FACULTY OF SCIENCE UNIVERSITY OF COPENHAGEN PhD thesis Lea Ellegaard-Jensen Fungal degradation of pesticides - construction of microbial consortia for bioremediation Academic advisor: Søren Rosendahl Submitted: November 2012 The PhD School of Science, Faculty of Science, University of Copenhagen, Denmark.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

F A C U L T Y O F S C I E N C E
U N I V E R S I T Y O F C O P E N H A G E N
PhD thesis
Lea Ellegaard-Jensen
Fungal degradation of pesticides
- construction of microbial consortia for bioremediation
Academic advisor: Søren Rosendahl
Submitted: November 2012 The PhD School of Science, Faculty of Science, University of Copenhagen, Denmark.

II
Name of department: Department of Biology Author: Lea Ellegaard-Jensen Title: Fungal degradation of pesticides - construction of
microbial consortia for bioremediation Academic advisor: Søren Rosendahl Submitted: November 2012

III
Preface
This thesis presents the results of my three years as a PhD student at the section of Terrestrial
Ecology at the University of Copenhagen. The PhD was part of the project on Microbial
Remediation of Contaminated Soil and Water Resources (MIRESOWA) funded by the Danish
Council for Strategic Research. The MIRESOWA project provided excellent opportunities for
collaboration between partners, and the research presented in this thesis was especially a result of
close collaboration between the section of Terrestrial Ecology at University of Copenhagen and the
department of Geochemistry at the Geological Survey of Denmark and Greenland (GEUS). The
work was therefore carried out both at the University of Copenhagen and at GEUS.
Acknowledgments
First of all, I wish to thank my two supervisors Søren Rosendahl and Jens Aamand for invaluable
guidance, support and also thoughtfulness during rough periods. I am truly grateful to you both for
the great experience and pleasure it has been working on this project: A project with bioremediation
as the core focus, which is one of my favorite scientific subjects.
I also want to thank all the kind people at the section of Terrestrial Ecology, many of whom I
already knew well from my years as a master student – starting my PhD work at the section really
made me feel at home though in a new role. Special thanks go to the very kind technician Lis and to
my fellow PhD students in the Journal Club for including me even though I spend much of my time
at GEUS.
To all the people at the department of Geochemistry at GEUS I am grateful for making me feel
welcome and for a great working environment. I appreciate the way that everyone there takes the
time to listen and discuss problems of both scientific and non-scientific nature. My great thanks go
to the skillful and caring technicians at GEUS Spire and Pia. I also wish to thank my fellow PhD
students (at the time): Meric, Erkin, Tina, Nora, Christian, Sarka and of course above all Berith.
Starting my PhD I knew that it was predetermined that 1/3 of the work should be in collaboration
with another PhD student on creating consortia of fungi and bacteria, which could create quite a
challenge. Amazingly this turned out to be not only one of the most interesting scientific parts, but

IV
also yielded lots of very enjoyable hours in the laboratory (and also outside it…) thanks to the way
two minds think alike.
I would also like to thank all the MIRESOWA partners for fruitful discussions and collaborations in
particular at our seminars and scientific meetings. Special thanks go to Anders Johansen from the
department of Environmental Science at Aarhus University and to Birthe B. Kragelund from the
Structural Biology and NMR Laboratory at the University of Copenhagen. Birthe thank you for
letting me visit and work in your laboratory.
Last but not least, I am very thankful to my family for all their support, and especially Michael and
Noa for their every-day loving support and understanding!
Lea Ellegaard-Jensen
Copenhagen, November 2012

Table of Contents
Preface ..................................................................................................................................... III
Acknowledgments ............................................................................................................................ III
List of manuscripts ..................................................................................................................... 3
English summary ........................................................................................................................ 5
Dansk resumé ............................................................................................................................ 8
Introduction and aims .............................................................................................................. 11
Background .............................................................................................................................. 13
Pesticide pollution – why should we care .......................................................................................... 13 Diuron, dichlobenil and BAM .................................................................................................................................. 13
Biodegradation, bioremediation and bioaugmentation ..................................................................... 15 Challenges for bioaugmentation ............................................................................................................................. 16
Biodegradation by fungi ................................................................................................................... 17 Filamentous fungi .................................................................................................................................................... 17 Mortierella .............................................................................................................................................................. 19 Fungal degradation of pesticides ............................................................................................................................ 20
Consortia for bioremediation ........................................................................................................... 24 Fungal-bacterial interactions .................................................................................................................................. 24 Fungal highway ....................................................................................................................................................... 25 Degradation by consortia ........................................................................................................................................ 26
Conclusions and perspectives ................................................................................................... 29
Reference list ........................................................................................................................... 31
Manuscript I ............................................................................................................................. 41
Manuscript II ............................................................................................................................ 61
Manuscript III ........................................................................................................................... 79
Appendix ................................................................................................................................ 104


3
List of manuscripts
List of enclosed article manuscripts:
Manuscript I
Ellegaard-Jensen, L., Aamand, J., Kragelund, B.B., Johnsen, A.H., and Rosendahl, S.
Degradation of the phenylurea herbicide diuron by soil fungi: different degradation potentials
of five Mortierella strains.
In revision for Biodegradation
Manuscript II
Knudsen, B.E., Ellegaard-Jensen, L., Albers, C.N., Rosendahl, S., and Aamand, J.
Transport of degrader bacteria via fungal hyphae increases degradation of 2,6-
dichlorobenzamide.
Submitted to Environmental Pollution
Manuscript III
Ellegaard-Jensen, L., Knudsen, B.E., Johansen, A., Albers, C.N., Aamand, J., and Rosendahl, S.
Fungal-bacterial consortium overcomes dry patches and increases diuron degradation.
In prep. for submission to Science of the Total Environment
Article manuscript not included in this thesis:
Knudsen, B.E., Ellegaard-Jensen, L., Albers, C.N., Harder, C.B., Ekelund, F., Aamand, J., and
Rosendahl, S. Characterization of the biota in sandfilter material from 11 Danish water works.
In prep. for submission to Water Research

4
Conference presentations:
Ellegaard-Jensen, L., Aamand, J., & Rosendahl, S. Fungal degradation of phenylurea herbicide
diuron. Presented at Symposium of the Danish Microbiological Society, Copenhagen 7.11.2011.
Poster Presentation
Knudsen, B.E., Ellegaard-Jensen, L., Albers, C., Rosendahl, S., & Aamand, J. Transport of BAM
degrading bacteria facilitated by fungal hyphae increases BAM mineralization in sand. Presented at
Symposium of the Danish Microbiological Society, Copenhagen 7.11.2011. Poster Presentation
Ellegaard-Jensen, L., Aamand, J., Kragelund, B.B., Johnsen, A.H., & Rosendahl, S. Degradation of
the herbicide diuron by Mortierella. Presented at The International Symposium on Microbial
Ecology – ISME14, Copenhagen 19.- 24.08.2012. Poster Presentation
Knudsen, B.E., Ellegaard-Jensen, L., Albers, C., Rosendahl, S., & Aamand, J. Transport of BAM
degrading bacteria facilitated by fungal hyphae increases BAM mineralization in sand. Presented at
The International Symposium on Microbial Ecology – ISME14, Copenhagen 19.- 24.08.2012. Poster
Presentation
Ellegaard-Jensen, L., Knudsen, B.E., Albers, C.N., Aamand, J., & Rosendahl, S. Three-member
consortium overcomes dry patches and increases diuron degradation. Presented at Copenhagen
Microbiology Center – CMC Symposium, Copenhagen 5.10.2012. Poster and Flash Presentation
Ellegaard-Jensen, L., Knudsen, B.E., Johansen, A., Albers, C.N., Aamand, J., & Rosendahl, S.
Three-member consortium overcomes dry patches and increases diuron degradation. Presented at
Symposium of the Danish Microbiological Society, Copenhagen 5.11.2012. Poster Presentation

5
English summary
Pesticides are used worldwide on agricultural land as well as in urban areas. This use has often led
to contamination of the environment with serious effects on our natural resources. Frequent
pesticide use and spills have led to deterioration of soil quality and pesticide leaching has resulted in
groundwater contamination. New technologies are therefore needed for cleaning up contaminated
soil and water resources. This PhD was part of the project entitled Microbial Remediation of
Contaminated Soil and Water Resources (MIRESOWA) where the overall aim is to develop
new technologies for bioremediation of pesticide contaminated soil and water. The objectives of
this PhD were to investigate fungal degradation of pesticides and following to construct microbial
consortia for bioremediation.
In Manuscript I the fungal degradation of the phenylurea herbicide diuron was studied.
Isolates of soil fungi of the genus Mortierella were tested for their ability to degrade diuron. The
fungi were incubated in liquid culture with diuron on an orbital shaker at 8
oC. The results showed
that three of the five strains tested could degrade diuron, and molecular analysis revealed that these
three strains constituted a closely related phylogenetic group, while the two non-degraders were
located more distantly on the phylogenetic tree. In addition, it was examined whether the fungi
utilized diuron as a carbon or nitrogen source. Here the most efficient diuron degrading strain
Mortierella sp. LEJ701 was applied in liquid cultures with different levels of carbon and nitrogen.
Degradation of diuron was fastest in carbon and nitrogen rich media while degradation was very
restricted at low nutrient levels, making it unlikely that Mortierella utilize diuron as carbon or
nitrogen source. The degradation kinetics of these experiments showed that diuron degradation was
followed by formation of the metabolites 1-(3,4-dichlorophenyl)-3-methylurea, 1-(3,4-
dichlorophenyl)urea and an hitherto unknown metabolite. This metabolite was subsequently
biosynthesised, purified and analyzed by nuclear magnetic resonance (NMR) and mass
spectroscopy (MS). On the basis of these analyses the metabolite was suggested to be 1-(3,4-
dichlorophenyl)-3-methylideneurea. This study especially brings new insights into the phylogenetic
link between fungal diuron degraders, but also to the fungal degradation pathway of diuron.

6
The next two manuscripts dealt with constructing fungal-bacterial consortia and examining
whether their degradation would be superior to that of the single strains in unsaturated systems. In
Manuscript II a consortium was created for degradation of the pesticide metabolite 2,6-
dichlorobenzamide (BAM). A consortium with Mortierella sp. LEJ702 and the BAM-degrading
Aminobacter sp. MSH1 as well as the single strains were introduced into small sand columns. The
sand was spiked with [ring-U-14
C]-BAM and mineralization could thus be determined from the
amount of evolved 14
CO2. In addition, the effect of moisture content was examined by adding water
corresponding to 0, 1.7, 5 or 10% of water holding capacity (WHC) to the sand. A faster
mineralization of BAM was obtained by the consortium compared to Aminobacter sp. MSH1 alone,
especially at the lower moisture contents. These results were supported by chemical analyses of 14
C
residues extracted from the sand. Additionally, it was investigated whether bacterial transport was
enhanced in the presence of Mortierella. This was done by extracting DNA from the top layer of the
sand followed by quantitative real-time polymerase chain reaction (qPCR) analysis. Results
demonstrated that the number of Aminobacter transported to the top sand was greatly enhanced in
the presence of Mortierella, suggesting that the fungal hyphae act as transport vectors for the
bacteria in the sand. Finally, the distribution of 14
C-BAM in the sand was studied in novel setup. It
was found that the presence of Mortierella slightly enhanced BAM distribution. From this work it is
evident that the fungal-bacterial consortium is capable of enhancing BAM-degradation in
unsaturated systems, and may therefore be a promising application for soil bioremediation.
In Manuscript III two- and three-member consortia were constructed with bacterial and
fungal diuron degraders. The purpose of this was to create a consortium which was superior for
diuron degradation in unsaturated system and to investigate the interactions between the
microorganisms in this consortium. The synergy leading to a more efficient degradation could either
be a result of co-operative catabolism or physical interactions between the organisms improving
growth and/or distribution of fungi and bacteria. The bacterial strains applied were Sphingomonas
sp. SRS2, Variovorax sp. SRS16 and Arthrobacter globiformis D47 and the fungal strains were
Mortierella sp. LEJ702 and Mortierella sp. LEJ703. In the experimental setup a layer of sterile
glass beads was added between the organisms and the sand column above; simulating air-filled gaps
in soil. [Ring-U-14
C]-diuron was mixed into the sand to a concentration of 100 μg diuron kg-1
.
Degradation was measured as the amount of 14
C-diuron mineralized and as 14
C residues in the sand
at experimental termination. Mineralization results established the three-member consortium

7
LEJ702/SRS16/D47 as the most efficient transforming 32% of the diuron to 14
CO2, while the single
strains or other consortia mineralized no more than 10%. Furthermore, analyses of 14
C residues in
the sand showed that production of diuron metabolites by this consortium was minimal. The
interactions between the organisms in the consortia were examined by phospholipid fatty acid
analysis (PLFA) and 16S rDNA PCR using strain-specific primers. The molecular results suggested
that the presence of Mortierella sp. LEJ702 enhanced distribution of SRS16 and D47 in the sand.
Only fungal phospholipid fatty acids could be quantified. From those, however, it was apparent that
the fungal growth was severely inhibited in the presence of A. globiformis D47. On the other hand,
this effect was somewhat alleviated in the three-member consortium LEJ702/SRS16/D47. This
study is the first to show that a three-member consortium of both fungal and bacterial degraders can
indeed increase pesticide degradation.

8
Dansk resumé
Pesticider anvendes over hele verden på landbrugsjorde såvel som i byområder. Denne anvendelse
har ofte medført forurening af miljøet med alvorlige følgevirkninger for vores naturressourcer.
Hyppig pesticid brug og utilsigtede udslip har ført til forringelse af jordkvalitet og udvaskning af
pesticider har resulteret i grundvandsforurening. Der er derfor brug for nye teknologier til rensning
af forurenet jord og vand. Denne PhD er udført som en del af forskningsprojektet Microbial
Remediation of Contaminated Soil and Water Resources (MIRESOWA), som har det
overordnede mål at udvikle nye teknologier til bioremediering af pesticidforurenet jord og vand.
Formålet med denne PhD var at undersøge svampes nedbrydning af pesticider og efterfølgende at
konstruere mikrobielle konsortier til bioremediering.
I Manuskript I blev svampes nedbrydning af phenylurea herbicidet diuron undersøgt.
Isolater af jordsvampe af Mortierella slægten blev testet for deres evne til at nedbryde diuron.
Svampene blev inkuberet i flydende kulturer med diuron på rystebord ved 8oC. Resultaterne viste at
tre ud af de fem testede stammer kunne nedbryde diuron, og molekulære analyser viste at disse tre
stammer fylogenetisk set udgjorde en nærtbeslægtet gruppe, mens de to ikke-nedbrydere var
placeret længere væk på det fylogenetiske træ. Derudover blev det undersøgt om svampene brugte
diuron som en kulstof eller kvælstof kilde. Her blev den mest effektive diuron-nedbryder
Mortierella sp. LEJ701 inkuberet i flydende medier med forskellige kulstof og kvælstof niveauer.
Diuron nedbrydning var hurtigst i medier med høje kulstof og kvælstof niveauer, mens
nedbrydningen var meget begrænset ved de lave næringsstofniveauer. Dette viser at Mortierella
sandsynligvis ikke bruger diuron som kulstof eller kvælstof kilde. Nedbrydningskinetikken set i
disse eksperimenter viste at diuron blev nedbrudt til metaboliterne 1-(3,4-dichlorophenyl)-3-
methylurea, 1-(3,4-dichlorophenyl)urea samt til et hidtil ukendt nedbrydningsprodukt. Dette
nedbrydningsprodukt blev efterfølgende biosyntesiseret, oprenset og analyseret ved kernemagnetisk
resonans (NMR) og massespektroskopi (MS). Ud fra disse analyser blev metaboliten med stor
sandsynlighed identificeret til at være 1-(3,4-dichlorophenyl)-3-methylideneurea. Dette studie har
bidraget med ny indsigt i fylogenetiske relationer mellem diuron-nedbrydende svampe, samt ny
viden om svampes nedbrydningsvej for diuron.

9
De to efterfølgende manuskripter beskæftigede sig med skabelsen af svampe-bakterie
konsortier og undersøge om disses nedbrydning ville være mere effektiv end renkulturernes
nedbrydning under umættede forhold. I Manuskript II blev et konsortium skabt med det formål at
nedbryde pesticidmetaboliten 2,6-dichlorobenzamide (BAM). Et konsortium bestående af
Mortierella sp. LEJ702 og den BAM-nedbrydende bakterie Aminobacter sp. MSH1, samt de
enkelte renkulturer, blev tilført små sandsøjler. Da sandet var blevet tilsat [ring-U-14
C]-BAM kunne
mineraliseringen således bestemmes ud fra mængden af dannet 14
CO2. Derudover blev effekten af
vandindholdet undersøgt ved at tilføre vandmængder svarende til 0, 1.7, 5 eller 10 % af den
vandholdende evne (WHC) til sandet. Konsortiet opnåede en hurtigere mineralisering af BAM
sammenlignet med Aminobacter sp. MSH1 alene, især ved de lavere vandindhold. Disse resultater
blev støttet af de kemiske analyser af 14
C forbindelser i sandet. Det blev yderligere undersøgt om
transporten af bakterier blev forøget når Mortierella var til stede. Dette blev gjort ved ekstrahering
af DNA fra det øverste sandlag efterfulgt af kvantitativ real-time polymerase chain reaction (qPCR)
analyse. Resultaterne viste at antallet af Aminobacter celler i det øverste sandlag var væsentligt
forøget ved tilstedeværelsen af Mortierella. Dette tyder på at svampehyferne fungerer som fysisk
transportvej for bakterierne i sandet. Herudover blev fordelingen af 14
C-BAM i sandet undersøgt i
en ny opstilling. Undersøgelsen viste at tilstedeværelsen af Mortierella forårsagede en svag
forøgelse af BAM spredningen. Dette studie viser tydeligt at det undesøgte svampe-bakterie
konsortium er i stand til at forøge nedbrydningen af BAM under umættede forhold, hvilket gør brug
af dette lovende i forbindelse med bioremediering af jord.
I Manuskript III blev konsortier med to eller tre organismer sammensat af bakterie og
svampe diuron-nedbrydere. Formålet med dette var at danne et konsortium, der var mere effektivt
med hensyn til diuron nedbrydning under umættede forhold og undersøge interaktionerne mellem
mikroorganismerne i dette konsortium. Synergien som fører til en mere effektiv nedbrydning kan
enten skyldes successiv nedbrydning eller fysiske interaktioner mellem organismerne, der forøger
vækst og/eller udbredelse af svampe og bakterier. De anvendte bakteriestammer var Sphingomonas
sp. SRS2, Variovorax sp. SRS16 og Arthrobacter globiformis D47 og svampe-stammerne var
Mortierella sp. LEJ702 og Mortierella sp. LEJ703. I det eksperimentelle set-up blev et lag af sterile
glasperler tilføjet mellem organismerne og den overliggende sandsøjle, for at simulere luftlommer i
jord. [Ring-U-14
C]-diuron blev blandet i sandet givende en koncentration på 100 µg kg-1
.
Nedbrydningen blev målt som mængden af mineraliseret 14
C-diuron og som koncentrationen af 14
C-

10
forbindelser i sandet ved eksperimentets afslutning. Mineraliseringsresultaterne påviste tre-
organisme konsortiet LEJ702/SRS16/D47 som det mest effektive, da det omdannede 32% af det
tilsatte diuron til 14
CO2, mens renkulturerne og de andre konsortier alle mineraliserede mindre end
10%. Ydermere viste analyserne af 14
C-forbindelser i sandet at dette konsortium kun havde
produceret minimale mængder af diuron-metaboliter. Interaktionerne mellem organismerne i
konsortierne blev undersøgt ved fosfolipid fedtsyre analyse (PLFA) og 16S rDNA PCR med
specifikke primere. De molekylære resultater indikerede at spredningen af bakterierne SRS16 og
D47 i sandet blev øget ved tilstedeværelsen af Mortierella sp. LEJ702. Kun svampenes fosfolipid
fedtsyrer kunne kvantificeres. Ud fra disse var det imidlertid tydeligt at svampenes vækst blev
kraftigt hæmmet ved tilstedeværelsen af A. globiformis D47. Denne effekt var dog mindsket i tre-
organisme konsortiet LEJ702/SRS16/D47. Dette studie er det første, der viser at et konsortium med
tre organismer af både svampe og bakterie nedbrydere kan forøge pesticidnedbrydning.

11
Introduction and aims
This PhD thesis consists of two submitted manuscripts and one manuscript in preparation.
Furthermore, it includes a background discussion of the most relevant literature regarding fungal
degradation of pesticides and what beneficial interactions may be achieved by employing fungal-
bacterial consortia for bioremediation. In these discussions the new findings of this PhD will be put
into context. Finally, there is a conclusion on the work, and perspectives are brought forth on future
research needs in the present area.
This PhD was part of the project entitled Microbial Remediation of Contaminated Soil and
Water Resources (MIRESOWA). In this setting it was predetermined that at least 1/3 of the work
should focus on constructing fungal-bacterial consortia for bioremediation of pesticides. The other
part of the work was focused on fungal degradation of pesticides. As it turned out these two parts of
the work became very well integrated into each other, and as such they both contributed to a deeper
understanding of the other. Another aspect of the work on consortia was that this was carried out in
close collaboration with a PhD student employed at the Geological Survey of Denmark and
Greenland (GEUS). The experimental work was therefore done both at the University of
Copenhagen and at GEUS.
The first aim of this PhD was to study the fungal degradation of pesticides with special focus
on the biology of the pesticide degrading strains found. For the biology behind fungal degradation
of pesticides the following scenarios could be envisioned: (i) utilization of the pesticide as a carbon
or nitrogen source either by uptake or extracellular breakdown of the compound, (ii) that it is a co-
metabolic process occurring by enzymes excreted by the fungus, where the pesticide does not serve
as a carbon and energy source for the fungus, or (iii) that it is a detoxification mechanism for the
fungus when exposed to toxic compounds. Manuscript I uses the phenylurea herbicide diuron as a
model compound to investigate these issues as well as the phylogenetic relationship between
pesticide degrading strains.

12
The second aim was to construct fungal-bacterial consortia for bioremediation of pesticides.
The hypothesis being that these would be more efficient than single strains for biodegradation in
water unsaturated heterogeneous environments. The interactions investigated in Manuscript II and
III were beneficial/antagonistic effects on growth and transport of bacteria via fungal hyphae. In
addition, the effects of these interactions on mineralization and metabolite development were
examined. In Manuscript II the pesticide metabolite 2,6-dichlorobenzamide (BAM) served as
model compound to study the effects of a non-degrading fungal strain on the transport and
performance of a BAM-degrading bacterial strain. Additionally, the influence of moisture content
on BAM mineralization by this consortium was examined. In Manuscript III the herbicide diuron
was used and two- and three-member consortia where both fungal and bacterial strains could
degrade diuron were constructed and studied. Giving in this regard a unique synergistic degradation
situation to investigate.

13
Background
Pesticide pollution – why should we care
Pesticides are chemical compounds used on agricultural land, but also in private gardens, along
railways and on other public areas. They are targeted to be toxic to certain groups of organisms; e.g.
insecticides target insects to protect crops from being eaten or infected, fungicides target fungi to
protect the crop from fungal attack, and herbicides target specific plants to protect the crops from
weed growth. Pesticide use worldwide is extensive, with the herbicides accounting for the largest
part of overall pesticide use (Grube et al., 2011).
Aside from being toxic to target organisms, pesticides often have side effects on other
organisms. Be it in the same ecosystem, but on non-target organisms, or in other adjacent
ecosystems such as streams and rivers, affecting the organisms there (Ayansina, 2008). Finally
some pesticides may reach the level of human consumption through the contamination of
groundwater from leaching of pesticides. The issue of pesticides in the groundwater is therefore
very serious. In Denmark groundwater is traditionally used for drinking water without any prior
treatment e.g. for pesticide contamination. Consequently, a number of water wells is being closed
annually, because of contamination by pesticides and/or their metabolites in concentrations above
the EU limit value (Brüsch, 2010). The EU maximum permissible value of a single pesticide in
ground- and drinking-water is 0.1 µg l-1
, whereas the sum of several pesticides in the water must not
exceed 0.5 µg l-1
(European Council, 1980). These values are not based upon direct assessments of
health or environmental hazards, but on the precautionary principle since the effects of many
metabolites and effects of mixtures of pesticides are unknown (Brüsch, 2010).
Diuron, dichlobenil and BAM
It is not unusual that pesticides are formulated and thus added as mixtures of two active compounds.
For instance, the two herbicides diuron and dichlobenil may be applied alone or in mixtures with
each other or alternative compounds (Tomlin, 1997). Diuron belongs to the phenylurea herbicides
while dichlobenil is a benzonitrile herbicide, and the properties of these two compounds are rather
different (see Appendix). Although both compounds have been banned from use as pesticides in
Denmark, they still contribute to the contamination of the environment in Denmark as well as in
other countries where they are still in use.

14
Diuron is on the EU Water Framework Directive´s list of priority substances (European
Parliament, 2008). One of the reasons being that it has serious effects on the environment and also
on human health as it is suspected of being carcinogenic (Cox, 2003; Giacomazzi and Cochet,
2004). In addition, diuron and its metabolites have been found in surface and groundwater in
concentrations above the EU limit value (Torstensson, 2001; Lapworth and Gooddy, 2006; Struger
et al., 2011).
For dichlobenil the largest problem is that it is readily degraded to the metabolite 2,6-
dichlorobenzamide (BAM), which is highly mobile and therefore leaches to the groundwater
(Clausen et al., 2004). BAM has frequently been detected in surface as well as groundwater in
concentrations above 0.1 µg l-1
(Porazzi et al., 2005; Eriksson et al., 2007; Brüsch, 2010), which has
led to the closure of several hundred water extraction wells in Denmark (Brüsch, 2010).
Though the compounds described above are moderately (diuron) to very (BAM) persistent in
the environment, microorganisms have been isolated that can degrade them. Simonsen et al. (2006)
and Sørensen et al. (2007) have isolated BAM mineralizing Aminobacter strains ASI1 and MSH1.
Holtze et al. (2007) proposed a degradation pathway for dichlobenil and BAM leading to
mineralization through the formation of new metabolites (see Appendix). Degradation of diuron
also occur primarily as a microbial process, and several organisms have been isolated that can
degrade diuron e.g. the bacterial strains Sphingomonas sp. SRS2 (Sørensen et al., 2001), Variovorax
sp. SRS16 (Sørensen et al., 2008), Arthrobacter globiformis D47 (Turnbull et al., 2001) and also a
number of fungal strains (see Table 1).

15
Biodegradation, bioremediation and bioaugmentation
Contamination of the environment with chemicals is a major issue in our time. Chemicals used
domestically, industrially or in agriculture may lead to environmental problems when deteriorating
the quality of soil and groundwater. In these cases bioremediation can be a possible solution for
remediating pollution in the environment. Bioremediation is simply the use of biological organisms
for cleaning up chemical pollution by reducing the concentrations and/or the toxicity of chemical
compounds and restoring natural conditions (Ahemad et al., 2008). The aim is to clean the
environment to protect both biodiversity (e.g. soil habitats and microbial composition) and
resources for human use and consumption (e.g. soil fertility and groundwater quality). As chemicals
are released into the environment in large amounts, both intentionally as is the case with spraying of
pesticides and accidentally through spills, leading to decrease in environmental quality the needs for
bioremediation is growing.
Numerous studies have focused on the use of either plants (see reviews by Alkorta and
Garbisu, 2001; Gerhardt et al., 2009) or microbes (see reviews by Sørensen et al., 2003; Singh,
2006; Ahemad et al., 2008; Das and Adholeya, 2012) for bioremediation of pollutants.
Nevertheless, bioremediation studies have not always yielded successful results showing that
thought bioremediation is a very promising biotechnology it also has its limitations (Juwarkar et al.,
2010). First, some chemicals are not readily susceptible to biological degradation due to their
chemical properties or strong sorption to the environmental matrix. Secondly, microbial degradation
of some compounds may yield metabolites which are more toxic than the starting compound.
Thirdly, many scientific resources have to go into discovering the best microorganism(s) for the job
(Boopathy, 2000). However, with a multidisciplinary approach, involving expertise from
microbiology, chemistry and geology to name a few, it should be possible to achieve satisfactory
bioremediation.
It goes without saying that bioremediation is more environmental friendly than conventional
remediation techniques. Bioremediation is considered a ‗green technology‘ as it only depends on
biological organisms and processes, and it does not require any chemical addition or heating
treatment (Juwarkar et al., 2010). It also presents several advantages over conventional remediation
techniques such as land filling and incineration, e.g. it can be done on site and is therefore less

16
disruptive, it is often less expensive, and it eliminates the pollutant permanently restoring the
vitality of the environment (Boopathy, 2000). Reviews by Juwarkar et al. (2010) and Leung et al.
(2007) describe applied bioremediation technologies e.g. bioaugmentation, biofilters, biostimulation
and establish the many factors that can affect the outcome of bioremediation e.g. temperature,
oxygen level, moisture and nutrients.
Challenges for bioaugmentation
Biodegradation can proceed as a process where the naturally occurring microorganisms degrade
pollutants, however the processes may be enhanced by special conditions in terms of adding
nutrients or electron acceptors (biostimulation) or by addition of specific degrader organisms
(bioaugmentation) (El Fantroussi and Agathos, 2005). The present work focuses on investigating
strains for bioaugmentation, as it is considered a microbial technology with a large potential for
bioremediation, but with great challenges in many regards (Thompson et al., 2005). These
challenges are in the physical and chemical sense the limited bioaccessibility of the compound due
to sorption or physical heterogeneity in the environment. In addition to this, biological challenges
for the introduced organisms are present, such as predation and competition from the naturally
occurring microorganisms. So the success of bioaugmentation not only depends on the ability of the
organism to degrade the pollutant, but very much also on the survival, growth and distribution of
the organism(s). Fungi alone or in combination with bacteria may be better suited to overcome these
challenges, and this subject will be the topic of the following sections.

