Expression of neurotrophin-3 in the growing velvet antler of the red deer Cervus elaphus R L Garcia, M Sadighi 1 , S M Francis 1 , J M Suttie 1 and J S Fleming Department of Physiology and Centre for Gene Research, Otago School of Medical Sciences, PO Box 913, Dunedin, New Zealand 1 AgResearch Invermay Agricultural Centre, Private Bag 50034, Mosgiel, New Zealand (Requests for offprints should be addressed to R L Garcia who is now at Rammelkamp Center for Research, MetroHealth Campus, Case Western Reserve University School of Medicine, 2500 MetroHealth Drive, Cleveland, Ohio 44109-1998, USA) ABSTRACT Antlers are organs of bone which regenerate each year from the heads of male deer. In addition to bone, support tissues such as nerves also regenerate. Nerves must grow at up to 1 cm/day. The control of this rapid growth of nerves is unknown. We examined the relative expression of neurotrophin-3 (NT-3) mRNA in the different tissues of the growing antler tip and along the epidermal/ dermal layer of the antler shaft of the red deer Cervus elaphus, using semi-quantitative reverse transcription-polymerase chain reaction. Expres- sion in the tip was found to be highest in the epidermal/dermal layer and lowest in the cartilaginous layer in all developmental stages examined. These data correlate well with the density and pattern of innervation of these tissues. Along the epidermal/dermal layer of the antler shaft, expression was highest in the segments subjacent to the tip and lowest near the base, arguing for differences in the temporal expression of NT-3 in these segments. The expression of NT-3 in cells isolated from the different layers of 60-day antlers did not mirror that observed when whole tissues were used and may suggest regional specificity of NT-3 expression within antler tissues. Journal of Molecular Endocrinology (1997) 19, 173–182 INTRODUCTION Antlers are organs of bone which regenerate each year from the heads of male deer. During growth, which takes place at the tip at about 1 cm/day, support tissues such as nerves must grow at the same rate. The control mechanism for this rapid growth rate, and indeed the pattern of regeneration, are unknown. Antlers are normally supplied only with sensory nerves (Wislocki & Singer 1946) and lack sym- pathetic innervation (Wika 1980, Rayner & Ewen 1981, Gray et al. 1992). Both myelinated and unmyelinated fibres have been observed in the epidermis and in the vascularised dermal tissue (Vacek 1955). The supraoptic and temporal branches of the trigeminal nerve innervate antlers (Wislocki & Singer 1946). Innervation occurs by the regeneration of pedicle nerves (Waldo et al. 1949). Unlike other cellular components of the antler which develop in situ, blood vessels and nerves invade the forming tissue and are able to grow at the same rapid rate as the antler (Gray et al. 1992). Denervation experiments show that although nerves affect the size and shape of the antler, they are not essential to the actual control of growth and regeneration. Antlers can develop or regenerate, harden and cast without a nerve supply (Suttie & Fennessy 1985, Li et al. 1993). Apart from these observations, however, little is known about the role nerves play in antler development. Immunohistochemical studies on antler tissues by Gray and colleagues (1992) showed the presence of nerve fibres immunoreactive to antisera raised against substance P and calcitonin gene-related peptide (CGRP). Both neuropeptides are synthe- sised by sensory neurons and are known to play a role in nociception (Woolf & Wiensenfeld-Hallin 1986). These authors, citing the parallel roles that substance P and CGRP play in other systems, suggested that nerve fibres containing these peptides may have a significant role in rapid 173 Journal of Molecular Endocrinology (1997) 19, 173–182 ? 1997 Journal of Endocrinology Ltd Printed in Great Britain 0952–5041/97/019–173 $08.00/0

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Expression of neurotrophin-3 in the growing velvet antlerof the red deer Cervus elaphus
R L Garcia, M Sadighi1, S M Francis1, J M Suttie1 and J S FlemingDepartment of Physiology and Centre for Gene Research, Otago School of Medical Sciences,PO Box 913, Dunedin, New Zealand
1AgResearch Invermay Agricultural Centre, Private Bag 50034, Mosgiel, New Zealand
(Requests for offprints should be addressed to R L Garcia who is now at Rammelkamp Center forResearch, MetroHealth Campus, Case Western Reserve University School of Medicine,2500 MetroHealth Drive, Cleveland, Ohio 44109-1998, USA)
ABSTRACT
Antlers are organs of bone which regenerate eachyear from the heads of male deer. In addition tobone, support tissues such as nerves also regenerate.Nerves must grow at up to 1 cm/day. The control ofthis rapid growth of nerves is unknown. Weexamined the relative expression of neurotrophin-3(NT-3) mRNA in the different tissues of thegrowing antler tip and along the epidermal/dermal layer of the antler shaft of the red deerCervus elaphus, using semi-quantitative reversetranscription-polymerase chain reaction. Expres-sion in the tip was found to be highest inthe epidermal/dermal layer and lowest in thecartilaginous layer in all developmental stages
