and exonucleolytic cleavages (pre-rRNA processing) which remove the transcribed spacers. Still, little is known about the mechanisms involved in the post-transcriptional steps of ribosome production. Due to their specific nucleolar location, snoRNAs have long been thought to be required for ribosome biogenesis. Indeed roles in pre-rRNA processing have been clearly demonstrated for U3, U14, snR30, snR10 and MRP RNA in Saccharomyces cerevisiae (3-8) and for U3, U8 and U22 in higher eukaryotes (9-13). According to the generally accepted model, snoRNAs function via direct interac- tions with the pre-rRNA. Consistent with this idea, base pairing interactions with pre-rRNA has been established in the cases of yeast and mammalian U3 (14-17), snR30 (5) and mammalian U17/E1, E2 and E3 (18). In addition, a subset of snoRNAs from higher eukaryotes, including U16, U18, U20 and U21, contains phylogenetically conserved extended regions of perfect comple- mentarity to rRNA sequences (reviewed in 19 and 20). This suggests that these snoRNAs might interact with various rRNA domains in the nucleolus. Factors able to modulate the structure of the pre-rRNA and the interactions between the pre-rRNA and snoRNAs or between pre-rRNA and proteins must clearly play an important role in ribosome synthesis. Indeed, two putative RNA helicases necess- ary for assembly of the 60S ribosomal subunit have been described (21,22). Futhermore, the GAR domain found in several nucleolar proteins such as nucleolin (23,24), fibrillarin/NOPI (25-28), SSB1 (29), NSR1 (30) and GARI (31) required for ribosome synthesis, has been reported to have the capacity to modify rRNA structure in vitro (32). The widespread occurrence of the GAR domain among nucleolar proteins characterized so far enabled us to undertake the isolation of novel nucleolar proteins from Schizosaccharomyces pombe using a cDNA fragment encoding the GAR domain of Xenopus fibrillarin as a probe. In this paper we report the characterization of a novel nucleolar protein of fission yeast called gar2. Although the overall structural organization of gar2 resembles of that of nucleolin, gar2 has only two RNA binding domains instead of the four found in nucleolin. The gar2+ gene * To whom correspondence should be addressed Present addresses: +The Center for Blood Research, Harvard Medical School, 200 Longwood Avenue, Boston, MA 02115, USA and §Division of Biochemistry and Molecular Biology, 401 Barker Hall, University of California at Berkeley, Berkeley, CA 94720, USA Q--Z) 1995 Oxford University Press

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

1912-1918 Nucleic Acids Research, 1995, Vol. 23, No. 11
gar2 is a nucleolar protein from Schizosaccharomycespombe required for 18S rRNA and 40S ribosomalsubunit accumulationMarie-Pierre Gulli, Jean-Philippe Girard+, Dan Zabetakis1, Bruno Lapeyre§, TeriMelesel and Michble Caizergues-Ferrer*
Laboratoire de Biologie Moleculaire Eucaryote du CNRS, 118 Route de Narbonne, 31062 Toulouse, France and1Columbia University, Department of Biological Sciences, New York, NY, USA
Received February 23, 1995; Revised and Accepted Aprl 27, 1995
ABSTRACT
Several nucleolar proteins, such as nucleolin, NOP1/fibrillarin, SSB1, NSR1 and GARI share a commonglycine and arginine rich structural motif called theGAR domain. To identify novel nucleolar proteins fromfission yeast we screened Schizosaccharomycespombe genomic DNA libraries with a probe en-compassing the GAR structural motif. Here we reportthe identification and characterization of a S.pombegene coding for a novel nucleolar protein, designatedgar2. The structure of the fission yeast gar2 isreminiscent of that of nucleolin from vertebrates andNSR1 from Saccharomyces cerevislae. In addition, likethese proteins, gar2 has a nucleolar localisatlon. Thedisruption of the gar2+ gene affects normal cellgrowth, leads to an accumulation of 35S pre-rRNA anda decrease of mature 18S rRNA steady state levels.Moreover, ribosomal profiles of the mutant show anincrease of free 60S ribosomal subunits and anabsence of free 40S ribosomal subunits. gar2 is able torescue a S.cerevsiae mutant lacking NSR1, thusestablishing gar2 as a functional homolog of NSRI. Wepropose that gar2 helps the assembly of pre-ribosomalparticles containing 18S rRNA.
INTRODUCTION
In eukaryotes, most steps of ribosome biogenesis occur within aspecialized nuclear domain, the nucleolus. The RNA polymeraseI transcibes a primary single transcript, the pre-rRNA. In additionto 18S, 5.8S and 25S rRNAs sequences which are maintained inthe mature rRNA forms, the pre-rRNA contains spacer regionsnot found in cytoplasmic ribosomes. Evidence exists that thegrowing pre-rRNA chain interacts with ribosomal proteins (1),non-ribosomal proteins and small nucleolar RNAs (snoRNAs)(2), leading to the formation of pre-ribosomal particles. In theseRNP complexes, the pre-rRNA undergoes base modification,
EMBL accession no. Z48166
methylation and a series of endo- and exonucleolytic cleavages(pre-rRNA processing) which remove the transcribed spacers.
Still, little is known about the mechanisms involved in thepost-transcriptional steps of ribosome production. Due to theirspecific nucleolar location, snoRNAs have long been thought tobe required for ribosome biogenesis. Indeed roles in pre-rRNAprocessing have been clearly demonstrated for U3, U14, snR30,snR10 andMRPRNA in Saccharomyces cerevisiae (3-8) and forU3, U8 and U22 in higher eukaryotes (9-13). According to thegenerally accepted model, snoRNAs function via direct interac-tions with the pre-rRNA. Consistent with this idea, base pairinginteractions with pre-rRNA has been established in the cases ofyeast and mammalian U3 (14-17), snR30 (5) and mammalianU17/E1, E2 and E3 (18). In addition, a subset of snoRNAs fromhigher eukaryotes, including U16, U18, U20 and U21, containsphylogenetically conserved extended regions of perfect comple-mentarity to rRNA sequences (reviewed in 19 and 20). Thissuggests that these snoRNAs might interact with various rRNAdomains in the nucleolus.