17
Biodegradation by fungi
Fungal diversity is globally estimated to 1.5 million species (Hawksworth, 1991, 2001) and consist
of an incredibly diverse group of organisms. Organisms studied by mycologists include members of
the fungal Kingdom but also others like Protozoa e.g. slime moulds (Carlile et al., 2001). The
Kingdom of Fungi is divided into three major classes: the zygomycetes, the ascomycetes and the
basidiomycetes. For a thorough review on the fungal diversity see Carlile et al. (2001).
Biodegradation by fungi is also known as mycodegradation. Likewise, bioremediation in
which fungi are employed is sometimes called mycoremediation (Singh, 2006). Fungi are
ubiquitous in the environment, and the literature on fungal ecology is vast. Despite this, the reports
of fungi in bioremediation are under-represented and as such represent the untapped potentials in
fungal bioremediation (Harms et al., 2011). Moreover, the biology and ecology of mycoremediation
have rarely been examined (Singh, 2006).
The overall aim of this chapter is to examine the manner and function of fungal pesticide
degradation. First, biological characteristics of the fungi which are relevant for successful
bioremediation will be highlighted. Then, characteristics of the fungal genus Mortierella, which
was used in all investigations of this thesis, are presented. Lastly, the most relevant studies on
fungal degradation of pesticides will be reviewed.
Filamentous fungi
Studies into the bioremediation potential of microorganisms have for the largest part focused on
bacterial degraders. However, filamentous fungi possess characteristics which are advantageous in
heterogeneous environments. Examples will be given below of fungal traits that in some instances
make them more successful than bacteria.
Although fungi are non-motile they can respond quickly to changing environmental
conditions to survive or escape them (Read, 2007). Abiotic factors that primarily affect the mycelial
growth include temperature, water potential, pH, oxygen accessibility, and nutrient status (Boddy
and Jones, 2007). However, filamentous fungi can escape unfavorable conditions which in
heterogeneous environments give them an advantage. For instance, in soil fungi do not require a
continuous water phase as they can bridge air filled gaps (Wösten et al., 1999; Ritz and Young,
2004), whereas bacteria in general are immobilized and their motility impeded at matric potentials

18
below -2 to -5 kPa (Wong and Griffin, 1976; Dechesne et al., 2010; Wang and Or, 2010). This
might be of importance in coarse soils for instance, where matric potentials may be relatively low,
approximately -5 kPa, causing discontinuities to occur in the water-filled pathways in the soil
(Wong and Griffin, 1976). Fungal hyphae are also able to penetrate solids and reach microhabitats
like water-filled micropores in soil (Gadd, 2007; Thorn and Lynch, 2007). Fungi may in this way
gain a much better access to the nutrients and contaminants in environments where the compounds
are heterogeneously distributed and inaccessible to bacteria in other ways e.g. trapped in micropores
(Harms et al., 2011). In addition, fungal hyphae transport nutrients between the different regions of
the mycelia, which enables fungal growth into otherwise inhospitable zones (Lindahl and Olsson,
2004).
Also, toxic conditions can be overcome by production of long sparsely-branched or
branchless explorative hyphae (Fomina et al., 2003). A growth response which enables expansive
search for less toxic sites with minimal resources spent by the fungus. We also observed this type of
growth when Mortierella sp. LEJ701 was exposed to diuron (Fig. 1D), suggesting that either diuron
or its metabolites were toxic to the fungus (Manuscript I). This type of growth was not observed
for the fungus in control treatments without diuron.
Saprotrophic fungi produce a wide range of extracellular enzymes which are essential for
degradation of plant materials (Kjøller and Struwe, 2002), and these enzymes may also enable
fungal degradation of organic pollutants. White-rot fungi have been considered as top fungal
candidates for bioremediation purposes for many years due to their potent enzymatic arsenal
(Pointing, 2001; Reddy and Mathew, 2001). However there are a number of drawbacks for this
group of organisms, for instance most white rot fungi require strict growth conditions (e.g. high
temperatures) and therefore have low competitive capabilities against indigenous organisms in the
environment (Baldrian, 2008; Gao et al., 2010). Furthermore, it has been shown that a number of
white-rot fungi have negative effects on soil bacteria e.g. inhibiting growth of indigenous bacteria
(Lang et al., 1997; Andersson et al., 2003). For bioremediation of certain compounds fungi from the
zygo- or ascomycetes might therefore be a superior choice. However, the effect on native bacteria
will probably be strain dependent.
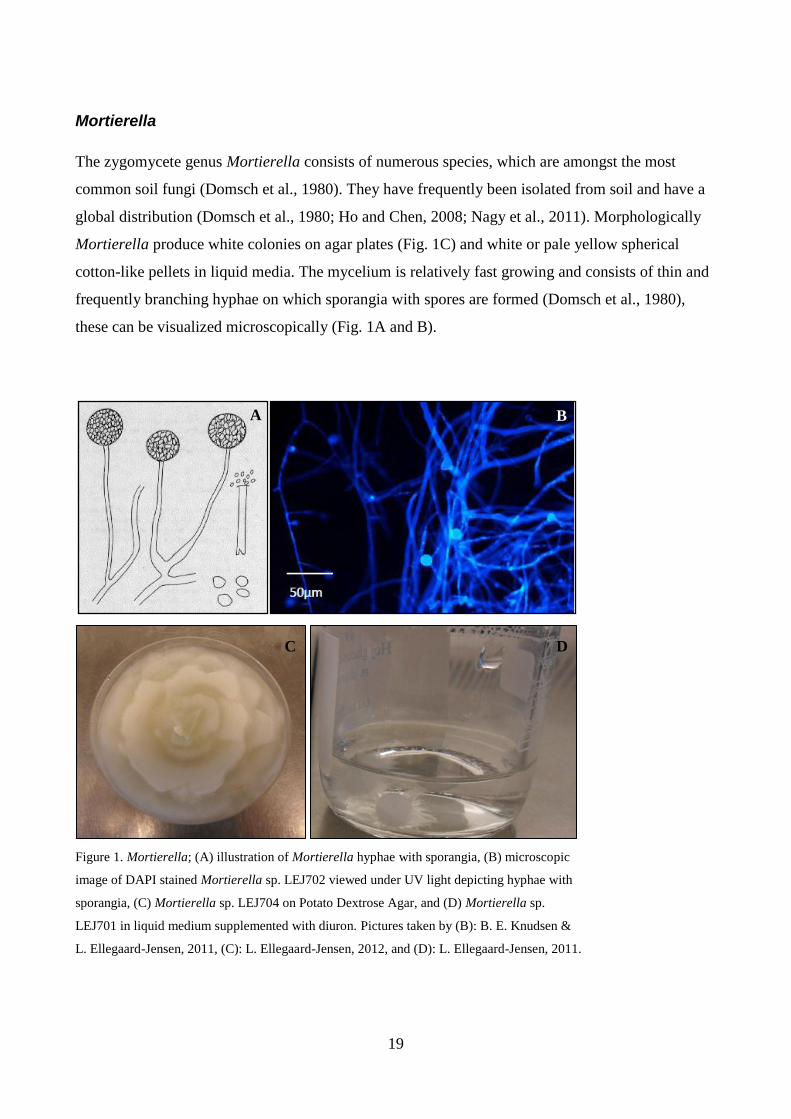
19
Mortierella
The zygomycete genus Mortierella consists of numerous species, which are amongst the most
common soil fungi (Domsch et al., 1980). They have frequently been isolated from soil and have a
global distribution (Domsch et al., 1980; Ho and Chen, 2008; Nagy et al., 2011). Morphologically
Mortierella produce white colonies on agar plates (Fig. 1C) and white or pale yellow spherical
cotton-like pellets in liquid media. The mycelium is relatively fast growing and consists of thin and
frequently branching hyphae on which sporangia with spores are formed (Domsch et al., 1980),
these can be visualized microscopically (Fig. 1A and B).
Figure 1. Mortierella; (A) illustration of Mortierella hyphae with sporangia, (B) microscopic
image of DAPI stained Mortierella sp. LEJ702 viewed under UV light depicting hyphae with
sporangia, (C) Mortierella sp. LEJ704 on Potato Dextrose Agar, and (D) Mortierella sp.
LEJ701 in liquid medium supplemented with diuron. Pictures taken by (B): B. E. Knudsen &
L. Ellegaard-Jensen, 2011, (C): L. Ellegaard-Jensen, 2012, and (D): L. Ellegaard-Jensen, 2011.
A B
C D

20
Mortierella are saprotrophic fungi degrading organic materials and utilizing sugars as well as
pectin, cellulose and chitin (Kjøller and Struwe, 1990). This distinguishes Mortierella from other
genera within the Mucorales, which are unable to degrade polysaccharides (Kjøller and Struwe,
2002). In addition, strains of Mortierella have been shown capable of degrading xenobiotic
compounds e.g. polycyclic aromatic hydrocarbons (PAHs) (Salicis et al., 1999) and pesticides
(Tixier et al., 2000; Rønhede et al., 2005; Vroumsia et al., 2005; Badawi et al., 2009; Kataoka et al.,
2011). Because of the abovementioned characteristics with regard to degradation potentials this
PhD focuses on studying the degradation of pesticides by Mortierella to gain a higher
understanding of the biological processes of fungal degradation of these compounds.
Fungal degradation of pesticides
As mentioned, biodegradation by fungi is not widely studied. However, a number of reports on
fungal degradation of pesticides are found in the literature. As it is the fungal degradation potential
of the phenylurea herbicide diuron that is studied in Manuscript I (and III) the literature on this
will be subjected to a short review. This is done with the aim of relating the findings in Manuscript
I to those of previous studies on fungal pesticide degradation, and further seeking to elucidate the
role that the fungal biology plays in diuron degradation.
The literature on fungal degradation of diuron is somewhat inconsistent with regard to what
parameters and which fungal species were studied (see Table 1). Khadrani et al. (1999) and
Vroumsia et al. (1996) both screened a large number of fungal species for their ability to degrade
phenylurea herbicides including diuron. Rhizoctonia solani (Vroumsia et al., 1996) and Bjerkandera
adusta (Khadrani et al., 1999), both species belonging to the basidiomycetes, were identified as the
most efficient degraders of diuron. However, care should be taken when choosing strains for
bioaugumentation and a plant pathogen as Rhizoctonia solani might therefore not be a sound
choice. Instead, others have reported species belonging to the zygomycetes as very efficient
degraders of phenylurea herbicides (e.g. Manuscript I; Tixier et al., 2000; Rønhede et al., 2005;
Hangler et al., 2007).
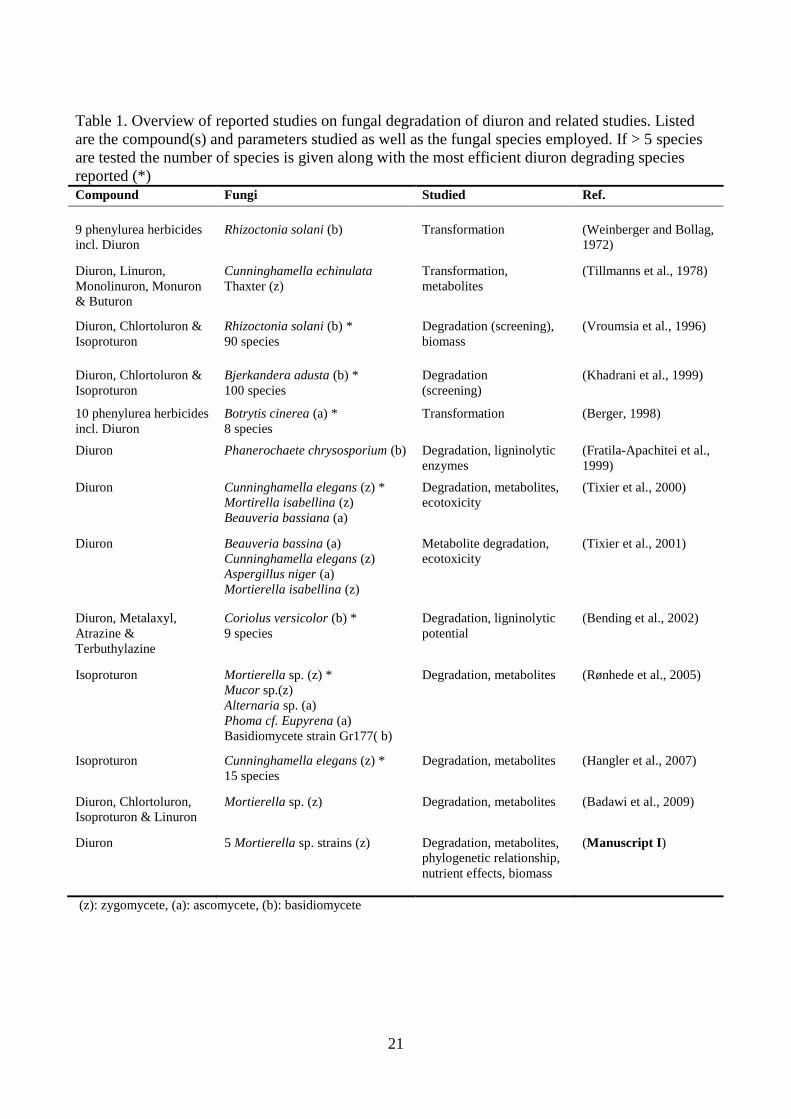
21
Table 1. Overview of reported studies on fungal degradation of diuron and related studies. Listed
are the compound(s) and parameters studied as well as the fungal species employed. If > 5 species
are tested the number of species is given along with the most efficient diuron degrading species
reported (*)
Compound Fungi Studied Ref.
9 phenylurea herbicides
incl. Diuron
Rhizoctonia solani (b)
Transformation
(Weinberger and Bollag,
1972)
Diuron, Linuron,
Monolinuron, Monuron
& Buturon
Cunninghamella echinulata
Thaxter (z)
Transformation,
metabolites
(Tillmanns et al., 1978)
Diuron, Chlortoluron &
Isoproturon
Rhizoctonia solani (b) *
90 species
Degradation (screening),
biomass
(Vroumsia et al., 1996)
Diuron, Chlortoluron &
Isoproturon
Bjerkandera adusta (b) *
100 species
Degradation
(screening)
(Khadrani et al., 1999)
10 phenylurea herbicides
incl. Diuron
Botrytis cinerea (a) *
8 species
Transformation (Berger, 1998)
Diuron Phanerochaete chrysosporium (b) Degradation, ligninolytic
enzymes
(Fratila-Apachitei et al.,
1999)
Diuron Cunninghamella elegans (z) *
Mortirella isabellina (z)
Beauveria bassiana (a)
Degradation, metabolites,
ecotoxicity
(Tixier et al., 2000)
Diuron Beauveria bassina (a)
Cunninghamella elegans (z)
Aspergillus niger (a)
Mortierella isabellina (z)
Metabolite degradation,
ecotoxicity
(Tixier et al., 2001)
Diuron, Metalaxyl,
Atrazine &
Terbuthylazine
Coriolus versicolor (b) *
9 species
Degradation, ligninolytic
potential
(Bending et al., 2002)
Isoproturon Mortierella sp. (z) *
Mucor sp.(z)
Alternaria sp. (a)
Phoma cf. Eupyrena (a)
Basidiomycete strain Gr177( b)
Degradation, metabolites (Rønhede et al., 2005)
Isoproturon Cunninghamella elegans (z) *
15 species
Degradation, metabolites (Hangler et al., 2007)
Diuron, Chlortoluron,
Isoproturon & Linuron
Mortierella sp. (z) Degradation, metabolites (Badawi et al., 2009)
Diuron 5 Mortierella sp. strains (z) Degradation, metabolites,
phylogenetic relationship,
nutrient effects, biomass
(Manuscript I)
(z): zygomycete, (a): ascomycete, (b): basidiomycete

22
Other authors have focused on the degradation kinetics of phenylurea herbicides (Tillmanns et
al., 1978; Badawi et al., 2009). A fungal degradation pathway has been established where diuron is
transformed by demethylation to 1-(3,4-dichlorophenyl)-3-methylurea (DCPMU) and then further
to 1-(3,4-dichlorophenyl)-urea (DCPU) (Tixier et al., 2000; Badawi et al., 2009). In addition, a
previously unidentified metabolite was reported in Badawi et al. (2009) and Manuscript I. By use
of NMR and mass spectrometry, the compound was suggested to be 1-(3,4-dichlorophenyl)-3-
methylideneurea (Manuscript I). Transformation beyond DCPU appears to occur mainly by
bacterial degradation. Where certain bacterial strains can transform diuron to 3,4-dichloroaniline
(3,4-DCA) (Turnbull et al., 2001) or mineralize it (Sørensen et al., 2008). Tixier et al. (2000; 2001;
2002) found that DCPMU, DCPU and 3,4-DCA were more toxic than diuron when tested in
Microtox assays, with the most toxic being 3,4-DCA. However, it has been shown that certain fungi
can transform 3,4-DCA to the less toxic 3,4-dichloroacetanilide (Tixier et al., 2002; Martins et al.,
2009; Martins et al., 2010; Silar et al., 2011; Manuscript III). Whether diuron or its metabolites are
toxic to fungi remains unclear, thought findings indicate that it could be the case (Manuscript I;
Vroumsia et al., 1996).
So it is clear from the above and from Table 1 that most studies on fungal degradation of
diuron have focused either on finding efficient degraders or on the kinetics of diuron degradation,
whereas fewer studies have investigated the biology to illuminate the function of fungal diuron
degradation. Pesticide degrading fungi have sometimes been shown able to utilize the compound as
a source of both carbon and nitrogen (Kulshrestha and Kumari, 2011). However, studies on
phenylurea herbicides show that fungi do not utilize these as a nutrient source (Berger, 1998;
Rønhede et al., 2005), which was further confirmed to also be the case for diuron (Manuscript I). It
therefore appears that it is a co-metabolic process: A process probably occurring in close proximity
of the fungal hyphae, where enzymes are excreted, since no further degradation occurred after
removal of the fungal mycelium (Manuscript I). Studies so far have not been able to determine
which enzymes are responsible for the process. Also, Bending et al. (2002) could not find any
relationship between ligninolytic activity and degradation of diuron.
Berger (1998) stated that phenylurea herbicides sorbed to the mycelia of certain fungal
species, though the results were somewhat incoherent as contrasting results were obtained when
using non-labeled and 14
C-labeled compounds. In Manuscript I it is shown that the total amount of

23
diuron can be accounted for as parent compound and metabolites at all times during the
degradation, so it is therefore evident that diuron does not sorb to the mycelia of Mortierella.
Previous studies suggest, on the basis of screening a large number of species, that the
degradation potential can vary greatly within a genus. From the results of Vroumsia et al. (2005) it
is seen that the degradation of 2,4-D varies between 0-52% for five Aspergillus species and 11-46%
for four Mortierella species. A similar picture is found for phenylurea herbicides degradation
(Vroumsia et al., 1996). In Manuscript I molecular tools were employed to investigate the
phylogenetic background with regard to diuron degradation within the genus of Mortierella. The
results revealed a link between the phylogenetic position of the fungi and their diuron degradation
potential under the experimental conditions, as the diuron degrading strains constituted a closely
related group. Similar results have also been found for bacterial phenylurea degraders (Hussain et
al., 2009).
All in all, from the above it is evident that fungal degradation of certain pesticides is mainly a
co-metabolic process. A process which is, under certain conditions, linked to specific strains of
Mortierella, probably due to an enzymatic system developed in those polygenetic groups. Whether
this enzyme is continuously produced or onset at the exposure to toxic compounds remains
uncertain.

24
Consortia for bioremediation
Communities of fungi and bacteria have vital functions in the environment, as they separately and
together degrade and mineralize organic compounds (Frey-Klett et al., 2011). It is therefore also
very plausible that consortia consisting of both fungi and bacteria can in some case be more suitable
in a bioremediation context than single strains. However, there are many things to consider when
seeking to construct efficient consortia for bioremediation. Especially the interactions between the
two groups are important to examine to understand the mechanisms within the consortia, avoid
mismatching of organisms, and creation of efficient consortia. In the following section the most
relevant issues are touched upon. Then the specific interaction known as ‗the fungal highway‘ will
be discussed. Finally, studies on consortia of fungal-bacterial degraders are reviewed.
Fungal-bacterial interactions
Fungal and bacterial organisms often share the same microhabitat (Warmink et al., 2009) and as
such interact with each other. In general terms such interactions may be categorized as antagonistic,
neutral or beneficial for the microbes. Antagonism may be the production of antibacterial or
antifungal compounds or simply the competition for nutrients. Competition for nutrients can lead to
great reduction of fungal biomass in the presence of competing bacteria and vice versa (Mille-
Lindblom and Tranvik, 2003), whereas other bacteria may even obtain nutrients by mycophagy i.e.
feeding on living fungi (de Boer et al., 2005; Höppener-Ogawa et al., 2008). A neutral interaction is
one where there is no positive or negative effect of the presence of the other organism, e.g. where
the organisms have different niches. However, what is desirable in a consortium is a beneficial
interaction for it to be efficient for degradation of pesticides. Beneficial interactions can either be
commensal, with one of the organisms benefiting and the other remaining unaffected, or mutual,
with both organisms benefiting from the interaction (Bennett and Feibelman, 2001). It has been
shown that soil fungi can create niches where bacteria can thrive (de Boer et al., 2005). Boersma et
al. (2010) found that the survival of the Variovorax paradoxus-like strain HB44 in sterilized soil
was significantly improved by the presence of the fungus Lyophyllum sp. strain Karsten, and that
the bacterial strain grew readily on compounds, in particular glycerol, released by the fungus. In
addition, Warmink et al. (2009) showed that bacteria found in the mycosphere (fungiphiles) could
utilize specific fungal exudates that bacteria found in the bulk soil could not. This gives the

25
interacting bacteria an advantage in the presence of the fungus where they gain a higher fitness.
However, information about the compounds released by the fungi is still limited (Nazir et al., 2010),
and this area needs further research.
Fungal highway
A more mechanistic type of interaction is the transport of bacteria along the fungal hyphae, termed
the fungal highway (Kohlmeier et al., 2005). Pollutant-degrading bacteria have limited accessibility
to the polluting compound, as well as water and nutrients, in heterogeneous and water-unsaturated
environments e.g. soil. Limited bioavailability is established as perhaps the most critical factor in
bioremediation (Bosma et al., 1996). The fungal highway may, however, improve this in several
ways. First, transport of bacteria can occur in the thin water film around the fungal hyphae (Wick et
al., 2010), consequently leading to a more homogenous distribution of these bacteria in the soil.
Warmink and van Elsas (2009) showed that soil bacteria were transported via the hyphae of
the basidiomycete Lyophyllum sp. strain Karsten. Likewise, Trifonova et al. (2009) found that
certain bacteria were translocated by the hyphae of the ascomycete Conochaeta ligniaria. Besides
transporting bacteria, fungal hyphae have also been shown to enhance transport of chemical
compounds such as sodium salicylate (Furuno et al., 2010), phenanthrene (Furuno et al., 2012), and
2,6-dichlorobenzamide (Manuscript II) in water-unsaturated systems.
Most importantly, it has also been shown that the transport of pollutant-degrading bacteria via
fungal hyphae enhanced the degradation of phenanthrene (Wick et al., 2007), 2,6-
dichlorobenzamide (Manuscript II) and diuron (Manuscript III). Recent findings confirm that
conditions of limited bacterial dispersal also limit degradation and that fungal networks may
drastically improve these adverse conditions for bioremediation in heterogeneous environments
(Banitz et al., 2011a; Banitz et al., 2011b). The ability to increase bacterial dispersal depends,
however, on the fungal strain and the surface hydrophobicity of its hyphae. Studies have shown
bacterial transport to occur almost exclusively along hydrophilic fungal mycelia (Kohlmeier et al.,
2005; Wick et al., 2007). Bacterial transport via the two Mortierella strains LEJ702 and LEJ703
was investigated in Manuscript III, and a clear difference was found for the two strains as bacterial
transport could only be observed in the presence of LEJ702. This could possibly be due to
differences in the surface hydrophobicity of these to Mortierella strains, though this was not
measured.

26
Degradation by consortia
In general, choosing ‗the right‘ organism for bioremediation, be it fungus or bacterium, is of great
importance. Members of both kingdoms have their advantages in a given environment. A possible
means to benefit from this fact could be to create consortia of both bacterial and fungal degraders
(See Fig. 2). Several authors have suggested this as a promising strategy to achieve more efficient
degradation (Bennett et al., 2007; Mikesková et al., 2012). However, very few studies are found in
the literature where isolates of fungi and bacteria are applied as consortia or co-cultures. Boonchan
et al. (2000) examined degradation of PAHs by Penicillium janthinellum in co-culture with S.
maltophilia both isolated from soil. Results showed that the co-culture degraded 5-ringed PAHs
more efficiently than either single strain alone, and from the compounds developed during
degradation the authors concluded that this was due to cooperative catabolism. On the other hand,
Arun and Eyini (2011) found that co-cultures entailing basidiomycetes plus B. pumilus could not
enhance the degradation of 2-4 ringed PAHs.
Examples are seen of fungi or bacteria degrading pesticide-metabolites produced by the other
organism, suggesting that cooperative catabolism could occur in fungal-bacterial consortia.
Rønhede et al. (2007) showed that fungal hydroxylation of isoproturon in soil generally enhanced
the bacterial mineralization of this compound. Furthermore, Tixier et al. (2002) found that fungi
could transform diuron metabolite 3,4-DCA produced by Arthrobacter sp. N2 to the less toxic 3,4-
dichloroacetanilide (3,4-DCAA). We also found 3,4-DCAA to be produced by Mortierella
especially when it was grown in consortia with Arthrobacter globiformis strain D47 (Manuscript
III), and though this transformation might not lead to mineralization the process reduces the
concentration of the most toxic diuron metabolite.
To my knowledge there are no previous reports on fungal-bacterial consortia constructed for
degradation of pesticides. However, it has been shown that bacterial co-cultures greatly enhance
degradation of phenylurea herbicides (Sørensen et al., 2002; Sørensen et al., 2008). In Manuscript
III it was examined whether consortia of fungal and bacterial diuron degraders could achieve more
efficient degradation than the single strains and the interactions between the organisms were
investigated. Diuron mineralization results showed that the three-member consortium entailing
Variovorax sp. SRS16, Arthrobacter globiformis strain D47 and Mortierella sp. LEJ702 was

27
superior to both the single strains and the other consortia tested in sand columns. In this experiment
the same advantage could not be found with the other fungal strain tested. This is somewhat in
agreement with Machín-Ramírez et al. (2010) who found that certain defined co-cultures of fungi
and bacteria could enhance the degradation of a 5-ringed PAH, while other combinations of fungal-
bacterial degraders did not.
Figure 2. The possible
synergistic effects within a
fungal-bacterial consortium
leading to enhanced
biodegradation of the
pesticide. Bacteria often
perform best in water
saturated systems were the
accessibility to the pesticide
is high. Filamentous fungi
are well adapted to
heterogeneous environments
like soil as the hyphae may
for instance grow through
air-filled gaps. As described
in the text interactions in a
fungal-bacteria consortium
can lead to superior
degradation efficiency.
In Figure 2 the most important synergistic effects within a fungal-bacterial consortium leading
to enhanced biodegradation are illustrated. Above all is the ability of the organisms to co-exist as it
is the foundation of any synergistic relationship. If one of the organisms in a consortium severely
inhibits the growth or activity of the other, then the benefit of having them together will be
eliminated. In Manuscript III, for instance, the presence of A. globiformis D47 inhibited the
growth of the two Mortierella strains. However, this effect was somewhat alleviated in the three-

28
member consortium Variovorax sp. SRS16, A. globiformis D47 and Mortierella sp. LEJ702
possibly contributing to enhanced degradation by this consortium.
From the results obtained in Manuscript III and the studies discussed above it is clear that
consortia of fungal-bacterial degraders can yield more efficient degradation of pollutants.
Nevertheless, these studies also show that care, consideration and research are needed to construct
and investigate these consortia as the organisms employed must be compatible in several ways.

