Systematic and Applied Microbiology 35 (2012) 485–495 Contents lists available at SciVerse ScienceDirect Systematic and Applied Microbiology jo u rn al homepage: www.elsevier.de/syapm Exploring the in situ accessibility of small subunit ribosomal RNA of members of the domains Bacteria and Eukarya to oligonucleotide probes Hatice E. Okten a , L. Safak Yilmaz b,∗ , Daniel R. Noguera c a Bahcesehir University, Engineering Faculty, Department of Environmental Engineering, 34349 Besiktas, Istanbul, Turkey b Department of Biochemistry and Molecular Pharmacology, University of Massachusetts Medical School, 364 Plantation Street, Worcester, MA 01605-2324, USA c Department of Civil and Environmental Engineering, University of Wisconsin - Madison, 1415 Engineering Drive, Madison, Wisconsin 53706-1691, USA a r t i c l e i n f o Article history: Received 22 June 2011 Received in revised form 4 November 2011 Accepted 9 November 2011 Keywords: Fluorescence in situ hybridization (FISH) Ssu rRNA Accessibility Thermodynamic affinity (G overall ◦ ) Nucleobase-specific quenching Dual-labeled probes a b s t r a c t The principle that the small subunit ribosomal RNA (ssu rRNA) is generally accessible to oligonu- cleotide probes designed to have high thermodynamic affinity was tested with Stenotrophomonas maltophilia, Rhodobacter sphaeroides, Bacillus subtilis, and Saccharomyces cerevisiae. Fluorescein-labeled probes, designed to have G overall ◦ = −14 ± 1 and to avoid the potential of nucleobase-specific quench- ing, were used to target 20 randomly selected sites in each organism. A site was considered accessible if probe brightness was at least 10 times the background signal. With 30-h hybridizations, 71 out of 80 target sites passed the accessibility criterion. Three additional sites were demonstrated to be accessible with either longer hybridizations, which seemed to have a negative effect on some probes, or the addition of formamide to the hybridization buffer. The remaining 6 sites were demonstrated to be accessible by changing the fluorophore to Cy5, slightly modifying probe lengths, using dual-labeled fluorescein probes, or a combination of these approaches. Probe elongations were only needed in 4 probes, indicating a 95% success in correctly predicting G overall ◦ , the key parameter for the design of high affinity probes. In addi- tion, 94% of the fluorescein labeled probes yielded bright signals, demonstrating that nucleobase-specific quenching of fluorescein is an important factor affecting probe brightness that can be predicted during probe design. Overall, the results support the principle that with a rational design of probes, it is possible to make most target sites in the ssu rRNA accessible. © 2011 Elsevier GmbH. All rights reserved. Introduction Fluorescence in situ hybridization (FISH) is a rapid cultiva- tion independent method for detecting and quantifying specific microorganisms within mixed microbial communities [1–3,24]. In most FISH applications, ribosomal RNA (rRNA) molecules are pre- ferred targets because of their abundance inside cells. In particular, the small subunit rRNA (ssu rRNA) is widely used as a phylogenetic marker [13]. The ssu rRNA sequence has conserved and variable regions [27], a characteristic that is used for defining target sites with the desired level of specificity, which could be from species to domain level [3]. Central to the development of useful FISH applications is the ability to produce hybridizations that maximize sensitivity (i.e., high signal-to-noise ratio) while maintaining specificity towards the targeted organisms. Sensitivity was initially thought to be a function of the location of the target site on the ssu rRNA molecule ∗ Corresponding author. Tel.: +1 508 856 2636; fax: +1 508 856 6231. E-mail addresses: [email protected] (L.S. Yilmaz), [email protected] (D.R. Noguera). [4,14] because of possible interference from higher order structures in the ribosome. Following this concept, Fuchs et al. [15] carried out a systematic study to evaluate the effect of target site location by performing hybridizations over the entire 16S rRNA molecule of Escherichia coli. Their results were interpreted as demonstrating the existence of inherently inaccessible regions on the 16S rRNA of E. coli. On a follow-up publication, Behrens et al. [9] used a sim- ilar approach to generate accessibility maps for three additional microorganisms, with the purpose of evaluating whether the loca- tion of inaccessible regions found in E. coli could be extended to other organisms. They produced a consensus map from the combi- nation of the accessibilities observed with the different organisms. Another publication from the same group of researchers inves- tigated the effect of RNA-RNA and RNA-protein interactions on hybridization efficiency [7]. However a correlation between flu- orescence signal intensity and ribosomal structure could not be found. Searching for alternative mechanisms to understand rRNA accessibility to DNA probes, Yilmaz and Noguera [28] demonstrated that the thermodynamic affinity of the probe to the target site, mea- sured as the overall free energy (G overall ◦ ) of a reaction including probe-target hybridization as well as probe and target self folding, 0723-2020/$ – see front matter © 2011 Elsevier GmbH. All rights reserved. doi:10.1016/j.syapm.2011.11.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Et
Ha
b
c
a
ARRA
KFSATND
I
tmmftmrwd
ahtf
n
0d
Systematic and Applied Microbiology 35 (2012) 485– 495
Contents lists available at SciVerse ScienceDirect
Systematic and Applied Microbiology
jo u rn al homepage: www.elsev ier .de /syapm
xploring the in situ accessibility of small subunit ribosomal RNA of members ofhe domains Bacteria and Eukarya to oligonucleotide probes
atice E. Oktena, L. Safak Yilmazb,∗, Daniel R. Noguerac
Bahcesehir University, Engineering Faculty, Department of Environmental Engineering, 34349 Besiktas, Istanbul, TurkeyDepartment of Biochemistry and Molecular Pharmacology, University of Massachusetts Medical School, 364 Plantation Street, Worcester, MA 01605-2324, USADepartment of Civil and Environmental Engineering, University of Wisconsin - Madison, 1415 Engineering Drive, Madison, Wisconsin 53706-1691, USA
r t i c l e i n f o
rticle history:eceived 22 June 2011eceived in revised form 4 November 2011ccepted 9 November 2011
eywords:luorescence in situ hybridization (FISH)su rRNAccessibilityhermodynamic affinity (�Goverall
◦)ucleobase-specific quenching
a b s t r a c t
The principle that the small subunit ribosomal RNA (ssu rRNA) is generally accessible to oligonu-cleotide probes designed to have high thermodynamic affinity was tested with Stenotrophomonasmaltophilia, Rhodobacter sphaeroides, Bacillus subtilis, and Saccharomyces cerevisiae. Fluorescein-labeledprobes, designed to have �Goverall
◦ = −14 ± 1 and to avoid the potential of nucleobase-specific quench-ing, were used to target 20 randomly selected sites in each organism. A site was considered accessibleif probe brightness was at least 10 times the background signal. With 30-h hybridizations, 71 out of 80target sites passed the accessibility criterion. Three additional sites were demonstrated to be accessiblewith either longer hybridizations, which seemed to have a negative effect on some probes, or the additionof formamide to the hybridization buffer. The remaining 6 sites were demonstrated to be accessible bychanging the fluorophore to Cy5, slightly modifying probe lengths, using dual-labeled fluorescein probes,
ual-labeled probes or a combination of these approaches. Probe elongations were only needed in 4 probes, indicating a 95%success in correctly predicting �Goverall
◦, the key parameter for the design of high affinity probes. In addi-tion, 94% of the fluorescein labeled probes yielded bright signals, demonstrating that nucleobase-specificquenching of fluorescein is an important factor affecting probe brightness that can be predicted duringprobe design. Overall, the results support the principle that with a rational design of probes, it is possibleto make most target sites in the ssu rRNA accessible.
ntroduction
Fluorescence in situ hybridization (FISH) is a rapid cultiva-ion independent method for detecting and quantifying specific
icroorganisms within mixed microbial communities [1–3,24]. Inost FISH applications, ribosomal RNA (rRNA) molecules are pre-
erred targets because of their abundance inside cells. In particular,he small subunit rRNA (ssu rRNA) is widely used as a phylogenetic
arker [13]. The ssu rRNA sequence has conserved and variableegions [27], a characteristic that is used for defining target sitesith the desired level of specificity, which could be from species toomain level [3].
Central to the development of useful FISH applications is thebility to produce hybridizations that maximize sensitivity (i.e.,
igh signal-to-noise ratio) while maintaining specificity towardshe targeted organisms. Sensitivity was initially thought to be aunction of the location of the target site on the ssu rRNA molecule∗ Corresponding author. Tel.: +1 508 856 2636; fax: +1 508 856 6231.E-mail addresses: [email protected] (L.S. Yilmaz),
[email protected] (D.R. Noguera).
723-2020/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.syapm.2011.11.001
© 2011 Elsevier GmbH. All rights reserved.
[4,14] because of possible interference from higher order structuresin the ribosome. Following this concept, Fuchs et al. [15] carriedout a systematic study to evaluate the effect of target site locationby performing hybridizations over the entire 16S rRNA moleculeof Escherichia coli. Their results were interpreted as demonstratingthe existence of inherently inaccessible regions on the 16S rRNAof E. coli. On a follow-up publication, Behrens et al. [9] used a sim-ilar approach to generate accessibility maps for three additionalmicroorganisms, with the purpose of evaluating whether the loca-tion of inaccessible regions found in E. coli could be extended toother organisms. They produced a consensus map from the combi-nation of the accessibilities observed with the different organisms.Another publication from the same group of researchers inves-tigated the effect of RNA-RNA and RNA-protein interactions onhybridization efficiency [7]. However a correlation between flu-orescence signal intensity and ribosomal structure could not befound.
