Eur. J. Biochem. 227, 715-722 (1995) 0 FEBS 1995 Hepatic fatty acid metabolism as a determinant of plasma and liver triacylglycerol levels Studies on tetradecylthioacetic and tetradecylthiopropionic acids Daniel K. ASIEDU’, Ayman AL-SHURBAJI’, A d d C. RUSTAN3, Ingemar BJORKHJ5M2,Lars BERGLUND’ and Rolf K. BERGE’ ’ Institute of Clinical Biology, Division of Biochemistry, University of Bergen, Haukeland University Hospital, Norway * Department of Clinical Chemistry Karolinska Institute, Huddinge University Hospital, Sweden ’ Department of Pharmacology, Institute of Pharmacy, University of Oslo, Norway (Received 21 October 1994) - EJB 94 1616/5 To investigate the importance of factors influencing substrate availability for triacylglycerol biosynthe- sis on lipoprotein metabolism, the effects of two opposite-acting sulphur-substituted fatty acid analogues, tetradecylthioacetic acid and tetradecylthiopropionic acid were studied. Administration of tetradecylthio- acetic acid to rats resulted in a reduction of plasma levels of triacylglycerols (44%) and cholesterol (26 %). This was accompanied by a reduction in very-low-density lipoprotein (VLDL) triacylglycerols (48 %), VLDL cholesterol (36% j, low-density lipoprotein (LDL) cholesterol (36 %) and high-density lipo- protein (HDL) triacylglycerols (50 %), whereas HDL cholesterol levels did not change. Subsequently, the HDLLDL-cholesterol ratio increased by 40 %. The cholesterol-lowering effect was accompanied by a reduction in hydroxymethylglutaryl CoA (HMG-CoA) reductase activity (37 %). Both mitochondrial and peroxisomal fatty acid oxidation increased (1.7-fold and 5.3-fold, respectively). Furthermore, there was a significant negative correlation between plasma triacylglycerols and mitochondrial fatty acid oxidation. Hepatic triacylglycerol synthesis was retarded, as indicated by a decrease in VLDL triacylglycerol secre- tion (40%), and by a reduced liver triacylglycerol content (29%). The activities of lipoprotein lipase and hepatic lipase in post-heparin plasma were not affected. Microsomal and cytosolic phosphatidate phosphohydrolase activities were inhibited (28 % and 70 %, respectively j. Hepatic malonyl-CoA levels decreased by 29% and the total activity of acetyl-CoA carboxylase was reduced (23%). In hepatocytes treated with tetradecylthioacetic acid, mitochondrial fatty acid oxidation increased markedly (100 %) and triacylglycerol secretion was reduced (40%). In tetradecylthiopropionic-acid-treated rats, a significant increase in both plasma and VLDL triacylglycerols was found (46 % and 72 %, respectively) but VLDL triacylglycerol secretion was unaffected. However, no effect on either plasma or lipoprotein cholesterol levels was seen. Mitochondrial fatty acid oxidation was decreased by 50 % and hepatic triacylglycerol levels increased by 33 %. In hepatocytes exposed to tetradecylthiopropionic acid, triacylglycerol synthesis increased (100 %) while triacylglycerol secretion and fatty acid oxidation remained unaltered. The results illustrate that lipoprotein triacylglycerol levels can be modulated by changes in the availability of fatty acid substrate for triacylglycerol biosynthesis, mainly by affecting mitochondrial fatty acid oxidation. In addition, we demonstrate that suppression of rat hepatic HMG-CoA reductase activity during treatment with tetradecylthioacetic acid may contribute to a cholesterol-lowering effect. Keywords. Very-low-density lipoprotein; hypolipidermic drug ; triacylglycerol and cholesterol synthesis ; lipogenic enzymes ; lipoprotein and hepatic lipases ; hydroxymethylglutaryl CoA reductase ; /?-oxidation. In the liver, mitochondrial fatty acid oxidation and triacyl- glycerol biosynthesis are the major competitors for the utiliza- tion of fatty acids as substrate [l, 21. The regulatory interrela- tionship between /?-oxidation and triacylglycerol biosynthesis Correspondence to D. K. Asiedu, Institute of Clinical Biology, Divi- sion of Biochemistry, University of Bergen, Haukeland University Hos- pital, N-5021 Bergen, Norway Abbreviations. ACAT, acyl-CoA :cholesterol acyltransferase ; HDL, high-density lipoprotein ; LDL, low-density lipoprotein ; VLDL, very- low-density lipoprotein ; HMGCoA, hydroxymethylglutaryl CoA. Enzymes. Acetyl-CoA carboxylase (EC 6.4.1.2) ; acyl-CoA:choles- terol acyltransferase (EC 2.3.1.26); ATP:citrate lyase (EC 4.1.3.8); cho- lesterol 7a-hydroxylase (EC 1.14.13.17); fatty acyl-CoA oxidase (EC 1.3.3.6); hepatic lipase (EC 3.1.1.34); HMG-CoA reductase (EC 1.1.1.88); lipoprotein lipase (EC 3.1.1.34); phosphatidate phosphohy- drolase (EC 3.1.3.4). has so far not been elucidated in detail. Growing evidence indi- cates that availability of triacylglycerols is a major driving force in the secretion of triacylglycerol-rich lipoproteins by the liver [3 -51. It is, therefore, conceivable that factors influencing tri- acylglycerol biosynthesis or fatty acid oxidation may ultimately influence plasma lipoprotein levels and metabolism. Sulphur- substituted fatty acids that profoundly affect /?-oxidation, have facilitated studies on the concerted regulation of fatty acid oxi- dation and triacylglycerol biosynthesis [6- 101. We have demon- strated that stimulation of P-oxidation may affect both triacyl- glycerol formation and plasma lipoprotein homeostasis under both normolipidemic and hyperlipidemic conditions [ll, 121. The different plasma lipoproteins are connected in a complex metabolic interrelationship. Administration of agents such as sulphur-substituted fatty acids that affect triacylglycerol forma-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 227, 715-722 (1995) 0 FEBS 1995
Hepatic fatty acid metabolism as a determinant of plasma and liver triacylglycerol levels Studies on tetradecylthioacetic and tetradecylthiopropionic acids
Daniel K. ASIEDU’, Ayman AL-SHURBAJI’, A d d C. RUSTAN3, Ingemar BJORKHJ5M2, Lars BERGLUND’ and Rolf K. BERGE’
’ Institute of Clinical Biology, Division of Biochemistry, University of Bergen, Haukeland University Hospital, Norway * Department of Clinical Chemistry Karolinska Institute, Huddinge University Hospital, Sweden ’ Department of Pharmacology, Institute of Pharmacy, University of Oslo, Norway
(Received 21 October 1994) - EJB 94 1616/5
