External brain morphology in rhesus macaques (Akcaca mulatta) Although endocasts from fossil primates have been extensiveiv examined in several previous studies, relatively little is currently known about the genetic and environmental factors that affect brain development in extant non-human primates. We examined the effects of age and sex on brain sizr and external morphology in a large sample of rhesus macaque endocastq from the Cave Santiago skeletal collection. Using a repeated mcasurcs design. we demonstrated that brain sulcus lengths can be reliably assessed with current computer input technology and careful trait defmttion. :\,gc- and sex variation in s&us lengths and endocranial capacity were also analyzed using ANCOVA and maximum likeiihood estimation methods. Our cross-sectional data indicated that total cranial capacity increases postnatally in the macaque, while the cortical sulci appear to have reached their adult length at or soon after birth. Cranial capacity reaches its adult volume at 3-57 years in females, but not until 6.08 years in males. We also, found considerable sexual dimorphism in brain sulcus lengths with males iarger than females, particularly for the more posteriorly located sulci. ‘I‘hr dimorphism is, however, a function of overall brain size. .&f&r controlling for cranial capacity, there is no sexual dimorphism for sulcal lengths. Introduction The evolution of the primate brain has a central role in many anthropological and natural historical debates (e.g., Armstrong, 1985; Armstrong & Falk, 1982; Falk, 1978a, 19786, 1982; Gould, 1975; Holloway, 1968, 1978; Jerison, 1973, 1982; Martin, 1982; Martin Sr Harvey, 1985; Noback, 1970; Passingham, 1973; Radinsky, 1975). Ultimately, the descriptions of brain morphology which must support these theoretical discussions are based upon phenotypic observations of brains in extant primates, or on endocast evidence from fossil forms. In either case, the brains are usually characterised by linear and volumetric measurements which are thought to accurately reflect the underlying genetic variation. Unfortunately, little attention has been paid to the repeatability or precision of these measurements. Additionally, because the majority of past studies have used sampies * ‘To whom correspondence should be addressed. 0047-2484/!30/030269 + 16 $03.00/O 0 1990 Academic Press Limited

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

External brain morphology in rhesus macaques (Akcaca mulatta)
Although endocasts from fossil primates have been extensiveiv examined in several previous studies, relatively little is currently known about the genetic and environmental factors that affect brain development in extant non-human primates. We examined the effects of age and sex on brain sizr and external morphology in a large sample of rhesus macaque endocastq from the Cave Santiago skeletal collection. Using a repeated mcasurcs design. we demonstrated that brain sulcus lengths can be reliably assessed with current computer input technology and careful trait defmttion. :\,gc- and sex variation in s&us lengths and endocranial capacity were also analyzed using ANCOVA and maximum likeiihood estimation methods. Our cross-sectional data indicated that total cranial capacity increases postnatally in the macaque, while the cortical sulci appear to have reached their adult length at or soon after birth. Cranial capacity reaches its adult volume at 3-57 years in females, but not until 6.08 years in males. We also, found considerable sexual dimorphism in brain sulcus lengths with males iarger than females, particularly for the more posteriorly located sulci. ‘I‘hr dimorphism is, however, a function of overall brain size. .&f&r controlling for cranial capacity, there is no sexual dimorphism for sulcal lengths.
Introduction
The evolution of the primate brain has a central role in many anthropological and natural
historical debates (e.g., Armstrong, 1985; Armstrong & Falk, 1982; Falk, 1978a, 19786, 1982; Gould, 1975; Holloway, 1968, 1978; Jerison, 1973, 1982; Martin, 1982; Martin Sr
Harvey, 1985; Noback, 1970; Passingham, 1973; Radinsky, 1975). Ultimately, the descriptions of brain morphology which must support these theoretical discussions are based upon phenotypic observations of brains in extant primates, or on endocast evidence
from fossil forms. In either case, the brains are usually characterised by linear and volumetric measurements which are thought to accurately reflect the underlying genetic variation. Unfortunately, little attention has been paid to the repeatability or precision of these measurements. Additionally, because the majority of past studies have used sampies
* ‘To whom correspondence should be addressed.
0047-2484/!30/030269 + 16 $03.00/O 0 1990 Academic Press Limited

270 1.. KONIGSBEK(; ET .\I..
of unknown age and genealogical relationship, there is no direct means for analyzing brain growth or partitioning phenotypic variation into genetic and environmental components.
We recently assessed variation in external brain morphology of the rhesus macaque
using two advancements in measurement and sampling which make this research unique. First, we used three-dimensional (3-D) computer digitisation to increase ~neasur~nlellt precision. Second, we sampled macaques of known demographic status and maternal relationships from the free-ranging colony on Cayo Santiago, Puerto Rico, so that heritability of neural traits could be estimated using the known maternal relationships. As
an additional benefit of using animals whose life histories are known, we were able to assess
the effects of age and gender on external brain morphology.
Because the use of 3-D computer digitisation is a novel approach to quantifying primate
brain morpholo~ (Falk et al., 1986), we first report the results of a repeated measures
analysis for a small subsample. Following this, we present analyses of the effects of age, sex
and size variation on cranial capacity and brain sulcus lengths. We use the results of these analyses to evaluate the feasibility of studying fossil endocasts when age-at-death and/or
sex are unknown, to discuss the developmental and growth patterns of the macaque brain, and to determine whether these factors need to be controlled in comparative studies and in studies assessing the level of genetic variation in neural traits.
Materials and methods
The Cayo Santiago rhesus macaque colony is composed of descendants of wild-trapped macaques who were placed on the island in 1939 (see Sade et al., 1985 for a history of the
colony). Systematic observation of the colony began in the mid 1950s and included the identification of genealogical relationships and collection of basic demographic data. For our current analysis we use the skeletal remains of330 animals of known sex and age at death. Ages were generally precise to within a week (Sade et al., 1985; Busse, personal communication), but because of uncertainty in exact date of birth, a few ages were known only to within a year (Falk, et al., 1989a).
Cranial endocasts were prepared by DF using standard methods (Falk, 1978a). A string of points were placed along each of 11 bilateral sulci at approximately every 2 mm by RH (see Appendix for definition of sulci and Figure 1 for their description) and were then digitised using a 3Space electromagnetic digitiser (~ildebolt & Vannier, 1988) operated by CW. The distances between points along a sulcus were summed to provide a measure of sulcus length. A total of 330 individuals were represented in the sample analyzed here.
As an initial step in the development of reliable cortical surface measures, a second, replicate endocast was prepared by DF from 15 of the 330 crania. Each of these endocast pairs was dotted once by RH for 24 bilateral sulci (see Appendix), and then digitised twice by CH. Following an initial repeatability study it was clear that 14 sulci could not be reliably measured or their end points reliably defined. As a result, these sulci were eliminated and definitions of some other sulci were altered to improve repeatability. We then undertook a second repeatability study on the ten remaining sulci. In this paper, we report the results of the second repeatability study, because it analyzes sulci as they were defined in the ensuing study. (It should be noted that the final study is based on 11 rather than 10 features because the superior temporal sulcus was divided into two parts after the second repeatability study took place.)
