Maturitas 60 (2008) 31–41 Available online at www.sciencedirect.com Ezrin is a key element in the human vagina A. Fadiel a , H.H. Lee a,1 , N. Demir b,c , S. Richman b , A. Iwasaki d , K. Connell a , F. Naftolin a,∗ a Department of Obstetrics and Gynecology, New York University, 550 First Avenue, TH528, New York, NY 10016, United States b Department of Obstetrics and Gynecology, Yale University School of Medicine, United States c Department of Histology and Embryology, Akdeniz University School of Medicine, 07070 Antalya, Turkey d Department of Immuno-biology, Yale University School of Medicine, United States Received 17 December 2007; received in revised form 14 March 2008; accepted 17 March 2008 Abstract Objective: The vagina is a complex tubular structure that has reproductive, support and barrier functions. These depend on the cytoarchitecture of the vaginal cells, which is controlled by key proteins. Cytoskeletal proteins determine cell polarity and membrane specializations by integrating the actin cytoskeleton with cell membranes. This integration is the domain of cytoskeletal proteins including the MERM protein family (moesin-ezrin-radixin-Merlin). Nothing is known about the cyto-localization of the MERM’s in the vaginal epithelium or how it influences the cytoarchitecture of the vaginal epithelium and stroma. Design: Full-thickness human vaginal fornix samples were obtained from 20 normal human specimens obtained at surgery for pelvic relaxation. Light- and electron microscopical immunohistochemistry (IHC) were used to identify and study activation and cellular localization of immuno-reactive-ezrin (ir-ezrin), a prototypical MERM. Results: Ir-ezrin was identified in the stratified squamous vaginal epithelium and connective tissue (fibroblasts, blood vessels and leucocytes). “H” scoring indicated that ir-ezrin staining is denser in the vaginal epithelium than in other layers, that the ir-ezrin staining was associated with increased keratinization and with the size of the tight junctions (p < 0.01). Both the amounts and localization of ir-ezrin were associated with high levels of estrogen, identified by the menstrual history and keratinization of the superficial vaginal epithelium. The density of stromal ir-ezrin was increased in the presence of dense epithelial keratinization. Immuno-reactive-ezrin staining was most pronounced near the cell membranes of both keratinized and non-keratinized epithe- lium, indicating that ezrin activation (unfolding and movement to the membrane) had occurred. Ultra-structural examination of the epithelium showed intra-cellular ir-ezrin to be localized to junctional complexes that have been associated with decreased mucosal penetration by microorganisms. Ir-ezrin was widely distributed throughout stromal fibro-muscular cell, vessels and immunocytes. Conclusions: MERM’s, represented by ezrin, are widely present in the vaginal wall. This has implications for the strength and resilience of this tubular structure and may be the case in other internal genital tissues. Ezrin’s localization and association with cell specializations indicate that in the vagina, as in other tissues, ezrin likely modulates vaginal cell–cell interactions including ∗ Corresponding author. E-mail addresses: [email protected], [email protected] (F. Naftolin). 1 Present address: Department of Obstetrics and Gynecology, School of Medicine, Soonchunhyang University, Bucheon 420-767, Republic of Korea. 0378-5122/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.maturitas.2008.03.007

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

A
OtmptDpaRaiatIltmiCrc
o
0
Maturitas 60 (2008) 31–41
Available online at www.sciencedirect.com
Ezrin is a key element in the human vagina
A. Fadiel a, H.H. Lee a,1, N. Demir b,c, S. Richman b,A. Iwasaki d, K. Connell a, F. Naftolin a,∗
a Department of Obstetrics and Gynecology, New York University, 550 First Avenue, TH528, New York, NY 10016, United Statesb Department of Obstetrics and Gynecology, Yale University School of Medicine, United States
c Department of Histology and Embryology, Akdeniz University School of Medicine, 07070 Antalya, Turkeyd Department of Immuno-biology, Yale University School of Medicine, United States
Received 17 December 2007; received in revised form 14 March 2008; accepted 17 March 2008
bstract
bjective: The vagina is a complex tubular structure that has reproductive, support and barrier functions. These depend onhe cytoarchitecture of the vaginal cells, which is controlled by key proteins. Cytoskeletal proteins determine cell polarity and
embrane specializations by integrating the actin cytoskeleton with cell membranes. This integration is the domain of cytoskeletalroteins including the MERM protein family (moesin-ezrin-radixin-Merlin). Nothing is known about the cyto-localization ofhe MERM’s in the vaginal epithelium or how it influences the cytoarchitecture of the vaginal epithelium and stroma.esign: Full-thickness human vaginal fornix samples were obtained from 20 normal human specimens obtained at surgery forelvic relaxation. Light- and electron microscopical immunohistochemistry (IHC) were used to identify and study activationnd cellular localization of immuno-reactive-ezrin (ir-ezrin), a prototypical MERM.esults: Ir-ezrin was identified in the stratified squamous vaginal epithelium and connective tissue (fibroblasts, blood vesselsnd leucocytes). “H” scoring indicated that ir-ezrin staining is denser in the vaginal epithelium than in other layers, that ther-ezrin staining was associated with increased keratinization and with the size of the tight junctions (p < 0.01). Both the amountsnd localization of ir-ezrin were associated with high levels of estrogen, identified by the menstrual history and keratinization ofhe superficial vaginal epithelium. The density of stromal ir-ezrin was increased in the presence of dense epithelial keratinization.mmuno-reactive-ezrin staining was most pronounced near the cell membranes of both keratinized and non-keratinized epithe-
ium, indicating that ezrin activation (unfolding and movement to the membrane) had occurred. Ultra-structural examination ofhe epithelium showed intra-cellular ir-ezrin to be localized to junctional complexes that have been associated with decreaseducosal penetration by microorganisms. Ir-ezrin was widely distributed throughout stromal fibro-muscular cell, vessels andmmunocytes.onclusions: MERM’s, represented by ezrin, are widely present in the vaginal wall. This has implications for the strength and
esilience of this tubular structure and may be the case in other internal genital tissues. Ezrin’s localization and association withell specializations indicate that in the vagina, as in other tissues, ezrin likely modulates vaginal cell–cell interactions including
∗ Corresponding author.E-mail addresses: [email protected], [email protected] (F. Naftolin).