To investigate the importance of factors influencing substrate availability for triacylglycerol biosynthe- sis on lipoprotein metabolism, the effects of two opposite-acting sulphur-substituted fatty acid analogues, tetradecylthioacetic acid and tetradecylthiopropionic acid were studied. Administration of tetradecylthio- acetic acid to rats resulted in a reduction of plasma levels of triacylglycerols (44%) and cholesterol (26 %). This was accompanied by a reduction in very-low-density lipoprotein (VLDL) triacylglycerols (48 %), VLDL cholesterol (36% j, low-density lipoprotein (LDL) cholesterol (36 %) and high-density lipo- protein (HDL) triacylglycerols (50 %), whereas HDL cholesterol levels did not change. Subsequently, the HDLLDL-cholesterol ratio increased by 40 %. The cholesterol-lowering effect was accompanied by a reduction in hydroxymethylglutaryl CoA (HMG-CoA) reductase activity (37 %). Both mitochondrial and peroxisomal fatty acid oxidation increased (1.7-fold and 5.3-fold, respectively). Furthermore, there was a significant negative correlation between plasma triacylglycerols and mitochondrial fatty acid oxidation. Hepatic triacylglycerol synthesis was retarded, as indicated by a decrease in VLDL triacylglycerol secre- tion (40%), and by a reduced liver triacylglycerol content (29%). The activities of lipoprotein lipase and hepatic lipase in post-heparin plasma were not affected. Microsomal and cytosolic phosphatidate phosphohydrolase activities were inhibited (28 % and 70 %, respectively j. Hepatic malonyl-CoA levels decreased by 29% and the total activity of acetyl-CoA carboxylase was reduced (23%). In hepatocytes treated with tetradecylthioacetic acid, mitochondrial fatty acid oxidation increased markedly (100 %) and triacylglycerol secretion was reduced (40%). In tetradecylthiopropionic-acid-treated rats, a significant increase in both plasma and VLDL triacylglycerols was found (46 % and 72 %, respectively) but VLDL triacylglycerol secretion was unaffected. However, no effect on either plasma or lipoprotein cholesterol levels was seen. Mitochondrial fatty acid oxidation was decreased by 50 % and hepatic triacylglycerol levels increased by 33 %. In hepatocytes exposed to tetradecylthiopropionic acid, triacylglycerol synthesis increased (100 %) while triacylglycerol secretion and fatty acid oxidation remained unaltered. The results illustrate that lipoprotein triacylglycerol levels can be modulated by changes in the availability of fatty acid substrate for triacylglycerol biosynthesis, mainly by affecting mitochondrial fatty acid oxidation. In addition, we demonstrate that suppression of rat hepatic HMG-CoA reductase activity during treatment with tetradecylthioacetic acid may contribute to a cholesterol-lowering effect.
Keywords. Very-low-density lipoprotein; hypolipidermic drug ; triacylglycerol and cholesterol synthesis ; lipogenic enzymes ; lipoprotein and hepatic lipases ; hydroxymethylglutaryl CoA reductase ; /?-oxidation.
In the liver, mitochondrial fatty acid oxidation and triacyl- glycerol biosynthesis are the major competitors for the utiliza- tion of fatty acids as substrate [l, 21. The regulatory interrela- tionship between /?-oxidation and triacylglycerol biosynthesis
Correspondence to D. K. Asiedu, Institute of Clinical Biology, Divi- sion of Biochemistry, University of Bergen, Haukeland University Hos- pital, N-5021 Bergen, Norway
Abbreviations. ACAT, acyl-CoA :cholesterol acyltransferase ; HDL, high-density lipoprotein ; LDL, low-density lipoprotein ; VLDL, very- low-density lipoprotein ; HMGCoA, hydroxymethylglutaryl CoA.
Enzymes. Acetyl-CoA carboxylase (EC 6.4.1.2) ; acyl-CoA:choles- terol acyltransferase (EC 2.3.1.26); ATP:citrate lyase (EC 4.1.3.8); cho- lesterol 7a-hydroxylase (EC 1.14.13.17); fatty acyl-CoA oxidase (EC 1.3.3.6); hepatic lipase (EC 3.1.1.34); HMG-CoA reductase (EC 1.1.1.88); lipoprotein lipase (EC 3.1.1.34); phosphatidate phosphohy- drolase (EC 3.1.3.4).
has so far not been elucidated in detail. Growing evidence indi- cates that availability of triacylglycerols is a major driving force in the secretion of triacylglycerol-rich lipoproteins by the liver [3 -51. It is, therefore, conceivable that factors influencing tri- acylglycerol biosynthesis or fatty acid oxidation may ultimately influence plasma lipoprotein levels and metabolism. Sulphur- substituted fatty acids that profoundly affect /?-oxidation, have facilitated studies on the concerted regulation of fatty acid oxi- dation and triacylglycerol biosynthesis [6- 101. We have demon- strated that stimulation of P-oxidation may affect both triacyl- glycerol formation and plasma lipoprotein homeostasis under both normolipidemic and hyperlipidemic conditions [ll, 121. The different plasma lipoproteins are connected in a complex metabolic interrelationship. Administration of agents such as sulphur-substituted fatty acids that affect triacylglycerol forma-

716 Asiedu et al. (EUK J. Biochem. 227)
Table 1. Effect of tetradecylthioacetic and tetradecylthiopropionic acids on plasma and lipoprotein lipid levels in rats. Values are mens 2 SEM (n = 6). Means in each row without a common superscript are significantly different at P < 0.05. To convert to mg/dl, multiply triacyl- glycerol by 88.5 and cholesterol by 38.7.
Composition Control Tetradecylthio- Tetradecylthio- acetic acid propionic acid
Plasma triacylglyceride cholesterol
triac ylgl yceride cholesterol
VLDL
LDL triacylgl yceride cholesterol
triac y lglyceride cholesterol
HDL cholesterol/ LDL cholesterol
HDL
1.02 2 0.1 1 a 2.14 20.05"
0.642 0.00" 0.36 2 0.01"
trace 0.89 2 0.21
0.38 20.02" 0.90 2 0.27"
1.01
mmolA
0.57 2 0.29h 1.60 2 O.OOb
0.33 2 0.11 0.23 2 0.01
trace 0.57 i 0.01
0.19i0.01b 0.80 2 0.10"
1.40
1.49 t_ 0.10' 2.2020.10"
1.0920.03' 0.42 2 0.03 a
trace 0.84 f 0.14"
0.40 20.03" 0.94 5 0.24"
1.12
tion, is likely to challenge the homeostasis of this metabolic net- work. In the present study, we have, therefore, investigated the effects of two opposite-acting sulphur-substituted fatty acid ana- logues, tetradecylthioacetic acid [CH,-CH,-(CH,),,-S-CH,- COOH], a 3-thia long-chain fatty acid which is non-/? but o oxidizable, and tetradecylthiopropionic acid [CH,-CH,-(CH,),,- S-CH,-CH,-COOH], a 4-thia P-oxidizable and o-oxidizable fatty acid, on plasma lipoproteins, triacylglycerol formation and triacylglycerol secretion. The results illustrate that the two fatty acid analogues modulate plasma lipoprotein levels by pro- foundly affecting the balance between fatty acid oxidation and glycerolipid biosynthesis, primarily by influencing the availabil- ity of fatty acids for triacylglycerol biosynthesis. Furthermore, tetradecylthioacetic acid also affect cholesterol synthesis and secretion.