examined. These data correlate well with thedensity and pattern of innervation of these tissues.Along the epidermal/dermal layer of the antlershaft, expression was highest in the segmentssubjacent to the tip and lowest near the base,arguing for differences in the temporal expressionof NT-3 in these segments. The expression ofNT-3 in cells isolated from the different layersof 60-day antlers did not mirror that observedwhen whole tissues were used and may suggestregional specificity of NT-3 expression withinantler tissues.Journal of Molecular Endocrinology (1997) 19, 173–182
INTRODUCTION
Antlers are organs of bone which regenerate eachyear from the heads of male deer. During growth,which takes place at the tip at about 1 cm/day,support tissues such as nerves must grow at thesame rate. The control mechanism for this rapidgrowth rate, and indeed the pattern of regeneration,are unknown.Antlers are normally supplied only with sensory
nerves (Wislocki & Singer 1946) and lack sym-pathetic innervation (Wika 1980, Rayner & Ewen1981, Gray et al. 1992). Both myelinated andunmyelinated fibres have been observed in theepidermis and in the vascularised dermal tissue(Vacek 1955). The supraoptic and temporalbranches of the trigeminal nerve innervate antlers(Wislocki & Singer 1946). Innervation occurs by theregeneration of pedicle nerves (Waldo et al. 1949).Unlike other cellular components of the antlerwhich develop in situ, blood vessels and nerves
invade the forming tissue and are able to grow at thesame rapid rate as the antler (Gray et al. 1992).Denervation experiments show that although
nerves affect the size and shape of the antler, theyare not essential to the actual control of growth andregeneration. Antlers can develop or regenerate,harden and cast without a nerve supply (Suttie &Fennessy 1985, Li et al. 1993). Apart from theseobservations, however, little is known about the rolenerves play in antler development.Immunohistochemical studies on antler tissues by
Gray and colleagues (1992) showed the presenceof nerve fibres immunoreactive to antisera raisedagainst substance P and calcitonin gene-relatedpeptide (CGRP). Both neuropeptides are synthe-sised by sensory neurons and are known to play arole in nociception (Woolf & Wiensenfeld-Hallin1986). These authors, citing the parallel rolesthat substance P and CGRP play in othersystems, suggested that nerve fibres containingthese peptides may have a significant role in rapid
173
Journal of Molecular Endocrinology (1997) 19, 173–182 ? 1997 Journal of Endocrinology Ltd Printed in Great Britain0952–5041/97/019–173 $08.00/0

antler growth by controlling vascular tone andneovascularisation.The expression of these peptides is in turn
modulated by nerve growth factor (NGF) which ismainly elaborated by target tissues (Lindsay 1994).Neurotrophin-3 (NT-3), on the other hand, is neces-sary for the survival and differentiation of anotherimportant subpopulation of sensory neurons, thelarge, myelinated proprioceptive type (Lindsay1994).The cascade of events leading to the rapid growth
and innervation of the antler merits further in-vestigation. As part of a bigger project aimed atidentifying and characterising trophic factorspresent in this unique organ, we have examined therelative expression of the NT-3 gene in the differenttissues of the growing antler tip at various stages ofdevelopment and along the velvet layer of the antlershaft, using semi-quantitative reverse transcriptionand polymerase chain reaction (RT-PCR). Focuson NT-3 derives from the observation that deerdemonstrate an acute spatial awareness of wheretheir antler points are, suggesting the presence ofproprioceptive sensory neurons which are NT-3-dependent.
MATERIALS AND METHODS
Animals, tissues and cell lines
Antlers were obtained from 4- to 5-year-old reddeer stags (Cervus elaphus) under conditionsapproved by the Invermay Animal Ethics Com-mittee and essentially as described by Sadighiet al. (1994). Antlers were removed 30, 60 and90 days after the casting of the previous hard antler(n=3–5 per developmental stage). These time pointsrepresent 25, 50 and 75% of growth completion. As100% growth is hard dead bone, it was not anappropriate tissue to examine. The tip of the mainbeam was roughly dissected by eye into four layersrepresenting different zones of differentiation: theepidermal/dermal (the velvet), subdermal/reservemesenchymal, pre-cartilaginous and cartilaginouslayers (Fig. 1A). Samples of the epidermal/dermallayer from along the lateral side of the antler shaft(L1, L2, L4, L5, L6 and L8) were also obtainedfrom two 60-day antlers as shown in Fig. 1B. L3and L7 are tines which are not normally examinedin our studies as we wish to concentrate our effortson the rapidly growing shaft of the main beam.All tissues were snap-frozen in liquid nitrogenwithin 15 min of antler removal and stored at"80 )C. Antler cells were obtained from 60-daydermal, reserve mesenchymal, pre-cartilaginous
and cartilaginous layers. Reserve mesenchymal,pre-cartilaginous and cartilaginous layers weredispersed, cultured and maintained as described(Sadighi et al. 1994). The dermal tissue wasdispersed in medium consisting of Dulbecco’sMinimal Essential Medium (DMEM: SigmaChemical Co., St Louis, MO, USA) containing0·1% (w/v) BSA (Bio-Rad Laboratories Ltd,Auckland, NZ), 2000 U/ml collagenase (Sigma),1000 U/ml penicillin (Gibco-BRL, Gaithersburg,MD, USA) and 100 mg/ml streptomycin (Gibco-BRL) and incubated for 24 h at 36 )C under 5%(v/v) CO2. The dispersed cells were then centri-fuged at 500 g for 5 min and cultured in DMEMcontaining 10% foetal bovine serum (Gibco-BRL),50 mg/ml ascorbic acid, 100 U/ml penicillin and100 mg/ml streptomycin.