Searching for alternative mechanisms to understand rRNA
accessibility to DNA probes, Yilmaz and Noguera [28] demonstratedthat the thermodynamic affinity of the probe to the target site, mea-sured as the overall free energy (�Goverall◦) of a reaction includingprobe-target hybridization as well as probe and target self folding,

486 H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495
F n nucl
wgcwmpmsehotsdc
pettitoiu[tn
M
M
LscaLaDt(m
gR0Rhwa[
for obtaining the scores yielding maximum correlation (Table 1)and the corresponding best fitting line that described the relation-ship between probe brightness and the total probe quenching factor(Fig. 2).
y = 16 .81x + 150 .7
0
20
40
60
80
100
120
140
-8. 0 -7. 0 -6. 0 -5. 0 -4. 0 -3. 0 -2. 0
Fluo
resc
ence
Inte
nsity
(CB
U)
Quenching Factor (Qf)
ig. 1. Illustration of quenching neighborhood where rRNA nucleotides play a role i
as a good predictor of probe brightness. In addition, it was sug-ested that the absence of formamide in the hybridization bufferould result in kinetic limitations when short hybridization periodsere used. Applying these concepts, Yilmaz et al. [30] used ther-odynamic principles for probe design and extended hybridization
eriods to demonstrate that the entire 16S rRNA of E. coli could beade accessible to DNA probes, and thus, that there were no intrin-
ically inaccessible sites on the ssu rRNA of this organism. Yilmazt al. [30] concluded that the most important criterion for efficientybridization was the establishment of DNA/RNA hybrids with anverall thermodynamic affinity of −13 kcal/mol or lower. In addi-ion, they also showed that even with high hybridization efficiency,ignal intensity of fluorescein-labeled probes might have been lowue to fluorophore quenching, which is known to depend on thelose proximity of specific nucleotides [12,23].
The thermodynamic affinity approach for the design of FISHrobes has so far been proven to be effective for E. coli [28,30], but anxperimental demonstration that the concept can be generalizedo other organisms has not been provided. Therefore, the objec-ive of this study was to determine whether random target sitesn the ssu rRNA of other organisms could also be made accessibleo DNA probes when thermodynamic and kinetic limitations werevercome. To achieve this goal, FISH experiments with four organ-sms representing different phylogenetic groups were conductedsing flow cytometry methods described in earlier publications29,30]. In order to minimize the problem of probe quenching inhis analysis, we also developed an empirical formulation to predictucleobase-specific fluorescein quenching.
aterials and methods
icrobial growth conditions
Stenotrophomonas maltophilia was grown aerobically on LennoxB broth (Fisher Biotech, Fair Lawn, NJ) at 25 ◦C. Rhodobacterphaeroides was grown either aerobically or photoheterotrophi-ally at 30 ◦C, with Sistrom’s medium [20] containing succinates the organic substrate. Bacillus subtilis was grown aerobically onB broth at 37 ◦C. Saccharomyces cerevisiae was grown at 37 ◦C on
medium that contained 20% BactoTM Yeast Extract (Becton andickinson and Company, Sparks, MD), 40% BactoTM peptone (Bec-
on and Dickinson and Company, Sparks, MD) and 40% d(+)-glucoseAcros, NJ). Growth curves were generated for each organism by
easuring optical density at 600 nm.For FISH experiments, cells were harvested in the exponential
rowth phase. Optical density values at fixation for S. maltophilia,. sphaeroides, B. subtilis, and Sc. cerevisiae were 0.40 ± 0.05,.70 ± 0.15, 0.85 ± 0.05, and 1.5 ± 0.1, respectively. S. maltophilia,. sphaeroides and B. subtilis cells were fixed with 4% paraformalde-
yde for 30 min at room temperature. Sc. cerevisiae was also fixedith 4% paraformaldehyde and at room temperature, but the fix-tion period was 3 h, following the procedures of Amann et al.6]. Longer fixation periods may potentially inactivate RNases that
eobase-specific quenching. Key positions are numbered in accordance with Table 1.
otherwise may degrade rRNA during hybridization. Fixed cellswere pelleted, resuspended in 1× PBS (phosphate buffered saline),and stored at 4 ◦C until hybridization. Paraformaldehyde fixationworked well with log-phase B. subtilis in this study, but in general,paraformaldehyde fixation is not recommended for Gram positiveorganisms, as its effectiveness is growth phase dependent.
Probe synthesis
Mono- and dual-labeled probes were used in this study. Allmono-labeled probes were synthesized at the University of Wis-consin Biotechnology Center with fluorescein phosphoramidite(FAM) (applicable to all probes) or Cy5-CE phosphoramidite (appli-cable only to probes suspected of experiencing nucleobase-specificquenching) attached to the 5′ end of the probe. Dual-labeled probeswith FAM attached to both 5′ and 3′ ends were purchased fromSigma–Aldrich (Saint Louis, MO).
Evaluation of fluorophore quenching
The co-variance between probe brightness and the identity ofnearby rRNA nucleotides to the 3′ end of target site (Fig. 1) was eval-uated with multi-way analysis of variance (ANOVAN), using the“anovan” function of MATLAB (The MathWorks Inc., Natick, MA).Statistical p-value output by this function characterized the rel-evance of nucleotide position and type to quenching. The “solver”routine of MS Excel (Microsoft Corp., Redmond, WA) was employed
Fig. 2. Maximized correlation between quenching factor and fluorescence intensityfor high affinity probes from Yilmaz et al. [30], upon 96 h hybridization periods.Solid curve is the best fitting line. Dashed line indicates the threshold used for probeselection in this study.
H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495 487
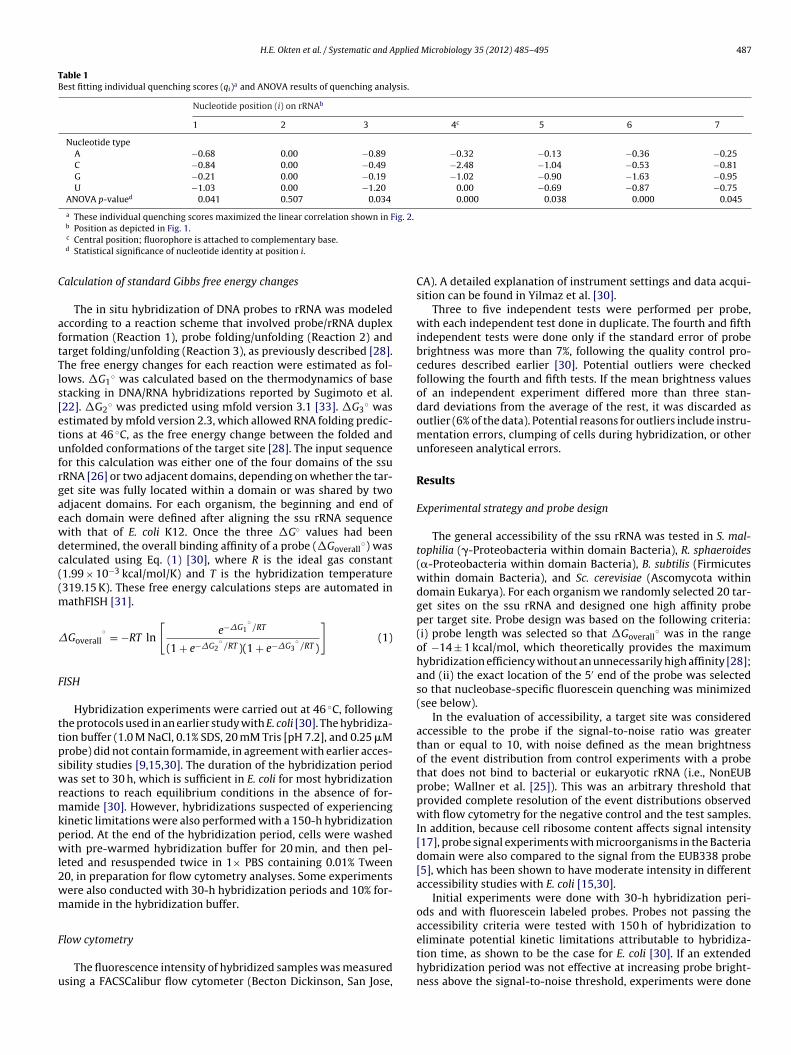
Table 1Best fitting individual quenching scores (qi)a and ANOVA results of quenching analysis.
Nucleotide position (i) on rRNAb
1 2 3 4c 5 6 7
Nucleotide typeA −0.68 0.00 −0.89 −0.32 −0.13 −0.36 −0.25C −0.84 0.00 −0.49 −2.48 −1.04 −0.53 −0.81G −0.21 0.00 −0.19 −1.02 −0.90 −1.63 −0.95U −1.03 0.00 −1.20 0.00 −0.69 −0.87 −0.75
ANOVA p-valued 0.041 0.507 0.034 0.000 0.038 0.000 0.045
a These individual quenching scores maximized the linear correlation shown in Fig. 2.
C
aftTls[etufrgaewdc((m
�
F
ttpswrmkpwl2wm
F
u
b Position as depicted in Fig. 1.c Central position; fluorophore is attached to complementary base.d Statistical significance of nucleotide identity at position i.
alculation of standard Gibbs free energy changes
The in situ hybridization of DNA probes to rRNA was modeledccording to a reaction scheme that involved probe/rRNA duplexormation (Reaction 1), probe folding/unfolding (Reaction 2) andarget folding/unfolding (Reaction 3), as previously described [28].he free energy changes for each reaction were estimated as fol-ows. �G1
◦ was calculated based on the thermodynamics of basetacking in DNA/RNA hybridizations reported by Sugimoto et al.22]. �G2
◦ was predicted using mfold version 3.1 [33]. �G3◦ was
stimated by mfold version 2.3, which allowed RNA folding predic-ions at 46 ◦C, as the free energy change between the folded andnfolded conformations of the target site [28]. The input sequenceor this calculation was either one of the four domains of the ssuRNA [26] or two adjacent domains, depending on whether the tar-et site was fully located within a domain or was shared by twodjacent domains. For each organism, the beginning and end ofach domain were defined after aligning the ssu rRNA sequenceith that of E. coli K12. Once the three �G◦ values had beenetermined, the overall binding affinity of a probe (�Goverall
◦) wasalculated using Eq. (1) [30], where R is the ideal gas constant1.99 × 10−3 kcal/mol/K) and T is the hybridization temperature319.15 K). These free energy calculations steps are automated in
athFISH [31].