1 Present address: Department of Obstetrics and Gynecology, School of Medicine, Soonchunhyang University, Bucheon 420-767, Republicf Korea.
378-5122/$ – see front matter © 2008 Elsevier Ireland Ltd. All rights reserved.doi:10.1016/j.maturitas.2008.03.007

3
trhaw©
K
1
ttfidbebvadiafcpof
oMtTtftfnrlsrso
e
2 A. Fadiel et al. / Maturitas 60 (2008) 31–41
he changing vaginal cellular interface with the external environment, the regulation of the elasticity of the vagina, and theegulation of microbial and chemical traffic that determine the pH and microbial environment of the vagina. In other work weave shown that ezrin expression is induced by estradiol. The increase of ir-ezrin staining during the appearance of keratinizationnd maturation of the vaginal cytology indicates that estrogen may regulate vaginal ezrin and thereby the properties of the vaginalall and epithelium.2008 Elsevier Ireland Ltd. All rights reserved.
arian c
opcbroroppcwGakciliapitctgrida
2
2
eywords: Ezrin; MERM; Pelvic floor; Vaginal cycle; Estrogen; Ov
. Introduction
Through a series of branches in mammalian evolu-ion the vagina has emerged as the interface betweenhe external environment and the interior milieu, a keyemale reproductive organ and an integrating nexusn the support of the pelvic floor [1]. While it is wellescribed as an anatomic structure, the molecularases of the function and changes of this indispensable,ndocrine-dependent and changing organ remain toe discovered. However, it is clear that the humanagina is a remarkably plastic organ that uses mech-nisms inherited from its predecessor excretory tracterivatives to maintain homeostatic function, mucosalmmunity, lack of excessive scarring/keloid formationnd support of the pelvic floor in an upright, internallyertilizing biped [2]. These functions require actinytoskeleton-linking proteins that regulate cell shape,olarization, membrane-specialization, variable ratesf proliferation and shedding and development ofunctional intermediate fibers such as keratin.
The MERM (moesin-ezrin-radixin-Merlin) familyf proteins is frequently involved in these actions.ERM’s are a highly conserved family of proteins
hat link the actin cytoskeleton to the cell membrane.he actions of the MERM’s are regulated by signal
ransduction pathways that are triggered by ligandsor membrane receptors [3]. Recently, we showedhat ezrin and moesin expression, activation andunction are regulated by estrogen, via both classicaluclear receptor mechanisms and membrane-boundeceptor-like molecules [3,4]. Because of the simi-arity between the actions of these membrane-actincaffolding proteins elsewhere in the body, and theepertoire displayed by the cells of the vagina, we
ought and confirmed the presence of ezrin in the cellsf the vaginal epithelium and stroma.Ezrin is one of the proteins that correlate withpithelial morphological changes. Ezrin is a member
fc
ycle; HRT
f the MERM family that function as scaffoldingroteins linking the cell membrane with the actinytoskeleton. This requires two phosphorylations thatond the C terminal to transmembrane proteins such aseceptors and the N terminal to other MERM proteinsr actin fibrils [5,6]. In this manner, ezrin plays a keyole in cellular processes such as motility, corticalrganization and cell proliferation [5,6] and cellolarity [7]. When ezrin is in its inactive or dormant,eri-nuclear form, its N and C terminals are held inlose conformation that stops ezrin from interactingith other proteins. Upon activation via the RhoTPase pathway, ezrin unfolds and the two sites are
vailable to phosphorylation [8,9]. These proteins arenown to be responsible for lateral specialization ofells, thereby playing a key role in modulating cell–cellnteraction and dynamics [10,11]. The intracellularocation of ezrin [12–14,9] allows inference of its rolen the cell: when first expressed, ezrin is peri-nuclearnd requires a signal, i.e. from the Rho GTPaseathway to move toward the cell membrane wheret organizes the relationship of the actin cytoskeletono the cell membrane, so that lateral specializationsan direct the cell’s activities. We have shown thathis process is directly influenced by E2, through bothenomic and non-genomic pathways [3,4]. In thiseport we document the presence and action of ezrinn the vaginal wall, including the ovarian cycle-relatedevelopment of intercellular membrane specializationss keratinization becomes the dominant feature.
. Materials and methods
.1. Tissue collection and preparation
Full thickness vaginal wall samples were obtainedrom 20 patients undergoing vaginal hysterectomy andolporraphy for pelvic relaxation. The protocol and

aturitas
cCwitc[
2e
toMwapnwDPctmaoALpVdVtcuifcmmvA
2
t
0mieot1adadtpcWCp
2
cebpaat2ws0vaJ1o3p3sngr
A. Fadiel et al. / M
onsent were approved by the Human Investigationommittee of Yale University. The mean patient ageas 35 ± 15 years (S.D.). Tissues were promptly fixed
n 4% paraformaldehyde, maintained in 70% alcohol,hen either embedded in paraffin for immunohisto-hemistry (IHC) or fixed in Somogyi–Takagi solution15] for electron microscopy.