Molecular biology
RNA extractionTotal cellular RNA was extracted following themethod of Chomcyznski and Sacchi (1987). Enrich-ment of polyadenylated messenger (m)RNA (polyA+ RNA) was carried out as described by Fleminget al. (1992). All samples had a ratio of absorbanceat OD260:OD280 of 1·6–2·0. Aliquots were run ina 1% agarose gel under denaturing conditions(Fleming et al. 1992) to show that the RNA(s) wasintact and that equal amounts of RNA from eachtissue were used for generation of first-strandcDNA.
Oligodeoxyribonucleotide primersThe NT-3 forward primer (5*-ACG GAT GCCATG GTT ACT T-3*) was a 19-mer based on thedistal end of exon 1b of the human (nt 3480–3498;GenBank Accession No. S55222) and mouse(nt 3542–3560; GenBank Accession No. S75812)NT-3 sequences. The reverse primer (5*-ACCTGG TGT CCC CGA ATG T-3*) was anantisense 19-mer corresponding to nt 575–593(GenBank Accession No. M37763) and nt 683–701(GenBank Accession No. X53257) of the humanand mouse NT-3 sequences, respectively. Theâ-actin forward primer (5*-CCC TAA GGC CAACCG TGA AAA GAT G-3*) was a 25-mercorresponding to nt 1661–1685 of the rat â-actinsequence and the reverse primer (5*-GTC CCGGCC AGC CAG GTC CAG-3*) was an antisense21-mer corresponding to nt 2324–2344 of the ratâ-actin sequence (GenBank Accession No. J00691).Both primer pairs cross a large intron thus rulingout contribution arising from amplification ofgenomic DNA.
and others · Neurotrophin-3 expression in velvet antler174
Journal of Molecular Endocrinology (1997) 19, 173–182

Reverse transcription-polymerase chain reactionReverse transcription was carried out as follows.Total or poly A+RNA (1 µg) and 25 m oligo-dTprimer (Boehringer Mannheim GmbH, Germany),in a total volume of 9 µl, was heated to 70 )C for10 min, to disrupt possible secondary structures.The mixture was then pre-incubated at 37 )C for2 min in 1#buffer (50 m Tris–HCl (pH 8·3),75 m KCl, 3 m MgCl2), 10 m dithiothreitoland 125 m of each dNTP (dATP, dCTP, dGTP,dTTP). Superscript II reverse transcriptase (1 µl;200 U/µl; Gibco-BRL) was added to a final reactionvolume of 20 µl and the mixture was incubated at37 )C for 60 min. The reaction was stopped by a5 min incubation at 94 )C, then snap-cooled on ice.First-strand cDNA (1 µl) was used for subsequentPCR reactions. A PCR cocktail (all componentsfrom GIBCO-BRL) was prepared containing 200 µof each dNTP, 1·5 m MgCl2, 40 pmol of each PCRprimer and 1·0 U Taq polymerase in a volume
of 19 µl. First-strand cDNA (1 µl) was added andthe mixture subjected to PCR amplification for 26(NT-3) or 16 (â-actin) cycles in an FTS 4000capillary thermocycler (Corbett Research, Sydney,Australia). Both reactions were within their pre-determined exponential phase of amplification. Thethermocycler was programmed to give an initial cycleconsisting of 94 )C denaturation for 3 min, 60 )Cannealing for 15 s and 72 )C extension for 40 s, fol-lowed by 25 (NT-3) or 15 (â-actin) cycles of 94 )C for10 s, 60 )C for 10 s and 72 )C for 40 s. A no templatecontrol and an RT (") control were performedalongside all experimental samples. In all manipu-lations, filtered pipette tips (Labcon, San Rafael,CA, USA) were used to prevent tissue-to-tissue andaerosol contaminations.
Analysis and relative quantitation of PCR productsâ-Actin, a ubiquitously expressed gene, was usedas an endogenous internal standard to determine
1. (A) Longitudinal section of the antler tip at 30, 60 and 90 days ofgrowth, showing the different zones of differentiation. Arrow head=epidermis/dermis, s=subdermis/reserve mesenchyme, p=pre-cartilage andc=cartilage. (B) Schematic diagram of a 60-day antler taken from a4-year-old stag, showing the regions from which samples of the velvet layerwere taken for comparison of NT-3 expression along the shaft. The majornerve supply is expected to enter the antler/pedicle from the outer lateral (L)side.