29
Conclusions and perspectives
This thesis has through the results obtained and presented in the enclosed manuscripts contributed
with new knowledge on the degradation of pesticides by fungi. In Manuscript I it was established
that Mortierella does not utilize the pesticide diuron as a carbon or nitrogen source, and it was
therefore concluded that the degradation occurs as a co-metabolic process. Furthermore, it was
shown that there, under the experimental conditions of this study, was a phylogenetic relationship
between the diuron degrading strains. Lastly, a previously unknown diuron metabolite, one that
might be specific for fungal degradation, was found and identified.
Future research should focus on examining the enzymatic activity of the fungal pesticide
degrades; seeking to elucidate the key enzyme(s) for the degradation. The activity of these enzymes
may be linked to certain phylogenetic groups as suggested in Manuscript I, and studying this could
bring us one step closer to discovering the genes responsible for the degradation.
Differences in the fungal degradation by individual strains (e.g. LEJ702 or LEJ703) in
Manuscript I, II and III points to that abiotic factors like temperature, moisture and growth matrix
influence fungal pesticide degradation. This is not surprising but may also be an area of further
study.
The other parts of the work focused on creating and investigating consortia for bioremediation
of pesticides. Successful construction of fungal-bacterial consortia for degradation of diuron and
BAM was achieved and presented in Manuscript II and III, respectively. In addition, the
hypothesis that these consortia would be more efficient than single strains for biodegradation in
unsaturated systems was confirmed, though this depended greatly on which organisms where
included in the specific consortium. It therefore also proved useful to examine the interactions
between the fungal and bacterial strains within the consortia to elucidate which mechanism(s)
where responsible for the differences in degradation efficiency (e.g. for consortia with Mortierella
sp. LEJ702 or LEJ703). For instance, Mortierella sp. LEJ702 mediated the transport not only of the
BAM-degrading Aminobacter sp. MSH1, but also of the diuron-degrading Variovorax sp. SRS16
and Arthrobacter globiformis D47, whereas no bacterial transport where found in consortia with
Mortierella sp. LEJ703. In addition, the growth of these two Mortierella strains was not equally
affected by the presence of the bacteria in the three-member consortia with Variovorax sp. SRS16
and Arthrobacter globiformis D47. Therefore, this underlines the importance of careful strain

30
selection when constructing consortia for bioremediation, as there are clearly differences between
strains, even within the same genus, with regard to the degradation efficiency and the extent of
interactions with the other members of the consortium. That Mortierella sp. LEJ702 worked well in
consortia for both diuron and BAM degradation suggests this as a superior candidate strain for
further research on fungal-bacterial consortia for biodegradation of pesticides.
The BAM degrading consortium and the most efficient diuron degrading consortium both
have potential for use for bioremediation of soils contaminated with these compounds. Indeed it is
possible that the synergistic effect may be even greater in soil where the pesticides are
heterogeneously distributed and sorbed to soil particles. Here the fungal mediated transport of
degrader bacteria could prove essential and the extent of this transport should be monitored along
with the survival of the introduced strains and the pesticide degradation.
A challenge when moving towards applying consortia such as these for soil bioremediation
may be that the heterogeneity of organic compounds and microbes in the soil environment might
destabilize the consortium, possibly leading to poor degradation and/or accumulation of toxic
metabolites. The interactions with indigenous soil microorganisms should therefore be thoroughly
investigated together with performance of the consortium in soil mesocosms with regard to its
degradation efficiency, before establishing its applicability for in situ bioremediation.
Formulation of the consortia in carriers could be a possible means to achieve successful
introduction and stabile growth of the consortia in the soil matrix. The microorganisms would be
embedded in synthetic beads supplemented with carbon and nitrogen sources supporting them in the
first phase of growth after introduction into the soil environment. The microorganisms may in this
way reach a sizeable biomass and be better fit for competition with indigenous organisms. Since the
strains applied in these consortia are isolated from soil there is a real possibility that they can in fact
be successfully re-inoculated into the soil matrix.

31
Reference list
Ahemad, M., Khan, M.S., Zaidi, A., Wani, P.A., 2008. Remediation of herbicides contaminated
soil using microbes. in: Khan, M.S., Zaidi, A., Mussarrat, J. (Eds.). Microbes in sustainable
agriculture. Nova Science Publishers Inc., New York, pp. 261-284.
Alkorta, I., Garbisu, C., 2001. Phytoremediation of organic contaminants in soils. Bioresour.
Technol. 79, 273-276.
Andersson, B.E., Lundstedt, S., Tornberg, K., Schnurer, Y., Oberg, L.G., Mattiasson, B.,
2003. Incomplete degradation of polycyclic aromatic hydrocarbons in soil inoculated with
wood-rotting fungi and their effect on the indigenous soil bacteria. Environ. Toxicol. Chem.
22, 1238-1243.
Arun, A., Eyini, M., 2011. Comparative studies on lignin and polycyclic aromatic hydrocarbons
degradation by basidiomycetes fungi. Bioresour. Technol. 102, 8063-8070.
Ayansina, A.D.V., 2008. Pesticide use in agriculture and microoganisms. in: Khan, M.S., Zaidi, A.,
Mussarrate, J. (Eds.). Microbes in sustainable agriculture. Nova Science Publishers, New
York, pp. 243-260.
Badawi, N., Ronhede, S., Olsson, S., Kragelund, B.B., Johnsen, A.H., Jacobsen, O.S., Aamand,
J., 2009. Metabolites of the phenylurea herbicides chlorotoluron, diuron, isoproturon and
linuron produced by the soil fungus Mortierella sp. Environ. Pollut. 157, 2806-2812.
Baldrian, P., 2008. Wood-inhabiting ligninolytic basidiomycetes in soils: Ecology and constraints
for applicability in bioremediation. Fungal Ecol. 1, 4-12.
Banitz, T., Fetzer, I., Johst, K., Wick, L.Y., Harms, H., Frank, K., 2011a. Assessing
biodegradation benefits from dispersal networks. Ecol. Model. 222, 2552-2560.
Banitz, T., Wick, L.Y., Fetzer, I., Frank, K., Harms, H., Johst, K., 2011b. Dispersal networks
for enhancing bacterial degradation in heterogeneous environments. Environ. Pollut. 159,
2781-2788.
Bending, G.D., Friloux, M., Walker, A., 2002. Degradation of contrasting pesticides by white rot
fungi and its relationship with ligninolytic potential. FEMS Microbiol. Lett. 212, 59-63.
Bennett, J.W., Connick, W.J., Daigle, D., Wunch, K., 2007. Formulation of fungi for in situ
bioremediation. in: Gadd, G.M., Watkinson, S.C., Dyer, P.S. (Eds.). Fungi in the
Environment. Cambridge University Press, Cambridge, UK, pp. 97-112.

32
Bennett, J.W., Feibelman, T., 2001. Fungal bacterial interactions. in: Hock, B. (Ed.). The Mycota
IX - Fungal Associations. Springer, Berlin.
Berger, B.M., 1998. Parameters Influencing Biotransformation Rates of Phenylurea Herbicides by
Soil Microorganisms. Pestic. Biochem. Phys. 60, 71-82.
Boddy, L., Jones, T.H., 2007. Mycelial responses in heterogeneous environments: parallels with
macroorganisms. in: Gadd, G.M., Watkinson, S.C., Dyer, P.S. (Eds.). Fungi in the
environment. Cambridge University Press, Cambridge, UK, pp. 112-140.
Boersma, F.G.H., Otten, R., Warmink, J.A., Nazir, R., van Elsas, J.D., 2010. Selection of
Variovorax paradoxus-like bacteria in the mycosphere and the role of fungal-released
compounds. Soil Biol. Biochem. 42, 2137-2145.
Boonchan, S., Britz, M.L., Stanley, G.A., 2000. Degradation and mineralization of high-
molecular-weight polycyclic aromatic hydrocarbons by defined fungal-bacterial cocultures.
Appl. Environ. Microbiol. 66, 1007-1019.
Boopathy, R., 2000. Factors limiting bioremediation technologies. Bioresour. Technol. 74, 63-67.
Bosma, T.N.P., Middeldorp, P.J.M., Schraa, G., Zehnder, A.J.B., 1996. Mass Transfer
Limitation of Biotransformation: Quantifying Bioavailability. Environ. Sci. Technol. 31,
248-252.
Brüsch, W., 2010. Pesticider. in: Thorling, L. (Ed.). Grundvandsovervågning 2009. GEUS,
Denmark, pp. 62-77.
Carlile, M.J., Watkinson, S.C., Gooday, G.W., 2001. The Fungi. Elsevier Academic Press,
London, UK.
Clausen, L., Larsen, F., Albrechtsen, H.-J., 2004. Sorption of the Herbicide Dichlobenil and the
Metabolite 2,6-Dichlorobenzamide on Soils and Aquifer Sediments. Environ. Sci. Technol.
38, 4510-4518.
Cox, C., 2003. Herbicide factsheet - DIURON. Journal of Pesticide Reform 23, 12-20.
Das, M., Adholeya, A., 2012. Role of Microorganisms in Remediation of Contaminated Soil. in:
Satyanarayana, T., Johri, B.N., Prakash, A. (Eds.). Microorganisms in Environmental
Management. Springer Netherlands, pp. 81-111.
de Boer, W., Folman, L.B., Summerbell, R.C., Boddy, L., 2005. Living in a fungal world: impact
of fungi on soil bacterial niche development. FEMS Microbiol. Rev. 29, 795-811.

33
Dechesne, A., Wang, G., Gulez, G., Or, D., Smets, B.F., 2010. Hydration-controlled bacterial
motility and dispersal on surfaces. Proc. Natl. Acad. Sci. U. S. A. 107, 14369-14372.
Domsch, K.H., Gams, W., Anderson, T.-H., 1980. Compendium of soil fungi. Academic Press,
London, UK.
El Fantroussi, S., Agathos, S.N., 2005. Is bioaugmentation a feasible strategy for pollutant
removal and site remediation? Curr. Opin. Microbiol. 8, 268-275.
Eriksson, E., Baun, A., Mikkelsen, P.S., Ledin, A., 2007. Risk assessment of xenobiotics in
stormwater discharged to Harrestrup Å, Denmark. Desalination 215, 187-197.
European Council, E.U., 1980. Drinking Water Directive (Directive 80/778/EEC). European
Council
European Parliament, E.U., 2008. Priority Substances Directive (Directive 2008/105/EC).
Official Journal of the European Union.
Fomina, M., Ritz, K., Gadd, G.M., 2003. Nutritional influence on the ability of fungal mycelia to
penetrate toxic metal-containing domains. Mycol. Res. 107, 861-871.
Fratila-Apachitei, L.E., Hirst, J.A., Siebel, M.A., Gijzen, H.J., 1999. Diuron degradation by
Phanerochaete chrysosporium BKM- F-1767 in synthetic and natural media. Biotechnol.
Lett. 21, 147-154.
Frey-Klett, P., Burlinson, P., Deveau, A., Barret, M., Tarkka, M., Sarniguet, A., 2011.
Bacterial-Fungal Interactions: Hyphens between Agricultural, Clinical, Environmental, and
Food Microbiologists. Microbiol. Mol. Biol. Rev. 75, 583-609.
Furuno, S., Foss, S., Wild, E., Jones, K.C., Semple, K.T., Harms, H., Wick, L.Y., 2012. Mycelia
Promote Active Transport and Spatial Dispersion of Polycyclic Aromatic Hydrocarbons.
Environ. Sci. Technol. 46, 5463-5470.
Furuno, S., Päzolt, K., Rabe, C., Neu, T.R., Harms, H., Wick, L.Y., 2010. Fungal mycelia allow
chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-
unsaturated systems. Environ. Microbiol. 12, 1391-1398.
Gadd, G.M., 2007. Geomycology: biogeochemical transformations of rocks, minerals, metals and
radionuclides by fungi, bioweathering and bioremediation. Mycol. Res. 111, 3-49.
Gao, D., Du, L., Yang, J., Wu, W.-M., Liang, H., 2010. A critical review of the application of
white rot fungus to environmental pollution control. Crit. Rev. Biotechnol. 30, 70-77.

34
Gerhardt, K.E., Huang, X.-D., Glick, B.R., Greenberg, B.M., 2009. Phytoremediation and
rhizoremediation of organic soil contaminants: Potential and challenges. Plant Sci. 176, 20-
30.
Giacomazzi, S., Cochet, N., 2004. Environmental impact of diuron transformation: a review.
Chemosphere 56, 1021-1032.
Grube, A., Donaldson, D., Kiely, T., Wu, L., 2011. Pesticides Industry Sales and Usage. U.S.
Environmental Protection Agency.
Hangler, M., Jensen, B., Rønhede, S., Sørensen, S.R., 2007. Inducible hydroxylation and
demethylation of the herbicide isoproturon by Cunninghamella elegans. FEMS Microbiol.
Lett. 268, 254-260.
Harms, H., Schlosser, D., Wick, L.Y., 2011. Untapped potential: exploiting fungi in
bioremediation of hazardous chemicals. Nat. Rev. Microbiol. 9, 177-192.
Hawksworth, D.L., 1991. The fungal dimension of biodiversity: magnitude, significance, and
conservation. Mycol. Res. 95, 641-655.
Hawksworth, D.L., 2001. The magnitude of fungal diversity: the 1.5 million species estimate
revisited. Mycol. Res. 105, 1422-1432.
Ho, S.Y., Chen, F., 2008. Genetic characterization of Mortierella alpina by sequencing the 18S-
28S ribosomal gene internal transcribed spacer region. Lett. Appl. Microbiol. 47, 250-255.
Holtze, M.S., Hansen, H.C.B., Juhler, R.K., Sørensen, J., Aamand, J., 2007. Microbial
degradation pathways of the herbicide dichlobenil in soils with different history of
dichlobenil-exposure. Environ. Pollut. 148, 343-351.
Höppener-Ogawa, S., Leveau, J.H.J., van Veen, J.A., De Boer, W., 2008. Mycophagous growth
of Collimonas bacteria in natural soils, impact on fungal biomass turnover and interactions
with mycophagous Trichoderma fungi. ISME J 3, 190-198.
Hussain, S., Sørensen, S.R., Devers-Lamrani, M., El-Sebai, T., Martin-Laurent, F., 2009.
Characterization of an isoproturon mineralizing bacterial culture enriched from a French
agricultural soil. Chemosphere 77, 1052-1059.
Juwarkar, A., Singh, S., Mudhoo, A., 2010. A comprehensive overview of elements in
bioremediation. Rev. Environ. Sci. Biotechnol. 9, 215-288.
Kataoka, R., Takagi, K., Sakakibara, F., 2011. Biodegradation of endosulfan by Mortieralla sp.
strain W8 in soil: Influence of different substrates on biodegradation. Chemosphere 85, 548-
552.

35
Khadrani, A., Seigle-Murandi, F., Steiman, R., Vroumsia, T., 1999. Degradation of three
phenylurea herbicides (chlortoluron, isoproturon and diuron) by micromycetes isolated from
soil. Chemosphere 38, 3041-3050.
Kjøller, A., Struwe, S., 1990. Decomposition of beech litter. A comparison of fungi isolated on
nutrien rich and nutrient poor media. Trans. Mycol. Soc. Japan 31, 5-16.
Kjøller, A.H., Struwe, S., 2002. Fungal communities, succession, enzymes, and decomposition. in:
Burns, R.G., Dick, R.P. (Eds.). Enzymes in the environment. Marcel Dekker, New York, pp.
267-284.
Kohlmeier, S., Smits, T.H.M., Ford, R.M., Keel, C., Harms, H., Wick, L.Y., 2005. Taking the
fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environ. Sci.
Technol. 39, 4640-4646.
Kulshrestha, G., Kumari, A., 2011. Fungal degradation of chlorpyrifos by Acremonium sp. strain
(GFRC-1) isolated from a laboratory-enriched red agricultural soil. Biol. Fertil. Soils 47,
219-225.
Lang, E.K., Kleeberg, I., Zadrazil, F., 1997. Competition of Pleurotus sp. and Dichomitus
squalens with soil microorganisms during lignocellulose decomposition. Bioresour.
Technol. 60, 95-99.
Lapworth, D.J., Gooddy, D.C., 2006. Source and persistence of pesticides in a semi-confined
chalk aquifer of southeast England. Environ. Pollut. 144, 1031-1044.
Leung, K.T., Nandakumar, K., Sreekumari, K., Lee, H., Trevors, J.T., 2007. Biodegradation
and bioremediation of organic pollutants in soil. in: van Elsas, J.D., Jansson, J.K., Trevors,
J.T. (Eds.). Modern Soil Microbiology. CRC Press, Florida, pp. 521-551.
Lindahl, B.D., Olsson, S., 2004. Fungal translocation – creating and responding to environmental
heterogeneity. Mycologist 18, 79-88.
Machín-Ramírez, C., Morales, D., Martínez-Morales, F., Okoh, A.I., Trejo-Hernández, M.R.,
2010. Benzo[a]pyrene removal by axenic- and co-cultures of some bacterial and fungal
strains. Int. Biodeter. Biodegr. 64, 538-544.
Martins, M., Dairou, J., Rodrigues-Lima, F., Dupret, J.-M., Silar, P., 2010. Insights into the
Phylogeny or Arylamine N-Acetyltransferases in Fungi. J. Mol. Evol. 71, 141-152.
Martins, M., Rodrigues-Lima, F., Dairou, J., Lamouri, A., Malagnac, F., Silar, P., Dupret, J.-
M., 2009. An Acetyltransferase Conferring Tolerance to Toxic Aromatic Amine Chemicals.
J. Biol. Chem. 284, 18726-18733.

36
Mikesková, H., Novotný, Č., Svobodová, K., 2012. Interspecific interactions in mixed microbial
cultures in a biodegradation perspective. Appl. Microbiol. Biotechnol., 1-10.
Mille-Lindblom, C., Tranvik, L.J., 2003. Antagonism between Bacteria and Fungi on
Decomposing Aquatic Plant Litter. Microb. Ecol. 45, 173-182.
Nagy, L.G., Petkovits, T., Kovács, G.M., Voigt, K., Vágvölgyi, C., Papp, T., 2011. Where is the
unseen fungal diversity hidden? A study of Mortierella reveals a large contribution of
reference collections to the identification of fungal environmental sequences. New Phytol.
191, 789-794.
Nazir, R., Warmink, J.A., Boersma, H., Van Elsas, J.D., 2010. Mechanisms that promote
bacterial fitness in fungal-affected soil microhabitats. FEMS Microbiol. Ecol. 71, 169-185.
Pointing, S.P., 2001. Feasibility of bioremediation by white-rot fungi. Appl. Microbiol. Biotechnol.
57, 20-33.
Porazzi, E., Pardo Martinez, M., Fanelli, R., Benfenati, E., 2005. GC–MS analysis of
dichlobenil and its metabolites in groundwater. Talanta 68, 146-154.
Read, N.D., 2007. Environmental sensing and the filamentous fungal lifestyle. in: Gadd, G.M.,
Watkinson, S.C., Dyer, P.S. (Eds.). Fungi in the environment. Cambridge University Press,
Cambridge, UK, pp. 38-57.
Reddy, C.A., Mathew, A., 2001. Bioremediation potential of white rot fungi. in: Gadd, G.M. (Ed.).
Fungi in bioremediation. Cambridge University Press, Cambridge, UK, pp. 52-78.
Ritz, K., Young, I.M., 2004. Interactions between soil structure and fungi. Mycologist 18, 52-59.
Rønhede, S., Jensen, B., Rosendahl, S., Kragelund, B.B., Juhler, R.K., Aamand, J., 2005.
Hydroxylation of the herbicide isoproturon by fungi isolated from agricultural soil. Appl.
Environ. Microbiol. 71, 7927-7932.
Rønhede, S., Sørensen, S.R., Jensen, B., Aamand, J., 2007. Mineralization of hydroxylated
isoproturon metabolites produced by fungi. Soil Biol. Biochem. 39, 1751-1758.
Salicis, F., Krivobok, S., Jack, M., Benoit-Guyod, J.L., 1999. Biodegradation of fluoranthene by
soil fungi. Chemosphere 38, 3031-3039.
Silar, P., Dairou, J., Cocaign, A., Busi, F., Rodrigues-Lima, F., Dupret, J.-M., 2011. Fungi as a
promising tool for bioremediation of soils contaminated with aromatic amines, a major class
of pollutants. Nat Rev Micro 9, 477-477.

37
Simonsen, A., Holtze, M.S., Sørensen, S.R., Sørensen, S.J., Aamand, J., 2006. Mineralisation of
2,6-dichlorobenzamide (BAM) in dichlobenil-exposed soils and isolation of a BAM-
mineralising Aminobacter sp. Environ. Pollut. 144, 289-295.
Singh, H., 2006. Mycoremediation: Fungal bioremediation. John Wiley & Sons Inc, New Jersey.
Sørensen, S.R., Albers, C.N., Aamand, J., 2008. Rapid mineralization of the phenylurea herbicide
diuron by Variovorax sp. SRS16 in pure culture and within a two-member consortium.
Appl. Environ. Microbiol. 74, 2332-2340.
Sørensen, S.R., Bending, G.D., Jacobsen, C.S., Walker, A., Aamand, J., 2003. Microbial
degradation of isoproturon and related phenylurea herbicides in and below agricultural
fields. FEMS Microbiol. Ecol. 45, 1-11.
Sørensen, S.R., Holtze, M.S., Simonsen, A., Aamand, J., 2007. Degradation and mineralization
of nanomolar concentrations of the herbicide dichlobenil and its persistent metabolite 2,6-
dichlorobenzamide by Aminobacter spp. isolated from dichlobenil-treated soils. Appl.
Environ. Microbiol. 73, 399-406.
Sørensen, S.R., Ronen, Z., Aamand, J., 2001. Isolation from agricultural soil and characterization
of a Sphingomonas sp. able to mineralize the phenylurea herbicide isoproturon. Appl.
Environ. Microbiol. 67, 5403-5409.
Sørensen, S.R., Ronen, Z., Aamand, J., 2002. Growth in Coculture Stimulates Metabolism of the
Phenylurea Herbicide Isoproturon by Sphingomonas sp. Strain SRS2. Appl. Environ.
Microbiol. 68, 3478-3485.
Struger, J., Grabuski, J., Cagampan, S., Rondeau, M., Sverko, E., Marvin, C., 2011.
Occurrence and distribution of sulfonylurea and related herbicides in central Canadian
surface waters 2006-2008. Bull. Environ. Contam. Toxicol. 87, 420-425.
Thompson, I.P., van der Gast, C.J., Ciric, L., Singer, A.C., 2005. Bioaugmentation for
bioremediation: the challenge of strain selection. Environ. Microbiol. 7, 909-915.
Thorn, R.G., Lynch, M.D.J., 2007. Fungi and eukaryotic algae. in: Paul, E.A. (Ed.). Soil
Microbiology, Ecology, and Biochemistry. Academic Press, Oxford, pp. 145-162.
Tillmanns, G.M., Wallnöfer, P.R., Engelhardt, G., Olie, K., Hutzinger, O., 1978. Oxidative
dealkylation of five phenylurea herbicides by the fungus Cunninghamella echinulata
thaxter. Chemosphere 7, 59-64.
Tixier, C., Bogaerts, P., Sancelme, M., Bonnemoy, F., Twagilimana, L., Cuer, A., Bohatier, J.,
Veschambre, H., 2000. Fungal biodegradation of a phenylurea herbicide, diuron : structure
and toxicity of metabolites. Pest Manag. Sci. 56, 455-462.

38
Tixier, C., Sancelme, M., Aït-Aïssa, S., Widehem, P., Bonnemoy, F., Cuer, A., Truffaut, N.,
Veschambre, H., 2002. Biotransformation of phenylurea herbicides by a soil bacterial
strain, Arthrobacter sp. N2: structure, ecotoxicity and fate of diuron metabolite with soil
fungi. Chemosphere 46, 519-526.
Tixier, C., Sancelme, M., Bonnemoy, F., Cuer, A., Veschambre, H., 2001. Degradation products
of a phenylurea herbicide, diuron: Synthesis, ecotoxicity, and biotransformation. Environ.
Toxicol. Chem. 20, 1381-1389.
Tomlin, C.D.S., 1997. The pesticide manual. The British Crop Protection Council, Surrey, UK.
Torstensson, L., 2001. Use of herbicides on railway tracks in Sweden. Pestic. Outlook 12, 16-21.
Trifonova, R., Postma, J., Van Elsas, J.D., 2009. Interactions of plant-beneficial bacteria with the
ascomycete Coniochaeta ligniaria. J. Appl. Microbiol. 106, 1859-1866.
Turnbull, G.T., Cullington, J.C., Walker, A.W., Morgan, J.M., 2001. Identification and
characterisation of a diuron-degrading bacterium. Biol. Fertil. Soils 33, 472-476.
Vroumsia, T., Steiman, R., Seigle-Murandi, F., Benoit-Guyod, J.L., Gedexe, 2005. Fungal
bioconversion of 2,4-dichlorophenoxyacetic acid (2,4-D) and 2,4-dichlorophenol (2,4-DCP).
Chemosphere 60, 1471-1480.
Vroumsia, T., Steiman, R., SeigleMurandi, F., BenoitGuyod, J.L., Khadrani, A., 1996.
Biodegradation of three substituted phenylurea herbicides (chlorotoluron, diuron, and
isoproturon) by soil fungi. A comparative study. Chemosphere 33, 2045-2056.
Wang, G., Or, D., 2010. Aqueous films limit bacterial cell motility and colony expansion on
partially saturated rough surfaces. Environ. Microbiol. 12, 1363-1373.
Warmink, J.A., Nazir, R., Van Elsas, J.D., 2009. Universal and species-specific bacterial
‗fungiphiles‘ in the mycospheres of different basidiomycetous fungi. Environ. Microbiol.
11, 300-312.
Warmink, J.A., van Elsas, J.D., 2009. Migratory Response of Soil Bacteria to Lyophyllum sp.
Strain Karsten in Soil Microcosms. Appl. Environ. Microbiol. 75, 2820-2830.
Weinberger, M., Bollag, J.-M., 1972. Degradation of Chlorbromuron and Related Compounds by
the Fungus Rhizoctonia solani. Appl. Microbiol. 24, 750-754.
Wick, L.Y., Furuno, S., Harms, H., 2010. Fungi as Transport Vectors for Contaminants and
Contaminant-Degrading Bacteria. in: Timmis, K.N. (Ed.). Handbook of Hydrocarbons and
Lipid Microbiology. Springer Berlin, pp. 1555-1561.

39
Wick, L.Y., Remer, R., Wurz, B., Reichenbach, J., Braun, S., Scharfer, F., Harms, H., 2007.
Effect of fungal hyphae on the access of bacteria to phenanthrene in soil. Environ. Sci.
Technol. 41, 500-505.
Wong, P.T.W., Griffin, D.M., 1976. Bacterial movement at high matric potentials—I. In artificial
and natural soils. Soil Biol. Biochem. 8, 215-218.
Wösten, H.A.B., van Wetter, M.A., Lugones, L.G., van der Mei, H.C., Busscher, H.J., Wessels,
J.G.H., 1999. How a fungus escapes the water to grow into the air. Curr. Biol. 9, 85-88.