.2. Immunohistochemistry and light microscopicxamination
Fixed tissue samples were processed for hema-oxylin and eosin staining or for IHC, and collectedn poly-l-lysine-coated slides (Sigma–Aldrich Inc.,O). Tissue antigens were unmasked by micro-aving at 750 W for 5 min. After cooling for 10 min
t room temperature, the sections were washed inhosphate-buffered saline (PBS; pH 7.4). Endoge-ous peroxidases were blocked by treating the sectionsith 3% hydrogen peroxide (Dako A/S, Glostrup,enmark) for 30 min then washing three times withBS. Slides were then incubated in a humidifiedhamber with blocking serum for 10 min at roomemperature (Lab Vision, CA). An ezrin monoclonal
ouse anti-human IgG1 (Sigma–Aldrich, CA) waspplied. Sections were incubated with the primary mAbvernight at 4 ◦C, and rinsed with PBS 3× 5 min.
biotinylated horse anti-mouse antibody (Vectoraboratories, CA) was applied for 1 h at room tem-erature. An avidin–biotin–peroxidase system (Labision) was used for antigen detection. DAB (3,3-iaminobenzidine tetrahydrochloride dehydrate; Labision) was used as a chromagen. Mayer’s hema-
oxylin (Merck, Darmstadt, Germany) was used forounterstaining. After staining, slides were mountedsing Permount (Fisher Chemicals, NJ). During themmunostaining process, a negative control was per-ormed using the above protocol (same conditions,oncentrations and processing) but with non-immuneouse IgG at the same concentration as the immuneAb. An AxioPlan fluorescent microscope was used to
isualize the slides. Images were captured using ZiessxioVision software (Ziess, Germany).
.3. H-scoring and statistical analysis
The intensity and distribution of ir-ezrin in vaginalissues was evaluated semi-quantitatively as follows:
fw1p
60 (2008) 31–41 33
, no staining; 1+, weak but detectable staining; 2+,oderate or distinct staining; and 3+, strong stain-
ng. An H-SCORE was calculated using the followingquation: H = �Pi (i + 1), where i is the intensityf staining with a value from 0 to 3 and Pi ishe percentage of stained cells, varying from 0 to00%. In each slide, five different random areasnd 100 cells in each area were evaluated by twoifferent blinded observers using an 40× objective,nd the average score was calculated. Since theata were normally distributed (as determined withhe Kolmogorov–Smirnov test), comparisons of sam-les were analyzed by the Student’s t-test. Statisticalalculations were performed using Sigma Stat forindows, version 3.0 (Jandel Scientific, San Rafael,A, USA). Statistical significance was defined as< 0.05.
.4. Electron microscopy
In order to demonstrate and determine the sub-ellular localization of ir-ezrin in human vaginalpithelium, tissue samples were immediately fixedy immersion in Somogyi–Takagi solution (4%araformaldehyde, 0.1% glutaraldehyde, 15% picriccid in 0.1 M PB (phosphate buffer, pH 7.3) [15]t room temperature for 2 h. Samples were kept inhe same fixative solution without glutaraldehyde forh at 4 ◦C. Following fixation; samples were washedith 0.1 M PB to remove debris of picric acid. The