Neurotrophin-3 expression in velvet antler · and others 175
Journal of Molecular Endocrinology (1997) 19, 173–182

the relative expression of NT-3 in different antlertissues. NT-3 and â-actin sequences from the sameRT reactions were amplified in two separate PCRreactions to avoid interference, because â-actinmRNA was much more abundant than NT-3 mRNA. Pilot experiments were performed todetermine the exponential phase of amplification forboth NT-3 and â-actin in different tissues. ForNT-3, this was found to be between cycles 22–28,and for â-actin between cycles 12–18. The expectedPCR product sizes were approximately 570 and220 bp for NT-3 and â-actin respectively. FollowingPCR amplification, the entire 20 µl reaction waselectrophoresed through a 1·8% agarose gel (GIBCO-BRL), then blotted onto Hybond N nylon membrane(Amersham International, Amersham, UK) over-night, using standard procedures (Sambrooket al. 1989). Southern blots were pre-hybridisedfor 2–4 h and hybridised overnight at 42 )C inhybridisation buffer (Fleming et al. 1992), with arat NT-3 probe (Regeneron Pharmaceuticals, Inc.,Tarrytown, NY, USA) labelled by random prim-ing (Ready-to-Go, Pharmacia Biotech, Uppsala,Sweden). Blots were washed in 0·2#SSC (1#SSC is 15 m sodium citrate plus 150 m sodiumchloride) at 42 )C and exposed to X-ray film (KodakXAR-5) for 2–20 h at "80 )C. The densities ofthe bands on both the autoradiographs and thephotographs of ethidium bromide-stained gels weredetermined densitometrically using a Bio-RadGS-700 imaging densitometer. The NT-3:â-actindensity ratio of each sample was determined andcompared. The data presented here are from threeseparate RNA extracts from each tissue, obtainedfrom three different antlers.
DNA sequencingThe NT-3 fragment amplified from deer antlerwas sequenced using the direct cycle sequencingmethod at the sequencing facility of the Universityof Otago Centre for Gene Research. Sequencecomparisons and analyses were carried out using theGCG package of sequence manipulation programs(Devereux et al. 1984) and the MSA, TRANS-LATE and BLAST (Altschul et al. 1990) programsavailable through the internet.
RESULTS
NT-3 mRNA expression in antler tip tissues
An amplified fragment of the expected size(2570 bp) was obtained from all tissues in all threedevelopmental stages examined, albeit in varyingamounts. As shown in Fig. 2, NT-3 expression was
higher in the epidermal/dermal layer of 30- and60-day antlers. The amounts of NT-3 mRNA inthe subdermal/reserve mesenchymal and pre-cartilaginous layers in all three developmental stageswere intermediate between those of the epidermal/dermal and cartilaginous layers (see discussion forcomments). The 60-day antler epidermal/dermallayer exhibited the highest amount of NT-3 mRNAand the 90-day antler cartilaginous layer the lowest.The higher levels of NT-3 PCR amplification
in epidermal/dermal layers could not be attributedto more cDNA having been used in the reaction,because the â-actin primers amplified a 220 bpfragment in all other antler tissues to levels equal toor even exceeding those in epidermal/dermal tissues(Fig. 2).
NT-3 mRNA expression in cultured antlercells
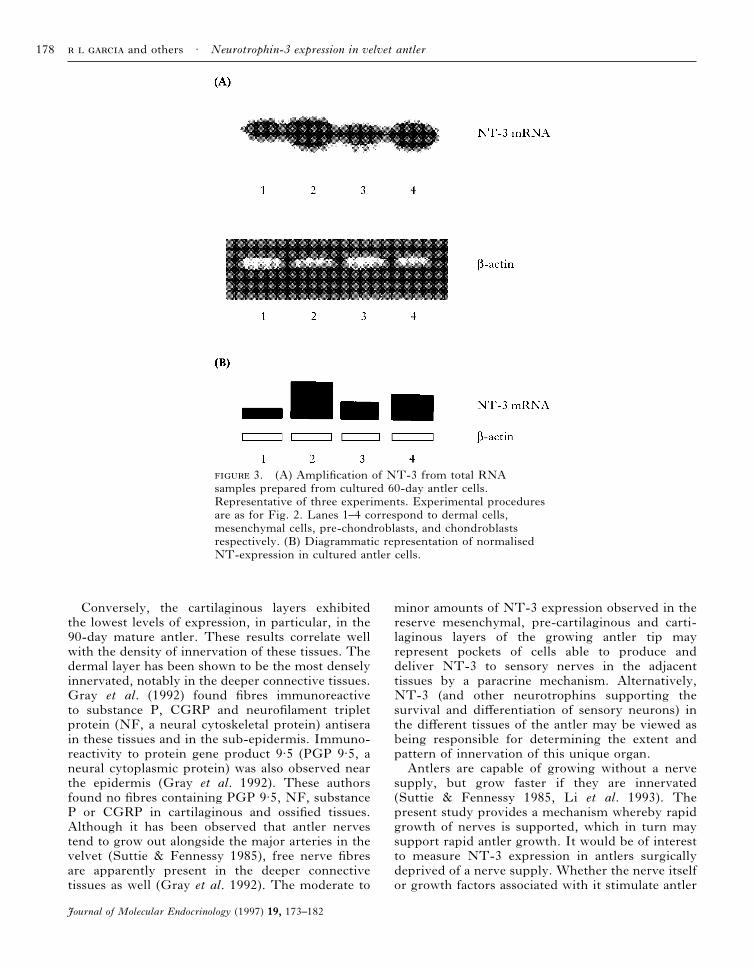
The expression of NT-3 mRNA in cells isolatedfrom the dermal, reserve mesenchymal, pre-cartilaginous and cartilaginous layers of 60-dayantlers was also investigated. A pattern of expres-sion was found which was different from thatobserved when whole tissue extracts were studied.Cells from the reserve mesenchymal and carti-laginous layers showed higher levels of expres-sion than those from the epidermal/dermal andpre-cartilaginous layers (Fig. 3).
