Exploring spike transfer through the thalamus using hybrid artificial-biological neuronal networks Damien Debay a , Jakob Wolfart a , Yann Le Franc a,b , Gwendal Le Masson b , Thierry Bal a, * a Unite ´ de Neurosciences Inte ´gratives et Computationnelles (UNIC), CNRS UPR 2191, Institut de Neurobiologie Alfred Fessard, 1, avenue de la Terrasse, Bat. 33, 91198 Gif-sur-Yvette Cedex, France b INSERM E 358, Institut F. Magendie, 1, Rue Camille Saint Saens, 33077 Bordeaux Cedex, France Abstract We use dynamic clamp to construct ‘‘hybrid’’ thalamic circuits by connecting a biological neuron in situ to silicon- or software- generated ‘‘neurons’’ through artificial synapses. The purpose is to explore cellular sensory gating mechanisms that regulate the transfer efficiency of signals during different sleep-wake states. Hybrid technology is applied in vitro to different paradigms such as: (1) simulating interactions between biological thalamocortical neurons, artificial reticular thalamic inhibitory interneurons and a simulated sensory input, (2) grafting an artificial sensory input to a wholly biological thalamic network that generates spontaneous sleep-like oscillations, (3) injecting in thalamocortical neurons a background synaptic bombardment mimicking the activity of corticothalamic inputs. We show that the graded control of the strength of intrathalamic inhibition, combined with the membrane polarization and the fluctuating syn- aptic noise in thalamocortical neurons, is able to govern functional shifts between different input/output transmission states of the tha- lamic gate. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Dynamic clamp; Feedback inhibition; Noise; Corticothalamic; Sleep; Arousal 1. Introduction The notion that activity of thalamic cells is governed by a combination of state-dependent membrane properties and synaptic feedback loops and the conclusion that those form a basis for the Ôthalamic gateÕ function has been the subject of many review articles and hypotheses [69,67,56,59]. Surprisingly, only little direct data has been published concerning the mechanisms at the cellular level by which thalamic circuits may operate as a gate, or how this gate might be controlled by top–down corticothalamic inputs [42,67]. Although extracellular data is available [51,71], the relative rarity of intracellular data to support these hypotheses are due to the technological difficulties of recording thalamic neurons intracellularly in the beha- ving animal during sensory processing across transitions between sleep states, wake and attentiveness. It has been hypothesized that feedback inhibition within the thalamic circuitry could be a mean for decoupling the brain from sensory inputs at the onset of sleep [81,1,67]. Indeed a decrease of the synaptic response of thalamocortical neu- rons to afferent volleys occurs during the spindle-related inhibitory post-synaptic potentials (IPSPs) [67] or other phases of slow-wave-sleep [73]. The intrathalamic feedback loop formed between thalamocortical (TC) and thalamic reticular (nRt/PGN) cells is a likely candidate for such a function. GABAergic nRt/PGN interneurons provide a strategically located inhibitory recurrent feedback to the afferent sensory pathway, activated by collaterals of the thalamocortical projection. Firing intensity in this feed- back loop will modulate the strength of the inhibitory feed- back to TC cells [28]. This mechanism has been generalized 0928-4257/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jphysparis.2005.09.008 * Corresponding author. Tel.: +33 1 69 82 34 08; fax: +33 1 69 82 34 27. E-mail address: [email protected] (T. Bal). www.elsevier.com/locate/jphysparis Journal of Physiology - Paris 98 (2004) 540–558

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/jphysparis
Journal of Physiology - Paris 98 (2004) 540–558
Exploring spike transfer through the thalamus using hybridartificial-biological neuronal networks
Damien Debay a, Jakob Wolfart a, Yann Le Franc a,b, Gwendal Le Masson b, Thierry Bal a,*
a Unite de Neurosciences Integratives et Computationnelles (UNIC), CNRS UPR 2191, Institut de Neurobiologie Alfred Fessard,
1, avenue de la Terrasse, Bat. 33, 91198 Gif-sur-Yvette Cedex, Franceb INSERM E 358, Institut F. Magendie, 1, Rue Camille Saint Saens, 33077 Bordeaux Cedex, France
Abstract