amples were immersed in 20% sucrose solution (in.1 M PB), until they descended to the bottom of theial, then embedded in cryo-matrix (Tissue-Tek, USA)nd cryosectioned (25 �m; 2800 Frigocut, Reichert-ung). The tissue sections were washed three times,0 min each with 0.1 M PB and the endogenous per-xidase activity were blocked by 0.3% H2O2 for0 min, then treated with 1% NaBHO solution pre-ared in 0.1 M PB for 15 min, then washed with PB× 10 min. Following freezing and thawing in 10%ucrose solution, and washing with PB 5× 10 min,on-specific binding was blocked with 2% normaloat serum (NGS). The tissues were incubated withabbit anti-human ezrin antibody (1:10,000 dilutions)
or 48 h at 4 ◦C. After washing 5× 10 min, sectionsere incubated with biotinylated goat anti-rabbit (IgG,:250) for 2 h and treated with an avidin–biotin com-lex (ABC) (1:100) (Vector Elite kit) for 2 h. After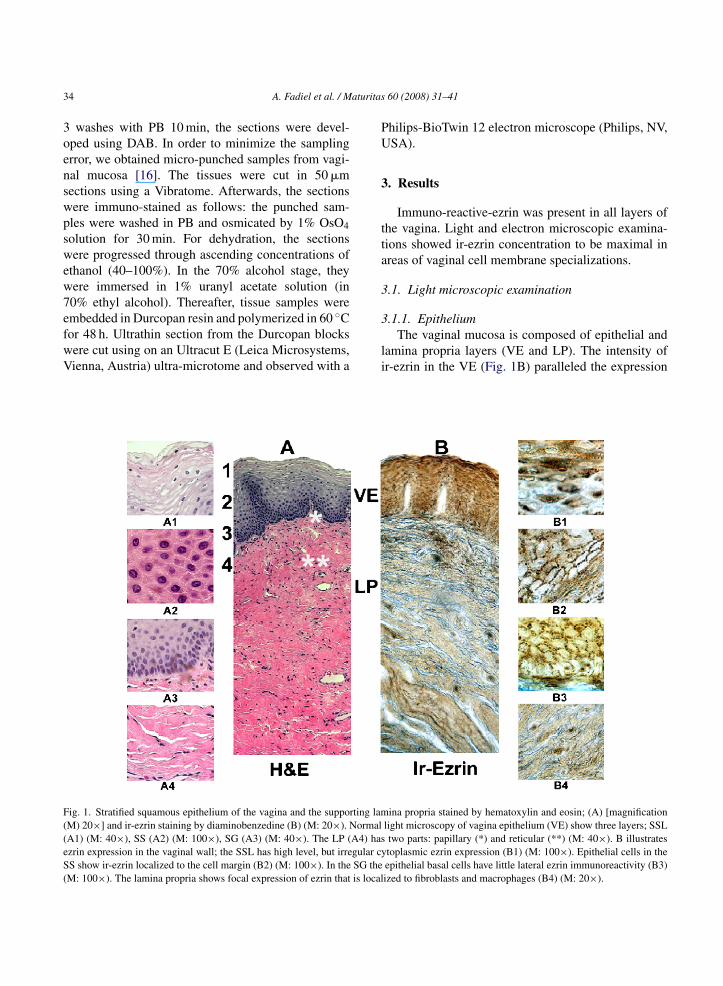
3 aturitas
3oenswpswew7efwV
PU
3
tta
3
F((eS(
4 A. Fadiel et al. / M
washes with PB 10 min, the sections were devel-ped using DAB. In order to minimize the samplingrror, we obtained micro-punched samples from vagi-al mucosa [16]. The tissues were cut in 50 �mections using a Vibratome. Afterwards, the sectionsere immuno-stained as follows: the punched sam-les were washed in PB and osmicated by 1% OsO4olution for 30 min. For dehydration, the sectionsere progressed through ascending concentrations of
thanol (40–100%). In the 70% alcohol stage, theyere immersed in 1% uranyl acetate solution (in0% ethyl alcohol). Thereafter, tissue samples were
mbedded in Durcopan resin and polymerized in 60 ◦Cor 48 h. Ultrathin section from the Durcopan blocksere cut using on an Ultracut E (Leica Microsystems,ienna, Austria) ultra-microtome and observed with a3
li
ig. 1. Stratified squamous epithelium of the vagina and the supporting lamM) 20×] and ir-ezrin staining by diaminobenzedine (B) (M: 20×). NormalA1) (M: 40×), SS (A2) (M: 100×), SG (A3) (M: 40×). The LP (A4) haszrin expression in the vaginal wall; the SSL has high level, but irregular cyS show ir-ezrin localized to the cell margin (B2) (M: 100×). In the SG theM: 100×). The lamina propria shows focal expression of ezrin that is local
60 (2008) 31–41
hilips-BioTwin 12 electron microscope (Philips, NV,SA).
. Results
Immuno-reactive-ezrin was present in all layers ofhe vagina. Light and electron microscopic examina-ions showed ir-ezrin concentration to be maximal inreas of vaginal cell membrane specializations.
.1. Light microscopic examination
.1.1. EpitheliumThe vaginal mucosa is composed of epithelial and
amina propria layers (VE and LP). The intensity ofr-ezrin in the VE (Fig. 1B) paralleled the expression
ina propria stained by hematoxylin and eosin; (A) [magnificationlight microscopy of vagina epithelium (VE) show three layers; SSLtwo parts: papillary (*) and reticular (**) (M: 40×). B illustratestoplasmic ezrin expression (B1) (M: 100×). Epithelial cells in theepithelial basal cells have little lateral ezrin immunoreactivity (B3)ized to fibroblasts and macrophages (B4) (M: 20×).
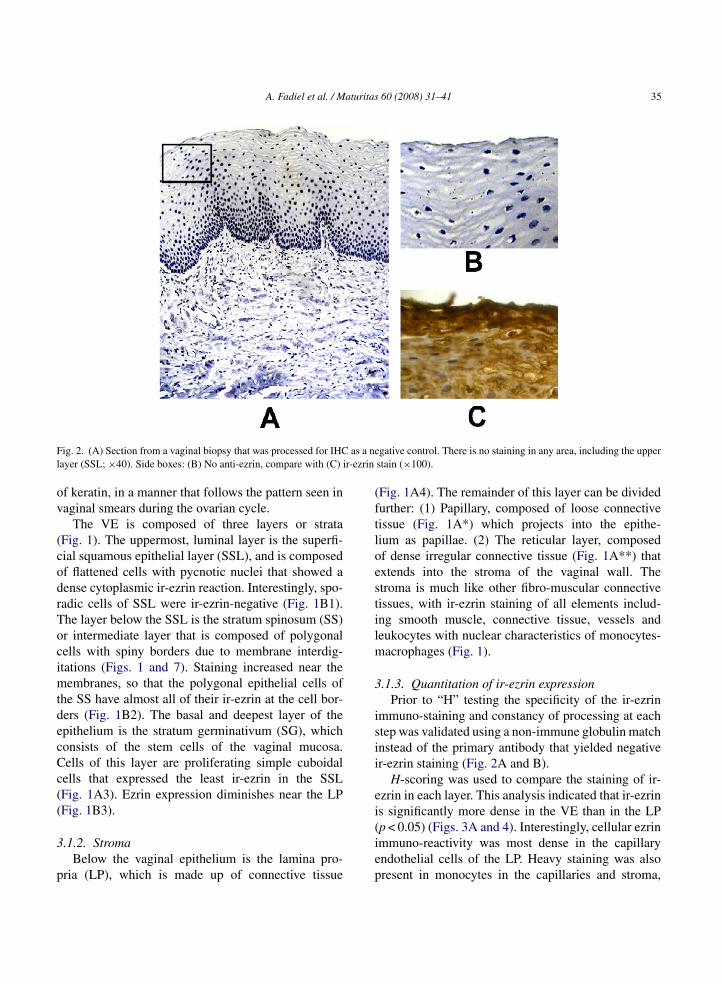
A. Fadiel et al. / Maturitas 60 (2008) 31–41 35
F as a nel r-ezrin
ov
(codrTocimtdecCc((
3
p
(ftloestilm
3
isii
ei
ig. 2. (A) Section from a vaginal biopsy that was processed for IHCayer (SSL; ×40). Side boxes: (B) No anti-ezrin, compare with (C) i
f keratin, in a manner that follows the pattern seen inaginal smears during the ovarian cycle.
