University of Kentucky University of Kentucky UKnowledge UKnowledge Theses and Dissertations--Animal and Food Sciences Animal and Food Sciences 2016 Fatty Acid Profile In Ruminal Content And Blood Plasma Of Fatty Acid Profile In Ruminal Content And Blood Plasma Of Finishing Beef Cattle, Supplemented With Different Sources Of Fat Finishing Beef Cattle, Supplemented With Different Sources Of Fat Amanda Pesqueira University of Kentucky, [email protected] Digital Object Identifier: http://dx.doi.org/10.13023/ETD.2016.077 Right click to open a feedback form in a new tab to let us know how this document benefits you. Right click to open a feedback form in a new tab to let us know how this document benefits you. Recommended Citation Recommended Citation Pesqueira, Amanda, "Fatty Acid Profile In Ruminal Content And Blood Plasma Of Finishing Beef Cattle, Supplemented With Different Sources Of Fat" (2016). Theses and Dissertations--Animal and Food Sciences. 62. https://uknowledge.uky.edu/animalsci_etds/62 This Master's Thesis is brought to you for free and open access by the Animal and Food Sciences at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Animal and Food Sciences by an authorized administrator of UKnowledge. For more information, please contact [email protected].

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

University of Kentucky University of Kentucky
UKnowledge UKnowledge
Theses and Dissertations--Animal and Food Sciences Animal and Food Sciences
2016
Fatty Acid Profile In Ruminal Content And Blood Plasma Of Fatty Acid Profile In Ruminal Content And Blood Plasma Of
Finishing Beef Cattle, Supplemented With Different Sources Of Fat Finishing Beef Cattle, Supplemented With Different Sources Of Fat
Amanda Pesqueira University of Kentucky, [email protected] Digital Object Identifier: http://dx.doi.org/10.13023/ETD.2016.077
Right click to open a feedback form in a new tab to let us know how this document benefits you. Right click to open a feedback form in a new tab to let us know how this document benefits you.
Recommended Citation Recommended Citation Pesqueira, Amanda, "Fatty Acid Profile In Ruminal Content And Blood Plasma Of Finishing Beef Cattle, Supplemented With Different Sources Of Fat" (2016). Theses and Dissertations--Animal and Food Sciences. 62. https://uknowledge.uky.edu/animalsci_etds/62
This Master's Thesis is brought to you for free and open access by the Animal and Food Sciences at UKnowledge. It has been accepted for inclusion in Theses and Dissertations--Animal and Food Sciences by an authorized administrator of UKnowledge. For more information, please contact [email protected].

STUDENT AGREEMENT: STUDENT AGREEMENT:
I represent that my thesis or dissertation and abstract are my original work. Proper attribution
has been given to all outside sources. I understand that I am solely responsible for obtaining
any needed copyright permissions. I have obtained needed written permission statement(s)
from the owner(s) of each third-party copyrighted matter to be included in my work, allowing
electronic distribution (if such use is not permitted by the fair use doctrine) which will be
submitted to UKnowledge as Additional File.
I hereby grant to The University of Kentucky and its agents the irrevocable, non-exclusive, and
royalty-free license to archive and make accessible my work in whole or in part in all forms of
media, now or hereafter known. I agree that the document mentioned above may be made
available immediately for worldwide access unless an embargo applies.
I retain all other ownership rights to the copyright of my work. I also retain the right to use in
future works (such as articles or books) all or part of my work. I understand that I am free to
register the copyright to my work.
REVIEW, APPROVAL AND ACCEPTANCE REVIEW, APPROVAL AND ACCEPTANCE
The document mentioned above has been reviewed and accepted by the student’s advisor, on
behalf of the advisory committee, and by the Director of Graduate Studies (DGS), on behalf of
the program; we verify that this is the final, approved version of the student’s thesis including all
changes required by the advisory committee. The undersigned agree to abide by the statements
above.
Amanda Pesqueira, Student
Dr. David L. Harmon, Major Professor
Dr. David L. Harmon, Director of Graduate Studies

FATTY ACID PROFILE IN RUMINAL CONTENT AND BLOOD PLASMA OF FINISHING BEEF CATTLE, SUPPLEMENTED WITH DIFFERENT SOURCES OF
FAT
THESIS
A thesis submitted in partial fulfillment of the requirements for the degree of Master of Science in the College of Agriculture, Food, and Environment at the University of
Kentucky
By
Amanda Pesqueira
Lexington, Kentucky
Director: Dr. David L. Harmon, Professor of Animal Science
Lexington, Kentucky
2016
Copyright © Amanda Pesqueira 2016

ABSTRACT OF THESIS
FATTY ACID PROFILE IN RUMINAL CONTENT AND BLOOD PLASMA OF
FINISHING BEEF CATTLE, SUPPLEMENTED WITH DIFFERENT SOURCES OF FAT
Eight steers were used in a replicated 4 x 4 Latin square design experiment to study the rumen metabolism of polyunsaturated fatty acids. The animals were fed a concentrate diet based on cracked corn grain, corn silage and hay fescue (K31). The dietary fat treatments were a negative control, tallow added at 60g/day, fish oil at 60g/day and heterotrophically grown microalgae high in DHA at 100g/day. Treatments were incorporated immediately prior to feeding each day and dosed through a ruminal fistula mixed with 0.45kg of the diet. Animals were fed at 1.75 x NEm once daily during adaptation then switched to twice daily during sample collection. Samples rumen, reticulum and omasal contents were used to determine ruminal digestion and metabolism of fatty acids while urine and feces were collected to determine diet digestibility and N balance. The experiment was analyzed as a Latin Square design with a 2 x 2 factorial using mixed models in SAS. There was no difference among treatments for DMI, urine or fecal excretion, N balance, total VFA concentrations, omasal or reticular flow, and apparent digestibility. Fat treatments had lower blood plasma C18:0 than control. Algae increased flow of C18:1 isomers and increased DHA in plasma.
KEYWORDS: Algae, Fat, Omasum, Polyunsaturated fatty acid, DHA
Amanda Pesqueira Student’s Signature
May 1st, 2016 Date

FATTY ACID PROFILE IN RUMINAL CONTENT AND BLOOD PLASMA OF FINISHING BEEF CATTLE, SUPPLEMENTED WITH DIFFERENT SOURCES OF
FAT
By Amanda Pesqueira
Dr. David L. Harmon Director of Thesis
Dr. David L. Harmon Director of Graduate Studies
May 1st, 2016 Date

iii
ACKNOWLEDGMENTS
I am grateful for the support and inspiration of my parents, Eliana Ionara de Oliveira
Pesqueira and Dalton dos Santos Pesqueira, without them I would not be the person I am
today. Also, my sister Juliana Pesqueira, for being my role model and always being
present in my life.
I would like to thanks my advisor Dr. David L. Harmon for believing in me and offering
me this opportunity. Thank you for pushing me to achieve my goals and work to my full
potential during this journey. His advice, guidance and input were absolutely essential
during the years seeking this degree.
Also, thank you to Dr. James L. Klotz, Dr. Kyle McLeod and Dr. Amanda Gehman for
being part of my committee and providing input.
Thank you to Lauren Clark, Winston Lin and Kirk Vanzant for providing support during
the research trial and laboratorial analysis at the University of Kentucky.
I am thankful for my good friend Suelen Capa de Avila for keeping me sane during the
intense sample collection times at the C. Oran Little Research Center Beef Unit and to
my animals used during this trial Snow, Buddy, Buggy, Little John, Rocket, Jolly,
Dummy, and Rumple.

iv
TABLE OF CONTENTS
ACKNOWLEDGMENTS ................................................................................................ iii LIST OF TABLES ..............................................................................................................v LIST OF FIGURES ...........................................................................................................vi Chapter One: Introduction ................................................................................................. 1 Chapter Two: Literature Review ....................................................................................... 5
Fat Supplementation ........................................................................................................5 Fat metabolism and biohydrogenation .......................................................................5 Exogenous fat ...……………………………………………………………..............8 Tallow ...……….........................................................................................................9 Fish Oil ...……………………...…………………………………………………..10
Marine Algae...…….......................................................................................................12 Omasum Sampling .........................................................................................................15 Conclusion .....................................................................................................................15 Objective ………………………………………………………………………………16
Chapter Three: Fatty acid profile in ruminal content and blood plasma of finishing beef cattle, supplemented with different sources of fat ..……………………………………..17
Introduction ....................................................................................................................17 Materials and Methods....................................................................................................17
Animals and Experimental Design...........................................................................17 Digestibility and N-balance……………………..…………………………………18 Ruminal Fatty Acid Metabolism………………………………………………..….19 Sample Analysis........................................................................................................20
Calculations…………………………………………………………………………….23 Statistical Analysis..........................................................................................................24 Results.............................................................................................................................25 Discussion ......................................................................................................................28
Chapter Four: Summary and Conclusions.........................................................................35 APPENDICES ………......................................................................................................54 REFERENCES .................................................................................................................57
VITA ………..………...…………………………………………………………………71

v
LIST OF TABLES
Table 1. Composition of high concentrate diet fed to animals with added tallow, fish oil
or high-DHA microalgae ...……………………………………………………….……..36
Table 2. Least square means for dry matter (DM) and organic matter (OM) intake and
digestibility, fecal and urine excretion, fecal fat, and chromium percent recovery in steers
fed a finishing diet with supplemental tallow, fish oil or high-DHA microalgae …….....37
Table 3. Least square means for N intake, fecal and urine excretion, and balance in steers
fed a finishing diet with supplemental tallow, fish oil or high-DHA microalgae …….....38
Table 4. Least square means for rumen volatile fatty acids (VFA) and pH in steers fed a
finishing diet with supplemental tallow, fish oil or high-DHA microalgae .……………39
Table 5. Least square means for omasal flow, apparent digestibility of dry matter (DM)
and apparent digestibility of organic matter (OM) in steers fed a finishing diet with
supplemental tallow, fish oil or high-DHA microalgae .……...........................................41
Table 6. Least square means for starch intake flow and digestibility, in steers fed a
finishing diet with supplemental tallow, fish oil or high-DHA microalgae ...…………..42
Table 7. Least square means for total tract and ruminal long chain fatty acid (LCFA)
digestibility in steers fed a finishing diet with supplemental tallow, fish oil or high-DHA
microalgae .........................................................................................................................43
Table 8. Least square means for omasal long chain fatty acid (LCFA) flow in steers fed a
finishing diet with supplemental tallow, fish oil or high-DHA microalgae .……….…...47
Table 9. Least square means for blood plasma long chain fatty acid (LCFA) flow in steers
fed a finishing diet with supplemental tallow, fish oil or high-DHA microalgae ...…......48

vi
LIST OF FIGURES
Figure 1.Biohydrogenation linoleic and linolenic acid to stearic acid ………………......49
Figure 2. Shift in the biohydrogenation process during the conversion of linoleic acid to
stearic acid ....………………………………………...………………………………….50
Figure 3. Omasum sampling vacuum pump ...…………………………………….…….51
Figure 4. Ruminal dry matter (DM) digestibility using chromium intake (%) or corrected
for fecal chromium recovery in steers fed a finishing diet with supplemental tallow, fish
oil or high-DHA microalgae …………….………………………………………………52
Figure 5ab. (a) Average omasum and reticulum DM flow (g/d) and (b) treatments average
of omasum and reticulum DM flow (g/d) corrected for fecal chromium in steers fed a
finishing diet with supplemental tallow, fish oil or high-DHA microalgae ...…………..53

1
CHAPTER ONE: INTRODUCTION
Because red meat contains a high concentration of saturated fatty acids,
supplementing cattle with a polyunsaturated fatty acid source in the diet could improve
the fatty acid profile of the meat (Duckett and Gillis, 2010). Saturated fat can increase the
risk of coronary heart disease in humans (Vartiainen et al., 2010), increase plasma
cholesterol (Keys et al. 1965) by elevating low-density lipoproteins (LDL) and
cholesterol content in the blood (Bronte-Stewart et al., 1956). High levels of saturated fat
in the diet can stimulate cholesterol synthesis by the liver (Keys et al. 1965), increasing
the production of very low-density lipoproteins (VLDL), that are oxidized in the blood
producing LDL and also by reducing LDL receptor activity in the cell membranes
(Bronte-Stewart et al., 1956). Consuming a diet rich in long-chain n-3 polyunsaturated
fatty acids (PUFA) could improve human health, because consuming a PUFA enriched
diet activates the peroxisome proliferator-activated receptor γ (PPARγ); (Xiao et al.,
2006) which up-regulates uncoupling protein 2 (UCP2). Mitochondrial uncoupling
protein 2 (UCP2) dissipate protons before it can be used in oxidative phosphorylation
pathway, generating heat instead of ATP, limiting the energy production capacity of β-
cells, inhibiting insulin secretion (Fleury et. al., 1997). When UCP2 is over expressed, it
reduces glucose-stimulated insulin secretion (GSIS), this could potentially impact type 2
diabetes, which is caused by insulin resistance when this hormone is produced in excess
by β-cells in the pancreas (Chan et al., 1999).
Mammals are not able to convert omega-6 fatty acid isomers to omega-3 due to
the absence of the enzyme omega-3 desaturase. When the consumption of omega-3
increases the content of omega-6 in the cell membrane is partially replaced (Simopoulos,