NT-3 expression in the velvet layer of theantler shaft
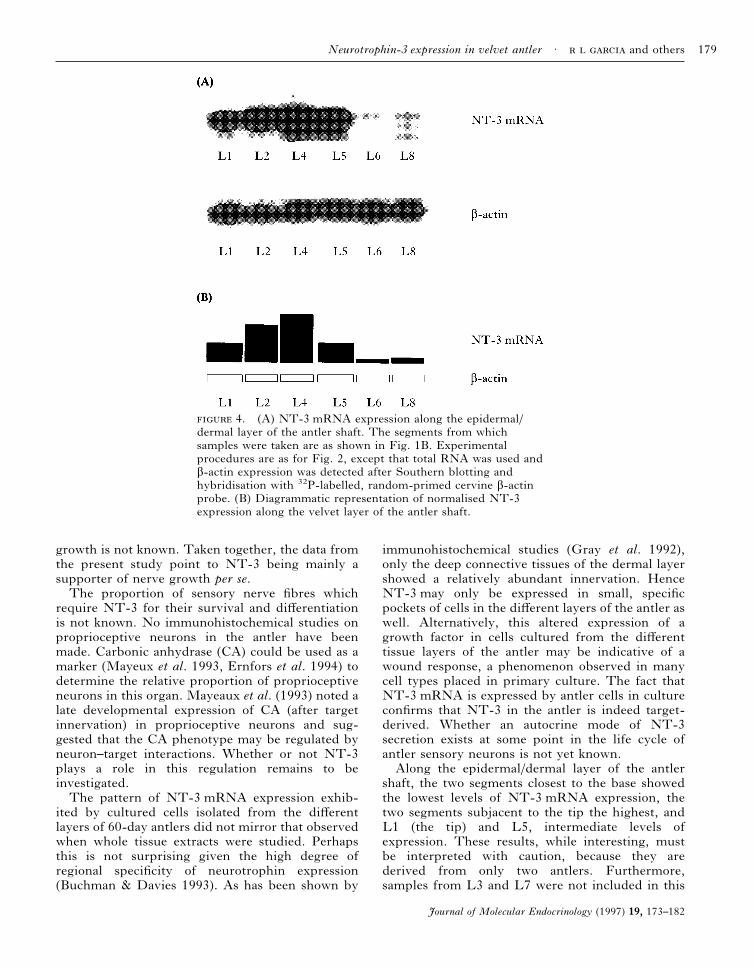
NT-3 mRNA expression was strongest in L4 andL2 (Fig. 4), the two segments subjacent to the antlertip (L1; refer to Fig. 1B). The segments closest tothe antler pedicle (L6 and L8) showed much lowerNT-3 expression compared with L4 and L2.â-Actin expression was fairly uniform in those partsof the main beam studied.
The deer NT-3 gene sequence
The PCR-amplified NT-3 fragment from antlercDNA was sequenced and revealed 82% amino acididentity with both human and mouse NT-3sequences (88% and 90% sequence similarity,respectively; Fig. 5). The cervine sequence is also atleast five amino acids shorter than either the humanor mouse sequence. The sequence amplified by theprimers used in this study included the canonicalcleavage site (RRKR, underlined in Fig. 5) for themature protein (Maisonpierre et al. 1990).
and others · Neurotrophin-3 expression in velvet antler176
Journal of Molecular Endocrinology (1997) 19, 173–182

DISCUSSION
We have employed RT-PCR coupled withSouthern blot analyses to examine tissue-specificdifferences in the expression of the NT-3 gene inthe growing antler. The sensitivity of the assayallowed the detection of minor expression in sometissues which may otherwise be missed by conven-tional methods such as Northern blot analyses.NT-3 mRNA expression was shown to be higher inthe epidermal/dermal (the velvet) layer of thegrowing antler tip, with the highest level at 60 days,the period of most rapid growth. Although the
90-day antler reserve mesenchymal layer appearedto show a higher level of expression than the 90-dayantler epidermal/dermal layer, it may have been theresult of the relatively crude dissection proceduresemployed in the interests of preparing the tissuesfor mRNA extraction as fast as possible. The tip of90-day antler is highly calcified and the layersare particularly difficult to separate. Moreover,because growth has almost stopped, there is little,if any, reserve mesenchymal tissue remaining inthe antler tip. What was designated as reservemesenchyme may therefore have been part of thedermal layer.