The VE is composed of three layers or strataFig. 1). The uppermost, luminal layer is the superfi-ial squamous epithelial layer (SSL), and is composedf flattened cells with pycnotic nuclei that showed aense cytoplasmic ir-ezrin reaction. Interestingly, spo-adic cells of SSL were ir-ezrin-negative (Fig. 1B1).he layer below the SSL is the stratum spinosum (SS)r intermediate layer that is composed of polygonalells with spiny borders due to membrane interdig-tations (Figs. 1 and 7). Staining increased near the
embranes, so that the polygonal epithelial cells ofhe SS have almost all of their ir-ezrin at the cell bor-ers (Fig. 1B2). The basal and deepest layer of thepithelium is the stratum germinativum (SG), whichonsists of the stem cells of the vaginal mucosa.ells of this layer are proliferating simple cuboidalells that expressed the least ir-ezrin in the SSLFig. 1A3). Ezrin expression diminishes near the LPFig. 1B3).
.1.2. StromaBelow the vaginal epithelium is the lamina pro-
ria (LP), which is made up of connective tissue
(iep
gative control. There is no staining in any area, including the upperstain (×100).
Fig. 1A4). The remainder of this layer can be dividedurther: (1) Papillary, composed of loose connectiveissue (Fig. 1A*) which projects into the epithe-ium as papillae. (2) The reticular layer, composedf dense irregular connective tissue (Fig. 1A**) thatxtends into the stroma of the vaginal wall. Thetroma is much like other fibro-muscular connectiveissues, with ir-ezrin staining of all elements includ-ng smooth muscle, connective tissue, vessels andeukocytes with nuclear characteristics of monocytes-
acrophages (Fig. 1).
.1.3. Quantitation of ir-ezrin expressionPrior to “H” testing the specificity of the ir-ezrin
mmuno-staining and constancy of processing at eachtep was validated using a non-immune globulin matchnstead of the primary antibody that yielded negativer-ezrin staining (Fig. 2A and B).
H-scoring was used to compare the staining of ir-zrin in each layer. This analysis indicated that ir-ezrins significantly more dense in the VE than in the LP
p < 0.05) (Figs. 3A and 4). Interestingly, cellular ezrinmmuno-reactivity was most dense in the capillaryndothelial cells of the LP. Heavy staining was alsoresent in monocytes in the capillaries and stroma,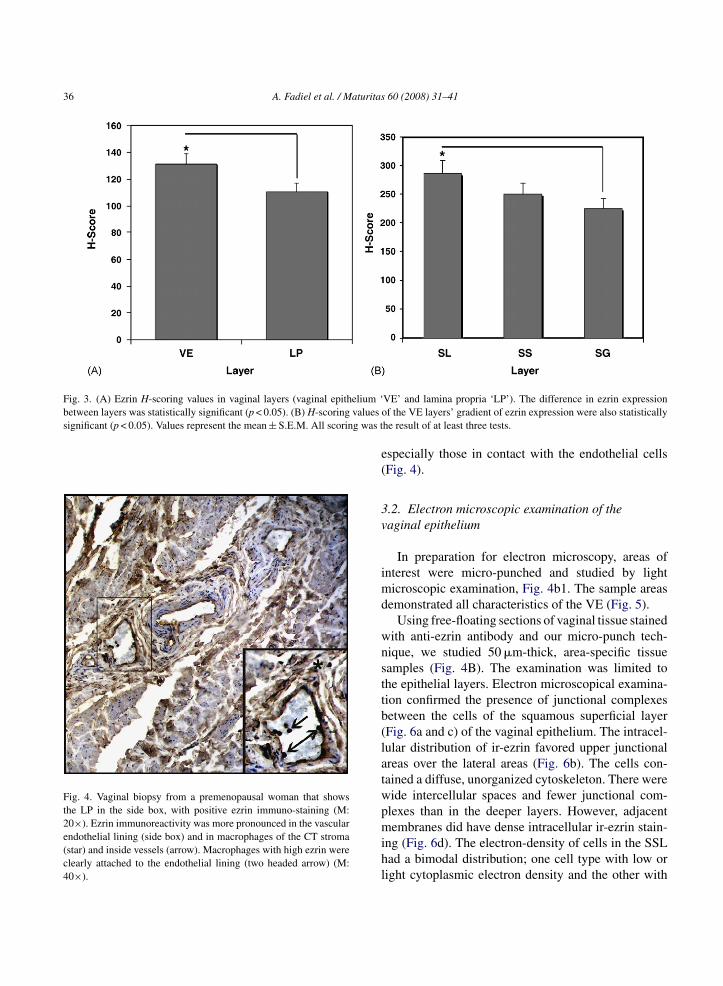
36 A. Fadiel et al. / Maturitas 60 (2008) 31–41
Fig. 3. (A) Ezrin H-scoring values in vaginal layers (vaginal epithelium ‘between layers was statistically significant (p < 0.05). (B) H-scoring values osignificant (p < 0.05). Values represent the mean ± S.E.M. All scoring was t
Fig. 4. Vaginal biopsy from a premenopausal woman that showsthe LP in the side box, with positive ezrin immuno-staining (M:20×). Ezrin immunoreactivity was more pronounced in the vascularendothelial lining (side box) and in macrophages of the CT stroma(star) and inside vessels (arrow). Macrophages with high ezrin wereclearly attached to the endothelial lining (two headed arrow) (M:40×).
e(
3v
imd
wnsttb(latwpmihl
VE’ and lamina propria ‘LP’). The difference in ezrin expressionf the VE layers’ gradient of ezrin expression were also statistically
he result of at least three tests.
specially those in contact with the endothelial cellsFig. 4).