2
1991). The omega-6 arachidonic acid produces lipoxins, prostaglandins, leukotrienes,
hydroxy fatty acids and thromboxanes that when produced in large quantities contribute
to the formation of thrombus and atheroma. Tromboxanes are vasoconstrictors produced
during the prostaglandin synthesis from arachidonic acid (C20:4n6) (Simopoulos, 2006).
Diets containing a high ratio of omega-6: omega-3 is related to high rates of
cardiovascular, inflammatory and autoimmune diseases, rheumatoid arthritis, asthma,
depression and cancer. Omega 6 isomers oxidize LDL present in the blood plasma,
leading to cholesterol accumulation (Simopoulos and Cleland, 2003). Meanwhile,
increasing the consumption of omega-3 PUFA decreases the incidence of cardiovascular
diseases because it reduces plaque build-up in the arterial walls (atherosclerosis). This
occurs because when LDL is oxidized by omega 6 isomers, macrophage lysosomes
hydrolyze lipoprotein cholesterol esters, accumulating unesterified cholesterol in its
lysosomal compartment (De Caterina et al., 1994). Blood pressure is significantly
reduced when humans consumed 3g/d of omega-3 fatty acids or more once omega 3
isomers reduce the production of thromboxane vasoconstrictor and increase the
production of prostacyclin vasodilator (Morris et al., 1993; Appel et al., 1993). Also,
omega-3 fatty acids are essential for the composition of brain lipids, retina and sperm
(Neuringer et al., 1988), the development of cortical function in infants (Albert et al.,
1998; Kang and Leaf, 1994; Agren et al., 1997), reduction of rheumatoid arthritis,
ulcerative colitis (Hawthorne et al., 1992), atopic dermatitis and psoriasis (Bittiner et al.,
1988). Consuming omega-3 eicosapentaenoic acid (EPA) and docosahexaenoic acid
(DHA) could be beneficial to human health. These fatty acid isomers prevent
hypertension due to their ability to lower blood pressure by inhibiting the activity of the

3
Δ5desaturase enzyme, which is required for the production of arachidonic acid (Engler,
1996). Also, plasma triglyceride can be reduced 25% to 30% by consuming high doses of
EPA and DHA (4g/d) due to reduction in VLDL synthesis and increase in LDL cell
membrane receptor activity (Harris, 1997). Docosahexaenoic acid is an essential
component of the central nervous system since it is a component of the brain membrane
(Crawford et al., 1997), and also increases brain development in children (Martinetz,
1992). These findings demonstrate that it is very beneficial to increase the consumption
of omega-3 fatty acids, and reduce the dietary intake of n-6 PUFA.
Wahle et al. (2004) suggested that long-chain fatty acids can prevent diseases by
altering the expression of inflammatory cytokines produced by macrophages. Conjugated
linoleic acids (CLA) were also identified as anticancer agents (Pariza et al., 2000),
reducing agents in the production of inflammatory cytokines (Song et al., 2004), and
regulators in vascular homeostasis (Torres-Duarte et al., 2003). Other benefits to
increasing PUFA consumption would be a decrease in body fat deposition by reducing
adipose cell fill and proliferation. Studies have shown that CLA consumption decreases
percent body fat by regulating enzymes involved in fat metabolism (Blankson et al.,
2000). Rats consuming conjugated linoleic acid had body fat decrease in response to an
increase in the activity of palmitoyl carnitine acyl transferase (CPT) which facilitate the
entry of fatty acids through the mitochondrial inner membrane, and a reduction of
lipoprotein lipase which hydrolyze triglycerides present in VLDL (Keim et al., 2003).
These studies demonstrate the importance of decreasing the consumption of
saturated fat and increasing PUFA intake in humans. Human health could benefit if the
fat profile in red meat was improved by increasing PUFA content, red meat has the

4
highest pounds per capita consumption in U.S. according to USDA’s Economic Research
Service.

5
CHAPTER TWO: LITERATURE REVIEW
FAT SUPPLEMENTATION
Fat Metabolism and Biohydrogenation
In ruminants dietary fat is hydrolyzed into free fatty acids (FFA) by microbial and
protozoal lipases, galactosidases and phospholipases in the rumen (Demeyer & Van
Nevel, 1995). The microbial population hydrolyzes the fatty acids from the glycerol
backbone followed by the hydrogenation of the FFA. Unsaturated fats suffer
biohydrogenation by the rumen bacteria converting linoleic acid (C18:2) and linolenic
acid (C18:3) into stearic acid (C18:0) (Figure 1) (Harfoot & Hazlewood, 1988).
Biohydrogenation is the reduction of double bonds in the fat molecule or the adding of
hydrogen atoms by the action of isomerases and reductases (Jenkins, 1993). High levels
of unsaturated fat supplemented in the diet can cause an incomplete biohydrogenation of
linoleic acid, leading to the production of isomers like trans-vaccenic acid (C18:l trans-
11), which will lead to a reduction in stearic acid (C18:0) content (Harfoot et al. 1973).
Microbial biohydrogenation is affected by low ruminal pH (Van Nevel and
Demeyer, 1996) which can be caused by feeding a high concentrate diet where the
greater amounts of nonstructural carbohydrates are rapidly fermented by the microbial
population in the rumen (Kalscheur et al., 1997). This decrease in pH alters the bacterial
profile of the rumen and results in an incomplete biohydrogenation of fatty acids
accumulating trans-C18:1 by decreasing the conversion of it to stearic acid. This
accumulation leads to milk fat depression in lactating cows once it can reduce de novo
synthesis in the mammary gland (Duckett and Gillis, 2010). A diet high in fat can reduce

6
structural carbohydrate digestion due to the shift in volatile fatty acid (VFA) production,
where acetate: propionate (A:P) is shifted due to an increase in propionate and decrease
in acetate and butyrate (Jenkins, 1993). The change in digestion is generated due to the
change in microbial population in the rumen, inhibiting microbial growth and reducing
the protozoa population. This reduction in structural carbohydrate digestion is not
observed when the diet is enriched with sources containing polyunsaturated fatty acids
with 20 to 22 carbons in the structure (Doreau and Chilliard, 1997) such as fish oil and
algae. Nonstructural carbohydrates appear less affected by dietary fat supplementation as
starch digestion was not compromised when cattle received fat supplementation. Fat
coating on feed particles impact is lower in nonstructural carbohydrates, and bacteria can
attach more efficiently, allowing microbial membrane lipase action (Bock et al., 1991).
Fat sources containing high levels of polyunsaturated fatty acids cause
disturbances in the rumen environment due to the toxic effect of this isomer to the
microbial population. Bacterial lipid membrane content is about 90% saturated fat, so
unsaturated fats can alter its membrane fluidity, reducing microbial lipase action,
possibly leading to a reduction in digestibility and consequently in dry matter intake
(DMI) (Jenkins, 1993). Animals receiving marine algae supplementation through a rumen
fistula had lower dry matter intake when compared to control animals, which can also
affect milk yield (Boeckaert et al. 2008). High contents of unsaturated fatty acids in the
diet can generate a shift in the biohydrogenation process converting linoleic acid (C18:2)
to C18:2 trans-10, cis-12, inducing milk fat depression caused by the reduction of
mammary gland de novo synthesis and reduction in enzyme expression in the cell (Figure
2) (Bauman and Griinari, 2003; Shingfield and Griinari, 2007). High polyunsaturated fat

7
supplementation causes a reduction in C18:0 supply to the mammary gland, which
decreases the endogenous synthesis of C18:1 cis-9 due to inhibition of the ∆9 enzyme,
limiting milk fat secretion (Glasser et al., 2007). Boeckaert et al. (2008) tested the
hypothesis that the inclusion of algae in the diet of cattle would generate shifts in the
rumen biohydrogenation process and alter the microbial ecosystem by conducting an
experiment using a control diet, a starch-rich diet, a bicarbonate-buffered starch-rich diet,
and a diet supplemented with DHA-enriched micro algae (43.0 g/kg of DMI), where
algae was supplemented directly through a rumen fistula. The study did not observe a
decrease in C18:0 in the rumen, and the level of C18:1 cis-9 in milk was not affected.
They suggest that there was an increase in endogenous synthesis of this isomer or that the
activity of the enzyme Δ9-desaturase decreased.
Milk fat is synthesized by the combination of fatty acid uptake, fatty acid
transport, desaturation of fatty acids, triacylglycerol synthesis, and de novo fatty acid
synthesis. The production of C18:2 trans-10, cis-12 (Figure 2) during the shift in the
biohydrogenation of linoleic acid has anti-lipogenic effects, inhibiting fat synthesis in the
mammary gland and adipose tissue due to reduction of ∆9 enzyme activity (Bauman and
Griinari, 2003). This inhibitory effect is possibly generated due to the double bond
position (Baumgard et al., 2000; Saebo et al. 2005a). The addition of a double bond in the
cis-6 position to the conjugated linoleic acid trans-19, cis-12 reduces the inhibitory effect
on fat synthesis (Park et al., 2004). Studies in vitro show that C18:2 trans-10, cis-12 and
C18:2 cis-10, trans-12 inhibit lipid accumulation in 3T3-L1 pre-adipocytes (Griinari et
al., 2005). This process is reversed when a carboxyl group or an alcohol is added to the
primary carbon, followed by an addition of a functional group or a double bond to the

8
carbon chain between the position 10, 12 double bond, indicating that fatty acid structure
play a key role on fat synthesis (Saebo et al., 2005b; Park et al., 2004). It is hypothesized
that other isomers besides conjugated linoleic acid trans-10, cis-12/ cis-10, trans- 12/
trans-9, cis-11 produced during the biohydrogenation process also affect milk fat
synthesis (Piperova et al., 2000; Piperova et al., 2002).
Exogenous Fat
Ruminants consuming a diet supplemented with fat tend to have a higher
absorption of FFA, which generates a decrease in de novo synthesis of free fatty acids
from acetate in adipocytes (Chilliard, 1993). Stearic acid (C18:0) and palmitic acid
(C16:0) are produced in greater quantities in the rumen when the free fatty acid content in
the rumen is increased (Jenkins, 1993). In vitro studies suggest that feeding fat inhibits
microbial growth and metabolism due to the formation of a coating over the feed particles
(Luvisetto et al., 1987; Galbraith et al., 1971). This layer reduces the action of bacterial
hydrolytic enzymes and consequently reduces structural carbohydrate digestibility
(Cheng et al., 1991; Immig et al. 1991). Bacterial membrane function can also be affected
by lipid supplementation where the hydrophobic and amphiphilic properties of the fatty
acids cause an attachment of the exogenous fat to the bacterial membrane (Gutknecht,
1988), disturbing the membrane function by uncoupling oxidative phosphorylation from
electron transfer (Luvisetto, 1987).
Another challenge encountered with fat supplementation in ruminants is the fatty
acids content that flows out of the rumen is different from the fatty acid profile of the
diet, due to the biohydrogenation process that occurs (Ward et al., 1964). The passage of

9
unsaturated fatty acids out of the rumen is reduced, delivering saturated fat to the small
intestine, which is absorbed and deposited in the body (Wu et al., 1991). This process can
be modified by offering a diet rich in nonstructural carbohydrates, where
biohydrogenation process can be reduced and increase the flow rate of unsaturated fatty
acids out of the rumen (Leat, 1977). Diets rich in readily fermented carbohydrates tend to
decrease rumen pH, which increases the production of trans isomers due to bacterial
activity and the incomplete biohydrogenation of dietary unsaturated fatty acids, leading to
a decrease in milk fat synthesis (Staples and Cullens, 2005). To avoid these negative
effects on ruminal digestion, fat supplementation is limited to 5% or less in the diet
(Byers and Schelling, 1988).
Tallow
Fat sources high in saturated fatty acids such as tallow do not alter microbial
fermentation as greatly. Saturated fat is more insoluble and the bacterial cell would be
less likely to attach to this isomer, decreasing digestibility (Chalupa et al., 1984). The
amount of fat supplemented to ruminants can generate different impacts in the overall
animal performance. Studies have shown that when supplementing 2% tallow in the diet
there was a decrease in dry matter intake of lactating cows (Onetti et al., 2001; Onetti et
al., 2002; Ruppert et al., 2003; Onetti et al., 2004). Onetti et al. (2001) observed a
depression in milk production when feeding half concentrate half corn silage to Holstein
cows and supplementing them with 2% tallow in the diet. However other studies
observed an increase in milk production by 2 kg/d when supplementing 2% tallow to
lactating Holstein cows consuming a forage diet (Smith et al., 1993; Onetti et al., 2002).