MATERIALS AND METHODS Chemicals and drugs. [l-'4C]Palmitic acid (56 Ci/mol),
acetyl-CoA (60 Ci/mol) and NaH['"]CO, (0.1 Ci/mol) were purchased from the Radiochemical Centre, Amersham. [4- ''C]Cholesterol and [3-'4C]hydroxymethylglutaryl CoA, with specific activities of 55-60 Ci/mol, were obtained from New England Corp. ,H,- labelled 7a-hydrocholesterol was synthesized as described earlier [13j. Arginine-free Dulbecco's modified Ea- gle's (DMEM) medium was obtained from Flow Lab. Collagen- ase (type 1) was delivered by Sigma Chem. Co. Tetradecylthio- acetic and tetradecylthiopropionic acids (Table 1) were prepared as described earlier [14, 151.
Fatty acid preparation. A sodium salt solution of oleic acid was prepared in distilled water, whereas tetradecylthioacetic and tetradecylthiopropionic acids were dissolved in 0.1 M NaOH. The sodium salts were then complexed with albumin.
Animals and treatments. Male Wistar rats from Mollegaard Breeding Laboratory, Ejby, Denmark, of approximately 150 g, were housed in metal cages and maintained under 12-h light/ dark cycles and at a constant temperature of 20 ? 3 "C. The ani-
mals were acclimatized for at least one week under these condi- tions before the start of the experiments. Tetradecylthioacetic and tetradecylthiopropionic acids were suspended in 0.5 % so- dium carboxymethylcellulose and administered by gastric intu- bation once a day for 5 days at a dose of 250mg . kg body mass-' . day-'. The animals had free access to water and food. There were no significant differences in food consumption or mass gain between the three experimental groups. At the end of the experiments (i.e. on the sixth day at 8.00 h), cardiac puncture was performed under neuroleptic anaesthesia and blood was col- lected in EDTA-containing tubes. Plasma was prepared by cen- trifugation at 3000 rpm for 20 min. For lipoprotein lipase and hepatic lipase assays, rats were injected intravenously with hepa- rin (1000 U k g body mass) 8 min prior to blood sampling.
Preparation of subcellular fractions. The livers were re- moved and immediately chilled on ice and the mass determined. Portions of the livers from the individual rats were homogenised in ice-cold sucrose medium (0.25 M sucrose in 10 mM Hepes, pH 7.4, and 1 mM EDTA) [8,9]. Mitochondrial and peroxisomal fractions were prepared as described earlier [8j. Microsomal fractions were prepared by centrifugating the homogenates at 20000Xg for 15 min and recentrifugation of the supernatant at lOOOOOXg for 60 min. The pellet was resuspended in the ice- cold sucrose medium. The resulting supernatant constituted the cytosolic fraction. Analysis of microsomal and cytoplasmic en- zyme activities is detailed below.
Enzyme assays. Phosphatidate phosphohydrolase activity in the microsomal fraction was determined by measuring the rate of release of free phosphate in the presence of 5 mM MgC1, [16]. ATP-citrate lyase [I71 was assayed in the cytosolic fraction and fatty acyl-CoA oxidase activity in the peroxisomal fraction was determined as described earlier [18, 191. Mitochondria1 p- oxidation was measured in a mitochondrial-enriched fraction using radiolabelled palmitoyl-L-carnitine or radiolabelled palmi- toyl-CoA as substrate [20]. Acetyl-CoA carboxylase activity was measured in the cytosolic fraction as the fixation of [14C]0, from NaH[14C]0, into malonyl-CoA, essentially as described by Tanabe et al. [21]. Preparation of microsomal fraction for assay of hydroxymethylglutaryl CoA (HMG-CoA) reductase and cho- lesterol 7a-hydroxylase has been described elsewhere [l l] . HMG-CoA reductase activity was assayed essentially as de- scribed by Brown et al. [22] and cholesterol 7a-hydroxylase ac- tivity was determined as described previously [ll, 131. Acyl- CoA :cholesterol acyltransferase (ACAT) activity was measured in the microsomal fraction essentially as described by Field et al. [23]. The assay was performed for 3 min at 37°C. Lipopro- tein and hepatic lipases activities were assayed as described [24].
Other analytical methods. Lipoprotein quantification was performed by a combination of ultracentrifugation and precipita- tion [25]. Briefly, plasma samples were centrifuged at a density 1.006 g/ml for 18 h at 35000 rpm in a Centrikon T-2060 ultra- centrifuge (Contron Roche) equipped with a 45.6 Ti rotor. The tubes were sliced, and the floating fractions (very-low-density lipoprotein; VLDL) as well as the low-density lipoprotein (LDL) and high-density lipoprotein (HDL) were analyzed for triacylglycerols and cholesterol. One portion of each infranatant was treated with phosphotungstic acid to precipitate apolipopro- tein-B-containing lipoproteins, and the resulting supernatant fraction was analyzed for triacylglycerols and cholesterol. By this procedure, it was possible to calculate the triacylglycerol and cholesterol contents in the VLDL, LDL and HDL fractions 1251.
Protein was assayed by the Bio-Rad protein assay kit (Bio- Rad). Lipid analyses were carried out by the Monotest choles- terol enzymic kit (Boehringer Mannheim), the Monospot phos-

Asiedu et al. ( E m J. Biochem. 227) 717
Table 2. Effect of tetradecylthioacetic and tetradecylthiopropionic acids on key enzymes of triacylglycerol biosynthesis and degradation. The reported values are means 5 SEM (n = 6). 1 mU enzyme activity is defined as the release of 1 nmol fatty acid from trioleidmin under the conditions of the assay. Means in each row without a common superscript are significantly different at P<O.O5.
Treatment Activity for
phosphatidate phosphohydrolase in lipoprotein hepatic
cytosolic fraction microsomal fraction lipase lipase
nmol . mg protein-' . min-' (nmol . g liver-' . min-') U/mI
Control 1.85 50.21" (111.40? 11.86") 23.27 50.95" (427.102 17.95") 3.94+0.14" 1.4610.03" Tetradecylthioacetic acid 0.5650.11b (52.61i10.30b) 16.65+0.26b (282.652 6.55b) 4.06?0.06" 1.6210.10" Tetradecylthiopropionic acid 1.27 20.09" (104.53 5 18.53") 21.95 i 0.86" (368.33 2 30.82") 4.35+0.19" 1.5410.02"
pholipid enzymic kit (Boehringer Mannheim) and the Biopak Triacylglycerol enzymic kit (Biotrol).
The VLDL triacylglycerol secretion rate was determined by measuring the increase in the plasma triacylglycerol concentra- tion after an intravenous injection of Triton WR 1339 as de- scribed before [26, 271.
Hepatic free CoASH and malonyl-CoA levels were deter- mined by an HPLC method as described by Corkey et al. [28].
Cell culture experiments. Hepatocytes were prepared from male albino rats (200-300 g) by collagenase perfusion and plated as described elsewhere [29]. After overnight incubation (18 h), the culture medium was removed and the cells were rinsed twice with serum-free Dulbecco's modified medium to wash off unattached and damaged cells. Some of the cells were then changed to a Dulbecco's modified medium containing Hepes (20mM), gentamycin (50mg/ml) and the various iso- topes and compounds indicated below. To hepatocytes cultured in plastic flasks (25 cm3) were added 2 ml incubation medium containing optically clear micellar solutions of [14C]palmitic acid (0.1 pCil20 pM), 200 pM oleic acid/80 pM bovine serum albu- min (2.5 : 1) in the absence (control) or presence of 200 pM tetra- decylthioacetic or 200 pM tetradecylthiopropionic acids. The hepatocytes were incubated for 4 h at 37°C. Synthesis and secre- tion of palmitic-acid-labelled lipids were linear over this time period.