.2. Electron microscopic examination of theaginal epithelium
In preparation for electron microscopy, areas ofnterest were micro-punched and studied by light
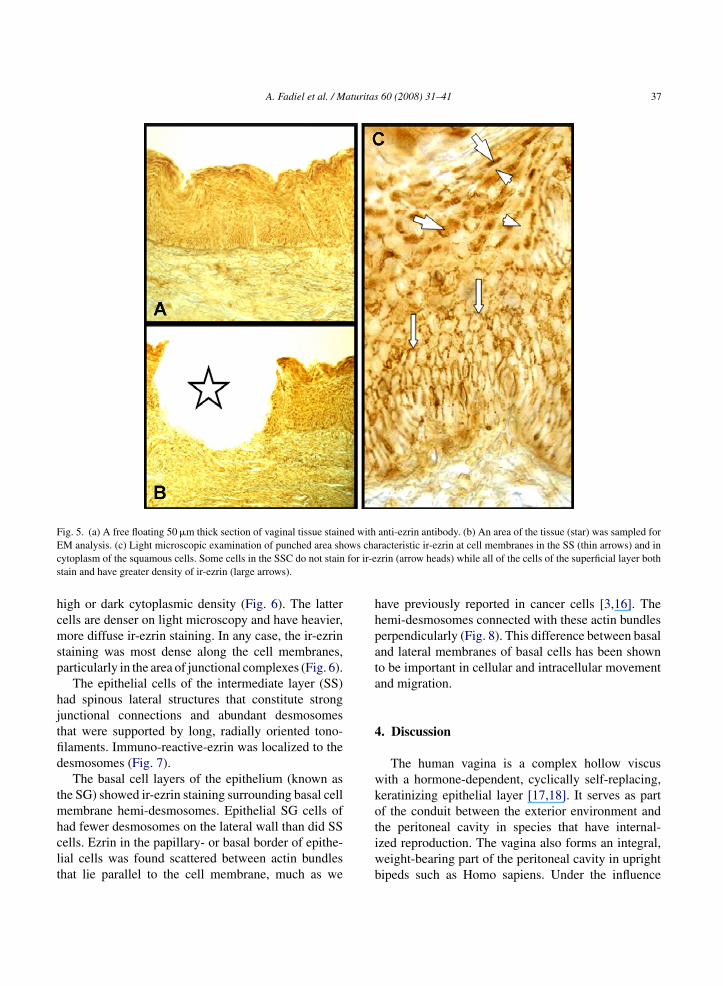
icroscopic examination, Fig. 4b1. The sample areasemonstrated all characteristics of the VE (Fig. 5).
Using free-floating sections of vaginal tissue stainedith anti-ezrin antibody and our micro-punch tech-ique, we studied 50 �m-thick, area-specific tissueamples (Fig. 4B). The examination was limited tohe epithelial layers. Electron microscopical examina-ion confirmed the presence of junctional complexesetween the cells of the squamous superficial layerFig. 6a and c) of the vaginal epithelium. The intracel-ular distribution of ir-ezrin favored upper junctionalreas over the lateral areas (Fig. 6b). The cells con-ained a diffuse, unorganized cytoskeleton. There wereide intercellular spaces and fewer junctional com-lexes than in the deeper layers. However, adjacent
embranes did have dense intracellular ir-ezrin stain-ng (Fig. 6d). The electron-density of cells in the SSLad a bimodal distribution; one cell type with low oright cytoplasmic electron density and the other with
A. Fadiel et al. / Maturitas 60 (2008) 31–41 37
Fig. 5. (a) A free floating 50 �m thick section of vaginal tissue stained with anti-ezrin antibody. (b) An area of the tissue (star) was sampled forE ows chac for ir-es
hcmsp
hjtfid
tmhclt
hhpata
4
wko
M analysis. (c) Light microscopic examination of punched area shytoplasm of the squamous cells. Some cells in the SSC do not staintain and have greater density of ir-ezrin (large arrows).
igh or dark cytoplasmic density (Fig. 6). The latterells are denser on light microscopy and have heavier,ore diffuse ir-ezrin staining. In any case, the ir-ezrin
taining was most dense along the cell membranes,articularly in the area of junctional complexes (Fig. 6).
The epithelial cells of the intermediate layer (SS)ad spinous lateral structures that constitute strongunctional connections and abundant desmosomeshat were supported by long, radially oriented tono-laments. Immuno-reactive-ezrin was localized to theesmosomes (Fig. 7).
The basal cell layers of the epithelium (known ashe SG) showed ir-ezrin staining surrounding basal cell
embrane hemi-desmosomes. Epithelial SG cells of
ad fewer desmosomes on the lateral wall than did SSells. Ezrin in the papillary- or basal border of epithe-ial cells was found scattered between actin bundleshat lie parallel to the cell membrane, much as wetiwb
racteristic ir-ezrin at cell membranes in the SS (thin arrows) and inzrin (arrow heads) while all of the cells of the superficial layer both
ave previously reported in cancer cells [3,16]. Theemi-desmosomes connected with these actin bundleserpendicularly (Fig. 8). This difference between basalnd lateral membranes of basal cells has been showno be important in cellular and intracellular movementnd migration.