10
This difference could be due to the reduction in the biohydrogenation process, producing
more C18:2 trans isomers. Even though milk production is affected differently depending
on the diet, studies demonstrated that milk fat decreases from 0.2% to 0.47% when
supplementing 2% tallow to lactating cows (Smith et al., 1993; Adams et al., 1995; Onetti
et al., 2001; Onetti et al., 2002; Ruppert et al., 2003; Onetti et al., 2004). This indicates
that concentrate diets tend to cause negative impacts on animal performance when
supplementing 2% tallow in the diet, as it was observed by Smith et al. (1993) that by
replacing corn silage for alfalfa hay would cease milk fat depression.
Fish Oil
Ashes et al. (1992) demonstrated sheep consuming fish oil at 2% of the diet had
an increase in serum lipids of 13% in eicosapentaenoic acid (EPA/C20:5n-3) and 5-fold
in docosahexaenoic acid (DHA/C22:6n-3), and a decrease in linoleic acid (C18:2) and
stearic acid (C18:0). No detectable amount of EPA and DHA were in the adipose tissue,
but EPA and DHA increased by 3- and 4-fold in muscle tissue. Also, a reduction in
vaccenic acid (C18:1 trans-11) concentrations in total muscle phospholipids was seen in
sheep. This suggests that the fatty acid content in fish oil is only biohydrogenated to a
certain extent, possibly due to specific ester linkage of the triglycerides being more prone
to microbial lipases. Eicosapentaenoic acid (EPA/C20:5n-3) and docosahexaenoic acid
(DHA/C22:6n-3) were not detected in ruminal microbial lipids of sheep supplemented
with fish oil for 30 days. These results indicate that ruminants can incorporate DHA and
EPA into muscle tissue, but that the amount is quite limited and may not be enough to
affect human health.

11
Fish oil inhibits the biohydrogenation of polyunsaturated fatty acids to stearic acid
(C18:0) due to the high concentrations of EPA (C20:5n-3) and DHA (C22:6n-3). This
increases the flow of the intermediates out of the rumen (Lee et al., 2005; Loor et al.,
2005a). It was hypothesized this inhibition occurs due to the reduction of Butyrivibrio
proteoclasticus population, which converts trans-11 18:1 to stearic acid (C18:0) in the
rumen (Lee et al., 2008). However, Huws et al. (2010) found no relationship between
inclusion of fish oil in the diet and a reduction of Butyrivibrio proteoclasticus population.
A linear regression analysis also showed no relationship between stearic acid (C18:0)
flow out of the rumen and Butyrivibrio proteoclasticus concentration. This suggests other
bacterial populations might be shifting the final step in the biohydrogenation process.
Scollan et al. (2001) observed steers consuming fish oil at 6% of total dietary
intake had higher carcass conformation score when compared to a control and lightly
bruised whole linseed. Animals consuming fish oil had lower subcutaneous adipose tissue
content with higher proportions of C12:0, C14:0, C16:0 and C18:1 trans but lower
C18:1n-9 and C18:2n-6 when compared to control animals. Fish oil increased the
proportion of palmitic acid (C16:0) in muscle neutral lipids, but did not affect the
proportions of stearic acid (C18:0). In muscle phospholipids the proportion of C18:1n-9
decreased due to the replacement of it with EPA (C20:5n-3) and DHA (22:6n-3). Both
muscle neutral lipids and phospholipids had a lower concentration of C18:2n-6.
Consuming 100g/d of meat from animals submitted to the fish oil treatment would
provide 103.5mg/d of n-3 fatty acids, 24 mg/d of EPA and 5.3mg/d of DHA.
Flaxseed oil fed to finishing cattle increased the percentage of carcasses grading
Choice and improved marbling scores due to its high content of α-linolenic acid

12
(LaBrune, 2000). However, steers receiving fish oil supplementation for 70-d, decreased
feed intake, but average daily gain and the gain: feed ratio did not differ in response to
this supplementation (Wistuba et al., 2006). It was observed in previous studies that
protected PUFA did not alter in situ forage degradability, but reduced dry matter intake in
beef cows (Cooke et al., 2011).
MARINE ALGAE
Marine algae can be used as a source of polyunsaturated fatty acids
supplementation in that it contains a large amount of long-chain n-3 fatty acids and high
amounts of EPA and DHA (Barsanti and Gualtieri, 2006). Feeding marine algae has been
shown to generate an incomplete biohydrogenation of fatty acids in vitro using rumen
samples of steers consuming a commercial pelleted grain based concentrate (Chow et al.,
2004) and in vivo feeding a high concentrate to forage ratio (65:35) (Loor et al., 2005b),
resulting in accumulation of several C18:1 trans fatty acids and CLA in the rumen
(Chilliard et al., 2007). Conjugated linoleic acid is produced in the rumen via
biohydrogenation of linoleic acid (C18:2) to stearic acid (C18:0) as observed in growing
ruminal bacterial cell cultures (Kim et al., 2000). Studies using fish oil have higher levels
of DHA, compared with plant oils, and it is more effective in elevating the milk fat cis-
9,trans-11 conjugated linoleic acid content when supplementing lactating ewes
consuming ad libitum hay, which leads to a reduction in production of C18:2 trans-
10,cis-12 that can inhibit fat synthesis by the mammary gland (Offer et al., 1999). This is
due to the inhibitory effect of DHA on the reduction of vaccenic acid (C18:1 trans-11) to
stearic acid in the rumen (Boeckaert et al., 2008). Marine algae has higher levels of DHA

13
and is expected to produce greater cis-9, trans-11 conjugated linoleic acid responses than
fish oil.
Inclusion of DHA enriched marine algae increased the content of trans-C18:1 in
the rumen, which could be due to the reduction in the population of Butyrivibrio
fibrisolvens and Butyrivibrio proteoclasticum (Lee et al., 2008). The reason why the
isomers C18:2 trans-11, cis-15 also increase is still unknown (Vlaeminck et al., 2007).
Harfoot and Hazlewood (1997) classified the bacteria involved in the biohydrogenation
process in two groups, where group A hydrogenate polyunsaturated fatty acids to C18:1
trans-11and group B complete the process by hydrogenating C18:1 trans-11 to C18:0 and
C18:2 trans-11, cis-15 to C18:1 trans-15- and cis-15. AbuGhazaleh et al. (2009)
demonstrated that substituting fish oil with DHA-microalgae in the diet of lactating
Holstein cows grazing on an alfalfa-grass had no effect on vaccenic acid and conjugated
linoleic acid cis-9, trans-11 concentrations in milk. It was also observed that there is a
linear relationship between the reduction of stearic acid (C18:0) and the increased
concentration of the previous isomers in the milk of animals supplemented with marine
algae. This suggests that fish oil has less effect in the final step of the rumen
biohydrogenation process once DHA levels are higher in marine algae and induce an
accumulation of trans C18:1 isomers in the rumen (Boeckaert et al., 2007).
Defaunated lambs fed a diet of hay chaff and crushed oats presented increased
levels of vaccenic acid (C18:1 trans-11) in the abomasum and conjugated linoleic acid in
the muscle (Dawson and Kemp, 1967), demonstrating the microbial population is directly
involved with the biohydrogenation process, where ciliates show higher content of
vaccenic acid and conjugated linoleic acid than bacteria (Devillard et al., 2006). The

14
different fatty acid isomers affect the biohydrogenation process differently, where EPA
and DHA tend to inhibit the hydrogenation of C18:2 n-6 and C18:3 n-3, producing more
C18:1 trans-11 and C18:2 trans-11cis-15 (Boeackaert et al., 2007). In vivo studies show
an accumulation of C18:1 trans- 11 in the rumen of cows supplemented with microalgae
and of C18:2 n-6 and C18:3 n-3 due to reduced rate of lipolysis. A higher concentration
of C18:2 trans-11, cis-15 and C18:1 trans-11 was also observed, indicating microalgae
supplementation affects the second and third step of the biohydrogenation of C18:3n-3 to
stearic acid (C18:0) and the second step of C18:2 n-6 to C18:0 (Boeckaert et al., 2007).
Butyrivibrio fibrisolvens is a bacterial strain associated with rumen
biohydrogenation of unsaturated C18 isomers (Polan et al., 1964; Wasowska et al., 2006),
but other species such as Butyrivibrio proteoclasticus are also involved in this process
(Maia et al. 2007; Paillard et al., 2007). Fat sources containing high levels of EPA and
DHA like algae can inhibit the production of stearic acid (C18:0) (AbuGhazaleh and
Jenkins, 2004) and reduce the population of ciliates in the rumen fauna (Boeckaert et al.,
2007). Boeckaert et al. (2008) demonstrated that DHA enriched marine algae
supplementation in cows changed the rumen fauna, thereby reducing the Butyrivibrio
population, consequently increasing the concentration of C18:1 trans-11 and C18:1
trans-10 and reducing C18:0. The rumen fluid of cows supplemented with marine algae
do not present different concentrations of palmitic acid (C16:0) when compared to
control animals, but the concentration of stearic acid (C18:0) is reduced in a linear
manner with supplementation level (Or-Rashid et al., 2008). This occurs due to the
inhibition in the conversion of C18:1 trans-11 to C18:0 in the presence of unsaturated
long chain fatty acids like DHA and EPA (Lee et al., 2005).

15
OMASAL SAMPLING
To understand the fatty acid biohydrogenation process in the rumen, it is
necessary to analyze the flow of fatty acids from out of the rumen, where the bacterial
and protozoal population is reduced (Smith, 1984; Punia and Leibholz, 1994). Typically
duodenal fistulas are used to collect post-ruminal samples. This technique can be difficult
due to long animal recovery time from surgery and it requires constant observation and
maintenance (Huhtanen et al., 1997). Omasal collection has been demonstrated to be a
more viable procedure to collect post-ruminal samples. Abomasum sample collections
are more invasive than omasal collections, and intestinal cannulas can cause distension of
the intestines and retention of digesta (Wenham and Wyburn, 1980).
CONCLUSION
A diet rich in polyunsaturated fatty acids, especially EPAand DHA can improve
human health. Unsaturated fats undergo biohydrogenation in the rumen, which can be
affected by rumen pH, type of diet offered to the animals, source of fat in the diet,
quantity of fat supplemented to the animal and others. Because of this process, absorption
and passage of unsaturated fat from the rumen is limited increasing the amount of
saturated fat absorbed in the small intestine. Fish oil and marine algae can be used as a
source of polyunsaturated fatty acids, inhibiting to some extent the biohydrogenation
process, accumulating several C18:1 trans fatty acids and conjugated linoleic acid (CLA)
in the rumen.

16
OBJECTIVE
The objective of this study was to study ruminal heterotrophically grown
microalgae high in DHA metabolism in a high grain finishing diet.

17
CHAPTER THREE: INTRODUCTION
Nicholson et al. (1992) reported that cattle receiving 230g/kg of fishmeal-
supplementation had a lower dry matter intake when compared to a control group, but the
animals gained weight at the same rate due to a more efficient feed conversion. These
results suggested there was no negative impact of fish meal supplementation on growth
performance, because of an increase in the efficiency of use of absorbed nutrients.
Kronberg et al. (2011) used flaxseed to supplement steers for 85-d finished on pasture.
They obtained an average daily gain of 1.04 kg for the flaxseed group, and 0.83 kg for the
control group, 25% greater for the supplemented steers than for the control. Methane
production is also reduced with lipid supplementation which will have a greater impact
on a high-fiber diet, since the amount of methane produced during microbial fermentation
is greater than in a high-grain diet (Zinn and Plascencia, 1993). The specific application
of PUFA sources in high-grain finishing diets has received limited attention. Therefore,
it is the objective to study ruminal algae PUFA metabolism in steers fed a high-grain
finishing diet.
MATERIALS AND METHODS
ANIMALS AND EXPERIMENTAL DESING
Eight rumen cannulated Holstein steers (BW = 337 ± 37 kg) were used in a
replicated 4 x 4 Latin square design experiment to study the rumen metabolism of fatty
acids incorporated into a high-grain finishing diet. Each experimental period was
composed of 21 days. Animals were adapted to the fat supplementation for 14 days and
placed in individual metabolism stalls for 7 days. The steers were fed a diet based on

18
75% cracked corn grain, 7.5% chopped hay and 7.5% corn silage (Table 1). Chromic
oxide was added to the diet to determine digestibility. To accomplish this, feed for each
experimental period was mixed in a single batch, placed in barrels and stored refrigerated
(10ºC).
The primary aim of the experiment was to evaluate the ruminal metabolism of
heterotrophically grown microalgae high in DHA. Treatments were a negative control,
with no fat supplementation (C), a positive control of tallow added at 60g/day (T), fish oil
at 60g/day (FO) and heterotrophically grown microalgae high in DHA at 100g/day (A).
The fish oil and the algae were analyzed for fatty acid composition and fish oil was added
to supply an equivalent amount of total fat (60g/day). The dietary fat treatments were
dosed in the rumen through the cannula mixed with 0.45 kg of the diet at the time of
feeding (0700) during the adaptation period, and twice a day (0700 and 1500) during the
7 days in individual metabolism stalls. Steers were fed at approximately 1.75 x NEm so
that intakes were equalized across all treatments while ensuring complete consumption of
an intake that promoted a high rate of gain. Animals received a Revalor XS implant
(Intervet, Summit, NJ) which is an implant containing 200 mg of trenbolone acetate and
40 mg estradiol; at the beginning of the experiment to stimulate growth and increase
nitrogen retention.
DIGESTIBILITY AND N-BALANCE
Intake was determined daily by weighing the feed offered and the orts remaining
for each animal. Feedstuffs and orts were sampled daily and stored at −4°C. Urine was
collected at 0700 daily from day 15-21 by continuous suction using a rubber funnel

