Physiologia Plantarum 143: 316 – 328. 2011 Copyright © Physiologia Plantarum 2011, ISSN 0031-9317 γ -Tocopherol methyltransferase from the green alga Chlamydomonas reinhardtii : functional characterization and expression analysis Gregorio G ´ alvez-Valdivieso, Rosa Carde ˜ nosa, Jos ´ e Manuel Vera, Manuel Pineda and Miguel Aguilar ∗ Departamento de Bot ´ anica, Ecolog´ ıa y Fisiolog´ ıa Vegetal, Instituto Andaluz de Biotecnolog´ ıa, Campus de Rabanales, Edif. C-4, 3 a Planta, Universidad de C ´ ordoba, 14071 C ´ ordoba, Spain Correspondence *Corresponding author, e-mail: [email protected] Received 1 December 2010; revised 14 March 2011 doi:10.1111/j.1399-3054.2011.01507.x γ -Tocopherol methyltransferase (γ -TMT) (EC 2.1.1.95) is a very important enzyme in tocopherol biosynthesis in all photosynthetic organisms. In this paper, we present the functional characterization and expression analysis of γ -TMT from the unicellular green alga Chlamydomonas reinhardtii. Recombinant TMT1 enzyme was purified and characterized. The size of TMT1 subunit was estimated as 37 kDa by sodium dodecylsulphate polyacrylamide gel electrophoresis (SDS-PAGE), in accordance with the predicted molecular size after TMT1 cDNA sequence. Recombinant TMT1 also showed an apparent molecular mass of 37 kDa in its native conformation, suggesting that native TMT1 has a monomeric structure similar to the plant TMTs already characterized. pH and temperature dependence of TMT1 activity were also similar to plant TMTs. Substrate specificity studies showed that Chlamydomonas TMT1 is responsible for the conversion of γ - and δ- tocopherol to α- and β -tocopherol, respectively. The kinetic properties of Chlamydomonas recombinant γ -TMT activity were studied and γ -TMT1 has a similar affinity for γ - and δ-tocopherol. Promoter sequence analysis and expression analysis by northern blot revealed that tmt1 expression is strongly upregulated by high light and downregulated by low temperature. This regulatory pattern of tmt1 expression supports the idea that γ - and α-tocopherol play specific roles in the adaptation to growth under low temperature and high light stress conditions. Introduction Living systems generate free radicals spontaneously. Those free radicals that are related to the activity of reactive oxygen species (ROS) have a particularly important biological relevance (Kehrer and Smith 1994). In addition, exogenous sources of free radicals must be taken into consideration; in plants, these include Abbreviations – ABA, abscisic acid; BHT, 2,6-di-tert-butyl-4-methylphenol; DTT, DL-Dithiothreitol; DMPBQ, 2,3-dimethyl- 6-phytil-1,4-benzoquinol; EDTA, ethylenediaminetetraacetic acid; GGDP, geranylgeranyldiphosphate; GST, glutathione S-transferase; HEPES, N-(2-Hydroxyethyl)piperazine-N -(2-ethanesulfonic acid); HGA, homogentisic acid; HPPD, p- hydroxyphenylpyruvate dioxygenase; HPT, homogentisate phytiltransferase; IPTG, isopropyl β -D-thiogalactopyranoside; MES, 2-(N-Morpholino)ethanesulfonic acid; MPBQ, 2-methyl-6-phytil-1,4-benzoquinol; PAGE, polyacrylamide gel electrophoresis; RT-PCR, reverse transcriptase-PCR; PDP, phytildiphosphate; ROS, reactive oxygen species; SDS, sodium dodecylsulphate; TMT, γ -tocopherol methyltransferase. pathogens infections, UV radiation, ozone and sulfur dioxide, herbicides and phytotoxic metals (Zn, Cu, Cd, etc.), and many stress-inducing factors like high or low salinity/temperature/light, nutrients deprivation, anoxia and mechanical stress (Wojtaszek 1997, Bolwell 1999, Mullineaux and Karpinski 2002). Oxidative stress can be defined as the result of an imbalance toward the oxidant 316 Physiol. Plant. 143, 2011

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Physiologia Plantarum 143: 316–328. 2011 Copyright © Physiologia Plantarum 2011, ISSN 0031-9317
γ -Tocopherol methyltransferase from the green algaChlamydomonas reinhardtii: functional characterizationand expression analysisGregorio Galvez-Valdivieso, Rosa Cardenosa, Jose Manuel Vera, Manuel Pineda and Miguel Aguilar∗
Departamento de Botanica, Ecologıa y Fisiologıa Vegetal, Instituto Andaluz de Biotecnologıa, Campus de Rabanales, Edif. C-4, 3a Planta, Universidadde Cordoba, 14071 Cordoba, Spain
Correspondence*Corresponding author,e-mail: [email protected]
Received 1 December 2010;revised 14 March 2011
doi:10.1111/j.1399-3054.2011.01507.x
γ -Tocopherol methyltransferase (γ -TMT) (EC 2.1.1.95) is a very importantenzyme in tocopherol biosynthesis in all photosynthetic organisms. In thispaper, we present the functional characterization and expression analysisof γ -TMT from the unicellular green alga Chlamydomonas reinhardtii.Recombinant TMT1 enzyme was purified and characterized. The size of TMT1subunit was estimated as 37 kDa by sodium dodecylsulphate polyacrylamidegel electrophoresis (SDS-PAGE), in accordance with the predicted molecularsize after TMT1 cDNA sequence. Recombinant TMT1 also showed anapparent molecular mass of 37 kDa in its native conformation, suggestingthat native TMT1 has a monomeric structure similar to the plant TMTsalready characterized. pH and temperature dependence of TMT1 activitywere also similar to plant TMTs. Substrate specificity studies showed thatChlamydomonas TMT1 is responsible for the conversion of γ - and δ-tocopherol to α- and β-tocopherol, respectively. The kinetic properties ofChlamydomonas recombinant γ -TMT activity were studied and γ -TMT1has a similar affinity for γ - and δ-tocopherol. Promoter sequence analysisand expression analysis by northern blot revealed that tmt1 expression isstrongly upregulated by high light and downregulated by low temperature.This regulatory pattern of tmt1 expression supports the idea that γ - andα-tocopherol play specific roles in the adaptation to growth under lowtemperature and high light stress conditions.
Introduction
Living systems generate free radicals spontaneously.Those free radicals that are related to the activityof reactive oxygen species (ROS) have a particularlyimportant biological relevance (Kehrer and Smith 1994).In addition, exogenous sources of free radicals mustbe taken into consideration; in plants, these include
Abbreviations – ABA, abscisic acid; BHT, 2,6-di-tert-butyl-4-methylphenol; DTT, DL-Dithiothreitol; DMPBQ, 2,3-dimethyl-6-phytil-1,4-benzoquinol; EDTA, ethylenediaminetetraacetic acid; GGDP, geranylgeranyldiphosphate; GST, glutathioneS-transferase; HEPES, N-(2-Hydroxyethyl)piperazine-N′-(2-ethanesulfonic acid); HGA, homogentisic acid; HPPD, p-hydroxyphenylpyruvate dioxygenase; HPT, homogentisate phytiltransferase; IPTG, isopropyl β-D-thiogalactopyranoside; MES,2-(N-Morpholino)ethanesulfonic acid; MPBQ, 2-methyl-6-phytil-1,4-benzoquinol; PAGE, polyacrylamide gel electrophoresis;RT-PCR, reverse transcriptase-PCR; PDP, phytildiphosphate; ROS, reactive oxygen species; SDS, sodium dodecylsulphate;TMT, γ -tocopherol methyltransferase.
pathogens infections, UV radiation, ozone and sulfurdioxide, herbicides and phytotoxic metals (Zn, Cu, Cd,etc.), and many stress-inducing factors like high or lowsalinity/temperature/light, nutrients deprivation, anoxiaand mechanical stress (Wojtaszek 1997, Bolwell 1999,Mullineaux and Karpinski 2002). Oxidative stress can bedefined as the result of an imbalance toward the oxidant
316 Physiol. Plant. 143, 2011

state of the equilibrium between prooxidant and antioxi-dant factors (Sies 1991). As a consequence of this imbal-ance, free radicals can cause structural damages on manydifferent molecules (Kehrer and Smith 1994, Gechevet al. 2003, Hurst et al. 2004). Two defense lines againstthe damaging effects of free radicals are postulated.One of them is based on repair/degradation mechanisms(Bray and West 2005). But the first defense line againstfree radicals involves two types of preventive mecha-nisms, some of them based on the action of enzymeslike superoxide dismutase, catalase, peroxidase and glu-tathione reductase (Scandalios 1994, Mallick and Mohn2000), and others entailing the direct action of antioxi-dant species, i.e. ascorbic acid, reduced glutathione orvitamin E (Fryer 1992, Noctor and Foyer 1998).