Termination of incubation and harvesting of cells. After incubation, the culture dishes were cooled on ice, the medium was collected, centrifuged at 2000 rpm for 5 min and a cell-free aliquot was further treated. The cells were scraped off the culture dish by using a rubber policeman into 2 ml NaCVP, (0.9% NaCl and 50mM potassium phosphate, pH 7.4). The cells were washed once with NaClP,. The cell suspension was centrifuged at 2000 rpm for 5 min and the supernatant was discarded. The cells were resuspended in 500 pl distilled water and frozen at -20°C. Protein was assayed by the Bio-Rad protein assay kit in an aliquot of the suspended cells after sonication.
Measurement of cell-associated and secreted lipids. Lipids from the cells and media were extracted with chlorofordmetha- no1 (2:1, by vol.) DO]. The cell suspension was mixed with 20vol. chlorofodmethanol (2:1, by vol.), 4 vol. 0.9% NaCl (pH adjusted to 2 with HC1) were added and the mixture was allowed to separate into two phases. The organic phase was col- lected and dried under a stream of nitrogen at 40°C. To 1 ml of medium devoid of cellular debris was added 4 vol. chloroform/ methanol (2:1, by vol.) and 2% serum as unlabeled carrier for the lipids. The water phase of the medium extract was reex- tracted once with 4 vol. chlorofodmethanol (2: 1, by vol.), and the combined organic phases were dried under nitrogen.
The residual lipids extracted from the media or the cellular fraction were redissolved in 200 p1 hexane and aliquots were separated by TLC using hexaneldiethyletherlacetic acid (80:20:1, by vol.) as developing solvent. Other aliquots were separated using hexane/diethylether/acetic acid (65 : 35 :1, by vol.) followed by hexane as a second developing solvent, for analysis of diacylglycerol and unesterified cholesterol. The TLC foils were cut and added to 8-ml scintillation vials and radio- activity determined in a liquid scintillation spectrometer (TRI- CARB 1900TR, Packard Instrument). The biosynthesis of tri- acylglycerols is defined as the triacylglycerol in cells and me- dium, while triacylglycerol secretion is defined as the radio- activity present in the triacylglycerol fraction in the medium.
Fatty acid oxidation. Measurement of total palmitic acid oxidation to acid-soluble products was carried out as detailed. Aliquots of medium (250 pl) from the above experiments were precipitated with 1.0 ml, 0.75 M perchloric acid. Bovine serum albumin (0.45 %) was added to ensure complete precipitation of free fatty acids and lipids. The ice-cold acidic extract was centri- fuged (3000 rpm for 10 min) and 500 pl supernatant was used for determination of radioactivity by liquid scintillation [20].
The counting efficiency for 14C was 92%, calculated by means of external standards; quenching was similar in all ex- periments. With this design, one cannot distinguish between mitochondria1 and peroxisomal fatty acid oxidation. However, in cultured hepatocytes, peroxisomal P-oxidation accounts for 10-15% of total fatty acid oxidation.
Statistical analysis. Data are presented as means .f SEM for six animals in each experimental group. One-way analysis of variance was used to determine significant differences among the different groups. Fisher's protected least-square difference was used to determine differences between means at the 95% confidence interval.
RESULTS
Plasma lipids and lipoproteins. As shown in Table 1, admin- istration of tetradecylthioacetic acid to rats resulted in a reduc- tion of plasma triacylglycerols (44 %) and plasma cholesterol (26%). The decrease in plasma lipids was accompanied by a reduction of both cholesterol and triacylglycerols in the VLDL fraction (36% and 48%, respectively), as well as a decrease in LDL cholesterol and HDL triacylglycerol levels (36% and 50%, respectively). In contrast, HDL cholesterol levels were not af- fected and, thus, the HDL cholesterol/LDL cholesterol ratio increased by 40 %. Administration of tetradecylthiopropionic acid increased plasma triacylglycerol levels (Table l), in
718 Asiedu et al. (Eul: J . Biochem. 2277
Oxidation r--
40 "id 0
Synthesis
4 - th i0 f a t t y acid - 3-thta
f a t t y acid U
Secretion
4 - t h i o 3-thio fa t t y f a t t y a c i d ac id - -
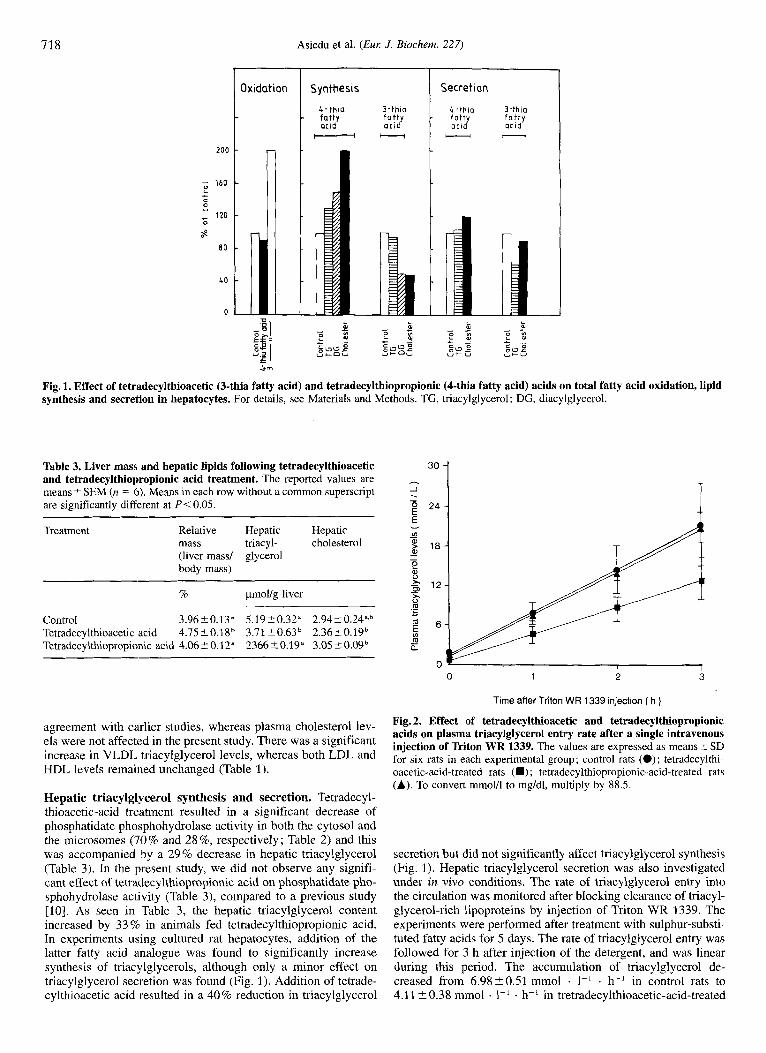
Fig. 1. Effect of tetradecylthioacetic (3-thia fatty acid) and tetradecylthiopropionic (4-thia fatty acid) acids on total fatty acid oxidation, lipid synthesis and secretion in hepatocytes. For details, see Materials and Methods. TG, triacylglycerol ; DG, diacylglycerol.