19
system attached to the ventral portion of the abdomen, collecting urine into a plastic
collection vessel. Urine acidity was reduced to pH < 3 by adding 1 L of a 23.5% solution
of H3PO4 to the collection vessel in order to prevent ammonia-N loss. Urine output
weight was recorded daily. Five percent of the urine was subsampled daily to contribute
approximately 250 g to a composite for each period and steer. This composite was stored
at 0°C until analysis. Fecal collection was accomplished by scraping fecal excretions
into a trough behind each animal several times daily. The wet weight of fecal output was
recorded at 0700 daily from day 15-21 for each steer. Fecal samples were weighed and
mixed and 10% of total fecal mass was sampled daily and frozen at 0°C until the end of
the experiment then composited by period and steer using a Hobart mixer (Model H-600,
Hobart Manufacturing Co, Troy, OH).
RUMINAL FATTY ACID METABOLISM
Reticulum and Omasal Sampling
Reticulum and omasal contents were collected as described by Huhtanen et al.
(1997) with modifications as described by Ahvenjarvi et al. (2000) using a plastic tube
(14 mm i.d.) connected to a vacuum sampling pump (Huhtanen et al., 1997) with the tube
manually passed thru the reticulo-omasal orifice and located in the omasum. The
reticulum samples were also collected as described by Fredin et al. (2014) by placing a
250 mL collection bottle in the reticulum at the reticulo-omasal orifice. After positioning
the bottle it was then unstopped and a sample collected. Approximately 750 mL of
digesta per sample was collected. Reticulum and omasal samples were obtained at 0700,
1000, 1300 and 1600 on day 16 with additional samples collected for 2 day, four times a

20
day 1 h later than on the previous day to obtain samples for each hour from 0700 – 1800
until day 18. Separating by location, all reticulum and omasal digesta samples were
mixed, homogenized in a blender and composited on an equal volume basis (500 mL per
sample) and stored frozen at −20°C prior to lyophilizing and grinding.
Rumen Samples
Rumen fluid (50 mL per sample) was collected on day 19 at 2 h intervals using a
suction strainer from 0700-1700 for pH and VFA analysis.
Blood Samples
Blood was collected on the last day of sample collection on day 21 as the animals
were weighed at 0900. A 20 mL sample was collected by jugular puncture using an
empty syringe and 19 gauge needle and immediately transferred into a 50 mL centrifuge
tube containing sodium heparin for the collection of plasma. Samples were placed
immediately on ice, centrifuged within 30 min after collection at 12,500 × g for 30 min at
4ºC, the plasma was separated and frozen at −80ºC.
SAMPLE ANALYSIS
Feed, Feces, Urine, Reticulum and Omasal Samples.
Composite feed and fecal samples were dried at 55°C in a forced-air oven.
Reticulum and omasal composites were freeze-dried. All dried samples were ground
through a 1 mm screen in a Wiley Mill (Model 4; Thomas Scientific, Swedesboro, NJ).
The dried ground samples were analyzed for dry matter using AOAC method 930.15.

21
Feed, feces, and urine were analyzed for N using method 990.03 (AOAC, 2005). Feed
and feces were analyzed for crude fat with ether extract using method 920.39 (AOAC,
1992). Feed, feces, reticulum, and omasal samples were analyzed for Cr (Williams et al.
1962). Feed, feces and omasal samples were analyzed for total fatty acid composition
(Sukhija and Palmquist 1988). Total long chain fatty acid analysis was modified by
including 50 mg/L butylated hydroxyl toluene (BHT) in benzene to protect fatty acids,
judicious use of N2 to all vials to prevent oxidation, addition of C19:0 isomer to standards
as an internal standard and incubation at 90°C to better hydrolyze soaps. Samples were
analyzed using gas chromatography analysis (Hewlett-Packard GC system 6890+ with an
FID) using a SP-2560 capillary column (100 m × 0.25 mm i.d. with 0.2 μm film
thickness; Supelco Inc., Bellefonte, PA). The initial GC oven temperature was 60°C for
28 min then ramped to 210°C at 2°C/min and held for 20 min. There was a second ramp
in temperature at 1°C/min to 230°C and held for 10 min. A third and final ramp at
2°C/min to 240°C with no hold in temperature. Injector and detector temperatures were
maintained at 240°C, the helium carrier gas flow rate was 1 ml/min, and the split ratio
was 100:1. Ruminal digestion of dry matter, ether extract, N and fatty acids were
calculated as described by Bock et al. (1991).
Ruminal Fluid.
After the ruminal fluid collection the pH was measured immediately (Corning
Model 12, Corning Scientific Instruments, Medfield, MA) and samples were prepared for
volatile fatty acids (VFA) analysis; 10 mL samples of ruminal fluid were placed into 50-
mL centrifuge tubes and 1.00 mL 85 mM 2-ethylbutyrate internal standard was added and

22
followed by 1.00 mL 50% metaphosphoric acid. The tubes were then capped, mixed and
frozen overnight (-4ºC). The following day the samples were thawed and centrifuged at
39,000 × g for 20 min before transferring to injection vials. The VFA concentrations
were determined by gas chromatography with a flame ionization detector (Agilent
HP6890 Plus GC with Agilent 7683 Series Injector and Auto Sampler; Agilent
Technologies, Santa Clara, CA) using a Supelco 25326 Nukol Fused Silica Capillary
Column (15 m × 0.53 mm × 0.5 µM film thickness; Sigma/Supelco, Bellefonte, PA). To
accomplish the analysis, 0.1 µL of sample was injected at 110ºC with a 2:1 split. After a
1 min hold, the temperature was increased at 5 ºC /min to 125 ºC for 2 min. The inlet
and injector are set at 260 ºC.
Blood Samples
Plasma was analyzed for total fatty acid profile as described by Corl et al. (2001).
Lipids were extracted from 2mL of plasma by adding 3.0 mL hexane: isopropanol (3:2).
This was washed with 2.0 mL sodium sulfate (67 g/L). The upper phase was removed
and dried over sodium sulfate 30 min and transferred to a clean tube and dried with N2.
Methylation was performed by adding 0.5 mL hexane and 40 µL methyl acetate. The
methylation reagent was prepared using 1.75 mL methanol: 0.4 mL 5.4 M sodium
methoxide, and 40 µL of the methylation reagent was added, layered with N2 and reacted
at room temp for 24 h. The termination reagent was prepared by combining 1.0 g oxalic
acid in 30 mL diethyl ether, 60 µL of the termination reagent was added, followed by the
addition of a few grains of CaCl2. The solution was transferred to vials for gas
chromatography analysis (Hewlett-Packard GC system 6890+ with an FID) using a SP-

23
2560 capillary column (100 m × 0.25 mm i.d. with 0.2 μm film thickness; Supelco Inc.,
Bellefonte, PA).
CALCULATIONS
Dry matter digestibility (%)
/ – /
/ x 100
Organic matter digestibility (%)
/ – /
/ x 100
DM Flow (g/d)
/
/ /
Corrected DM Flow (g/d)
/
/
Corrected apparent ruminal digestibility (%)
/ / – /
/ / x 100

24
Corrected apparent post ruminal digestibility (%)
/ – / /
/ / x 100
Total long chain fatty acid intake (g/d)
⁄ ⁄ +
⁄ ⁄
Total long chain fatty acid output (g/d)
⁄ ⁄
Total long chain fatty acid omasal flow (g/d)
⁄ ⁄
STATISTICAL ANALYSIS
The experiment was analyzed as a replicated Latin square design with mixed
models in SAS (version 9.2; SAS Inst. Inc., Cary, NC). Volatile fatty acid data were
analyzed using the least square means repeated measures of SAS (version 9.2; SAS Inst.
Inc., Cary, NC), where time was included as a variable. The remaining data were
analyzed using steer as the experimental unit and treatments were compared using

25
contrasts; Control vs. others, Tallow vs. Fishes (Algae + Fish oil), Fish oil vs. Algae
where P < 0.05 was considered significant.
RESULTS
Addition of fat, regardless of form did not affect dry matter (DM) or organic
matter (OM) intake, DM or OM digestibility (Table 2). Algae treatment had a tendency
(P = 0.06) to lower fecal excretion when compared to fish oil (Table 2). No differences
were observed in urine excretion or fecal recovery of Cr. Animals consuming algae had a
lower fecal crude fat percentage (P = 0.02) and higher fecal crude fat digestibility (P
=0.0368) when compared to fish oil. Control animals had lower fecal crude fat
digestibility (P = 0.003) compared to the other treatments.
There were no differences among treatments in N intake, fecal N, urinary N
excretion or N retention (Table 3). The percentage N in urine and feces were also
unaffected by treatment.
Rumen fermentation variables (Table 4) were influenced by time after feeding (P
< 0.05); however, there were no time × treatment interactions so the focus will be on
daily means for all samples collected. There was no difference in acetate and propionate
concentrations, acetate molar proportions or total VFA concentration. Propionate molar
proportion was higher in control (P = 0.01) and tallow (P = 0.002), but was not different
between fish oil and algae treatments. Isobutyrate concentrations were greater (P = 0.02)
for fat treatments than control. Butyrate concentration (control vs others P = 0.004;
tallow vs fishes P < 0.001; algae vs fish oil P = 0.016), and molar proportion (control vs
others P = 0.02; tallow vs fishes P < 0.001; algae vs fish oil P = 0.28) were affected by fat

26
treatments being lower for tallow than the fish sources (P < 0.001) and greater for algae
than fish oil (P = 0.02). Isovalerate concentrations (control vs others P < 0.001) and
proportions (P = 0.01) were greater for fat treatments than control. Valerate
concentrations were greater for the fat treatments (control vs others; P = 0.0285) and
greater for algae than fish oil (algae vs fish oil; P = 0.003). Acetate:propionate was not
affected by treatment whereas pH was increased for all fat treatments (control vs others;
P = 0.001).
Corrected ruminal DM digestibility (corrected for fecal recovery of Cr) was on
average 60.6% whereas uncorrected ruminal DM digestibility was 34.3% (Figure 4).
Samples collected in the reticulum consistently (Figure 5A) produced higher DM flows
out of the rumen (Figure 5B). Omasal flow, apparent ruminal digestibility of DM and
OM were not affected by fat supplementation (Table 5); however, ruminal DM flow
tended (P = 0.09) to be greater for algae than fish oil. Animals receiving the fat
treatments tended to have greater DM (P = 0.09) and OM (P = 0.08) digested post
ruminally and animals receiving algae tended (P = 0.06) to have greater DM and had
greater (P = 0.04) OM digested post ruminally than those receiving fish oil.
Intake, fecal excretion and total tract digestion of starch were not affected by
treatment (Table 6). Animals consuming tallow had higher ruminal starch digestibility
when compared with fish oil and algae treatments (P = 0.04) and had a tendency for
lower starch flow out of the rumen (P = 0.06).
Tallow had the lowest ruminal and total tract digestibility of total long chain fatty
acid (Table 7). The medium chain fatty acids from C13:0 to C17:1, with the exception of
C16:0, were present in small amounts and only in some diets and when present were