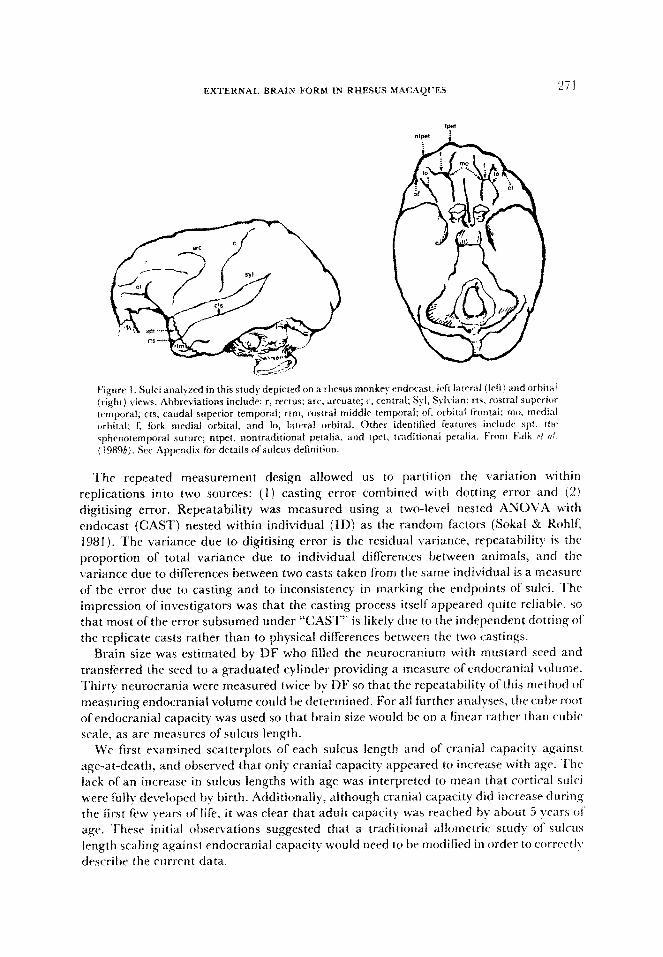
EXTERNAL BRAIN FORM IN RHESUS MAC.kQC??.S “71
E’igurr 1. Sulci analyed in this study depicted on a rhesus monkry endncast, left lawral (irft) and orbital (right) views. Abbreviations include: r, rectus; arc. arruate; c, central; $1, St Ivian; rts, rostra1 sup~i~ temporal: cts, caudal superior tmporal; rtm, rostra1 middle temporal; of, orbital frontal: mo. medial orbital; 1; fork medial orbital, and lo, lateral orbital. Other identified features inrludr spt. tlw s~b~~~ternp~~ra~ suture; ntpet, nontraditional petalia, and tpct, traditirmai pf%alia. From b’alk ri oi (198%). SW Appendix fbr details ofsuicus definition.
The repeated measurement design allowed us to partition the variation within replications into two sources: (1) casting error combined with dotting error and (2) digitising error. Repeatability was measured using a two-level nested ANOVA with
endocast (CXST) nested within individual (ID) as the random factors (Sokal & Rohlf, 1981). The variance due to digitising error is the residual variance, repeatability is the
proportion of total variance due to individual differences between animals, and the variance due to differences between two casts taken from the same indiv~dua1 is a measure of the error due to casting and to inconsistency in marking the endpoints of sulci. The impression of investigators was that the casting process itself appeared quite reliable, so that most of the error subsumed under “CAST” is likely due to the independent dotting of
the replicate casts rather than to physical differences between the two castings. Brain size was estimated by DF who filled the neurotranium with mustard seed and
transferred the seed to a graduated cylinder providing a measure of endocranial volume.
Thirty ne~~rocrania were measured twice by DF so that the repeatability of this method of measuring endocranial volume could be determined. For all further analyses, the cube root of endocranial capacity was used so that brain size would be on a linear rather than cubic scale? as are measures of sulcus length.
We first examined scatterplots of each sulcus length and of cranial capacity against
age-at-death, and observed that only cranial capacity appeared to increase with age. The lack of an increase in sulcus lengths with age was interpreted to mean that cortical sulci were fuIlv developed by birth. Additionally, although cranial capacity did increase during the first few years of life, it was clear that adult capacity was reached by about 5 years 01 age. These initial observations suggested that a traditional allometric study of sulcus length scaling against endocranial capacity would need to be modified in order to correctI>, describe the current data.

Ixbeau rt 01. iiY86: l W) arg-11' that if‘agr i\i kno\~~rt it should 1~ rrtili0r.d in an il~~Ol~Jf'~l ic
analysis, and that the scaling 4‘ one org;in ;lgaiml ;tnothcr is “ii tlsrfitl hut irnprrliv~l substitute for the more corrrplrtc description ttt’ growth which can Iw giwn wht.n ;tgr i5 known.‘” Because age-at-death is known for the C:ayo macaques. we perli~rmcd a~ mai\-sis of “time allomrtry,” or regression of trait size on age (‘JolicoPur. 1 YII5l. Hccaure WC takr
endocranial capacity as an overall measure of brain size. w(‘ 1nust tirst ;itttmpt to modri thr
growth of endocranial capacity, then groM.th of the sulci, and finall!. attrrr~pt to dclinratc
the proportionality between brain sizr and sulcus lengths 3s a hv-product 01 rVlativc
growth.
In Huxley’s \ 1932 j originai statement of the allometry equation an organ’s growth was seen as multiplicati~~e throughout the fife of an organism. Thus. viewed against age 10, an organ’s size tp) would be:
p = 13’: (I)
where R and C are the constants describing the exponential curve. Many organs do not grow continuously and exponentially throughout life (see Chapprll, 1989). As a
consequence. growth curves which feature upper asymptotes have been proposed (summarised in Jolicoeur & Pirlot, 1988). To model the growth of endocranial capacity, wt
selected one of the simplest curves. a linear response and plateau (LRP) function that is
written as:
y = d + bBt t (l-6)&. ($2 1
where 6 equals zero when t is greater than or equal to t, and b equals 1 when t is less than 1,. With this function, organ size begins at size A (t = 0) and increases linearly with age, at rate B, until growth ceases (at age tC). This is the linear response part ofthe function. From the
age of growth cessation (lC) and onward the size of the organ is fixed at a plateau. The function has three parameters: an intercept (.4}, slope jB), and age ofgrowth cessation it,). Prunty (1983) showed that the LRP function is a special cast of a continuous function, which becomes the piece-wise linear LRP function in its limit. ‘To Gt the LRP function to endocranial capacity, we separately regressed the cube root of cranial capacity in
millimeters on age. Because the LRP function is intrinsically non-linear, it cannot be estimated using the
ordinary least-squares regression approach. The parameters in this function can, however, be obtained by maximum likelihood estimation (IMLE), and indeed any statistical package
which contains a non-linear regression module and the ability to handle a parameter used in an inequality (i.e., I, in equation (2)] . can be used to fit the LRP function. For example, Systat’s NONLIN module (Wilkinson, 1988) can be used to do piece-wise linear regression, of which the LRP is a special case. However, Systat, as well as many other statistical packages, cannot handle the problem of’ boundaries on parameters. For example, t, should not be allowed to become negative, although it should he possible t(J
evaluate the likelil~ood when t, is on this boundary (i.e., when t, equals zero). For out
anaiysis, we fit the LRP using standard maximum likelihood methods. The log-likelihood of obtaining the observed data given a candidate set of parameters can be written as:
In(r,) = -0.5 X ln(V? -I l/(21/3 j, (~1, -_jji)’
wheregi is the observed cube root of cranial capacity for individual i, the predicted values are taken from equation (2}, and C’refers to variance around the regression line. ?Be maximum likelihood estimates of ,4, K and 1, in equation (2) are found by iterativelv

EXTERNAL BRAIN F‘ORM IX RHESLFi M4t:AQl’E” 273
evaluating the log-likelihood function with different values of A, B and /< until the marimum log-likelihood is identified. In order to search the log-likelihood fimctictn VW’
used the FORTRAN subroutine GEMINI (T,alouel, 1979). which uses a “quasi-Newton”
or variable metric method. The interested reader can find a short account of this rncthod
and further references in Press et al. 11986: 307 312).