Goverall◦ = −RT ln
[e−�G1
◦/RT
(1 + e−�G2◦
/RT )(1 + e−�G3◦
/RT )
](1)
ISH
Hybridization experiments were carried out at 46 ◦C, followinghe protocols used in an earlier study with E. coli [30]. The hybridiza-ion buffer (1.0 M NaCl, 0.1% SDS, 20 mM Tris [pH 7.2], and 0.25 �Mrobe) did not contain formamide, in agreement with earlier acces-ibility studies [9,15,30]. The duration of the hybridization periodas set to 30 h, which is sufficient in E. coli for most hybridization
eactions to reach equilibrium conditions in the absence of for-amide [30]. However, hybridizations suspected of experiencing
inetic limitations were also performed with a 150-h hybridizationeriod. At the end of the hybridization period, cells were washedith pre-warmed hybridization buffer for 20 min, and then pel-
eted and resuspended twice in 1× PBS containing 0.01% Tween0, in preparation for flow cytometry analyses. Some experimentsere also conducted with 30-h hybridization periods and 10% for-amide in the hybridization buffer.
low cytometry
The fluorescence intensity of hybridized samples was measuredsing a FACSCalibur flow cytometer (Becton Dickinson, San Jose,
CA). A detailed explanation of instrument settings and data acqui-sition can be found in Yilmaz et al. [30].
Three to five independent tests were performed per probe,with each independent test done in duplicate. The fourth and fifthindependent tests were done only if the standard error of probebrightness was more than 7%, following the quality control pro-cedures described earlier [30]. Potential outliers were checkedfollowing the fourth and fifth tests. If the mean brightness valuesof an independent experiment differed more than three stan-dard deviations from the average of the rest, it was discarded asoutlier (6% of the data). Potential reasons for outliers include instru-mentation errors, clumping of cells during hybridization, or otherunforeseen analytical errors.
Results
Experimental strategy and probe design
The general accessibility of the ssu rRNA was tested in S. mal-tophilia (�-Proteobacteria within domain Bacteria), R. sphaeroides(�-Proteobacteria within domain Bacteria), B. subtilis (Firmicuteswithin domain Bacteria), and Sc. cerevisiae (Ascomycota withindomain Eukarya). For each organism we randomly selected 20 tar-get sites on the ssu rRNA and designed one high affinity probeper target site. Probe design was based on the following criteria:(i) probe length was selected so that �Goverall
◦ was in the rangeof −14 ± 1 kcal/mol, which theoretically provides the maximumhybridization efficiency without an unnecessarily high affinity [28];and (ii) the exact location of the 5′ end of the probe was selectedso that nucleobase-specific fluorescein quenching was minimized(see below).
In the evaluation of accessibility, a target site was consideredaccessible to the probe if the signal-to-noise ratio was greaterthan or equal to 10, with noise defined as the mean brightnessof the event distribution from control experiments with a probethat does not bind to bacterial or eukaryotic rRNA (i.e., NonEUBprobe; Wallner et al. [25]). This was an arbitrary threshold thatprovided complete resolution of the event distributions observedwith flow cytometry for the negative control and the test samples.In addition, because cell ribosome content affects signal intensity[17], probe signal experiments with microorganisms in the Bacteriadomain were also compared to the signal from the EUB338 probe[5], which has been shown to have moderate intensity in differentaccessibility studies with E. coli [15,30].
Initial experiments were done with 30-h hybridization peri-ods and with fluorescein labeled probes. Probes not passing theaccessibility criteria were tested with 150 h of hybridization to
eliminate potential kinetic limitations attributable to hybridiza-tion time, as shown to be the case for E. coli [30]. If an extendedhybridization period was not effective at increasing probe bright-ness above the signal-to-noise threshold, experiments were done
4 pplied
wattwti
H
[(otrvtoorb(iotoutaEp
Q
nettoon
tcbsatiahtibsblpcsli
88 H.E. Okten et al. / Systematic and A
ith Cy5-labeled probes, since it has been shown that cyanine dyesre less affected by nucleobase-specific quenching [18] and thathey improve duplex stability [19,21]. For probes not passing thehreshold test with either fluorophore, a more detailed analysisas conducted with probes re-designed for the same target sites
o potentially minimize probe quenching, increase �Goverall◦, or
ncreasing fluorophore concentration by using dual labeled probes.
euristic model to calculate nucleobase-specific quenching
The dataset of probes targeting E. coli described in Yilmaz et al.30] contains 73 probes designed with �Goverall
◦ < −13.5 kcal/molsee Supplementary Material, Table S1). The fluorescence intensityf these probes, upon 96-h hybridizations that allowed equilibriumo be established, varied from as low as 12 to as high as 125 on aelative scale. Since kinetic and thermodynamic limitations wereirtually eliminated in the study of Yilmaz et al. [30], we assumedhat signal intensity variability resulted from the quenching of flu-rescein and developed a heuristic approach to calculate the effectf specific nucleotides located in the neighborhood of the fluo-ophore. Basically, we aimed at establishing a linear correlationetween fluorescence intensity and a total probe quenching factorQf) calculated with Eq. (2) as the sum of the individual quench-ng contributions (qi) of the ribonucleotides in the neighborhoodf the fluorophore. This neighborhood corresponds to the 3′ end ofhe target site for probes labeled at the 5′ end (Fig. 1). The numberf ribonucleotides to include in the neighborhood was determinedsing multivariate analysis of variance tests, which showed a statis-ically significant relationship (p < 0.05) between probe brightnessnd the type of nucleotide in 6 out of the 7 positions shown in Fig. 1.xtending the neighborhood in either direction did not reveal theresence of other statistically significant quenching interactions.
f =7∑
i=1
qi (2)
As a control test, we evaluated the covariance of probe bright-ess and nucleotide type in an equivalent neighborhood at the 5′
nd of the target site, which is far away from the fluorophore. Inhis analysis, p-values for each of the seven positions were greaterhan 0.05, and thus, no statistically significant correlations werebserved. This control test provides confidence that the covariancebserved in the neighborhood of the fluorophore was representingucleobase-specific quenching interactions.
Individual quenching scores (qi) for the different nucleotideypes in the defined ribonucleotide neighborhood were then cal-ulated using an iterative approach so that (i) a linear correlationetween the probe quenching factor (Qf) and probe signal inten-ity was obtained, as demonstrated in Fig. 2, (ii) quenching scorest position 2 were equal to zero, as they were shown to be not sta-istically significant (p-values are shown in Table 1), and (iii) allndividual quenching scores were less than or equal to zero (anrbitrary rule). Thus, the more negative the individual score, theigher its influence on fluorescein quenching. Table 1 summarizeshe quenching scores that maximized the linear correlation shownn Fig. 2. This set of qi values is not unique, as many different com-inations can be shown to provide the same level of linearization ofignal intensity versus Qf. Nevertheless, the linear correlation cane used to define a quenching criterion to avoid designing probes
ikely to be limited by fluorescein quenching. Thus, for the design ofrobes in this study, we selected Qf > −5.4, which as shown in Fig. 2,
orresponds to a signal intensity greater than 60 in the brightnesscale used by Yilmaz et al. [30]. This intensity corresponds to theower end of the two upper brightness classes of probes as definedn earlier accessibility studies [15,30].Microbiology 35 (2012) 485– 495
The usefulness of the applied quenching rule is dependent onthe predictive power of the heuristic model. Since a total of 24parameters (Table 1) were used to fit the model to 73 observa-tions, it is possible that the model is overparameterized and unableto predict independent observations. Furthermore, the set of 73probes used in model training does not capture potential quenchingrelationships with all nucleotide permutations. Thus, we evalu-ated the model further using two validation tests as explained inSupplementary Material. First, we performed leave-one-out crossvalidation (LOOCV) [11], wherein a probe in the training set is leftoutside the best-fitting and predicted independently based on themodified model trained by the remaining 72 probes (Fig. S1). Com-pared to the average absolute predictive error of 8.9 CBU in thebest-fit (i.e., Fig. 2), average LOOCV prediction error increased onlyto 12 CBU, and was somewhat evenly distributed for all probes(Fig. S1). Thus, the model did not seem to be highly overparameter-ized. Second, we used two independent sets for validation (a total of23 probes), both obtained from published data [29,30] (Table S1).Although one set had slightly lower free energy values than thetraining set (−13.0 kcal/mol < �Goverall
◦ < −13.5 kcal/mol) and theother was evaluated at shorter hybridization periods (24 h), themajority of these probes were consistent with the trend in Fig. 2with only two exceptions (see Fig. S2). Furthermore, the lowestintensity obtained from 17 independent probes that conformed tothe design criteria of this study (i.e., predicted intensity > 60 CBU)was 51 CBU, a moderate level of brightness. Therefore these val-idation tests show that the relationship in Fig. 2 represents thegeneral trend in the probe population studied and that the applieddesign criterion effectively removes quenching potential, althoughquenching by unknown nucleotide combinations or other factorsshould still be expected for a small fraction of new probes.