We use dynamic clamp to construct ‘‘hybrid’’ thalamic circuits by connecting a biological neuron in situ to silicon- or software-generated ‘‘neurons’’ through artificial synapses. The purpose is to explore cellular sensory gating mechanisms that regulate the transferefficiency of signals during different sleep-wake states. Hybrid technology is applied in vitro to different paradigms such as: (1) simulatinginteractions between biological thalamocortical neurons, artificial reticular thalamic inhibitory interneurons and a simulated sensoryinput, (2) grafting an artificial sensory input to a wholly biological thalamic network that generates spontaneous sleep-like oscillations,(3) injecting in thalamocortical neurons a background synaptic bombardment mimicking the activity of corticothalamic inputs. We showthat the graded control of the strength of intrathalamic inhibition, combined with the membrane polarization and the fluctuating syn-aptic noise in thalamocortical neurons, is able to govern functional shifts between different input/output transmission states of the tha-lamic gate.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Dynamic clamp; Feedback inhibition; Noise; Corticothalamic; Sleep; Arousal
1. Introduction
The notion that activity of thalamic cells is governed bya combination of state-dependent membrane propertiesand synaptic feedback loops and the conclusion thatthose form a basis for the �thalamic gate� function has beenthe subject of many review articles and hypotheses[69,67,56,59]. Surprisingly, only little direct data has beenpublished concerning the mechanisms at the cellular levelby which thalamic circuits may operate as a gate, or howthis gate might be controlled by top–down corticothalamicinputs [42,67]. Although extracellular data is available[51,71], the relative rarity of intracellular data to supportthese hypotheses are due to the technological difficulties
0928-4257/$ - see front matter � 2005 Elsevier Ltd. All rights reserved.doi:10.1016/j.jphysparis.2005.09.008
* Corresponding author. Tel.: +33 1 69 82 34 08; fax: +33 1 69 82 34 27.E-mail address: [email protected] (T. Bal).
of recording thalamic neurons intracellularly in the beha-ving animal during sensory processing across transitionsbetween sleep states, wake and attentiveness. It has beenhypothesized that feedback inhibition within the thalamiccircuitry could be a mean for decoupling the brain fromsensory inputs at the onset of sleep [81,1,67]. Indeed adecrease of the synaptic response of thalamocortical neu-rons to afferent volleys occurs during the spindle-relatedinhibitory post-synaptic potentials (IPSPs) [67] or otherphases of slow-wave-sleep [73]. The intrathalamic feedbackloop formed between thalamocortical (TC) and thalamicreticular (nRt/PGN) cells is a likely candidate for such afunction. GABAergic nRt/PGN interneurons provide astrategically located inhibitory recurrent feedback to theafferent sensory pathway, activated by collaterals of thethalamocortical projection. Firing intensity in this feed-back loop will modulate the strength of the inhibitory feed-back to TC cells [28]. This mechanism has been generalized

D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 541
as a ‘‘variable gain regulator’’ hypothesis for modulatingthe retino-geniculate transmission of sensory informationto the cortex according to the level of arousal [1].
To address these questions in the more accessible experi-mental conditions in vitro, we have developed a new hybridtechnology based on dynamic clamp recording technique[52,54,55], allowing to mimic connections between identi-fied components of neural networks and realistic conduc-tance-based model neurons with a unique control ofsynaptic and membrane properties. This technology hasbeen tested by construction of a canonical retino-thalamichybrid network [31], for a review see [48], and applied toexamine the dynamics of temporally patterned synaptic sig-nal transfer through the thalamus and its modulation bycorticothalamic synaptic influences [79].
In a first part, we show in vitro, that spindle-like oscilla-tions generated in the biological thalamic network preventthe faithful transmission of pseudo-sensory inputs pro-duced either by the stimulation of retinal ganglion cellaxons in the optic tract or by purely artificial synapticinputs. In a second part we will detail the dynamics ofthe ‘‘sensory gating’’ process by the reconstruction of a‘‘hybrid retino-thalamic’’ circuit, made of biological andartificial neurons coupled together through artificial syn-apses. Finally we will extend these notions by introducingthe hypothesis that corticothalamic pathways may dynam-ically control the thalamic gate by means of ongoing synap-tic background activity.
2. Materials and methods
2.1. Biological preparations and recording techniques
Thalamocortical (TC) neurons were recorded at 34.5–36 �C in slices ofthe LGNd in guinea pig (1–4 months old) or ferrets (4–15 months old)maintained in an interface style recording chamber. Recording solutioncontained (in mM): NaCl, 126; KCl, 2.5; MgSO4, 1.2; NaH2PO4, 1.25;CaCl2, 2; NaHCO3, 26; dextrose, 10. Intracellular recordings were madewith glass micropipettes filled with 1.2 M K-acetate (90–100 MX after bev-elling) [2].