'I he MLE method provides an ideaf means fi)r statistically testing altcrnatil-TV hypotheses describing brain growth, and consequently WC’ considered a number ofcliffrr~n I regressions in our analysis. In the most ,general regressir>n. the slopes, i~t~rceJ~t% a!p 01’
cessation and variances around the regre$sicJl~ lines were allowed to difrer fi)r males and f&males, so that eight parameters were estimated. In 1116rc’ restricted models, some OL thc.sc*
paritmeters werr set equal in the two sexes. and only ;t c‘tJmutJn parameter was t%‘timatcd it! pfacr of the previous two parameters. Because reduced modtls ~JT special caces of‘thr me)rr‘
g-encraf regression equation, we can test whethrt. the reduction in number of parani(cff*r5 betM,een two regressions leads t(J a signifir.ant reduction in the ability of’ the ~no&i IO
describe the data. Negative two times the di&rence in thra log-likelihoods bet\$.t.rn tilt. t\\o rcgrcssions is asvmptotically distributed as 21 chi-square, with degrees of frcedonl cq11:11 10 the dif%rencr in number of parameters included in the tuxr regressions.
At’ter fitting the LRP function to endocranial capacity for males and females. M-V
tastimated separate analyses of covariance (:~NC:C)L.rl) 1. ix endocranial capacity and ~a&
sulcus against age for indi~~iduals less than 5 scars old and then for indisiduafs aged 5 vears
or older. The 5 k-ear age cut-off corresponded approsimatrlp to the age ofcranial capacit) grab th cessation for males and females. Thr full ANCX>VA model contained the 1~~t.tor.h age, sex and age by sex interaction. This analysis indicated. as we had anticipatt>d. that
only cranial capacity grew during the early (less than 5 )‘c*ars old) period, thilt no nt*llral
structures grew during the later period. and that there was considerable sr,sual
dimorphism for cranial capacity and suicus lengths throughout life. Because of the sexual dirnorptlis~l obsc*rved in man) neural characters, WC wvrc
interested in whether this dimorphism was due solely to overall size differences. In a classic
bivariate allometric analysis sulcus lengths would bc rrgrrssed on cranial capacities and
tested for slope or intercept differences between ttle sexes. (:are is necessary here. hcJw(‘\.t’r., to distimguish between cross-sectional ontogenetic: allometr). and static (adult) allomrtr!~ (Clock, 1966). Because sulcus lengths do not appear to increase after birth, the relationship between any pair of sulci aCrUsS the entire lif@ span describes the static allometr~ ofthes?
traits. Scaling between the sulci would cons~qu~,ntl~ be viewed as an end result of th(t
growlb rate of each sulcus against time, the relative times of cessation ofgrowth fi)r tsach sulcus, or a conglomeration of both ef’fects. On the other hand. if the relationship bt-twt.t~~ cranial capacity and a given sulcus across the entire age-span is considered. then th(
analysis would incorrectly mix ontogenetic with static allometry, as the cranial capacit)
wol~ltl be growing during the early years. Consequently, the scaling ofeach sulcus against CraJliil~ capacity was considered only for those individuals who wcrc 5 years or older ;rt time of death.
Results
Repeatabilip
Repeatability analyses were performed to guide in the precise definition of measurements and to help provide guidelines for scoring sulci as missing. Table 1 ContrCns the proportions of variance due to individual, cast and digitisirig error for each of ten paired sulci. Data are
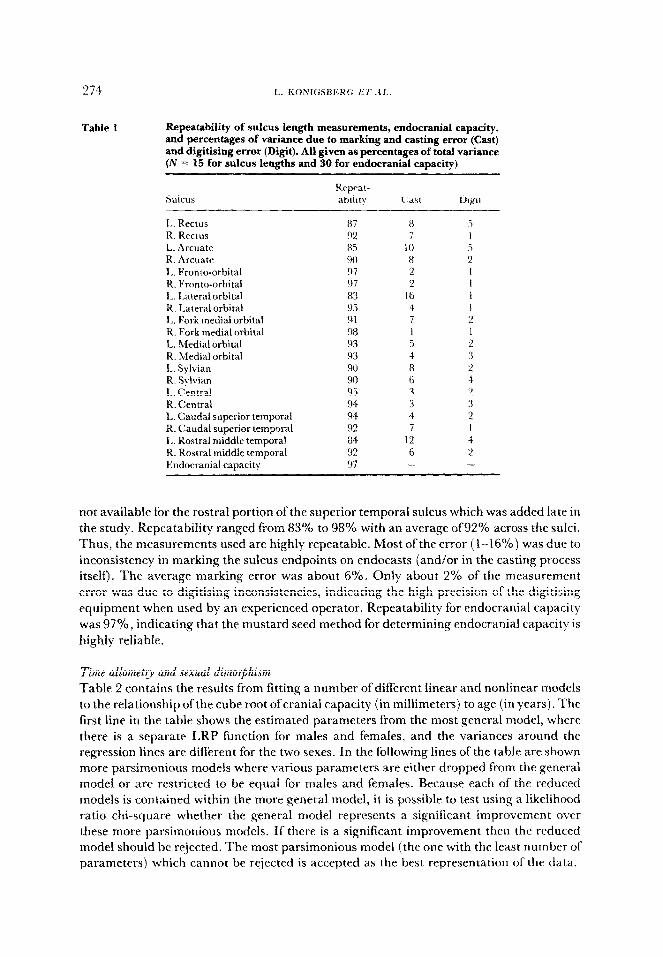
274
Table 1 Repeatability of sulcus length measurements, endocranial capacity, and percentages of variance due to marking and casting error (Cast) and digitising error (Digit). All given as percentages of total variance (N = 15 for sulcus lengths and 30 for endocranial capacity}
1,. Rectus R. Rectus L. Arcuate R. Arcuate L. Fronto-orbital R. Fronto-orbital I,. Lateral orbital R. Lateral orbital 1,. Fork medial orbital R. Fork medial orbital L. Medial orbital R. Medial orbital L. Sylvian R. Sylvian 1,. Central R. Central L. Caudal superior temporal R. Caudal superior temporal L. Rostra1 middle temporal R. Rostra1 middle temporal Endocranial caaacitv
7 I 0 8 2
2 I6
2
4
8
6
4
12 6
2
2 1
3 2
not available for the rostra1 portion of the superior temporal sulcus which was added late in the study. Repeatability ranged from 83% to 98% with an average of92% across the sulci.
Thus, the measurements used are highly repeatable. Most of the error ( i-16%) was due to inconsistency in marking the sulcus endpoints on endocasts (and/or in the casting process itself). The average marking error was about 6%. Only about 2% of the measurement error was due to digitising inconsistencies, indicating the high precision of the digitising equipment when used by an experienced operator. Repeatability for endocranial capacity
was 97%, indicating that the mustard seed method for determining endocranial capacity is
highly reliable.
Time al~~rn~t~~ and sexual dimorphism Table 2 contains the results from fitting a number of different linear and nonlinear models
to the relationship of the cube root ofcranial capacity (in millimeters) to age (in years). The first line in the table shows the estimated parameters from the most general model, where there is a separate LRP function for males and females, and the variances around the regression lines are different for the two sexes. In the following lines of the table are shown more parsimonious modets where various parameters are either dropped from the general model or are restricted to be equal for males and females. Because each of the reduced models is contained within the more general model, it is possible to test using a likelihood ratio chi-square whether the general model represents a significant improvement over these more parsimonious models. If there is a significant improvement then the reduced mode1 should be rejected. The most parsimonious model (the one with the least number of parameters) which cannot be rejected is accepted as the best representation of the data.

.?