The term vitamin E makes reference to a groupof strong lipid-soluble antioxidants known as tocols,consisting of a hydrophilic chromanol ring and a longhydrophobic phytol chain. There are eight differentmolecules with vitamin E activity: α-, β-, γ - andδ-tocopherol, and α-, β-, γ - and δ-tocotrienol. Alltocopherols have a 16-carbon saturated isoprenoid chainwith three chiral centers (at positions 2′, 4′ and 6′)all adopting the R configuration, while tocotrienolshave three double bonds (at positions 3′, 7′ and 11′).The different tocopherols and tocotrienols differ in thenumber and position of methyl groups on the chromanolring (Herrera and Barbas 2001).
Vitamin E is synthesized only by photosyntheticorganisms, and its enzymatic pathway was eluci-dated through biochemical methods during the pastdecades. More recent genomic approaches allowedthe identification of the genes involved (DellaPennaand Pogson 2006). Homogentisic acid (HGA) is acommon substrate for the synthesis of tocopherols,tocotrienols and plastoquinones. HGA is synthe-sized from p-hydroxyphenylpyruvate via the cytoso-lic p-hydroxyphenylpyruvate dioxygenase (Garcıa et al.1997). The rest of enzymes are located in the innerchloroplast membrane. Isopentenyldiphosphate is usedfor the synthesis of geranylgeranyldiphosphate (GGDP)by GGDP synthase (Okada et al. 2000). GGDP reduc-tase reduces three out of the four double bonds ofGGDP to yield phytildiphosphate (PDP) (Keller et al.1998). Alternatively, PDP can be generated from phytoland ATP (Ischebeck et al. 2006). Homogentisate phytil-transferase (HPT) catalyzes the condensation of HGAand PDP to form 2-methyl-6-phytil-1,4-benzoquinol(MPBQ) (Collakova and DellaPenna 2001). MPBQcan be methylated to yield 2,3-dimethyl-6-phytil-1,4-benzoquinol (DMPBQ) by MPBQ methyltransferase(Marshall et al. 1985). MPBQ and DMPBQ are convertedto δ- and γ -tocopherol, respectively, by tocopherol
cyclase (Stocker et al. 1996). Finally, γ -tocopherolmethyltransferase (TMT) catalyzes methylation of δ- andγ - tocopherol to yield β- and α-tocopherol, respectively(D’Harlingue and Camara 1985).
A major protective function has always been attributedto tocopherols due to their antioxidant properties. Thechloroplast is a very important source of ROS inplants (Fryer et al. 2002). α-Tocopherol accumulatesin chloroplast membranes (Fryer 1992), and this factwas correlated with a putative role of α-tocopherol inthe maintenance of the photosynthetic apparatus (Trebstet al. 2002), though other tocopherols and also otherantioxidant molecules are involved (Munne-Bosch andFalk 2004). But tocopherols are found also in non-photosynthetic tissues, which suggests that tocopherolsmust play other functions non-related to the protectionof the photosynthetic apparatus. One of these functionscould be the prevention of the oxidation of unsaturatedfatty acids in stored lipids and in cell membranes(Munne-Bosch and Alegre 2002, Sattler et al. 2004),which is particularly important in the adaptation ofcells to growth at low temperature (Maeda et al. 2006).Other functions of tocopherols include the inhibition ofenzymatic activity, regulation of gene transcription andsignaling (Russin et al. 1996, Hofius et al. 2004, Sakuragiet al. 2006).
Tocopherols biosynthesis has been subject to a thor-ough analysis both at the molecular and physiologicallevel. Most of the genes involved in this process havealready been cloned and characterized in differentorganisms. In our laboratory, we have been studyingtocopherols biosynthesis in the unicellular green algaChlamydomonas reinhardtii for the last years. As part ofthis study, we approached the analysis of tmt1, the genecoding for TMT. A cDNA of this gene was already clonedand sequenced in our laboratory and the sequence wasdeposited in the Genbank under the accession numberAJ884948 before the sequencing of Chlamydomonasgenome was completed and published (Merchant et al.2007). In this paper, we present the overexpression oftmt1 in Escherichia coli and a characterization of therecombinant enzyme. We also analyze the effect of lightand temperature on the expression of this gene.
Materials and methods
Biological material and growth conditions
Chlamydomonas reinhardtii wild-type 6145c strain(mt−) was supplied by Dr Ruth Sager (Sydney FarberCancer Center, Boston). Cells were cultured in growthchambers either in liquid minimal media with 5% (v:v)CO2-enriched air or on solid Tris-acetate-phosphatemedia, both containing 10 mM ammonium chloride as
Physiol. Plant. 143, 2011 317

the only nitrogen source, as detailed elsewhere (Aguilaret al. 1991). The growth chamber was kept at 25◦Cunder constant illumination with day light provided byfluorescent tubes with an average photon irradiance of110 μmol m−2 s−1 at the center of the chamber.
RNA extraction and northern blot analysis
Chlamydomonas cells were recovered from a 10-mlculture by centrifugation at 3000 g for 5 min andresuspended in 500 μl extraction buffer A (50 mM Tris-HCl, pH 8.0, 5 mM EDTA, 300 mM NaCl). Sodiumdodecylsulphate (SDS) and proteinase K were addedto the suspension at a final concentration of 2% (w:v)and 40 μg ml−1, respectively, thoroughly mixed andincubated at 45◦C for 15 min. After three extractions withphenol:chloroform:isoamyl alcohol (25:24:1, v:v:v), theaqueous phase was recovered and RNA was precipitatedby adding one volume of 8 M LiCl and incubation at−20◦C for 14–16 h. RNA was pelleted by centrifugationat 15 000 g for 20 min at 4◦C, washed with 1 ml of2 M LiCl, redissolved in 100 μl sterile MilliQ waterand precipitated again by adding 0.1 volumes of 3 MNaAcO (pH 4.8) and 2.5 volumes of 95% ethanol. Afterthorough mixing and incubation at −20◦C for 2 h, RNAwas pelleted by centrifugation at 15 000 g for 20 min at4◦C. Finally, the RNA pellet was washed with 1 ml 70%ethanol, dried out at room temperature and redissolvedin 50 μl of sterile MilliQ water.
For northern blot analysis, samples containing 15 μgof total RNA were mixed with 25 μl deionizedformamide, 5 μl 10 × MOPS and 9 μl formaldehyde,and denatured by incubation at 65◦C for 15 min andquickly cooled down on ice. After RNA denaturing,samples were supplemented with the appropriateloading buffer and subjected to electrophoresis indenaturing formaldehyde agarose gel. Fractionated RNAwas transferred to nylon membranes, prehybridizedfor 2 h at 42◦C with hybridization solution (50%formamide, 5 × SSC, 50 mM sodium phosphate, pH7.0, 5 × Denhardt, 1% SDS and 0.1 mg ml−1 salmonsperm DNA), and hybridized overnight at 42◦C withfresh hybridization solution containing a probe that waspreviously denatured by boiling for 5 min. To make theprobe, a 406-bp fragment of tmt1 cDNA was amplifiedby PCR using TOCO11 (5′-GCG GGG CTG CTA TGGAG-3′) and TOCO12 (5′-GCG TGA TGC AGC CCGAAG ACA-3′) oligonucleotides, and later the antisensechain was labeled using TOCO12 oligonucleotide. Thisfragment contains part of the 3′-untranslated region fromnucleotide 1074 to nucleotide 1479 of tmt1 cDNA.Finally, photograph films were exposed to membranesin cassettes at −80◦C for the appropriate time beforebeing developed.
Construction of recombinant expression vectors
A cDNA containing the whole open reading frameof TMT1 was cloned by RT-PCR. Total RNA (3 μg)was subjected to reverse transcription with PowerScript(Clontech, Palo Alto, CA) following the manufacturer’sinstructions. The resulting total cDNA was used in PCRexperiments with specific oligonucleotides TOCO15(5′-AGA TTA AAA CAA AAC CGC TGA GAA-3′)and TOCO17 (5′-CCA GCC AAC TTG CAA TAGCCT CAG-3′). The sequence of TOCO17 primerwas directly determined after the sequence of anEST (AV38892) (Asamizu et al. 2000) that showeda significant similarity with Arabidopsis thaliana andSynechocystis PCC6803 γ -TMT. To obtain the sequenceof TOCO15, it was necessary to reconstruct the5′-sequence of Chlamydomonas tmt1 coding region byanalyzing the sequenced genome of C. reinhardtii. Thefollowing PCR program was used: initial denaturation at95◦C for 2 min, 35 cycles (95◦C, 30 s/60◦C, 1 min/72◦C,1.5 min), and a final extension step at 72◦C for 6 min.A single amplification product was obtained, clonedinto pGEM-T (Promega, Madison, WI) and double-strand sequenced. The sequence encoding TMT1 proteinwas amplified by PCR from the recombinant plasmidpGTMT1 using primers GEXTMT-5 (5′-GGT GGT GGAATT CTA ATG CCC AGC ACT GCG CTG-3′) andGEXTMT-3 (5′-TGA ATT AAG CTT CTA CTG CTGCAG CTT GCG-3′). GEXTMT-5 contained the first 18nucleotides of tmt1 coding region plus an upstreamsequence with a restriction site for EcoRI. GEXTMT-3included the last 15 nucleotides of the coding regionplus the stop codon and an upstream sequence witha Hind III restriction site. The following PCR programwas used: initial denaturation at 95◦C for 5 min, 30cycles (95◦C, 1 min/65◦C, 1 min/72◦C, 3 min), and afinal extension at 72◦C for 10 min. A 1044 bp fragmentwas amplified, cloned into pGEM-T and double-strandsequenced. The resulting plasmid was transformedinto E. coli DH5′αF′, positive colonies were isolatedand recombinant plasmids analyzed by digestion withrestriction enzymes to identify a recombinant plasmidcontaining the right insert, which was released by doubledigestion with EcoRI and Hind III and cloned into pGEX-KG to yield the recombinant plasmid pKGTMT1.