In the ferret, LGNd slices were cut in a plane parallel to the most proxi-mal extent of the optic tract (Fig. 1A). This procedure preserved a portionof the optic radiation (OR) containing corticothalamic axons and theirsynaptic connections to thalamic cells, as well as several millimeters ofthe optic tract (OT). Extracellular multiunit recordings from LGN lami-nae revealed periodic spontaneous spindle waves that were indistinguish-able from those obtained from sagittal slices used previously [77,4]. Thepresence of this network activity indicates that synaptic connectionsbetween perigeniculate and thalamocortical cells were functionally intact.The OR was stimulated at a distance of 400–800 lm from the PGN, usingbipolar tungsten electrodes similar to those used for extracellular record-ings, spaced 200–450 mm apart, and oriented perpendicular to the cortico-thalamic fiber bundles [2]. The OT was stimulated at a distance of up to 1–2 mm from the LGNd. Delayed stimuli of OT (see Fig. 4) were triggeredby the activity of thalamic relay cells using a custom data acquisition soft-ware (Acquis1/Elphy, developed by Sadoc, UNIC, Centre National de laRecherche Scientifique Gif-sur-Yvette, Agence Nationale pour la Valori-sation des Applications de la Recherche Biological). The software detectedthe TC discharges in intracellular recordings by a voltage threshold, andset the latency at which a command was sent to an OT stimulating unit
(A360; WPI). Stimulation intensities ranged from 5 to 50 lA (0.1 ms dura-tion). In ferret slices, a physical disconnection between PGN and LGNnuclei was performed by a blade cut under microscopic visual guidancein cold sucrose solution followed by 2 h of recovery in standard solutionsand temperature [2]. In contrast, in the guinea pig LGNd the slicing pro-cedure synaptically isolated TC cells from afferent (nRT or cortical)inputs. Ferret and guinea pig slices were used for the construction ofhybrid networks as indicated in figure legends. Data analysis was per-formed using Elphy.
2.2. Hybrid network method
Based on the dynamic-clamp principle [52,54,55], the hybrid methodallows to connect in real time, biological neurons with artificial neuronsvia modeled synapses. Running real time simulations in the hybrid net-work method requires that the time needed to solve the whole set of modelequations should be at least equal to the integration time step. This con-strain, essential for following the dynamics of living neurons can beachieved by using two different technical approaches. The first, describedin the present paper, is a numerical modeling approach based on two dif-ferent system prototypes where conductance-based models are software-implemented in a computer associated with a programmable digital signalprocessor (DSP) board. Synaptic input (background and signal) was gen-erated using either a first protoype of the hybrid system [30,31] or a morerecent prototype based upon a real time version (designed by Le Masson,Foutry, Le Franc and updated by Sadoc) of the NEURON simulationenvironment [23]. The hybrid RT-NEURON simulator runs in the Win-dows 2000 operating system (Microsoft) on a PC equipped with a PentiumIV processor. A PCI device, DSP board with 4 A/D and 4 D/A converters(M67, Innovative Instruments) is used to constrain the input/output signalprocessing and the simulation time to occur in real time by sending highpriority interruption request to the system. The DSP board allows input(for instance the Vm of the real cell incorporated in the equations of themodels) and output signals (the synaptic current to be injected into thecell) to be processed at regular intervals (dt = 0.1 ms or smaller). A cus-tom-made interface is used to connect the digital and analog inputs/out-puts signals of the DSP board with the intracellular amplifier and thedata acquisition system.
The second approach involves analog silicon integrated circuits(ASICs) which perform real time processing independently of the model�scomplexity. The analog implementation of model neurons on programma-ble microelectronic VLSI (very large-scale integration) circuits will not bedetailed here [32,31].
Data acquisition and current injections were done with an Axoclamp-2B amplifier (Axon instruments). Discontinuous current-clamp configura-tion was used for the dynamic clamp.
2.3. Models implementation
The formalism and the parameters describing the dynamics of cellularmodels (TC, nRt) and synaptic conductances (AMPA, GABAA andGABAB) were based on models by Destexhe et al. [17,15]. Details are in[31]. A NMDA receptor-mediated component of the retino-geniculateEPSP was not included in the present model based on the fact that NMDAreceptor activation is largely reduced at hyperpolarized membrane volt-ages. These receptors could nonetheless participate to the gating of visualtransmission for membrane voltage near spike threshold [53].
To mimic background synaptic bombardment we used independentexcitatory and inhibitory fluctuating conductances by generated a point-conductance model [19] representing a population of stochastically releas-ing synapses. The total conductance of synaptic background activity wasadjusted such that it reduced the input resistance of TC cells to approxi-mately 50%, similar to the effect of corticothalamic barrages in vivo[12]. Discrete inputs, different from the background activity, were also gen-erated in NEURON via a simulation of excitatory conductance (gAMPA)with random amplitudes at 5–20 Hz.