From ‘I’ablr 2, it is clear that the variances around tlw L,RP tiln&ons in ~nalcs .u~rl
fernah can be constrained to be equal v = 0,7572), and that furthermore the \loprs it’
equal &jr males and females Ir, = 0.5597). The test ofequality of’interccpts shown in ‘T&II.
2 contains an unusual feature in that the age oi‘ growth cessation ii)r- females wrnt to a
bound of’zero. Because a11 ages are grearer than zero, this means that the equal intercept
model forces the females to have only a plateau phase, and no linear Iresponse. in any event,
the log-likelihood for fitting the equal intercept model was -98O.S-23, which easily leads to
its rejection against the general model. Moving down to the restricted case where 1, is Ibrced
to he equal for males and females, this model also must be rejectrd (p =I 0.0250). The linal
model, an ordinary linear regression with separate slopes and intercepts l’or males a11d
females (and t, dropped from the analysis), also can he rejected (p < OWOl ). 2’0
summarise these results, the most ~arsinl~)niolls model which adequately fits the data is
one where the male and female slopes and residual variances are equal, but the intercepts
and ages ofcessation differ. The ages of’male and female cessation are estimated at 6.08 and
3.57 years respectively, and are significantly different (~2 for model with equal slopes versus
model with equal slopes and fr’s is 8.1870 with one degree of freedom, p = @0042). Because
there is a considerable amount ofscatter around the regression lines (see Figures 2 and 3)
this model yields a relatively low coeffiri~nt of determination equal to 0.3807 across the
combined male and female sampies.
For females and males, we applied an ANCOVA to cranial capacity and sutcal Iengths
for individuals below 5 years of age and those above 5 years. Our decision to divide the
sample into these two categories was based on our findings from the LRP function that
cranial capacity did not increase after the age of 6.08 years in males and 3.57 years in
females. The results fiiom the ANC:C)VAs are reported in Tables 3 and 4, where all
measurements are expressed in centimeters and age is taken in )ICYUS. Prior to the
ANCC)VAs, we tested for homogeneity of slopes using the Li~lly saturated model including
an ~rlteraction between age and sex. l’his model provides a test of growth rate (slope)
difference between males and females tix the neural traits. In Tables 3 and 4, when the
interaction term was significant (at the OQ5 level), the male and f‘zmale slopes arc listed
separately with indications of statistical significance. If the slopes were not significantI)
different from each other, then the ANCO\‘A model was estimated without the interaction
term. In this model there was a common male-female slope; thus only the intercepts could
differ. The male and female intercepts (at age Zen,) are shown in Tables 3 and 4, along with
an indication ofwhether these intercepts were signi~cantl~ different from one another. For
traits which could be fit only with a full model (including age by sex interaction’). intercept
difrerences cannot be evaluated.
Table 3 indicates that age by ses interactions are uncommon for neural traits in
juveniles, so a single trait-against-age slope can he used for males and females. ‘This
indicates that there is no difference in growth rates between males and f’rmales for rnost
neural characters. Only two traits, the leli arcuate and right medial orbital sulcus lengths,
show significantly different male and female growth rates.
Considering the levels of significance for the regressions of neural traits on age, it appears
that only endocranial capacity is growing during the juvenile period. Cranial capacity
regressed on age has a positive slope 01~0.03, which is significantly diEerent from zero. In
contrast to brain size (as measured by cranial capacity), few of the sulci show significant
age-related variation. Regarding sex differences, cranial capacity and a number of the sulci
are significantly greater in males (contrcjlling for age as the covariate), as shown in the
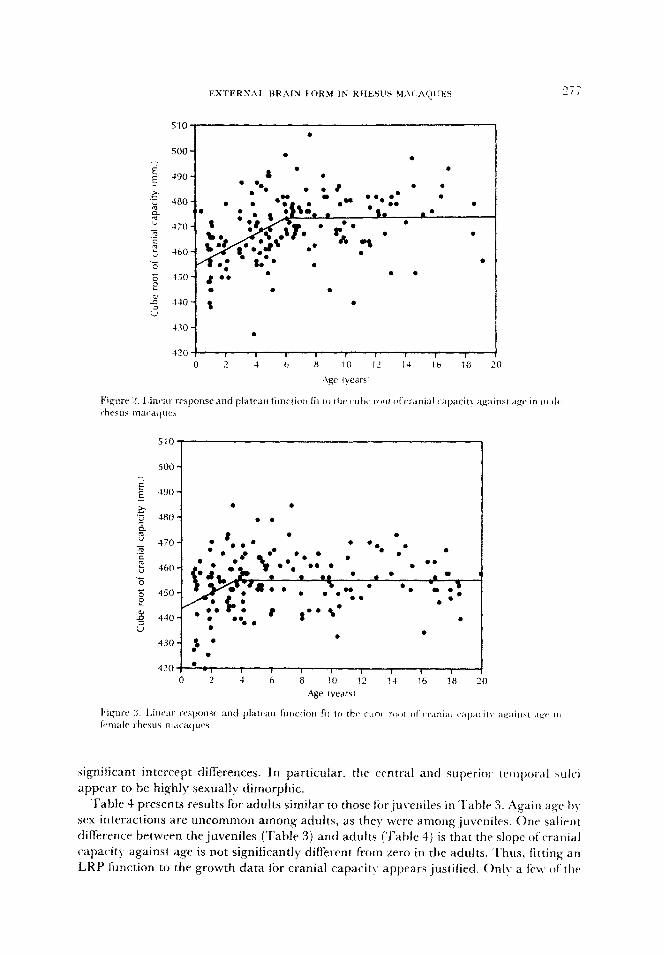
. .
480 * .
?(
significant intercept differences. In particular. the central and superior temporal ~Ici
appear to be highly sexually dimorphic.