. Discussion
The human vagina is a complex hollow viscusith a hormone-dependent, cyclically self-replacing,eratinizing epithelial layer [17,18]. It serves as partf the conduit between the exterior environment and
he peritoneal cavity in species that have internal-zed reproduction. The vagina also forms an integral,eight-bearing part of the peritoneal cavity in uprightipeds such as Homo sapiens. Under the influence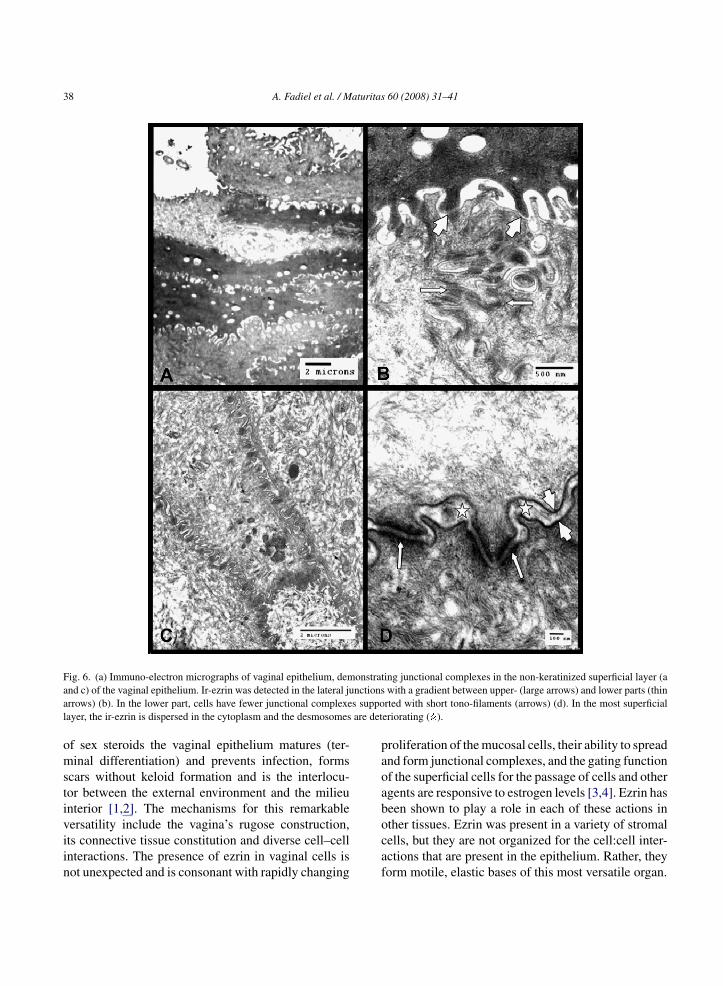
38 A. Fadiel et al. / Maturitas 60 (2008) 31–41
F onstrata nctionsa s suppol are det
omstiviin
paoab
ig. 6. (a) Immuno-electron micrographs of vaginal epithelium, demnd c) of the vaginal epithelium. Ir-ezrin was detected in the lateral jurrows) (b). In the lower part, cells have fewer junctional complexeayer, the ir-ezrin is dispersed in the cytoplasm and the desmosomes
f sex steroids the vaginal epithelium matures (ter-inal differentiation) and prevents infection, forms
cars without keloid formation and is the interlocu-or between the external environment and the milieunterior [1,2]. The mechanisms for this remarkable
ersatility include the vagina’s rugose construction,ts connective tissue constitution and diverse cell–cellnteractions. The presence of ezrin in vaginal cells isot unexpected and is consonant with rapidly changingocaf
ing junctional complexes in the non-keratinized superficial layer (awith a gradient between upper- (large arrows) and lower parts (thinrted with short tono-filaments (arrows) (d). In the most superficialeriorating ( ).
roliferation of the mucosal cells, their ability to spreadnd form junctional complexes, and the gating functionf the superficial cells for the passage of cells and othergents are responsive to estrogen levels [3,4]. Ezrin haseen shown to play a role in each of these actions in
ther tissues. Ezrin was present in a variety of stromalells, but they are not organized for the cell:cell inter-ctions that are present in the epithelium. Rather, theyorm motile, elastic bases of this most versatile organ.
A. Fadiel et al. / Maturitas 60 (2008) 31–41 39
Fig. 7. (a) Immuno-electron micrographs of vaginal epithelium demonstrating that cells of the SS with strong spinous junctional connectionsh us juncs (arrowj evelopeT fibrils.
bciait
pta
ave ezrin expression (arrows). (b) High magnification of the spinoupportive, long tono-filaments (arrows). (c) Clear ezrin expressionunctional connections. (d) As shown here, there is frequent, well dhere are associations with long tono-filaments that are intermediate
Important lateral specializations regulate trafficetween vaginal epithelial cells throughout the ovarianycle [17]. Immuno-reactive-ezrin was demonstrated
n the areas of these tight junctions, zona adherensnd desmosomes. There is also a correlation betweenr-ezrin and cellular fate, as vaginal cells maturingowards the surface change from a round shape to amflcm
tional connections indicated ezrin localization and the presence ofs) is shown on the transverse and longitudinal section of spinousd specialization in the form of desmosomes surrounding each cell.These junctional complexes stain heavily with ezrin (arrows).
olygonal shape, flatten, express keratin and undergoerminal differentiation, including nuclear pyknosisnd desquamation. Others have shown that different
echanisms may control trans-cellular transport ofuids and electrolytes [18], however, our findings indi-ates that ezrin may be involved through its structuralodification of the junctures between vaginal epithe-
40 A. Fadiel et al. / Maturitas 60 (2008) 31–41
F Cells hb of (a),b e hemi-(
lireef
ds
ig. 8. (a) Immuno-electron micrographs of cells from SG and LP.undle in lateral cytoplasm (large arrow). (b) High magnificationundles. (c) Epithelial cells of the papillae have ir-ezrin around thstars). (d) High magnification of (c).
ial cells. These findings suggest that ezrin may be anmportant protein that contributes to vaginal epithelial
emodeling. This may explain the relationship betweenzrin expression and location to the superficial lay-rs of the vaginal epithelium, a characteristic that wasound to be associated with proliferation/decreased celltbsa
ave ir-ezrin around the hemi-desmosomes (thin arrows) and largeshows hemi-desmosomes (arrows) and intra-cytoplasmic filamentdesmosomes (arrows) and with connected bundles of actin fibrils
eath [10,19–21]. At the ultra-structural level, ir-ezrintaining was most dense in the area of lateral specializa-
ions, the desmosomes and other junctional complexesetween vaginal cells cell–cell junctions. These lateralpecializations may support the vaginal epithelium andssist in the maintenance of its functions, including the
aturitas
rpaosfsfiecon
bit
A
l
C
i
R
[
[
[
[
[
[
[
[
[
[
[
[
tion are stimulated by interleukin 1 alpha and epidermal growth
A. Fadiel et al. / M
egulation of the vaginal pH, flora and might influenceermeability of vaginal epithelium [22]. Ezrin playsimportant role on cell migration and reorganization
f cytoskeleton for cellular movement. Ezrin expres-ion by both the VE and lamina propria may be a keyactor in the development and maturation of this tis-ue. In addition to effects on epithelial growth andunction, the vaginal wall is important for structuralntegrity and support of the pelvic floor. The finding ofzrin in fibroblasts, vascular endothelium and leuco-ytes implies similar functions of ezrin, and possiblyther MERM proteins in these components of the vagi-al wall.