Hybridizations at 20 randomly generated sites
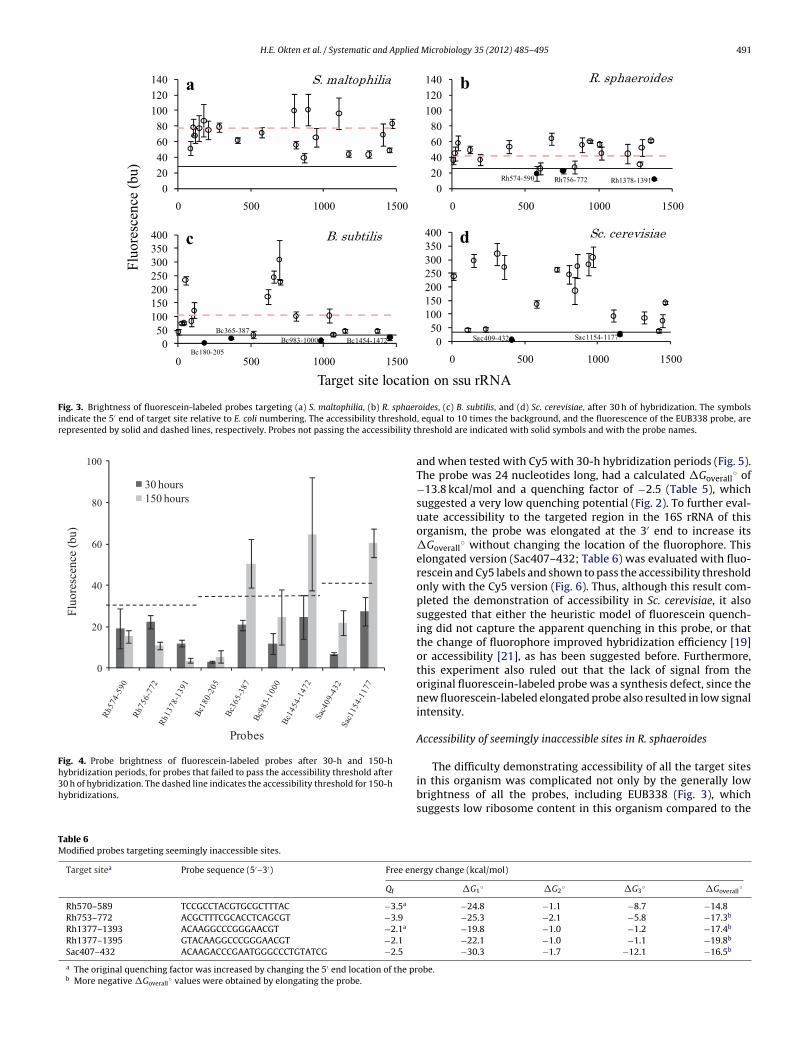
Tables 2–5 show detailed information of the 20 probes designedfor each organism, including their thermodynamic parameters andquenching factors. Fig. 3 summarizes the resulting probe bright-ness for each organism in comparison to the accessibility threshold(i.e., signal greater than or equal to 10 times the noise) and thesignal from the EUB338 probe, when applicable. As shown in thefigure, the overall signal intensity was lower for S. maltophilia andR. sphaeroides compared to B. subtilis and Sc. cerevisiae. In addition,the signal from EUB338 was significantly lower in R. sphaeroidescompared to B. subtilis and S. maltophilia, suggesting lower ribo-some content in R. sphaeroides compared to the other two membersof the domain Bacteria. Regarding accessibility, all probes target-ing S. maltophilia produced fluorescence intensities higher than thethreshold, while 3 probes targeting R. sphaeroides, 4 probes target-ing B. subtilis, and 2 probes targeting Sc. cerevisiae had signals lowerthan the selected threshold. The accessibility of the sites targetedby these 9 probes was then further investigated.
Effect of hybridization period on probes not passing the brightnessthreshold
The first level of evaluation of probes that did not pass thebrightness threshold with a 30-h hybridization period was anextension of hybridization to 150 h. Fig. 4 shows the comparisonof probe brightness values for the two hybridization periods. Thefigure demonstrates that an extended hybridization period did notimprove the brightness of any of the probes targeting R. sphaeroides,
but significant brightness increases were obtained for 2 out of 4probes targeting B. subtilis and 1 out of 2 probes targeting Sc. cere-visiae, suggesting that these sites are accessible to oligonucleotideprobes, but accessibility may be kinetically limited.
H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495 489
Table 2Characteristics of probes targeting 20 randomly chosen sites on the ssu rRNA of S. maltophilia.
Target sitea Probe sequence (5′–3′) Free energy change (kcal/mol) Quenchingfactor (Qf)
Probe brightnessb (bu)
�G1◦ �G2
◦ �G3◦ �Goverall
◦ 30 h 150 h
NonEUBc ACTCCTACGGGAGGCAGC – – – – – 2.9 ± 0.2 3.2 ± 0.2EUB338 GCTGCCTCCCGTAGGAGT −25.1 −1.6 −5.4 −18.1 −6.0 77 ± 7 179 ± 65St87–105 CCACTCGCCACCCAGAGAG −24.5 −0.4 −9.7 −14.1 −5.0 52 ± 9 180 ± 23St106–122 TATTCCTCACCCGTCCG −21.1 3.3 −7.2 −13.9 −4.4 79 ± 11 166 ± 10St115–131 ATTCCGATGTATTCCTC −16.5 1.5 −1.7 −14.7 −3.3 68 ± 9 170 ± 10St146–165 AAGTTTCCCTACGTTATCCC −21.9 0.8 −7.4 −14.4 −3.5 77 ± 16 217 ± 37St178–195 TCACCCGTAGGTCGTATG −19.9 −0.8 −5.8 −13.2 −2.2 87 ± 21 232 ± 14St210–227 TCAATCGCGCAAGGTCCG −19.6 1.4 −6.1 −13.5 −3.5 75 ± 12 195 ± 44St282–302 CCTCTTAGACCAGCTACGGAT −23.2 0.0 −9.3 −13.5 −3.4 79 ± 6 194 ± 42St411–433 CTTTACAACCCGAAGGCCTTCTT −24.7 −0.7 −9.9 −13.9 −5.0 62 ± 4 183 ± 44St575–598 ACTTAAACGACCACCTACGCACGC −25.0 0.6 – −14.2 −3.4 72 ± 7 126 ± 11St796–813 AGGGCGTGGACTACCAGG −21.4 −0.1 −7.0 −13.8 −2.1 100 ± 21 266 ± 12St812–832 ATCCAGTTCGCATCGTTTAG −20.3 1.0 −6.9 −13.3 −3.6 56 ± 4 175 ± 36St865–884 AGGCGGCGAACTTAACGCGT −21.7 −0.9 −6.4 −14.3 −4.2 39 ± 6 83 ± 8St891–909 TTTCAGTCTTGCGACCGTA −20.2 0.0 −6.8 −13 −4.2 101 ± 20 235 ± 7St947–971 CGTTGCATCGAATTAAACCACATAC −20.5 0.6 −6.9 −13.3 −5.1 66 ± 12 165 ± 18St1105–1127 CAACTAAGGACAAGGGTTGCGCT −22.8 0.3 −8 −14.5 −4.3 96 ± 20 122 ± 27St1172–1187 CCCCACCTTCCTCCGG −24.2 1.4 −9.9 −14.2 −3.4 44 ± 4 266 ± 13St1309–1332 TCATGGAGTCGAGTTGCAGACTCC −26.7 −3.0 – −13.5 −3.7 44 ± 5 107 ± 41St1406–1426 CAACAAACTCCCATGGTGTGA −19.9 −0.3 −6.3 −13 −4.6 69 ± 14 203 ± 25St1453–1470 CGTGGCAAGCGCCCTCCC −26.8 −1.6 – −15 −5.1 49 ± 3 189 ± 12St1470–1487 CCAGTCATCGGCCACACC −23.0 0.6 −9.1 −13.7 −5.3 83 ± 6 147 ± 35
a Target sites indicate start and end nucleotides, according to E. coli numbering.b
miaaut
TC
Average and standard deviation of at least three independent experiments.c Control probe used to determine background fluorescence.
Interestingly, contrary to the expectations from E. coli experi-ents [30], an extended hybridization period decreased the signal
ntensity of most of the probes targeting R. sphaeroides (Table 3) andbout half of the probes targeting Sc. cerevisiae (Table 5). Therefore,
n extension of the hybridization period when formamide is notsed in the hybridization buffer cannot be generalized as a strategyo increase signal intensity by eliminating kinetic limitations.able 3haracteristics of probes targeting 20 randomly chosen sites on the ssu rRNA of R. sphaer
Target sitea Probe sequence (5′–3′) Free energy change (kc
�G1◦ �G2
◦
NonEUBc ACTCCTACGGGAGGCAGC – –
EUB338 GCTGCCTCCCGTAGGAGT −25.1 −1.6
Rh8–26 TGAGCCAGGATCAAACTCT −18.0 0.6
Rh18–35 CGTTCATTCTGAGCCAGG −19.3 0.5
Rh40–56 ATGTGTTAGGCCTGCCG −20.3 0.6
Rh122–138 AGGGCACGTTCCCACGC −22.5 −0.9
Rh192–208 TCTTTCCCCCAAAGGG −18.3 −1.1
Rh389–406 CACGCGGCATGGCTAGAT −20.2 0.7
Rh574–590 ATCCGCCTACGTGCGCT −22.3 −1.1
Rh601–621 TGAGCCCTGGGATTTCACCTC −25.5 0.5
Rh678–703 CTACGAATTTCACCTCTACACTCGGA −25.4 0.4
Rh756–772 ACGCTTTCGCACCTCAG −19.3 0.1
Rh836–855 CACCGAACAGCATGCTGCC −22.1 −0.6
Rh883–897 GGCCGTACTCCCCAG −21.0 1.5
Rh939–954 CCACATGCTCCACCGC −20.8 1.1
Rh1006–1029 ACTGAAGGAACCATCTCTGGAACC −23.1 −1.2
Rh1017–1036 CCAGCCGAACTGAAGGAACCA −21.9 −1.1
Rh1197–1216 CCGTAAGGGCCATGAGGACT −22.8 −1.0
Rh1280–1301 ACTGAGACAGCTTTTTGGGAT −19.8 0.3
Rh1292–1315 AGACCCCAATCCGAACTGAGACAG −24.9 0.5
Rh1354–1374 TCACCGCGTCATGCTGTTACG −23.7 −0.5
Rh1378–1391 AAGGCCCGGGAACG −15.4 −0.7
a Target sites indicate start and end nucleotides, according to E. coli numbering.b Average and standard deviation of at least three independent experiments.c Control probe used to determine background fluorescence.