27
completely digested in the rumen and the total tract. Algae had significant higher C16:0
digestibility compared to fish oil (P = 0.03). Tallow, fish oil and algae treatments had
negative C18:0 ruminal and total tract digestibility indicating a net production of this
fatty acid in the rumen. Digestibility of EPA (C20:5n3) and DHA (22:6n3) were 100% in
algae and fish oil treatments for ruminal and total tract digestibility indicating that the
amount provided in the supplementation were absorbed or biohydrogenated in the rumen.
The control diet provided the lowest levels of C16:0 total tract digestibility (P = 0.003).
Algae and fish oil had a tendency for greater total tract digestibility of total long chain
fatty acid when compared with tallow (P = 0.08; Table 7).
Total long chain fatty acid omasal flow was higher for tallow, followed by algae,
control and fish oil (tallow vs fishes P = 0.02; Table 8). Algae had the highest C16:0
omasal flow among treatments (algae vs fish oil; P = 0.002) and control had the lowest
(control vs. others; P = 0.003). Levels of vaccenic acid (C18:1 trans-11) were non-
detectable in omasal flow samples. Algae had a tendency to lower omasal flow of C18:0
(P = 0.10) and tallow had the highest (P = 0.0002). Algae had a tendency for higher
omasal flow of C18:1cis-6 (algae vs fish oil; P = 0.09) and C18:1 trans-9 (algae vs fish
oil; P = 0.08) than fish oil. Omasal flow of C18:1 cis-9 was higher in the algae treatment
when compared with fish oil (P = 0.03).
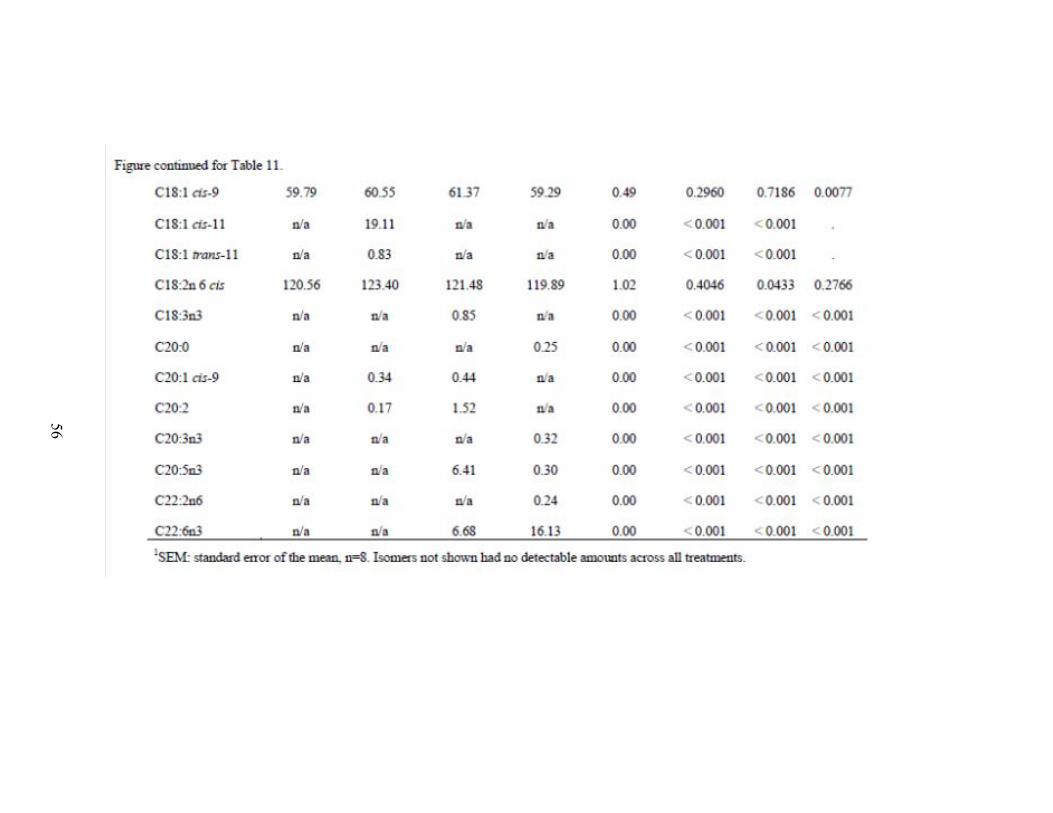
The total long chain fatty acid profile of blood plasma showed that fish oil and
algae had lower plasma C18:0 percent when compared with control (P = 0.01) and tallow
was higher than the fish oils (P = 0.0009; Table 9). The C18:1 trans-9 plasma percent
was greater for the fish oil and algae treatments when compared with tallow (P = 0.03)
and tended (P = 0.08) to be lower for control than the other treatments. The C18:2n6

28
trans percent was greatest for control (P < 0.001) with the tallow lower than the fish oil
(P < 0.001). The control treatment had lower (P = 0.04) C18:3n3 percent when compared
to the other treatments. The algae treatment had the highest C20:4n6 plasma percent (P =
0.005) and control the lowest (P = 0.01). The weight percent C22:6n3 (DHA) in algae
was 4.22% while fish oil only had 1.31% (P = 0.02). Control had zero percent plasma
DHA (P = 0.03). Fish oil and algae treatments gave the highest levels of C24:0 (P = 0.02)
in blood plasma when compared with tallow and the control tended (P = 0.07) to be lower
than the other treatments.
DISCUSSION
Omasal samples were obtained using a sampling tube connected to a vacuum
pump, an Erlenmeyer vacuum flask was attached and used as a collection vessel for the
omasum digesta (Figure 3). This system provided alternating suction and pressure to
prevent clogs. The sampling tube was placed by hand in the omasum by passing the
reticulum-omasum orifice, accessed through the rumen fistula. Once the sampling device
was placed in the omasum, the vacuum pump was initiated resulting in digesta flow to the
Erlenmeyer flask. This technique has shown to be efficient to obtain omasum samples
and useful to analyze digestibility parameters (Ahvenjarvi et al., 2000; Choi et al., 2002;
Ahvenjarvi et al., 2003; Shingfield et al., 2003; Reynal and Broderick, 2005; Broderick et
al., 2008).
By including chromium oxide in the diet, we could determine flow of fatty acids
out of the rumen by correlating fatty acid composition in the omasal samples and omasal
flow. Lundy et al. (2004) obtained consistent results using Cr as a marker and analyzing

29
omasum samples to determine rumen digestibility. All digestibility calculations used
chromium excreted in feces instead of chromium consumed in the diet. It was felt this
provided the best estimate of Cr intake based on estimates of rumen DM digestibility
obtained for both measures of Cr intake (Figure 4). Corrected ruminal DM digestibility
gave a more realistic measurement of rumen DM digestibility for steers fed a high
concentrate diet. Knowlton et al. (1998) obtained 57.3% ruminal dry matter digestibility,
May et al. (2009) obtained 78.84% total tract dry matter digestibility for rolled dry corn,
which is similar to corrected ruminal DM digestibility data in this study.
We next compared DM flow between the two sampling locations, the reticulum
vs. the omasum. The reticulum flow is higher than omasum flow across treatments
(Figure 5a). It was felt that flows determined from reticulum samples were unrealistically
high. This was based on estimates of rumen starch digestibility with omasal samples
providing more reasonable estimates (Table 6), observing previous studies where dry
rolled corn rumen starch digestibility was 85.6% (Streeter et al., 1989), 78% (Galyean et
al., 1976), 75.9% (Zinn , 1987), 72% (Cole et al., 1976), 70.6% (Zinn et al., 1995), and
total tract starch digestibility was 89.8% (Zinn et al., 1995), 94% (Cole et al., 1976), 96%
(Galyean et al., 1976) and 92.4% (Streeter et al., 1989).
It was observed in previous studies that animals consuming high levels of
protected PUFA supplementation had a decreased DM intake (Araujo et al., 2010; Cooke
et al., 2011), but Duckett et al. (2002) did not observe a this decrease in intake when
limiting the fish oil supplementation to 1% of the diet. In this study fat intake was limited
to 60g of fat/day and the dry matter intake was approximately 1.75 x NEm to equalize

30
intake among treatments. This would equate to only 1% added fat so it is not surprising
intake was not affected.
There were no differences in N metabolism. Previous studies have shown that
lipid supplementation does not influence N metabolism (Doreau et al., 1991; Kucuk et
al., 2004; Montgomery et al., 2014). Nitrogen degradability could increase with fat
supplementation, due to a shift in proteolytic action of the microbes (Broderick et al.,
1991). It was demonstrated that this occurs due to a drop in protozoal population caused
by the added fat in the diet reducing the rate of engulfment of bacteria (Sutton et al.,
1983).
A molar proportion was higher in tallow than in fish oil and algae treatments,
which could be due to the fact that saturated fatty acids can shift VFA proportions,
increasing propionate production (Jenkins, 1993) due to shifts in the microbial population
and reductions in the protozoal population and inhibition of microbial growth (Doreau
and Chilliard, 1997). Increases in the butyrate concentration and molar proportion
observed in fish oil and algae treatments were observed previously in vitro by Lodge-
Ivey et al. (2013) when incubating lipid extracted algae Chlorella spp. Palmquist and
Griinari (2006) also observed an increase in butyrate concentration in vivo when
supplementing cattle with fish oil at 10 g/kg of DM. The presence of DHA in the rumen
can decrease propionate and increase butyrate molar proportions once it reduces the
protozoa population in the rumen (Vlaeminck et al., 2007), both effects have been
observed in fat treatments containing high levels of DHA. Isobutyrate, isovalerate and
valerate concentrations were higher when animals received fat supplementation, and
valerate concentration was particularly increased by algae supplementation. Fat sources

31
containing high levels of C18:1 can increase the isobutyrate, isovalerate and valerate
proportions in the rumen (Hristov et al. 2005). Rumen pH was lower for control animals
which suggest that the fat supplementation was beneficial for maintaining a higher
ruminal pH. Van Emon et al. (2015) did not observe differences in rumen pH when
supplementing cattle with deoiled microalgae meal, suggesting the increased pH was a
response from the added fat.
The ruminal and total tract starch digestion was similar to results obtained in
previous studies (Orskov, 1986, Buchanan-Smith et al., 1986; Holmes et al., 1970;
McNeill et al., 1971; Hinman and Johnson, 1974a; Hinman and Johnson, 1974b, Garcia
et al., 1981a; Hibberd et al., 1983). Increasing fat intake can reduce starch digestibility,
once carbohydrate digestion decreases and shift VFA proportions (Jenkins, 1993). This
study did not observe the reduction in acetate and butyrate ruminal concentrations that
could lead to a compromised starch digestion. Bock et al. (1991) also did not observe
differences in ruminal and total tract starch digestion when supplementing 3.5% soybean
oil or 3.5% tallow to steers fed high concentrate diets.
A negative rumen digestibility represents fatty acids that had higher rumen output
than intake, due to microbial activity in the rumen. Tallow, fish oil and algae treatments
all had negative C18:0 ruminal digestibility due to microbial fermentation saturating the
unsaturated fatty acids generating a higher output than intake. This production of stearic
acid is a result of the biohydrogenation process. The bacterial lipid profile is 90%
composed of saturated fatty acids, especially palmitic (C16:0) and stearic (C18:0) acids.
Unsaturated fatty acids also can be toxic to the microbial population and the
biohydrogenation process in the rumen is performed by the microbial population to avoid

32
the toxicity and produce the saturated fatty acids necessary for bacterial membrane
integrity (Kozloski, 2011).
Previous studies did not observe differences between control and marine algae
supplemented animals in rumen fluid C16:0 content (Or-Rashid et al., 2008). In this study
the fatty acid content of rumen fluid was not analyzed, but intact algae had higher flow of
C16:0 in omasal samples when compared to other treatments. A diet high in fat can
increase the production of palmitic acid (C16:0) if the content of free fatty acids present
in the rumen increases (Jenkins, 1993). Previous studies have shown that animals
consuming algae had a greater content of C18:1 trans-9 in rumen samples resulting from
the inhibition of the reduction step from C18:1 trans to C18:0 due to the presence of
PUFA (Lee et al., 2005; Loor et al., 2005a). In this study similar results were observed.
The algae treatment had a higher level of C18:1 cis-6 and C18:1 trans-9, and a lower
C18:0 flowing out of the rumen when compared to the other treatments. It has been
suggested that the high DHA content in the diet inhibits the conversion of C18:1 trans-11
to C18:0, due to the reduction in the Butyrivibrio fibrisolvens (Polan et al., 1964;
Wasowska et al., 2006) and Butyrivibrio proteoclasticum (Maia et al., 2007; Paillard et
al., 2007) population in the rumen. These bacterial strains are classified as part of a
microbial group that completes the biohydrogenation process of C18:1 trans-11 to C18:0
(Harfoot and Hazlewood et al., 1997). No omasal flow of C18:1 trans-11 was detected,
but the reduction of stearic acid content flowing out of the rumen could be caused by this
shift in microbial population, but this hypothesis cannot be confirmed as a bacterial strain
analysis was not performed in this study. Tallow had a higher stearic acid (C18:0) content
flowing out of the rumen when compared to fish oil and algae treatments, which was

33
expected since this fat source contains a higher concentration of stearic acid. Duckett and
Gillis (2010) detected a higher duodenal flow of C20:5 isomers in animals consuming 1%
fish oil, but in this study there was no detectable omasal flow of this isomer in any
treatment.
Blood plasma weight percentage of C16:0 was not different among treatments,
which differs from the higher omasal outflow of this isomer in algae treatment. Algae and
fish oil treatments presented higher levels of blood plasma C18:1 trans-9 and lower
levels of C18:0 demonstrating that the presence of PUFA inhibited the reduction step
from C18:1 trans to C18:0 and affected the blood plasma fatty acid concentrations.
Cooke et al. (2011) also observed a reduction in stearic acid (C18:0) content when
supplementing rumen-protected PUFA (10% DM basis) to 72 Angus × Hereford steers
consuming a grain-based diet. Kitessa et al. (2001) obtained similar results where C18:0
levels were lower in blood plasma of 5 sheep consuming unprotected and protected tuna
oil in a hay-oat grains diet. Similar to this study, AbuGhazaleh et al. (2003) observed an
increase in linolenic acid (C18:3n3) plasma levels when supplementing 1% fish oil plus
2% fat from flax seeds to 4 Holstein cows consuming 50% concentrate mix, 25% corn
silage, 12.5% alfalfa silage, and 12.5% alfalfa hay. The presence of EPA and DHA
inhibit the hydrogenation of C18:2n6 and C18:3n3 due to the reduction in lipolysis
(Boeckaerta et al., 2007). In this study a reduction in C18:2n6 trans was observed in
algae and fish oil treatments when compared to control. The percentage of C18:3n3 was
higher in fat supplemented animals which could be a result of inhibition of
biohydrogenation of linolenic acid (C18:3n3) to stearic acid (C18:0). Scollan et al. (2001)
supplemented steers with 6% fish oil and obtained higher C16:0 and C18:1 trans content

34
in adipose tissue; which is similar to omasal flow data in this study; and lower C18:2n6
content in muscle neutral lipids and phospholipids; similar to blood plasma data obtained
in this study. This demonstrates how fatty acids are incorporated differently depending on
the nature of the tissue. Arachidonic acid (C20:4n6) can be considered an essential fatty
acid in cattle, and it is used for prostaglandin synthesis, but when present in excess can
cause negative impacts such as vasoconstriction due to the production of thromboxanes
during prostaglandin synthesis (Simopoulos, 2006). Linoleic acid (C18:2) is converted to
C20:4n6 by chain elongation and desaturation (Church, 1988). Arachidonic acid (20:4n-
6) was higher in the blood plasma of animals consuming the algae treatment, which is
similar to previous studies (Kitessa et al., 2001; Cooke et al., 2011; Hess et al., 2012).
The levels of blood plasma DHA were higher in animals consuming the algae treatment.
This increased concentration was also observed by Hess et al. (2012) when
supplementing 38 g of algae and fish oil to 21 mares that consumed a hay and barley diet;
by Cooke et al. (2011) when supplementing rumen-protected PUFA; and by Kitessa et al.
(2001) when supplementing protected and unprotected tuna oil. This indicates that DHA
is absorbed and transported to the blood when animals consume this isomer in the diet.
Similar results were obtained by Ashes et al. (1992) when supplementing sheep with 2%
fish oil in the diet, presenting higher contents in serum lipids of DHA.