Factors able to modulate the structure of the pre-rRNA and theinteractions between the pre-rRNA and snoRNAs or betweenpre-rRNA and proteins must clearly play an important role inribosome synthesis. Indeed, two putative RNA helicases necess-ary for assembly of the 60S ribosomal subunit have beendescribed (21,22). Futhermore, the GAR domain found in severalnucleolar proteins such as nucleolin (23,24), fibrillarin/NOPI(25-28), SSB1 (29), NSR1 (30) and GARI (31) required forribosome synthesis, has been reported to have the capacity tomodify rRNA structure in vitro (32).The widespread occurrence of the GAR domain among
nucleolar proteins characterized so far enabled us to undertake theisolation of novel nucleolar proteins from Schizosaccharomycespombe using a cDNA fragment encoding the GAR domain ofXenopus fibrillarin as a probe. In this paper we report thecharacterization of a novel nucleolar protein of fission yeastcalled gar2. Although the overall structural organization of gar2resembles of that of nucleolin, gar2 has only two RNA bindingdomains instead of the four found in nucleolin. The gar2+ gene
* To whom correspondence should be addressed
Present addresses: +The Center for Blood Research, Harvard Medical School, 200 Longwood Avenue, Boston, MA 02115, USA and §Division ofBiochemistry and Molecular Biology, 401 Barker Hall, University of California at Berkeley, Berkeley, CA 94720, USA
Q--Z) 1995 Oxford University Press

Nucleic Acids Research, 1995, Vol. 23, No. 11 1913
disruption affects normal cell growth at 300C. Additionally, agar2- mutant accumulates 35S pre-rRNA and has a decrease inthe steady-state levels of 18S rRNA. Expression of gar2 in aS.cerevisiae nsrl- mutant restores the wild-type phenotype,suggesting that S.pombe gar2 is a functional homolog ofS.cerevisiae NSR1.
MATERIALS AND METHODS
Plasmids
pBL1242. A 1.8 kb PCR product corresponding to the gar2+cDNA was digested with BamHI and cloned into the BamHI siteof the vector pAR3040, for expression of recombinant protein inEscherichia coli under the T7 promoter (33).pBL1252. A 2.1kb insert corresponding to the Hindlll genomicfragment of gar2+ was cloned into the BamHI-Sall site of thepBM272 vector (30).pBL1263. Contains a HindlIl fragment of 11 kb that correspondsto the entire rDNA repeated unit (34). This plasmid was eitherdouble digested with KpnI and NdeI (the resulting 1022 bpfragment was used as a 5' ETS probe) or digested with XbaI (theresulting 1460 bp fragment was used as a 18S probe).pWL1O. Contains a 1.6 kb BamHI-SalI fragment that correspondsto the entire coding region ofNSR1 inserted into the BamHI-SalIsite of pBM272 (30).
All DNA manipulations and bacterial transformations weredone according to published procedures (35).
Yeast strains
Sp972. h-, this haploid wild-type strain was first described by U.Leupold.SplS. h+, ade6M210, leul-32, ura4DJ8 (gift from P. Fantes).SpJ6. h, ade6M216, leul-32, ura4DJ8 (generous gift from P.Fantes).SpJ8. h+/h-, ade6M210/ade6M216, 1eu132/1eu132, ura4D18-/ura4DJ8 (obtained by conjugation of SplS and Spl6).SpJ9. h+/h, ade6M210/ade6M216, leul-32/leul-32, ura4D18/ura4D18, gar2+/gar2::ura4+ (obtained by transformation ofSpl8).Sp22. h-, ade6M216, leul-32, ura4DJ8 (gar2+).Sp24. h+, ade6M216, leul-32, ura4D18, gar2::ura4+(both latterstrains were obtained after sporulation of the Spl9 diploid strain).WLY353. MATa, ade2-1, canl-100, ura3-1, leu2-3,112, trpl-1,his3-11,15, nsrl::HIS3 (30).S.pombe cells were cultured either in rich medium YES: 0.5%
yeast extract, 3% glucose plus supplements or in EMM minimalmedium (36).
S.cerevisiae media used were YPG: 1% yeast extract (DifcoLaboratories), 2% Bacto peptone (Difco) and 2% galactose, orSG: 0.67% yeast nitrogen base without amino acids (Difco) and2% galactose. EMM and SD media were both supplemented asrequired.S.pombe and S.cerevisiae were transformed by standard
techniques (36,37).
Isolation of the gene encoding the S.pombe gar2 protein
A 2.1 kb HindHu fragment containing the gar2+ gene was firstdetected by Southern blot analysis of S.pombe genomic DNA, atlow stringency with a cDNA probe corresponding to the GAR
domain ofXenopus fibrillarin (26,38,39). A S.pombe subgenomiclibrary was then constructed with size selected (2.1 kb) HindIllgenomic fragments, following a strategy previously described(31,38,39). The isolated clone was characterized by restrictionmapping and DNA sequencing using a sequenase kit (USBCorporation). The EMBL accession number is Z48166. A gar2+cDNA was cloned using a PCR-based strategy previouslydescribed (38,39). Briefly, 1 ,ug of S.pombe total RNA was mixedwith 1 ,ug of a BamHI-oligo(dT) oligonucleotide [CCCGGAT-CC(T)17] and denatured for 10' at 70°C before returning to ice.Reverse transcription was then performed using a BRL kit. A PCRamplification of the cDNA thus obtained with the BamHI-oligo(dT)primer and a second BamHI-G2P6 primer (5'-CCCGGATCCAT-GGCAAAAAAGGATAAAACC-3') complementary to the 5' endof the gar2+ ORF allowed us to obtain a 1.8 kb PCR productcorresponding to the gar2 + cDNA.
Bacterial expression of the gar2 protein and obtentionof polyclonal serum
The plasmid pBL1242 was introduced into Ecoli strain BL21(DE3) LysS. Recombinant gar2 expression was induced withisopropyl P-D-thiogalactopyranoside (IPTG) to a final concentra-tion of 0.4 mM (33). Induced E.coli protein extract was analyzedby 8% SDS-PAGE. Recombinant gar2 protein was electroelutedfrom 8% polyacrylamide gel and used to raise antibodies inrabbits (40). Antibodies against gar2 were purified from immuneserum using recombinant protein as an affinity ligand. For theaffinity column, 1 mg of gar2 was coupled with CNBr activatedSepharose (Pharmacia). In this case, the gar2 protein was purifiedfrom an E.coli extract by three successive chromatographies,Heparin Sepharose, Q Sepharose and Mono Q Sepharose.
Iumunofluorescence microscopy
The procedure followed was based on that of Hagan and Hyams(41). Cells were fixed in PEMS (100 mM PIPES pH 6.9, 1 mMEGTA, 1 mM MgSO4, 1 M Sorbitol) containing 3.7% formalde-hyde for 1.5 h at room temperature. They were then washed threetimes in PEMS. For digestion of the cell wall, cells wereresuspended at a density of 107 cells/ml in PEMS containing 0.3mg/ml Lysing enzyme (Sigma) and 0.15 mg/ml Zymolyase 20T(ICN Biomedicals) and incubated 30 min at room temperature.After three washes in PEMS, cells were applied to poly-L-lysinecoated slides, air dried and permeabilized by three quick washesin acetone at room temperature. Afterwards, cells were incubatedat room temperature with primary anti-gar2 antibody at 1/4dilution in PBS plus 0.05% Tween 20 for 1 h. Following threewashes in the same buffer, cells were incubated with secondaryantibody (anti-rabbit Ig-Texas Red linked from donkey; Amer-sham) in PBS plus 0.05% Tween 20, for 1 h at room temperature.After three more washes in PBS plus 0.05% Tween 20 and onein PBS, a drop of mounting medium containing 1 mg/mlp-phenylene diamine and 1 ,ug/ml 4',6-diamidino-2-phenylindole(DAPI) was applied to the air-dried cells. Slides were viewed witha Zeiss photomicroscope.