Expression and purification of recombinantTMT1 protein
The recombinant plasmid pKGTMT1 was introducedin the E. coli strain BL21 (DE3). A single colonyof a transformant containing this plasmid was usedto inoculate 2 ml of Luria-Bertani broth (LB) mediumand the culture was incubated overnight at 37◦C
318 Physiol. Plant. 143, 2011

in a rotary shaker (Aerotron, InforsHT, Bottmingen,Switzerland). One milliliter of this overnight culturewas used to inoculate 50 ml of fresh LB mediumand the new culture was incubated at 37◦C untilit reached an A600 of 0.5. At this point, isopropylβ-D-thiogalactopyranoside (IPTG) was added at a finalconcentration of 1 mM to induce the expression ofTMT1. After incubating the culture under the sameconditions for 4 h, it was cooled down on ice and cellswere recovered by centrifugation at 3000 g for 5 min,resuspended in 10 ml PBS, centrifuged again and finallyresuspended in 5 ml PBS supplemented with DNAase,RNAase, 1 mM phenylmethylsulfonyl fluoride (PMSF)and 10 mM ethylenediamine tetraacetate (EDTA). Cellswere disrupted by sonication with three pulses of 90 Wfor 5 s each with a Vibra Cell™ sonicator (Sonics &Materials, Newton, CT). The suspension of lysed bacteriawas centrifuged at 12 000 g for 10 min at 4◦C to discardthe insoluble fraction and recover the crude extract.A bacterial extract (4.5 ml) was chromatographed on aglutathione-agarose column. After convenient washingto discard contaminant proteins, GST-TMT1 fusionprotein was eluted with a buffer containing 50 mMTris–HCl (pH 8.0) and 10 mM reduced glutathione.Finally, approximately 1 mg of the fusion protein wastreated with 10 U of thrombin for 2 h. The thrombin-treated protein was passed through a glutathione-agarose column so that glutathione S-transferase andthe remaining non-lysed fusion protein were retained inthe column and pure TMT1 protein was eluted.
DNA sequencing and analysis
DNA Sequences were obtained with the ‘dye terminator’method using an ABI PRISM™ 373 automatic sequencer(Applied Biosystems, Foster City, CA). Sequence analysisand oligonucleotide design were done with the DNASTAR
software package. Sequence motifs were identifiedwith PLACE (http://www.dna.affrc.go.jp/PLACE/) at theNational Institute of Agrobiological Sciences, Japan.tmt1 cDNA sequence was deposited in the Genbank(accession number AJ884948).
Determination of enzymatic activities
γ -Tocopherol methyltransferase activity was assayed aspreviously described (D’Harlingue and Camara 1985)with minor modifications. The assay is based on themethylation of γ - or δ-tocopherol to yield α- orβ-tocopherol, respectively. A standard reaction bufferincluded 50 mM HEPES (pH 7.5), 5 mM DTT, 50 μMspecific substrate (γ - or δ-tocopherol), 50 μM S-adenosylmethionine (SAM). The reaction was initiated by addingthe appropriate amount of enzyme in a final volume
of 1 ml, and it was carried out for a convenienttime, after which it was stopped by adding 1 mlisooctane supplemented with BHT (1 mg ml−1). Aftermixing vigorously, the isooctane fraction was recoveredby centrifuging at 12 000 g for 2 min. The reactionproduct (α- or β-tocopherol) was determined by highperformance liquid chromatography (HPLC).
Tocopherols determination
Tocopherols were determined by HPLC on a SystemGold (Beckman, Madrid, Spain), equipped with a 126high pressure pump, a 20 μl Redone 7125 injectionvalve (Cotati, California) and a Jasco FP-920 fluorescencedetector (Jasco, Tokyo, Japan) with λexc = 294 nm andλem = 340 nm. Samples were chromatographed on aLichrospher 60 SI normal phase column (25 × 0.4 cmi.d.; Teknocroma, Barcelona, Spain) at a constant flowof 2 ml min−1 with a mobile phase of isooctane:ethylacetate (96.5:3.5, v:v). The column was kept at aconstant temperature of 35◦C with a Gecko 2000column heater (Inacom Instrument B.V., Veenendaal,The Netherlands) and the whole system was controlledwith a System Gold software package (Beckman).Actual concentration of tocopherols in samples wasdetermined as previously described (Mohamed et al.2007) considering the response factor of the tocopheroland the response factor of an internal standard.
Protein determination
Protein was determined according to Bradford (1976)with ‘Protein Assay’ kit (BioRad, Hercules, CA), usingbovine serum albumin (Sigma-Aldrich, St Louis, MO) asstandard.
Protein electrophoresis
Protein extracts were fractionated by polyacrylamide gelelectrophoresis (PAGE) in a Mini-Protean II Electrophore-sis Cell (BioRad) as previously described (Laemmli1970). For PAGE under denaturing conditions, both acry-lamide gels and electrophoresis buffer contained 0.1%SDS, and protein samples were subjected to a dena-turing treatment in Laemmli’s buffer at 95–100◦C for5 min before electrophoresis. High molecular weightSDS-PAGE markers (Sigma) were used: rabbit mus-cle myosin (205 kDa), β-galactosidase from E. coli(116 kDa), rabbit muscle phosphorylase b (97.4 kDa),bovine serum albumin (66 kDa), chicken ovalbumin(42.7 kDa) and carbonic anhydrase from bovine erythro-cytes (29 kDa). Protein Marker IV (Merck, Darmstadt,Germany) was also used: ovotransferrin (78 kDa), bovineserum albumin (66 kDa), chicken ovalbumin (42.7 kDa),
Physiol. Plant. 143, 2011 319

carbonic anhydrase from bovine erythrocytes (29 kDa),equine myoglobin (17 kDa) and equine cytochromec (12.3 kDa). For pore-gradient PAGE, the followingsize markers were used: Canavalia ensiformis urease(272 and 545 kDa), bovine serum albumin (66 and132 kDa) and chicken ovalbumin (45 kDa). After elec-trophoresis, protein bands were visualized by stainingwith Coomassie brilliant blue R-250. Alternatively, pro-teins were stained with silver nitrate (Heukeshoven andDernick 1985).
Results and discussion
Chlamydomonas TMT1 overexpressionand characterization
Lack of purified enzymes involved in the tocopherolpathway has restrained detailed analysis of the enzymesand the overall tocopherol synthesis and regulation.Earlier reports include the partial purification ofγ -TMT from bell pepper (Capsicum annuum) fruits(D’Harlingue and Camara 1985), spinach (Ishiko et al.1992) and Euglena (Shigeoka et al. 1992). Purificationto homogeneity was not possible for the bell pepper,Euglena and spinach enzymes because of the instabilityof the solubilized enzyme. This is the reason whyγ -TMTs from diverse origins have been overexpressedin E. coli (Koch et al. 2003). We have used the samestrategy to study Chlamydomonas γ -TMT. A cDNAcontaining the whole ORF of Chlamydomonas tmt1cDNA was isolated by RT-PCR. This cDNA was clonedinto pGEM-T and the resulting recombinant plasmid(pGTMT1) was sequenced. The largest ORF of this cDNAcodes for a 338 AA polypeptide with a molecular massof 36 791 Da. A sequence analysis with TARGETP Server(Emanuelsson et al. 2000) showed that the 27 aminoacids of TMT1 N-end constitute a transit peptide for itsimport into the plastid. So, after processing this transitpeptide, the functional protein would have 311 residueswith a molecular mass of 33 971 Da. The alignment ofTMT1 protein sequence with other TMTs from differentorigins revealed that TMT1 shares with all of themtwo fundamental motifs that represent an S-adenosylmethionine-binding domain (SAM) (Kagan and Clarke1994). A phylogenetic analysis revealed that at least atthe sequence level, C. reinhardtii TMT1 is closer to dicotplant TMTs (49.4% similarity with Brassica oleraceaTMT and 48.5% with Arabidopsis thaliana TMT) thanto bacterial TMTs (40.2% similarity with Synechocystissp. TMT).