35
CHAPTER FOUR: SUMMARY AND CONCLUSIONS
The supplementation of heterotrophically grown microalgae high in DHA
(100g/d) through a rumen fistula in steers consuming a high grain finishing diet can be
used as a tool to modify the fatty acid profile flowing out of the rumen and present in the
blood plasma. Algae supplementation provided a polyunsaturated fatty acid profile that
inhibits to some extent the biohyodrogenation process in the rumen. This was indicated
by the higher omasal outflow of C18:1 isomers; C18:1 cis-6, C18:1 trans-9 and C18:1
cis-9; and lower outflow of C18:0. The high DHA content in this heterotrophically grown
microalgae could affect the steps in the biohydrogenation process that occurs in the
rumen. Blood plasma weight percent of C18:2n6 trans significantly lower in animals
consuming algae. This could indicate a partial inhibition of the biohydrogenation process
of C18:2n6. Algae successfully modified the fatty acid profile in blood plasma, where
animals presented a lower weight percent of C18:0 and a higher DHA content, even
though the flow of DHA out of the rumen was not detectable in omasal samples. Algae
treatment also was efficient in maintaining a higher rumen pH when compared to the
control diet, which can be beneficial on high grain diets. These data indicate that algae
feeding may have potential to alter the fatty acid profile of steers consuming a high grain
finishing diet.

36

37

38

39

40

41

42

43

44

45

46

47

48

49
Figure 1. Biohydrogenation linoleic and linolenic acid to stearic acid.

50
Figure 2. Shift in the biohydrogenation process during the conversion of linoleic acid to
stearic acid.

51
Figure 3. Omasum sampling vacuum pump.

52
Figure 4. Ruminal dry matter (DM) digestibility using chromium intake (%) or corrected
for fecal chromium recovery in steers fed a finishing diet with supplemental tallow, fish
oil or high-DHA microalgae.

53
Figure 5ab. (a) Average omasum and reticulum DM flow (g/d) and (b) treatments average
of omasum and reticulum DM flow (g/d) corrected for fecal chromium in steers fed a
finishing diet with supplemental tallow, fish oil or high-DHA microalgae.

54

55
56

57
REFERENCES AbuGhazaleh, A. A., D. J. Schingoethe, A. R. Hippen, and K. F. Kalscheur. 2003.
Conjugated linoleic acid and vaccenic acid in rumen, plasma and milk of cows fed fish oil and fats differing in saturation of 18 carbon fatty acids. J. Dairy Sci. 86:3648-3660.
AbuGhazaleh, A. A., and T. C. Jenkins. 2004. Disappearance of docosahexaenoic and eicosapentaenoic acids from cultures of mixed ruminal microorganisms. J. Dairy Sci. 87:645-651.
AbuGhazaleh, A. A., R. B. Potu, and S. Ibrahim. 2009. Short communication: The effect of substituting fish oil in dairy cow diets with docosahexaenoic acid-micro algae on milk composition and fatty acids profile. J. Dairy Sci. 92:6156-6159.
Adams, A. L., B. Harris, Jr., H. H. Van Horn, and C. J. Wilcox. 1995. Effects of varying forage types on milk production responses to whole cottonseed, tallow, and yeast. J. Dairy Sci. 78:573-581.
Agren, J. J., S. Vaisanen, O. Hannien, A. D. Muller, and G. H. Hornstra. 1997. Hemostatic factors and platelet aggregation after a fish-enriched diet or fish oil or docosahexenoic acid supplementation. Prostaglandins Leukot. Essent. Fatty Acids. 57:419-421
Ahvenjarvi, S., A. Vanhatalo, P. Huhtanen, and T. Varvikko. 2000. Determination of reticulo- rumen and whole-stomach digestion in lactating cows by omasal canal or duodenal sampling. Br. J. Nutr. 83:67-77.
Ahvenjarvi, S., A. Vanhatalo, K. J. Schingfield, and P. Huhtanen. 2003. Determination of digesta flow entering the omasal canal of dairy cows using different marker systems. Br. J. Nutr. 90:41-52.
Albert, C. M., C. H. Hennekens, C. J. O’Donnell, U. A. Ajani, V. J. Carey, W. C. Willett, J. N. Ruskin, and J. E. Manson. 1998. Fish consumption and risk of sudden cardiac death. JAMA. 279:23-28.
AOAC. 1992. Official Methods of Analysis. 3rd ed. Assoc. Anal. Chem., Arlington, VA.
AOAC. 2005. Official Methods of Analysis. 18th ed. Assoc. Anal. Chem., Arlington, VA.
Appel, L. J., E. R. Miller 3rd, and A. J. Seidler. 1993. Does supplementation of diet with ‘fish oil’ reduce blood pressure? A meta-analysis of controlled clinical trials. Arch. Intern. Med. 153:1429-1438.

58
Araujo, D. B., R. F. Cooke, G. R. Hansen, C. R. Staples, and J. D. Arthington. 2010. Effects of rumen-protected polyunsaturated fatty acid supplementation on performance and physiological responses of growing cattle following transportation and feedlot entry. J. Anim. Sci. 88:4120-4132.
Ashes, J. R., B. D. Siebert, S. K. Gulati, A. Z. Cuthbertson, and T. W. Scott. 1992. Incorporation of n−3 fatty acids of fish oil into tissue and serum lipids of ruminants. Lipids. 8:629-631.
Barsanti, L., and P. Gualtieri. 2006. Algae: Anatomy, Biochemistry, and Biotechnology. CRC Press - Boca Raton, FL.
Bauman, D. E., and J. M. Griinari. 2003. Nutritional regulation of milk fat synthesis. Annu. Rev. Nutr. 23:203-227.
Baumgard, L. H., B. A. Corl, D. A. Dwyer, A. Saebo, and D. E. Bauman. 2000. Identification of the conjugated linoleic acid isomer that inhibits milk fat synthesis. Am. J. Physiol. 278:179-184.
Bittiner, S. B., I. Cartwright, W. F. G. Tucker, and S. S. Bleehen. 1988. A double-blind randomized placebo-controlled trial of fish oil in psoriasis. Lancet. 1:378-380.
Blankson, H., J. A. Stakkestad, H. Fagertun, E. Thom, J. Wadstein, and O. Gudmundsen. 2000. Conjugated linoleic acid reduces body fat mass in overweight and obese humans. J. Nutr. 130:2943-2948.
Bock, B. J., D. L. Harmon, R. T. Brandt Jr., and J. E. Schneider. 1991. Fat source and calcium level effects on finishing steer performance, digestion, and metabolism. J. Anim. Sci. 69:2211-2224.
Boeckaert, C., V. Fievez, D. Van Hecke, W. Verstraete, and N. Boon. 2007. Changes in rumen biohydrogenation intermediates and ciliate protozoa diversity after algae supplementation to dairy cattle. Eur. J. Lipid. Sci. Technol. 109:767-777.
Boeckaert, C., B. Vlaeminck, J. Dijkstra, A. Issa-Zacharia, T. Van Nespen, W. Van Straalen, and V. Fievez. 2008. Effect of dietary starch or micro algae supplementation on rumen fermentation and milk fatty acid composition of dairy cows. J. Dairy Sci. 91:4714-4727.
Broderick, G. A., R. J. Wallace, and E. R. Orskov. 1991. Control of rate and extent of protein degradation. Physiological Aspects of Digestion and Metabolism in Ruminants. Academic Press Inc. San Diego, CA. p. 541-594.

59
Broderick, G. A., N. D. Luchini, S. M. Reynal, G. A. Varga, and V. A. Ishler. 2008. Effect on production of replacing dietary starch with sucrose in lactating dairy cows. J. Dairy Sci. 91:4801-4810.
Bronte-Stewart, B., A. Antonis, L. Eales, and J. F. Brock. 1956. Effects of feeding different fats on serum-cholesterol level. Lancet. 1:521.
Byers, F.M., and G. T. Schelling. 1988. Lipids in ruminant nutrition. Ruminant Animal, Digestive Physiology and Nutrition. 298-312.
Buchanan-Smith, J. G., R. Totusek, and A. D. Tillman. 1968. Effect of methods of processing on digestibility and utilization of grain sorghum by cattle and sheep. J. Anim. Sci. 27:525.
Chalupa, W., B. Rickabaugh, D. S. Kronfeld, and D. Sklan. 1984. Rumen fermentation in vitro are influenced by long-chain fatty acids. J. Dairy Sci. 67:1439.
Chan, C. B., P. E. MacDonald, M. C. Saleh, D. C. Johns, E. Marban, and M. B. Wheeler. 1999. Overexpression of uncoupling protein 2 inhibits glucose-stimulated insulin secretion from rat islets. Diabetes. 48:1482-1486.
Cheng, K.J., C. W. Forsberg, H. Mnato, and J. W. Costerton. 1991. Microbial ecology and physiology of feed degradation without the rumen. Physiological Aspects of Digestion and Metabolism in Ruminants: Proceedings of the Seventh International Symposium on Ruminant Physiology. 595.
Chilliard, Y., and F. Bocquier. 1993. Effects of fat supplementation on milk yield and composition in dairy goats and ewes. La Qualita nelle Produzioni dei Piccoli Ruminanti: 61-78.
Chilliard, Y., F. Glasser, A. Ferlay, L. Bernard, J. Roueland, and M. Doreau. 2007. Diet, rumen biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid Sci. Technol. 109:828-855.
Choi, C. W., S. Ahvenjarvi, A. Vanhatalo, V. Toivonen, and P. Huhtanen. 2002. Quantitation of the flow of soluble non-ammonia nitrogen entering the omasal canal of dairy cows fed grass silage based diets. Animal Feed Science and Technology. 96:203-220.
Chow, T. T., V. Fievez, A. P. Moloney, K. Raes, D. Demeyer, and S. De Smet. 2004. Effect of fish oil on in vitro rumen lipolysis apparent biohydrogenation of linoleic and linolenic acid and accumulation of biohydrogenation intermediates. Anim. Feed Sci. Technol. 117: 1–12.

60
Church, D. C. 1988. The ruminant animal. Digestive physiology and nutrition of ruminants. 2nd ed. Academic Press, Englewood Cliffs, NJ. p. 298-312.
Cole, N. A., R. R. Johnson, and F. N. Owens. 1976. Influence of roughage level and corn processing method on the site and extent of digestion by beef steers. J. Anim. Sci. 43:490.
Cooke, R. F., D. W. Bohnert, P. Moriel, B. W. Hess, and R. R. Mills. 2011. Effects of polyunsaturated fatty acid supplementation on ruminal in situ forage degradability, performance, and physiological responses of feeder cattle. J. Anim. Sci. 89:3677-3689.
Corl, B. A., L. H. Baumgard, D. A. Dwyer, J. M. Griinari, B. S. Phillips, and D. E. Bauman. 2001. The role of Δ9-desaturase in the production of cis-9, trans-11 CLA. J. Nutr. Biochem. 12:622-630.
Crawford, M. A., K. Costeloe, K. Ghebremeskel, A. Phylactos, L. Skirvin, and F. Stacey. 1997. Are deficits of arachidonic and docosahexaenoic acids responsible for the neural and vascular complications of preterm babies? Am. J. Clin. Nutr. 66:1032-1041.
Dawson, R. M. C., and P. Kemp. 1967. The aminoethylphosphonate-containing lipids of rumen protozoa. Biochem. J. 105:837-842.
De Caterina, R., M. I. Cybulsky, S. K. Clinton, M. A. Gimbrone Jr., and P. Libby. 1994. The omega-3 fatty acid docosahexaenoic reduces cytokine-induced expression of proatherogenic and proinflammatory proteins in human endothelial cells. Arterioscler. Thromb. Vasc. Biol. 14:1829-1836.
Demeyer, D. I., and C. J. Van Nevel. 1995. Transformations and effects of lipids in the rumen: three decades of research at Gent University. Archives of Animal Nutrition. 48:119-134.
Devillard, E., F. M. McIntosh, C. J. Newbold, and R. J. Wallace. 2006. Rumen ciliate protozoa contain high concentrations of conjugated linoleic acids and vaccenic acid, yet do not hydrogenate linoleic acid or desaturate stearic acid. Br. J. Nutr. 96:697-704.
Doreau, M., F. Legay, and D. Bauchart. 1991.Effect of Source and level of Supplemental Fat on Total and Ruminal Organic Matter and Nitrogen Digestion in Dairy Cows. J. Dairy. Sci. 74:2233-2242.