Tabie 4 presents results for adults similar to those Lhr,juvenilcs in Table 3. Again age h!,
sex interactions are uncommon among adults, as they were among juveniles. Onr salient
difference between the juveniles (Table 3) and adults (Table 4) is that the slope ofcranial
capacit? against age is not significantly different from zero in the adults. Thus, fitting an
L,RP function to the growth data f&r cranial capacity appears justified. Only a few of thr,
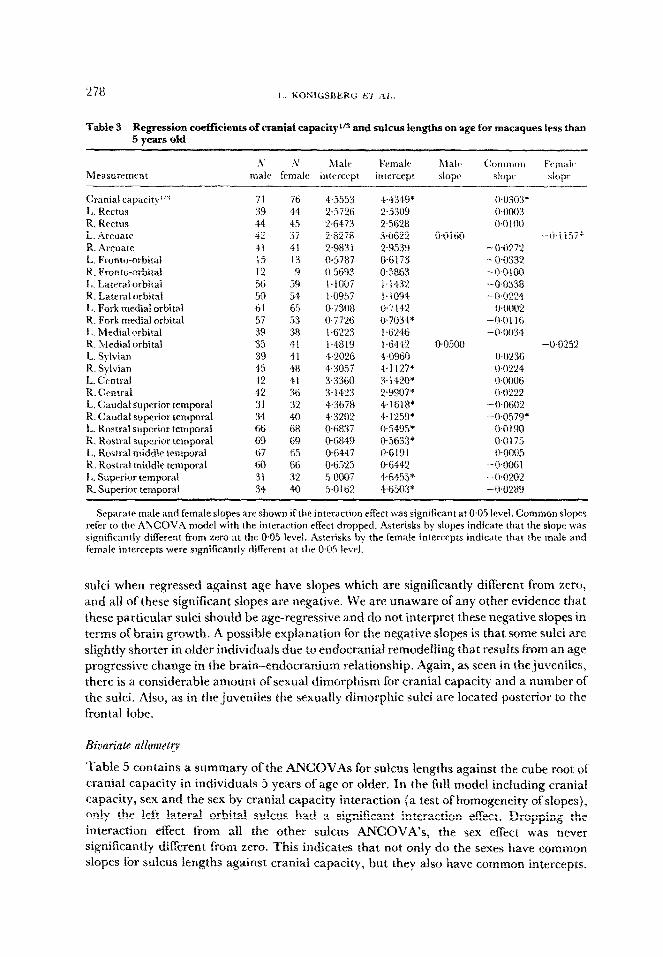
Table 3 Regression coeffkients of cranial capa~ity”~ and sulcus lengths on age for macaques less than 5 old years
h’ !V Male E’emalt~ h~falc (:otnmon f’cnndt~ Measurement male female intercept intercept slope slope siopr
Cranial capacity”” 71 76 4.5553 44349* Wo303* L. Rectus 39 44 2.5726 2.5309 0.0003 R. Rectus 44 45 2+i4?3 2,5628 c~~nloo L. Arcuate f2 37 2.8278 3Otj22 o-Ol60 -$I 157* R. Arcuate 4I 41 2.9831 2-9539 -0-0272 t. Fronto-orbitaL 15 13 0.5787 IT6173 -0.0332 R. pronto-orbital 12 9 0.5693 @5%63 -O-0100 L. Laterai orbitai 56 59 L-IO07 I-1432 -0-033X R. Lateral arbitai 50 54 1.0957 l.iO94 -WY124 L. Fork medial orbital 61 65 0.730% 0.7142 0.0002 R. Fork medial orbital 57 53 0.7726 0~7034* -0,0116 I,. Medial orbital 39 3% iW’27 L . 1.6246 -0-0034 R. Medial orbital 35 41 14819 I.6442 @050% -Wf25% L. Sylvian 39 41 4.2026 4.0960 0.0236 R. Sylvian 45 48 4.3057 “4~112P 0.0224 L. Central 42 41 3.3360 3.14!?0* WOO06 R. Central 42 36 3.14’S 2~9907” 0.0222 L. Caudal superior tcmporat 31 32 4.3678 4.161%* -0+602 R. Caudal superior temporal 34 40 4.3202 4.1259* -0.0579* L. Rostrai superior temporat 66 6% 06837 O?i495’ 0~019tIl R. Rostra1 superior temporal 69 69 &6849 056W OGL 75 L. Rost~aL middle temporal 67 65 0.644 7 O-iii91 0~0005 R. Rostrai middle temporal 60 66 WzQ5 06441 -00061 I,. Superior temporal 31 32 5Gx?7 +64.55* -OWX! R. superior temporal 34 40 5-O Iii2 :4~65tB* -0~0289
Separate male and female slopes are shown if the interaction effect was significant at 0.05 level. Common slopes refer to the ANCOVA model with the interaction effect dropped. Asterisks by slopes indicate that the slope was significantly different from zero at the 0.05 level. Asterisks hy the female intercepts indicate that the male and female intercepts were si~ni~cantly different at the 0.05 level.
sulci when regressed against age have slopes which are significantiy diflerent from zero, and ail of these significant slopes are negative, We are unaware of any other evidence that these particular sulci should be age-regressive and do not interpret these negative slopes in terms of brain growth. A possible explanation for the negative slopes is that some sulci are slightly shorter in oIder indiv~dua~s due to endocranial remodeffing that results from an age progressive change in the brain~ndocranium relationship. Again, as seen in the juveniles, there is a considerable amount of sexual dimorphism for cranial capacity and a number of the sulci. Also, as in the juveniles the sexuaily dimorphic suici are located posterior to the frontal lobe.
&variate atlomety
Table 5 contains a summary of the ANCOVAs for sulcus lengths against the cube root of cranial capacity in individuals 5 years of age or older. In the full model including cranial capacity, sex and the sex by crania1 capacity interaction {a test of homogeneity of slopes), only the left lateral orbitat ~ulcus had a significant interaction effect. Dropping the interaction effect from all the other sulcus ANCOVA’s, the sex effect was never si~n~~~nt~y different from zero. This indicates that not only do the sexes have common slopes for sulcus lengths against cranial capacity, but they also have common intercepts.
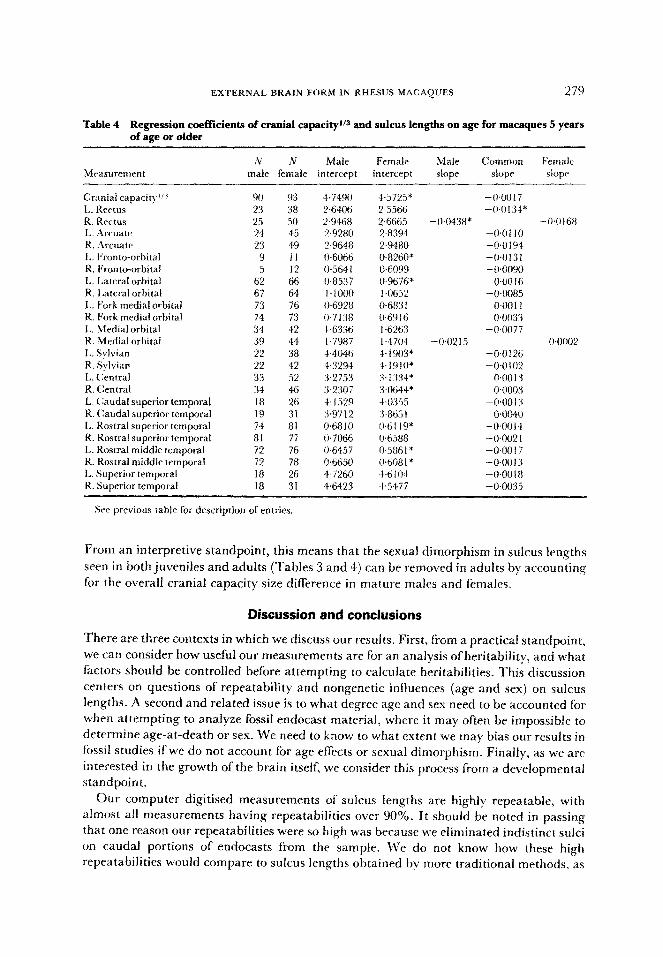
EXTERNAL BRAIN FORM IN RHESUS MACAQUES 279
Table 4 Regression coefficients of cranial capacityl’s and sulcus lengths on age for macaques 5 years of age or older
‘V N Male Female Malt ~ornrn(ln Fern&
male female intercept intercept slope slope slopr
Cranial capacity’i’i 1,. Rertus R. Rcctus L. .hYlate R. .2rcuatc 1,. ~r[~nto-orbital R. Fronto-orbital I,. Lateral orbital R. Lateral orbital L. Fork medial orbital R. Fork medial orbital L. .Medial orbital R. Medial orbital I.. S+ian R. Sylvian L. Chtral R. (:entral L. Chudal superior temporal R. Caudal superior temporal I,. Rostra1 superior temporal R. Rostra1 superior temporal L. Rostra1 middle temporal R. Rostrai middle temporal L. Superior temporal R. Superior temporal
90 93 23 38 25 50 24 45 23 49
9 I1
6; 2
67 64 73 76 74 73 34 42 39 44 22 38 22 42 33 52 34 46 18 26 19 31 74 81 81 77 72 76 72 78 18 26 18 31
47490 4~572.Y -rho0 17 2.6406 2.5566 -0~0134* 2.94ti8 2.666.5 -lw438* -0~olti8 2.9280 2.8394 -0~0 I IO 2.9648 2.9480 -o+l194 (1.6066 0+x160* -@OiYf 0.564 I 0~6099 - O.O(?9lf 0.853 7 0.9676* 0.00 I6 1~1000 1 ~0652 -O+O85 06928 @68:i I 0~0011 0.7338 0.6916 00033 I ,6336 I .6263 -0.0077 1.7987 I .4704 -0.0215 0~0002 44046 4- 1903* -0.0126 4.3294 -P1910* -@0102 :x,2753 3.1:134* @OO 13 3.2307 3.0644’ 4~0003 4~152!2 +0355 -0w1 .Y 3.9712 3%5 I @OO40 0.68 10 WI 19* -0GO14 0~7066 OG88 -0.002 I 0.6457 0.586 I * -0~0017 II.6650 0608 I * -O-OOl3 4.726(1 +- ti I 1J4 -o~ooIs 4.6423 4.5477 -0.0035
Se previous table for description of entries.
