Proc. Nat. Acad. Sci. USA Vol. 71, No. 5, pp. 2033-2037, May 1974 Ferritin-Conjugated Antibodies Used for Labeling of Organelles Involved in the Cellular Synthesis and Transport of Procollagen (ferritin-antibody conjugates/Golgi vacuoles/protein secretion) BJORN R. OLSEN AND DARWIN J. PROCKOP Department of Biochemistry, The Rutgers Medical School, College of Medicine and Dentistry of New Jersey, Piscataway, N.J. 08854 Communicated by G. E. Palade, February 7, 1974 ABSTRACT An improved procedure s4as used to conjugate ferritin to antibodies specific for the NH2- terminal extensions on the precursor form of collagen known as procollagen. The ferritin-antibody conjugates were then used to determine the localization of procollagen in fibroblasts isolated from chick-embryo tendons. Pro- collagen was found in the cisternae of the endoplasmic reticulum, indicating that the protein passes into this compartment early in its biosynthesis. Specific labeling of large Golgi vacuoles was also observed, suggesting that this compartment is also involved in the secretory process. When cells were incubated with colchicine so that the secretion of procollagen was delayed, there was an increase of large, smooth-surfaced vacuoles in the cells and these vacuoles were labeled with the ferritin-antibody conju- gate. Largely on the basis of classical studies on exocrine cells of the pancreas (1-3), it is generally accepted that proteins syn- thesized for "export" into the extracellular space pass into the cisternae of the endoplasmic reticulum, and that from there they are transported through Golgi vesicles to large Golgi vacuoles before secretion via secretory vesicles or gran- ules. Some experimental evidence, largely morphological (4-9), has suggested that this general scheme also applies to the synthesis and secretion of the precursor form of collagen known as procollagen. However, it has been difficult to obtain definitive proof for this hypothesis. For example, autoradio- graphic studies with different types of cells synthesizing pro- collagen have raised the possibility that some or all of the protein passes into the cytoplasm and is secreted through the plasma membrane (10-12). Alternatively, it has been sug- gested that procollagen may pass directly from the cisternae of the endoplasmic reticulum into the extracellular space (5). We have recently prepared and purified specific antibodies (13) to the NH2-terminal extensions on the procollagen synthesized and secreted by chick-embryo fibroblasts. We here report on the use of these antibodies, together with an improved procedure for preparation of ferritin-antibody con- jugates, to demonstrate the localization of procollagen in fibroblasts. MATERIALS AND METHODS Preparation and Purification of Antibodies to Procollagen. Procollagen was prepared from matrix-free fibroblasts (14) and was used to immunize a rabbit (13). Specific antibodies to the NH2-terminal extensions were then purified with an im- munoadsorbent gel containing the isolated, disulfide-linked NH2-terminal extensions of procollagen. The gel was mixed with an equal amount of Sephadex G-25 (Pharmacia; fine grade) and poured into a column (0.9 X 5 cm) that was equilibrated at 40 with 0.1 M sodium phosphate buffer, pH 7.3. Three ml of antisera absorbed with 3 ml of fetal-calf serum were passed through the column. The column was washed with several volumes of 0.1 M sodium phosphate buffer and the antibody was eluted with 3 M KSCN. The eluted antibody was desalted on a Sephadex G-25 column equilibrated at 40 with 0.1 M sodium phosphate buffer, pH 7.3. Conjugation of the Purified Antibodies with Ferritin. Ferritin was activated with glutaraldehyde (15) by mixing 4.3 mg of the activated ferritin with 0.8 mg of the specific antibody and 0.3 mg of nonimmune IgG that had been previously labeled with ["25I]iodine (16). The mixture, in a final volume of 14.8 ml of 0.1 M sodium phosphate buffer, pH 7.3, was con- centrated at 40 to 2.5 ml with an Amicon PM-30 filter. The concentrate was allowed to stand at 4° for 100 hr, and the con- jugate was isolated by gel filtration on an 6% agarose column (Bio-Gel A-5m; 200-400 mesh) (1.5 X 80 cm) that was equili- brated and eluted with 0.1 M Tris * HCl buffer, pH 7.5. Preparation of Fibroblasts for Electron Microscopy. Fibro- blasts were isolated from tendons of 17-day-old chick