The results of this study implicate a relationshipetween structural proteins such as the MERM fam-ly and the specialized function and cytoarchitecture ofhe vagina.
cknowledgement
We give special thanks to T.J. Scalise for her excel-ent assistance in the preparation of this manuscript.
onflicts of Interest
The authors confirm that there are no conflicts ofnterest.
eferences
[1] Lavy G, Whitten PL, Naftolin F. Introduction to vertebratereproductive endocrinology. In: Pang PKT, Schreibman MP,editors. Vertebrate endocrinology: fundamentals and biomed-ical implications, vol. 4. New York: Academic Press; 1991. p.1–22.
[2] Richman S, Naftolin F. Evolution of the mammalian reproduc-tive tract and placentation. In: Mor G, editor. Immunology ofPregnancy. Eurekah.com; 2004. p. 1–8.
[3] Song J, Fadiel A, Edusa V, et al. Estradiol-induced ezrin overex-pression in ovarian cancer: a new signaling domain for estrogen.Cancer Lett 2005;220:57–65.
[4] Simoncini T, Scorticati C, Mannella P, et al. Estrogenreceptor alpha interacts with Galpha13 to drive actin remod-
eling and endothelial cell migration via the RhoA/Rhokinase/moesin pathway. Mol Endocrinol 2006;20(8):1756–71.[5] Louvet-Vallee S. ERM proteins: from cellular architecture tocell signaling. Biol Cell 2000;92:305–16.
[
60 (2008) 31–41 41
[6] Saotome I, Curto M, McClatchey AI. Ezrin is essential forepithelial organization and villus morphogenesis in the devel-oping intestine. Dev Cell 2004;6:855–64.
[7] Algrain M, Turunen O, Vaheri A, et al. Ezrin containscytoskeleton and membrane binding domains accounting forits proposed role as a membrane-cytoskeletal linker. J Cell Biol1993;120:129–39.
[8] Li JG, Chen C, Liu-Chen LY. Ezrin-radixin-moesin-binding phosphoprotein-50/Na+/H+ exchanger regulatoryfactor (EBP50/NHERF) blocks U50,488H-induced down-regulation of the human kappa opioid receptor by enhancingits recycling rate. J Biol Chem 2002;277:27545–52.
[9] Bretscher A, Edwards K, Fehon RG. ERM proteins and mer-lin: integrators at the cell cortex. Nat Rev Mol Cell Biol2002;3:586–99.
10] Otey CA, Rachlin A, Moza M, et al. The palladin/myotilin/myopalladin family of actin-associated scaffolds. Int Rev Cytol2005;246:31–58.
11] Kluger M. Vascular endothelial cell adhesion and signalingduring leukocyte recruitment. Adv Dermatol 2004;20:163–201.
12] McRobert EA, Tikoo A, Gallicchio MA, et al. Localization ofthe ezrin binding epitope for glycated proteins. Ann N Y AcadSci 2005;1043(June):617–24.
13] Baumgartner M, Patel H, Barber DL. Na(+)/H(+) exchangerNHE1 as plasma membrane scaffold in the assembly of signal-ing complexes. Am J Physiol Cell Physiol 2004;287(October(4)):C844–50.
14] Miller KG. A role for moesin in polarity. Trends Cell Biol2003;13(April (4)):165–8.
15] Somogyi P, Takagi H. A note on the use of picric acid-paraformaldehyde-glutaraldehyde fixative for correlated lightand electron microscopic immunocytochemistry. Neuroscience1982;7:1779–83.
16] Eichenbaum K, Eichenbaum J, Fadiel A, et al. Minimally inva-sive method for murine brain fixation. BioTechniques 2005:39.
17] Kress A, Regli C, Spornitz UM, Morson G. Changes in theepithelium of the vaginal complex during the estrous cycle ofthe marsupial Monodelphis domestica. 2. Scanning electronmicroscopy study. Cells Tissues Organs 2004;178:48–59.
18] Gorodeski GI. Aging and estrogen effects on transcervical-transvaginal epithelial permeability. J Clin Endocrinol Metab2005;90(January (1)):345–51 [Epub October 13, 2004].
19] Regli C, Kress A. Changes in the epithelium of the vaginalcomplex during the estrous cycle of the marsupial Monodelphisdomestica. 1. Transmission electron microscopy study. CellsTissues Organs 2002;172:276–96.
20] Fais S, De Milito A, Lozupone F. The role of FAS to ezrin asso-ciation in FAS-mediated apoptosis. Apoptosis 2005;10(October(5)):941–7.
21] Chen Z, Fadiel A, Feng Y, et al. Ovarian epithelial carcinomatyrosine phosphorylation, cell proliferation, and ezrin transloca-
factor. Cancer 2001;92:3068–75.22] King BF. The permeability of nonhuman primate vaginal epithe-
lium: a freeze-fracture and tracer-perfusion study. J UltrastructRes 1983;83:99–110.
Related Documents