From an interpretive standpoint, this means that the sexual dimorphism in sulcus lengths
seen in both juveniles and adults (Tables 3 and 4) can be removed in adults by accounting
for the overall cranial capacity size difference in mature males and females.
Discussion 8nd conclusions
There are three contexts in which we discuss our results. First, from a practical standpoint, we can consider how useful our measurements are for an analysis ofheritability, and what
factors should be controlled before attempting to calculate heritabihties. This discussion centers on questions of repeatability and nongenetic influences (age and sex) on sulcus lengths. A second and related issue is to what degree age and sex need to be accounted for when attempting to analyze fossil endocast material, where it may often be impossible to determine age-at-death or sex. We need to know to what extent we may bias our results in fossil studies if we do not account for age effects or sexual dimorphism. Finally, as we are
interested in the growth of the brain itself, we consider this process from a developmental standpoint.
Our computer digitised measurements of sulcus lengths are highly repeatable, with almost all measurements having repeatabilities over 90%. It should be noted in passing that one reason our repeatabilities were so high was because we eliminated indistinct sulci on caudal portions of endocasts from the sample. We do not know how these high repeatabilities would compare to sulcus lengths obtained by more traditional methods, as
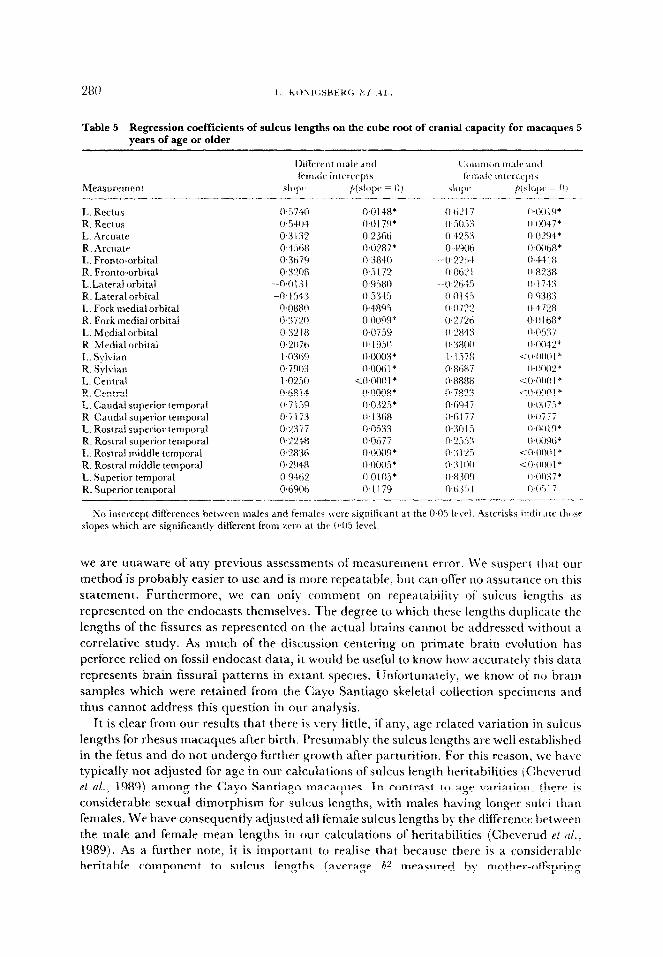
Table 5 Regression coefficients of sulcus lengths on the cube root of cranial capacity for macaques 5 years of age or older
L. Rectus R. Rectus L. Arcuate R. Arcuate I.. Front+orbital R. Front+orbital L,Lateral orbital R. Lateral orbital I.. Fork mediai orbital R. Fork medial orbital I,. Medial orbital R. Medial orbital L.. Sylvian R. Sylvian L. Central R. Central L. Caudal superior temporal R. Caudal superior temporal I,. Rostra1 superior temporal R. Rostra1 superior temporal 1.. Rostra1 middle temporal R. Rostra1 middle temporal I,. Sup&or temporal R. Superior temporal
we are unaware of any previous assessments of measurement error. \Ve suspert that auf
method is probably easier to use and is more repeatable, but can offer no assurance on this statement. Furthermore, we c‘an only comment on repeatability of sulcus lengths as
represented on the endocasts themselves. The degree to which these lengths duplicate the lengths of the fissures as represented on the actual brains cannot be addressed without a correlative study. As much of the discussion centering on primate brain evolution has perforce relied on fossil endocast data. it would be useful to know how accurateI)- this data represents brain fissural patterns in extant species. Unfortunate~y~ we know of no brain
samples which were retained from the Cayo Santiago skeletal collection specimens and thus cannot address this question in our analysis.
It is clear from our results that there is ver) little, if any, age related variation in sulcus
lengths for rhesus macaques after birth. Presumably the sulcus lengths are well established in the fetus and do not undergo further growth after parturition. For this reason, we have typically not adjusted for age in our calculations of sulcus length heritabilities iC:hevcrud et al., 19891 among the Cay0 Santiago macaques. In contrast to age variation, there is considerable sexual dimorphism for sulcus lengths, with males having longer sulci than females. We have consequently adjusted ail female sulcus lengths by the difference between the ma.le and female mean lengths in our calculations of heritabilities (Cheverud et cl/., 1989). As a further note, it is important to realise that because there is a considerable heritable component to sulcus lengths (average h” measured by mother-of&spring

EXTERNAI. BRAIN FORM IN RHES17!, MAC:AQI:ES “81
regressions across 20 sulci equal 0.3 1, Cheverud et al., 1989), our finding that these traits do
not vary with age is likely to be robust to neglecting the non-independence among related
animals. Ignoring non-independence should inflate the probability ofa type I error, so that
thr null hypothesis of no age effect would be falsely rejected at a greater rate than the
nominal level of 0.05. Therefore, if an age eff‘ect is not significant here, then it should not be
significant in the complete design where both heritability and age effects arc
simultaneously estimated.
The observation of age-stable sulcus lengths in macaques is encouraging i?n
comparative studies and studies of fossil primate endocasts. Because sulcus lengths do not
appear to increase with age (even in the very young). it appears reasonable to use
spcscimens of differing ages-at-death without correcting for age differences. Of (XILI~SC, ‘IV
there is sexual dimorphism among the macaques for sulcus lengths, errd~,cdsts should nc)t
be compared without regard to the sex of the indiCduals. Howe\-er. the lack of&pr and
intercept differences between the sexes suggests that a mc’asur~ ofbrain size c,ould be used
to remove the sexual dimorphism in sulcus lengths (see Corruccini, 1987 for a discussion 01
\rarious methods for size corrections). Becausr the relationship between sulcus lengths and
cranial capacity- changes in,juveniles (see above), such ;I procedure should br applied onl!
in adults.