542 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
3. Results and discussion
3.1. The gating of input signals during sleep-like oscillation in
biological networks in vitro
A decade ago [77], we have shown that intracellular andextracellular multiple unit recordings from slices of the dor-sal lateral geniculate nucleus (LGNd) of ferrets revealedthe spontaneous occurrence of spindle waves. We thendemonstrated that spindle waves were endogenously gener-ated in the network by synchronized rebound burst firing inrelay cells. This rebound burst firing resulted from inhibi-tory post-synaptic potentials arriving from the perigenicu-late nucleus, the cells of which was activated in turn byburst firing of relay neurons. Spontaneous spindle wavegeneration was preserved, although with varying degreeof robustness, when slices were cut from different sagittal,coronal or horizontal angles although this occurred witha varying degree of robustness.
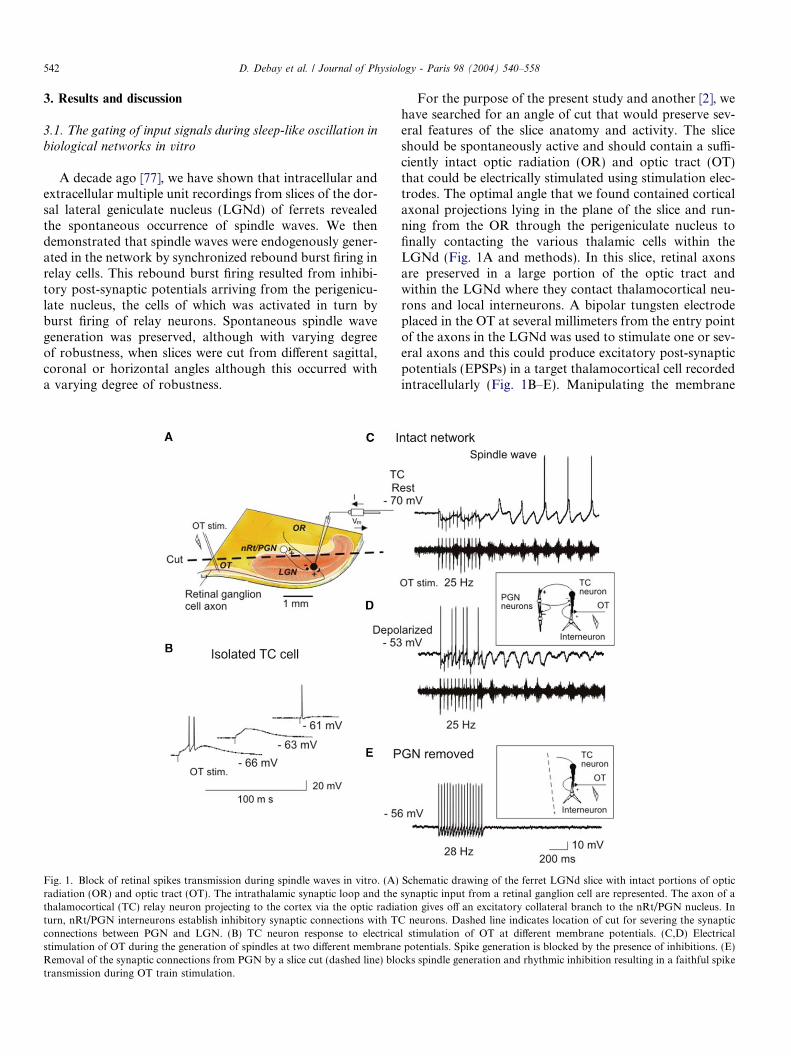
Fig. 1. Block of retinal spikes transmission during spindle waves in vitro. (A)radiation (OR) and optic tract (OT). The intrathalamic synaptic loop and thethalamocortical (TC) relay neuron projecting to the cortex via the optic radiaturn, nRt/PGN interneurons establish inhibitory synaptic connections with Tconnections between PGN and LGN. (B) TC neuron response to electricastimulation of OT during the generation of spindles at two different membraneRemoval of the synaptic connections from PGN by a slice cut (dashed line) blotransmission during OT train stimulation.