em- bryos by digestion with collagenase and trypsin, and the cells were incubated for 3 hr at 370 in modified Krebs medium containing 10% fetal-calf serum (14). The cells were. fixed for E o 0.4 0 wu 0.3 0 m ir °o 0.2 IDI 1000 ,_ .n CY 2 a. 20 30 40 50 60 70 80 90 100 110 120 FRACTION NUMBER FIG. 1. Gel filtration of ferritin conjugated to antibodies specific for the NH2-terminal extensions on procollagen. Acti- vated ferritin was reacted with antibodies to procollagen and gel filtration was carried out as described in text. The fraction size was 1.3 ml, and the void volume was about 50 ml. Symbols: Elution of ferritin as assayed by absorbance at 410 nm (O O); elution of IgG (@--*) as assayed by 125I in carrier IgG added to the specific antibody (see text). 2033

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript
Proc. Nat. Acad. Sci. USAVol. 71, No. 5, pp. 2033-2037, May 1974
Ferritin-Conjugated Antibodies Used for Labeling of Organelles Involvedin the Cellular Synthesis and Transport of Procollagen
(ferritin-antibody conjugates/Golgi vacuoles/protein secretion)
BJORN R. OLSEN AND DARWIN J. PROCKOP
Department of Biochemistry, The Rutgers Medical School, College of Medicine and Dentistry of New Jersey, Piscataway, N.J. 08854
Communicated by G. E. Palade, February 7, 1974
ABSTRACT An improved procedure s4as used toconjugate ferritin to antibodies specific for the NH2-terminal extensions on the precursor form of collagenknown as procollagen. The ferritin-antibody conjugateswere then used to determine the localization of procollagenin fibroblasts isolated from chick-embryo tendons. Pro-collagen was found in the cisternae of the endoplasmicreticulum, indicating that the protein passes into thiscompartment early in its biosynthesis. Specific labeling oflarge Golgi vacuoles was also observed, suggesting thatthis compartment is also involved in the secretory process.When cells were incubated with colchicine so that thesecretion of procollagen was delayed, there was an increaseof large, smooth-surfaced vacuoles in the cells and thesevacuoles were labeled with the ferritin-antibody conju-gate.
Largely on the basis of classical studies on exocrine cells of thepancreas (1-3), it is generally accepted that proteins syn-thesized for "export" into the extracellular space pass intothe cisternae of the endoplasmic reticulum, and that fromthere they are transported through Golgi vesicles to largeGolgi vacuoles before secretion via secretory vesicles or gran-ules. Some experimental evidence, largely morphological(4-9), has suggested that this general scheme also applies tothe synthesis and secretion of the precursor form of collagenknown as procollagen. However, it has been difficult to obtaindefinitive proof for this hypothesis. For example, autoradio-graphic studies with different types of cells synthesizing pro-collagen have raised the possibility that some or all of theprotein passes into the cytoplasm and is secreted through theplasma membrane (10-12). Alternatively, it has been sug-gested that procollagen may pass directly from the cisternaeof the endoplasmic reticulum into the extracellular space (5).We have recently prepared and purified specific antibodies
(13) to the NH2-terminal extensions on the procollagensynthesized and secreted by chick-embryo fibroblasts. Wehere report on the use of these antibodies, together with animproved procedure for preparation of ferritin-antibody con-jugates, to demonstrate the localization of procollagen infibroblasts.
MATERIALS AND METHODS
Preparation and Purification of Antibodies to Procollagen.Procollagen was prepared from matrix-free fibroblasts (14)and was used to immunize a rabbit (13). Specific antibodies tothe NH2-terminal extensions were then purified with an im-munoadsorbent gel containing the isolated, disulfide-linkedNH2-terminal extensions of procollagen. The gel was mixedwith an equal amount of Sephadex G-25 (Pharmacia; fine
grade) and poured into a column (0.9 X 5 cm) that wasequilibrated at 40 with 0.1 M sodium phosphate buffer, pH7.3. Three ml of antisera absorbed with 3 ml of fetal-calf serumwere passed through the column. The column was washedwith several volumes of 0.1 M sodium phosphate buffer andthe antibody was eluted with 3 M KSCN. The eluted antibodywas desalted on a Sephadex G-25 column equilibrated at 40with 0.1 M sodium phosphate buffer, pH 7.3.