Plasmid pKGTMT1 was used to express the Chlamy-domonas tmt1 cDNA in E. coli BL21 and cells carryingpGEX-KG were taken as a control. Bacterial strains
Fig. 1. Purification of recombinant TMT1. (A) An Escherichia colitransformant carrying plasmid pKGTMT1 was cultured at 37◦C andconstant shaking up to an absorbance of 0.5 at 600 nm. IPTG was addedat a final concentration of 1 mM and the culture was incubated for anadditional 4 h period. Cells were harvested, lysed by sonication andthe protein crude extract was subjected to affinity chromatography onglutathione-agarose. Samples representing every purification step wereanalyzed by 7.5% SDS-PAGE and Coomassie blue staining. Cr1, controlcrude extract of strain carrying pGEX-KG. Cr2, crude extract of straincarrying pKGTMT1. NR, proteins not retained by glutathione-agarose.L, proteins eluted after washing with PBS buffer. E, proteins eluted bywashing with 10 mM reduced glutathione in 50 mM Tris–HCl (pH 8.0).(B) A 1 mg sample of purified GST-TMT1 fusion protein (E) was digestedwith thrombin protease (T), and then subjected to glutathione-agarosechromatography to separate pure TMT1 from undigested fusion protein(TMT). Samples were analyzed by 10% SDS-PAGE and Coomassie bluestaining. Position and size of protein markers are shown.
carrying pKGTMT1 and pGEX-KG, respectively, werecultured and IPTG induced. Protein extracts of thesecell cultures were analyzed by SDS-PAGE and an addi-tional protein band of 63 kDa was detected in extractsfrom induced cells carrying pKGTMT1. TMT activitywas also assayed as previously described (D’Harlingueand Camara 1985) and this activity was specificallydetected in whole extracts of cells carrying pKGTMT1,thus demonstrating that Chlamydomonas tmt1 encodesa functional enzyme with TMT activity.
Chlamydomonas GST-TMT1 fusion protein waspurified by affinity chromatography on glutathione-agarose (Fig. 1A). The molecular mass of GST-TMT1fusion protein was estimated by SDS-PAGE and it was63 kDa (Fig. 1B). Considering that the GST moietyhas a molecular mass of 26 kDa, the molecularmass of TMT1 must be 37 kDa, as it was predictedby tmt1 cDNA sequence analysis. The size of therecombinant fusion protein GST-TMT1 in its nativeconformation was also determined by pore-gradient
320 Physiol. Plant. 143, 2011
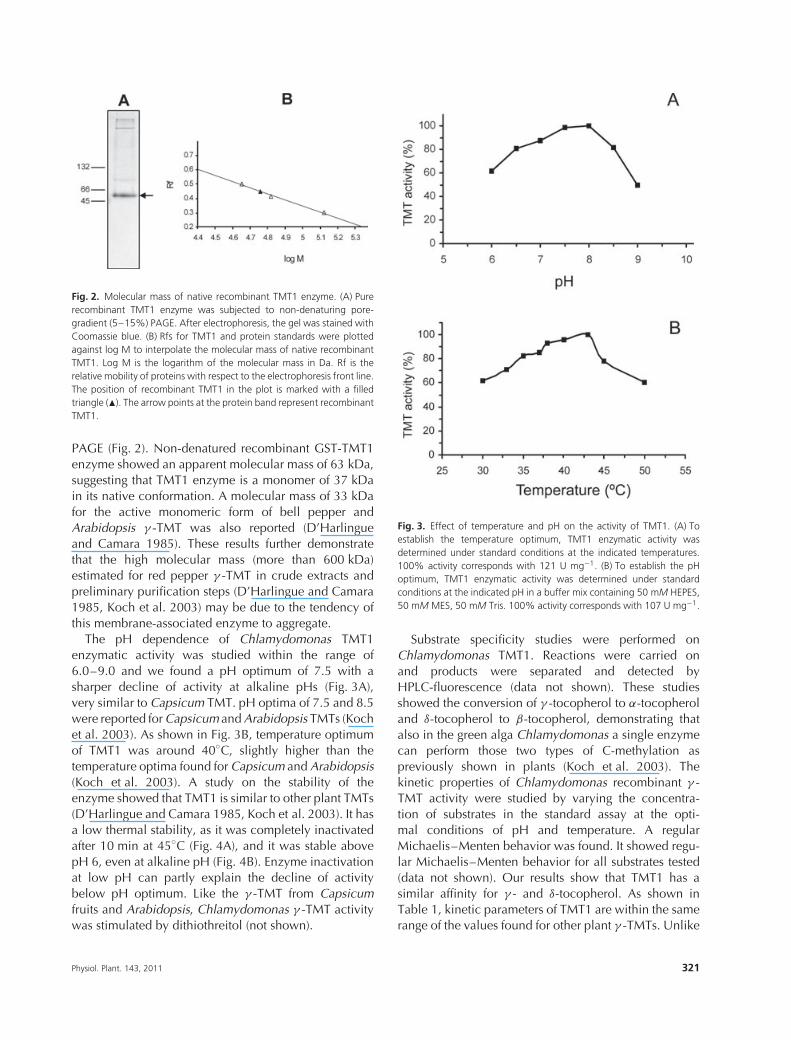
Fig. 2. Molecular mass of native recombinant TMT1 enzyme. (A) Purerecombinant TMT1 enzyme was subjected to non-denaturing pore-gradient (5–15%) PAGE. After electrophoresis, the gel was stained withCoomassie blue. (B) Rfs for TMT1 and protein standards were plottedagainst log M to interpolate the molecular mass of native recombinantTMT1. Log M is the logarithm of the molecular mass in Da. Rf is therelative mobility of proteins with respect to the electrophoresis front line.The position of recombinant TMT1 in the plot is marked with a filledtriangle (�). The arrow points at the protein band represent recombinantTMT1.
PAGE (Fig. 2). Non-denatured recombinant GST-TMT1enzyme showed an apparent molecular mass of 63 kDa,suggesting that TMT1 enzyme is a monomer of 37 kDain its native conformation. A molecular mass of 33 kDafor the active monomeric form of bell pepper andArabidopsis γ -TMT was also reported (D’Harlingueand Camara 1985). These results further demonstratethat the high molecular mass (more than 600 kDa)estimated for red pepper γ -TMT in crude extracts andpreliminary purification steps (D’Harlingue and Camara1985, Koch et al. 2003) may be due to the tendency ofthis membrane-associated enzyme to aggregate.
The pH dependence of Chlamydomonas TMT1enzymatic activity was studied within the range of6.0–9.0 and we found a pH optimum of 7.5 with asharper decline of activity at alkaline pHs (Fig. 3A),very similar to Capsicum TMT. pH optima of 7.5 and 8.5were reported for Capsicum and Arabidopsis TMTs (Kochet al. 2003). As shown in Fig. 3B, temperature optimumof TMT1 was around 40◦C, slightly higher than thetemperature optima found for Capsicum and Arabidopsis(Koch et al. 2003). A study on the stability of theenzyme showed that TMT1 is similar to other plant TMTs(D’Harlingue and Camara 1985, Koch et al. 2003). It hasa low thermal stability, as it was completely inactivatedafter 10 min at 45◦C (Fig. 4A), and it was stable abovepH 6, even at alkaline pH (Fig. 4B). Enzyme inactivationat low pH can partly explain the decline of activitybelow pH optimum. Like the γ -TMT from Capsicumfruits and Arabidopsis, Chlamydomonas γ -TMT activitywas stimulated by dithiothreitol (not shown).
Fig. 3. Effect of temperature and pH on the activity of TMT1. (A) Toestablish the temperature optimum, TMT1 enzymatic activity wasdetermined under standard conditions at the indicated temperatures.100% activity corresponds with 121 U mg−1. (B) To establish the pHoptimum, TMT1 enzymatic activity was determined under standardconditions at the indicated pH in a buffer mix containing 50 mM HEPES,50 mM MES, 50 mM Tris. 100% activity corresponds with 107 U mg−1.