40

Manuscript I
DEGRADATION OF THE PHENYLUREA HERBICIDE DIURON BY SOIL FUNGI:
DIFFERENT DEGRADATION POTENTIALS OF FIVE MORTIERELLA STRAINS
Lea Ellegaard-Jensen, Jens Aamand, Birthe B. Kragelund, Anders H. Johnsen and Søren Rosendahl
In revision for Biodegradation


43
Degradation of the phenylurea herbicide diuron by soil
fungi: different degradation potentials of five Mortierella
strains
Lea Ellegaard-Jensena,b *, Jens Aamandb, Birthe B. Kragelundc, Anders H. Johnsend and Søren Rosendahla.
a Department of Biology, Copenhagen University, Universitetsparken 15, DK-2100 Copenhagen Ø,
Denmark. b Department of Geochemistry, Geological Survey of Denmark and Greenland (GEUS), Øster
voldgade 10, DK-1350 Copenhagen K, Denmark.c Structural Biology and NMR Laboratory, Department of
Biology, University of Copenhagen, Ole Maaloes Vej 5, DK-2200 Copenhagen N, Denmark. d Department of
Clinical Biochemistry, Rigshospitalet, Blegdamsvej 9, DK-2100 Copenhahagen Ø, Denmark
* Corresponding author: Tel.: 45 3532 2257; Fax: 45 3532 2321; E-mail: [email protected]
Abstract Microbial pesticide degradation studies have until now mainly focused on bacteria, although fungi
have also been shown to degrade pesticides. In this study we clarify the background for the ability of the
common soil fungus Mortierella to degrade the phenylurea herbicide diuron. Diuron degradation potentials
of five Mortierella strains were compared, and the role of carbon and nitrogen for the degradation process
was investigated. Results showed that strains able to degrade diuron constituted a closely related group,
indicating that diuron degradation was not a general feature, but limited to a phylogenetic group within
Mortierella. Degradation of diuron was fastest in carbon and nitrogen rich media while suboptimal nutrient
levels restricted degradation, making it unlikely that Mortierella utilize diuron as carbon or nitrogen sources.
Degradation kinetics showed that diuron degradation was followed by formation of the metabolites 1-(3,4-
dichlorophenyl)-3-methylurea, 1-(3,4-dichlorophenyl)urea and an hitherto unknown metabolite suggested to
be 1-(3,4-dichlorophenyl)-3-methylideneurea.
Keywords Fungal biodegradation, Phenylurea herbicide, Diuron, Fungal genetics, Phylogenetic
relationships

44
Introduction
Pesticide pollution is one of the major concerns regarding contamination of the environment.
Pesticides are mainly used on agricultural land, but also in private gardens, along railways and other
public areas. Herbicides account for the largest part of overall pesticide use worldwide (Grube et al.
2011). Among these are the phenylurea herbicides which are frequently found as environmental
pollutants (Eriksson et al. 2007; Lapworth and Gooddy 2006; Struger et al. 2011). The phenylurea
herbicide diuron [N-(3,4-dichlorophenyl)-N,N-dimethyl-urea] is on the EU Water Framework
Directive´s list of priority substances (European Parliament 2008) and as it is often found in
groundwater at concentrations exceeding the EU limit of 0.1µg l-1
(Torstensson 2001; Lapworth and
Gooddy 2006) biodegradation of this compound is a central issue.
The majority of studies on microbial pesticide degradation have focused on bacteria (e.g. El-
Bestawy and Albrechtsen 2007; El-Deeb et al. 2000; Sørensen et al. 2008; Simonsen et al. 2006),
whereas fewer studies examine the role of fungi (Harms et al. 2011). Several fungi are known to
produce enzymes that are able to degrade aromatic herbicides and have therefore been suggested as
candidates for bioremediation (Entry et al. 1996).
Fungi can account for up to 75% of soil microbial biomass (Harms et al. 2011) and their
hyphae can grow to a length of 102-10
4 m g
-1 soil (Ritz and Young 2004). Fungi may play an
important role in bioremediation of soil as the fungal hyphae can grow into micropores between soil
aggregates and through air filled gaps (Wösten et al. 1999) and in this way gain a better access to
the compound to be degraded.
Earlier studies have shown that certain fungal species/strains have the ability to degrade
diuron (Badawi et al. 2009; Khadrani et al. 1999; Tixier et al. 2000; Tixier et al. 2001; Vroumsia et
al. 1996). These studies have given insight into formation of different metabolites as well as toxicity
of the parent compound and the metabolites. The studies usually include a number of species and
consequently compare differences in diuron degradation potential among fungal species. Such
differences may likely reflect differences in enzymatic abilities and adaptation to different habitats,
but little is known about the variation within fungal genera or species concerning their ability to
degrade pesticides.
The common soil fungus Mortierella is one of the fungi that are capable of degrading
phenylurea herbicides e.g. isoproturon (Rønhede et al. 2005) and diuron (Tixier et al. 2001; Tixier
et al. 2000; Vroumsia et al. 1996). Nevertheless, it is not known whether this ability is a general
characteristic of the genus or the tested isolates were unique in this respect. In the present study we

45
therefore compare Mortierella isolates, originating from the same agricultural field, to investigate if
the ability to degrade diuron is restricted to a specific phylogenetic group or it is a general feature in
the genus.
Though many microorganisms have been shown to degrade pesticides, the physiological
significance of the degradation remains unclear. For some pesticide degrading bacteria it has been
shown that they are able to utilize the compound as a carbon or a nitrogen source (e.g. El-Deeb et
al. 2000). Whether the same could be the case for fungal degradation remains unclear.
Alternatively, degradation could also be a co-metabolic detoxification process, as some compounds
may share characteristics with natural compounds. To answer these questions we investigate if
Mortierella can utilize diuron as a carbon and a nitrogen source, or whether degradation is caused
by co-metabolism.
Materials and methods
Chemicals and media
Analytical-grade standards were purchased from Dr. Ehrenstorfer GmbH (Augsburg, Germany):
Diuron [N-(3,4-dichlorophenyl)-N,N-dimethyl-urea] (CAS no. 330-54-1; 97.5% purity), DCPMU
[1-(3,4-dichlorophenyl)-3-methylurea] (CAS no. 3567-62-2; 97.5% purity), DCPU [1- (3,4-
dichlorophenyl) urea] (CAS no. 2327-02-8; 99% purity), and 3,4-DCA [3,4-dichloroaniline] (CAS
no. 95-76-1; 99% purity). 3,4-DCAA [3,4-dichloroacetanilide] (CAS no. 2150-93-8) was purchased
from Sigma-Aldrich (St. Louis, Missouri). Acetonitrile-D3 was from Eurisotop (99.8% D).
For fungal isolation the following growth media were used: corn meal agar (CMA), synthetic
nutrient deficient agar (SNA), and potato dextrose agar (PDA) all autoclaved and supplied with
ampicillin (5ml l-1
) and streptomycin (5ml l-1
). In addition modified Melin Norkans medium
(MMN), with the fungicide Benomyl (trade name: Benlate, DuPont), was synthesised according to
Kjøller and Bruns (2003). After isolation all fungi were maintained on PDA plates. Liquid glucose-
mineral media used for degradation experiments were made according to Badawi et al. (2009).

46
Isolation and characterization of fungi
Fungi were isolated from an agricultural field in Græse, Denmark (55° 51' 51 N, 12° 5' 44 E) with a
previous history of phenylurea (isoproturon) application. For isolation CMA, SNA, PDA and MMN
media all with ampicillin and streptomycin were used to select for different fungal strains with
different growth requirements.
To characterize the isolates, DNA was extracted from fungal mycelia using the following
protocol: 20µl Chelex, 40µl TE buffer and mycelia heated to 95oC for 2 min followed by
centrifugation at 13.000g for 2 min, DNA now in the supernatant was transferred to clean tubes and
frozen (-18 oC). PCR amplification of the internal transcribed spacer (ITS) region was performed as
described by Gardes and Bruns (1996) with the following modifications: Amplification for 35
cycles consisting of denaturation for 35 s at 94°C, annealing for 55 s at 55°C and extension at 72°C
for 45 s rising with 4 s per cycle. 10 × diluted fungal DNA was amplified with primers ITS 1F
(Gardes and Bruns 1993) and ITS4 (White et al. 1990). PCR products were sequenced by Macrogen
(Seuol, Korea) and the sequences aligned in MEGA version 5 (www.megasoftware.net/index.php).
The most similar sequences were found by BLAST search in Genbank. The phylogenetic
relationship between the isolates and related sequences from Genbank was made by Maximum
likelihood in MEGA version 5 (Tamura et al. 2011), and the robustness of the phylogenetic tree
tested by 1000 bootstraps.
Fungal degradation potential
Five Mortierella isolates were chosen for further studies (see table 1). The experiment was
conducted to test the potential of the different fungal strains for degradation of diuron. It was
carried out in 100ml screw-cap flasks with Teflon-lined lids. Initially 25µl of stock solution diuron
(5000ppm) dissolved in acetonitrile was added to flasks, acetonitrile was allowed to evaporate, and
25ml of glucose-mineral medium was added to each flask to give a final concentration of 5mg
diuron l-1
. Inoculation for each flask was done with three agar plugs (ø = 4mm) with mycelium cut
from PDA plates. The experiment was carried out in triplicates of each treatment, and with an
abiotic treatment to serve as control.
Fungal cultures were incubated on an orbital shaker at 110 rpm in the dark at 8oC. Repeated
aliquots sampling from flasks were done with 1ml sterile syringes, transferring samples to UPLC

47
vials through a syringe filter (0.2 µm PTFE membrane, 15mm). Samples were frozen (-18oC) until
UPLC analysis.
Diuron degradation by Mortierella sp. LEJ701 at different C and N concentrations
One fungal strain, Mortierella sp. LEJ 701, was used to investigate the effect of different carbon
and nitrogen concentrations on fungal diuron degradation. Experimental setup was done as above
with the following changes: for different glucose treatments three different media were used with 5
g, 0.5 g or 0 g glucose l-1
(only glucose available to the fungi is from the agar plug ≤ 0.045 g l-1
).
For all treatments (three glucose levels and with/without diuron) 12 flasks were prepared for
destructive sampling of triplicates at four pre-determined sampling times. At sampling aliquots
were taken for UPLC analysis as described above.
In addition, effects of different nitrogen concentrations were also studied. Again the setup had
six treatments (three nitrogen levels and with/without diuron). NH4Cl in the media was (l-1
): 1 g,
0.04 g or 0 g (only N available to the fungi is from the agar plug ≤ 0.001 g l-1
). Three additional
flasks were used where fungal mycelia were removed after 10 days to test if extracellular enzymes
would continue the diuron degradation in the media. In both experiments described above fungal
biomasses were collected on filters and dry-weights were measured (80oC for 20h).
UPLC analysis of diuron and metabolites
Ultra performance liquid chromatography (UPLC) was performed for quantitative analysis of
diuron and its metabolites in the culture media. The system used was an Acquity UPLC (Waters,
Milford, MA) equipped with a UV detector (200-225 nm absorbance) and an Acquity BEH C18
column (1.7 µm particle size, 2.1 mm inner diameter, and 100 mm length) set at a constant flow rate
of 0.4 ml min-1
, column temperature of 43oC, and an injection volume of 10µl. The initial mobile
phase was composed of 30% acetonitrile and 70% water with a 5 min gradient ending with 35 %
acetonitrile and 65% water. Hereafter conditions returned to the starting values giving a total
analysis time of 6 min per sample.
Retentions times for the compounds analyzed were: DCPU 2.45 min, DCPMU 3.33 min, 3,4-
DCAA 3.96 min, diuron 4.24 min, and 3,4-DCA 4.72 min. In addition, an unknown metabolite with
a retention time of 2.16 min was also detected.

48
Biosynthesis of the unidentified metabolite
Mortierella sp. LEJ701 was inoculated in flasks containing glucose-mineral medium and diuron (5
mg l-1
), as described above, for large scale production of the unknown metabolite for identification
by nuclear magnetic resonance (NMR) and mass spectroscopy (MS). Flasks were incubated on an
orbital shaker at 110 rpm in dark at 8oC for 20 days. Sampling was done with 1ml sterile syringes,
transferring samples to 2 ml vials through syringe filters (0.2 µm PTFE membrane, 15mm).
Samples were frozen (-18oC) until HPLC analysis.
Preparative HPLC
Reverse phase HPLC was used to purify the unknown metabolite, and the identity was determined
by NMR spectroscopy and MS. The system used was an ÄKTApurifier (GE Healthcare, Munich,
Germany) equipped with a UV detector (210-212 nm absorbance) and a Phenomenex Luna C18
column (5µm, 100Å; 4.6 mm x 100 mm). Preparative HPLC was performed using a gradient
program with a flow rate of 1.0 ml min-1
. The initial mobile phase was composed of 30%
acetonitrile and 70% water with a 30 min gradient ending with 50 % acetonitrile and 50% water.
The injection volume was 1.8 ml. Fractions of 0.5 ml were collected. The identity and elution
positions of the known compounds, diuron, DCPU, DCPMU and 3,4-DCA, were confirmed by
mass spectrometry.
NMR spectroscopy
Fractions containing the unknown metabolite were isolated, pooled, lyophilized and dissolved in
CD3CN (99.80 %D) to a volume of 600 µL and transferred to a 545 NMR tube. A parallel sample
of diuron was prepared from lyophilized powder and analyzed and a comparative sample of the
same volume of HPLC buffer at a matched % of acetonitrile was lyophilized and dissolved in
CD3CN (99.80 %D). For all samples 1D 1H NMR spectra were recorded on a Varian INOVA 750
MHz NMR spectrometer equipped with a 5 mm triple-resonance probe and Z-field gradient at 298
K utilizing 128 (diuron), 80.000 (solvent) or 80.000 (metabolite) transients. The spectra were

49
transformed and analyzed using MesTreNova (Mestrec Research, Spain) and referenced to internal
CD3CN at 1.94 ppm.
MS analyses
Two µl fractions from the preparative HPLC were diluted with 8 µl 0.5 % formic acid and analyzed
by LC-MS. The set-up consisted of an EASY nanoLC (Proxeon, Odense, Denmark) in front of a
micrOTOF Q II (Bruker, Bremen, Germany). HPLC conditions were: A 75 µm x 100 mm 3 µm
C18 column (Thermo Scientific, Copenhagen, Denmark) eluted at 300 nl/min with a 10 min
gradient from 10 – 50 % B (A: 0.5 % HCOOH in H2O, B: 0.5 HCOOH in CH3CN).
Statistics
Statistical analyses were done in SigmaPlot version 11.0 (Systat Software Inc). Results from
different treatments were tested for significance with ANOVA including Tukey Test for pairwise
multiple comparison procedures. Linear regression was performed to test the correlation between
amount of fungal biomass and diuron degradation. Differences are found statistically significant
when p ≤ 0.05. Data are presented as means ± standard error (SE) unless stated otherwise.
Strain deposition and nucleotide sequence accession numbers
The five Mortierella strains LEJ701-LEJ705 are deposited at the Centraalbureau voor
Schimmelcultures under the accession numbers CBS 133175-CBS 133179 in the CBS Collection of
Fungi. In addition, the sequences determined in this study have been deposited in the Genbank
database under accession numbers JX206802-JX206806.

50
Results
Phylogeny of fungal pesticide degraders
The five strains showed very different potential for degradation of diuron (Table 1). The most
efficient diuron degrader Mortierella sp. LEJ701 was able to degrade all diuron in the medium and
the two closely related Mortierella sp. isolates LEJ702 and LEJ704 showed some potential for
diuron degradation removing 33.6% and 9.2% of the diuron, respectively. The two strains that did
not degrade diuron were found to be only distantly related to the diuron degraders (Fig. 1). The
most efficient diuron degrader Mortierella sp. LEJ701 had a sequence similarity of 97% and 96%
with the two other degraders Mortierella sp. LEJ702 and LEJ704, respectively. Whereas
Mortierella sp. LEJ701 only had a sequence similarity of 92% and 91% with the non-degraders
Mortierella sp. LEJ703 and LEJ705, respectively.
Substrate effects on diuron degradation
Mortirella sp. LEJ701 clearly responded to the level of glucose in the medium. Diuron degradation
was faster and more complete in the medium with a high glucose concentration compared to the
degradation in media with intermediate and low glucose concentrations (Fig. 2). The diuron
degradation rate in the low glucose medium was very slow, whereas the degradation rates were
significantly higher for intermediate and high glucose concentration media (p < 0.001) (see Table
3). No significant difference between degradation rates was found for intermediate and high glucose
media (p = 0.154).
Fungal biomass differed significantly between the three glucose treatments at experimental
termination (Table 2, ANOVA Tukey test; all p < 0.001). Diuron degradation was positively
correlated with fungal biomass (p < 0.001, r2
= 0.683; Fig. 3), and the degradation rates per unit of
biomass differed significantly for the three treatments (i.e. carbon concentration) (Table 3).
The results showed no effect of diuron on fungal biomass quantity at experimental
termination (p = 0.607; Table 2), although an alteration in pellet structure was observed. The fungal
pellets in treatments with diuron had long hyphal outgrowths from the otherwise spherical pellets.
The effect of nitrogen on diuron degradation was very similar to the effects observed with
glucose concentrations. Diuron degradation was slow in the treatment with low nitrogen
concentration and significantly higher in the treatments with intermediate nitrogen concentration (p

51
< 0.001). Finally the treatment with high nitrogen concentration showed the fastest degradation rate
(Table 3; p < 0.001).
There was no degradation in the abiotic controls. Furthermore no further degradation was
seen by possible exudates after removal of fungal biomass (data not shown).
Accumulation and identification of diuron metabolites
Diuron degradation and accumulation of the two metabolites DCPMU and DCPU corresponded
with the patterns shown in previous studies (Badawi et al. 2009). The first metabolite DCPMU
accumulated as the diuron concentration decreased and after approximately ten days the second
metabolite DCPU appeared (Fig. 2a). In addition, to these compounds, our results showed an
unknown metabolite with a retention time of 2.16 min observed by UPLC (Fig. 4). When this
metabolite was included we achieved complete recovery of the diuron initially added to the system
(Fig. 2a & b).
To identify the unknown metabolite it was produced in large-scale and analyzed by NMR
spectroscopy and MS analyses. From the analytical MS analyses it was directly apparent that the
unknown metabolite-fraction from the preparative fractionation contained two different species of
molecular mass (m.m.) 204.004 Da and 215.996 Da, (monoisotopc values), corresponding to DCPU
(theoretical m.m. = 203.986 Da) and the unknown metabolite as well as trace amounts of intact
diuron (m.m. = 232.019 Da compared to the theoretical 232.017 Da). Hence any quantitative NMR
analysis would be meaningless. Despite the very low concentration, and hence low signal-to-noise
in the NMR spectrum, it was however immediately apparent that compared to the NMR spectrum of
diuron, several sets of peaks originating from the aromatic ring suggested this to be intact also in the
metabolite with both chloride atoms as well as the amide proton present. The two chloride atoms
were corroborated from the 35
Cl-37
Cl isotope ‖signature‖ in the MS spectra. In addition, from the
molecular mass and isotopic pattern using the software application SmartFormula (Bruker
Daltonics) the composition of the unknown metabolite was suggested to be C8H6ON2 (theoretical
m.m. = 215.986 Da) with high confidence. None of the methyl groups were present in the NMR
spectrum as concluded from lack of these signals in the aliphatic region, suggesting that the fraction
of diuron in the NMR sample was below detection. A small but significant change in the chemical
shifts for all the aromatic protons ortho-positioned relative to the amide group suggested a change
in the chemistry of this side group compared to diuron, although the structure was not elucidated by

52
NMR due to the sparse amount of sample. The shorter retention time on the UPLC excluded the
metabolite as either DCPMU or DCPU, so based on the NMR data, the molecular mass, the
suggested atomic composition of the metabolite, as well as the lack of methyl groups, we suggest
the metabolite to be 1-(3,4-dichlorophenyl)-3-methylideneurea (DCPMDU). Figure 5 depicts the
proposed degradation pathway of diuron, including the most likely position of 1-(3,4-
dichlorophenyl)-3-methylideneurea based the degradation kinetics (Fig. 2). The molecular masses
were calculated as m/z-values – 1.0078 (H+).
Discussion
Our results show that the ability to degrade diuron among the Mortierella strains was linked to their
phylogenetic position. This is in agreement with a phylogenetic analysis by Hussain et al. (2009)
who demonstrated that bacterial strains degrading isoproturon were closely related. The most
efficient degrader Mortierella sp. LEJ701 and the two other degraders Mortierella sp. LEJ702 and
LEJ704 showed 96-97% genetic similarity, suggesting that though they are closely related they may
belong to separate species. Nevertheless these closely related strains share a particular trait enabling
them to degrade diuron by demethylation. Vroumsia et al. (1996) found, among the 90 fungal
strains they screened for degradation potential, that four Mortierella isolates had degraded 19 - 54%
diuron after 7 days incubation at 24oC. These values are within the same range as in the present
study, though we achieved this at a much lower temperature – one comparable to average yearly
temperature in a temperate climate. Furthermore, their work was not supported by molecular data
on the fungal phylogeny, and it is therefore not possible to evaluate the phylogenetic relationship
between the isolates.
The intra- and inter-specific differences in ability to degrade pesticides clearly demonstrate
the need for further studies to clarify the genetic background for this characteristic. Future
functional genetic studies should clarify these differences and may facilitate more focused isolation
strategies for potential pesticide degraders. For biotechnological purposes this will be important as
it may be possible to isolate strains with even higher potential for bioremediation under natural
conditions.
Our results show that it is unlikely that the fungus utilizes diuron as a carbon and/or nitrogen
source, since growth was not stimulated by diuron addition even at the lowest levels of glucose and
NH4Cl where diuron C and N would otherwise contribute significantly to biomass quantity. On the

53
contrary our results showed that Mortierella sp. LEJ 701 required an additional carbon and nitrogen
source to be able to degrade diuron. Likewise, Rønhede et al. (2005) found that isoproturon did not
serve as an energy or nutrient source for the fungi used in their experiment. In contrast, Kulshrestha
and Kumari (2011) found that an Acremonium strain could utilize the organophosphate chlorpyrifos
as a source of both carbon and nitrogen. Since the Mortierella strain cannot utilize diuron as a N or
C source it seems plausible, that the degradation process is co-metabolic mediated by enzymes
excreted by the fungus during growth leading to successive demethylation through the metabolite
DCPMDU.
We did not detect a decrease in fungal biomass quantity in treatments with diuron (Table 2).
This is in contrast to Vroumsia et al. (1996) who reported a minor growth inhibition of Rhizoctonia
solani exposed to a four times higher diuron concentration. However, we did observe an altered
morphology of the fungal pellets during the experiment. The growth response characterized by long
hyphal outgrowths from the pellet is often a response to stress e.g. toxic or lack of nutrients
(Fomina et al. 2003). Toxicity of diuron or its metabolites towards the fungus can therefore not be
excluded, and the co-metabolic degradation process could be a detoxification mechanism.
Ecotoxicity studies by Tixier et al. (2000) have shown that the metabolites DCPU and
DCPMU are three times more toxic than diuron. It is consequently of concern that the degradation
of diuron by common soil fungi leads to accumulation of these compounds (see Fig. 2). We have no
information on the toxicity of the new metabolite since only small amounts could be synthesized,
but seen in a regulatory context all these metabolites should be included in monitoring programs as
well as in approval of pesticides on the basis of their toxicity and potential accumulation in the
environment.
In conclusion, we have shed light on the phylogenetic relationship within the Mortierella
genus in respect to diuron degradation potential. Also we have shown that diuron degradation by
Mortierella is a co-metabolic process resulting in accumulation of its metabolites. Further research
should include degradation of these metabolites. We suggest a possible way of achieving fast diuron
degradation beyond these intermediate metabolites could be by developing consortia incorporating
both fungal and bacterial strains.
Acknowledgments The authors thank Nora Badawi and Spire Kiersgaard for guidance on the UPLC method and Signe
Sjørup and Allan Kastrup for expert technical assistance. The work was supported by the MIRESOWA project funded
by the Danish Council for Strategic Research (grant number 2104-08-0012).

54
References
Badawi N, Ronhede S, Olsson S, Kragelund BB, Johnsen AH, Jacobsen OS, Aamand J (2009) Metabolites of the
phenylurea herbicides chlorotoluron, diuron, isoproturon and linuron produced by the soil fungus Mortierella
sp. Environ Pollut 157 (10):2806-2812. doi:10.1016/j.envpol.2009.04.019
El-Bestawy E, Albrechtsen H-J (2007) Effect of nutrient amendments and sterilization on mineralization and/or
biodegradation of 14C-labeled MCPP by soil bacteria under aerobic conditions. Int Biodeterior Biodegrad 59
(3):193-201
El-Deeb B, Soltan S, Ali A, Ali K (2000) Detoxication of the herbicide diuron by Pseudomonas sp. Folia Microbiol 45
(3):211-216. doi:10.1007/bf02908946
Entry JA, Donnelly PK, Emmingham WH (1996) Mineralization of atrazine and 2,4-D in soils inoculated with
Phanerochaete chrysosporium and Trappea darkeri. Appl Soil Ecol 3 (1):85-90
Eriksson E, Baun A, Mikkelsen PS, Ledin A (2007) Risk assessment of xenobiotics in stormwater discharged to
Harrestrup Å, Denmark. Desalination 215 (1-3):187-197
European Parliament EU (2008) Priority Substances Directive (Directive 2008/105/EC). Official Journal of the
European Union.
Fomina M, Ritz K, Gadd GM (2003) Nutritional influence on the ability of fungal mycelia to penetrate toxic metal-
containing domains. Mycol Res 107 (7):861-871
Gardes M, Bruns TD (1993) ITS primers with enhanced specificity for basidiomycetes - application to the identification
of mycorrhizae and rusts. Mol Ecol 2 (2):113-118. doi:10.1111/j.1365-294X.1993.tb00005.x
Gardes M, Bruns TD (1996) ITS-RFLP matching for identification of fungi. In: Clapp JP (ed) Methodes in molecular
biologi: Species diagnostics protocols - PCR and other nucleic acid methods, vol 50. Humana Press Inc.,
Totowa, New Jersey, pp 177-186
Grube A, Donaldson D, Kiely T, Wu L (2011) Pesticides Industry Sales and Usage. U.S. Environmental Protection
Agency,
Harms H, Schlosser D, Wick LY (2011) Untapped potential: exploiting fungi in bioremediation of hazardous chemicals.
Nat Rev Microbiol 9 (3):177-192. doi:10.1038/nrmicro2519
Hussain S, Sørensen SR, Devers-Lamrani M, El-Sebai T, Martin-Laurent F (2009) Characterization of an isoproturon
mineralizing bacterial culture enriched from a French agricultural soil. Chemosphere 77 (8):1052-1059
Khadrani A, Seigle-Murandi F, Steiman R, Vroumsia T (1999) Degradation of three phenylurea herbicides
(chlortoluron, isoproturon and diuron) by micromycetes isolated from soil. Chemosphere 38 (13):3041-3050
Kjøller R, Bruns TD (2003) Rhizopogon spore bank communities within and among California pine forests. Mycologia
95 (4):603-613