Regarding the developmental aspects of the macacluc brain, we can onI> comment WI
postnatal growth. As the macaque neonate’s brain volume is already approximateI>. 60%
the size of the adult’s (Holt et al., 1975). our postnatal data postdates much uf‘the ,grow t h and development of brain external anatomy. In both humans (Dabbing & Sands. 197Si
and macaques (Holt et al., 1975). the inflection point 01’ the Ggmoidal growth curvf‘ lor
brain weight is reached at or near birth. Consequently, our use CJf an LRP fimction to lit
postnatal growth in cranial capacity for macaques seems justified. Less data are availablr
concerning prenatal development of the sulci (see (:hi et al.. 1977 for human data I, ttlougl~ it stems cleat, from our analysis that the sulci are Ml>, developed h) or \oon after birth ill
the macaque.
l‘hc lack of increase in cortical sulcus lengths with age gi\,en growth in cranial capacir!
mav be due to differences among brain parts in timing ofgrowth. The c-erebrurn, on \\hich
the sulci appear, reaches its adult size much earlier than the cerebellum, which accounts
for much of thr later growth in brain size (Tanner, 1978;). Therefore, the ohserved increase
in i>rain size over the juvenile years ma)’ be due primaril?. to non-cerebral ft.aturrs and,
thus, be unrelated to cortical sulcus lengths.
The sexual dimorphism in the time of growth cessation (t, ) reported here is, as far as MY’
can determine, a unique finding. We are unaware of other studies on relativel\. large
samples which have even considered sex differences in the de\!elopnlent of gross brdin
morphology in primates. Because the male intercept (cranial capacit) at birth J is
significantly higher than that for females, our data indicate that the development of sexual
dimorphism in brain size is in part a prenatal event. Lacking prenatal data \vc’ cannot
detc.rmine direc.tl>, whether the dimorphism at birth is due to different growth late5 or
diffcrent de\~elopmental timing. Our data indicate that post-natally males and females
undergo further brain growth at identical rates, but that this growth is maintained for ;L
longer period in males. Thus, the difference in adult male and female brain size would by,
due to both different prenatal growth pattcbrns and a prolonged developmental period irl
the male. Given the known effects of sex hormones on brain de\,elopment (Gesch\~ind 8r
Galaburda. 1987), these differences in brain de\.elopmclrt ljerween the sexes are not too

surprising. Furthermore, the existence of different male and female growth trajectories ilr baboons for body weight, length, and adiposity (Coelho, 1985) and in macaques Ibr J,od)
and non-neural organ weights (Larson, 1985) suggests that similar sesual dimorphism in growth patterns could exist for the rhesus brain.
Sexual dimorphism in sulcus lengths occurs for all but the orbital sulci, which art” not
different between the sexes. The observed sexual dimorphism for sulcus lengths can be entirely explained as a result ofoverall brain size differences between the sexes, but until we
obtain prenatal data, we cannot suggest whether this brain size difference at birth is due to different prenatal growth rates or different times of growth initiation. From a
developmental standpoint, there remains much to be learned about the primate brain.
Acknuwl~gments
This research is supported by Public Health Service grant 7 RO 1 NS24904. We thank the
University of Puerto Rico for free access to the Cayo Santiago skeletal collection. We also thank Allen Moore and Cashell Jacquish for their help in the analysis, and John Blangero and Sarah Williams-Blangero for their comments on earlier drafts of this paper.
References
Armstrong, E. (1985). Ailometric consideration% of the adult mammalian brain, with special emphasis on primates. In (W. I.. jungers, Ed.) Size u~~~ca~jn~ in Primate Biofog~, pp. i 15-146. Pfenum: New So&.
Armstrong, E. & Falk, D. (Eds) (1982). Pr&r& BY& ~ziol~t~o~: ;Wefhods end ~o~epts. Plenum: New York. Chappell, R. (1989). Fitting bent fines to data, with applications to allometry. J. thee. &of. L38, 235-256. Cheverud, J., Falk, D., Vannier, M., Konigsberg. L., Helmkamp, R. C, & Hildebolt, C:. (1989). Heritability ol
brain size and surface features in rhesus macaques (Xfacaca mulath). J Hered@ (in press) Chi, j., Dooling, E. & Gilles, F. (1977). Gyral development of the human brain. Ann. Newof. I, 86-93. Cock, A. (1966). Genetical aspects of metrtcal growth and form in animals. Q. Rev. Biol. 41, 131-190. Coelho, A. (1985). Baboon dimorphism: growth in weight, length and adiposity from birth to 8 years of age. In
(E. S. Watts, Ed.) Nonhuman Primate Modelsfor Human Growth and Development, pp. 125-159. Alan R. Liss: NW York.
Corruccini, R. (1987). Shape in morphometrics: comparative analyses. Ant. ./. p/y. Anfhrop. 73, 289-303.
Dobhing, J. & Sands, .J. (1973). Q uantitative growth and development of human hrain. Arch. Dis. Child. 48, 757-767.
Falk, D. (1978a). External Nemoanatomy ofOld World Monkeys (Cercopithecoidea). (;unfn’b. Primafol. vol. 1.5. S.
Karger: New York. F&k, D. (1978b). Brain evolution in Old World monkeys. Am. J. p.@. Anthrop. 48, 315-320. Falk, D. (1982). Mapping fossil endocasts. In (E. Armstrong & D. Falk, Eds) Prim& Brain ~~~lo/?ifj~~~ .tfcfhads aad
Conr~pfs, pp. 21 J-226. Plenum: New York. Falk, D., Cheverud, J., Vannier, M. 12’. & Conroy, G. C. (1986). Advanced c~)mputergraphics technology reveals
cortical asymmetry in endocasts of rhesus monkeys. Folia primat. 46, 98-103.
Falk, D., Konigsberg, L., Helmkamp, C., Cheverud, J., Vannier, M. & Hiideboh, C. (1989n). Endocranial suture closure in rhesus macaques (~%facaca mulatfa). Am.J. p/y.?. .4nthrop. (in press).
Falk, D., Hildeholt, C., Cheverud, J., Vannier, M., Helmkamp, C. & Konigshcrg, I,. (198%). Cortical asymmetries in frontal lobes of rhesus monkeys (,2~ncactr mulatln). Suhmitted for publication.
Geschwind. N. & Galaburda, A. (1987). Cerebral Laferalization. MIT Press: Gambridge. MA. Gould, S. (1975). Allometry in primates, with emphasis on scaling and the evolution ofthe brain. In (F. Szalav,
Ed.) Approaches to Primate Palrobiolog), pp. 244-292. Contrib. Primalof. vol. 5. S. Karger: Nev+ York. Hildebolt, C. & Vannier, M. (1988). Three-dimensional measurement accuracy of skull surface landmarks. :lm.
J. p/y. Anthrop. 76, 497-504. Holloway, R. (1968). The evolution of the primate brain: some aspects of quantitative relations. Bruin Res. 7,
t21-172. Holloway, R. (1978). The relevance ofendocasts for studying primate brain rvohrtion. in CC. Nohack. Ed.) .%RSO:)’
Sjstms ~~Pr~rnute~, pp. 181-200. Plenum: New York. Hoh, A., Cheek, D., Mellits, E. & Hill, D. (1975j. Brain size and the trfation ofthr primate to the nonprimatc. in
(D. B. Cheek, Ed.) Fetal and Postnatal Cellular Growth, pp. 23-44. John Wiley and Sons: New York. Huxley, J. (1932). Problems of Relative Growth. Dial Press: New York. Jerison, H. (1973). ~z~olu~ion qf the Rrnin and 1nteili~mw. ncademic Press: New Sork

EXT~~~AL~~A~~ FORM IN RHESUS MACAQUES 283
Jerison. H. (1982). Allometry, brain size, cortical surface, and convolutedness. In (E. Armstrong & I). Falk. Eds~ f’rimate Brain IGolution: .\fethods and Concepts, pp. 77-84. Plenum: New I.ork.
jolicoeur. P. (1985). A flexible 3-parameter curve for limited or unlimited somatic growth. Grouth 49, 785-794.
loticocur. P. & Pirlot, P. (1988). Asymptotic growth and complex allometry ofthe brain and body in tht* white rat.
hfedical Biophysics and Gomputing, University of Utah, Technical Rrport No. 14. I.arwn, S. (1985). Organ weight scaling in Primates. In (M’. L.Jungers. Ed.) .C’ ~ZP and .Scalin,~ in Prim& Hrnl~~pr. pi’.