2. Expression of NT-3 mRNA in antler tissues.Representative of three experiments. (A) A 570 bp NT-3 fragmentwas amplified from antler tissue poly A+ RNA(s) via RT-PCR.PCR products were Southern blotted and hybridised with a32P-labelled rat NT-3 random-primed probe. A 220 bp â-actinfragment was amplified from the same RT reaction to serve as anendogenous internal control. The intensities of autoradiographicsignals and of bands in ethidium bromide-stained gels werequantified by densitometry and the ratio of NT-3 to â-actinexpression was calculated for each tissue. A diagrammaticrepresentation of normalised NT-3 expression in deer antler tissuesis shown in (B). Lanes 1–12 correspond to 30-day epidermis/dermis,30-day subdermis/reserve mesenchyme, 30-day pre-cartilage, 30-daycartilage, 60-day epidermis/dermis, 60-day subdermis/reservemesenchyme, 60-day pre-cartilage, 60-day cartilage, 90-dayepidermis/dermis, 90-day subdermis/reserve mesenchyme, 90-daypre-cartilage and 90-day cartilage respectively.
Neurotrophin-3 expression in velvet antler · and others 177
Journal of Molecular Endocrinology (1997) 19, 173–182
Conversely, the cartilaginous layers exhibitedthe lowest levels of expression, in particular, in the90-day mature antler. These results correlate wellwith the density of innervation of these tissues. Thedermal layer has been shown to be the most denselyinnervated, notably in the deeper connective tissues.Gray et al. (1992) found fibres immunoreactiveto substance P, CGRP and neurofilament tripletprotein (NF, a neural cytoskeletal protein) antiserain these tissues and in the sub-epidermis. Immuno-reactivity to protein gene product 9·5 (PGP 9·5, aneural cytoplasmic protein) was also observed nearthe epidermis (Gray et al. 1992). These authorsfound no fibres containing PGP 9·5, NF, substanceP or CGRP in cartilaginous and ossified tissues.Although it has been observed that antler nervestend to grow out alongside the major arteries in thevelvet (Suttie & Fennessy 1985), free nerve fibresare apparently present in the deeper connectivetissues as well (Gray et al. 1992). The moderate to
minor amounts of NT-3 expression observed in thereserve mesenchymal, pre-cartilaginous and carti-laginous layers of the growing antler tip mayrepresent pockets of cells able to produce anddeliver NT-3 to sensory nerves in the adjacenttissues by a paracrine mechanism. Alternatively,NT-3 (and other neurotrophins supporting thesurvival and differentiation of sensory neurons) inthe different tissues of the antler may be viewed asbeing responsible for determining the extent andpattern of innervation of this unique organ.Antlers are capable of growing without a nerve
supply, but grow faster if they are innervated(Suttie & Fennessy 1985, Li et al. 1993). Thepresent study provides a mechanism whereby rapidgrowth of nerves is supported, which in turn maysupport rapid antler growth. It would be of interestto measure NT-3 expression in antlers surgicallydeprived of a nerve supply. Whether the nerve itselfor growth factors associated with it stimulate antler
3. (A) Amplification of NT-3 from total RNAsamples prepared from cultured 60-day antler cells.Representative of three experiments. Experimental proceduresare as for Fig. 2. Lanes 1–4 correspond to dermal cells,mesenchymal cells, pre-chondroblasts, and chondroblastsrespectively. (B) Diagrammatic representation of normalisedNT-expression in cultured antler cells.