55
Kulshrestha G, Kumari A (2011) Fungal degradation of chlorpyrifos by Acremonium sp. strain (GFRC-1) isolated from
a laboratory-enriched red agricultural soil. Biol Fertil Soils 47 (2):219-225. doi:10.1007/s00374-010-0505-5
Lapworth DJ, Gooddy DC (2006) Source and persistence of pesticides in a semi-confined chalk aquifer of southeast
England. Environ Pollut 144 (3):1031-1044
Ritz K, Young IM (2004) Interactions between soil structure and fungi. Mycologist 18 (2):52-59
Rønhede S, Jensen B, Rosendahl S, Kragelund BB, Juhler RK, Aamand J (2005) Hydroxylation of the herbicide
isoproturon by fungi isolated from agricultural soil. Appl Environ Microbiol 71 (12):7927-7932.
doi:10.1128/aem.71.12.7927-7932.2005
Simonsen A, Holtze MS, Sørensen SR, Sørensen SJ, Aamand J (2006) Mineralisation of 2,6-dichlorobenzamide (BAM)
in dichlobenil-exposed soils and isolation of a BAM-mineralising Aminobacter sp. Environ Pollut 144 (1):289-
295
Sørensen SR, Albers CN, Aamand J (2008) Rapid mineralization of the phenylurea herbicide diuron by Variovorax sp.
SRS16 in pure culture and within a two-member consortium. Appl Environ Microbiol 74:2332-2340.
doi:10.1128/aem.02687-07
Struger J, Grabuski J, Cagampan S, Rondeau M, Sverko E, Marvin C (2011) Occurrence and distribution of
sulfonylurea and related herbicides in central Canadian surface waters 2006-2008. Bull Environ Contam
Toxicol 87 (4):420-425
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S (2011) MEGA5: Molecular evolutionary genetics
analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol
28 (10):2731-2739. doi:10.1093/molbev/msr121
Tixier C, Bogaerts P, Sancelme M, Bonnemoy F, Twagilimana L, Cuer A, Bohatier J, Veschambre H (2000) Fungal
biodegradation of a phenylurea herbicide, diuron : structure and toxicity of metabolites. Pest Manag Sci 56
(5):455-462
Tixier C, Sancelme M, Bonnemoy F, Cuer A, Veschambre H (2001) Degradation products of a phenylurea herbicide,
diuron: Synthesis, ecotoxicity, and biotransformation. Environ Toxicol Chem 20 (7):1381-1389
Torstensson L (2001) Use of herbicides on railway tracks in Sweden. Pestic Outlook 12 (1):16-21
Vroumsia T, Steiman R, SeigleMurandi F, BenoitGuyod JL, Khadrani A (1996) Biodegradation of three substituted
phenylurea herbicides (chlorotoluron, diuron, and isoproturon) by soil fungi. A comparative study.
Chemosphere 33 (10):2045-2056
White TJ, Bruns TD, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal ribosomal RNA genes for
phylogenetics. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ (eds) PCR protocols: A guide to methodes and
applications. Academic Press Inc., San Diego, California, pp 315-322
Wösten HAB, van Wetter MA, Lugones LG, van der Mei HC, Busscher HJ, Wessels JGH (1999) How a fungus escapes
the water to grow into the air. Curr Biol 9 (2):85-88
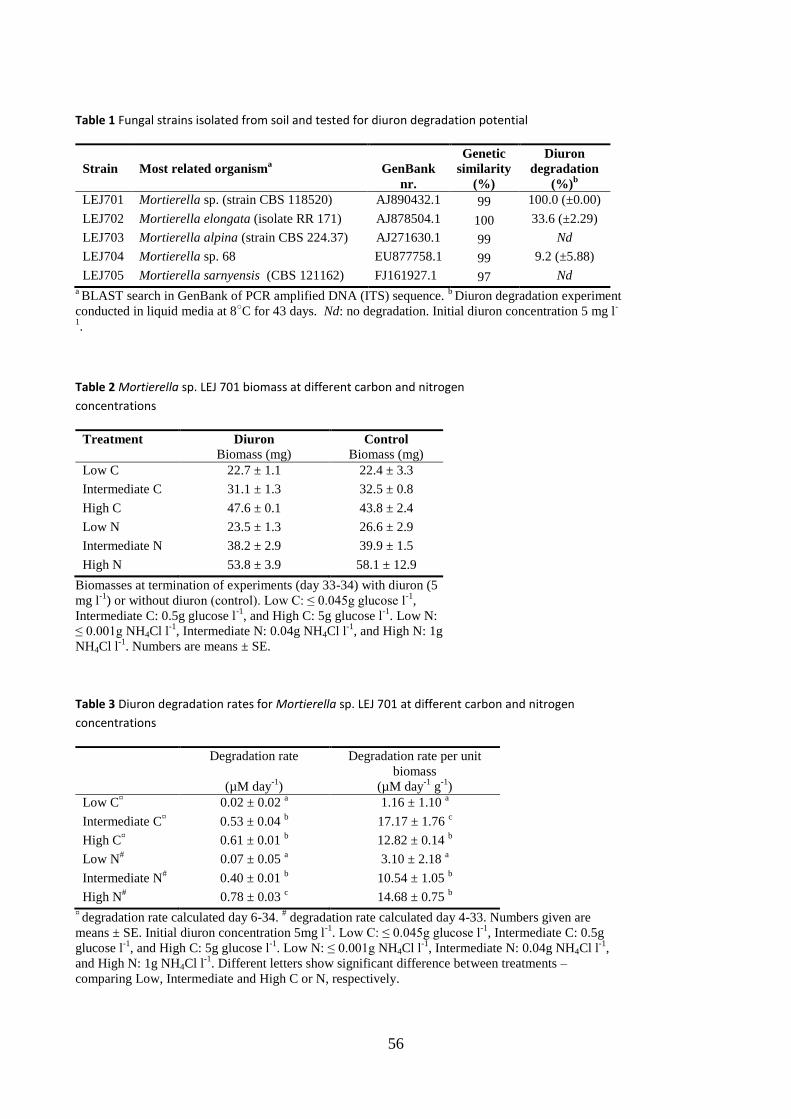
56
Table 1 Fungal strains isolated from soil and tested for diuron degradation potential
Strain
Most related organisma
GenBank
nr.
Genetic
similarity
(%)
Diuron
degradation
(%)b
LEJ701 Mortierella sp. (strain CBS 118520) AJ890432.1 99 100.0 (±0.00)
LEJ702 Mortierella elongata (isolate RR 171) AJ878504.1 100 33.6 (±2.29)
LEJ703 Mortierella alpina (strain CBS 224.37) AJ271630.1 99 Nd
LEJ704 Mortierella sp. 68 EU877758.1 99 9.2 (±5.88)
LEJ705 Mortierella sarnyensis (CBS 121162) FJ161927.1 97 Nd a BLAST search in GenBank of PCR amplified DNA (ITS) sequence.
b Diuron degradation experiment
conducted in liquid media at 8○C for 43 days. Nd: no degradation. Initial diuron concentration 5 mg l
-
1.
Table 2 Mortierella sp. LEJ 701 biomass at different carbon and nitrogen
concentrations
Treatment
Diuron
Biomass (mg) Control
Biomass (mg)
Low C
22.7 ± 1.1 22.4 ± 3.3
Intermediate C
31.1 ± 1.3 32.5 ± 0.8
High C
47.6 ± 0.1 43.8 ± 2.4
Low N
23.5 ± 1.3 26.6 ± 2.9
Intermediate N
38.2 ± 2.9 39.9 ± 1.5
High N
53.8 ± 3.9 58.1 ± 12.9
Biomasses at termination of experiments (day 33-34) with diuron (5
mg l-1
) or without diuron (control). Low C: ≤ 0.045g glucose l-1
,
Intermediate C: 0.5g glucose l-1
, and High C: 5g glucose l-1
. Low N:
≤ 0.001g NH4Cl l-1
, Intermediate N: 0.04g NH4Cl l-1
, and High N: 1g
NH4Cl l-1
. Numbers are means ± SE.
Table 3 Diuron degradation rates for Mortierella sp. LEJ 701 at different carbon and nitrogen
concentrations
Degradation rate
(µM day-1
)
Degradation rate per unit
biomass
(µM day-1
g-1
)
Low C¤
0.02 ± 0.02 a
1.16 ± 1.10 a
Intermediate C¤
0.53 ± 0.04 b
17.17 ± 1.76 c
High C¤
0.61 ± 0.01 b
12.82 ± 0.14 b
Low N#
0.07 ± 0.05 a
3.10 ± 2.18 a
Intermediate N#
0.40 ± 0.01 b
10.54 ± 1.05 b
High N#
0.78 ± 0.03 c
14.68 ± 0.75 b
¤ degradation rate calculated day 6-34.
# degradation rate calculated day 4-33. Numbers given are
means ± SE. Initial diuron concentration 5mg l-1
. Low C: ≤ 0.045g glucose l-1
, Intermediate C: 0.5g
glucose l-1
, and High C: 5g glucose l-1
. Low N: ≤ 0.001g NH4Cl l-1
, Intermediate N: 0.04g NH4Cl l-1
,
and High N: 1g NH4Cl l-1
. Different letters show significant difference between treatments –
comparing Low, Intermediate and High C or N, respectively.
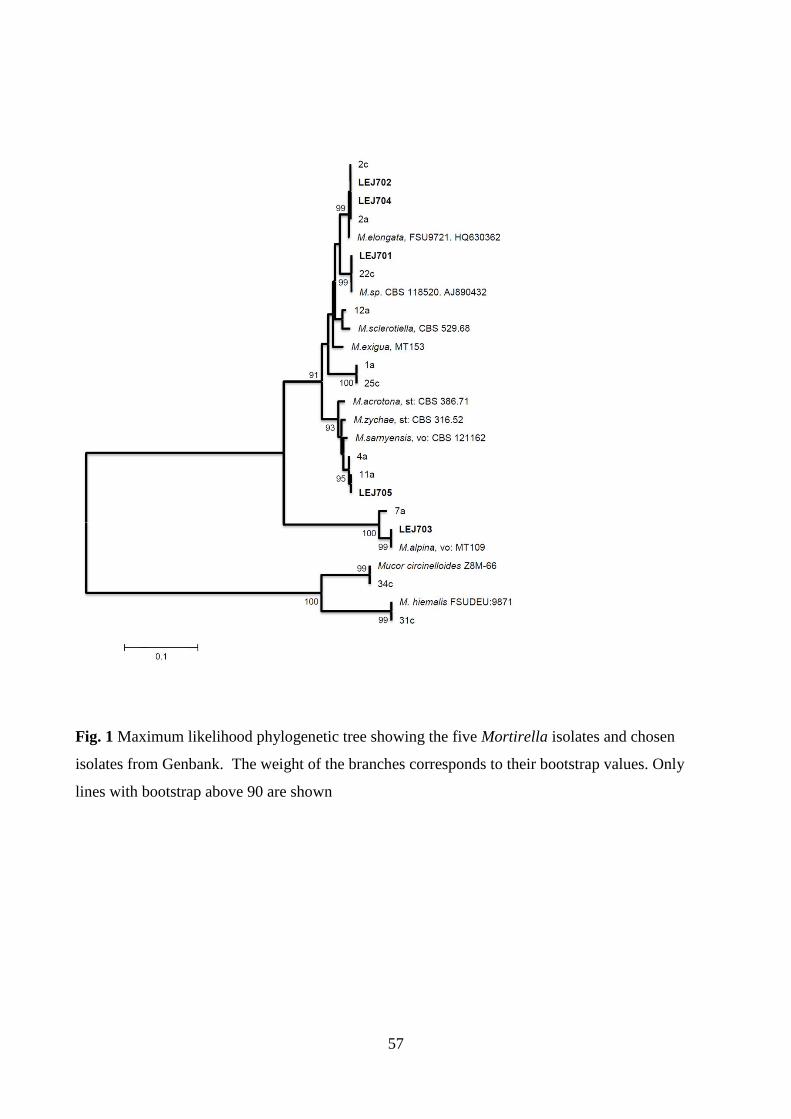
57
Fig. 1 Maximum likelihood phylogenetic tree showing the five Mortirella isolates and chosen
isolates from Genbank. The weight of the branches corresponds to their bootstrap values. Only
lines with bootstrap above 90 are shown

58
(a)
Days
0 10 20 30
µm
ol l-
1
0
5
10
15
20
Are
a
0
10000
20000
30000
40000
(b)
Days
0 10 20 30
µm
ol l-
1
0
5
10
15
20A
rea
0
10000
20000
30000
40000
(c)
Days
0 10 20 30
µm
ol l-1
0
5
10
15
20
Are
a
0
10000
20000
30000
40000
Diuron
DCPMU
DCPU
Unknown
Total
Fig. 2 Diuron degradation kinetics by Mortierella sp. LEJ 701 at (a) high, (b) intermediate and (c)
low glucose concentrations. Diuron: [N-(3,4-dichlorophenyl)-N,N-dimethyl-urea], DCPMU: [1-
(3,4-dichlorophenyl)-3-methylurea], DCPU: [1-(3,4-dichlorophenyl) urea], and unknown metabolite
(RT 2.16min)
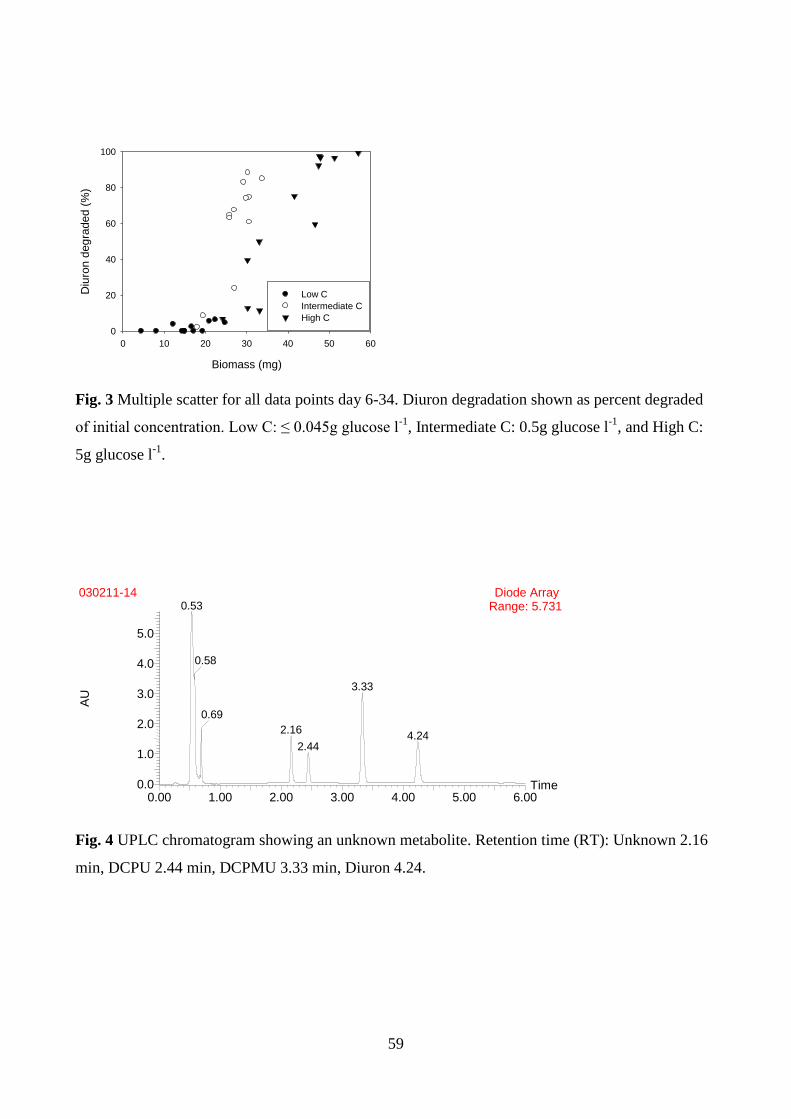
59
Biomass (mg)
0 10 20 30 40 50 60
Diu
ron d
eg
rade
d (
%)
0
20
40
60
80
100
Low C
Intermediate C
High C
Fig. 3 Multiple scatter for all data points day 6-34. Diuron degradation shown as percent degraded
of initial concentration. Low C: ≤ 0.045g glucose l-1
, Intermediate C: 0.5g glucose l-1
, and High C:
5g glucose l-1
.
Fig. 4 UPLC chromatogram showing an unknown metabolite. Retention time (RT): Unknown 2.16
min, DCPU 2.44 min, DCPMU 3.33 min, Diuron 4.24.
51
Time0.00 1.00 2.00 3.00 4.00 5.00 6.00
AU
0.0
1.0
2.0
3.0
4.0
5.0
030211-14 Diode Array Range: 5.7310.53
0.58
3.33
0.69
2.16
2.444.24

60
Fig. 5 Proposed degradation pathway of diuron. Diuron: [N-(3,4-dichlorophenyl)-N,N-dimethyl-
urea], DCPMU: [1-(3,4-dichlorophenyl)-3-methylurea], DCPU: [1-(3,4-dichlorophenyl) urea], 3,4-
DCA: [3,4-dichloroaniline] and DCPMDU: [1-(3,4-dichlorophenyl)-3-methylideneurea]. 3,4- DCA
was only found as trace amounts by MS. The structure and placement of DCPMDU, in brackets, are
suggested based on the composition C8H6ON2 and the degradation kinetics.

Manuscript II
TRANSPORT OF DEGRADER BACTERIA VIA FUNGAL HYPHAE INCREASES
DEGRADATION OF 2,6-DICHLOROBENZAMIDE
Berith E. Knudsen, Lea Ellegaard-Jensen, Christian N. Albers, Søren Rosendahl and Jens Aamand
Submitted to Environmental Pollution


63
Transport of degrader bacteria via fungal hyphae increases degradation of 2,6-
dichlorobenzamide
Berith Elkær Knudsena,b,§,*
, Lea Ellegaard-Jensena,b,§
, Christian Nyrop Albersa, Søren Rosendahl
b, and Jens Aamand
a
a) Department of Geochemistry, Geological Survey of Denmark and Greenland (GEUS), Øster Voldgade 10, DK-1350
Copenhagen K, Denmark. b) Department of Biology, Copenhagen University, Universitetsparken 15, DK-2100
Copenhagen Ø, Denmark.
§) Joint first author
*) Corresponding author: [email protected], Phone: +45 38142316, Fax: +45 38142050
Abstract
Introduction of specific degrading microorganisms into polluted soil or aquifers is a promising
remediation technology provided that the organisms survive and spread in the environment. We
suggest that consortia, rather than single strains, may be better suited to overcome these challenges.
Here we introduced a fungal-bacterial consortium consisting of Mortierella sp. LEJ702 and the
2,6-dichlorobenzamide (BAM)-degrading Aminobacter sp. MSH1 into small sand columns. A more
rapid mineralisation of BAM was obtained by the consortium compared to MSH1 alone especially
at lower moisture contents. Results from quantitative real-time polymerase chain reaction (qPCR)
demonstrated better spreading of Aminobacter when Mortierella was present suggesting that fungal
hyphae are transport routes for the bacteria. Extraction and analysis of BAM showed that also the
compound was transported by the fungal hyphae in the sand. This suggests that fungal-bacterial
consortia are promising for successful bioremediation of pesticide contamination.
Key words: 2,6-dichlorobenzamide (BAM), consortium, fungal highway, pesticide biodegradation,
fungal-bacterial interactions.
Capsule: This study brings new knowledge to the benefits of applying bacterial-fungal consortia for
bioremediation.

64
1. Introduction
Extensive use of pesticides on agricultural and urban areas poses a threat to groundwater
resources. This is of great concern, especially in countries where the majority of the drinking water
originates from groundwater. The most frequently detected pollutant in Danish groundwater is 2,6-
dichlorobenzamide (BAM), a degradation product of the herbicide dichlobenil, the use of which has
been banned in Denmark since 1997 (Thorling et al., 2010). Several water supply wells have been
closed due to detection of BAM at concentrations exceeding the threshold value of 0.1 µg l-1
. The
closing of wells, however, does not solve the problem, as the pollutants still remain in the water
resources.
Bioremediation, especially bioaugmentation, has been suggested as a possible means of
cleaning pesticide-polluted sites (Thompson et al., 2005). Various bacterial strains capable of
degrading different pesticides have been identified, e.g. the BAM-degrading Aminobacter sp. strain
MSH1 (Sørensen et al., 2007). There are, however, still major challenges that need to be overcome
before bioaugmentation can be efficient (El Fantroussi & Agathos, 2005; Thompson et al., 2005).
These challenges include: i) securing the survival of the introduced strains, ii) development of
efficient tools for spreading degrader organisms in the polluted environment and iii) providing
access for the microorganisms to the pollutants sorbed to the sediment and organic matter or
trapped in micropores, e.g. small cracks or voids on the particle surfaces. It is difficult to ensure
survival of the introduced strains (van Veen et al., 1997; Pepper et al., 2002), since various factors
like temperature, moisture content and nutrient availability in the new environment may differ from
their original habitat (Vogel, 1996). Furthermore, studies have shown that bacteria applied to soil
surfaces rarely are transported more than 5 cm into the soil without the help of a transport agent,
such as percolating water (Edmonds, 1976; Madsen and Alexander, 1982). Even if the strains are
successfully added to the environment, the pollutants are often only present at low concentrations
and are not easily accessible for the degrader bacteria (Harms and Bosma, 1997; Wick et al., 2007).
It has been suggested that fungal hyphae may function as a transport vector for bacteria capable of
degrading hydrophobic PAHs (Furuno et al., 2010). Also, many bacteria can be found in the
hyphosphere; thus they have better access to nutrients from fungal exudates, ensuring their survival
in the environment (Boersma et al., 2010; de Boer et al., 2005). Therefore, adding fungal-bacterial
consortia instead of single bacterial strains may be a way to overcome these challenges. Previous
studies by Kohlmeier et al. (2005) and Wick et al. (2007) have moreover shown that fungal hyphae
may facilitate transport of PAH-degrading bacteria in soil. The water film produced by some fungi

65
around their hyphae allows a better transport of the bacteria present in the hyphosphere. Motile
bacteria can utilise the water film as a transport vector enabling transport along the hyphae − a
transport mechanism termed the fungal highway (Kohlmeier et al., 2005). Thus fungal growth may
enable the bacteria to access areas and thereby potentially access pesticides they otherwise would
not be able to reach. The water film may also facilitate transport of the pollutant. However, the
above mentioned studies have focused on hydrophobic compounds like PAHs and it has therefore
not been examined whether bacteria degrading hydrophilic compounds like pesticides are
transported in the same way. The transport of hydrophilic compounds along the fungal hyphae will
also potentially differ from the transport of the hydrophobic PAHs.
In the present study we used the common soil fungus Mortierella and the BAM-degrading
bacterium Aminobacter sp. MSH1. Mortierella is known to produce hydrophilic mycelia (Chau et
al., 2010), a feature which has been shown to facilitate transport of motile bacteria (Kohlmeier et
al., 2005). The aim of this study was to examine whether the presence of Mortierella sp. LEJ702
affected BAM degradation and mineralization by the motile Aminobacter sp. MSH1 in sand
columns. Moreover, it is investigated whether BAM would be transported along the hyphae of
Mortierella. Our hypothesis was that presence of fungal hyphae facilitates transport of both bacteria
and BAM, thereby increasing the accessibility of BAM to the degrader bacteria, leading to an
increased degradation.
2. Materials and Methods
2.1. Chemical and media.
[Ring-U-14
C]- 2,6-dichlorobenzamide (BAM) (25.2 mCi mmol-1
) was purchased from Izotop,
(Institute of Isotopes Co., Ltd., Hungary). At the time of use, the standard had a radiochemical
purity >99% as determined by Thin Layer Chromatography (TLC). A stock solution of radiolabeled
BAM (3,000,000 DPM ml-1
) was prepared in acetonitrile.
Potato Dextrose Agar (PDA) (Becton, Dickinson and Company, Franklin Lakes, NJ, USA) was
used for cultivation of Mortierella sp. LEJ702. Minimal medium (MSNC) containing 0.2 g L-1
Na-
succinate (CAS 6106-21-4), 0.476 g L-1
(NH4)2SO4, 0.1 g L-1
KNO3 and 0.08% glycerol (Sørensen
and Aamand 2003) was used for growing Aminobacter sp. MSH1.

66
The liquid medium used for the experimental setup was buffered MilliQ water. The buffer
contained 136 g L-1
KH2PO4 and 178 g L-1
Na2HPO4.2H2O. The buffer was mixed 1:99 with MilliQ
water, giving a final phosphate concentration in the medium of 6 mM (pH 7).
Agar and media were sterilized by autoclaving at 121oC for 30 minutes.
2.2. Strains/organisms
The BAM-degrading bacterial strain used in the experiments was the motile Aminobacter sp.
MSH1 isolated and described by Sørensen et al. (2007). The strain was pre-grown from a -80oC
stock culture in MSNC medium at 20oC on an orbital shaker (125 rpm). The cells were harvested in
the exponential growth phase, determined by OD600nm measurements. The cells were harvested by
centrifugation at 8000 x g for 10 minutes and washed twice in buffered MilliQ water.
The fungus Mortierella sp. LEJ702 was isolated by Ellegaard-Jensen et al. (in preparation). It
was grown and maintained at 8oC on PDA plates.
2.3. Sand
The sand used was oven-dried quartz sand purchased from Saint Gobain Weber A/S (Optiroc,
Randers, Denmark) with grain sizes between 0.3 and 1 mm. However, before experimental use it
was sieved to remove particles between 0.3 and 0.6 mm. Thus, the sand used had a grain size
between 0.6 and 1 mm giving a less densely packed matrix in the experimental setup. The sand was
sterilized by autoclaving at 121oC for 30 minutes. The water holding capacity (WHC) was
determined (28.7%) according to Danish Standards; DS/ISO 14238-1.
2.4. Mineralization experiment
Mineralization experiments were set up in glass vials (24 mm Ø, 95 mm h) containing 500 μl
PDA agar. Then either Aminobacter sp. MSH1, Mortierella sp. LEJ702, or the fungal-bacterial
consortium was added to the agar. Mortierella was added as an agar plug (Ø = 4mm) and the
bacteria were added to reach 105 cells g
-1 sand. Vials without microorganisms served as abiotic
controls. The sand was weighed into portions of 10 g and added [ring-U-14
C]-BAM dissolved in
acetonitrile, giving a concentration of 100 μg kg-1
. The sand was left for a few minutes in order for
the acetonitrile to evaporate before adding buffered MilliQ water. The amount of water added in the
treatments corresponded to 10%, 5%, 1.7% and 0% of WHC, respectively. A small glass tube
containing 1 ml 1M NaOH was added to each vial to trap evolved 14
CO2. The experiment was

67
incubated at 20oC. The NaOH was replaced approximately once a week and mixed (1:5) with
Optiphase ‗hisafe‘ 3 scintillation liquid (PerkinElmer Inc., Waltham, Massachusetts, USA). The
amount of radioactivity in the alkaline solution was measured on a Wallac 1409 DSA Liquid
scintillation Counter (PerkinElmer Inc., Waltham, Massachusetts, USA). All experiments were
carried out in triplicate. At termination of the experiments, the columns were analysed for BAM
residues using TLC (see below). To follow the removal of BAM and accumulation of degradation
products a parallel series of vials was set up with the same inocula, but at a WHC of 1.7% only.
Vials were harvested once a week for TLC analysis. The vials were harvested by removing the
CO2-trap and adding 6 ml MeOH-MilliQ water (50:50). The vials were then shaken vigorously for
two hours, followed by centrifugation for 10 min at 200 x g. From the supernatant, 2 ml was
transferred to Eppendorf tubes and centrifuged for 2 min at 13000 x g. The supernatant was used for
scintillation counting (500 μl) and for TLC analysis (40 μl).
2.5. DNA extraction and qPCR
At termination of the mineralization experiment, DNA was extracted from the top of the sand
layer (0.25 g) to investigate whether there had been transport of Aminobacter cells up through the
sand. DNA extractions were performed using the PowerLyzer PowerSoil DNA isolation kit (Mobio
Laboratories Inc., Carlsbad, CA, USA). Aminobacter specific primers described by Sjøholm et al.
(2010) were used for quantitative real-time polymerase chain reaction (qPCR). The amplifications
were performed in volumes of 20 μl, containing 2 μl BSA, 10 μl SiBir master mix (Bioron GmbH,
Ludwigshafen, Germany) and 0.4 μM of each primer as described by Sjøholm et al. (2010). The
conditions for the reactions were as follows: an initial denaturation step at 95 oC for 5 min. fifty
cycles at 95oC for 30 s, 53
oC for 30 s and 72
oC for 30 s, followed by an elongation step at 72
oC for
6 min and 71 cycles at 60oC for 30 s with a 0.5
oC increase per cycle. Final step was at 72
oC for 45 s.
Results of the qPCR were processed using the Bio-Rad iQ5 program (www.discover.bio-rad.com).
2.6. Transport of BAM
An experiment was set up to study whether Mortierella sp. LEJ702 facilitated the transport of
BAM through the sand. The experiment was set up in 20 ml sterile plastic syringes (Terumo
Corporation, Belgium). The pistons were set at the 20 ml marking, 500 μl PDA and a Mortierella
sp. LEJ702 plug (Ø = 4 mm) were added to the syringes. Five grams quartz sand was mixed with
buffered MilliQ-water and [Ring-U-14
C]-BAM giving a concentration of 100 μg kg-1
as described

68
above and added to the syringes followed by 5 g of unspiked quartz sand mixed with buffered
MilliQ-water. The experiment was carried out at the same moisture conditions as in the
mineralization experiment. The syringes were sealed with rubber plugs and wrapped in parafilm and
incubated at 20oC. Once a week the syringes were opened for aeration. At termination of the
experiment the sand was pushed to the top of the syringe using the piston. The unspiked sand was
divided into two equal size fractions, while the 14
C-spiked portion was kept as one portion.
Radiolabeled carbon was extracted from the top fraction of the unspiked sand as described above
with the modification that 3 ml MeOH:MilliQ-water was used for the extraction. After
centrifugation, 500 μl of the supernatant was mixed with 3 ml Optiphase ‗hisafe‘ 3 scintillation
liquid (PerkinElmer Inc., Waltham, Massachusetts, USA) and measured on a Liquid Scintillation
Analyzer, Tri-Carb 2810 TR (PerkinElmer Inc., Waltham, Massachusetts, USA). Subsamples of the
remaining extract were used for TLC analysis.
2.7. Thin Layer Chromatography
A total of 40 µL of each sample was spotted onto a 10x20 cm silica gel 60 normal phase TLC
plate (Merck, Darmstadt, Germany) and eluted with CH3CN:H2O:CH2Cl2;HCOOH (70:20:5:5) in a
CAMAG Horizontal Developing Chamber (CAMAG, Berlin, Germany). After air-drying, the plates
were developed on phosphor imaging screens and analyzed autoradiographically on a Cyclone
Scanner (Packard Instrument Company, Meriden, CT). The chromatograms obtained were analyzed
using the OptiQuant Image Analysis software (Packard Instrument Company) to integrate the
different bands. Rf-values were: BAM (0.95), 2,6-dichlorobenzoic acid (0.88) and unidentified
degradation products (0.37, 0.61, 0.80 and 0.99). [ring-U-14
C]-BAM and [Carboxy-14
C]-2,6-DCBA
(Moravek Biochemicals Inc., Brea, Ca) were used as analytical standards and analyzed on the first
line of each TLC plate.
3. Results
3.1. Mineralization of BAM
The most rapid mineralization of BAM was by the consortium with Aminobacter sp. MSH1 and
Mortierella sp. LEJ702 at all moisture contents. In the vials with moisture content of 10% of WHC
(fig. 1A) about 29% of the added BAM was mineralized by the consortium after 14 days, whereas
MSH1 alone had only mineralized 19%. No BAM was mineralized by Mortierella alone or in the

69
abiotic controls at any of the moisture contents. The stimulating effect on mineralization by
growing MSH1 with the fungus was even more pronounced at 5% of WHC (see fig. 1B) where
more than 50% of the added BAM was recovered as CO2. MSH1 alone only mineralized 16% of the
BAM at this WHC. Moreover, the mineralization rate was significantly increased by the consortium
compared to MSH1 alone.
A stimulation of the mineralization by the consortia was also seen at the lower moisture
contents of 1.7% and 0% of WHC (fig. 1C and D), although at an overall lower level with a
maximum of 35% and 27% recovered as CO2, respectively.
3.2. Transport of Aminobacter
The qPCR analysis revealed that Aminobacter sp. MSH1 had been transported to the very top of
the sand layer in all vials (fig. 2). For all moisture contents the greatest transport of the bacterium
through the sand was seen when grown with the fungus. The difference was most pronounced at a
moisture content of 5% of WHC, where the number of MSH1 detected in the top layer was more
than 2000 times higher in the consortium vials than in vials containing MSH1 alone. In the vials
with MSH1 alone the numbers of bacteria detected were approximately at the detection limit of 400
cells g-1
. At the highest moisture content, the MSH1 count for the consortium was 600 times higher
than for MSH1 alone.
3.3. Transport of BAM
The pesticide residue BAM was likewise transported up through the sand layer in both
experiments with Mortierella and in the abiotic controls (see fig. 3). Even in the syringes with a
moisture content of 0% of WHC, BAM was detected in the top layer of the sand, though at lower
levels than in syringes with higher moisture contents. More BAM appears to be transported to the
top sand in the syringes with Mortierella at 5%, 1.7% and 0% of WHC, though the difference is
only statistically significant at 0% of WHC. Thus, the ability of the Mortierella to facilitate
transport of BAM becomes increasingly pronounced the lower the moisture content. At the driest
condition (0% of WHC) about 10% of the added BAM was found in the top sand with Mortierella
compared to only 5.7% in the abiotic control.