Disruption of the gar2+ gene and tetrad analysis
The chromosomal copy of the gar2+ gene was replaced with apartial gar2 deletion by the one-step gene disruption method (42).The 0.49 kb SacI-KpnI fragment of the gar2 + gene (aa 163-325)

1914 Nucleic Acids Research, 1995, Vol. 23, No. 11
was cut and replaced by the 1.8 kb SacI-KpnI fragment containingthe S.pombe ura4+ gene from the plasmid pBKS+ura4 + (gift fromG. Basi). A linear 3.7 kb fragment containing the ura4+ gene
flanked by the sequences ofthe gar2 + gene (0.672 kb at 5' end and0.860 kb at 3' end) was isolated and used to transform the diploidura4- strain Spl8 obtained from SpiS and Spl6. ura4+transformants were checked by Southern analysis to confirm thatthe interrupted gene was integrated into the homologous gar2+locus. Transformants which contained one wild-type copy andone disrupted copy of the gar2+gene (gar2+1gar2:: ura4+) were
sporulated, the tetrads dissected and analyzed according to (43).The steady-state levels ofrRNAs were analyzed by Northern blot(35).
Ribosomal profiles
Analysis of ribosomes was done essentially as described (44).S.pombe strains were grown in YES medium, while S.cerevisiaestrains were grown overnight in SG medium to select for theplasmid, and then for several generations in YPG medium. Cellswere collected and washed with bufferA (10mM Tris-Cl pH 7.4,100 mM NaCl, 30 mM MgCl2 and 50 ig of cycloheximide/ml).Cells were lysed with glass beads and the supernatants collected.Ribosomes were analyzed by centrifugation through a sucrose
gradient (50 mM Tris-acetate pH 7, 50 mM NH4Cl, 12 mMMgCl2, 1 mM dithiothreitol and 7, 17, 27, 37 and 47% sucrose).Gradient were centrifuged at 89 000 g for 5.5 h in a SW27 rotor(Beckman). Samples were analyzed with an ISCO model 640gradient fractionator and the effluent was monitored as OD254.
RESULTS
Isolation and characterization of the gar2+ gene
The similarities between the GAR domains of the two nucleolarproteins nucleolin and fibrillarin are sufficient to obtain, in a lowstringency hybridization experiment, a cross reaction between theDNA sequences encoding these domains (26). In order tocharacterize new genes encoding proteins containing GARdomains in S.pombe, we first performed Southern analysis usingtotal genomic DNA from S.pombe digested either by EcoRl orbyHindIu and a fragment of the Xenopus laevis fibrillarin cDNA as
a probe (38,39). Four strong hybridization signals were detected,cwrresponding to HinduI fragments of 5.2 kb, 3.5, 2.2 and 2.1 kb(38). In order to clone the corresponding fragments, we preparedand screened different subgenomic libraries, each containing thesized fragments 5.2, 3.5, 2.2 and 2.1 kb. The isolated clones havebeen analyzed and sequenced. The 5.2 kb fragment has not beenfurther characterized, the 3.5 kb fragment contains the S.pombegene for fibrillarin (39), the 2.2 kb fragment contains the S.pombegari + gene (38), the equivalent of a S.cerevisiae gene that wehave characterized (31) and the 2.1 kb fragment corresponds toa new S.pombe gene.This 2.1 kb HindIl fragment has been mapped and sequenced
on both strands. Sequence analysis reveals the presence of a 1500bp open reading frame (ORF) that could encode a protein of 500residues. After data base comparison, it appeared to be a new
protein of S.pombe containing a GAR domain located at theC-terminus of the protein; therefore, we have called it gar2. Sincemany genes of S.pombe, as opposed to S.cerevisiae, do containone or more introns, we have cloned a gar2 + cDNA from
and Methods). Detailed restriction mapping, followed bysequencing using different primers, allowed us to confirm theabsence of introns within this gene (data not shown). A Northernblot was performed using total RNA from S.pombe to define thesize of the gar2 mRNA. A unique S.pombemRNA of 1750 nt wasdetected (39), which is consistent with the size of the gar2+ORFand the absence of intron within the gene.
gar2 is structurally related to NSR1 and nucleolin
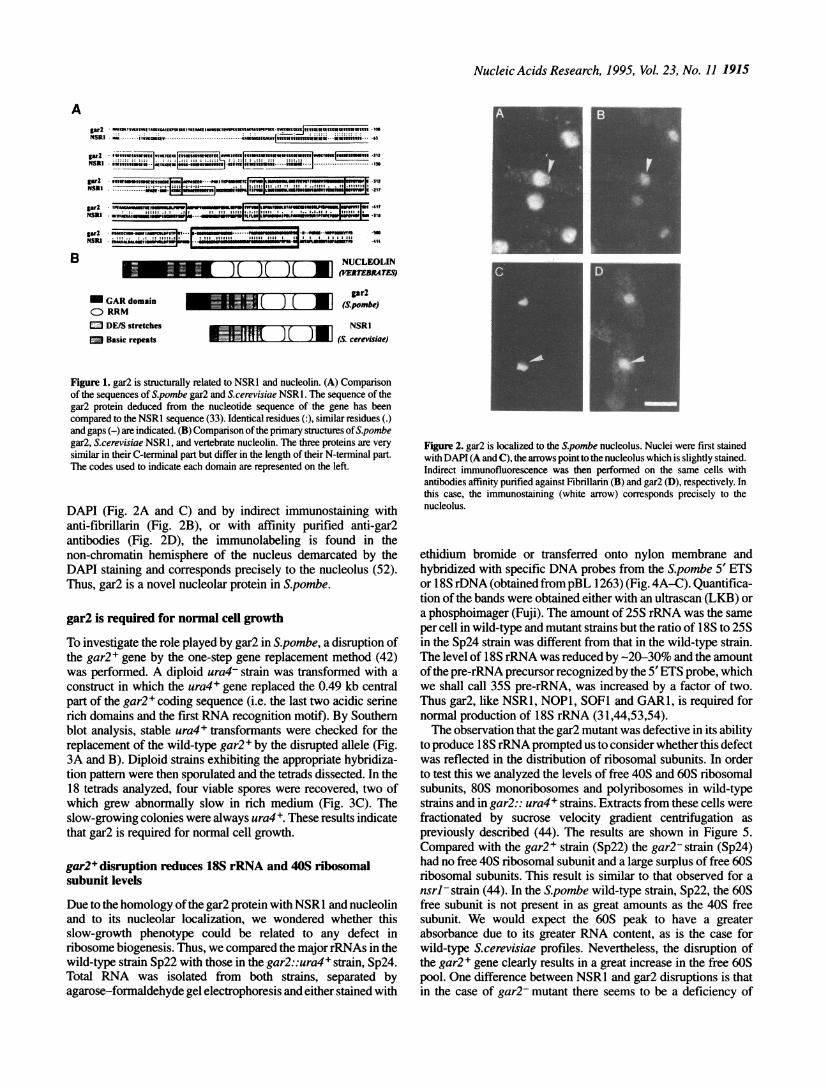