and others · Neurotrophin-3 expression in velvet antler178
Journal of Molecular Endocrinology (1997) 19, 173–182
growth is not known. Taken together, the data fromthe present study point to NT-3 being mainly asupporter of nerve growth per se.The proportion of sensory nerve fibres which
require NT-3 for their survival and differentiationis not known. No immunohistochemical studies onproprioceptive neurons in the antler have beenmade. Carbonic anhydrase (CA) could be used as amarker (Mayeux et al. 1993, Ernfors et al. 1994) todetermine the relative proportion of proprioceptiveneurons in this organ. Mayeaux et al. (1993) noted alate developmental expression of CA (after targetinnervation) in proprioceptive neurons and sug-gested that the CA phenotype may be regulated byneuron–target interactions. Whether or not NT-3plays a role in this regulation remains to beinvestigated.The pattern of NT-3 mRNA expression exhib-
ited by cultured cells isolated from the differentlayers of 60-day antlers did not mirror that observedwhen whole tissue extracts were studied. Perhapsthis is not surprising given the high degree ofregional specificity of neurotrophin expression(Buchman & Davies 1993). As has been shown by
immunohistochemical studies (Gray et al. 1992),only the deep connective tissues of the dermal layershowed a relatively abundant innervation. HenceNT-3 may only be expressed in small, specificpockets of cells in the different layers of the antler aswell. Alternatively, this altered expression of agrowth factor in cells cultured from the differenttissue layers of the antler may be indicative of awound response, a phenomenon observed in manycell types placed in primary culture. The fact thatNT-3 mRNA is expressed by antler cells in cultureconfirms that NT-3 in the antler is indeed target-derived. Whether an autocrine mode of NT-3secretion exists at some point in the life cycle ofantler sensory neurons is not yet known.Along the epidermal/dermal layer of the antler
shaft, the two segments closest to the base showedthe lowest levels of NT-3 mRNA expression, thetwo segments subjacent to the tip the highest, andL1 (the tip) and L5, intermediate levels ofexpression. These results, while interesting, mustbe interpreted with caution, because they arederived from only two antlers. Furthermore,samples from L3 and L7 were not included in this
4. (A) NT-3 mRNA expression along the epidermal/dermal layer of the antler shaft. The segments from whichsamples were taken are as shown in Fig. 1B. Experimentalprocedures are as for Fig. 2, except that total RNA was used andâ-actin expression was detected after Southern blotting andhybridisation with 32P-labelled, random-primed cervine â-actinprobe. (B) Diagrammatic representation of normalised NT-3expression along the velvet layer of the antler shaft.
Neurotrophin-3 expression in velvet antler · and others 179
Journal of Molecular Endocrinology (1997) 19, 173–182

study. The proximal-to-distal growth pattern of theantler probably provides the best clue as to whatthese differences in NT-3 expression along the shaftimply. If NT-3 was maximally expressed at the tipand the expression progressively decreased towardsthe base, this would favour a ‘nerve puller’hypothesis. That is, it would imply the presence ofan NT-3 gradient guiding neurons towards their
target. However, the two segments subjacent tothe tip exhibited higher levels of NT-3 mRNAexpression than the tip itself. While classical ex-periments (Levi-Montalcini 1975, Gundersen &Barett 1980) showed elegantly that NGF is able toattract sympathetic and sensory neurons, morerecent evidence indicates that the onset of bothneurotrophin and neurotrophin receptor synthesis
5. (A) Nucleotide and derived amino acid sequences of NT-3 cDNA obtained by RT-PCR. The sequenceshave been deposited in the GenBank/EMBL database (Accession No. U66715). (B) Alignment of deer NT-3 derivedamino acid sequence with those of human and mouse. Dots have been introduced for alignment and do not representgaps in sequence. The canonical cleavage site for the mature protein (Maisonpierre et al. 1990) is underlined.
and others · Neurotrophin-3 expression in velvet antler180
Journal of Molecular Endocrinology (1997) 19, 173–182

coincides with the start of target field innervation(Buchman & Davies 1993). Synthesis of neuro-trophins by target tissues starts only at the arrival ofthe earliest nerve fibres. Expression of neurotrophinreceptors in nerve terminals, on the other hand, isnot triggered by target encounter (Buchman &Davies 1993). Taken together, these results sug-gest that during development neurotrophins donot attract nerve fibres towards their target field,but are only involved in their survival anddifferentiation.An alternative explanation is possible if the
different segments of the antler are seen asrepresenting different time points in development.The lower level of NT-3 mRNA expression nearthe base (L6 and L8, and probably even L5) mayrepresent neurons past their period of neurotrophindependence. The higher levels of expression in thetwo segments subjacent to the tip may represent theperiod of maximal NT-3-responsiveness, the periodafter the initial target encounter, until just beforethe onset of naturally occurring cell death. Theexpression in the tip may represent the period whenaxons are still growing towards their target up to theinitial target encounter. Buchman and Davies (1993)observed this developmental sequence of events intrigeminal ganglion neurons. These neurons surviveindependently of neurotrophins when their axonsare still growing towards their target. This isfollowed by a period of responsiveness to brain-derived neurotrophic factor (BDNF) and NT-3,then later to NGF. The neurons later lose theirresponsiveness first to BDNF then to NT-3, but theNGF survival response is maintained through tothe naturally occurring cell death phase (Buchman& Davies 1993). Whether it is valid to cite thissequence of events to explain differential neuro-trophin expression along the antler shaft remains tobe seen. A detailed study of antler innervation at thecellular level and at closely staged intervals indevelopment is warranted.To verify its identity, the PCR fragment
amplified using NT-3 primers was sequenced andrevealed strong sequence conservation with othermammalian NT-3 genes. It is expected that theremainder of the coding region will be very similarto those of other mammalian NT-3 sequences,although the regulatory region of the deer NT-3 gene may well be organised differently. Genomicsequencing may reveal unique sequences respon-sible for the temporal and tissue-specific expressionof NT-3 in the antler.In summary, we have shown that NT-3 expres-
sion in tissues of the rapidly growing antler tipcorrelates well with the density and pattern ofinnervation of this organ. Along the velvet layer
of the antler shaft, NT-3 expression was shown tobe highest in the segments subjacent to the tip andlowest near the base, arguing for differences in thetemporal expression of NT-3 in these segments.Expression of NT-3 by antler cells grown in culturesubstantiates the general idea that neurotrophins aretarget-derived. In situ hybridisation studies arerequired to demonstrate the cellular distributionwithin antler tissues of NT-3 mRNA.