Evaluation of accessibility with Cy5-labeled probes
A total of 6 probes remained dim after extended periods ofhybridization with fluorescein-labeled probes (Fig. 4). Possible rea-
sons for the low brightness of these probes are: (i) quenching ofthe fluorophore if the heuristic model was unable to identify apotential quenching condition; (ii) low �Goverall◦ value resulting
oides.
al/mol) Quenchingfactor (Qf)
Probe brightnessb (bu)
�G3◦ �Goverall
◦ 30 h 150 h
– – – 2.6 ± 0.5 3.1 ± 0.8−7.0 −16.5 −6.0 42 ± 5 24 ± 1−3.5 −14.2 −3.2 36 ± 6 25 ± 2−4.6 −14.5 −5.1 45 ± 9 22 ± 3−5.5 −14.6 −3.2 58 ± 9 31 ± 4−7.4 −14.0 −3.8 49 ± 5 31 ± 1−3.1 −14.0 −3.5 37 ± 7 24 ± 1−5.4 −14.6 −4.9 54 ± 8 30 ± 1−6.7 −14.4 −4.7 19 ± 10 15 ± 3
−10.5 −14.8 −3.1 26 ± 7 22 ± 3−10.3 −14.8 −3.4 64 ± 8 34 ± 1−5.8 −13.1 −3.9 23 ± 3 10 ± 2−6.3 −15.0 −3.3 27 ± 8 30 ± 2−6.9 −14.0 −4.9 56 ± 9 30 ± 1−6.2 −14.5 −3.7 60 ± 2 33 ± 1−7.3 −14.5 −3.2 56 ± 3 34 ± 3−7.2 −13.5 −3.9 45 ± 8 31 ± 1−7.2 −14.5 −4.4 45 ± 12 26 ± 1−4.8 −14.7 −3.2 31 ± 4 17 ± 3
−11.2 −13.5 −2.4 53 ± 10 31 ± 2−8.6 −14.4 −2.0 61 ± 2 40 ± 3−1.1 −13.3 −4.1 12 ± 2 4 ± 1

490 H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495
Table 4Characteristics of probes targeting 20 randomly chosen sites on the ssu rRNA of B. subtilis.
Target sitea Probe sequence (5′–3′) Free energy change (kcal/mol) Quenchingfactor (Qf)
Probe brightnessb (bu)
�G1◦ �G2
◦ �G3◦ �Goverall
◦ 30 h 150 h
NonEUBc ACTCCTACGGGAGGCAGC – – – – – 3.2 ± 0.5 3.5 ± 0.4EUB338 GCTGCCTCCCGTAGGAGT −25.1 −1.6 −5.4 −18.1 −6.0 104 ± 9 244 ± 50Bc5–22 CCAGGATCAAACTCTCCG −18.1 0.9 −4.0 −13.9 −4.8 44 ± 11 72 ± 8Bc26–41 CGCCAGCGTTCGTCCT −21.8 0.6 −7.6 −14.0 −4.1 75 ± 5 83 ± 30Bc42–60 TTGCATGTATTAGGCACGC −19.0 −0.3 −4.3 −14.1 −4.0 76 ± 6 133 ± 14Bc52–72 TCCGCTCGACTTGCATGTA −21.4 0.7 −7.4 −13.8 −3.7 234 ± 16 257 ± 16Bc94–111 CGTCCGCCGCTAACATC −20.1 0.7 −6.1 −13.9 −5.0 84 ± 20 101 ± 24Bc110–127 CCACGTGTTACTCACCCG −21.3 −0.3 −6.8 −13.9 −3.4 123 ± 29 156 ± 34Bc180–205 CCACCTTTTATGTTTGAACCATGCGGTTC −31.4 −2.5 −15.1 −13.8 −4.1 3.2 ± 0.5 5.6 ± 2.9Bc365–387 AGACTTTCGTCCATTGCGGAAGA −23.6 −2.4 −7.8 −13.4 −2.8 21 ± 3 50 ± 12Bc519–532 TACCGCGGCTGCTG −17.4 0.0 −3.2 −13.8 −2.7 33 ± 13 49 ± 10Bc617–632 CCCTCCCCGGTTGAGC −23.9 0.6 −9.9 −13.8 −3.4 173 ± 28 197 ± 9Bc658–673 CACTCTCCTCTTCTGC −18.5 2.8 −4.7 −13.8 −4.4 246 ± 22 189 ± 16Bc697–713 CTCCACATCTCTACGCA −18.5 0.0 −4.4 −13.8 −3.6 308 ± 72 298 ± 27Bc704–721 TGGTGTTCCTCCACATCT −21.1 −0.1 −6.3 −14.3 −4.2 227 ± 9 225 ± 14Bc808–826 GCACTCATCGTTTACGGCG −20.8 0.0 −5.9 −14.4 −4.2 101 ± 18 134 ± 15Bc983–1000 AGGATGTCAAGACCTGGT −18.2 0.3 −4.3 −13.6 −4.4 12 ± 5 25 ± 13Bc1038–1053 CACCACCTGTCACTCT −18.7 0.2 −4.8 −13.5 −4.6 104 ± 24 157 ± 46Bc1066–1088 CCAACATCTCACGACACGAGCTG −23.6 0.0 −9.2 −14.0 −4.8 34 ± 6 155 ± 24Bc1147–1167 TCACCGGCAGTCACCTTAGAG −24.2 0.6 −10.4 −13.7 −2.4 47 ± 8 80 ± 26Bc1367–1387 CCCGGGAACGTATTCACCGCG −25.5 −1.0 −10.1 −14.2 −4.4 48 ± 7 77 ± 13Bc1454–1472 ACCTTCGGCGGCTGGCTCC −27.5 −0.3 −13.4 −13.5 −4.3 25 ± 11 65 ± 27
a Target sites indicate start and end nucleotides, according to E. coli numbering.
ftetpttsfis
TC
b Average and standard deviation of at least three independent experiments.c Control probe used to determine background fluorescence.
rom an underestimated free energy change for the unfolding ofhe target site (i.e., �G3
◦, which is the parameter with the great-st uncertainty in the model of Yilmaz and Noguera [28]); (iii) theargeted site is truly inaccessible to oligonucleotide probes; or (iv)robe synthesis was defective. Thus, the next step in the evalua-ion of accessibility was done with probes labeled with Cy5, since
his dye is less prone to quenching [18] and may increase duplextability [19,21]. Fig. 5 shows that the Cy5-labeled probes were suf-cient to demonstrate accessibility in the two remaining B. subtilisites, but were not effective at the R. sphaeroides and Sc. cerevisiaeable 5haracteristics of probes targeting 20 randomly chosen sites on the ssu rRNA of Sc. cerevi
Target sitea Probe sequence (5′–3′) Free energy change
�G1◦ �G2
◦
NonEUBc ACTCCTACGGGAGGCAGC – –
Sac13–34 ATATGACTACTGGCAGGATCAA −19.4 1.1
Sac109–135 ATAAACGATAACTGATTTAATGAGCCAT −19.7 1.1
Sac149–173 ATTAGCTCTAGAATTACCACAGTTATA −22.6 0.7
Sac231–247 AGTTGATAGGGCAGAAATT −15.0 1.8
Sac311–326 CTCAGGCTCCCTCTCC −22.4 0.1
Sac358–378 AGGATTGGGTAATTTGCGCGC −22.1 0.7
Sac409–432 ACAAGACCCGAATGGGCCCTGTAT −27.8 −1.7
Sac583–600 CCAACCGGGCCCAAAGTTCAAC −25.0 0.7
Sac720–741 CCTTGGCAAATGCTTTCGCAGTAG −25.3 −0.6
Sac804–825 TCGGCATAGTTTATGGTTAAGA −19.2 0.1
Sac844–859 AAGGTGCCGAGTGGGTCATTA −23.2 0.1
Sac861–882 ACCCAAAGACTTTGATTTCTCG −19.7 −0.7
Sac938–954 CCGCAGGCTCCACTCCT −24.4 −0.3
Sac966–983 TGAGTTTCCCCGTGTTGAG −21.5 1.2
Sac1108–1131 ACCACTATTTAGTAGGTTAAGGTCTC −24.6 −0.1
Sac1154–1177 CATCGGCTTGAAACCGATAGTCCC −26.2 −2.9
Sac1321–1347 CTAGGAATTCCTCGTTGAAGAGCAATA −24.2 −0.9
Sac1420–1448 AGATCCTGAGGCCTCACTAAGCCATTC −31.5 −1.2
Sac1446–1458 CTCCGCTCTGAGATGGAGTTGCCCCCT −37.8 −0.6
Sac1464–1485 CCTCTAAATGACCAAGTTTGTCC −22.2 0.7
a Target sites indicate start and end nucleotides, according to E. coli numbering.b Average and standard deviation of at least three independent experiments.c Control probe used to determine background fluorescence.
target sites. Therefore, modifications to the probes were made toeither circumvent potential quenching limitations or to increasethe �Goverall
◦.
Accessibility of Sc. cerevisiae site targeted by probe Sac409–432
Probe Sac409–432 was the only probe targeting Sc. cerevisiaefor which accessibility to the target site remained to be demon-strated. Probe signal was below the accessibility threshold whenused with fluorescein at 30-h and 150-h hybridizations (Fig. 4),
siae.
(kcal/mol) Quenchingfactor (Qf)
Probe brightnessb (bu)
�G3◦ �Goverall
◦ 30 h 150 h
– – – 3.3 ± 0.3 4.1 ± 0.2−5.3 −14.0 −3.7 240 ± 13 78 ± 5−6.2 −13.4 −4.8 42.2 ± 8 93 ± 5−7.8 −14.6 −2.8 297 ± 24 166 ± 5
0.0 −14.5 −3.5 45 ± 8 33 ± 5−8.1 −13.9 −3.7 322 ± 41 222 ± 10−8.7 −13.2 −4.9 273 ± 44 192 ± 10
−12.3 −13.8 −2.5 7 ± 1 22 ± 6−11.5 −13.3 −4.8 137 ± 13 70 ± 5−11.0 −13.5 −3.5 263 ± 8 209 ± 5−3.9 −14.9 −3.5 245 ± 35 235 ± 8−9.5 −13.3 −3.7 186 ± 48 151 ± 24−5.3 −13.5 −2.4 277 ± 45 183 ± 3−9.3 −14.5 −4.1 283 ± 40 203 ± 4−6.8 −14.6 −3.5 309 ± 37 205 ± 9−9.3 −14.8 −3.4 93 ± 22 151 ± 7−9.4 −13.9 −3.9 27 ± 7 60 ± 7−8.9 −14.3 −4.0 87 ± 22 116 ± 9
−16.4 −13.8 −3.3 39 ± 8 63 ± 6−23.0 −14.0 −3.4 77 ± 24 149 ± 4−7.8 −14.2 −2.7 142 ± 4 200 ± 5
H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495 491
Target site loca tion on ssu rRNA
Fluo
resc
ence
(bu)
020406080
100120140
0 500 100 0 150 0
b R. sphaeroides
Rh574 -590 Rh756-772 Rh1378-13910
20406080
100120140
0 500 1000 150 0
a S. maltophili a
050
100150200250300350400
0 500 10 00 15 00
c B. subtilis
Bc180 -205
Bc365-387Bc983-1000 Bc14 54-1472 0
50100150200250300350400
0 500 10 00 1500
d Sc. cerevisiae
Sac409-432 Sac1154-1177
Fig. 3. Brightness of fluorescein-labeled probes targeting (a) S. maltophilia, (b) R. sphaerindicate the 5′ end of target site relative to E. coli numbering. The accessibility thresholdrepresented by solid and dashed lines, respectively. Probes not passing the accessibility t
0
20
40
60
80
100
Fluo
resc
ence
(bu)
Probes
30 hours150 hours
Fig. 4. Probe brightness of fluorescein-labeled probes after 30-h and 150-hhybridization periods, for probes that failed to pass the accessibility threshold after30 h of hybridization. The dashed line indicates the accessibility threshold for 150-hhybridizations.