Conjugation of the Purified Antibodies with Ferritin. Ferritinwas activated with glutaraldehyde (15) by mixing 4.3 mg ofthe activated ferritin with 0.8 mg of the specific antibodyand 0.3 mg of nonimmune IgG that had been previouslylabeled with ["25I]iodine (16). The mixture, in a final volumeof 14.8 ml of 0.1 M sodium phosphate buffer, pH 7.3, was con-centrated at 40 to 2.5 ml with an Amicon PM-30 filter. Theconcentrate was allowed to stand at 4° for 100 hr, and the con-jugate was isolated by gel filtration on an 6% agarose column(Bio-Gel A-5m; 200-400 mesh) (1.5 X 80 cm) that was equili-brated and eluted with 0.1 M Tris * HCl buffer, pH 7.5.
Preparation of Fibroblasts for Electron Microscopy. Fibro-blasts were isolated from tendons of 17-day-old chick em-bryos by digestion with collagenase and trypsin, and the cellswere incubated for 3 hr at 370 in modified Krebs mediumcontaining 10% fetal-calf serum (14). The cells were. fixed for
E
o 0.40
wu 0.30
mir°o 0.2IDI
1000 ,_.nCY
2a.
20 30 40 50 60 70 80 90 100 110 120FRACTION NUMBER
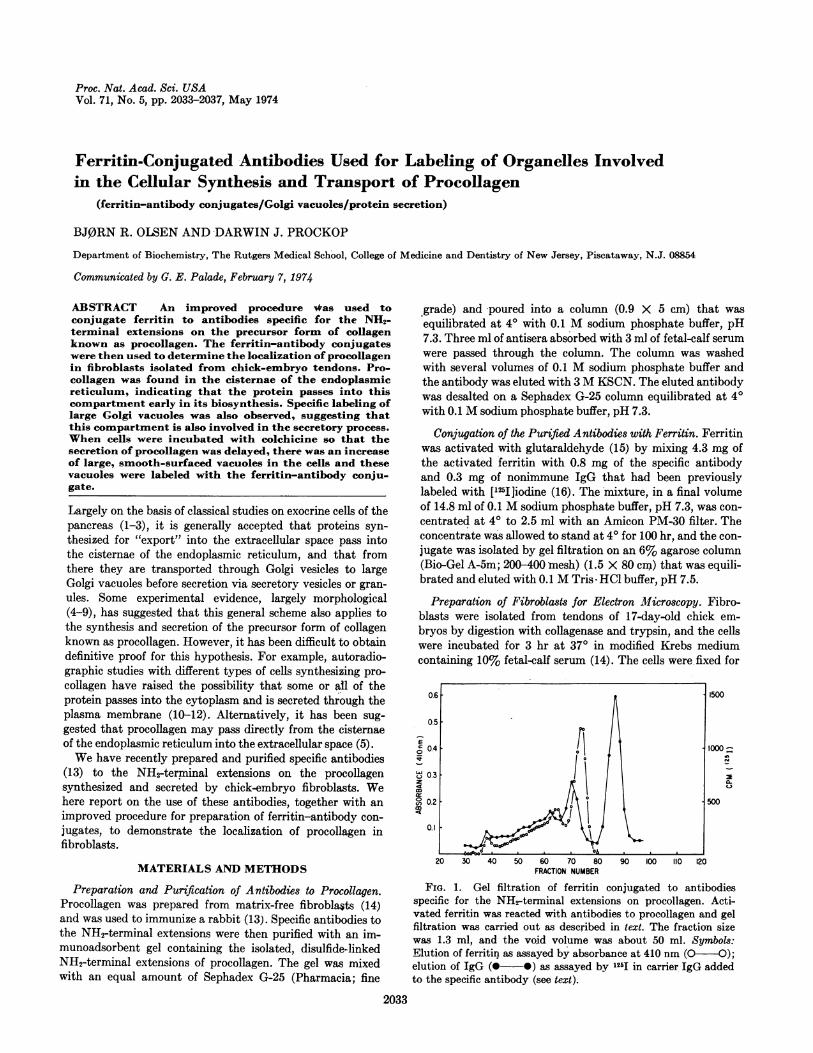
FIG. 1. Gel filtration of ferritin conjugated to antibodiesspecific for the NH2-terminal extensions on procollagen. Acti-vated ferritin was reacted with antibodies to procollagen and gelfiltration was carried out as described in text. The fraction sizewas 1.3 ml, and the void volume was about 50 ml. Symbols:Elution of ferritin as assayed by absorbance at 410 nm (O O);elution of IgG (@--*) as assayed by 125I in carrier IgG addedto the specific antibody (see text).