61
Doreau, M., and Y. Chilliard. 1997. Effects of ruminal or post ruminal fish oil supplementation on intake and digestion in dairy cows. Reproduction Nutrition Development. 37:113-124.
Duckett, S. K., J. G. Andrae, and F. N. Owens. 2002. Effect of high-oil corn or added corn oil on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finishing diets. J. Anim. Sci. 80:3353-3360.
Duckett, S. K., and M. H. Gillis. 2010. Effects of oil source and fish oil addition on ruminal biohydrogenation of fatty acids and conjugated linoleic acid formation in beef steers fed finishing diets. J. Anim. Sci. 88:2684-2691.
Engler, M. M. 1996. γ-Linolenic acid: a potent blood pressure lowering nutrient. γ-Linolenic acid: metabolism and its roles in nutrition and medicine. AOCS Press. 200-217.
Fleury, C., M. Neverova, S. Collins, S. Raimbault, O. Champigny, C. Levi-Meyrueis, F. Biuillaud, M. F. Seldin, R. S. Surwit, D. Ricquier, and C. H. Warden. 1997. Uncoupling protein-2: a novel gene linked to obesity and hyperinsulinem Nat. Genet. 15:269–272.
Fredin, S. M., L. F. Ferraretto, M. S. Akins, and R. D. Shaver. 2014. Comparison of omasal and reticular sampling methods on ruminal nutrient outflow and digestion in lactating dairy cows. J. Anim. Sci. 92:1601.
Galbraith, H., T. B. Miller, A. M. Paton, and J. K. Thompson. 1971. Antibacterial activity of long chain fatty acids and the reversal with calcium, magnesium, ergocalciferol and cholesterol. J. Appl. Bacteriol. 34:803.
Galyean, M. L., D. G. Wagner, and R. R. Johnson. 1976. Site and extent of starch digestion in steers fed processed corn rations. J. Anim. Sci. 43:1088.
Garcia, J. A., C. B, Theurer, and W. H. Hale. 1981a. Digestao ruminal e pos-ruminal do amido, em novilhos alimentados corn dois niveis de grao de sorgo floculado. Revista da Sociedade Brasileira de Zootecnia.10:235.
Glasser, F., M. Doreau, A. Ferlay, J. J. Loor, and Y. Chilliard. 2007. Milk fatty acids: Mammary synthesis could limit transfer from duodenum in cows. Eur. J. Lipid Sci. Technol. 109:817-827.
Griinari, J. M., L. Granlund, J. I. Pedersen, P. Delmonte, K. Shingfield, and A. Saebo. 2005. Cis-10, trans-12 conjugated linoleic acid inhibits lipid accumulation during adipocyte differentiation. In: Proceedings of the International Society of Fat Research.

62
Gutknecht, J. 1988. Proton conductance caused by long-chain fatty acids in phospholipid bilayer membranes. J. Membr. Biol. 106:83.
Harfoot, C. G., R. C. Noble, and J. H. Moore. 1973. Food particles as a site for biohydrogenation of unsaturated fatty acids in the rumen. Biochem. J. 132:829-832.
Harfoot, C.G., and G. P. Hazlewood. 1988. Lipid Metabolism in the Rumen, in The
Rumen Microbial Ecosystem. Elsevier Applied Science. 285-322. Harfoot, C. G. and G. P. Hazlewood. 1997. Lipid Metabolism in the Rumen. The Rumen
Microbial Ecosystem. 382-426. Harris, W. S. 1997. N-3 Fatty acids and serum lipoproteins: human studies. Am. J. Clin.
Nutr. 65:1645-1654. Hawthorne, A. B., T. K. Daneshmend, C. J. Hawkey, A. Belluzzi, S. J. Everitt, G. K.
Holmes, C. Malkinson, M. Z. Shaheen, and J. E. Willars. 1992. Treatment of ulcerative colitis with fish oil supplementation: a prospective 12- month randomized controlled trial. Gut. 33:922-928.
Hess, T. M., J. K. Rexford, D. K. Hansen, M. Harris, N. Schauermann, T. Ross, T. E.
Engle, K. G. D. Allen, and C. M. Mulligan. 2012. J. Anim. Sci. 90:3023-3031. Hibberd, C. A., D. G. Wagner, R. L. Hintz, and D. D. Griffin. 1983. Effect of sorghum
grain variety and processing method on the site and extent of starch digestion in steers. Anim. Sci. Res. 114:28.
Hinman, D. D., and R. R. Johnson. 1974a. Influence of processing methods on digestion
of sorghum starch in high concentrate beef cattle rations, J. Anim. Sci. 39:417. Hinman, D. D, and R. R. Johnson. 1974b. Influence of degree of micronization on the site
and extent of sorghum starch digestion in beef cattle fed high concentrate rations. J. Anim. Sci. 39:958.
Holmes, J.H.G., M. J. Drennan, and W. N. Garrett. 1970. Digestion of steam-processed
milo by ruminants. J. Anim. Sci. 31:409. Hristov, A. N., L. R. Kennington, M. A. McGuire, and C. W. Hunt. 2005. Effect of diets
containing linoleic acid- or oleic acid-rich oils on ruminal fermentation and

63
nutrient digestibility, and performance and fatty acid composition of adipose and muscle tissues of finishing cattle. J. Anim. Sci. 83:1312-1321.
Huhtanen, P., P. G. Brotz, and L. D. Satter. 1997. Omasal sampling technique for
assessing fermentative digestion in the forestomach of dairy cows. J. Anim. Sci. 75:1380-1392.
Huws, S. A.,M. R. F. Lee, S. M. Muetzel, M. B. Scott, R. J. Wallace, and N. D. Scollan.
2010. Forage type and fish oil cause shifts in rumen bacterial. Microbiol. Ecol. 73:396-407.
Immig, V. I., S. J. Wirth, G. A. Wolf, and H. Abel. 1991. Quantifizierung der
Cellulaseaktivitat und Nachweis von Fettsaure-Coating-Effekten im Pansen von Schafen. J. Anim. Physiol. Anim. Nutr. 66:45.
Jenkins, T. C. 1993. Lipid metabolism in the rumen. Journal of Dairy Science. 76:3851-
3863. Kalscheur, K. F., B. B. Teter, L. S. Piperova, and R. A. Erdman. 1997. Effect of dietary
forage concentration and buffer addition on duodenal flow of trans-C-18:1 fatty acids and milk fat production in dairy cows. J. Dairy Sci. 80:2104-2114.
Kang, J. X., and A. Leaf. 1994. Effects of long chain polyunsaturated fatty acids on the contraction of neonatal rat cardiac myocytes. Proc. Nat. Acad. Sci. 91:9886-9890.
Keim, N. L., J. L. Sebedio, W. W. Christie, and R. Adolf. 2003. Advances in conjugated linoleic acid research. AOCS Press. 2:316-324.
Keys, A., J. T. Anderson, and F. Grande. 1965. Serum cholesterol response to change in diet. I. Iodine value of dietary fat verses 2S-P. Metabolism. 14:747.
Knowlton, K. F., B. P. Gleen, and R. A. Erdman. 1998. Performance, ruminal fermentation, and site of starch digestion in early lactation cows fed corn grain harvested and processed differently. J. Dairy Sci. 81:1972–1984.
Kim, Y. Z., R. H. Lui, D. R. Bond, and J. B. Russel. 2000. Effect of linoleic acid concentration on conjugated linoleic acid produced by Butyrivibrio fibrisolvens A38. Applied and Environmental Microbiology. 66:5226-5230.
Kitessa, S. M., S. K. Gulati, J. R. Ashes, E. Fleck, T. W. Scott, and P. D. Nichols. 2001. Utilization of fish oil in ruminants: Fish oil metabolism in sheep. Animal Feed Science and Technology. 89:189-199.

64
Kozloski, G. V. 2011. Bioquimica de Ruminantes. 3th ed. Academic Press, Santa Maria, RG – Brazil. p. 37-44.
Kronberg, S. L., E. J. Scholljegerdes, A. N. Lepper, and E. P. Berg. 2011. The effect of flaxseed supplementation on growth, carcass characteristics, fatty acid profile, retail shelf life, and sensory characteristics of beef from steers finished on grasslands of the northern Great Plains. J. Anim. Sci. 89:2892-2903.
Kucuk, O., B. W. Hess, and D. C. Rule. 2004. Soybean oil supplementation of a high-concentrate diet does not affect site and extent of organic matter, starch, neutral detergent fiber, or nitrogen digestion, but influences both ruminal metabolism and intestinal flow of fatty acids in limit-fed lambs. J. Anim. Sci. 82:2985-2994.
LaBrune, H. J. 2000. The effects of grain processing and lipid addition on blood glucose, performance, carcass characteristics, and meat quality in finishing cattle. M.S. Thesis, Kansas State Univ., Manhattan.
Leat, W. M. F. 1977. Depot fatty acids of aberdeen Angus and friesian cattle reared in hay and barley diets. J. Agric. Sci. Camb. 89:575.
Lee, M. R. F, J. K. S. Tweed, A. P. Moloney, and N. D. Scollan. 2005. The effects of fish oil supplementation on rumen metabolism and the biohydrogenation of unsaturated fatty acids in beef steers given diets containing sunflower oil. Anim. Sci. 80:361-367.
Lee, M. R. F., K. J. Shingfield, J. K. S. Tweed, V. Toivonen, S. A. Huws, and N. D. Scollan. 2008. Effect of fish oil on ruminal biohydrogenation of C18 unsaturated fatty acids in steers fed grass or red clover silages. Animal. 12:1859-1869.
Lodge-Ivey, S. L., L. N. Tracey, and A. Salazar. 2013. The utility of lipid extract algae as a protein source in forage or starch-based ruminant diets. J. Anim. Sci. 92:1331-1342.
Loor, J. J., K. Ueda, A. Ferlay, Y. Chilliard, and M. Doreau. 2005a. Intestinal flow and digestibility of trans fatty acids and conjugated linoleic acids (CLA) in dairy cows fed a high concentrate diet supplemented with fish oil, linseed oil, or sunflower oil. Anim. Feed Sci. Tech. 119:203-225.
Loor, J. J., A. Ferlay, A. Ollier, K. Ueda, M. Doreau, and Y. Chilliard. 2005b. High- concentrate diets and polyunsaturated oils alter trans and conjugated isomers in bovine rumen, blood, and milk. J. Dairy Sci. 88:3986-3999.

65
Lundy, III, F. P., E. Block, W. C. Bridges Jr., J. A. Bertrand, and T. C. Jenkins. 2004. Ruminal biohydrogenation in Holstein cows fed soybean fatty acids as amides or calcium salts. J. Dairy Sci. 87:1038-1046.
Luvisetto, S., D. Pietrobon, and G. F. Azzone. 1987. Uncoupling of oxidative phosphorylation: 1. Protonophoric effects account only partially for un-coupling. Biochemistry. 26:7332.
Maia, M. R. G., L. C. Chaudhary, L. Figueres, and R. J. Wallace. 2007. Metabolism of polyunsaturated fatty acids and their toxicity to the microflora of the rumen. J. Gen. Mol. Microbiol. 91:303-314.
Martinetz, M. 1992. Tissue levels of polyunsaturated fatty acids during early human development. J. Pediatr. 120:129-138.
May, M. L., M. J. Quinn, C. D. Reinhardt, L. Murray, M. L. Gibson, K. K. Karges, and J. S. Drouillard. 2009. Effects of dry-rolled or steam-flaked corn finishing diets with or without twenty-five percent dried distillers grains on ruminal fermentation and apparent total tract digestion. J. Anim. Sci. 87:3630-3638.
McNeill, J. W., G. D. Potter, and J. K. Riggs. 1971. Ruminal and post ruminal carbohydrate utilization in steers fed processed sorghum grain. J. Anim. Sci. 33:1371.
Mir, P. S., C. M. Kalnin, and S. A. Garvey. 1989. Recovery of fecal chromium used as a digestibility marker in cattle. J. Dairy Sci. 72:2549-2553.
Montgomery, S. P., J. S. Drouillard, T. G. Nagaraja, E. C. Titgemeyer, and J. J. Sindt. 2014. Effects of supplemental fat source on nutrient digestion and ruminal fermentation in steers. J. Anim. Sci. 2008. 86:640-650.
Morris, M. C., F. Sacks, and B. Rosner. 1993. Does fish oil lower blood pressure? A meta-analysis of controlled trials. Circulation. 88:523–533.
Neuringer, M., G. J. Anderson, and W. E. Connor. 1988. The essentiality of n-3 fatty acids for the development and function of the retina and brain. Annu. Rev. Nutr. 8:517-541.
Nicholson, J. W., G. E. Charmley, and R. S. Bush. 1992. The effect of supplemental protein source on ammonia levels in rumen fluid and blood and intake of alfalfa silage by beef cattle. Can. J. Anim. Sci. 72:853-862.
Offer, N. W., M. Marsden, J. Dixon, B. K. Speake, and F. E. Thacker. 1999. Effect of dietary fat supplements on levels of n-3 poly-unsaturated fatty acids, trans acids and conjugated linoleic acid in bovine milk. J. Anim. Sci. 69:613-625.