70
3.4. Determination of degradation products
The results of the TLC analyses confirmed the results of the mineralization experiments, as the
most rapid dissipation of BAM was seen in vials containing both the bacterium and the fungus (fig.
4). Furthermore, the results show that degradation of BAM is a biological process as BAM was not
degraded in the abiotic controls.
The TLC analysis at termination of the mineralization experiment enabled elaboration of the
results, making it possible to account for the fraction of BAM that was not mineralized (fig. 5). At
5% of WHC a great deal more 14
C was recovered as BAM when MSH1 was grown alone compared
to the consortium (fig. 5B). The TLC analysis revealed that there was a significantly larger amount
of BAM remaining in the MSH1 treatment and that there had been a very limited partial
degradation of BAM. Similar to the results at the highest moisture contents, a small fraction of the
radiolabelled compound was detected as 2,6-dichlorobenzoic acid (2,6-DCBA).
The vials containing Mortierella alone were included in the TLC analyses at 1.7% moisture
content (fig. 5C). Results showed that approximately 80% of the initially added BAM could be
recovered in the vials with Mortierella. The TLC analysis also revealed the presence of an unknown
compound not present at the higher moisture content. The compound was only detected in the vials
containing Mortierella, indicating that it could be a fungal metabolite (designated ―F‖ in figure 5).
This potential fungal metabolite was also detected in the TLC analysis of the weekly harvested vials
in concentrations up to 5% of the added BAM (data not shown). More BAM remained in the
consortium vials at this moisture content than at higher moisture contents.
The distribution pattern at 0% moisture content was similar to that at 1.7% with less BAM
having been mineralized and more remaining as BAM. The unknown compound only detected in
the Mortierella vials was also detected here.
4. Discussion
One of the abiotic factors that has proven to strongly influence survival and ability to degrade
pollutants is moisture content (Cattaneo et al., 1997; Ronen et al., 2000). Ronen et al. (2000)
showed that low water contents led to poor bacterial survival, and at a water content of 10%
population size declined rapidly. This does not seem to be the case for Aminobacter sp. MSH1. In
fact, our results indicate that MSH1 was growing, as a high number could be detected at the top of
the sand layer at the end of the experiment (fig. 2). Growth of MSH1 seemed to be enhanced by the

71
presence of Mortierella sp. LEJ702, probably because of the exudates produced by the fungus being
utilised by the bacteria.
The mineralization results for MSH1 showed high activity by MSH1 at 10% of WHC (fig. 1A)
and an increase in activity at the lower moisture contents in the presence of Mortierella. Results by
Cattaneo et al. (1997) showed that moisture contents of between 30% and 90% of WHC were
required in order to have optimal biodegradation. The high activity of MSH1 even at low moisture
contents will likely increase the chance of successful bioremediation in natural heterogeneous
environments.
A fungal presence may not just increase activity. It has previously been demonstrated that fungi
produce exudates that in some cases can be utilised by bacteria and thereby increase their chance of
survival (de Boer et al., 2005; Boersma et al., 2010; Furuno et al., 2010). This could prove
especially important under stressful conditions such as drought or starvation. Fungi require less
moisture, and results by Cattaneo et al. (1997) showed that fungi dominate the microbial population
under dry conditions. Moreover, the ability of fungi to transport nutrients through the mycelium,
thus supporting hyphal growth in otherwise low-nutrient environments, renders the fungi less
affected by conditions that are stressful for other organisms, for example bacteria (Furuno et al.,
2010). This also seems to be the case in our experiments. Less BAM is mineralized by MSH1 when
the moisture content is reduced (fig. 1). The presence of the fungus reduces this effect. However,
detection of unknown compounds at the two lowest moisture contents and only in the presence of
Mortierella may indicate that the fungus was itself also affected at the two lower moisture contents
(fig. 5). Thus these compounds could be the result of a fungal response to increased stress.
Besides potentially providing nutrients, the fungal hyphae can function as transport vectors for
bacteria. Several experiments have shown that bacteria need the help of a transporting agent in
order to move through the soil (Madsen and Alexander, 1982; Goldstein et al., 1985; Kohlmeier et
al., 2005). The fungi-mediated transport may help the introduced bacteria survive as it enables
relocation, potentially to areas with better nutrient supply or less competition. Hydrophilic hyphae
provide a network of liquid pathways capable of bridging air-filled gaps, thus allowing dispersion
of motile bacteria to otherwise unavailable areas (Ritz and Young, 2004; Wick et al., 2007; Furuno
et al., 2010).
We have shown that moisture content plays an important role in the relocation of the BAM-
degrading MSH1 bacteria, as this declined with decreasing moisture content. We also showed that
bacterial transport was strongly increased in the presence of Mortierella, to some degree alleviating

72
the effects of the driest conditions, which corresponds well with previous findings. Furuno et al.
(2010), for example, found that both bacteria and chemicals could be translocated via the fungal
hyphae.
Studies on hydrophobic compounds have shown that fungal hyphae increase the access to
compounds (Kohlmeier et al., 2005; Wick et al., 2007). Other studies have shown that spatial
variability in pesticide degradation is correlated with heterogenous distribution of degraders rather
than with soil properties (Gonod et al., 2003; Gonod et al., 2006; Sjøholm et al. 2010). Our results
obtained with MSH1 also indicate that the presence of fungal hyphae increases the bioavailability of
BAM as greater rate of mineralization and degradation takes place with the consortium. These
findings support the hypothesis that accessibility to the pollutant is the main limitation for
successful removal of the compound and that a fungal-bacterial consortium is better equipped to
overcome this challenge. Hence, for bioremediation of pesticides and their metabolites fungal-
bacterial consortia may prove more efficient and future research should focus on investigating
effects of biotic as well as abiotic factors on these consortia.
Acknowledgements
The work was supported by the ―Microbial Remediation of Contaminated Soil and Water Resources
(MIRESOWA)‖ project funded by the Danish Council for Strategic Research (grant number 2104-
08-0012). The authors wish to thank Pia Bach Jakobsen for help with the qPCR method.
References
Boersma, F.G.H., Otten, R., Warmink, J.A., Nazir, R. and van Elsas, J.D., 2010. Selection of
Variovorax paradoxus-like bacteria in the mycosphere and the role of fungal-released
compounds. Soil Biology & Biochemistry 42, 2137-2145.
Cattaneo, M. V., Masson, C. and Greer C. W., 1997. The influence of moisture on microbial
transport, survival and 2,4-D biodegradation with a genetically marked Burkholderia cepacia
in unsaturated soil columns. Biodegradation 8, 87-96.
Chau, H.W., Goh, Y.K., Si, B.C. and Vujanovic, V., 2010. Assessment of alcohol percentage test
for fungal surface hydrophobicity measurement. Letters in Applied Microbiology 50, 295-300.

73
de Boer, W., Folman, L. B., Summerbell, R.C. and Boddy, L., 2005. Living in a fungal world:
impact of fungi on soil bacterial niche development. FEMS Microbiology Reviews 29(4),
795-811.
Edmonds, R. L., 1976. Survival of coliform bacteria in sewage sludge applied to a forest clearcut
and potential movement into groundwater. Applied and Environmental Microbiology 32 (4),
537-546.
El Fantroussi, S. and Agathos, S. N., 2005. Is bioaugmentation a feasible strategy for pollutant
removal and site remediation? Current opinion in Microbiology 8, 268-275
Furuno, S., Päzolt, K., Rabe, C., Neu, T. R., Harms, H. and Wick, L. Y., 2010. Fungal mycelia
allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-
unsaturated systems. Environmental Microbiology 12(6), 1391-1398.
Goldstein, R. M., Mallory, L. M. and Alexander, M., 1985. Reasons for possible failure of
inoculation to enhance biodegradation. Applied and Environmental Microbiology 50 (4), 977-
983.
Gonod, L. V., Chenu, C. and Soulas, G., 2003. Spatial variability of 2,4-dichlorophenoxyacetic acid
(2,4-D) mineralization potential at a millimeter scale in soil. Soil Biology & Biochemistry 35,
373-382.
Gonod, L. V., Chadoeuf, J. and Chenu, C., 2006. Spatial distribution of microbial 2,4-
Dichlorophenoxy acetic acid mineralization from field to microhabitat scales. Soil Science
Society of America 70, 64-71.
Harms, H. and Bosma, T. N. P., 1997. Mass transfer limitation of microbial growth and pollutant
degradation. Journal of Microbiology & Biotechnology 18, 97-105.
Kohlmeier, S., Smits, T. H. M., Ford, R. M., Keel, C., Harms, H. and Wick, L. Y., 2005. Taking the
fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environmental
Science and Technology 39, 4640-4646.
Madsen, E. L. and Alexander, M., 1982. Transport of Rhizobium and Pseudomonas through soil.
Soil Science Society of America Journal 46, 557-560.
Pepper, I. L., Gentry, T. J., Newby, D. T., Roane, T. M. and Josephson K. L., 2002. The role of cell
bioaugmentation and gene bioaugmentation in the remediation of co-contaminated soils.
Environmental Health Perspectives 110 (6), 943-946.
Ritz, K. and Young, I. M., 2004. Interactions between soil structure and fungi. Mycologist 18, 52-
59.

74
Ronen, Z., Vasiluk, L., Abeliovich, A. and Nejidat, A., 2000. Activity and survival of
tribromophenol-degrading bacteria in a contaminated desert soil. Soil Biology &
Biochemistry 32, 1643-1650.
Sjøholm, O. R., Aamand, J., Sørensen, J. and Nybroe, O., 2010. Degrader density determines spatial
variability of 2,6-dichlorobenzamide mineralization in soil. Environmental Pollution 158,
292-298.
Sørensen, S. R., Holtze, M. S., Simonsen, A. and Aamand, J., 2007. Degradation and mineralization
of nanomolar concentrations of the herbicide dichlobenil and its persistent metabolite 2,6-
dichlorobenzamide by Aminobacter spp. isolated from dichlobenil-treated soils. Applied and
Environmental Microbiology 73 (2), 399-406.
Thompson, I. P., van der Gast, C. J., Ciric, L. and Singer A. C., 2005. Bioaugmentation for
bioremediation: the challenge of strain selection. Environmental Microbiology 7(7), 909-915.
Thorling, L., Hansen, B., Langtofte, C., Brüsch, W., Møller, R. R., Mielby, S. and Højberg, A. L.,
2010. Grundvand. Status og udvikling 1989 – 2009. Teknisk rapport, GEUS 2010.
Van Veen, J. A., van Overbeek, L. S. and van Elsas, J. D., 1997. Fate and activity of
microorganisms introduced into soil. Microbiology and Molecular Biology Reviews 61 (2),
121-135.
Vogel, T. M., 1996. Bioaugmentation as a soil remediation approach. Current Opinion in
Biotechnology 7, 311-316.
Wick, L. Y., Remer, R., Würz, B., Reichenbach, J., Braun, S., Schäfer, F. and Harms, H., 2007.
Effect of fungal hyphae on the access of Bacteria to phenanthrene in soil. Environmental
Science and Technology 41, 500-505.
75
Figure 1. Results of the mineralization experiment at different moisture contents: A) 10% of WHC,
B) 5% of WHC, C) 1.7% of WHC and D) 0% of WHC. : MSH1 + Mortierella consortium, :
Aminobacter sp. MSH1, : Mortierella sp. LEJ702 and : Abiotic control.
76
Figure 2. Bacterial cell numbers in the top sand layer at the different moisture contents. M+M:
Aminobacter sp. MSH1 + Mortierella sp. LEJ702 consortium, and MSH1: Aminobacter sp.
MSH1. Numbers are averages.
Moisture content (% of WHC)
10 5 1.7 0
% o
f a
dd
ed
14C
-BA
M
0
5
10
15
20
25
Mortierella
abiotic
Figure 3. Percent of initially added 14
C-BAM transported to the top fraction of the sand columns.
Data are presented as means ± SE.
77
Days
0 10 20 30 40 50 60
% o
f in
itia
lly a
dde
d 1
4C
-BA
M
0
20
40
60
80
100
Figure 4. Data from the TLC analysis of the weekly harvested vials. Moisture contents: 1.7% of
WHC. : Aminobacter sp. strain MSH1 + Mortierella consortium (n=3), : Aminobacter sp. strain
MSH1 (n=3), and : Abiotic control (n=1). Data are means ± SE.
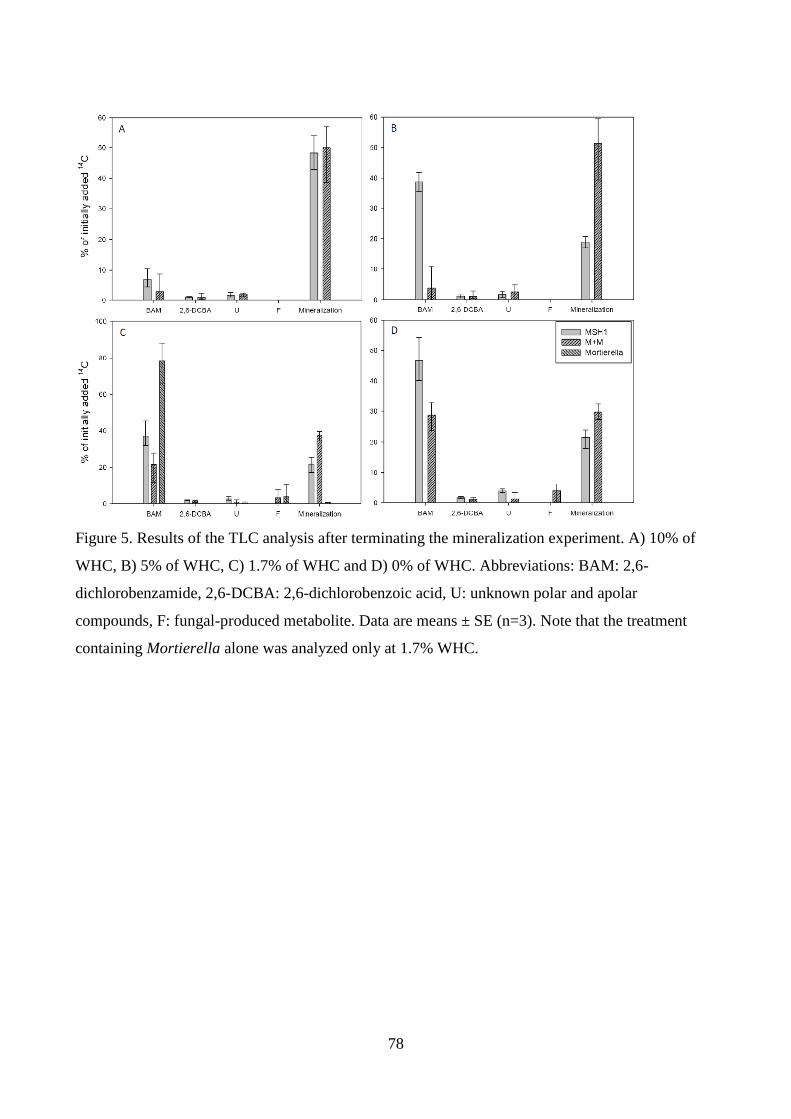
78
Figure 5. Results of the TLC analysis after terminating the mineralization experiment. A) 10% of
WHC, B) 5% of WHC, C) 1.7% of WHC and D) 0% of WHC. Abbreviations: BAM: 2,6-
dichlorobenzamide, 2,6-DCBA: 2,6-dichlorobenzoic acid, U: unknown polar and apolar
compounds, F: fungal-produced metabolite. Data are means ± SE (n=3). Note that the treatment
containing Mortierella alone was analyzed only at 1.7% WHC.

Manuscript III
FUNGAL-BACTERIAL CONSORTIUM OVERCOMES DRY PATCHES AND
INCREASES DIURON DEGRADATION
Lea Ellegaard-Jensen, Berith E. Knudsen, Anders Johansen,
Christian Nyrop Albers, Jens Aamand and Søren Rosendahl
In prep. for submission to Science of the Total Environment


81
Fungal-bacterial consortium overcomes dry patches and increases diuron
degradation
Lea Ellegaard-Jensena,b,§,*
, Berith Elkær Knudsena,b,§
, Anders Johansenc, Christian Nyrop Albers
b,
Jens Aamandb, and Søren Rosendahl
a.
aDepartment of Biology, Copenhagen University, Universitetsparken 15, DK-2100 Copenhagen Ø,
Denmark. bDepartment of Geochemistry, Geological Survey of Denmark and Greenland (GEUS),
Øster Voldgade 10, DK-1350 Copenhagen K, Denmark. cDepartment of Environmental Science,
Aarhus University, Frederiksborgvej 399, DK-4000, Roskilde, Denmark.
§Joint first authors
*Corresponding author: Lea Ellegaard-Jensen
Phone: +45 38142304, Email: [email protected]
Abstract
Consortia of fungal-bacterial strains could prove more efficient for bioremediation of
pesticide contaminations than individual strains alone. This may be due to co-operative catabolism
along the degradation pathway. The strains should, however, also be able to survive and preferably
spread into the environmental matrix e.g. soil. Fungal hyphae can act as transport vectors for
degrading bacteria. This may lead to a higher accessibility to the pollutant and ultimately to a more
efficient degradation.
To test this we chose the phenylurea herbicide diuron as our model compound. We
constructed a small column system in which diuron spiked sand and organisms were divided by a
layer of sterile glass beads for spatial separation. Fungal-bacterial consortia were created by
different combinations of the bacterial strains Sphingomonas sp. SRS2, Variovorax sp. SRS16 and
Arthrobacter globiformis D47 and the fungal strains Mortierella sp. LEJ702 and LEJ703.
Results showed that, of the consortia tested for degradation of 14
C-labeled diuron, the three-
member consortium LEJ702/SRS16/D47 achieved the overall highest mineralization measured as 14
CO2. In addition, production of diuron metabolites by this consortium was minimal. Molecular
results suggested that bacteria were transported more efficiently by LEJ702 than by LEJ703.
Finally, it was found that the fungal growth differed for LEJ702 and LEJ703 in the three-member
consortia. This study demonstrates new possibilities for creating efficient fungal-bacterial consortia
for bioremediation.
Keywords: Phenylurea herbicide, microbial consortia, synergistic interactions, fungal highway,
pesticide, biodegradation.

82
1. Introduction
The phenylurea herbicide diuron [N-(3,4-dichlorophenyl)-N,N-dimethyl-urea] is mainly used
for control of broad-leaved weeds on agricultural land, but also for weed control on non-cultivated
areas like roads, railways and parks. As a consequence of its use diuron is found in surface- and
groundwater at concentrations above the EU threshold limit of 0.1 µg l-1
(Lapworth and Gooddy,
2006; Struger et al., 2011; Torstensson, 2001). In addition, diuron has adverse effects on the
environment and presumably also on human health (Cox, 2003; Giacomazzi and Cochet, 2004), and
diuron is therefore on the EU Water Framework Directive´s list of priority substances (European
Parliament, 2008). In order to alleviate persistent diuron contamination in terrestrial environments,
bioremediation has been suggested as a possible means to remove diuron from soils.
Bioaugmentation, i.e. introduction of specific degrading microorganisms into the
environment, may be used as a technology to remediate diuron polluted soils. Finding strains
suitable for bioremediation is, however, challenging (Thompson et al., 2005). Not only should the
strains be able to degrade the contaminant, but they must also survive, proliferate and preferably be
spread into the soil matrix to obtain higher accessibility to the pollutants and ultimately a more
efficient degradation. It has been suggested that fungal-bacterial consortia might perform better than
single degrader strains in a heterogeneous soil matrix (Bennett et al., 2007; Mikesková et al., 2012);
this has been demonstrated for degradation of both hydrophobic (Wick et al., 2007) and hydrophilic
compounds (Knudsen et al., submitted). This synergistic effect has partly been explained by an
increased dispersal of the degrader bacteria along the hyphae of the fungal partner – a mechanism
referred to as the ´Fungal Highway´ (Furuno et al., 2010; Kohlmeier et al., 2005; Warmink and van
Elsas, 2009). The fungal hyphae may act exclusively as physical vectors for bacterial transport
enabling the bacteria to cross air-filled gaps in the matrix and in this way access otherwise
inaccessible contaminants. Certain fungi, however, also have potentials for degradation of
pollutants like phenylurea herbicides (Badawi et al., 2009; Ellegaard-Jensen et al., submitted;
Rønhede et al., 2005; Tixier et al., 2000) and may therefore, besides mediating bacterial transport,
also directly contribute to the degradation process.
We propose that consortia, consisting of both bacterial and fungal pesticide degraders might
lead to faster and more complete pesticide degradation in spatial heterogeneous systems, compared
to single strains. A synergistic effect on degradation by consortia may partly be due to the
individual degrader strains being metabolically active at different parts of the degradation pathway.
Recently, Sørensen et al. (2008) showed a synergistic effect on diuron degradation by a two-

83
member bacterial consortium consisting of Variovorax sp. SRS16 and Arthrobacter globiformis
D47.
Though microorganisms may exhibit mutual growth stimulating effects that may lead to
enhanced pesticide degradation, fungi and bacteria are also known to be antagonists (Höppener-
Ogawa et al., 2008; Mille-Lindblom and Tranvik, 2003; Trifonova et al., 2009). Clearly, knowledge
of such effects is necessary for evaluating the function of constructed consortia.
In this study we construct consortia of fungi and bacteria using different combinations of five
diuron-degrading organisms – the bacterial strains Sphingomonas sp. SRS2, Variovorax sp. SRS16
and Arthrobacter globiformis D47 and the fungal strains Mortierella sp. LEJ702 and LEJ703 to
elucidate the controlling mechanisms leading to enhanced degradation. The following mechanisms
will be examined: fungal mediated bacterial transport, co-operative catabolism i.e. the organisms
complementing each other along the metabolic degradation pathway, and physical interactions
between the organisms effecting growth.
2. Materials and methods
2.1. Chemicals and media
[Ring-U-14
C] diuron (36.3 mCi mmol-1
) was purchased from Izotop, (Institute of Isotopes Co.,
Ltd., Hungary) and had a radiochemical purity of 98.9%. A stock solution of radio labeled diuron
(1,700,000 DPM ml-1
) was prepared in acetonitrile.
Analytical-grade diuron [3-(3,4-dichlorophenyl)-1,1-dimethyl-urea] (CAS no. 330-54-1;
97.5% purity), DCPMU [1-(3,4-dichlorophenyl)-3-methylurea] (CAS no. 3567-62-2; 97.5% purity),
DCPU [1- (3,4-dichlorophenyl) urea] (CAS no. 2327-02-8; 99% purity), 3,4-DCA [3,4-
dichloroaniline] (CAS no. 95-76-1; 99% purity), and linuron [3-(3,4-dichlorophenyl)-1-methoxy-1-
methylurea] (CAS no. 330-55-2; 99.5% purity) were purchased from Dr. Ehrenstorfer GmbH
(Augsburg, Germany). 3,4-DCAA [3,4-dichloroacetanilide] (CAS no. 2150-93-8) was purchased
from Sigma-Aldrich (St. Louis, Missouri).
Growth media: Potato Dextrose Agar (PDA) (Difco, BD Diagnostics, Sparks, MD), Luria-
Bertani broth (LB) (Difco, BD Diagnostics, Sparks, MD), and R2A-based broth (R2B) prepared
according to (Reasoner and Geldreich, 1985).