While the characterization of gar2 was being carrried out, thesequence of a protein of S.cerevisiae, called NSR1, was reported(30). The alignment of the amino-acid sequences of the twoproteins reveals that gar2 and NSR1 exhibit 53% identity (Fig.lA). In addition, the structure of these two proteins is stronglyreminiscent of that of nucleolin from vertebrates. Like nucleolinand NSR1, gar2 has four different domains (Fig. iB). The firstdomain of 80 residues is reminiscent of the N-terminal domain ofnucleolin, containing numerous doublets of lysine surrounded byhydrophobic residues and a potential cdc2 phosphorylation site(S50PKK). In nucleolin, eight threonine residues in the N-terminaldomain are phosphorylated in vitro and in vivo by the cdc2 kinase(45,46). The major difference between nucleolin or gar2 andNSR1, is that the latter has only a very short N-terminal basicdomain and does not contain a cdc2 phosphorylation site. Thesecond domain in gar2 is formed by long stretches of serines andacidic residues. Similar serine/acidic rich domains are found inNSR1 while the four acidic domains of nucleolin are different inthe sense that they contain more acidic than serine residues. Ingar2, NSR1 and nucleolin some serine residues in the acidicdomain are CKII phosphorylation sites (47; Gulli, Faubladier andCaizergues-Ferrer, unpublished data). The third domain is theputative RNA binding domain, formed by the repetition of a longconserved domain of -80 residues, which is present in a largefamily of nuclear RNA binding proteins and is referred to as theRRM (for RNA recognition motif; 48,49). gar2 and NSR1contain twoRRMs and nucleolin contains fourRRMs. The fourthdomain is the GAR domain. The typical motif FGGRGG, foundin multiple copies in the nucleolar GAR proteins (31) is repeatedtwice in the gar2 protein. This motif has been described as a targetfor post-translational modifications such as methyl incorporationin the form of N9,N9-dimethylarginine in various nuclear RNAbinding proteins, e.g. hnRNP A and B proteins, fibrillarin andnucleolin (50 and references therein).
gar2 is a nucleolar protein of S.pombeThe expression of the gar2+ gene in S.pombe, was analyzed byWestern blots. Rabbit antibodies against gar2 have been producedand affinity purified (see Materials and Methods). Western blotanalysis of crude protein extracts from S.pombe with theseantibodies revealed a single protein that migrates on SDS-PAGEas a 72 kDa polypeptide (result not shown). This apparentmolecular mass of the gar2 protein is larger than the predicted 52kDa calculated from the sequence. Such an abnormal mobilityduring electrophoresis is probably due to the presence in gar2 oflong stretches of serine/acidic residues, as previously shown forother nucleolar proteins containing acidic stretches (51). Theaffinity-purified antibodies were used to localize gar2 byimmunofluorescence. 4',6-diamidino-2-phenylindole (DAPI)stains both mitochondrial and nuclear DNA in fixed S.pombecells. When double labeling of the same cells was performed withS.pombe total RNA using a PCR-based strategy (see Materials
Nucleic Acids Research, 1995, Vol. 23, No. 11 1915
Agarl FM,tSwK.MStaNSRI -..TTMGWKef
gar2 nu,sfsssicsvwg.:::::: :: ::::
"
NSRI * fsss"nnfi.il-f,
gr2lNSRIt
pr2-NSRIt
gir2NSRI
. . . . 22*:: ::: :: .:: ::: ::.:.. .*wFTw"f ~ .... " m .In....
@st_t ,,.,1ttfctwhtlw.-* ;*:* *6"it?I'21?
TKAXMMN tU _I _lwotwm I_11OON -41?
*znfu;"~~~~~~~~~~ wag ,-t-- -| ...|_;;1 lo -Si
.l_ItSttmv-omalco * -. ..... ......................@ _"*--_--_:. :!::: L",Rt t? -u :
*DIOMNf*mAliW_ ..-- " w_ins
B NUCLEOLINR N A. (VERTEBRATS
M GAR domainCD RRMM DE/S stretches
m Basic repeats
gakr2(S.pombe)
NSRI19 ) ( ~~~(S. cerevisiae)
Figure 1. gar2 is structurally related to NSRl and nucleolin. (A) Comparisonof the sequences of Spombe gar2 and S.cerevisiae NSRl. The sequence of thegar2 protein deduced from the nucleotide sequence of the gene has beencompared to the NSRI sequence (33). Identical residues (:), similar residues (.)and gaps (-) are indicated. (B) Comparison of the primary structures ofS.pombegar2, S.cerevisiae NSRl, and vertebrate nucleolin. The three proteins are verysimilar in their C-terminal part but differ in the length of their N-terminal part.The codes used to indicate each domain are represented on the left.
DAPI (Fig. 2A and C) and by indirect immunostaining withanti-fibrillarin (Fig. 2B), or with affinity purified anti-gar2antibodies (Fig. 2D), the immunolabeling is found in thenon-chromatin hemisphere of the nucleus demarcated by theDAPI staining and corresponds precisely to the nucleolus (52).Thus, gar2 is a novel nucleolar protein in S.pombe.