Table 3. Liver mass and hepatic lipids following tetradecylthioacetic and tetradecylthiopropionic acid treatment. The reported values are means L SEM (n = 6). Means in each row without a common superscript are significantly different at P< 0.05.
Treatment Relative Hepatic Hepatic mass triacyl- cholesterol (liver mass/ glycerol body mass)
% prnoVg liver
Control 3.96+0.13" 5.19?0.32" 2.94i-0.24".b Tetradecylthioacetic acid 4.75 irO.lgb 3.71 -+0.63b 2.36?0.19b Tetradecylthiopropionic acid 4.06 t- 0.12" 2366 2 0.19" 3.05 t O.Ogh
agreement with earlier studies, whereas plasma cholesterol lev- els were not affected in the present study. There was a significant increase in VLDL triacylglycerol levels, whereas both LDL and HDL levels remained unchanged (Table 1).
Hepatic triacylglycerol synthesis and secretion. Tetradecyl- thioacetic-acid treatment resulted in a significant decrease of phosphatidate phosphohydrolase activity in both the cytosol and the microsomes (70% and 2896, respectively; Table 2) and this was accompanied ,by a 29 % decrease in hepatic triacylglycerol (Table 3). In the present study, we did not observe any signifi- cant effect of tetradecylthiopropionic acid on phosphatidate pho- sphohydrolase activity (Table 3), compared to a previous study [lo]. As seen in Table 3, the hepatic tnacylglycerol content increased by 33 % in animals fed tetradecylthiopropionic acid. In experiments using cultured rat hepatocytes, addition of the latter fatty acid analogue was found to significantly increase synthesis of triacylglycerols, although only a minor effect on triacylglycerol secretion was found (Fig. 1). Addition of tetrade- cylthioacetic acid resulted in a 40% reduction in triacylglycerol
0 1 2 3
Time after Triton WR 1339 injection ( h )
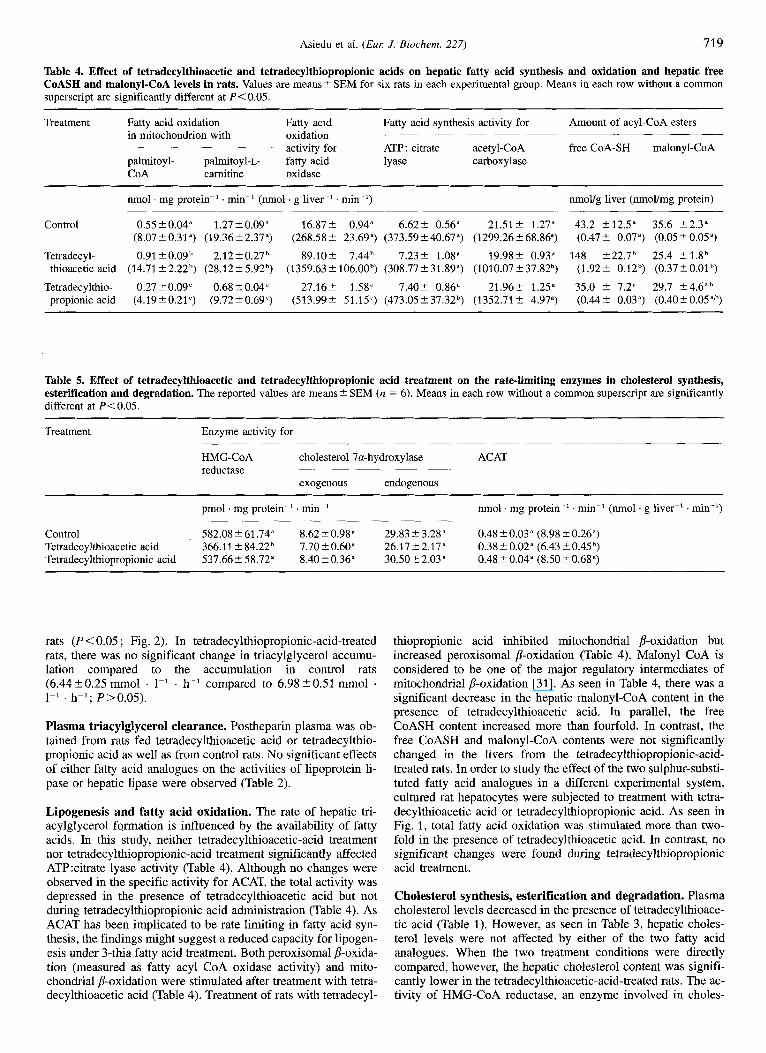
Fig. 2. Effect of tetradecylthioacetic and tetradecylthiopropionic acids on plasma triacylglycerol entry rate after a single intravenous injection of Triton WR 1339. The values are expressed as means L SD for six rats in each experimental group; control rats (0); tetradecylthi- oacetic-acid-treated rats (W); tetradecylthiopropionic-acid-treated rats (A). To convert mmolll to mg/dl, multiply by 88.5.
secretion but did not significantly affect triacylglycerol synthesis (Fig. 1). Hepatic triacylglycerol secretion was also investigated under in vivo conditions. The rate of triacylglycerol entry into the circulation was monitored after blocking clearance of triacyl- glycerol-rich lipoproteins by injection of Triton WR 1339. The experiments were performed after treatment with sulphur-substi- tuted fatty acids for 5 days. The rate of triacylglycerol entry was followed for 3 h after injection of the detergent, and was linear during this period. The accumulation of triacylglycerol de- creased from 6.98-+0.51 mmol . 1-' . h-' in control rats to 4.11 '. 0.38 mmol . 1-' . h-' in tretradecylthioacetic-acid-treated
Asiedu et al. ( E m J. Biochem. 227) 719
Table 4. Effect of tetradecylthioacetic and tetradecylthiopropionic acids on hepatic fatty acid synthesis and oxidation and hepatic free CoASH and malonyl-CoA levels in rats. Values are means 2 SEM for six rats in each experimental group. Means in each row without a common superscript are significantly different at P < 0.05.
Treatment Fatty acid oxidation Fatty acid Fatty acid synthesis activity for Amount of acyl-CoA esters in mitochondrion with oxidation
palmitoyl- palmitoyl-L- fatty acid lyase carboxylase CoA camitine oxidase
activity for ATP: citrate acetyl-CoA free CoA-SH malonyl-CoA
nmol . mg protein-' . min-' (nmol . g liver-' . min-') nmol/g liver (nmol/mg protein)
Control 0.5550.04" 1.2720.09" 16.875 0.94" 6.622 0.56" 21.51t 1.27" 43.2 212.5" 35.6 52.3"
Tetradecyl- 0.91 t0.09' 2.1250.27b 89.102 7.44b 7.23" 1.08" 19.982 0.93" 148 222.7b 25.4 51.8b
Tetradecylthio- 0.27 t 0.09" 0.68 2 0.04' 27.16t 1.58' 7.40? 0.86" 21.962 1.25" 35.0 5 7.2" 29.7 24.6".b
(8.07 20.31") (19.3622.37") (268.58 2 23.69") (373.59240.67") (1299.265 68.86") (0.472 0.07") (0.05 k0.05")
thioacetic acid (14.71 22.22b) (28.125 5.92b) (1359.63 k 106.00b) (308.77 5 31.89") (1010.07 t 37.82') (1.922 0.12') (0.37 20.01')
propionic acid (4.1950.21') (9.7250.69') (513.992 51.15") (473.05537.32') (1352.71 t 4.97") (0.442 0.03") (0.4050.05"~')
Table 5. Effect of tetradecylthioacetic and tetradecylthiopropionic acid treatment on the rate-limiting enzymes in cholesterol synthesis, esterification and degradation. The reported values are means 5 SEM (n = 6). Means in each row without a common superscript are significantly different at P<O.O5.