84
MilliQ water with a phosphate-buffer (136 g l-1
KH2PO4; 178 g l-1
Na2HPO4.2H2O), giving a
final phosphate concentration of 6mM (pH 7), was used as liquid medium in experimental set-up.
Clean quartz sand (grain size 0.8-1.4 mm, 99.0% SiO2) was used as a growth matrix in the
experiments (DKI A/S, Denmark). The water holding capacity (WHC) was determined according to
Danish Standards; DS/ISO 14238-1 to be 25.2%. All media and sand were sterilized by autoclaving
at 121oC for 30 minutes before use.
2.2. Microorganisms
The diuron degrading fungal strains Mortierella sp. LEJ702 and Mortierella sp. LEJ703 were
isolated and described by Ellegaard-Jensen et al. (submitted). Both strains were maintained on PDA
at 8oC.
The diuron degrading bacterial strains Sphingomonas sp. SRS2 and Variovorax sp. SRS16
were both isolated and characterized in our laboratory (Sørensen et al., 2005; Sørensen et al., 2001),
while the diuron degrading Arthrobacter globiformis D47 (Turnbull et al., 2001) was kindly
provided by A. Walker. All strains were stored in 40% glycerol stock solutions at -80 °C.
Prior to the experiments, each bacterial strain was thawed and grown as follows: SRS2 in R2B,
SRS16 in R2B supplemented with 25 mg l-1
linuron, and D47 in LB supplemented with 25 mg l-1
diuron. Immediately before used for experiments, the strains were washed twice in buffered MilliQ
water.
2.3. Experiment 1: degradation of diuron
Degradation of diuron was studied using a setup described by Knudsen et al. (submitted) with
modifications (See Fig. S1). In short, the experiment was set up in sterile glass vials (Ø= 24 mm, h=
95 mm). PDA (500 μl) was added to each vial and surface-inoculated with either single strains, two-
member consortia or three-member consortia (See Table 1).
The fungi were added as agar plugs with mycelia (Ø = 4 mm). The bacteria were added in
suspensions corresponding to approximately 5x106 cells g
-1 sand. Vials without microorganisms
served as abiotic controls. A layer of sterile glass beads (2.97g; Ø = 1 mm, Assistent, Rhön,
Germany) was added on top of the inoculated PDA to separate the microbes from the contaminated
sand, simulating air-filled spaces in soil.
[Ring-U-14
C]-diuron dissolved in acetonitrile was added to 10 grams of quartz sand, giving a
concentration of 100 μg kg-1
. After evaporation of the acetonitrile, the sand was added buffered

85
MilliQ (corresponding to 10% of quartz sand WHC) and transferred to the vials. To trap the 14
CO2
evolved from the mineralization of the labeled diuron one small glass tube (Ø = 10.5 mm, h = 40
mm) with 1 ml 1M NaOH was added to each vial. The NaOH was replaced at regular intervals,
mixed with 5 ml scintillation liquid (Optiphase ‗hisafe‘ 3, PerkinElmer Inc, Waltham,
Massachusetts, USA), and the radioactivity in the solution was measured for 10 min on a Liquid
Scintillation Analyzer (Tri-Carb 2810 TR, PerkinElmer Inc., Waltham, Massachusetts, USA). All
treatments were carried out in triplicates. The experiment was incubated in the dark at 20o C.
At experimental termination sand from the top layer in each vial was gently mixed and 0.25 g
of this was used for DNA extraction (see Section 2.6). Subsequently, chemical extractions were
made from the vials to quantify diuron residues and metabolites using thin-layer chromatography
(TLC). All the vials were added 7ml MeOH:MilliQ (75:25), shaken for 2 hours (200rpm) and
centrifuged at 200 x g for 10 min. The supernatant was transferred to Eppendorf tubes and
centrifuged at 13 000 x g for 2 min. One ml of the supernatant was used for TLC (see Section 2.5)
and 500 µl was mixed with 5 ml Optiphase ‗hisafe‘ 3 scintillation liquid (PerkinElmer Inc) for
measurement of radioactivity in the solution.
2.4. Experiment 2: effect of bacteria on fungal growth
Growth of the fungal mycelium in the presence of the bacteria was examined in columns of
similar composition as described in experiment 1 (Section 2.3). Since the difference between the
three-member consortia LEJ702/SRS16/D47 and LEJ703/SRS16/D47 resides in the choice of
fungal strain, we wished to investigate the growth of these fungal strains within these consortia. The
experiment also included two-member consortia with the fungi and either of the bacterial strains
(see Table 1).
The experiment was set up in 20 ml sterile plastic syringes (Terumo Corporation, Leuven,
Belgium) with the front end of the barrel cut off. Syringes standing in a vertical position with the
cut-off end upwards had the pistons set at the 20-ml marking. PDA was added to each syringe and
inoculated with microorganisms (see Table 1). As above a layer of sterile glass beads (2.97g; Ø = 1
mm, Assistent, Rhön, Germany) was added on top of this for spatial separation of the microbes
from the diuron-spiked sand. Ten grams of quartz sand with a concentration of 100μg kg-1
[Ring-U-
14C]-diuron and a moisture content of 10% of WHC was then added to the syringes. Syringes were
sealed with rubber stoppers and parafilm. The experiment was incubated in the dark at 20oC. Twice
a week the syringes were opened for aeration in a sterile flow bench.

86
After 14 and 24 days triplicates from each treatment were harvested destructively. The sand
was pushed upwards using the piston, and the sand was divided into two fractions (i.e. top and
bottom) of approximately equal size. The fractions were instantly placed on ice and kept frozen
until phospholipid fatty acid (PLFA) analysis.
2.5. Thin-layer chromatography
Analysis of diuron and its degradation products were performed using TLC as previously
described (Sørensen et al., 2008). Briefly, 30 µl of centrifuged extract (see Section 2.3) was spotted
on a 10x20 cm Silica Gel 60 thin-layer chromatography plate (Merck, Darmstadt, Germany) and
eluted with a mixture of dichloromethane and methanol (95:5, vol/vol) in a CAMAG horizontal
developing chamber (CAMAG, Berlin, Germany). After air drying, the plates were developed on
phosphor imaging screens and analyzed autoradiographically on a Cyclone Scanner (Packard
Instrument Company, Meriden, CT). The chromatograms obtained were analyzed using the
OptiQuant Image Analysis software (Packard Instrument Company) to integrate the different bands.
Rf-values of identified compounds were: DCPU, 0.24; DCPMU, 0.30; 3,4-DCAA, 0.35; diuron,
0.37 and 3,4-DCA, 0.61.
2.6. Detection of bacteria with specific primers
DNA from 0.25 g top-sand from each vial (only experiment 1) was extracted using the
PowerLyzer PowerSoil® Isolation Kit (Mobio Laboratories Inc, Carlsbad, CA, USA) according to
the manufacturer‘s instructions. For detection of the bacteria the following specific primer sets were
used: (i) Sphingomonas, Sphingo108f - Sphingo420r (Leys et al., 2004), (ii) Variovorax, VarF –
VarR (Bers et al., 2011), and (iii) Actinobacteria/Arthrobacter, Com2xf – Ac1186r (Schäfer et al.,
2010). The PCR mixture and protocol (modified to 35 cycles) described by Leys et al. (2004) were
used with the Sphingo108f - Sphingo 420r primer pair to detect Sphingomonas sp. SRS2. Likewise,
for detection of Variovorax sp. SRS16 the PCR mixture and protocol described by Bers et al. (2011)
were used with the VarF – VarR primer set. For Arthrobacter globiformis D47 the PCR mixture
contained 1U Taq DNA polymerase, 62.5 pmol of each primer (Com2xf – Ac1186r), 25 nmol
dNTP, and 1 x PCR buffer in a final volume of 20 µl. The Arthrobacter PCR protocol was as
follows: an initial denaturation step at 95 o
C for 5 min, followed by 35 cycles consisting of
denaturation for 30 sec at 95 o
C, annealing for 30 sec at 55 o C and extension at 72
o C for 30 sec,

87
and finalized by an elongation step at 72 o C for 6 min. DreamTaq DNA polymerase and buffer
(Fermentas, St. Leon-Rot, Germany) were used in all PCR reactions.
2.7. Phospholipid fatty acid analysis (PLFA)
The procedure for extraction of phospholipid fatty acids was as described by Frostegård et al.
(1993) and modified by Johansen and Olsson (2005). Approximately 5 g of quartz-sand sample was
placed in Teflon centrifuge tubes (Oak Ridge, Nalge Nunc Int., US) and extracted in 10 ml of
dichloromethane/methanol/citrate buffer (0.15 M; pH4.0; 1:2:0.8,vol:vol:vol). Supernatants from
two repeated extractions were pooled and split into two phases by the addition of dichloromethane
and citrate buffer. Polar lipids from the lower phase were purified and derivatized according to
Joner et al. (2001). Samples were analyzed on an Agilant 7890 GC (Agilant, CA, USA) equipped
with an autoinjector (splitless mode), a flame ionization detector, and a 60-m HP5 column, using H2
as carrier (2 mL min-1
). The initial oven temperature was 80 °C (5 min), increased at 20 °C min-1
to
160 °C and at 5 °C min-1
to 270 °C (maintained for 5 min). Inlet and detector temperatures were
230 and 270 °C, respectively. In addition, PLFAs of the bacterial and fungal pure strains were
extracted (as described above) and the individual fatty acids identified using the Midi Sherlock
system (Mansfeld-Giese et al., 2002).
2.8. Statistics
Statistical analysis of PLFA data was performed using SYSTAT v. 9.0 using the GLM
procedure to test for main effects of consortium type and time of incubation (14 and 24 days) as
well as interactions. Bonferroni Tukey multiple comparison analysis was used to determine
significance of differences (presented as P values) between consortia at day 24.
3. Results
3.1. Mineralization and transformation by consortia
The LEJ702/SRS16/D47 consortium mineralized 32.2 ± 5.7 % of the total amount of diuron
after 54 days (Fig. 1), where the single strains or other consortia reached no more than 10 %
mineralization (Fig. 1 and Table 1). Low mineralization rates does not imply that diuron was not
degraded by some of the stains or consortia as metabolites (DCPMU, DCPU and 3,4-DCA) were
found in several treatments (Fig. 2 and Fig. S2). In the abiotic treatments, no degradation products

88
were detected. The degradation pathway of diuron is illustrated in Fig. 3, where the steps performed
by either fungal or bacterial strains are also seen.
Both Mortierella strains degraded diuron to DCPMU and DCPU (Fig. 2A and Fig. S2D),
whereas the bacterial single strains and consortia with solely bacterial strains degraded only
minimal amounts of diuron (Fig. S2). With the LEJ702/SRS16 consortium, a small amount of the
diuron was mineralized while the two demethylated metabolites were also present (Fig. 2B).
Similarly, in the LEJ702/D47 consortium DCPMU and DCPU were present, in addition 3,4-DCA
and 3,4-DCAA (Fig. 2C) were found. The three-member LEJ702/SRS16/D47 consortium was the
most effective combination of microorganisms as only small amounts of intermediary metabolites
could be detected and one third of the diuron was mineralized to CO2 (Fig. 2D).
Some 14
C did not migrate on the TLC plate and was therefore termed non-extractable fraction
(NEF). This fraction varied in the different treatments and was generally largest in treatments with
LEJ703 where it constituted as much as 50% of the 14
C originating from diuron compared to 10-
25% in treatments without this strain and 0% in the abiotic control (Fig. S2). Part of the NEF
fraction (50-80%) was found to be able to move in a more polar solvent (MeOH:CH2Cl2 ratio
reversed from 5:95 to 95:5). This method could not be used for quantification since diuron and its
metabolites were not chromatographically separated, but it indicates that the NEF includes one or
more unidentified polar degradation products rather than being 14
C bound in non-dissolvable
organic macro structures as observed in a soil system by (Sørensen et al., 2008).
3.2. Transport of bacteria
PCR analysis with Variovorax-specific primers revealed that Variovorax sp. SRS16 could be
detected in the top sand layer, but only in the vials containing the three-member consortium
LEJ702/SRS16/D47. Likewise, Arthrobacter globiformis D47 was only detected in samples from
the top sand layer of the vials containing the two-member consortium LEJ702/D47 when analyzed
with Actinobacteria/Arthrobacter-specific primers (data not shown).
These observations indicate that bacteria were only transported to the top of the sand column
when Mortierella sp. LEJ702 was present. Interestingly, in the vials with Mortierella sp. LEJ703 no
bacteria could be detected in the top sand independent of the treatments. Analysis with the
Sphingomonas-specific primers did not detect Sphingomonas sp. SRS2 in the top sand layer in any
of the treatments.

89
3.3. Growth of Mortierella
PLFAs from Mortierella sp. LEJ702 and LEJ703 were measured in high amounts in the
presence of Variovorax sp. SRS16, with significantly more in the uppermost sand layer than in the
bottom (Fig. 4A, B; main effect top vs. bottom P<0.001), but with no statistically significant
difference between day 14 and 24 for any of the consortia. In contrast, the sand content of PLFAs
indicative of both the Mortierella strains were much lower when A. globiformis D47 was present,
both in the top and bottom of the sand column (Fig. 4; main effect of consortia P<0.001). For
Mortierella sp. LEJ703 this D47-induced decrease in growth was still very pronounced in the three-
member consortium. In comparison, the inhibition effect on the biomass of Mortierella sp. LEJ702
was lessened in the three-member consortium at day 24 (Fig. 4B; P<0.022). This indicates that the
presence of SRS16 alleviates the growth inhibition that D47 causes on Mortierella sp. LEJ702. The
PLFAs indicative of Variovorax sp. SRS16 and A. globiformis D47 could not be detected in
significant proportions (above detection level) in any of the treatment (data not shown).
4. Discussion
Reports of consortia including both fungal and bacterial degraders are scarce, and our finding
that a fungal-bacterial three-member consortium can indeed increase pesticide degradation has to
our knowledge not been reported previously. The three-member consortium LEJ702/SRS16/D47
was mineralizing three times more diuron than either of the single strains or other consortia tested.
The two-member consortium SRS16/D47 did not mineralize more than 8.3 ± 2.6 % of the added
diuron in our experiment. This is in contrast to previous studies where the two bacterial strains in
co-culture have been reported to enhance degradation of diuron with approximately 60 % of the
added 14
C-diuron mineralized in liquid cultures (Sørensen et al., 2008). This suggests that the
bacteria might not have been able to cross the air-filled gap created by the glass-bead-layer and
reach the diuron in the sand in the present study.
The fungal hyphae may have acted as transport vector for the degrading bacteria enabling the
bacteria to cross air-filled gaps to reach the diuron in the sand. Such hyphal mediated bacterial
transport, known as ´the fungal highway´(Kohlmeier et al., 2005), has been demonstrated by other
authors previously, where transport of bacteria was shown to occur in the thin water film around
the fungal hyphae (Furuno et al., 2010; Wick et al., 2010).

90
In heterogeneous and water-unsaturated environments, e.g. soil, bacteria are almost
completely immobilized and can, unlike fungi, not cross over air-filled gaps without a transport
vector (Banitz et al., 2011a; Banitz et al., 2011b). Therefore, bioavailability has been suggested to
be a critical factor for bioremediation success (Bosma et al., 1996) - a challenge which may be
overcome by applying fungal-bacterial consortia.
In agreement with our findings that fungal transport of bacterial degraders increase
degradation of diuron, Wick et al. (2007) found that transport of phenanthrene degrading bacteria
via fungal hyphae increased the degradation of this compound. Likewise, Knudsen et al. (submitted)
showed for the pesticide metabolite 2,6-dichlorobenzamide (BAM) that transport of the degrader
bacterium also increased BAM mineralization. Transport of bacteria was only detected in consortia
with Mortierella sp. LEJ702, and not in consortia with Mortierella sp. LEJ703. Whether this is due
to morphological or physiological differences between the two fungal strains is not known.
Kohlmeier et al. (2005) showed that fungal transport of Achromobacter sp. SK1 strongly depended
on the surface hydrophobicity of the fungal hyphae, with enhanced transport along the hydrophilic
hyphae of Fusarium oxysporum. It could be speculated that this could also be the principle cause in
the present experiment.
A co-operative catabolism may be expected when inspecting the degradation pathway of
diuron (Fig. 3). Here it is seen that Sphingomonas sp. SRS2 and Mortierella degrade diuron by
demethylation (Badawi et al., 2009; Ellegaard-Jensen et al., submitted; Sørensen and Aamand,
submittet), while Variovorax sp. SRS16 and A. globiformis D47 degrades diuron directly to 3,4-
dichloroaniline (Sørensen et al., 2008).
The single bacterial strains degraded only limited amounts of diuron in the present study (Fig.
S2 A-C). Low concentrations of 3,4-DCA were detected with D47 alone and in the two-member
consortium LEJ702/D47. However, in the two-member consortium the demethylated metabolites
DCPU and DCPMU along with the metabolite 3,4-DCAA were also present (Fig. 2C). This can be
seen as a clear indication of co-operative catabolism since this metabolite is created from 3,4-DCA
by the fungus (Fig. 3). This process was also demonstrated by Tixier et al. (2002) testing the ability
of four fungal strains to degrade synthesized 3,4-DCA in pure cultures. Though the process does not
lead to complete mineralization, it still leads to detoxification as 3,4-DCAA is less toxic than 3,4-
DCA (Tixier et al., 2002). Variovorax sp. SRS16 is the organism with the highest potential for
complete mineralization of diuron among the tested strains (Sørensen et al., 2008). Although some
mineralization was seen in all treatments with SRS16 (Table 1), it was only in the three-member

91
consortium LEJ702/SRS16/D47 that the mineralization was markedly increased (Fig. 1). This
suggests that there were other distinctive interactions between these three organisms, besides
cooperative catabolism and bacterial transport, which enabled them to perform superior compared
to the other consortia.
The two Mortierella strains differed in their growth response in the consortia. In the three-
member consortia with SRS16 and D47, the growth of LEJ703 was clearly restricted compared to
that of LEJ702 (Fig. 4). This outcome seemed to be caused by a D47-inhibitory effect on both
fungi, which was alleviated by SRS16, although only for LEJ702. This may, at least partly, explain
the increased diuron mineralization in the three-member consortium LEJ702/SRS16/D47 compared
to the consortium LEJ703/SRS16/D47 (Table 1).
The two bacteria SRS16 and D47 exerted different effects on the growth of the fungi. Bacteria
are known to potentially inhibit fungal growth. Trifonova et al. (2009) found that four out of seven
bacterial species changed the mycelium morphology and reduced colony size of the ascomycete
Coniochaeta ligniaria grown on agar plates. We did, however, not detect any inhibitory effects on
fungal morphology and growth on agar plates with the bacterial and fungal strains used in the
present work (data not shown). The PLFA method enabled us to follow the growth dynamics of
fungi in the constructed consortia and it has, to our knowledge, not been used in this way before.
The PLFA profiles of the two fungal strains were identical and very different from those of the
bacterial strains. PLFAs from the bacterial partners were not quantifiable in any of the consortia.
From the PCR analysis we know that Variovorax sp. SRS16 and, Arthrobacter globiformis D47
were present in the sand compartment, although this must have been in proportions of biomass
which were far less than the fungi. Hence, the present results indicate that efficient degradation of
contaminants by microbial consortia can proceed even with large differences in amount of biomass
between the microbial partners involved in the degradation.
Fungal exudates of Mortierella sp. LEJ702 might also have contributed to the synergistic
effect on diuron degradation. Exudates have been shown to be an important source of carbon for
bacteria in the hyphosphere (Boersma et al., 2010; de Boer et al., 2005; Nazir et al., 2010;
Trifonova et al., 2009), and since diuron was the only added carbon source available for the
bacteria in the sand they were likely to depend on the fungal hyphae for transport and release of
water and nutrients. Previously, it has been demonstrated that the mineralization of diuron by
SRS16 only occurred when it was supplemented with appropriate growth substrates or in co-culture
with D47 (Sørensen et al., 2008).

92
Our study underlines the importance of careful strain selection in constructions of consortia
for bioremediation. Fungal-bacterial consortia for degradation of PAHs have been created with
varying success; either showing no positive effect (Arun and Eyini, 2011) or enhanced PAH
degradation by specific consortia (Boersma et al., 2010; Machín-Ramírez et al., 2010). The last case
supports our findings that fungal-bacterial consortia may be useful for the remediation of polluting
compounds, but also that more research is needed to unravel the complex interactions within the
consortia. It is valuable to gain knowledge about these interactions if the ultimate aim is
bioaugmentation of soil where competition with indigenous microorganisms will add to the
complexity. The interactions with indigenous soil microorganisms should therefore be thoroughly
investigated including performance of the consortium in soil mesocosms with regard to its
degradation efficiency, before establishing its applicability for in situ bioremediation.
In conclusion, the present study shows that microbial consortia may be much more efficient in
degrading chemical contaminants compared to the consortium members individually. However, it
also shows that the traits of the member organisms are important to obtain the synergism necessary
for optimal remediation in a heterogeneous environment like soil. Our results also show that a
fungal partner is crucial for spatial distribution of the bacterial degraders.
Acknowledgments
This work was supported by grant 2104-08-0012 from the Danish Council for Strategic Research
given through the MIRESOWA project. Anders Johansen was supported by the SOILEFFECTS
project funded by the Norwegian Research Council. We thank Dr. Sebastian R. Sørensen for
providing the strains SRS2 and SRS16 and Pia B. Jakobsen, Pernille Stockmarr and Spire
Kiersgaard for skillful technical assistance in the laboratory.
References
Arun A, Eyini M. Comparative studies on lignin and polycyclic aromatic hydrocarbons degradation by basidiomycetes fungi. Bioresource Technology 2011; 102: 8063-8070.
Badawi N, Ronhede S, Olsson S, Kragelund BB, Johnsen AH, Jacobsen OS, et al. Metabolites of the phenylurea herbicides chlorotoluron, diuron, isoproturon and linuron produced by the soil fungus Mortierella sp. Environmental Pollution 2009; 157: 2806-2812.
Banitz T, Fetzer I, Johst K, Wick LY, Harms H, Frank K. Assessing biodegradation benefits from dispersal networks. Ecological Modelling 2011a; 222: 2552-2560.

93
Banitz T, Wick LY, Fetzer I, Frank K, Harms H, Johst K. Dispersal networks for enhancing bacterial degradation in heterogeneous environments. Environmental Pollution 2011b; 159: 2781-2788.
Bennett JW, Connick WJ, Daigle D, Wunch K. Formulation of fungi for in situ bioremediation. In: Gadd GM, Watkinson SC, Dyer PS, editors. Fungi in the Environment. Cambridge University Press, Cambridge, UK, 2007, pp. 97-112.
Bers K, Sniegowski K, Albers P, Breugelmans P, Hendrickx L, De Mot R, et al. A molecular toolbox to estimate the number and diversity of Variovorax in the environment: application in soils treated with the phenylurea herbicide linuron. FEMS Microbiology Ecology 2011; 76: 14-25.
Boersma FGH, Otten R, Warmink JA, Nazir R, van Elsas JD. Selection of Variovorax paradoxus-like bacteria in the mycosphere and the role of fungal-released compounds. Soil Biology and Biochemistry 2010; 42: 2137-2145.
Bosma TNP, Middeldorp PJM, Schraa G, Zehnder AJB. Mass Transfer Limitation of Biotransformation: Quantifying Bioavailability. Environmental Science & Technology 1996; 31: 248-252.
Cox C. Herbicide factsheet - DIURON. Journal of Pesticide Reform 2003; 23: 12-20. de Boer W, Folman LB, Summerbell RC, Boddy L. Living in a fungal world: impact of fungi on soil bacterial
niche development. FEMS Microbiology Reviews 2005; 29: 795-811. Ellegaard-Jensen L, Aamand J, Kragelund BB, Johnsen AH, Rosendahl S. Degradation of the phenylurea
herbicide diuron by soil fungi: different degradation potentials of five Mortierella strains. submitted.
European Parliament EU. Priority Substances Directive (Directive 2008/105/EC): Official Journal of the European Union., 2008.
Frostegård Å, Tunlid A, Bååth E. Phospholipid Fatty Acid Composition, Biomass, and Activity of Microbial Communities from Two Soil Types Experimentally Exposed to Different Heavy Metals. Applied and Environmental Microbiology 1993; 59: 3605-3617.
Furuno S, Päzolt K, Rabe C, Neu TR, Harms H, Wick LY. Fungal mycelia allow chemotactic dispersal of polycyclic aromatic hydrocarbon-degrading bacteria in water-unsaturated systems. Environmental Microbiology 2010; 12: 1391-1398.
Giacomazzi S, Cochet N. Environmental impact of diuron transformation: a review. Chemosphere 2004; 56: 1021-1032.
Höppener-Ogawa S, Leveau JHJ, van Veen JA, De Boer W. Mycophagous growth of Collimonas bacteria in natural soils, impact on fungal biomass turnover and interactions with mycophagous Trichoderma fungi. ISME J 2008; 3: 190-198.
Johansen A, Olsson S. Using Phospholipid Fatty Acid Technique to Study Short-Term Effects of the Biological Control Agent Pseudomonas fluorescens DR54 on the Microbial Microbiota in Barley Rhizosphere. Microbial Ecology 2005; 49: 272-281.
Joner EJ, Johansen A, Loibner AP, dela Cruz MA, Szolar OHJ, Portal J-M, et al. Rhizosphere Effects on Microbial Community Structure and Dissipation and Toxicity of Polycyclic Aromatic Hydrocarbons (PAHs) in Spiked Soil. Environmental Science & Technology 2001; 35: 2773-2777.
Knudsen BE, Ellegaard-Jensen L, Albers CN, Rosendahl S, Aamand J. Transport of degrader bacteria via fungal hyphae increases degradation of 2,6-dichlorobenzamide. submitted.
Kohlmeier S, Smits THM, Ford RM, Keel C, Harms H, Wick LY. Taking the fungal highway: Mobilization of pollutant-degrading bacteria by fungi. Environmental Science & Technology 2005; 39: 4640-4646.
Lapworth DJ, Gooddy DC. Source and persistence of pesticides in a semi-confined chalk aquifer of southeast England. Environmental Pollution 2006; 144: 1031-1044.
Leys NMEJ, Ryngaert A, Bastiaens L, Verstraete W, Top EM, Springael D. Occurrence and Phylogenetic Diversity of Sphingomonas Strains in Soils Contaminated with Polycyclic Aromatic Hydrocarbons. Applied and Environmental Microbiology 2004; 70: 1944-1955.
Machín-Ramírez C, Morales D, Martínez-Morales F, Okoh AI, Trejo-Hernández MR. Benzo[a]pyrene removal by axenic- and co-cultures of some bacterial and fungal strains. International Biodeterioration & Biodegradation 2010; 64: 538-544.

94
Mansfeld-Giese K, Larsen J, Bødker L. Bacterial populations associated with mycelium of the arbuscular mycorrhizal fungus Glomus intraradices. FEMS Microbiology Ecology 2002; 41: 133-140.
Mikesková H, Novotný Č, Svobodová K. Interspecific interactions in mixed microbial cultures in a biodegradation perspective. Applied Microbiology and Biotechnology 2012: 1-10.
Mille-Lindblom C, Tranvik LJ. Antagonism between Bacteria and Fungi on Decomposing Aquatic Plant Litter. Microbial Ecology 2003; 45: 173-182.
Nazir R, Warmink JA, Boersma H, Van Elsas JD. Mechanisms that promote bacterial fitness in fungal-affected soil microhabitats. FEMS Microbiology Ecology 2010; 71: 169-185.
Reasoner DJ, Geldreich EE. A new medium for the enumeration and subculture of bacteria from potable water. Applied and Environmental Microbiology 1985; 49: 1-7.
Rønhede S, Jensen B, Rosendahl S, Kragelund BB, Juhler RK, Aamand J. Hydroxylation of the herbicide isoproturon by fungi isolated from agricultural soil. Appl. Environ. Microbiol. 2005; 71: 7927-7932.
Schäfer J, Jäckel U, Kämpfer P. Development of a new PCR primer system for selective amplification of Actinobacteria. Fems Microbiology Letters 2010; 311: 103-112.
Sørensen SR, Aamand J. Degradation and mineralisation of diuron by Sphingomonas sp. SRS2 and its potential for remediating at a realistic µg l-1 diuron concentration. submittet.
Sørensen SR, Albers CN, Aamand J. Rapid mineralization of the phenylurea herbicide diuron by Variovorax sp. SRS16 in pure culture and within a two-member consortium. Appl. Environ. Microbiol. 2008; 74: 2332-2340.
Sørensen SR, Rasmussen J, Jacobsen CS, Jacobsen OS, Juhler RK, Aamand J. Elucidating the Key Member of a Linuron-Mineralizing Bacterial Community by PCR and Reverse Transcription-PCR Denaturing Gradient Gel Electrophoresis 16S rRNA Gene Fingerprinting and Cultivation. Applied and Environmental Microbiology 2005; 71: 4144-4148.
Sørensen SR, Ronen Z, Aamand J. Isolation from agricultural soil and characterization of a Sphingomonas sp. able to mineralize the phenylurea herbicide isoproturon. Applied and Environmental Microbiology 2001; 67: 5403-5409.
Struger J, Grabuski J, Cagampan S, Rondeau M, Sverko E, Marvin C. Occurrence and distribution of sulfonylurea and related herbicides in central Canadian surface waters 2006-2008. Bulletin of environmental contamination and toxicology 2011; 87: 420-5.
Thompson IP, van der Gast CJ, Ciric L, Singer AC. Bioaugmentation for bioremediation: the challenge of strain selection. Environmental Microbiology 2005; 7: 909-915.
Tixier C, Bogaerts P, Sancelme M, Bonnemoy F, Twagilimana L, Cuer A, et al. Fungal biodegradation of a phenylurea herbicide, diuron : structure and toxicity of metabolites. Pest Management Science 2000; 56: 455-462.
Tixier C, Sancelme M, Aït-Aïssa S, Widehem P, Bonnemoy F, Cuer A, et al. Biotransformation of phenylurea herbicides by a soil bacterial strain, Arthrobacter sp. N2: structure, ecotoxicity and fate of diuron metabolite with soil fungi. Chemosphere 2002; 46: 519-526.
Torstensson L. Use of herbicides on railway tracks in Sweden. Pesticide Outlook 2001; 12: 16-21. Trifonova R, Postma J, Van Elsas JD. Interactions of plant-beneficial bacteria with the ascomycete
Coniochaeta ligniaria. Journal of Applied Microbiology 2009; 106: 1859-1866. Turnbull GT, Cullington JC, Walker AW, Morgan JM. Identification and characterisation of a diuron-
degrading bacterium. Biology and Fertility of Soils 2001; 33: 472-476. Warmink JA, van Elsas JD. Migratory Response of Soil Bacteria to Lyophyllum sp. Strain Karsten in Soil
Microcosms. Appl. Environ. Microbiol. 2009; 75: 2820-2830. Wick LY, Furuno S, Harms H. Fungi as Transport Vectors for Contaminants and Contaminant-Degrading
Bacteria. In: Timmis KN, editor. Handbook of Hydrocarbons and Lipid Microbiology. Springer Berlin, 2010, pp. 1555-1561.
Wick LY, Remer R, Wurz B, Reichenbach J, Braun S, Scharfer F, et al. Effect of fungal hyphae on the access of bacteria to phenanthrene in soil. Environmental Science & Technology 2007; 41: 500-505.