gar2 is required for normal cell growth
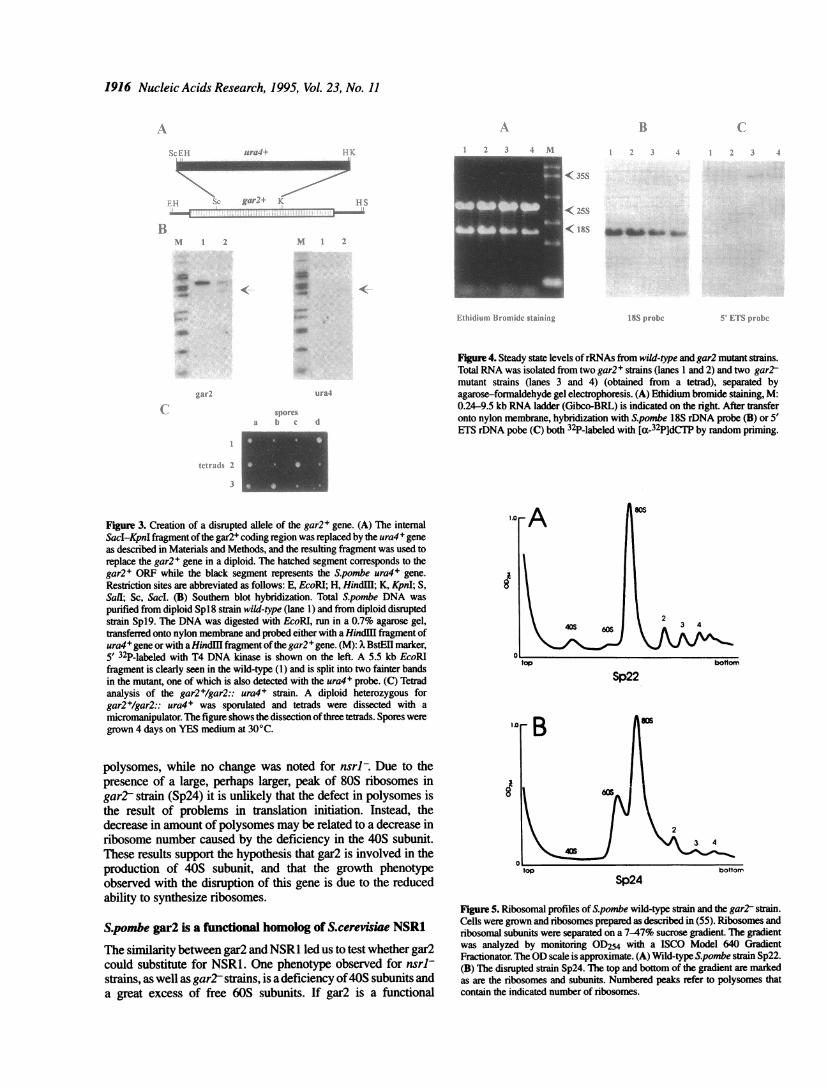
To investigate the role played by gar2 in S.pombe, a disruption ofthe gar2+ gene by the one-step gene replacement method (42)was performed. A diploid ura4- strain was transformed with aconstruct in which the ura4+ gene replaced the 0.49 kb centralpart of the gar2+ coding sequence (i.e. the last two acidic serinerich domains and the first RNA recognition motif). By Southernblot analysis, stable ura4+ transformants were checked for thereplacement of the wild-type gar2+ by the disrupted allele (Fig.3A and B). Diploid strains exhibiting the appropriate hybridiza-tion pattern were then sporulated and the tetrads dissected. In the18 tetrads analyzed, four viable spores were recovered, two ofwhich grew abnormally slow in rich medium (Fig. 3C). Theslow-growing colonies were always ura4+. These results indicatethat gar2 is required for normal cell growth.
gar2+ disruption reduces 18S rRNA and 40S ribosomalsubunit levels
Due to the homology ofthe gar2 protein with NSR1 and nucleolinand to its nucleolar localization, we wondered whether thisslow-growth phenotype could be related to any defect inribosome biogenesis. Thus, we compared the major rRNAs in thewild-type strain Sp22 with those in the gar2::ura4+ strain, Sp24.Total RNA was isolated from both strains, separated byagarose-formaldehyde gel electrophoresis and either stained with
Figure 2. gar2 is localized to the Spombe nucleolus. Nuclei were first stainedwith DAPI (A and C), the arrows point to the nucleolus which is slightly stained.Indirect immunofluorescence was then performed on the same cells withantibodies affinity purified against Fibrillarin (B) and gar2 (D), respectively. Inthis case, the immunostaining (white arrow) corresponds precisely to thenucleolus.
ethidium bromide or transferred onto nylon membrane andhybridized with specific DNA probes from the S.pombe 5' ETSor 18S rDNA (obtained frompBL 1263) (Fig. 4A-C). Quantifica-tion of the bands were obtained either with an ultrascan (LKB) ora phosphoimager (Fuji). The amount of 25S rRNA was the sameper cell in wild-type and mutant strains but the ratio of 18S to 25Sin the Sp24 strain was different from that in the wild-type strain.The level of 18S rRNA was reduced by -20-30% and the amountofthe pre-rRNA precursor recognized by the 5'ETS probe, whichwe shall call 35S pre-rRNA, was increased by a factor of two.Thus gar2, like NSR1, NOPI, SOFI and GARI, is required fornormal production of 18S rRNA (31,44,53,54).The observation that the gar2 mutant was defective in its ability
to produce 18S rRNA prompted us to consider whether this defectwas reflected in the distribution of ribosomal subunits. In orderto test this we analyzed the levels of free 40S and 60S ribosomalsubunits, 80S monoribosomes and polyribosomes in wild-typestrains and in gar2:: ura4+ strains. Extracts from these cells werefractionated by sucrose velocity gradient centrifugation aspreviously described (44). The results are shown in Figure 5.Compared with the gar2+ strain (Sp22) the gar2-strain (Sp24)had no free 40S ribosomal subunit and a large surplus of free 60Sribosomal subunits. This result is similar to that observed for ansrl strain (44). In the S.pombe wild-type strain, Sp22, the 60Sfree subunit is not present in as great amounts as the 40S freesubunit. We would expect the 60S peak to have a greaterabsorbance due to its greater RNA content, as is the case forwild-type S.cerevisiae profiles. Nevertheless, the disruption ofthe gar2 + gene clearly results in a great increase in the free 60Spool. One difference between NSR1 and gar2 disruptions is thatin the case of gar2- mutant there seems to be a deficiency of
'WO-414
1916 Nucleic Acids Research, 1995, VoL 23, No. 11
2 j 4 \l
II
< 35S
< 25S
< I18%olIbI4 Iaw 0e
R V. S prol)VS ro(b).
Figure 4. Steady state levels ofrRNAs from wild-type and gar2 mutant strains.Total RNA was isolated from two gar2+ strains (lanes 1 and 2) and two gar2tmutant strains (lanes 3 and 4) (obtained from a tetrad), separated byagarose-formaldehyde gel electrophoresis. (A) Ethidium bromide staining, M:0.24-9.5 kb RNA ladder (Gibco-BRL) is indicated on the right. After transferonto nylon membrane, hybridization with S.pombe 18S rDNA probe (B) or 5'ETS rDNA pobe (C) both 32P-labeled with [a-32P]dCTP by random priming.
Figure 3. Creation of a disrupted allele of the gar2+ gene. (A) The internalSacI-KpnI fragment ofthe gar2+ coding region was replaced by the ura4 + geneas described in Materials and Methods, and the resulting fragment was used toreplace the gar2+ gene in a diploid. The hatched segment corresponds to thegar2+ ORF while the black segment represents the S.pombe ura4+ gene.