Table 6Modified probes targeting seemingly inaccessible sites.
Target sitea Probe sequence (5′–3′) Free ene
Qf
Rh570–589 TCCGCCTACGTGCGCTTTAC −3.5a
Rh753–772 ACGCTTTCGCACCTCAGCGT −3.9
Rh1377–1393 ACAAGGCCCGGGAACGT −2.1a
Rh1377–1395 GTACAAGGCCCGGGAACGT −2.1
Sac407–432 ACAAGACCCGAATGGGCCCTGTATCG −2.5
a The original quenching factor was increased by changing the 5′ end location of the prb More negative �Goverall
◦ values were obtained by elongating the probe.
oides, (c) B. subtilis, and (d) Sc. cerevisiae, after 30 h of hybridization. The symbols, equal to 10 times the background, and the fluorescence of the EUB338 probe, arehreshold are indicated with solid symbols and with the probe names.
and when tested with Cy5 with 30-h hybridization periods (Fig. 5).The probe was 24 nucleotides long, had a calculated �Goverall
◦ of−13.8 kcal/mol and a quenching factor of −2.5 (Table 5), whichsuggested a very low quenching potential (Fig. 2). To further eval-uate accessibility to the targeted region in the 16S rRNA of thisorganism, the probe was elongated at the 3′ end to increase its�Goverall
◦ without changing the location of the fluorophore. Thiselongated version (Sac407–432; Table 6) was evaluated with fluo-rescein and Cy5 labels and shown to pass the accessibility thresholdonly with the Cy5 version (Fig. 6). Thus, although this result com-pleted the demonstration of accessibility in Sc. cerevisiae, it alsosuggested that either the heuristic model of fluorescein quench-ing did not capture the apparent quenching in this probe, or thatthe change of fluorophore improved hybridization efficiency [19]or accessibility [21], as has been suggested before. Furthermore,this experiment also ruled out that the lack of signal from theoriginal fluorescein-labeled probe was a synthesis defect, since thenew fluorescein-labeled elongated probe also resulted in low signalintensity.
Accessibility of seemingly inaccessible sites in R. sphaeroides
The difficulty demonstrating accessibility of all the target sitesin this organism was complicated not only by the generally lowbrightness of all the probes, including EUB338 (Fig. 3), whichsuggests low ribosome content in this organism compared to the
rgy change (kcal/mol)
�G1◦ �G2
◦ �G3◦ �Goverall
◦
−24.8 −1.1 −8.7 −14.8−25.3 −2.1 −5.8 −17.3b
−19.8 −1.0 −1.2 −17.4b
−22.1 −1.0 −1.1 −19.8b
−30.3 −1.7 −12.1 −16.5b
obe.

492 H.E. Okten et al. / Systematic and Applied Microbiology 35 (2012) 485– 495
Fig. 5. Probe brightness of Cy5-labeled probes after 30-h hybridization periods, forthb
osot
ss
Fttsbft
0
5
10
15
20
25
Rh57 4-59 0* Rh75 6-77 2* Rh137 8-1391*
Signal- to-no
isera�o
Probes
FL_orig Cy 5_orig DL_orig FL_mod
Cy5_mod DL_mod DL_mod2
Fig. 7. Probe brightness of fluorescein-labeled (FL orig, Rh574–590, Rh756–772,and Rh1378–1391), Cy5-labeled (Cy5 orig), and dual-labeled (DL orig,) original andmodified (mod, Rh570–589, Rh753–772, Rh1377–1393, and mod2, Rh1377–1395)probes targeting the three R. sphaeroides sites that did not pass the accessibilitythreshold with the original probes labeled with either fluorescein or Cy5. For com-parison of signal from different fluorophores, probe fluorescence was normalizeddividing by the fluorescence of the NonEUB controls in each experiment (2.6 for DL,
hose probes that failed to pass the accessibility threshold after 30-h and 150-hybridizations with fluorescein-labeled probes. Dashed lines represent the accessi-ility thresholds for Cy5 experiments.
ther organisms tested, but also by the unexpected decrease inignal intensity upon extended hybridization periods (Fig. 4). Flu-rescence of the three probes was below the arbitrary accessibilityhreshold even if Cy5-labeled derivatives were used (Fig. 5).
Thus, the following tests were performed to explore if it was pos-ible to overcome the accessibility threshold in the remaining threeites by altering the experimental conditions. Since R. sphaeroides
0
10
20
30
40
50
60
70
Sac409-432 Sac407-432
Signal-to-no
isera�o
Probes
FL
Cy5
ig. 6. Probe brightness of original (Sac409–432) and modified (Sac407–432) probesargeting the only Sc. cerevisiae site that did not pass the accessibility threshold withhe original probes labeled with either fluorescein (FL) or Cy5. For comparison ofignal from two different fluorophores, probe fluorescence was normalized dividingy the fluorescence of the NonEUB controls in each experiment (3.5 for FL, and 6.0or Cy5). The hybridization time was 30 h. Dashed line represents the accessibilityhreshold.
2.4 for FL, and 3.7 for Cy5). The hybridization time was 30 h. Dashed line representsthe accessibility threshold, and asterisks indicate that probes were also evaluatedin a modified version.
is a photoheterotrophic organism, experiments were conducted tocompare signal intensity obtained from cultures grown under aer-obic and photosynthetic conditions, in case the ribosome contentin these cells vary depending on the growth conditions. However,these experiments did not show a significant difference in probesignal (data not shown). Furthermore, since extended hybridizationperiods negatively affected probe brightness, experiments with themore conventional 3-h hybridization period were done, but alsofailed to increase signal intensity.
Therefore, probe modifications were then made to further assessaccessibility. For all three sites, versions of the original probes weredual labeled [21] with fluorescein and tested with 30-h hybridiza-tion periods. However, this strategy did not result in signals abovethe threshold at any of the three sites (Fig. 7), but helped rule out thepossibility that the low signal with the original fluorescein-labeledprobes could be due to a defective batch of probes. Elongated probeswere not effective with fluorescein either, but when labeled withCy5 probes Rh570–589 and Rh753–772 passed the threshold, whileRh1377–1393 remained dim (Fig. 7). A dual-labeled version ofRh1377–1393 resulted in fluorescent signals just above the acces-sibility threshold, while a dual-labeled probe with an additionalelongation (Rh1377–1395; Table 6) was well above the threshold,completing the demonstration of accessibility in the four organismstested.
Discussion
Probe design strategy
In addition to evaluating the accessibility of random target sitesin four different organisms, the methodologies used in this studyalso allow to evaluate the effectiveness of a FISH probe design strat-egy that uses thermodynamic calculations [28,31] and an empiricalformulation of nucleobase-specific quenching to achieve high sig-
nal intensities from hybridized cells. Probes were designed toyield an overall thermodynamic affinity (�Goverall◦) in the rangeof −14 ± 1 kcal/mol and to avoid placing the fluorophore in rRNAneighborhoods with high fluorescein quenching potential. Probes

pplied
wsow8mspeptocfitSt(dpms
T
mhwaTndti−(o3aldwiwtbHp[fcflmttttpl−atii
H.E. Okten et al. / Systematic and A
ere considered to be successfully hybridized if their signal inten-ity was greater than or equal to the conservative threshold valuef 10 times the background noise. In the first set of experiments,ith no formamide and 30-h hybridization periods, 71 out of
0 probes produced signal intensities above the threshold. All S.altophilia-targeted probes passed the threshold, whereas the R.
phaeroides, B. subtilis and Sc. cerevisiae probe sets contained dimrobes. When the hybridization period was extended to 150 h,ffective hybridization was obtained for an additional set of threerobes. Thus, when potential kinetic limitations were eliminated,he design strategy was proven to be 92.5% effective. That is, 74ut of 80 probes were appropriately designed to yield high fluores-ence signals, a high success rate. The length of these probes rangedrom 14 to 28 nucleotides long because the key design parameters �Goverall
◦ instead of length. Long probes were necessary to reachhe required �Goverall
◦ in target sites with low GC content (e.g.,ac109–135; Table 5) or to overcome strong secondary structures inhe ssu rRNA, which are represented by large negative �G3
◦ valuese.g., Sac1446–1458). Although all sites requiring long probes wereemonstrated to be accessible, they may not be preferred sites forrobe design from a specificity perspective, since shorter probes areore desirable to minimize hybridizations to mismatched target
ites.