2033
2034 Cell Biology: Olsen and Prockop
St~~~~~~t
4~~~ ~ ~ ~ ~~~~~~~r
t v .Ii 49
I=
<<;g X > r x. j-g , ;Xs _,b
491z-
Proc. Nat. Acad. Sci. USA 71 (1974)

Synthesis and Transport of Procollagen 2035
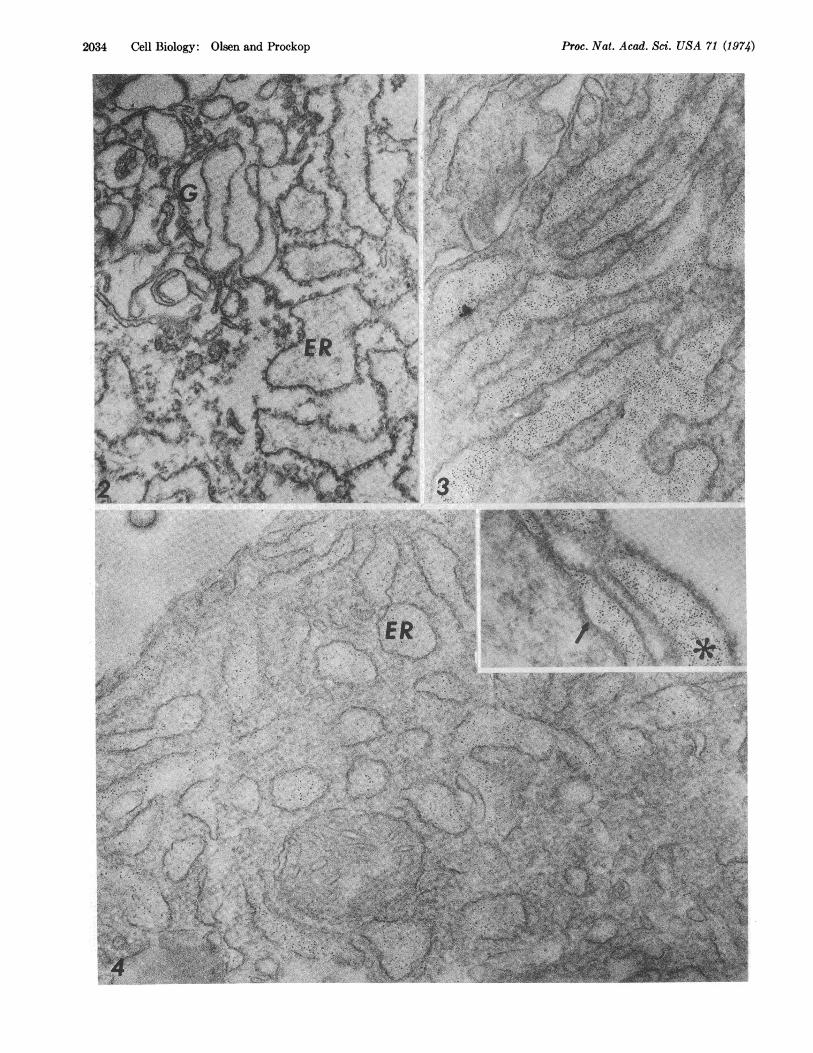
FIG. 2. Electron micrograph from control experiments in which cell fragments fixed for 3 hr with 1% formaldehyde were incubatedwith ferritin conjugated to nonimmune IgG. Washing of the cell fragments with phosphate buffer removed essentially all of the ferritinconjugates so that there was essentially no ferritin over the Golgi complex (G) or endoplasmic reticulum (ER). Magnification X 62,500.
FIG. 3. Electron micrograph of cell fragments that were fixed for 3 hr with 1% formaldehyde and then incubated with ferritin con-jugated to antibodies to procollagen. Heavy labeling of the endoplasmic reticulum is seen. Magnification X50,000.
FIG. 4. Electron micrograph of cell fragments that were fixed for 15 hr with 1% formaldehyde and then incubated with ferritin conju-gated to antibodies to procollagen. Because of the longer fixation time, the cell morphology was relatively well preserved, but the frag-ments were apparently less penetrable by the ferritin-conjugate. However, the specific labeling of the cisternae of the endoplasmic reticu-lum (ER) is still apparent. Inset: Labeling of perinuclear space (arrow) as well as endoplasmic reticulum (asterisk) in the same cell frag-ments. Magnification X62,500.