Treatment Enzyme activity for
HMG-COA cholesterol 7a-hydroxylase ACAT reductase
exogenous endogenous
pmol . mg protein-' . min-' nmol . mg protein-' . min-' (nmol . g liver-' . min-')
Control 582.08 t 61.74" 8.62 2 0.98" 29.83 2 3.28" 0.48 50.03" (8.98 20.26") Tetradecylthioacetic acid 366.11 284.22b 7.70t0.60" 26.17t2.17" 0.3820.02" (6.4320.45b) Tetradecylthiopropionic acid 537.66 5 58.72" 8.40 t 0.36" 30.50 t 2.03 a 0.48 20.04" (8.502 0.68")
rats (P < 0.05 ; Fig. 2). In tetradecylthiopropionic-acid-treated rats, there was no significant change in triacylglycerol accumu- lation compared to the accumulation in control rats (6.44 t 0.25 mmol . I-' . h-' compared to 6.98 _+ 0.51 mmol 1-' . h-'; P>0.05).
Plasma triacylglycerol clearance. Postheparin plasma was ob- tained from rats fed tetradecylthioacetic acid or tetradecylthio- propionic acid as well as from control rats. No significant effects of either fatty acid analogues on the activities of lipoprotein li- pase or hepatic lipase were observed (Table 2).
Lipogenesis and fatty acid oxidation. The rate of hepatic tri- acylglycerol formation is influenced by the availability of fatty acids. In this study, neither tetradecylthioacetic-acid treatment nor tetradecylthiopropionic-acid treatment significantly affected ATP:citrate lyase activity (Table 4). Although no changes were observed in the specific activity for ACAT, the total activity was depressed in the presence of tetradecylthioacetic acid but not during tetradecylthiopropionic acid administration (Table 4). As ACAT has been implicated to be rate limiting in fatty acid syn- thesis, the findings might suggest a reduced capacity for lipogen- esis under 3-thia fatty acid treatment. Both peroxisomal p-oxida- tion (measured as fatty acyl CoA oxidase activity) and mito- chondrial @-oxidation were stimulated after treatment with tetra- decylthioacetic acid (Table 4). Treatment of rats with tetradecyl-
thiopropionic acid inhibited mitochondria1 @-oxidation but increased peroxisomal @-oxidation (Table 4). Malonyl CoA is considered to be one of the major regulatory intermediates of mitochondria1 P-oxidation [31]. As seen in Table 4, there was a significant decrease in the hepatic malonyl-CoA content in the presence of tetradecylthioacetic acid. In parallel, the free CoASH content increased more than fourfold. In contrast, the free CoASH and malonyl-CoA contents were not significantly changed in the livers from the tetradecylthiopropionic-acid- treated rats. In order to study the effect of the two sulphur-substi- tuted fatty acid analogues in a different experimental system, cultured rat hepatocytes were subjected to treatment with tetra- decylthioacetic acid or tetradecylthiopropionic acid. As seen in Fig. 1, total fatty acid oxidation was stimulated more than two- fold in the presence of tetradecylthioacetic acid. In contrast, no significant changes were found during tetradecylthiopropionic acid treatment.
Cholesterol synthesis, esterification and degradation. Plasma cholesterol levels decreased in the presence of tetradecylthioace- tic acid (Table 1). However, as seen in Table 3, hepatic choles- terol levels were not affected by either of the two fatty acid analogues. When the two treatment conditions were directly compared, however, the hepatic cholesterol content was signifi- cantly lower in the tetradecylthioacetic-acid-treated rats. The ac- tivity of HMG-CoA reductase, an enzyme involved in choles-

720 Asiedu et al. ( E m J. Biochem. 227)
terol metabolism, decreased significantly in rats treated with tet- radecylthioacetic acid, but was not affected by tetradecylthio- propionic acid (Table 5). Neither ACAT nor cholesterol 7a-hy- droxylase activities were affected by administration of the fatty acid analogues.
DISCUSSION
A major new finding in the present work was the effect of tetradecylthioacetvc and tetradecylthiopropionic acids on lipo- protein fractions (Table 1). A decrease in both VLDL triacyl- glycerol and LDL cholesterol levels was noted after administra- tion of tetradecylthioacetic acid. The decrease in VLDL triacyl- glycerol was associated with a reduction in the secretion of newly synthesized triacylglycerol (Fig. 1). Since inhibition of fatty acid and triacylglycerol synthesis decrease VLDL triacyl- glycerol secretion [31], the reduced secretion of VLDL triacyl- glycerol (Fig. 1) after tetradecylthioacetic acid treatment is well in line with the decreased triacylglycerol synthesis under these conditions. It is interesting to note that the reduction in plasma triacylglycerol (44 %) was approximately equal to the reduction in VLDL triacylglycerol secretion (40 %). This suggests that a reduction in synthesis might be the dominating factor behind the lowering of plasma and lipoprotein triacylglycerol levels. In agreement with this, the activities of plasma lipoprotein lipase and hepatic lipase were unaffected (Table 2). This is in contrast to the findings on 3-thiadicarboxylic acid, another 3-thia fatty acid where the reduction in plasma triacylglycerol levels could not be explained by a decrease in VLDL triacylglycerol secre- tion, and treatment slightly increased the activities of lipoprotein lipase and hepatic Lipase [ 1 I]. Thus, the hypotriglyceridemia ob- served after tetradecylthioacetic acid treatment was solely due to effects on triacylglycerol production.
In the tetradecylthiopropionic acid treatment group, the level of triacylglycerol in plasma as well as VLDL triacylglycerol lev- els were higher than those in control groups (Table 1). However, the secretion of hepatic VLDL triacylglycerol (Fig. 2) and its subsequent clearance (Table 2) from the circulation was unaf- fected, compared to in control rats. It can, however, not be ex- cluded that under the present conditions, with a modest hy- pertriglyceridemia, the Triton method may not be sensitive enough to detect subtle differences in VLDL triacylglycerol secretion. The hypocholesterolemic effect of tetradecylthioacetic acid was accompanied by a decrease in VLDL and LDL choles- terol levels. As VL.DL is a precursor of LDL, it is conceivable that a reduction of VLDL secretion and of plasma VLDL levels will secondarily affect LDL formation, which might contribute to the lowering of LDL cholesterol levels. No significant changes in HDL cholesterol were found. As the HDL fraction is prominent in rats, it is particularly noteworthy that plasma total cholesterol levels decreased after tetradecylthioacetic acid ad- ministration, in spite of unchanged HDL cholesterol levels.