66
Onetti, S. G., R. D. Shaver, M. A. McGuire, and R. R. Grummer. 2001. Effect of type and level of dietary fat on rumen fermentation and performance of dairy cows fed corn silage-based diets. J. Dairy Sci. 84:2751-2759.
Onetti, S. G., R. D. Shaver, M. A. McGuire, D. L. Palmquist, and R. R. Grummer. 2002. Effect of supplemental tallow on performance of dairy cows fed diets with different corn silage: alfalfa silage ratios. J. Dairy Sci. 85:632-641.
Onetti, S.G., S. M. Reynal, and R. R. Grummer. 2004. Effect of alfalfa forage preservation method and particle length on performance of dairy cows fed corn silage-based diets and tallow. J. Dairy Sci. 87:652-664.
Or-Rashid, M. M., J. K. G. Kramer, M. A. Wood, and B. W. McBride. 2008. Supplemental algal meal alters the ruminal trans-18:1 fatty acid and conjugated linoleic acid composition in cattle. J. Anim. Sci. 86:187-196.
Orskov, E. R. 1986. Starch digestion and utilization in ruminants. J. Anim. Sci. 63:1624-1633.
Paillard, D., N. McKain, M. T. Rincon, K. J. Shingfield, D. I. Givens, and R. J. Wallace. 2007. Quantification of ruminal Clostridium proteoclasticum by real-time PCR using a molecular beacon approach. J. Appl. Microbiol. 103:1251-1261.
Palmquist, D. L., and J. M. Griinari. 2006. Milk fatty acid composition in response to reciprocal combinations of sunflower and fish oils in the diet. Anim. Feed Sci. Tech. 131:358-369.
Pariza, M. W., Y. Park, and M. E. Cook. 2000. Mechanisms of action of conjugated linoleic acid: evidence and speculation. Proceedings of Society for Experimental Biology in Medicine. 223:8-13.
Park, Y. J., M. Storkson, W. Liu, K. J. Albright, M. E. Cook, and M. W. Pariza. 2004. Structure-activity relationship of conjugated linoleic acid and its cognates in inhibiting heparin-releasable lipoprotein lipase and glycerol release from fully differentiated 3T3-L1 adipocytes. Nutr. Biochem.15:561-568.
Piperova, L. S., B. B. Teter, I. Bruckental, J. Sampugna, S. E. Mills, M. P. Yurawecz, J. Fritsche, K. Ku, and R. A. Erdman. 2000. Mammary lipogenic enzyme activity, trans fatty acids and conjugated linoleic acids are altered in lactating dairy cows fed a milk fat-depressing diet. J. Nutr. 130:2568-2574.
Piperova, L. S., J. Sampugna, B. B. Teter, K. F. Kalscheur, M. P. Yurawecz, Y. Ku, K. M. Morehouse and R. A. Erdman. 2002. Duodenal and milk trans octadecenoic acid and conjugated linoleic acid (CLA) isomers indicate that post absorptive

67
synthesis is the predominant source of cis-9-containing CLA in lactating dairy cows. J Nutr. 132: 1235–1241.
Polan, C. E., S. B. Tove, and J. J. McNeill. 1964. Biohydrogenation of unsaturated fatty acids by rumen bacteria. J. Bacteriol. 88:1056-1064.
Punia, B. S., and J. Leibholz. 1994. Effect of level of intake of kikuyu (Pennisetum clandestinum) grass hay on the flow of protozoal nitrogen to the omasum of cattle. Anim. Feed Sci. Tech. 47:77.
Reynal, S. M., and G. A. Broderick. 2005. Effect of dietary level of rumen-degraded protein on production and nitrogen metabolism in lactating dairy cows. J. Dairy Sci. 88:4045-4064.
Ruppert, L. D., J. K. Drackley, D. R. Bremmer, and J. H. Clark. 2003. Effects of tallow in diets based on corn silage or alfalfa silage on digestion and nutrient use by lactating dairy cows. J. Dairy Sci. 86:593-609.
Saebo, A., P. Saebo, J. M. Griinari and K. J. Shingfield. 2005a. Effect of abomasal infusion of geometric isomers of 10,12 conjugated linoleic acid on milk fat synthesis in dairy cows. Lipids. 40:823-832.
Saebo, A., J. W. Perfield II, P. Delmonte, M. P. Yurawecz, P. Lawrence, J. T. Brenna, and D. E. Bauman. 2005b. Milk fat synthesis is unaffected by abomasal infusion of the conjugated diene 18:3 isomers cis-6, trans-10, cis-12 and cis-6, trans-8, cis- 12. Lipids. 40:89-95.
Scollan, N. D., N. I. Choi, E. Kurt, A. V. Fisher, M. Enser, and J. D. Wood. 2001. Manipulating the fatty acid composition of muscle and adipose tissue in beef cattle. Br. J. Nutr. 85:115-124.
Shingfield, K. J., S. Ahvenjarvi1, V. Toivonen, A. Arola, K. V. V. Nurmela, P. Huhtanen, and J. M. Griinari. 2003. Effect of dietary fish oil on biohydrogenation of fatty acids and milk fatty acid content in cows. J. Anim. Sci. 77:165-179.
Shingfield, K. J., and J. M. Griinari. 2007. Role of biohydrogenation intermediates in milk fat depression. Eur. J. Lipid Sci. Technol. 109:799-816.
Simopoulos, A. P. 1991. Omega-3 fatty acids in health and disease and in growth and development. Am. J. Clin. Nutr. 54:438-63.
Simopoulos, A. P., and L. G. Cleland. 2003. Omega-6/omega-3 essential fatty acid ratio: the scientific evidence World. Nutr. Diet. 92.

68
Simopoulos, A. P. 2006. Evolutionary aspects of diet, the omega-6/omega-3 ratio and genetic variation: nutritional implications for chronic diseases. Biomedicine & Pharmacotherapy. 60:502-507.
Streeter, M. N., D. G. Wagner, F. N. Owens, and C. A. Hibberd. 1989. Combinations of high-moisture harvested sorghum grain and dry-rolled corn: effects on site and extent of digestion in beef heifers. J. Anim. Sci. 67:1623-1633.
Smith, R. H. 1984. Microbial activity in the omasum. Proc. Nutr. Soc. 43:63.
Smith, W.A., B. Harris, Jr., H.H. Van Horn, and C.J. Wilcox. 1993. Effects of forage type on production of dairy cows supplemented with whole cottonseed, tallow, and yeast. J. Dairy Sci. 76:205-215.
Song, H. J., I. Grant, D. Rotondo, I. Mohede, N. Sattar, and S. D. Heys. 2004. Effect of CLA supplementation on immune function in young healthy volunteers. Eur. J. Clin. Nutr. 59:508-517.
Staples, C. R, and F. M. Cullens. 2005. Implications of fat-feeding practices for lactating dairy cows – effects on milk fat. Advances in Dairy Technology. 17:277-295.
Sukhija, P. S., and D. L. Palmquist. 1988. Rapid method for determination of total fatty acid content and composition of feedstuffs and feces. J. Agric. and Food Chem. 36:1202-1206.
Sutton, J. D., R. Knight, A. B. McAllan, and R. H. Smith. 1983. Digestion and synthesis in the rumen of sheep given diets supplemented with free and protected oils. Br. J. Nutr. 49: 419-432.
Torres-Duarte, A. P., and J. Y. Vander Hoek. 2003. Conjugated linoleic acid exhibits stimulatory and inhibitory effects on prostanoid production in human endothelial cells and platelets. Biochim. Biophys. Acta. Molec. Cell. Res. 1640:69-76.
Van Emon, M. L., D. D. Loy, and S. L. Hansen. 2015. Determining the preference, in vitro digestibility, in situ disappearance, and grower period performance of steers fed a novel algae meal derived from heterotrophic microalgae. J. Anim. Sci. 93:3121-3129.
Van Nevel, C. J, and D. I. Demeyer. 1996. Influence of pH on lipolysis and biohydrogenation of soybean oil by rumen contents in vitro. Reprod. Nutr. Dev. 36:53-63.
Vartiainen, E., T. Laatikainen, and M. Peltonen. 2010. Thirty-five-year trends in
cardiovascular risk factors in Finland. Int. J. Epidemiol. 39:504-518.

69
Vlaeminck, B., G. Mengistu, V. Fievez, L. de Jonge, and J. Dijkstra. 2007. Effect of in vitro docosahexaenoic acid supplementation to marine algae-adapted and unadapted rumen inoculum on the biohydrogenation of unsaturated fatty acids in freeze-dried grass. J. Dairy Sci. 91:1122–1132
Wahle, K. W. J., S. D. Heys, and D. Rorondo. 2004. Conjugated linoleic acids: Are they beneficial or detrimental to health? Prog. Lipid Res. 43:553-587.
Ward, P. F. V., T. W. Scott, and R. M. C. Dawson. 1964. The hydrogenation of
unsaturated fatty acids in the ovine digestive tract. Biochem. J. 9250. Wasowska, I., M. R. G. Maia, K. M. Niedzwiedzka, M. Czauderna, J. M. C. R. Ribeiro,
E. Devillard, K. J. Shingfield, and R. J. Wallace. 2006. Influence of fish oil on ruminal biohydrogenation of C18 unsaturated fatty acids. Br. J. Nutr. 95:1199-1211.
Wenham, G., and R. S. Wyburn. 1980. A radiological investigation of the effects of
cannulation on intestinal motility and digesta flow in sheep. Camb. J. Agric. Sci. 95:539.
Williams, C. H., D. J. David, and O. Iisma. 1962. The determination of chromic oxide in
feces samples by atomic absorption spectrophotometry. J. Agr. Sci. 59:381-385. Wistuba, T. J., E. B. Kegley, and J. K. Apple. 2006. Influence of fish oil in finishing diets
on growth performance, carcass characteristics, and sensory evaluation of cattle. J. Anim. Sci. 84:902-909.
Wu, Z., 0. A. Ohajuruka, and D. L. Palmquist. 1991. Ruminal synthesis,
biohydrogenation, and digestibility of fatty acids by dairy cows. J. Dairy Sci. 74:3025.
Xiao C., A. Giacca, A. Carpentier, and G. F. Lewis. 2006. Differential effects of
monounsaturated, polyunsaturated and saturated fat ingestion on glucose-stimulated insulin secretion, sensitivity and clearance in overweight and obese, non-diabetic humans. Diabetologia 49:1371-1379.
Zinn, R. A. 1987. Influence of lasalocid and monensin plus tylosin on comparative
feeding value of steam-flaked versus dry-rolled corn in diets for feedlot cattle. J. Anim. Sci .65:256-266.

70
Zinn, R. A., and A. Plascencia. 1993. Interaction of whole cottonseed and supplemental fat on digestive function in cattle. J. Anim. Sci. 71:11–17.
Zinn, R. A., C. F. Adam, and M. S. Tamayo. 1995. Interaction of feed intake level on comparative ruminal and total tract digestion of dry-rolled and steam-flaked corn. J. Anim. Sci. 73:1239-1245.

71
VITA
Birthplace: São Paulo - SP, Brazil
Academic Background:
2013 - 2016 M.S. Graduate Research Assistant
University of Kentucky - Lexington/KY
Animal & Food Sciences - Ruminant Nutrition
Adviser David L. Harmon, PhD
2008 - 2013: Universidade Estadual de Maringa – Maringa/PR – Brazil
Bachelor of Science – Zootecnia / Animal & Food Sciences
Adviser Antonio Ferriani Branco, PhD
Awards:
2013 Alltech Young Scientist Competition
Global 1st Place Undergraduate winner
North America Region 1st Place Undergraduate winner
Research Projects:
2012: Bovine lateral saphenous veins exposed to ergopeptine alkaloids do not relax.
University of Kentucky / USDA
Advisers: Dr. David L. Harmon - University of Kentucky and Dr. James L. Klotz – USDA
2010 - 2011: Levels of substitution of feed grade urea for optigen in the diet of confined
sheep: microbial efficiency.
Universidade Estadual de Maringa – Brazil
Adviser Antonio Ferriani Branco, phD
2010 - 2011: Sugar cane silage treated or not with different non-protein nitrogen sources:

72
uric plasmatic nitrogen.
Universidade Estadual de Maringa– Brazil
Adviser Antonio Ferriani Branco, phD
2009 - 2010: Evaluation of two propolis production systems of Apis mellifera, and the correlation between the hygienic behavior and the Varroa destructor infestation in the swarms.
Universidade Estadual de Maringa– Brazil
Adviser Lucimar Pontara Peres de Moura, phD
Publications:
Pesqueira, A., D. L. Harmon, A. F. Branco, and J. L. Klotz. 2014. Bovine lateral saphenous veins exposed to ergopeptine alkaloids do not relax. J. Anim. Sci. 92:1213-1218.
Amanda Pesqueira
Related Documents