3 hr or 15 hr at 40 with 1% formaldehyde in 60 mM sodiumphosphate buffer, pH 7.3, containing 0.14 M sucrose. Aftercentrifugation at 600 X g for 6 min, the cells were resuspendedin 0.1 M sodium phosphate buffer and stored at 40 for 15-24hr. The fixed-cells were then homogenized with from 50 to 100strokes in a Teflon and glass homogenizer (15). The cell frag-ments were sedimented by centrifugation at 20,000 X g for10 min and the pellet was incubated with ferritin-conjugatedantibodies to procollagen for 24-48 hr at 4°. Fragments from50 to 100 million cells were incubated with 100 ,l of 0.1 MTris- HCl, pH 7.4, containing the ferritin-labeled specificantibodies (1 mg of ferritin and 0.3 mg of antibody per ml).The cell fragments were washed twice with 5 ml of 0.1 Msodium phosphate bufferj pH 7.3; fixed at 40 for 1 hr with 3%glutaraldehyde in 60 mM sodium phosphate buffer, pH 7.3,containing 0.14 M sucrose; post-fixed at 4° for 1 hr with 1%osmium tetroxide in 60 mM sodium phosphate buffer, pH 7.3,containing 0.16 M sucrose; block-stained with 0.5% mag-nesium uranyl acetate for 30 min at room temperature;dehydrated in ethanol; and embedded in Araldite (Taab).Ultra-thin sections were examined in a JEM-1OOB electronmicroscope.
RESULTS
Properties of the Ferritin-Conjugated Antibodies. The ferritin-conjugate of the purified antibodies was largely free of higherpolymers of antibody, and gel filtration of the conjugate(Fig. 1) made it possible to select fractions (fractions 56-75)that were free both of unconjugated IgG and of polymericferritin. A large fraction of the ferritin was linked to the spe-cific antibodies, since the molar ratio of ferritin to '25I-labeledIgG in the peak fractions (fractions 68-70 in Fig. 1) was about0.7. The amount of free ferritin was relatively small and it wasreadily removed from the cell fragments by washing withphosphate buffer (see below).
Distribution of Ferritin-Conjugated Antibodies to Procollagenin Fibroblasts. A series of conditions was examined to estab-lish an optimal method for preparing cell fragments. Fibro-blasts were incubated with ['4C]proline for 3 hr so that theintracellular procollagen contained [14C]hydroxyproline (14).The medium was removed by centrifugation (14), and thecells were fixed in 1% formaldehyde for 1, 3, or 15 hr; Assaysfor nondialyzable ['4C]hydroxyproline indicated that about80% of the initial [14C]procollagen was recovered in the cellfragments regardless of the fixation time. However, if the fixa-tion was carried out for only 1 hr, there was a heavy back-ground of the ferritin-conjugated antibody around the cellfragments as well as within them. This observation was prob-ably explained by diffusion of the procollagen out of cellcompartments because of insufficient crosslinking of the pro-
teins in the fragments. If the fixation with 1% formaldehydewas extended to 3 or 15 hr, this background staining wasalmost completely eliminated. To test the effect of the fixa-tion on antigenicity of procollagen, the cells were fixed with1% formaldehyde for various times and the cells were rupturedby N2 cavitation (18) to make the procollagen available forreaction with antibody. Assay of the fragments by hemagglu-tination inhibition with erythrocytes coated with isolateddisulfide-linked NH2-terminal extension from procollagen (13)indicated that fixation with 1% formaldehyde for up to 15 hrhad no detectable effect on the antigenicity of the procollagen.
Further control experiments were carried out with ferritinconjugated to nonimmune IgG from rabbits. When the cellfragments were incubated with nonimmune conjugate and notwashed, some ferritin was seen in all cellular organelles suchas nuclei, mitochondria, cisternae of the endoplasmic retic-ulum, and Golgi vacuoles. The results indicated, therefore,that the conjugate penetrated into these compartments.However, essentially all of this nonimmune conjugate was re-moved from the fragments by washing with phosphate bufferas described above (Fig. 2).