Restriction sites are abbreviated as follows: E, EcoRl; H, HindIl; K, KpnI; S,SalI; Sc, Sacl. (B) Southem blot hybridization. Total Spombe DNA was
purified from diploid Spl8 strain wild-type (lane 1) and from diploid disruptedstrain Spl9. The DNA was digested with EcoRI, run in a 0.7% agarose gel,transferred onto nylon membrane and probed either with a HindU fragment ofura4+ gene or with a Hindli fragment ofthe gar2 + gene. (M): xBstEII marker,
5' 32P-labeled with T4 DNA kinase is shown on the left. A 5.5 kb EcoRlfragment is clearly seen in the wild-type (1) and is split into two fainter bandsin the mutant, one of which is also detected with the ura4+ probe. (C) Tetradanalysis of the gar2+/gar2:: ura4+ strain. A diploid heterozygous forgar2+/gar2:: ura4+ was sporulated and tetrads were dissected with a
micromanipulator. The figure shows the dissection ofthree tetrads. Spores weregrown 4 days on YES medium at 30°C.
polysomes, while no change was noted for nsrl-. Due to thepresence of a large, perhaps larger, peak of 80S ribosomes ingart strain (Sp24) it is unlikely that the defect in polysomes isthe result of problems in translation initiation. Instead, the
decrease in amount of polysomes may be related to a decrease inribosome number caused by the deficiency in the 40S subunit.These results support the hypothesis that gar2 is involved in theproduction of 40S subunit, and that the growth phenotypeobserved with the disruption of this gene is due to the reducedability to synthesize ribosomes.
S.pombe gar2 is a functional homolog of S.cerevisiae NSR1
The similarity between gar2 and NSR1 led us to test whether gar2could substitute for NSR1. One phenotype observed for nsrl-strains, as well as gar2 strains, is a deficiency of40S subunits anda great excess of free 60S subunits. If gar2 is a functional
.orA
2
top
Sp22
88
B
3 4
Sp24
FigureS. Ribosomal profiles of S.pombe wild-type strain and the gart strain.Cells were grown and ribosomes prepared as described in (55). Ribosomes andribosomal subunits were separated on a 7-47% sucrose gradient. The gradientwas analyzed by monitoring OD254 with a ISCO Model 640 GradientFractionator. TheOD scale is approximate. (A) Wild-type S.pombe strain Sp22.(B) The disrupted strain Sp24. The top and bottom of the gradient are markedas are the ribosomes and subunits. Numbered peaks refer to polysomes thatcontain the indicated number of ribosomes.
¼
ICL.:H ura4 i
gar K
Bi
I s7-El-
xi 2
.I-
N1
a4; _
aw
u rat4
'I) () r d
11
1 (I [II f1fB1i1- l'II ( t. Stkli II 1we
(vt li;t] 2
Nucleic Acids Research, 1995, Vol. 23, No. 11 1917
8
8
A806
2 3 4
B
pBLl252
control
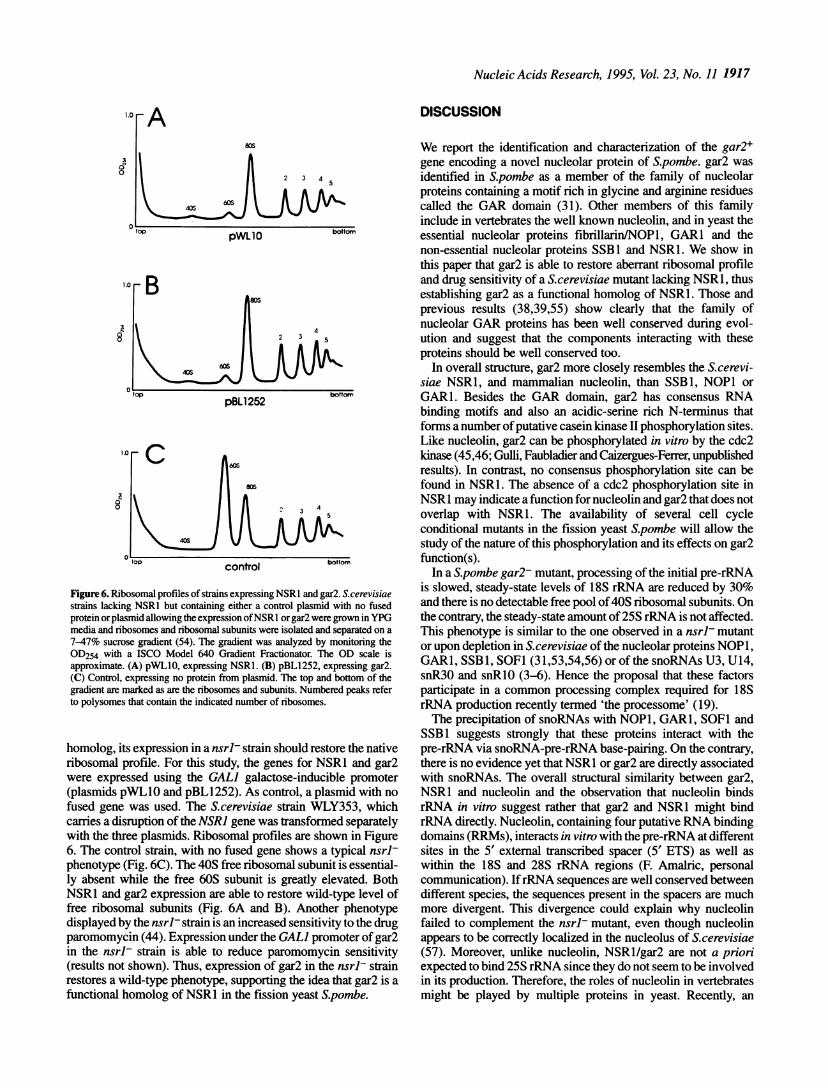
Figure 6. Ribosomal profiles of strains expressing NSRl and gar2. S.cerevisiaestrains lacking NSRI but containing either a control plasmid with no fusedprotein orplasmid allowing the expression ofNSR I or gar2 were grown in YPGmedia and ribosomes and ribosomal subunits were isolated and separated on a7-47% sucrose gradient (54). The gradient was analyzed by monitoring theOD254 with a ISCO Model 640 Gradient Fractionator. The OD scale isapproximate. (A) pWLIO, expressing NSRl. (B) pBL1252, expressing gar2.(C) Control, expressing no protein from plasmid. The top and bottom of thegradient are marked as are the ribosomes and subunits. Numbered peaks referto polysomes that contain the indicated number of ribosomes.
homolog, its expression in a nsrl- strain should restore the nativeribosomal profile. For this study, the genes for NSR1 and gar2were expressed using the GAL] galactose-inducible promoter(plasmids pWLIO and pBLI252). As control, a plasmid with nofused gene was used. The S.cerevisiae strain WLY353, whichcarries a disruption of the NSRJ gene was transformed separatelywith the three plasmids. Ribosomal profiles are shown in Figure6. The control strain, with no fused gene shows a typical nsrl-phenotype (Fig. 6C). The 40S free ribosomal subunit is essential-ly absent while the free 60S subunit is greatly elevated. BothNSR1 and gar2 expression are able to restore wild-type level offree ribosomal subunits (Fig. 6A and B). Another phenotypedisplayed by the nsrl- strain is an increased sensitivity to the drugparomomycin (44). Expression under the GALI promoter of gar2in the nsrl- strain is able to reduce paromomycin sensitivity(results not shown). Thus, expression of gar2 in the nsrl- strainrestores a wild-type phenotype, supporting the idea that gar2 is afunctional homolog of NSR1 in the fission yeast S.pombe.