arget site accessibility
In regards to accessibility, the design strategy and the imple-ented probe modifications resulted in 100% of the targeted sites
aving sufficiently high probe signal to be called accessible, albeitith probe elongations required for 3 out of the 80 tested sites
nd dual-labeling plus elongation required for an additional site.hree sites were in R. sphaeroides and one site in Sc. cerevisiae. Theeed for probe elongation suggests that a problem with the initiallyesigned probe was an overprediction of �Goverall
◦ most likely dueo an underestimated �G3
◦ [28]. The target site of Rh1378–1391s particularly interesting from this point of view (�G3
◦ is only1.1 kcal/mol) and was analyzed with three probes in this study
Fig. 7, Table 6). It spans most of helix 31 (in numbering systemf Brosius et al. [10]), which connects the two terminals of the′ major domain of 16S rRNA [26], at positions 921–936 (5′ end)nd 1379–1396 (3′ end). Although mfold captures most of theseong-range interactions in the optimal predicted structure of thisomain [28], it also predicts an alternative (suboptimal) structureith a similar folding free energy but without helix 31, whose elim-
nation requires propagating conformational changes that disagreeith the native state. Since helix 31 is non-existent in the subop-
imal structure, the calculated �G3◦ only reflects the difference
etween optimal and suboptimal structures, which is minimal.owever, such conformational changes may not be easily takinglace in this highly packed (i.e., protein-rich) domain of 16S rRNA26,32], thus causing a theoretical underestimation of the actualree energy penalty. To check if this mechanism explains the dis-repancy between �Goverall
◦ values (Table 6) and probe-conferreduorescence (Fig. 7), we calculated �G3
◦ by reducing the whole 3′
ajor domain to helix 31 and a polyA loop (i.e., a stem-loop struc-ure with positions 921–936 followed by a polyA sequence andhen by positions 1379–1396), where the polyA loop representshe rest of the structure, thereby eliminating global conforma-ional changes. The resulting �G3
◦ was −7.7 kcal/mol for all threerobes, which has a greater magnitude than the originally calcu-
ated −1.1 kcal/mol (Table 6), thus yielding �Goverall◦ values of −6.6,
11.0, and −13.3 kcal/mol for probes Rh1378–1391, Rh1377–1393,
nd Rh1377–1395, respectively. This result explains the observa-ion in Fig. 7, which shows probe brightness increasing as the probes elongated. Therefore, �G3◦ calculations involving long rangenteractions as in helix 31 should be handled with care, especially if
Microbiology 35 (2012) 485– 495 493
estimations are low in magnitude. The need for elongated probes inthe three other regions (Table 6) cannot be explained by this type ofmechanism, but smaller gains in �Goverall
◦ may have contributedto an improved binding affinity. However, this is not conclusive,as these sites passed the accessibility threshold only when theywere labeled with Cy5, and therefore, quenching effects and poten-tial accessibility improvements with the change of fluorophore areadditional factors that may have affected probe brightness.
The kinetics of probe/target hybridization can be a majorcomponent of target accessibility. It has been shown that poten-tial kinetic limitations can be overcome by either increasing thehybridization period [30] or adding formamide to the hybridiza-tion buffer [28]. Since all initial hybridizations were conductedwithout formamide in this study, as in earlier accessibility studies[9,15,30], the possibility of kinetic limitations affecting probe signalwas evaluated with 150-hybridization periods. With this approach,3 out of the 9 initially dim probes passed the accessibility thresh-old (Bc365–387, Bc1454–1472, and Sac1154–1177). However, asurprising result was the decrease of signal upon the exten-sion of hybridization in multiple probes targeting R. sphaeroidesand Sc. cerevisiae. To explore whether this decrease could be theresult of rRNA degradation during the long hybridization peri-ods due to residual RNase activity, additional experiments withphotosynthetically-grown R. sphaeroides were conducted to assessthe effect of longer (2.5 h) and shorter (10 min) fixation times,which could be more or less effective at inactivating RNases, respec-tively. However, these experiments did not support a correlationbetween fixation period and signal intensity (data not shown).Also, the decrease in signal intensity at longer hybridization peri-ods could not be repeated in these additional tests, although therewas not a significant increase either. Given that the only dif-ference from the original tests was the mode of growth of R.sphaeroides (anoxic photosynthetic versus aerobic), the reasons forthe observed decrease in signal intensity are not clear. Regardless,long hybridization periods may not be considered practical dueto increased background signal with complex samples, and maynot be necessary for overcoming kinetic limitations, since typi-cal FISH protocols involve formamide in the hybridization buffer,which can do the same job by denaturing the secondary or higher-order ribosomal structure at the target site [16,28]. Accordingly weevaluated the three probes facing potential kinetic barriers alsowith 30-h hybridizations and with formamide in the hybridiza-tion buffer. By adding 10% formamide, probe brightness increased2.1 fold for Bc365–387, 1.7 fold for Bc1454–1472, and 2.8 fold forSac1154–1177. Such brightness increases were sufficient to passthe accessibility threshold with all three probes, and therefore,150-h hybridizations were not essential to demonstrate accessi-bility. Based on this study and earlier datasets [28–30] we believehybridizations performed with an incubation period of up to a dayand at least 10% formamide can effectively remove kinetic limita-tions in most cases.
Quenching
Out of the 80 probes tested, 5 (6.2%) passed the threshold ofaccessibility only when evaluated with Cy5, suggesting that flu-orescein quenching may be limiting the signal intensity of someprobes, even after using the heuristic model to prevent placing thefluorophore in a high quenching neighborhood and after elongat-ing the probes to increase �Goverall
◦. However, although not 100%efficient, the heuristic model appropriately eliminated from con-sideration quenching neighborhoods that resulted in dim probes
in the dataset of Yilmaz et al. [30]. For instance, 10 of the initiallydim probes in the E. coli study had a GT starting sequence at the 5′ ofthe probe. This corresponds to having an adenine in position 3 anda cytosine in position 4 of the quenching neighborhood defined in
4 pplied
FtQoc
dsYsetctrwwthitingwpplefqftwptesnso
fllltnmRsipfmtn
iitFbhiew
94 H.E. Okten et al. / Systematic and A
ig. 1. From Table 1 follows that because of the large magnitude ofhe corresponding qi values, this combination could easily result inf < −5.4, which would be indicative of quenching. As a result, nonef the probes tested in this study had that particular nucleotideombination at the 5′ end of the probe.
The assumption that variations in signal intensity of probesesigned to have high affinity to the target site is due to nucleobase-pecific quenching of fluorescein agrees with the conclusions ofilmaz et al. [30], but contradicts the conclusions of the nucleobase-pecific quenching study of Behrens et al. [8]. In that study, Behrenst al. tested the hypothesis that guanines in high proximity tohe fluorophore of a hybridized probe would decrease the fluores-ence intensity of fluorescein-labeled probes during FISH. To testhe hypothesis, they hybridized a set of 17 probes to a guanine-richegion of the 16S rRNA of E. coli. Some quenching was observedhen probes were hybridized to purified rRNA, but no quenchingas observed during in situ hybridizations. Thus, they concluded
hat nucleobase-specific quenching had a negligible effect on FISHybridizations. An alternative explanation for the lack of quench-
ng during in situ hybridizations in the study of Behrens et al. [8] ishat the evaluated region was not a high quenching region. Accord-ng to the heuristic model of quenching, a guanine-rich region doesot necessarily imply a high quenching neighborhood. For instance,uanines in positions 1 and 3 of the neighborhood defined in Fig. 1ould have a relatively small quenching effect compared to otherossible nucleotides in the same positions (Table 1). Guanines inositions 4–7 would have a stronger quenching effect. The calcu-
ated overall quenching factor, Qf, for the 17 probes used by Behrenst al. [8] resulted in 16 of the probes having Qf > −5.4, and there-ore, the heuristic model predicts that these probes would not beuenched. Only one probe in the dataset had an overall quenchingactor below the threshold. Probe Eco649, with guanines in posi-ions 4–7 of the quenching neighborhood, resulted in Qf = −6.06,hich according to Fig. 2, would be moderately quenched (for com-arison, Qf for EUB338 was −6.0). Interestingly, Probe Eco649 washe probe with the lowest signal intensity in the study of Behrenst al. [8]. Thus, our experimental data is consistent with nucleobase-pecific quenching of fluorescein being a factor that cannot beeglected, and the heuristic model provides a reasonable and con-istent explanation for the lack of significant quenching in the studyf Behrens et al. [8].
On the other hand, the inability to predict the low signals of fiveuorescein-labeled probes in this study could be explained by the
imited size of the quenching modeling dataset [30], which couldack some nucleotide combinations with high quenching poten-ial. Moreover, this simple linear model with a local quenchingeighborhood cannot take into account tertiary interactions thatay create longer range but strong quenching. Nevertheless, probe
h756–772 coincidentally has an almost identical probe in thetudy of Yilmaz et al. [30] (E755–772), and both probes have andentical quenching neighborhood. Probe E755–772 was a brightrobe, ranked in the second highest brightness class, so at leastor this probe, it is reasonable to conclude that the quenching
odel correctly predicted a low quenching potential, suggestinghat a miscalculation of �Goverall
◦ was the likely cause of low sig-al.
If quenching potential was correctly predicted, then themproved signal with Cy5 labeling could be the result of a moderatencrease in probe/target duplex stability [19] represented by a con-ribution to �Goverall
◦ not included in the thermodynamic model.or most probes in the study, the design requirement of �Goverall
◦
eing in the range of −14 ± 1 kcal/mol would place the theoretical
ybridization efficiency at ∼100% with no label, where a moderatencrease in duplex stability would not be significant. However, theffect of the fluorophore change on stability is more likely to be seenhen the actual �Goverall
◦ value is around −8.5 to −10.5 kcal/mol,
Microbiology 35 (2012) 485– 495
where, hybridization efficiency rapidly varies from 14% to 79% [28],and therefore, a moderate free energy addition from Cy5 label-ing could significantly improve hybridization. Hence, in theory,the affinity-based explanation for the fluorescence gains with Cy5would require a ∼3–5 kcal/mol overestimation of �Goverall
◦ (inmagnitude) for the two B. subtilis probes (Fig. 5 and Table 4) and∼6–8 kcal/mol overestimation for the Sc. cerevisiae probe (Fig. 6 andTable 5), which cannot be ruled out without detailed systematicstudies of the effect of labeling on hybridization efficiency.