YI-I 13. Plenum: New York. Lcbrau, R., Jolicorur. P., Pageau. G. & Grossman, E. (1986). Aqmptotic growth. qg production and tri\arialc.
allomrtry in I& ma.rquinong>~ Mitchill. Gomth 50, 18.5-200. Xlartin. R. (198’lj. Allometric approaches to the evolution oftht primate nwvous system. In C,E. Armstrong: & I).
F.&, Eds) Prinwtr ~r~~~ Euoiution: ,~~~t~~~s nnd Concepts_ pp. 3%-.X. Plenum: h‘et-t- York. ,\fa&n, R. & Harvq, P. (1985). Brain sizeaflometry: ontogeny and phylogen>-. In (bV. L.~jungrrs. Ed.) .Sii;utmci
.\infitt~g irz Prima/p Aiolo,~r, pp, 147-l 73. Plenum: NW York. Nohack, C:. (Ed.) (1970). The Primate Rrain. Plenum: New \‘ork. Passingham, R. ( 1973). Anatomical differences hetwwn thr nrocortrs of man and other primates. Rmin. Mar.
Gtmhridge Linivcrsity: N~M- York.
85 i-866, Radinsky. I,. ( 197.5). Primate brain evolution. rlnr. Sciezt. 63, 656-663. Sadc. I>.. Chepko-Sadc, D., Schneider, J., Roberts, S. & Richtsmeier, J. (1985). &sir Demo,pmphir 0hczrr~vztion.r on
k’rrr-Ranging Rhr.w .\fonkey. Human Relations .4rea Files: New Haven, CT. Sokal, R. & RohlF, F. (1981). Biometcv (2nd Ed.). W. H. Freeman: San Francisco. Tanner. j. ( 1978). I’oeiw into Man. Harvard University Press: C:amhridge, hL4. \Silkin&. I,. (14RRt. S’wtaf: Mr .6$~.rtem,fir Sf&siic.r. Systat. Inc.: Evanston. II..
fdenGJTc.ntion of sulci on endocasts
Appendix t
IVeIl-defined portions of 1 I sulci shown in Figure 1 were digitised on endocasts prepared from skulls of rhesus monkeys. Dots were placed on the endpoints of each sulcus and ~ipproximatel~ 2 mm apart along their intervening lengths by RCH. The dots were verified
hy DF for each sulcus on each endocast and any questionable sulci were discarded from the study. Thus, only portions of suici whose entire lengths were clearly and identical11
identified by two workers were digitised and analyzed. Below the criteria for defining rach sulcus or portion of sulcus are listed:
1. Rectus. This was one of the easier sulci to dot, i.e., its endpoints and configuration of the inte~~~renin~~ sulcal length were usually clearly represented on the endocasts. The entire sulctts was dotted and digitised.
2. :4rcuate. The medial end of the arcuatc sometimes converged with a transverse sulcus
located dorsomedially in the frontal lobe (i.e., somewhat parallel to the central sulcus). When this occurred, the medial end of the arcuate would appear to be arched caudally. However. the intersection of the two sulci was usually clear and we defined the medial endpoint of the arcuate as the point of intersection in these cases. The other end of the
arcuate and the intervening pathway were usually well-defined.
3. Central. The lateral end of the central sulcus was frequently obscured by a small vessel, and its medial end simply did not reproduce well in numerous cases. The intervening portion of-the sulcus was usually clear. and only central sulci whose endpoints were clearly visible were analyzed.
4. $Waiatr. This fissure was problematic at both ends. The rostra1 endpoint had to be arbitrarily d-Abed, since it is not clear even on real brains exactly where this fissure ends. 11’~ chose to define the anterior endpoint as the most rostrally located point along the

transitional curve from the lateral to basal surfat~e of the end~tcast. I’his point is lrtcart-d somewhat more medially than the traditional endpoint defined from a stl ictly lateral point
of view, i.e., the Sylvian as we define it cur\‘cs around a lit& underneath the ti,ontal loll. ‘The caudal end of’ the Sylvian usuall?- does not reproduce well on rhesus nlonke\- endocasts. We therefore dotted only those Sylvian fissures that mrr,~ed with superiot
temporal sulci, and considered the point of intersection as the caudal end of’both sulci.
5. rostrum srtperior ~e~~~ra~. The portion of the superior temporal sulcus rostra1 to rhe
sphenotemporal suture. Both endpoints and the Icr+$l of this short sulc~~s were cstrem~ly
clear on most endocasts.
6. Cuudal superior temporal. The rostra1 endpoint of this portion of the superior temporal
sulcus was the same as the caudal endpoint of No. 5 and was clearly defined on most of the casts. The caudal endpoint was considered to he the point where this sulcus merged with
the Sylvian fissure. Because the caudal endpoint of‘cts was difficult to see in other casts, cts was analyzed only in endocasts where Sjl and cls merged. The intervening portion of~ts was
usually quite clear.
7. Rostra1 middle temporal. Because the caudal portion of the middle temporal sulcus was extremely variable and hard to define in many endocasts, we chose to analyze onl?- the clearly defined portion rostra1 to the sphenotemporal suture. Both endpoints and the short
intervening length were extremely clear.
8. Orbitofioncal. This sulcus incises the orbitolateral border of rhesus endocasts, so it is visible in both views of Figure I. This sulcus was dotted and analyzed only if both
endpoints were clear and the sulcus was separate from the lateral orbital sulcus.
9. Medial orbital. This sulcus was problematic at both ends. Sometimes the caudal end was crossed by a suture that obscured its endpoint. The rostra1 end frequently tapered oB‘in such a way that an endpoint was not clearly defined. The intervening portion usually
reproduced clearly.
10. Fork of the rn~di~l orbital. This is one of the ciearest sulci on rhesus monkey endocasts. If
the orbital pattern was an H configuration, the fork was defined as the crossbar on the H. If the configuration was a k’ pattern, the fork was the lateral arm of the Y. Endpoints and the
short intervening lengths were always clear. The rostra1 endpoint of the arm of the Y configuration was also clear.
Il. Z,uter~~ orbital. This was a di&uIt sulcus to define because, unlike the fork, there was a
good deal of variation in its pattern. Sometimes, this sulcus extended onto the lateral surface of the frontal lobe. In these instances, the entire length was coded as the lateral
orbital sulcus (i.e., none of it was defined as the orbitofrontal sulcus). In the H orbital pattern, the lateral arm of the H was defined as the lateral orbital sulcus.
Other sulci which were digitised in the first round of repeatability nleasuren~ents but later excluded from the study because they could not be reliabiy defined on a sufficient sample of casts included: frontal superior complex (lateral to medial), frontal superior complex (medial to lateral), frontal superior complex proper, tail of the arcuate, width of
the temporal pole, precentral superior, subcentral anterior, postcentral superior, intraparietal, inferior occipital, lunate, parietal-occipital, lateral calcarine and branch of the lateral calcarine.
Related Documents