DISCUSSION
We report the identification and characterization of the gar2+gene encoding a novel nucleolar protein of S.pombe. gar2 wasidentified in S.pombe as a member of the family of nucleolarproteins containing a motif rich in glycine and arginine residuescalled the GAR domain (31). Other members of this familyinclude in vertebrates the well known nucleolin, and in yeast theessential nucleolar proteins fibrillarin/NOPI, GARI and thenon-essential nucleolar proteins SSB1 and NSR1. We show inthis paper that gar2 is able to restore aberrant ribosomal profileand drug sensitivity of a S.cerevisiae mutant lacking NSR1, thusestablishing gar2 as a functional homolog of NSR1. Those andprevious results (38,39,55) show clearly that the family ofnucleolar GAR proteins has been well conserved during evol-ution and suggest that the components interacting with theseproteins should be well conserved too.
In overall structure, gar2 more closely resembles the S.cerevi-siae NSR1, and mammalian nucleolin, than SSB1, NOPI orGARL. Besides the GAR domain, gar2 has consensus RNAbinding motifs and also an acidic-serine rich N-terminus thatforms a number ofputative casein kinase II phosphorylation sites.Like nucleolin, gar2 can be phosphorylated in vitro by the cdc2kinase (45,46; Gulli, Faubladier and Caizergues-Ferrer, unpublishedresults). In contrast, no consensus phosphorylation site can befound in NSR1. The absence of a cdc2 phosphorylation site inNSR1 may indicate a function for nucleolin and gar2 that does notoverlap with NSR1. The availability of several cell cycleconditional mutants in the fission yeast S.pombe will allow thestudy of the nature of this phosphorylation and its effects on gar2function(s).
In a S.pombe gar2- mutant, processing of the initial pre-rRNAis slowed, steady-state levels of 18S rRNA are reduced by 30%and there is no detectable free pool of40S ribosomal subunits. Onthe contrary, the steady-state amount of25S rRNA is not affected.This phenotype is similar to the one observed in a nsrl- mutantor upon depletion in S.cerevisiae of the nucleolar proteins NOP 1,GARI, SSB1, SOFI (31,53,54,56) or of the snoRNAs U3, U14,snR30 and snRIO (3-6). Hence the proposal that these factorsparticipate in a common processing complex required for 18SrRNA production recently termed 'the processome' (19).The precipitation of snoRNAs with NOPI, GARI, SOF1 and
SSB1 suggests strongly that these proteins interact with thepre-rRNA via snoRNA-pre-rRNA base-pairing. On the contrary,there is no evidence yet that NSR1 or gar2 are directly associatedwith snoRNAs. The overall structural similarity between gar2,NSR1 and nucleolin and the observation that nucleolin bindsrRNA in vitro suggest rather that gar2 and NSR1 might bindrRNA directly. Nucleolin, containing four putative RNA bindingdomains (RRMs), interacts in vitro with the pre-rRNA at differentsites in the 5' external transcribed spacer (5' ETS) as well aswithin the 18S and 28S rRNA regions (F. Amalric, personalcommunication). IfrRNA sequences are well conserved betweendifferent species, the sequences present in the spacers are muchmore divergent. This divergence could explain why nucleolinfailed to complement the nsrl- mutant, even though nucleolinappears to be correctly localized in the nucleolus of S.cerevisiae(57). Moreover, unlike nucleolin, NSRl/gar2 are not a prioriexpected to bind 25S rRNA since they do not seem to be involvedin its production. Therefore, the roles of nucleolin in vertebratesmight be played by multiple proteins in yeast. Recently, an

1918 Nucleic Acids Research, 1995, Vol. 23, No. 11
essential nucleolar protein termed NOP4/NOP77 containingthree canonical RRMs and required for 25S production andpre-rRNA methylation was characterized (58,59).We propose that gar2 might interact with the pre-rRNA via its
two RRMs in combination with the GAR domain. Futhermoregar2, like NSRl or nucleolin, might directly help recruit orassemble some highly basic ribosomal proteins with the pre-rRNA via its acidic/serine rich N-terminus (60). To test theseassumptions, we are currently investigating by mutationalanalyses the roles of the gar2 subdomains, searching for its RNAtarget(s) and protein partners.
ACKNOWLEDGEMENTS
We thank Y. Henry, T. Kiss and P. Ferrer for critical reading of themanuscript and other members of our group for fruitful dis-cussions. We are grateful to J. Feliu and M. Faubladier forexcellent technical assistance, to Y. de Preval for oligonucleotidessynthesis, and P. Fantes and B. Ducommun for providing strainsand vectors. We are indebted to F. Amalric for his continuoussupport. MPG and JPG had a fellowship from the Ministere del'Enseignement Superieur et de la Recherche (MESR). Thisresearch was carried out under contract by the Association pourla Recherche sur le Cancer and by the Universite Paul Sabatier.
REFERENCES1 Chooi, W. Y., and Leiby, K. R. (1981) Proc. NatI. Acad. Sci. USA 78,
4823-4827.2 Mougey, E. B., O'Reilly, M., Osheim, Y., Miller Jr, 0. L., Beyer, A., and
Sollner-Webb, B. (1993) Genes Dev. 7, 1609-1619.3 Hughes, J. M. X., and Ares, M. Jr (1991) EMBO J. 10, 4231-4239.4 Li, H. V., Zagorski, J., and Fournier, M.J. (1990) Mol. Cell. Biol. 10,
1145-1152.5 Morrissey, J., and Tollervey, D. (1993) Mol. Cell. Biol. 13, 2469-2477.6 Tollervey, D. (1987) EMBO J. 6, 4169-4175.7 Schmitt, M. E., and Clayton, D. A. (1993) Mol. Cell. Biol. 13, 7935-7941.8 Lygerou, Z., Mitchell, P., Petfalski, E., Seraphin, B., and Tollervey, D.
(1994) Genes Dev. 8, 1423-1433.9 Savino, R., and Gerbi, S. (1990) EMBO J. 9, 2299-2308.10 Kass, S., Tyc, K., Steitz, J.A., and Sollner-Webb, B. (1990) Cell 60,
897-908.11 Mougey, E. B., Pape, L. K., and Sollner-Webb, B. (1993) Mol. Cell. Biol.