In contrast, the ferritin conjugated to purified antibodiesto procollagen specifically labeled the cisternae of the endo-plasmic reticulum (Figs. 3 and 4) and some of the largeGolgi vacuoles (Fig. 5). No significant labeling was seen in thecytoplasmic matrix except for a few sites at which there wasdiscontinuity of the membrane of the endoplasmic reticulum.Also, no specific labeling was seen over the ribosomes free inthe cytoplasm or over ribosomes attached to the membranesof the endoplasmic reticulum (Fgis. 3 and 4). The distributionof label in the cisternae of the endoplasmic reticulum wasrelatively uniform and was similar to that previously seen insimilar cell fragments with ferritin-conjugates of antibodies toprolyl hydroxylase (15). Of special interest was the findingthat ferritin-conjugates were present in the perinuclear space(Fig. 4, inset). Some large Golgi vacuoles were clearly labeledwith the ferritin-conjugate (Fig. 5) but it was difficult todemonstrate specific labeling of the small vesicles within theGolgi complex. Incubating the cells with 1 AM colchicine for3 hr greatly increased the number and size of the Golgivacuoles (Figs. 6 and 7). The large vacuoles in the cells treatedwith colchicine were extensively labeled with the ferritin-conjugated antiprocollagen (Fig. 6).
DISCUSSIONThe purified antibodies used here are specific for the NH2-terminal extensions on procollagen containing interchaindisulfide bonds or on individual pro-a chains in which the ex-tensions contain intrachain disulfide bonds (13). The anti-body does not react with pro-a chains if these are completelyreduced and alkylated. The latter observation suggests that
Proc. Nat. Acad. Sci. USA 71 (1974)

2036 Cell Biology: Olsen and Prockop
- -.~~~~~~~~~~~~~~~..~~~. .-
.7?1~~~~~~~~~~~-
EUjk.-.
wfA
Proc. Nat. Acad. Sci. -USA 71 (1974)

Synthesis and Transport of Procollagen 2037
FIG. 5. Electron micrograph of cell frag nts prepareas4,described in the legend of Fig. 4. Some labeling of large Golgi vacuoles(arrows) is apparent. Magnification X62,500.
FIG. 6 Electron micrograph from an experiment in which cells were incubated with 1 ,uM1I colchicine for 3 hr at 370, fixed with 1%formaldehyde for 3 hr, and then incubated with ferritin-antibody conjugates as in Figs. 4 and 5. The treatment with colchicine has pro-duced distended, smooth-surfaced vacuoles (see Fig. 7) which are heavily labeled with ferritin (arrows). Magnification X62,500.
FIG. 7. Electron micrograph of tendon fibroblasts incubated with 1 1AM colchicine at 370 for 3 hr. The cells were fixed with 3% glu-taraldehyde for 1 hr and post-fixed with 1% osmium tetroxide as described in text. The section was stained with 1% uranyl acetate in70% methanol. Note the presence of large, smooth-surfaced vacuoles (V) similar to those seen in the cell fragments (see Fig. 6). Magnifi-cation X25,000.
the antibody would not react with nascent forms of thepolypeptides in which the intrachain disulfide bonds had notyet been synthesized, but they would react with single pro-achains containing intrachain disulfide bonds before such chainsare linked together with interchain bonds. The antibody alsomight react with NHrterminal extensions released fromprocollagen by proteolytic degradation of the protein if suchproteolysis released the extensions in a largely intact form.However, previous studies with the cells used here indicatethat this does not present a problem in interpreting the presentdata. In particular, pulse-label and chase experiments with thecells have shown that there is little if any intracellular deg-radation of the procollagen which would lead to release ofpeptides containing intact antigenic determinants (14, 20).Therefore, most of the antigen in the cells reacting with theferritin-antibody conjugates must consist of either procollagenor of pro-a chains in which the NHrterminal extensions are in-tact and contain intrachain disulfide bonds.The results demonstrated specific labeling of the cisternae
of the endoplasmic reticulum and no significant labeling of thecytoplasmic matrix or any other cellular compartment. Theseobservations conclusively establish that the newly synthesizedpolypeptide chains of procollagen pass into the cisternae dur-ing or after their assembly on ribosomes. Ferritin-antibodyconjugates also labeled the perinuclear space, indicating thatsome procollagen passes into this compartment early in itsbiosynthesis. The presence of procollagen in the cisternae ofthe endoplasmic reticulum agrees with the recent demonstra-tion (15) that prolyl hydroxylase, the enzyme that syn-thesizes the hydrox-yproline in collagen by the hydroxylationof peptide-bound proline, is also found in the cisternae of theendoplasmic reticulum. Labeling with ferritin-antibody con-jugates was also seen over the large Golgi vacuoles, suggestingthat these cellular organelles participate in the secretoryprocess. Specific labeling was not seen over the small vesiclesin the Golgi complex, an observation that might be explainedby incomplete penetration of these vesicles whose diameter(about 500 A) is not much larger than the anticipated size ofthe ferritin-antibody conjugates (200-300 A). It is also possi-ble, however, that these vesicles, which are smaller than thelargest dimension of the triple helix of procollagen (about3000 A), do not participate in the secretion of this protein inthese cells.The data presented here did not conclusively establish that
all the procollagen in the cells was secreted through the Golgivacuoles since it was not possible to carry out kinetic studieson the transfer of protein from one compartment to another.