Substrate specificity studies were performed onChlamydomonas TMT1. Reactions were carried onand products were separated and detected byHPLC-fluorescence (data not shown). These studiesshowed the conversion of γ -tocopherol to α-tocopheroland δ-tocopherol to β-tocopherol, demonstrating thatalso in the green alga Chlamydomonas a single enzymecan perform those two types of C-methylation aspreviously shown in plants (Koch et al. 2003). Thekinetic properties of Chlamydomonas recombinant γ -TMT activity were studied by varying the concentra-tion of substrates in the standard assay at the opti-mal conditions of pH and temperature. A regularMichaelis–Menten behavior was found. It showed regu-lar Michaelis–Menten behavior for all substrates tested(data not shown). Our results show that TMT1 has asimilar affinity for γ - and δ-tocopherol. As shown inTable 1, kinetic parameters of TMT1 are within the samerange of the values found for other plant γ -TMTs. Unlike
Physiol. Plant. 143, 2011 321
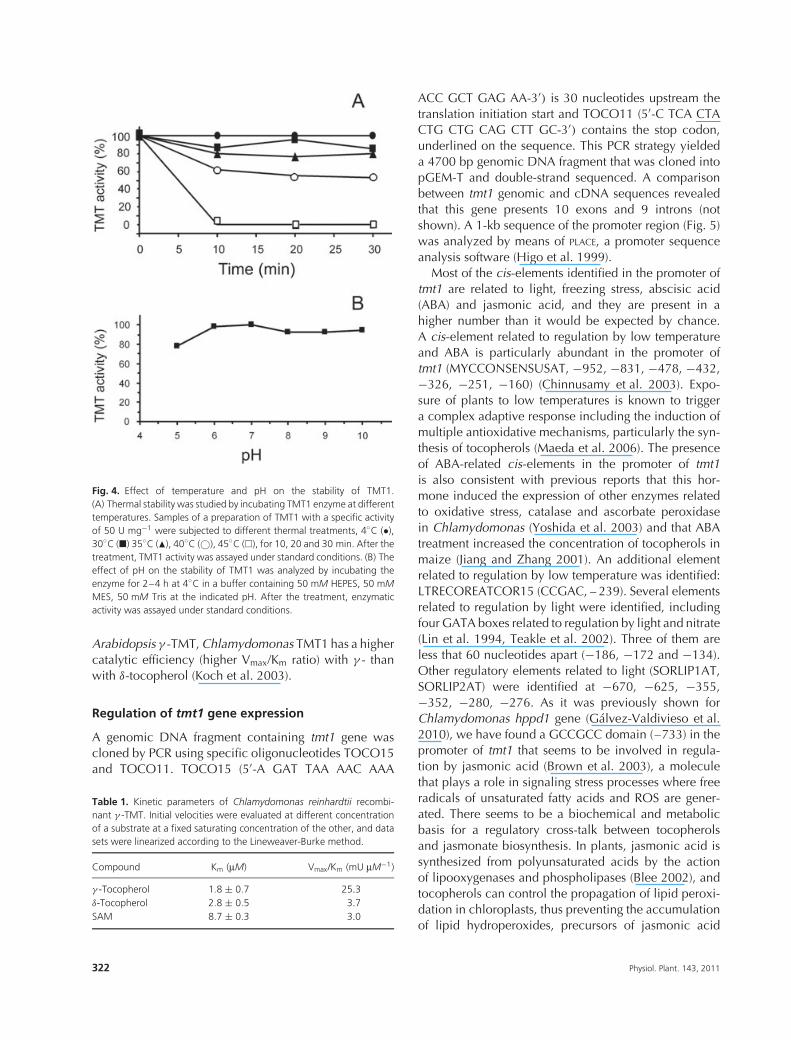
Fig. 4. Effect of temperature and pH on the stability of TMT1.(A) Thermal stability was studied by incubating TMT1 enzyme at differenttemperatures. Samples of a preparation of TMT1 with a specific activityof 50 U mg−1 were subjected to different thermal treatments, 4◦C (•),30◦C (�) 35◦C (�), 40◦C (©), 45◦C (�), for 10, 20 and 30 min. After thetreatment, TMT1 activity was assayed under standard conditions. (B) Theeffect of pH on the stability of TMT1 was analyzed by incubating theenzyme for 2–4 h at 4◦C in a buffer containing 50 mM HEPES, 50 mMMES, 50 mM Tris at the indicated pH. After the treatment, enzymaticactivity was assayed under standard conditions.
Arabidopsis γ -TMT, Chlamydomonas TMT1 has a highercatalytic efficiency (higher Vmax/Km ratio) with γ - thanwith δ-tocopherol (Koch et al. 2003).
Regulation of tmt1 gene expression
A genomic DNA fragment containing tmt1 gene wascloned by PCR using specific oligonucleotides TOCO15and TOCO11. TOCO15 (5′-A GAT TAA AAC AAA
Table 1. Kinetic parameters of Chlamydomonas reinhardtii recombi-nant γ -TMT. Initial velocities were evaluated at different concentrationof a substrate at a fixed saturating concentration of the other, and datasets were linearized according to the Lineweaver-Burke method.
Compound Km (μM) Vmax/Km (mU μM−1)
γ -Tocopherol 1.8 ± 0.7 25.3δ-Tocopherol 2.8 ± 0.5 3.7SAM 8.7 ± 0.3 3.0
ACC GCT GAG AA-3′) is 30 nucleotides upstream thetranslation initiation start and TOCO11 (5′-C TCA CTACTG CTG CAG CTT GC-3′) contains the stop codon,underlined on the sequence. This PCR strategy yieldeda 4700 bp genomic DNA fragment that was cloned intopGEM-T and double-strand sequenced. A comparisonbetween tmt1 genomic and cDNA sequences revealedthat this gene presents 10 exons and 9 introns (notshown). A 1-kb sequence of the promoter region (Fig. 5)was analyzed by means of PLACE, a promoter sequenceanalysis software (Higo et al. 1999).
Most of the cis-elements identified in the promoter oftmt1 are related to light, freezing stress, abscisic acid(ABA) and jasmonic acid, and they are present in ahigher number than it would be expected by chance.A cis-element related to regulation by low temperatureand ABA is particularly abundant in the promoter oftmt1 (MYCCONSENSUSAT, −952, −831, −478, −432,−326, −251, −160) (Chinnusamy et al. 2003). Expo-sure of plants to low temperatures is known to triggera complex adaptive response including the induction ofmultiple antioxidative mechanisms, particularly the syn-thesis of tocopherols (Maeda et al. 2006). The presenceof ABA-related cis-elements in the promoter of tmt1is also consistent with previous reports that this hor-mone induced the expression of other enzymes relatedto oxidative stress, catalase and ascorbate peroxidasein Chlamydomonas (Yoshida et al. 2003) and that ABAtreatment increased the concentration of tocopherols inmaize (Jiang and Zhang 2001). An additional elementrelated to regulation by low temperature was identified:LTRECOREATCOR15 (CCGAC, – 239). Several elementsrelated to regulation by light were identified, includingfour GATA boxes related to regulation by light and nitrate(Lin et al. 1994, Teakle et al. 2002). Three of them areless that 60 nucleotides apart (−186, −172 and −134).Other regulatory elements related to light (SORLIP1AT,SORLIP2AT) were identified at −670, −625, −355,−352, −280, −276. As it was previously shown forChlamydomonas hppd1 gene (Galvez-Valdivieso et al.2010), we have found a GCCGCC domain (–733) in thepromoter of tmt1 that seems to be involved in regula-tion by jasmonic acid (Brown et al. 2003), a moleculethat plays a role in signaling stress processes where freeradicals of unsaturated fatty acids and ROS are gener-ated. There seems to be a biochemical and metabolicbasis for a regulatory cross-talk between tocopherolsand jasmonate biosynthesis. In plants, jasmonic acid issynthesized from polyunsaturated acids by the actionof lipooxygenases and phospholipases (Blee 2002), andtocopherols can control the propagation of lipid peroxi-dation in chloroplasts, thus preventing the accumulationof lipid hydroperoxides, precursors of jasmonic acid
322 Physiol. Plant. 143, 2011

Fig. 5. Regulatory elements of the tmt1 gene. The sequence covers about 1000 nt of the promoter. Sequences that match consensus motifs ofcis-regulatory elements are underlined. Element related to regulation by low temperature: MYCCONSENSUSAT (CANNTG), LTRECOREATCOR15(CCGAC). Elements related to regulation by light: GATABOX (GATA), SORLIP1AT (GCCAC), SORLIP2AT (GGGCC). Element related to regulation byjasmonic acid ( ). Element that binds Ca2+/calmodulin: (CGCGBOXAT, ). Element involved in regulation by carbon metabolism(CACTFTPPCA1, ). Translation start (ATG) is in bold.
(Schaller 2001). Interestingly, there are recent evidencesthat tocopherols might influence stress signaling by reg-ulating jasmonic acid levels in plants (Munne-Boschet al. 2007). In addition, Arabidopsis tyrosin aminotrans-ferase (enzyme that precedes HPPD in the pathway) andbarley hppd are positively regulated by jasmonic acid(Falk et al. 2002, Sandorf and Hollander-Czytko 2002).A cis-element that binds Ca2+/calmodulin, involved inmany signaling processes, was also identified (CGCG-BOXAT, −1033, −1028, −1026, −922, −920, −863,−846, −717, −703, −697, −613, −566, −506) (Yangand Poovaiah 2002). We also found several motifsrelated with the regulation of genes involved in carbonmetabolism (CACTFTPPCA1, −770, −649, −422, −285,−184, −112) (Gowik et al. 2004). This reinforces theidea that tocopherols might also be regulatory molecules.
Overall, the analysis of tmt1 promoter sequencesuggests that transcriptional regulation of this geneis related with light and stress, particularly with lowtemperature, what helps to better understand the
functions of γ - and α-tocopherol, the end-products ofthe anabolic pathway where tmt1 is integrated.