13, 5990-5998.12 Peculis, B.A., and Steitz, J.A. (1993) Cell 73, 1233-1245.13 Tycowski, K., Shu, M. D., and Steitz, J. A. (1994) Science 266,
1558-1561.14 Maser, R. L., and Calvet, J. P. (1989) Proc. Natl. Acad. Sci. USA 86,
6523-6527.15 Stroke, I. L., and Weiner, A. M. (1989) J. Mol. Biol. 210, 497-512.16 Tyc, K., and Steitz, J.A. (1992) Nucleic Acids Res. 20, 5375-5382.17 Beltrame, M., and Tollervey, D. (1992) EMBO J. 11, 1531-1542.18 Rimoldi, 0. J., Raghu, B., Mihir, K. N., and Eliceiri, G. L. (1993) Mol.
Cell. Bio. 13, 4382-4390.19 Foumier, M. J., and Maxwell, E. S. (1993) Trends Biochem. Sci. 18,
131-135.20 Sollner-Webb, B. (1993) Cell 75, 403-405.21 Sachs, A. B., and Davies, R. W. (1990) Science 247, 1077-1079.22 Ripmaster, T. L., Vaughin, G. P., and Woolford Jr., J. L. (1992) Proc. Natl.
Acad. Sci. USA 89, 11131-11135.23 Lapeyre, B., Bourbon, H. M., and Amalinc, F (1987) Proc. Natl. Acad. Sci.
USA 84, 1472-1476.
24 Caizergues-Ferrer, M., Mariottini, P., Curie, C., Lapeyre, B., Gas, N.,Amalric, F., and Amaldi, F. (1989) Genes Dev. 3, 324-333.
25 Shimmang, T., Tollervey, D., Kern, H., Frank, R., and Hurt, E.C. (1989)EMBO J. 8, 489-497.
26 Lapeyre, B., Mariottini, P., Mathieu, C., Ferrer, P., Amaldi, F., Amalric, F.,and Caizergues-Ferrer, M. (1990) Mol. Cell. Biol. 10, 430-434.
27 Henriquez, R., Blobel, G., and Afis, J. P. (1990) J. Bio. Chem. 265,2209-2215.
28 Aris, J. P., and Blobel G. (1991) Proc. Natl. Acad. Sci. USA 88,931-935.29 Jong, A. Y. S, Clark M. W., Gilbert, M., Oehm, A., and Campbell, J. L.
(1987) Mol. Cell. Biol. 7, 2947-2955.30 Lee, W.C., Xue, Z., and Melese, T. (1991) J. Cell Biol. 113, 1-12.31 Girard, J. P., Lehtonen, H., Caizergues-Ferrer, M., Amalric, F., Tollervey,
D., and Lapeyre, B. (1992) EMBO J. 11, 673-682.32 Ghisolfi, L., Joseph, G., Amalric, F., and Erard, M. (1992) J. Biol. Chem.
267, 2955-2959.33 Studier, F. W., Rosenberg, A. H., Dunn, J. J., and Dubendorf J. W. (1990)
Methods Enzymol. 185, 60-68.34 Lapeyre, B., Michot, B., Feliu, J., and Bachellenie, J.P. (1993) Nucleic
Acids Res. 21, 3322.35 Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989) Molecular Cloning: A
Laboratory Manual, 2nd ed.. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
36 Moreno, S., Klar, A., and Nurse, P. (1991) Methods Enzymol. 194,795-823.
37 Ito, H., Fukuda, Y., Murata, K., and Kimura, A. (1983) J. Bacteriol. 153,163-168.
38 Girard, J. P., Caizergues-Ferrer, M., and Lapeyre, B. (1993) Nucleic AcidsRes. 21, 2149-2155.
39 Girard, J. P., Feliu, J., Caizergues-Ferrer, M. and Lapeyre, B. (1993)Nucleic Acids Res. 21, 1881-1887.
40 Harlow, E., and Lane, D. (1988) Antibodies: A Laboratory Manual. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
41 Hagan, I. M., and Hyams, J. S. (1988) J. Cell. Sci. 89, 343-357.42 Rothstein, R.J. (1983) Methods Enzymol. 101, 203-211.43 Sherman, F., Hicks, J. B., and Fink, G. R. (1986) Methods in Yeasts
Genetics. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY.44 Lee, W. C., Zabetakis, D., and Melese, T. (1992) Mol. Cell. Biol. 12,
3865-3871.45 Belenguer, P., Caizergues-Ferrer, NI., Labbe, J. C., Doree, M., and Amalric,
F. (1990). Mol. Cell. Biol. 10, 3607-3618.46 Peter, M., Nakagawa, J., Doree, M., Labbe, J. C., and Nigg, E. A. (1990)
Cell 60, 791-801.47 Caizergues-Ferrer, M., Belenguer, P., Lapeyre, B., Amalric, F, Wallace, M.
O., and Olson, M. 0. J. (1987) Biochemistry 26, 7876-7883.48 Mattaj, I. W. (1993) Cell 73, 827-840.49 Kenan, D. J., Query, C. C., and Keene, J. D. (1991) Trends Biochem. Sci.
16, 214-220.50 Najbauer, J., Jonhson, B. A., Young, A. L., and Aswad, D. W. (1993) J.
Biol. Chem. 268, 10501-10509.51 Alderuccio, F., Chan, E. K. L., and Tan, E. M. (1991) J. Exp. Med. 173,
941-952.52 Hirano, T., Konoha, G., Toda, T., and Yanagida, M (1989) J. Cell Biol.
108, 243-253.53 Tollervey, D., Lehtonen, H., Carmo-Fonseca, M., and Hurt, E. C. (1991)
EMBO J. 10, 573-5837.54 Jansen, R., Tollervey, D., and Hurt, E. C. (1993) EMBO J. 12, 2549-2558.55 Jansen, R. P., Hurt, E. C., Kern, H., Lehtonen, H., Carmo-Fonseca, M.,
Lapeyre, B., and Tollervey, D. (1991) J. Cell Biol. 113, 715-729.56 Clark, M. W., Yip, M. L. R., Campbell, J., and Abelson, J. (1990) J. Cell
Biol. 111, 1741-175 1.57 Xue, Z., Shan, X., Lapeyre, B., and Melese, T. (1993) Eur J. Cell Biol. 62,
13-21.58 Sun, C., and Woolford, J. L. Jr (1994) EMBO J. 13, 3127-3135.59 Berges, T., Petfalski, E., Tollervey, D., and Hurt, E. C. (1994) EMBO J. 13,
3136-3148.60 Xue, Z., and Melese, T (1994) Trends Biol. Cell 4, 414-417.
Related Documents