However, the observations suggesting that secretion involvedthe large Golgi vacuoles were further supported by the experi-ments with colchicine. Previous studies demonstrated thattreatment of the cells with colchicine delaved the secretion ofprocollagen without affecting its rate of synthesis and thereforeled to an intracellular accumulation of the protein (14, 21,22). The results presented here showed that this intracellularaccumulation was accompanied by a distension of the Golgivacuoles and by an increase in the amount of procollagen insuch vacuoles.
This work was supported in part by N.I.H. Grant AM-16,516from the U.S. Public Health Service. We gratefully acknowledgethe expert technical assistance of Mrs. Nancy Doerr.
1. Palade, G. E., Siekevitz, P. & Caro, L. G. (1961) in CibaFoundation Symposium on the Exocrine Pancreas, eds. deReuck, A. V. S. & Cameron, M. P. (J. & A. Churchill Ltd.,London), pp. 23-49.
2. Jamieson, J. D. & Palade, G. E. (1967) J. Cell Biol. 34, 577-596.
3. Jamieson, J. D. & Palade, G. E. (1971) J. Cell Biol. 50,135-158.
4. Revel, J. P. & Hay, E. D. (1963) Z. Zellforsch. Mikroskop.Anat. 61, 110-144.
5. Ross, R. & Renditt, E. P. (1965) J. Cell Biol. 27, 83-106.6. Trelstad, R. L. (1971) J. Cell Biol. 48, 689-694.7. Weinstock, M. (1972) Z. Zellforsch. Mikroskop. Anat. 129,
455-470.8. Coulombre, A. J. & Coulombre, J. L. (1972) Develop. Biol.
28, 183-190.9. Hay, E. D. & Dodson, J. W. (1973) J. Cell Biol. 57, 190-213.
10. Salpeter, M. M. (1968) J. AMorphol. 124, 387-421.11. Cooper, G. W. & Prockop, D. J. (1968) J. Cell Biol. 38,
523-537.12. Reith, E. J. (1968) J. Ultrastruct. Res. 21, 383-414.13. Dehm, P., Olsen, B. R. & Prockop, D. J. (1974) Eur. J.
Biochem., in press.14. Dehm, P. & Prockop, D. J. (1972) Biochim. Biophys. Acta
264, 375-382.15. Olsen, B. R., Berg, R. A., Kishida, Y. & Prockop, D. J.
(1973) Science 182, 825-827.16. Marchalonis, J. J. (1969) Biochem. J. 113, 299-305.17. Kraehenbuhl, J. P. & Jamieson, J. D. (1972) Proc. Nat.
Acad. Sci. USA 69, 1771-1775.18. Hunter, M. J. & Commerford, S. L. (1961) Biochim. Biophys.
Acta 47, 580-586.19. Schofield, J. D. & Prockop, D. J. (1973) Clin. Orthop. Relat.
Res. 97, 175-195.20. Schofield, J. D., Uitto, J. & Prockop, D. J. (1974) Biochem-
istry, in press.21. Ehrlich, H. P. & Bornstein, P. (1972) Nature New Biol. 238,
257-260.22. Diegelmann, R. F. & Peterkofsky, B. (1972) Proc. Nat.
Acad. Sci. USA 69, 892-896.
Proc. Nat. Acad. Sci. USA 71 (1974)
Related Documents