Northern blot analyses were performed on samples ofChlamydomonas cultures subjected to different growthconditions to study the effect of light and temperatureon the expression of tmt1 at the RNA level. To avoidpossible artifacts related to cell culture growth duringthe experiments, we used cell cultures at an earlystationary phase. Regulation by light was studied bychanging culture conditions from low to moderatephoton irradiance and from moderate photon irradianceto darkness. Cells were cultured up to the beginning ofthe stationary phase in MM-A medium at 25◦C with aphoton irradiance of 30 μmol m−2 s−1. When these cellcultures were transferred from low photon irradiance(30 μmol m−2 s−1) to moderate photon irradiance(110 μmol m−2 s−1), we observed a fast induction oftmt1 to double the initial level (Fig. 6A). On the otherhand, when cells were transferred from moderate photonirradiance to darkness, a significant 70% reduction of
Physiol. Plant. 143, 2011 323
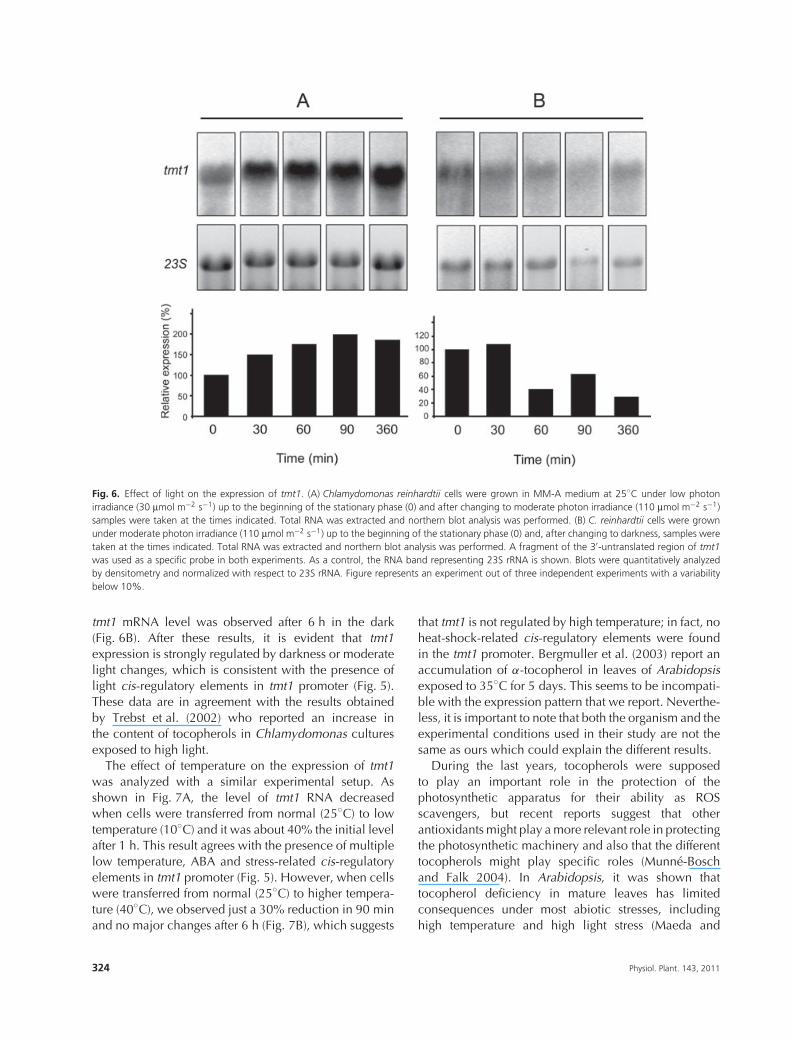
Fig. 6. Effect of light on the expression of tmt1. (A) Chlamydomonas reinhardtii cells were grown in MM-A medium at 25◦C under low photonirradiance (30 μmol m−2 s−1) up to the beginning of the stationary phase (0) and after changing to moderate photon irradiance (110 μmol m−2 s−1)samples were taken at the times indicated. Total RNA was extracted and northern blot analysis was performed. (B) C. reinhardtii cells were grownunder moderate photon irradiance (110 μmol m−2 s−1) up to the beginning of the stationary phase (0) and, after changing to darkness, samples weretaken at the times indicated. Total RNA was extracted and northern blot analysis was performed. A fragment of the 3′-untranslated region of tmt1was used as a specific probe in both experiments. As a control, the RNA band representing 23S rRNA is shown. Blots were quantitatively analyzedby densitometry and normalized with respect to 23S rRNA. Figure represents an experiment out of three independent experiments with a variabilitybelow 10%.
tmt1 mRNA level was observed after 6 h in the dark(Fig. 6B). After these results, it is evident that tmt1expression is strongly regulated by darkness or moderatelight changes, which is consistent with the presence oflight cis-regulatory elements in tmt1 promoter (Fig. 5).These data are in agreement with the results obtainedby Trebst et al. (2002) who reported an increase inthe content of tocopherols in Chlamydomonas culturesexposed to high light.
The effect of temperature on the expression of tmt1was analyzed with a similar experimental setup. Asshown in Fig. 7A, the level of tmt1 RNA decreasedwhen cells were transferred from normal (25◦C) to lowtemperature (10◦C) and it was about 40% the initial levelafter 1 h. This result agrees with the presence of multiplelow temperature, ABA and stress-related cis-regulatoryelements in tmt1 promoter (Fig. 5). However, when cellswere transferred from normal (25◦C) to higher tempera-ture (40◦C), we observed just a 30% reduction in 90 minand no major changes after 6 h (Fig. 7B), which suggests
that tmt1 is not regulated by high temperature; in fact, noheat-shock-related cis-regulatory elements were foundin the tmt1 promoter. Bergmuller et al. (2003) report anaccumulation of α-tocopherol in leaves of Arabidopsisexposed to 35◦C for 5 days. This seems to be incompati-ble with the expression pattern that we report. Neverthe-less, it is important to note that both the organism and theexperimental conditions used in their study are not thesame as ours which could explain the different results.
During the last years, tocopherols were supposedto play an important role in the protection of thephotosynthetic apparatus for their ability as ROSscavengers, but recent reports suggest that otherantioxidants might play a more relevant role in protectingthe photosynthetic machinery and also that the differenttocopherols might play specific roles (Munne-Boschand Falk 2004). In Arabidopsis, it was shown thattocopherol deficiency in mature leaves has limitedconsequences under most abiotic stresses, includinghigh temperature and high light stress (Maeda and
324 Physiol. Plant. 143, 2011
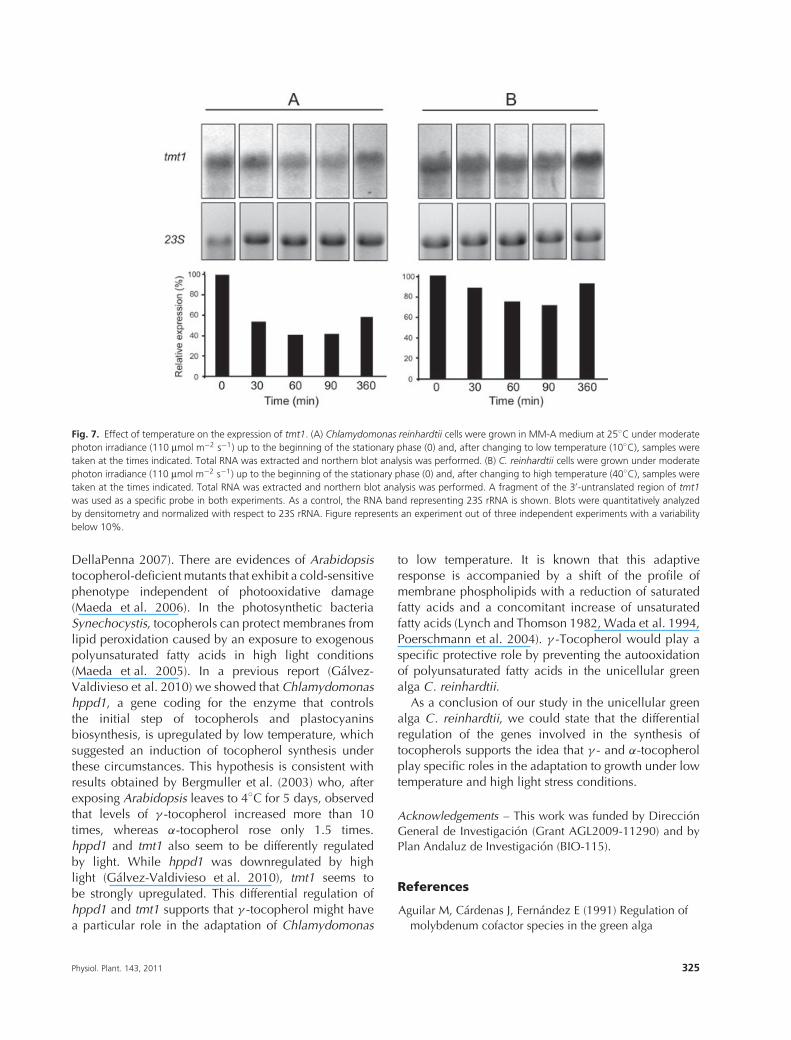
Fig. 7. Effect of temperature on the expression of tmt1. (A) Chlamydomonas reinhardtii cells were grown in MM-A medium at 25◦C under moderatephoton irradiance (110 μmol m−2 s−1) up to the beginning of the stationary phase (0) and, after changing to low temperature (10◦C), samples weretaken at the times indicated. Total RNA was extracted and northern blot analysis was performed. (B) C. reinhardtii cells were grown under moderatephoton irradiance (110 μmol m−2 s−1) up to the beginning of the stationary phase (0) and, after changing to high temperature (40◦C), samples weretaken at the times indicated. Total RNA was extracted and northern blot analysis was performed. A fragment of the 3′-untranslated region of tmt1was used as a specific probe in both experiments. As a control, the RNA band representing 23S rRNA is shown. Blots were quantitatively analyzedby densitometry and normalized with respect to 23S rRNA. Figure represents an experiment out of three independent experiments with a variabilitybelow 10%.