Another new finding in this study was that, in tetradecylthio- acetic-acid-treated rats, the activity of HMG-CoA reductase, the rate-limiting enzyme in cholesterol synthesis, was inhibited. Furthermore, in vitro experiments also showed that the admin- istration of tetradec ylthioacetic acid to hepatocytes resulted in a reduction of cholesterol ester synthesis (Fig. 1). These effects may contribute to lowering plasma cholesterol levels and sug- gest that different mechanisms may be in operation to lower rat plasma total cholesterol levels after tetradecylthioacetic acid treatment. Indeed, the availability of cholesterol in addition to triacylglycerols, has been shown to be of importance for lipopro- tein formation [32, 331. Thus, tetradecylthioacetic acid may af- fect lipoprotein metabolism by interfering with both triacylglyc-
'E ! 06 0'5/
Mitochondria1 p - owidation nmol . mg protein-1 . min-1
Fig. 3. Correlation between plasma triacylglycerol level and mito- chondrial /&oxidation after feeding rats with tetradecyltbioacetic and tetradecylthiopropionic acids (r = -0.87; P = 0.0025).
erol and cholesterol formation and availability. In contrast, ad- ministration of tetradecylthiopropionic acid had no effect on he- patic or plasma cholesterol levels. In agreement with this, the activities of HMG-CoA reductase, ACAT and cholesterol 7a- hydroxylase, as well as lipoprotein cholesterol levels, were unaf- fected in tetradecylthiopropionate-treated rats.
In previous studies, we have demonstrated that 3-thia fatty acids affect mitochondrial and peroxisomal P-oxidation. This was confirmed in the present study, as tetradecylthioacetic acid stimulated these pathways. The underlying mechanisms for these effects have not been clarified. It is conceivable that the increase in mitochondrial P-oxidation after tetradecylthioacetic acid treatment could be due to the low levels of malonyl-CoA (Table 41, as a decrease in the hepatic malonyl-CoA content in- creases carnitine palmitoyltransferase I activity [34], thereby increasing mitochondrial fatty acid oxidation. However, 3-thia- acyl-CoA can be formed in vivo (Berge, R. K., Skorve, J. and Vaagenes, H., unpublished results). Thus, an alternative mecha- nism may be that carnitine palmitoyltransferase I might be acti- vated and desensitized towards malonyl-CoA, possibly by com- petition of 3-thiaacyl-CoA for the malonyl-CoA-binding site of carnitine palmitoyltransferase I. Further studies are, however, needed to explore these mechanisms. The importance of modu- lation of P-oxidation for lipoprotein levels were underscored by the significant negative correlation found between plasma tri- acylglycerol levels and mitochondrial /?-oxidation ( r = -0.87, P = 0.0025; Fig. 3). Together, these results suggest that there is a causal relationship between the tetradecylthioacetic-acid-stim- ulated oxidation rate and the reduced hepatic triacylglycerol bio- synthesis and secretion. Thus, by stimulating fatty acid oxidation and decreasing lipogenesis, tetradecylthioacetic acid interferes with the availability of fatty acid for triacylglycerol synthesis.
In contrast to tetradecylthioacetic acid, repeated administra- tion of tetradecylthiopropionic acid to normolipidemic rats increased plasma triacylglycerol (46 %) and decreased mito- chondrial P-oxidation. In spite of the reduction of the latter, there was no change in hepatic malonyl-CoA levels. It is, therefore, apparent that the regulation of hepatic mitochondrial fatty acid oxidation after tetradecylthiopropionic acid administration may be independent of malonyl-CoA levels. It has recently been re- ported that tetradecylthiopropionic acid is metabolized to tetra- decylthioacrylic acid and that this metabolite inhibits mito- chondrial &oxidation [35]. Furthermore, key enzymes involved in fatty acid and triacylglycerol synthesis, namely, acetyl-CoA carboxylase and phosphatidate phosphohydrolase, respectively, were unaffected by tetradecylthiopropionic acid (Tables 2 and 4). In a cultured hepatocyte model system, tetradecylthiopropi- onic acid, however, increased triacylglycerol synthesis (Fig. 1).

Asiedu et al. ( E m J. Biochem. 227) 721
The data suggest that inhibition of mitochondrial /3-oxidation and normal lipogenesis may increase the availability of fatty acids for triacylglycerol synthesis, and this could account, in part, for the observed hypertriglyceridemia after administration of tetradecylthiopropionic acid to normolipidemic rats.
In summary, the results show that the following events occur after tetradecylthioacetic acid treatment : increased fatty acid ox- idation ; decreased triacylglycerol synthesis ; inhibition of cho- lesterol synthesis ; decreased VLDL and cholesterol ester secre- tion. In contrast, tetradecylthiopropionic acid inhibits mito- chondrial j?-oxidation and stimulates triacylglycerol formation but does not affect cholesterol metabolism.
The authors are grateful to Miss Randi Sandvik, Ms Anita Lofgren, Mr Svein Kriiger and Mr Terje BjGrndal for excellent technical assis- tance. The work was supported by grants from the Norwegian Council on Cardiovascular Disease, Bergen Health Foundation, The Swedish Medical Research Council (Projects 10349 and 3141) and Nordisk lnsu- linfond. Dr Berglund is a Florence Irving Associate Professor of Medi- cine and supported by grants from the Irving Endowment.
REFERENCES 1. Schulz, H. (1991) Oxidation of fatty acids, in Biochemistry of lipids,
lipoproteins and membranes (Vance, D. E. & Vance, J., eds) pp. 87-110, Elsevier Science Publishers B. V., Amsterdam.
2. Brindley, D. (1991) Metabolism of triacylglycerol, in Biochemistry oflipids (Vance, D. E. & Vance, J., eds) pp. 171 -203, Amsterdam Elsevier Science Publishers B. V.
3. Bortn, J., Wettesten, M., Rustaeus, S., Anderson, M. & Olofsson, S.-0. (1993) The assembly and secretion of apo B-100-containing lipoproteins, Biochem. Soc. Truns. 21, 487-493.
4. Gibbous, G. F. (1990) Assembly and secretion of hepatic very-low- density lipoprotein, Biochem. J. 286, 1 - 13.
5. Dixon, J. L., Furukawa, S . , Ginsberg, H. N. (1991) Oleate stimulates secretion of apolipoprotein B-containing lipoproteins from HepG2 cells by inhibiting early intracellular degradation of apoli- poprotein B, J. Bid. Chem. 266, 5080-5086.
6. Aarsland, A,, Aarsaether, N., Bremer, J. & Berge, R. K. (1989) Al- kylthioacetic acids (3-thia fatty acids) as non-p-oxidizable fatty acid analogues: a new group of hypolipidemic drugs. 111. Dissoci- ation of cholesterol- and triacylglycerol-lowering effects and the induction of peroxisomal p-oxidation, J. Lipid Res. 30, 1711- 1718.
7. Berge, R. K., Aarslaud, A,, Kryvi, H., Bremer, J. & Aarsaether, N. (1989) Alkylthioacetic acids (3-thia fatty acids) - a new group of non-l)-oxidizable peroxisome-inducing fatty acid analogues. I. Dose-response studies on hepatic peroxisomal and mitochondrial changes and long-chain fatty acid metabolizing enzymes in rats, Biochem. Pharmacol. 38, 3969-3979.