Finally, dual-fluorescein-labeled probes were used in this studyas suggested by Stoecker et al. [21] for two reasons. First was topotentially minimize the effect of quenching with the expectationthat it was unlikely to have both dyes quenched. Second, to evalu-ate whether the signal-to-noise threshold set in this study could bereached without probe elongation. Since the change from mono- todual-labeled probes did not increase signal to reach the threshold(Fig. 7), this result also suggests that quenching was less likely to bethe cause of low signals. Therefore, overestimations of �Goverall
◦,along with moderate improvements in affinity provided by Cy5seem to be the likely explanations for needing extended probes(with dual-labeling in one case) and Cy5 to complete the acces-sibility demonstrations in the sites that required the additionalanalyses.
Concluding remarks
The general accessibility of the ssu rRNA to DNA probes initiallydemonstrated with E. coli [30] was also shown to be the case forS. maltophilia, R. sphaeroides, B. subtilis, and Sc. cerevisiae. Over-all, 80 target sites were tested in this study. Out of these sites 3were shown to be accessible only after extended incubation orafter adding formamide to the hybridization, 5 sites required afluorophore change, and 4 sites required probe elongations (withone of them requiring dual labeling) before accessibility could bedemonstrated, but the majority of them were easily shown to beaccessible.
The design strategy used in this study, which included thecalculation of �Goverall
◦ to ensure a high thermodynamic affin-ity, and the minimization of strongly quenching neighborhoods,was highly effective at producing hybridizations with high flu-orescence signals. Ninety-two percent of the fluorescein-labeledprobes produced high signals. Cy5-labeled probes demonstratedaccessibility of 5 sites, some of which may have had nucleobase-specific fluorescein quenching, although the results from usingdual-labeled and elongated probes are more consistent with theoriginal probes having lower affinity than predicted by free energycalculations. Thus, combining the design approach described hereand automated in mathFISH [31] with appropriate evaluations ofprobe specificity (which was not relevant in this study), quenchingpotential (may be included in future versions of mathFISH), and acareful consideration of potential site-specific free energy miscal-culations should be a step forward into improving the effectivenessof probe design for FISH applications.
Acknowledgments
This research was supported by National Science Foundationgrant CBET-0606894. We thank Allie Sanders and Lorene Calvanofor their help with flow cytometry analyses.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.syapm.2011.11.001.

pplied
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
Microbiol. 77, 1118–1122.
H.E. Okten et al. / Systematic and A
eferences
[1] Amann, R. (1995) In situ identification of microorganisms by whole cellhybridization with rRNA-targeted nucleic acid probes. In: Akkermans, A.D.L.,van Elsas, D.J., de Bruijn, F.J.S. (Eds.), Molecular Microbial Ecology Manual,Kluwer Academic Publishers, Dordrecht, pp. 1–15.
[2] Amann, R., Fuchs, B.M. (2008) Single-cell identification in microbial commu-nities by improved fluorescence in situ hybridization techniques. Nat. Rev.Microbiol. 6, 339–348.
[3] Amann, R., Fuchs, B.M., Behrens, S. (2001) The identification of microorganismsby fluorescence in situ hybridisation. Curr. Opin. Biotechnol. 12, 231–236.
[4] Amann, R., Ludwig, W., Schleifer, K.-H. (1995) Phylogenetic identification andin situ detection of individual microbial cells without cultivation. Microbiol.Rev. 59, 143–169.
[5] Amann, R.I., Binder, B.J., Olson, R.J., Chisholm, S.W., Devereux, R., Stahl, D.A.(1990) Combination of 16S rRNA-targeted oligonucleotide probes with flowcytometry for analyzing mixed microbial populations. Appl. Environ. Microbiol.56, 1919–1925.
[6] Amann, R.I., Krumholz, L., Stahl, D.A. (1990) Fluorescent-oligonucleotide prob-ing of whole cells for determinative, phylogenetic, and environmental studiesin microbiology. J. Bacteriol. 172, 762–770.
[7] Behrens, S., Fuchs, B., Mueller, L., Amann, R. (2003) Is the in situ accessibilityof the 16S rRNA of Escherichia coli for Cy3-labeled oligonucleotide probes pre-dicted by a three-dimensional structure model of the 30S ribosomal subunit?Appl. Environ. Microbiol. 69, 4935–4941.
[8] Behrens, S., Fuchs, B.M., Amann, R. (2004) The effect of nucleobase-specificfluorescence quenching on in situ hybridization with rRNA-targeted oligonu-cleotide probes. Syst. Appl. Microbiol. 27, 565–572.
[9] Behrens, S., Ruhland, C., Inacio, J., Huber, H., Fonseca, A., Spencer-Martins,I., Fuchs, B., Amann, R. (2003) In situ accessibility of small-subunit rRNAof members of the domains Bacteria, Archaea, and Eucarya to Cy3-labeledoligonucleotide probes. Appl. Environ. Microbiol. 69, 1748–1758.
10] Brosius, J., Dull, T.J., Sleeter, D.D., Noller, H.F. (1981) Gene organization andprimary structure of a ribosomal RNA operon from Escherichia coli. J. Mol. Biol.148, 107–127.
11] Christensen, R. 2002 Plane Answers to Complex Questions: The Theory of LinearModels, 3rd ed., Springer, New York.
12] Crockett, A.O., Wittwer, C.T. (2001) Fluorescein-labeled oligonucleotides forreal-time PCR: using the inherent quenching of deoxyguanosine nucleotides.Anal. Biochem. 290, 89–97.
13] DeLong, E., Wickham, G., Pace, N. (1989) Phylogenetic stains: ribosomal RNA-based probes for the identification of single cells. Science 243, 1360–1363.
14] Frischer, M.E., Floriani, P.J., Nierzwicki-Bauer, S.A. (1996) Differential sensitiv-ity of 16S rRNA targeted oligonucleotide probes used for fluorescence in situhybridization is a result of ribosomal higher order structure. Can. J. Microbiol.42, 1061–1071.
15] Fuchs, B.M., Wallner, G., Beisker, W., Schwippl, I., Ludwig, W., Amann, R. (1998)Flow cytometric analysis of the in situ accessibility of Escherichia coli 16S rRNA
for fluorescently labeled oligonucleotide probes. Appl. Environ. Microbiol. 64,4973–4982.16] Gamper, H.B., Cimino, G.D., Hearst, J.E. (1987) Solution hybridization ofcrosslinkable DNA oligonucleotides to bacteriophage M13 DNA. J. Mol. Biol.197, 349–362.
[
[
Microbiology 35 (2012) 485– 495 495
17] Hoshino, T., Yilmaz, L.S., Noguera, D.R., Daims, H., Wagner, M. (2008) Quantifi-cation of target molecules needed to detect microorganisms by fluorescencein situ hybridization (FISH) and catalyzed reporter deposition-FISH. Appl. Envi-ron. Microbiol. 74, 5068–5077.
18] Marras, S.A.E., Kramer, F.R., Tyagi, S. (2002) Efficiencies of fluorescence res-onance energy transfer and contact-mediated quenching in oligonucleotideprobes. Nucleic Acids Res. 30, e122.
19] Moreira, B.G., You, Y., Behlke, M.A., Owczarzy, R. (2005) Effects of fluorescentdyes, quenchers, and dangling ends on DNA duplex stability. Biochem. Biophys.Res. Commun. 327, 473–484.
20] Sistrom, W.R. (1965) Effect of oxygen on growth and the synthesisof bacteriochlorophyll in Rhodospirillum molischianum. J. Bacteriol. 89,403–408.
21] Stoecker, K., Dorninger, C., Daims, H., Wagner, M. (2010) Double labelingof oligonucleotide probes for fluorescence in situ hybridization (DOPE-FISH)improves signal intensity and increases rRNA accessibility. Appl. Environ.Microbiol. 76, 922–926.
22] Sugimoto, N., Nakano, S., Katoh, M., Matsumura, A., Nakamuta, H., Ohmichi,T., Yoneyama, M., Sasaki, M. (1995) Thermodynamic parameters to pre-dict stability of RNA/DNA hybrid duplexes. Biochemistry 34, 11211–11216.
23] Torimura, M., Kurata, S., Yamada, K., Yokomaku, T., Kamagata, Y., Kanagawa,T., Kurane, R. (2001) Fluorescence-quenching phenomenon by photoinducedelectron transfer between a fluorescent dye and a nucleotide base. Anal. Sci.17, 155–160.
24] Wagner, M., Horn, M., Daims, H. (2003) Fluorescent in situ hybridization forthe identification characterization of prokaryotes. Curr. Opin. Microbiol. 6,302–309.
25] Wallner, G., Amann, R., Beisker, W. (1993) Optimizing fluorescent in situhybridization with rRNA-targeted oligonucleotide probes for flow cytometricidentification of microorganisms. Cytometry 14, 136–143.
26] Wimberly, B.-T., Brodersen, D.-E., Clemons, V.-M.-J., Morgan-Warren, R., vonRhein, C., Hartsch, T., Ramakrishnan, V. (2000) Structure of the 30S ribosomalsubunit. Nature 407, 327–339.
27] Woese, C.R. (1987) Bacterial evolution. Microbiol. Rev. 51, 221–271.28] Yilmaz, L.S., Noguera, D.R. (2004) Mechanistic approach to the problem of
hybridization efficiency in fluorescent in situ hybridization. Appl. Environ.Microbiol. 70, 7126–7139.
29] Yilmaz, L.S., Noguera, D.R. (2007) Development of thermodynamic models forsimulating probe dissociation profiles in fluorescence in situ hybridization.Biotechnol. Bioeng. 96, 349–363.
30] Yilmaz, L.S., Okten, H.E., Noguera, D.R. (2006) Making all parts of the 16S rRNA ofEscherichia coli accessible in situ to single DNA oligonucleotides. Appl. Environ.Microbiol. 72, 733–744.
31] Yilmaz, L.S., Parnerkar, S., Noguera, D.R. (2011) mathFISH, a web tool thatuses thermodynamics-based mathematical models for in silico evaluation ofoligonucleotide probes for fluorescence in situ hybridization. Appl. Environ.
32] You, Y., Moreira, B.G., Behlke, M.A., Owczarzy, R. (2006) Design of LNA probesthat improve mismatch discrimination. Nucleic Acids Res. 34.
33] Zuker, M. (2003) Mfold web server for nucleic acid folding and hybridizationprediction. Nucleic Acids Res. 31, 3406–3415.
Related Documents