DellaPenna 2007). There are evidences of Arabidopsistocopherol-deficient mutants that exhibit a cold-sensitivephenotype independent of photooxidative damage(Maeda et al. 2006). In the photosynthetic bacteriaSynechocystis, tocopherols can protect membranes fromlipid peroxidation caused by an exposure to exogenouspolyunsaturated fatty acids in high light conditions(Maeda et al. 2005). In a previous report (Galvez-Valdivieso et al. 2010) we showed that Chlamydomonashppd1, a gene coding for the enzyme that controlsthe initial step of tocopherols and plastocyaninsbiosynthesis, is upregulated by low temperature, whichsuggested an induction of tocopherol synthesis underthese circumstances. This hypothesis is consistent withresults obtained by Bergmuller et al. (2003) who, afterexposing Arabidopsis leaves to 4◦C for 5 days, observedthat levels of γ -tocopherol increased more than 10times, whereas α-tocopherol rose only 1.5 times.hppd1 and tmt1 also seem to be differently regulatedby light. While hppd1 was downregulated by highlight (Galvez-Valdivieso et al. 2010), tmt1 seems tobe strongly upregulated. This differential regulation ofhppd1 and tmt1 supports that γ -tocopherol might havea particular role in the adaptation of Chlamydomonas
to low temperature. It is known that this adaptiveresponse is accompanied by a shift of the profile ofmembrane phospholipids with a reduction of saturatedfatty acids and a concomitant increase of unsaturatedfatty acids (Lynch and Thomson 1982, Wada et al. 1994,Poerschmann et al. 2004). γ -Tocopherol would play aspecific protective role by preventing the autooxidationof polyunsaturated fatty acids in the unicellular greenalga C . reinhardtii.
As a conclusion of our study in the unicellular greenalga C . reinhardtii, we could state that the differentialregulation of the genes involved in the synthesis oftocopherols supports the idea that γ - and α-tocopherolplay specific roles in the adaptation to growth under lowtemperature and high light stress conditions.
Acknowledgements – This work was funded by DireccionGeneral de Investigacion (Grant AGL2009-11290) and byPlan Andaluz de Investigacion (BIO-115).
References
Aguilar M, Cardenas J, Fernandez E (1991) Regulation ofmolybdenum cofactor species in the green alga
Physiol. Plant. 143, 2011 325

Chlamydomonas reinhardtii. Biochim Biophys Acta1073: 463–469
Asamizu E, Miura K, Kucho K, Inoue Y, Fukuzawa H,Ohyama K, Nakamura Y, Tabata S (2000) Generation ofexpressed sequence tags from low-CO2 and high-CO2
adapted cells of Chlamydomonas reinhardtii. DNA Res7: 305–307
Bergmuller W, Porfirova S, Dormann P (2003)Characterization of an Arabidopsis mutant deficient inγ -tocopherol methyltransferase. Plant Mol Biol 52:1181–1190
Blee E (2002) Impact of phyto-oxylipins in plant defense.Trends Plant Sci 7: 315–322
Bolwell GP (1999) Role of active oxygen species and NOin plant defense responses. Curr Opin Plant Biol 2:287–294
Bray CM, West CE (2005) DNA repair mechanisms inplants: crucial sensors and effectors for the maintenanceof genome integrity. New Phytol 168: 511–28
Brown RL, Kazan K, McGrath KC, Maclean DJ,Manners JM (2003) A role for the GCC-box injasmonate-mediated activation of the PDF1.2 gene ofArabidopsis. Plant Physiol 132: 1020–1032
Chinnusamy V, Ohta M, Kanrar S, Lee BH, Hong X,Agarwal M, Zhu JK (2003) ICE1: a regulator ofcold-induced transcriptome and freezing tolerance inArabidopsis. Genes Dev 17: 1043–1054
Collakova E, DellaPenna D (2001) Isolation and functionalanalysis of homogentisate phytyltransferase fromSynechocystis sp. PCC 6803 and Arabidopsis. PlantPhysiol 127: 1113–1124
D’Harlingue A, Camara B (1985) Plastid enzymes ofterpenoid biosynthesis. Purification and characterizationof gamma-tocopherol methyltransferase from Capsicumchromoplasts. J Biol Chem 260: 15200–15203
DellaPenna D, Pogson BJ (2006) Vitamin synthesis inplants: tocopherols and carotenoids. Ann Rev Plant Biol57: 711–738
Emanuelsson O, Nielsen H, Brunak S, von Heijne G(2000) Predicting subcellular localization of proteinsbased on their N-terminal amino acid sequence. J MolBiol 300: 1005–1016
Falk J, Kraub N, Dahnhardt D, Krupinska K (2002) Thesenescence associated gene of barley encoding4-hydroxyphenylpyruvate dioxygenase isexpressed during oxidative stress. J Plant Biol 159:1245–1253
Fryer MJ (1992) The antioxidant effects of thylakoidvitamin E (α-tocoferol). Plant Cell Environ 15: 381–392
Fryer MJ, Oxborough K, Mullineaux PM, Baker NR (2002)Imaging of photo-oxidative stress responses in leaves. JExp Bot 53: 1249–1254
Galvez-Valdivieso G, Pineda M, Aguilar M (2010)Functional characterization and expression analysis ofp-hydroxyphenylpyruvate dioxygenase from the green
alga Chlamydomonas reinhardtii (Chlorophyta). J Phycol46: 297–308
Garcıa I, Rodgers M, Lenne C, Rolland A, Sailland A,Matringe M (1997) Subcellular localization andpurification of a p-hydroxyphenylpyruvate dioxygenasefrom cultured carrot cells and characterization of thecorresponding cDNA. Biochem J 325: 761–769
Gechev T, Willekens H, Van Montagu M, Inze D, VanCamp W, Toneva V, Minkov (2003) Different responsesof tobacco antioxidant enzymes to light and chillingstress. J Plant Physiol 160: 509–515
Gowik U, Burscheidt J, Akyildiz M, Schlue U, Koczor M,Streubel M, Westhoff P (2004) cis-Regulatory elementsfor mesophyll-specific gene expression in the C4 plantFlaveria trinervia, the promoter of the C4phosphoenolpyruvate carboxylase gene. Plant Cell 16:1077–1090
Herrera E, Barbas C (2001) Vitamin E: action, metabolismand perspectives. J Physiol Biochem 57: 43–56
Heukeshoven J, Dernick R (1985) Simplified method forsilver staining of proteins in polyacrylamide gels and themechanisms of silver staining. Electrophoresis 6:103–112
Higo K, Ugawa Y, Iwamoto M, Korenaga T (1999) Plantcis-acting regulatory DNA elements (PLACE) database.Nucl Acids Res 27: 297–300
Hofius D, Hajirezaei MR, Geiger M, Tschiersch H,Melzer M, Sonnewald U (2004) RNAi-mediatedtocopherol deficiency impairs photoassimilate exportin transgenic potato plants. Plant Physiol 135:1256–1268
Hurst AC, Grams TEE, Ratajczak R (2004) Effects ofsalinity, high irradiance, ozone, and ethylene on modeof photosynthesis, oxidative stress and oxidative damagein the C-3/CAM intermediate plant Mesembryanthemumcrystallinum L. Plant Cell Environ 27: 187–197
Ischebeck T, Zbierzak AM, Kanwischer M, Dormann P(2006) A salvage pathway for phytol metabolism inArabidopsis. J Biol Chem 281: 2470–2477
Ishiko H, Shigeoka S, Nakano Y, Mitsunaga T (1992) Someproperties of γ -tocopherol methyltransferase solubilizedfrom spinach chloroplasts. Phytochemistry 31:1499–1500
Jiang Q, Zhang J (2001) Effect of abscisic acid on activeoxygen species, antioxidative defense system andoxidative damage in leaves of maize seedlings. PlantCell Physiol 42: 1265–1273
Kagan RM, y Clarke S (1994) Widespread occurrence ofthree sequence motifs in diverseS-adenosylmethionine-dependent methyltransferasessuggests a common structure for these enzymes. ArchBiochem Biophys 310: 417–427
Kehrer JP, Smith CS (1994) Free radicals in biology:sources, reactivities, and roles in the etiology of humandiseases. In: Frei B (ed) Natural Antioxidants in Human