95
Table 1. Accumulated 14
CO2 produced from 14
C-labeled
diuron during 54 days incubation.
Treatment
Mineralization
(% of added 14
C diuron ± SE)
Sphingomonas sp. SRS2 2.2 ± 0.4 %
Variovorax sp. SRS16 4.4 ± 0.8 %
Arthrobactor sp. D47 4.8 ± 1.0 %
Mortierella sp. LEJ702 0.4 ± 0.1 %
Mortierella sp. LEJ703 1.6 ± 0.2 %
Two member consortia
SRS2/SRS16 7.3 ± 1.3 %
D47/SRS16* 8.3 ± 2.6 %
LEJ702/SRS2 0.6 ± 0.2 %
LEJ702/SRS16* 7.3 ± 2.8 %
LEJ702/D47* 1.3 ± 0.4 %
LEJ703/SRS2 1.1 ± 0.2 %
LEJ703/SRS16* 10.0 ± 3.5 %
LEJ703/D47* 0.9 ± 0.1 %
Three member consortia
LEJ702/SRS16/SRS2 7.0 ± 1.9 %
LEJ702/SRS16/D47* 32.2 ± 5.7 %
LEJ703/SRS16/SRS2 9.1 ± 4.1 %
LEJ703/SRS16/D47* 7.2 ± 1.4 %
* treatments also used in experiment 2

96
Days
0 10 20 30 40 50
% 1
4C
O2 o
f added 1
4C
-diu
ron
0
10
20
30
40
SRS16
D47
LEJ702
SRS16&D47
LEJ702&SRS16
LEJ702&D47
LEJ702&SRS16&D47
Figure 1. Mineralization of 14
C-labeled diuron by single strains and consortia. 14
C-diuron
mineralization by (●) Variovorax sp. SRS16, (○) Arthrobacter globiformis D47, (▼) Mortierella
sp. LEJ702, (∆) consortium SRS16/D47, (■) consortium LEJ702/SRS16, (□) consortium
LEJ702/D47, and (♦) consortium LEJ702/SRS16/D47. Data are presented as means ± SE.
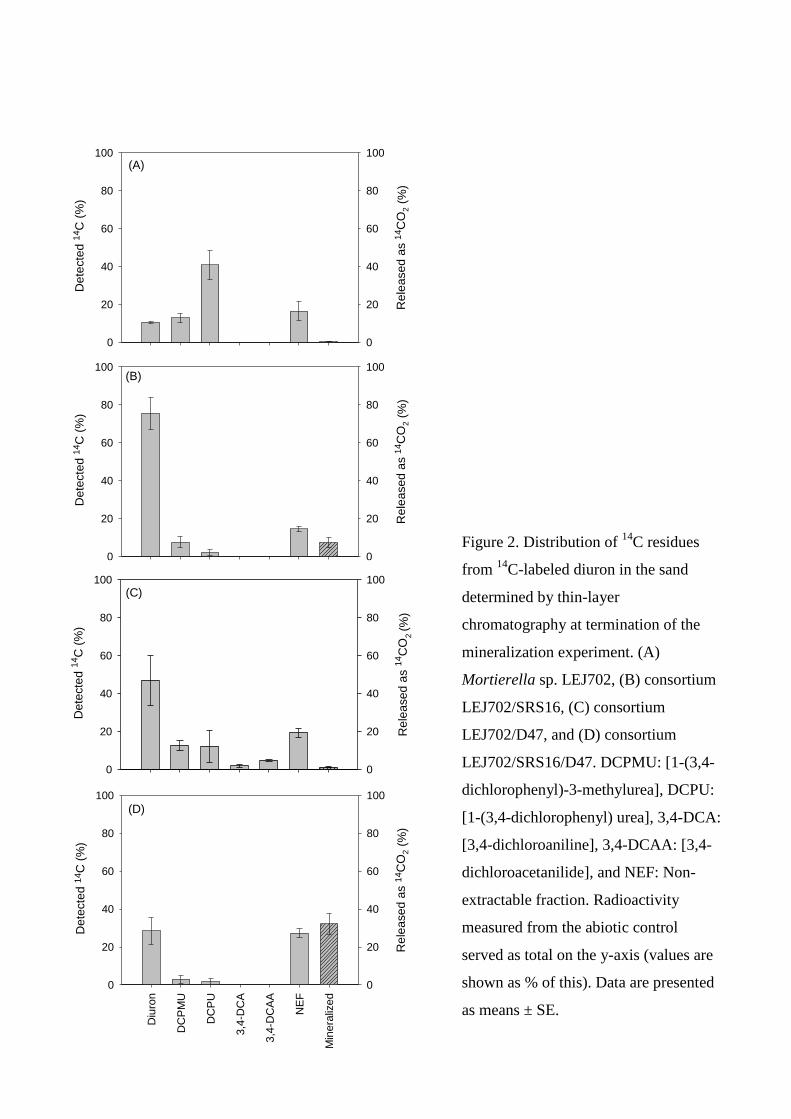
97
(A)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(B)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(C)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lease
d a
s 1
4C
O2
(%
)
0
20
40
60
80
100
(D)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
3,4
-DC
AA
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lease
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
Figure 2. Distribution of 14
C residues
from 14
C-labeled diuron in the sand
determined by thin-layer
chromatography at termination of the
mineralization experiment. (A)
Mortierella sp. LEJ702, (B) consortium
LEJ702/SRS16, (C) consortium
LEJ702/D47, and (D) consortium
LEJ702/SRS16/D47. DCPMU: [1-(3,4-
dichlorophenyl)-3-methylurea], DCPU:
[1-(3,4-dichlorophenyl) urea], 3,4-DCA:
[3,4-dichloroaniline], 3,4-DCAA: [3,4-
dichloroacetanilide], and NEF: Non-
extractable fraction. Radioactivity
measured from the abiotic control
served as total on the y-axis (values are
shown as % of this). Data are presented
as means ± SE.
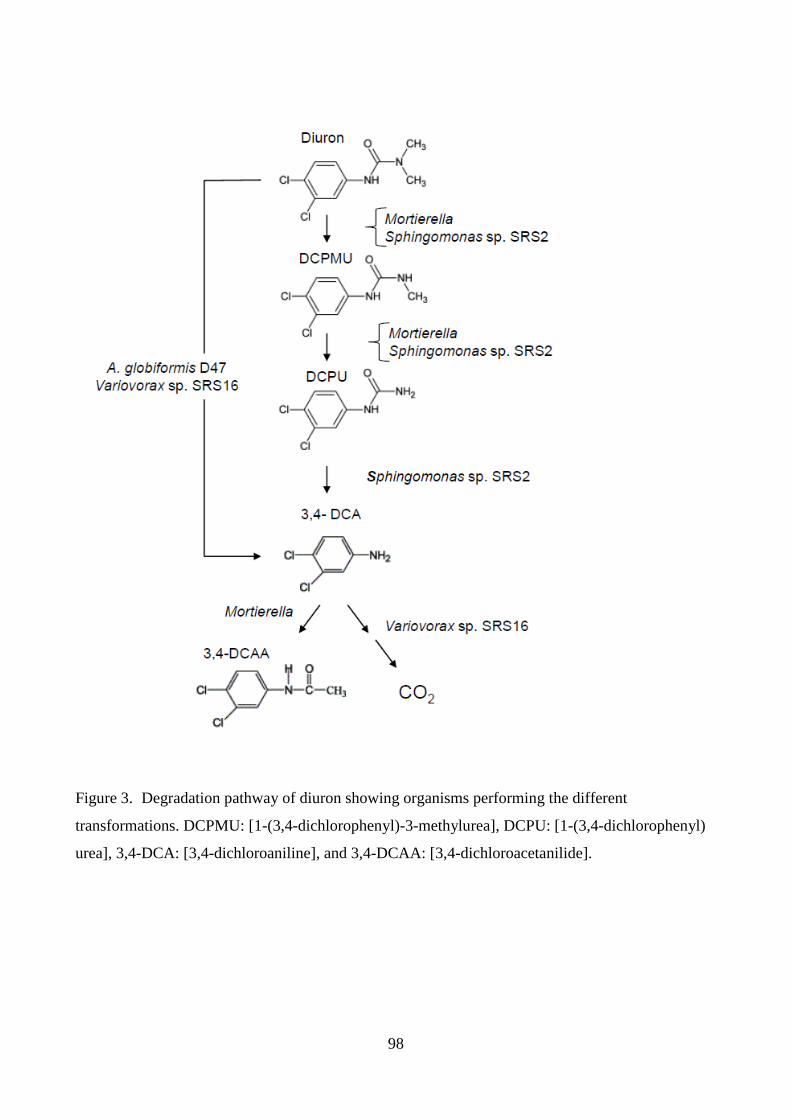
98
Figure 3. Degradation pathway of diuron showing organisms performing the different
transformations. DCPMU: [1-(3,4-dichlorophenyl)-3-methylurea], DCPU: [1-(3,4-dichlorophenyl)
urea], 3,4-DCA: [3,4-dichloroaniline], and 3,4-DCAA: [3,4-dichloroacetanilide].
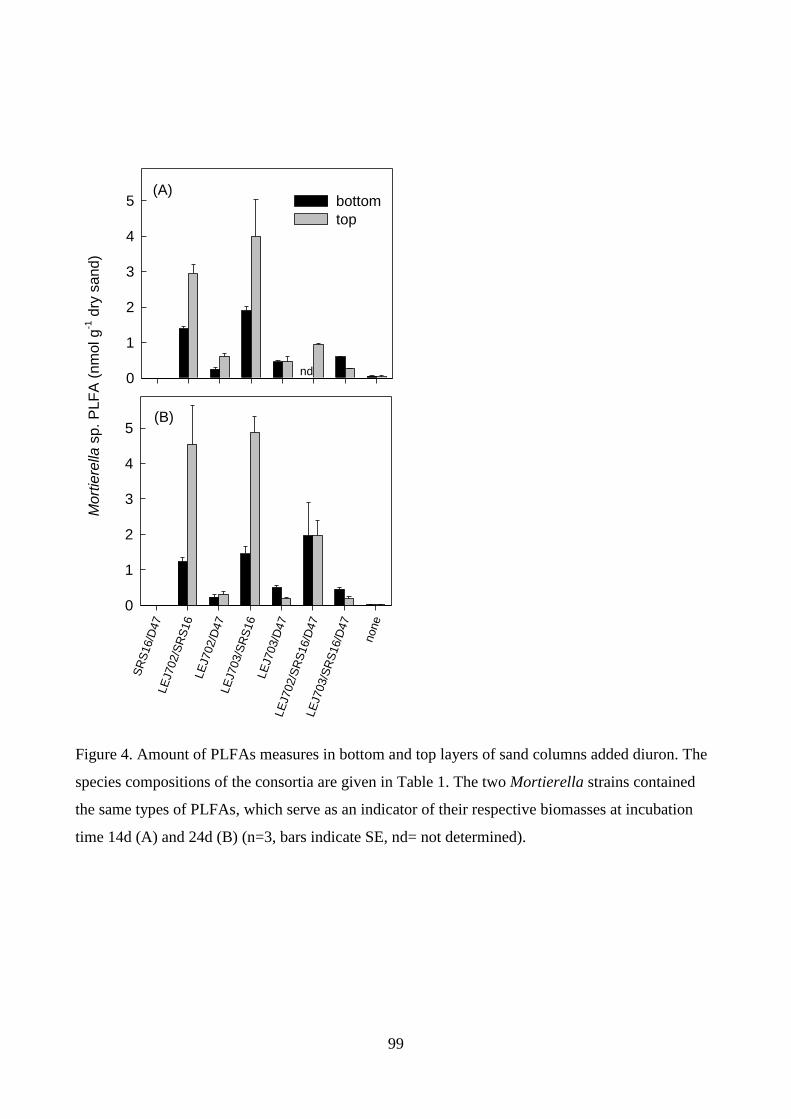
99
Mort
iere
lla s
p.
PLF
A (
nm
ol g
-1 d
ry s
and)
0
1
2
3
4
5 bottom
top
SR
S16/D
47
LE
J702/S
RS
16
LE
J702/D
47
LE
J703/S
RS
16
LE
J703/D
47
LE
J702/S
RS
16/D
47
LE
J703/S
RS
16/D
47
none
0
1
2
3
4
5
nd
(A)
(B)
Figure 4. Amount of PLFAs measures in bottom and top layers of sand columns added diuron. The
species compositions of the consortia are given in Table 1. The two Mortierella strains contained
the same types of PLFAs, which serve as an indicator of their respective biomasses at incubation
time 14d (A) and 24d (B) (n=3, bars indicate SE, nd= not determined).

100
Fig. S1. Illustration of mineralization experiment
setup. See Section 2 for written description.
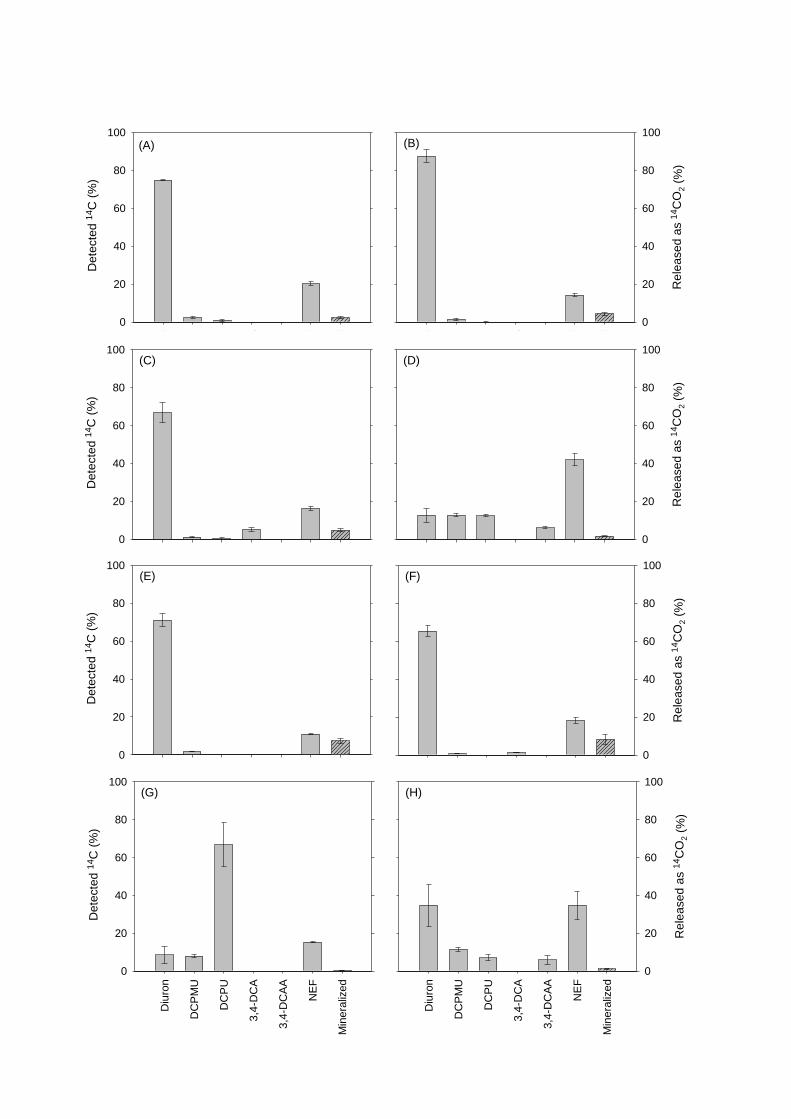
(B)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(A)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(C)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(D)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(F)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(E)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(G)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
3,4
-DC
AA
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(H)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
3,4
-DC
AA
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
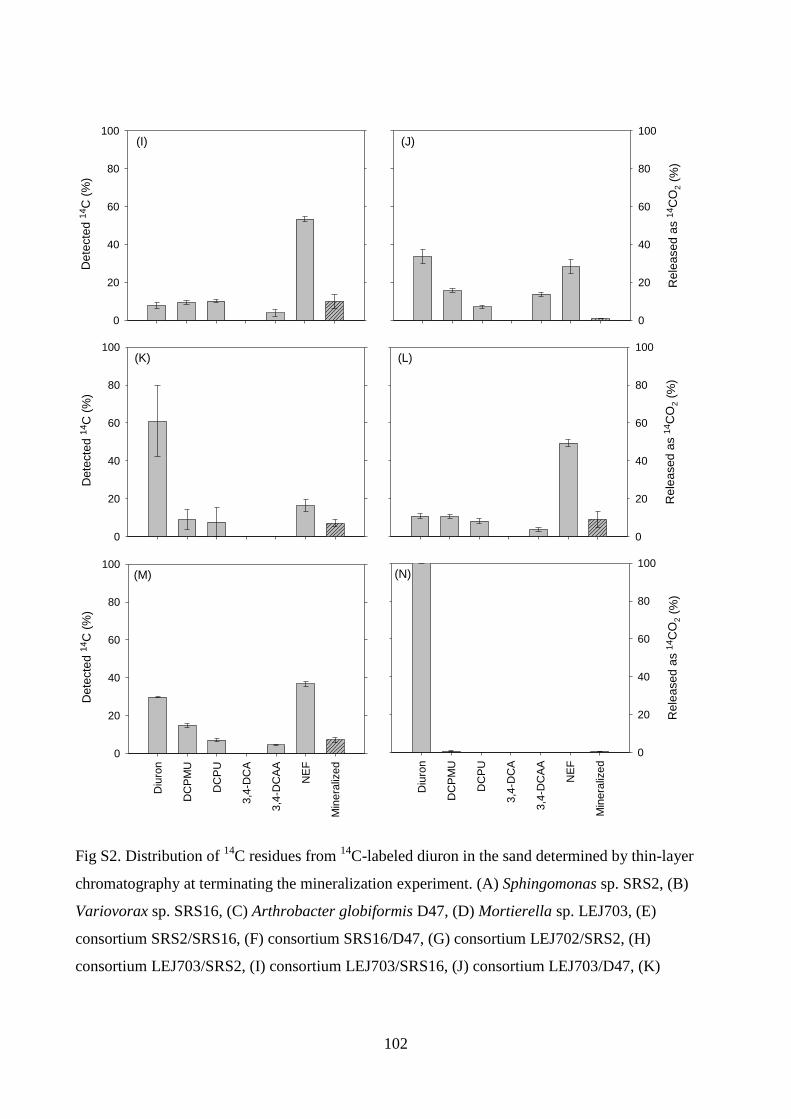
102
Fig S2. Distribution of 14
C residues from 14
C-labeled diuron in the sand determined by thin-layer
chromatography at terminating the mineralization experiment. (A) Sphingomonas sp. SRS2, (B)
Variovorax sp. SRS16, (C) Arthrobacter globiformis D47, (D) Mortierella sp. LEJ703, (E)
consortium SRS2/SRS16, (F) consortium SRS16/D47, (G) consortium LEJ702/SRS2, (H)
consortium LEJ703/SRS2, (I) consortium LEJ703/SRS16, (J) consortium LEJ703/D47, (K)
(I)D
iuro
n
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(J)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(K)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(L)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
UK
NE
F
Min
era
lize
d
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100
(N)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
3,4
-DC
AA
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100(M)
Diu
ron
DC
PM
U
DC
PU
3,4
-DC
A
3,4
-DC
AA
NE
F
Min
era
lized
De
tecte
d 1
4C
(%
)
0
20
40
60
80
100
Re
lea
se
d a
s 1
4C
O2 (
%)
0
20
40
60
80
100

103
consortium LEJ702/SRS2/SRS16, (L) consortium LEJ703/SRS2/SRS16, (M) consortium
LEJ703/SRS16/D47, and (N) abiotic treatment.
DCPMU: [1-(3,4-dichlorophenyl)-3-methylurea], DCPU: [1-(3,4-dichlorophenyl) urea], 3,4-DCA:
[3,4-dichloroaniline], 3,4-DCAA: [3,4-dichloroacetanilide], and NEF: Non-extractable fraction.
Data are presented as means ± SE.
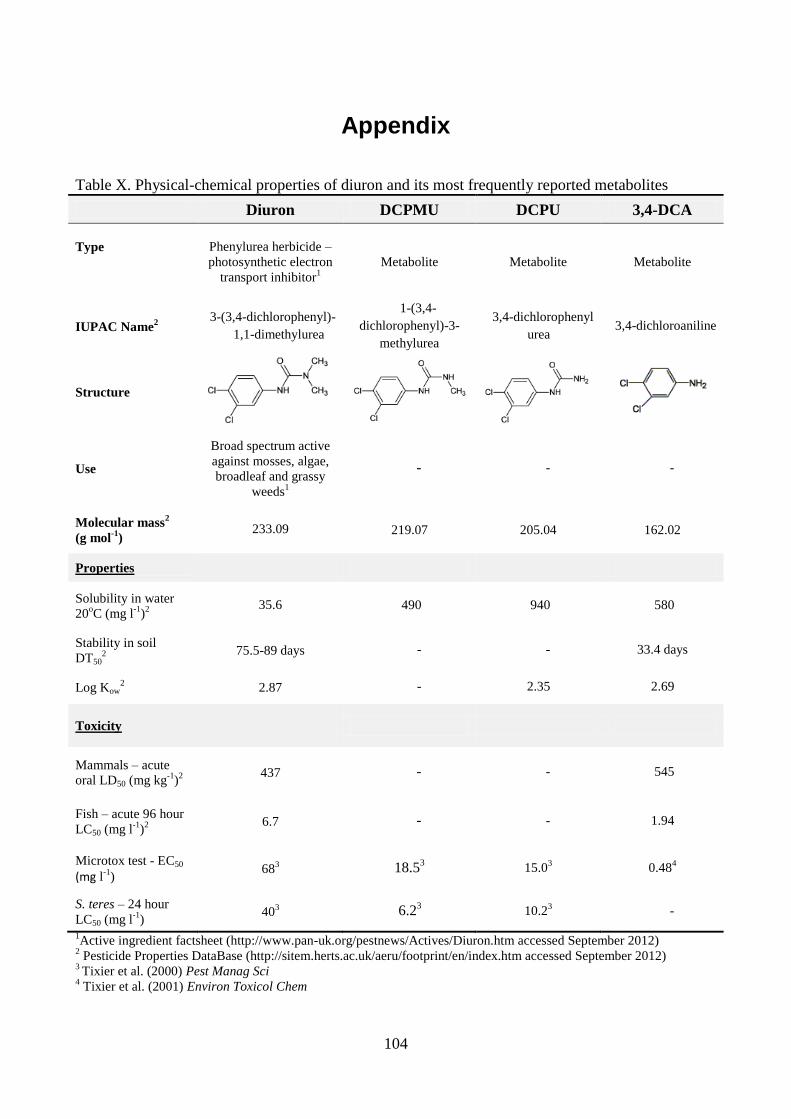
104
Appendix
Table X. Physical-chemical properties of diuron and its most frequently reported metabolites
Diuron
DCPMU DCPU
3,4-DCA
Type
Phenylurea herbicide –
photosynthetic electron
transport inhibitor1
Metabolite Metabolite Metabolite
IUPAC Name2
3-(3,4-dichlorophenyl)-
1,1-dimethylurea
1-(3,4-
dichlorophenyl)-3-
methylurea
3,4-dichlorophenyl
urea 3,4-dichloroaniline
Structure
Use
Broad spectrum active
against mosses, algae,
broadleaf and grassy
weeds1
- - -
Molecular mass2
(g mol-1
) 233.09 219.07 205.04 162.02
Properties
Solubility in water
20oC (mg l
-1)
2
35.6
490 940 580
Stability in soil
DT502 75.5-89 days - - 33.4 days
Log Kow2
2.87 - 2.35 2.69
Toxicity
Mammals – acute
oral LD50 (mg kg-1
)2 437 - - 545
Fish – acute 96 hour
LC50 (mg l-1
)2 6.7 - - 1.94
Microtox test - EC50
(mg l-1
) 68
3 18.53
15.03
0.484
S. teres – 24 hour
LC50 (mg l-1
) 40
3 6.23
10.23
-
1Active ingredient factsheet (http://www.pan-uk.org/pestnews/Actives/Diuron.htm accessed September 2012)
2 Pesticide Properties DataBase (http://sitem.herts.ac.uk/aeru/footprint/en/index.htm accessed September 2012)
3 Tixier et al. (2000) Pest Manag Sci
4 Tixier et al. (2001) Environ Toxicol Chem
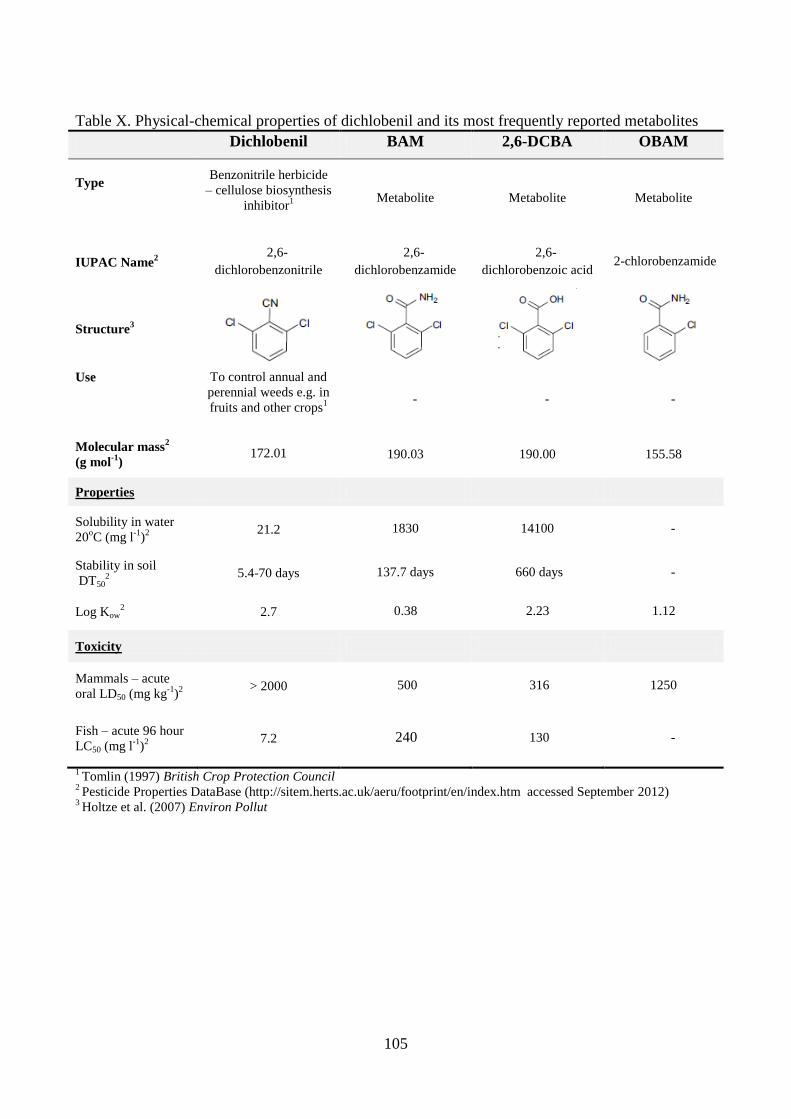
105
Table X. Physical-chemical properties of dichlobenil and its most frequently reported metabolites
Dichlobenil
BAM 2,6-DCBA OBAM
Type
Benzonitrile herbicide
– cellulose biosynthesis
inhibitor1
Metabolite Metabolite Metabolite
IUPAC Name2
2,6-
dichlorobenzonitrile
2,6-
dichlorobenzamide
2,6-
dichlorobenzoic acid 2-chlorobenzamide
Structure3
Use
To control annual and
perennial weeds e.g. in
fruits and other crops1
- - -
Molecular mass2
(g mol-1
) 172.01 190.03 190.00 155.58
Properties
Solubility in water
20oC (mg l
-1)
2 21.2 1830 14100 -
Stability in soil
DT502 5.4-70 days 137.7 days 660 days -
Log Kow2
2.7 0.38 2.23 1.12
Toxicity
Mammals – acute
oral LD50 (mg kg-1
)2 > 2000 500 316 1250
Fish – acute 96 hour
LC50 (mg l-1
)2 7.2 240 130 -
1 Tomlin (1997) British Crop Protection Council
2 Pesticide Properties DataBase (http://sitem.herts.ac.uk/aeru/footprint/en/index.htm accessed September 2012)
3 Holtze et al. (2007) Environ Pollut
Related Documents