REFERENCES
Altschul SF, Gish W, Miller W, Myers EW & Lipman DJ1990 Basic local alignment search tool. Journal of MolecularBiology 215 403–410.
Buchman VL & Davies AM 1993 Different neurotrophins areexpressed and act in a developmental sequence to promotethe survival of embryonic sensory neurons. Development 118989–1001.
Chomcyznski P & Sacchi N 1987 Single-step method of RNAisolation by guanidinium thiocyanate-phenol-chloroformextraction. Analytical Biochemistry 162 156–159.
Devereux J, Haeberli P & Smithies O 1984 A comprehensiveset of sequence analysis programs for the VAX. Nucleic AcidsResearch 12 387–395.
Ernfors P, Lee KF, Kucera J & Jaenisch R 1994 Lack ofneurotrophin-3 leads to deficiencies in the peripheral nervoussystem and loss of limb proprioceptive afferents. Cell 77503–512.
Fleming JS, Greenwood PJ & Chen CLC 1992 Expression ofthe clusterin gene in the tissues of Booroola sheep whichwere homozygous or non-carriers of the fecundity geneFecB. Journal of Molecular Endocrinology 9 207–211.
Gray C, Hukkanen M, Kontinnen YT, Terenghi G, ArnettTR, Jones SJ, Burnstock G & Polak JM 1992 Rapidneural growth: calcitonin gene-related peptide andsubstance P-containing nerves attain exceptional growthrates in regenerating deer antler. Neuroscience 50953–963.
Gundersen RW & Barett JN 1980 Characterisation of theturning response of dorsal root neurites toward nerve growthfactor. Journal of Cell Biology 87 546–554.
Levi-Montalcini R 1975 NGF: An uncharted route. In TheNeurosciences: Paths of Discovery, pp 245–265. Eds FGWorden, JP Swazey & G Adelman. Cambridge, MA: MITPress.
Li C, Sheard PW, Corson ID & Suttie JM 1993 Pedicle andantler development following sectioning of the sensorynerves to the antlerogenic region of red deer (Cervuselaphus). Journal of Experimental Zoology 267 188–197.
Lindsay RM 1994 Neurotrophins and receptors. Progress inBrain Research 103 3–14.
Maisonpierre PC, Belluscio L, Squinto S, Ip NY, Furth ME,Lindsay RM & Yancopoulos GD 1990 Neurotrophin 3: aneurotrophic factor related to NGF and BDNF. Science 2471446–1451.
Mayeux V, Tafti M, Baldy-Moulinier M & Valmier J 1993Developmental regulation of carbonic anhydrase expressionin mouse dorsal root ganglia. Developmental Brain Research71 201–208.
Rayner V & Ewen SWB 1981 Do the blood vessels of theantler velvet of the red deer have an adrenergicinnervation? Quarterly Review of Experimental Physiology66 81–90.
Neurotrophin-3 expression in velvet antler · and others 181
Journal of Molecular Endocrinology (1997) 19, 173–182

Sadighi M, Haines SR, Skottner A, Harris AJ & Suttie JM1994 Effects of insulin-like growth factor-I (IGF-I) andIGF-II on the growth of antler cells in vitro. Journal ofEndocrinology 143 461–469.
Sambrook J, Fritsch EF & Maniatis T 1989 Molecular Cloning:A Laboratory Manual. Cold Spring Harbor: Cold SpringHarbor Laboratory Press.
Suttie JM & Fennessy PF 1985 Regrowth of amputated antlerswith and without innervation. Journal of ExperimentalZoology 234 359–366.
Vacek Z 1955 Innervace lyci rostoucia parohu u cervidu. CslkaMorf 3 249–264.
Waldo CM, Wislocki GB & Fawcett DW 1949 Observations onthe blood supply of growing antlers. American Journal ofAnatomy 84 27–61.
Wika M 1980 On growth of reindeer antlers. In Proceedingsof the 2nd International Reindeer/Caribou Symposium,pp 416–421. Eds E Reimers, E Garre & S Skjenneberg.Trondheim, Norway: Direktoratet for vilt og ferskvannsfisk,Trondheim.
Wislocki GB & Singer M 1946 The occurrence and function ofnerves in the growing antlers of deer. Journal of ComparativeNeurology 85 1–19.
Woolf C & Wiensenfeld-Hallin Z 1986 Substance P andcalcitonin gene-related peptide synergistically modulate thegain of the nociceptive withdrawal reflex in the rat.Neuroscience Letters 66 226–230.
17 December 1996
and others · Neurotrophin-3 expression in velvet antler182
Journal of Molecular Endocrinology (1997) 19, 173–182
Related Documents