8. Berge, R. K., Aarsland, A,, Kryvi, H., Bremer, J. & Aarsrether, N. (1989) Alkylthioacetic acids (3-thia fatty acids) - a new group of non-p-oxidizable peroxisome-inducing fatty acid analogues. I. A study on the structural requirements for proliferation of peroxi- somes and mitochondria in rat liver, Biochim. Biophys. Acta 1004,
9. Asiedu, D. K., Skorve, J., Willumsen, N., Demoz, A. & Berge, R. K. (1993) Early effects on mitochondrial and peroxisomal p-oxi- dation by the hypolipidemic 3-thia fatty acids in rat livers, Bio- chim. Biophys. Actu 1166, 73-76.
10. Skorve, J., Asiedu, D., Rustan, A. C., Drevon, C. A., Al-Shurbaji, A. & Berge, R. K. (1990) Regulation of fatty acid oxidation and triacylglycerol and phospholipid metabolism by the hypolip- idemic sulfur-substituted fatty acid analogues, J. Lipid Res. 31, 1627- 1635.
11. Skorve, J., Al-Shurbaji, A., Asiedu, D., Bjorkhem, I., Berglund, L. & Berge, R. K. (1993) On the mechanism of the hypolipidemic ef- fect of sulfur-substituted hexadecaenoic acid (3-thiadicarboxylic acid), J. Lipid Res. 34, 1177-1185.
12. Al-Shurbaji, A,, Skorve, J., Berge, R. K., Rudling, M., Bjorkhem, I. & Berlgund, L. (1993) The effect of 3-thiadicarboxylic acid on
343-356.
lipid metabolism in experimental nephrosis, Arterioscler Tlzromb.
13. Bjorkhem, 1. & Kallner, A. (1976) Hepatic 7 a-hydroxylation of cholesterol in ascorbate-deficient and ascorbate-supplemented guinea pigs, J. Lipid Res. 17, 360-365.
14. Aarsland, A., Berge, R. K., Bremer, J. & Aarssether, N. (1990) The hypolipidemic peroxisome proliferating drug bis(carboxymethy1- thio) 1.10 decane, a dicarboxylic metabolite of tiadenol, is acti- vated to an acylcoenzyme A thioester, Biochim. Biophys. Actu 1033, 176-183.
15. Spydevold, 0. & Bremer, J. (1989) Induction of peroxisomal 8- oxidation in 7800 C1 Moms hepatoma cells in steady state by fatty acids and fatty acid analogues, Biochim. Biophys. Acta 1003,
16. Mavis, R. D., Finkelstein, J. N. & Hall, B. P. (1978) Pulmonary surfactant synthesis. A highly active microsomal phospatidate phosphohydrolase in the lungs, J. Lipid Res. 19, 467-477.
17. Rose-Khan, G. & Bar-Tana, I. (1989) Inhibition of lipid synthesis by j?’ . 8’-tetramethyl-substituted C,,-C,,, a, w-dicarboxylic acids in cultured rat hepatocytes, J. Biol. Chem. 260, 8411-8415.
18. Berge, R. K., Nilsson, A. & Husoy, A. M. (1988) Rapid stimulation of liver palmitoyl-CoA, synthetase, carnitine palmitoyltransferase and glycerophosphate acyltransferase compared to peroxisomal p- oxidation and palmitoyl-CoA hydrolase in rats fed high-fat diets, Biochim. Biophys. Acta 960, 417-426.
19. Berge, R. K., Flatmark, T. & Christiansen, E. N. (1987) Effect of a high-fat diet with partially hydrogenated fish oil on long-chain fatty acid metabolizing enzymes in subcellular fractions of rat liver, Arch. Biochem. Biophys. 252, 269-276.
20. Asiedu, D., Aarsland, A,, Skorve, J., Svardal, A. M. & Berge, R. K. (1990) Fatty acid metabolism in liver of rats treated with hypolip- idemic sulphur-substituted fatty acid analogues, Biochim. Bio- phys. Acta 1044, 211-221.
21. Tanabe, T. & Nakanishi, S . , Hashimoto, T., Nikowa, J.-I. & Numa, S . (1981) Acetyl-CoA carboxylase from rat liver, Methods Enzy-
22. Brown, M. S., Goldstein, J. L. & Diestschy, J. M. (1979) Active and inactive forms of 3-hydroxy-3-methylglutaryl Coenzyme A reductase in the liver of rat: comparison with the rate of choles- terol synthesis in different physiological states, J. Biol. Chem.
23. Field, F. J., Albright, E. & Methur, S . (1991) Inhibition of acyl coenzyme A: cholesterol acyltransferase activity by PD 128042. Effect on cholesterol metabolism and secretion in CaCo-2 cells, Lipids 26, 1-8.
24. Nilsson-Ehle, P. & Ekman, R. (1977) Rapid, simple and specific assays for lipoprotein lipase and hepatic lipase, Artery 3, 194- 209.
25. Carlson, K. (1973) Lipoprotein fractionation, J. Clin. Path. 26, 32- 37.
26. Otway, S . & Robinson, D. S . (1967) A non-ionic detergent (Triton WR 1339) to determine rates of triacylglycerol entry into the cir- culation of rats under different physiological conditions, J. Phy- siol. 190, 321 -323.
27. Al-Shurbaji, A,, Berglund, L. & Bjorkhem, I. (1990) The effect of Acipimox on triacylglycerol metabolism in rat, Scand. J. Clin. Invest. 50, 203 - 208.
28. Corkey, B. E., Brand, M., Williams, R. J. & Williams, J. R. (1981) Assay of short-chain acyl-CoA intermediates in tissue extracts in high pressure chromatography, Anal. Biochem. 118, 30-41.
29. Drevon, C. A,, Weinstein, D. B. & Steinberg, D. (1980) Regulation of cholesterol esterification and biosynthesis in monolayer cul- tures of normal adult rat hepatocytes, J. Biol. Chem. 255, 9128- 9137.
30. Folch, J., Lees, M. & Sloane-Stanley, G. H. (1957) A simple method for the isolation and purification of total lipids from animal tis- sues, J. Bid. Chem. 226, 497-509.
31. Arbeeny, C. M., Meyers, D. S . , Bergquist, K. E. & Gregg, R. E. (1992) Inhibition of fatty acid synthesis decreases very low den- sity lipoproptein secretion in the hamster, J. Lipid Res. 33, 843- 850.
32. Khan, B., Wilcox, H. G. & Heimberg, M. (1989) Cholesterol is required for secretion of very low density lipoproteins by rat liver, Biochem. J. 258, 807-816.
13, 1580-1586.
72-79.
m01. 71, 5-16.
254, 5144-5149.

722 Asiedu et al. (Eus J. Biochem. 227)
33. Cianflone, K. M., Yasruel, Z., Rodriguez, M. A., Vas, D. & Snider- man, A. D. (1990) Regulation of apo B secretion from Hep G2 cells : evidence for a critical role for cholesteryl ester synthesis in the response to fatty acid challenge, J . Lipid Res. 31,2045 -2055.
34. McGarry, J. D., Mannaerts, G. & Foster, D. W. (1977) A possible role for malonyl CoA in the regulation of hepatic fatty acid oxida- tion and ketogenesis, J, Clin. Invest. 60, 265-270.
35. Skrede, S. & Bremer, .I. (1993) Tetradecylthioacrylic acid, a beta- oxidation or metabolite of tetradecylthiopropionic acid, inhibits hepatic fatty acid activation and oxidation in rat, in Fatty acids and lipids from cell biology to human disease, p. 61, Lugano, Switzerland.
Related Documents