326 Physiol. Plant. 143, 2011

Health and Disease. Academic Press, San Diego, pp25–62
Keller Y, Bouvier F, d’Harlingue A, Camara B (1998)Metabolic compartmentation of plastid prenyllipidbiosynthesis – evidence for the involvement of amultifunctional geranylgeranyl reductase. Eur J Biochem251: 413–417
Koch M, Lemke R, Heise K-P, Mock H-P (2003)Characterization of γ -tocopherol methyltransferasesfrom Capsicum annuum L and Arabidopsis thaliana. EurJ Biochem 270: 84–92
Laemmli UK (1970) Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature227: 680–685
Lin Y, Hwang C-F, Brown JB, y Cheng C-L (1994) 50proximal regions of Arabidopsis nitrate reductase genesdirect nitrate-induced transcription in transgenictobacco. Plant Physiol 106: 477–484
Lynch DV, Thompson GA (1982) Low temperature-induced alterations in the chloroplast and microsomalmembranes of Dunaliella salina. Plant Physiol 69:1369–1375
Maeda H, DellaPenna D (2007) Tocopherol functions inphotosynthetic organisms. Curr Opin Plant Biol 10:260–265
Maeda H, Sakuragi Y, Bryant DA, Dellapenna D (2005)Tocopherols protect Synechocystis sp. strain PCC 6803from lipid peroxidation. Plant Physiol 138: 1422–1435
Maeda H, Songa W, Sagee TL, DellaPenna D (2006)Tocopherols play a crucial role in low-temperatureadaptation and phloem loading in Arabidopsis. PlantCell 18: 2710–2732
Mallick N, Mohn FH (2000) Reactive oxygen species:response of algal cells. J Plant Physiol 157: 183–193
Marshall PS, Morris SR, Threlfall DR (1985) Biosynthesis oftocopherols: a re-examination of the biosynthesis andmetabolism of 2-demethyl-6-phytyl-1,4-benzoquinol.Phytochemistry 24: 1705–1711
Merchant SS, Prochnik SE, Vallon O, Harris EH,Karpowicz SJ, Witman GB, Terry A, Salamov A,Fritz-Laylin LK, Marechal-Drouard L, Marshall WF, QuLH, Nelson DR, Sanderfoot AA, SpaldingMH, Kapitonov VV, Ren Q, Ferris P, Lindquist E,Shapiro H, Lucas SM, Grimwood J, Schmutz J, Cardol P,Cerutti H, Chanfreau G, Chen CL, Cognat V, Croft MT,Dent R, Dutcher S, Fernandez E, Fukuzawa H,Gonzalez-Ballester D, Gonzalez-Halphen D,Hallmann A, Hanikenne M, Hippler M, Inwood W,Jabbari K, Kalanon M, Kuras R, Lefebvre PA,Lemaire SD, Lobanov AV, Lohr M, Manuell A, Meier I,Mets L, Mittag M, Mittelmeier T, Moroney JV, Moseley J,Napoli C, Nedelcu AM, Niyogi K, Novoselov SV,Paulsen IT, Pazour G, Purton S, Ral JP, Riano-PachonDM, Riekhof W, Rymarquis L, Schroda M, Stern D,Umen J, Willows R, Wilson N, Zimmer SL, Allmer J,
Balk J, Bisova K, Chen CJ, Elias M, Gendler K, Hauser C,Lamb MR, Ledford H, Long JC, Minagawa J, Page MD,Pan J, Pootakham W, Roje S, Rose A, Stahlberg E,Terauchi AM, Yang P, Ball S, Bowler C, Dieckmann CL,Gladyshev VN, Green P, Jorgensen R, Mayfield S,Mueller-Roeber B, Rajamani S, Sayre RT, Brokstein P,Dubchak I, Goodstein D, Hornick L, Huang YW,Jhaveri J, Luo Y, Martınez D, Ngau WC, Otillar B,Poliakov A, Porter A, Szajkowski L, Werner G, Zhou K,Grigoriev IV, Rokhsar DS, Grossman AR (2007) TheChlamydomonas genome reveals the evolution ofkey animal and plant functions. Science 318: 245–250
Mohamed R, Pineda M, Aguilar M (2007) Antioxidantcapacity of extracts from wild and crop plants of theMediterranean region. J Food Sci 72: S59–S63
Mullineaux P, Karpinski S (2002) Signal transduction inresponse to excess light: getting out of the chloroplast.Curr Opin Plant Biol 5: 43–48
Munne-Bosch S, Alegre L (2002) The function oftocopherols and tocotrienols in plants. Crit Rev Plant Sci21: 31–57
Munne-Bosch S, Falk J (2004) New insights into thefunction of tocopherols in plants. Planta 218: 323–326
Munne-Bosch S, Weiler EW, Alegre L, Muller M, DuchtingP, Falk J (2007) Alpha-tocopherol may influence cellularsignaling by modulating jasmonic acid levels in plants.Planta 225: 681–91
Noctor G, Foyer CH (1998) Ascorbate and glutathione:keeping active oxygen under control. Annu Rev PlantPhysiol Plant Mol Biol 49: 249–279
Okada K, Saito T, Nakagawa T, Kawamukai M, Kamiya Y(2000) Five geranylgeranyl diphosphate synthasesexpressed in different organs are localized into threesubcellular compartments in Arabidopsis. Plant Physiol122: 1045–1056
Poerschmann J, Spijkerman E, Langer U (2004) Fatty acidpatterns in Chlamydomonas sp. as a marker fornutritional regimes and temperature under extremelyacidic conditions. Microbial Ecol 48: 78–89
Russin WA, Evert RF, Vanderveer PJ, Sharkey TD,Briggs SP (1996) Modification of a specific class ofplasmodesmata and loss of sucrose export ability in thesucrose export defective1 maize mutant. Plant Cell 8:654–658
Sakuragi Y, Maeda H, Dellapenna D, Bryant DA (2006)Alpha-tocopherol plays a role in photosynthesis andmacronutrient homeostasis of the cyanobacteriumSynechocystis sp. PCC 6803 that is independentof its antioxidant function. Plant Physiol 141:508–521
Sandorf I, Holander-Czytko H (2002) Jasmonate isinvolved in the induction of tyrosin aminotransferaseand tocopherol biosynthesis in Arabidopsis thaliana.Planta 216: 173–179
Physiol. Plant. 143, 2011 327

Sattler SE, Gilliland LU, Magallanes-Lundback M,Pollard M, DellaPenna D (2004) Vitamin E is essentialfor seed longevity and for preventing lipid peroxidationduring germination. Plant Cell 16: 1419–1432
Scandalios JG (1994) Regulation and properties of plantcatalases. In: Foyer CH, Mullineaux P (eds) Causes ofPhotooxidative Stress and Amelioration of DefenseSystem in Plants. CRC Press Inc, Boca Raton, pp275–315
Schaller F (2001) Enzymes of the biosynthesis ofoctadecanoid-derived signaling molecules. J Exp Bot 52:11–23
Shigeoka S, Ishiko H, Nakano Y, Mitsunaga T (1992)Isolation and properties of γ -tocopherolmethyltransferase in Euglena gracilis. Biochim BiophysActa 1128: 220–226
Sies H (1991) Oxidative stress: Introduction. In: Sies H (ed)Oxidative Stress: Oxidants and Antioxidants. AcademicPress, San Diego, pp 15–22
Stocker A, Fretz H, Frick H, Ruttimann A, Woggon WD(1996) The substrate specificity of tocopherol cyclase.Bioorg Med Chem 4: 1129–1134
Teakle GR, Manfield IW, Graham JF, y Gilmartin PM(2002) Arabidopsis thaliana GATA factors: organisation,
expression and DNA-binding characteristics. Plant MolBiol 50: 43–57
Trebst A, Depka B, Hollander-Czytko H (2002) A specificrole for tocopherol and of chemical singlet oxygenquencher in the maintenance of photosystem II structureand function in Chlamydomonas reinhardtii. FEBS Lett516: 156–160
Wada H, Gombos Z, Murata N (1994) Contribution ofmembrane lipids to the ability of the photosyntheticmachinery to tolerate temperature stress. Proc Natl AcadSci USA 91: 4273–4277
Wojtaszek P (1997) Oxidative burst: an early plantresponse to pathogen infection. Biochem J 322:681–692
Yang T, Poovaiah BW (2002) A calmodulin-binding/CGCG box DNA-binding protein familyinvolved in multiple signaling pathways in plants. J BiolChem 277: 45049–45058
Yoshida K, Igarashi E, Mukai M, Hirata K, Miyamoto K(2003) Induction of tolerance to oxidative stress in thegreen alga, Chlamydomonas reinhardtii, by abscisicacid. Plant Cell Environ 26: 451–457
Edited by V. Shulaev
328 Physiol. Plant. 143, 2011
Related Documents