For the purpose of the present study and another [2], wehave searched for an angle of cut that would preserve sev-eral features of the slice anatomy and activity. The sliceshould be spontaneously active and should contain a suffi-ciently intact optic radiation (OR) and optic tract (OT)that could be electrically stimulated using stimulation elec-trodes. The optimal angle that we found contained corticalaxonal projections lying in the plane of the slice and run-ning from the OR through the perigeniculate nucleus tofinally contacting the various thalamic cells within theLGNd (Fig. 1A and methods). In this slice, retinal axonsare preserved in a large portion of the optic tract andwithin the LGNd where they contact thalamocortical neu-rons and local interneurons. A bipolar tungsten electrodeplaced in the OT at several millimeters from the entry pointof the axons in the LGNd was used to stimulate one or sev-eral axons and this could produce excitatory post-synapticpotentials (EPSPs) in a target thalamocortical cell recordedintracellularly (Fig. 1B–E). Manipulating the membrane
Schematic drawing of the ferret LGNd slice with intact portions of opticsynaptic input from a retinal ganglion cell are represented. The axon of ation gives off an excitatory collateral branch to the nRt/PGN nucleus. InC neurons. Dashed line indicates location of cut for severing the synapticl stimulation of OT at different membrane potentials. (C,D) Electricalpotentials. Spike generation is blocked by the presence of inhibitions. (E)cks spindle generation and rhythmic inhibition resulting in a faithful spike
A
B
C
D
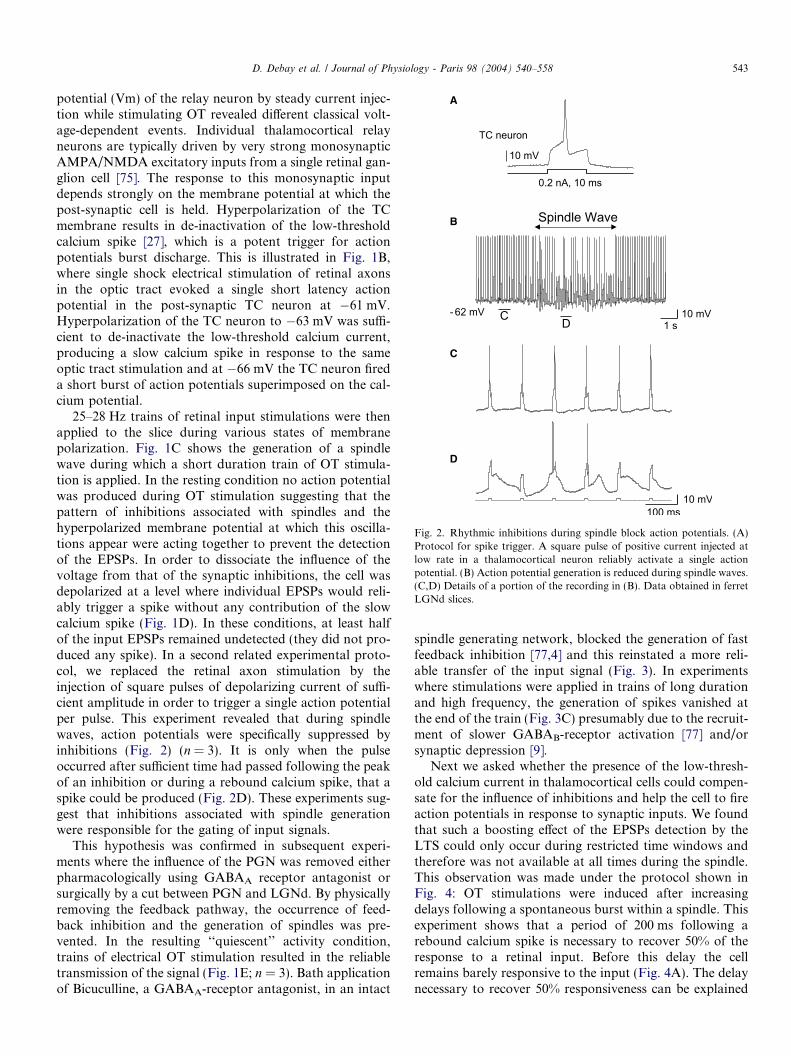
Fig. 2. Rhythmic inhibitions during spindle block action potentials. (A)Protocol for spike trigger. A square pulse of positive current injected atlow rate in a thalamocortical neuron reliably activate a single actionpotential. (B) Action potential generation is reduced during spindle waves.(C,D) Details of a portion of the recording in (B). Data obtained in ferretLGNd slices.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 543
potential (Vm) of the relay neuron by steady current injec-tion while stimulating OT revealed different classical volt-age-dependent events. Individual thalamocortical relayneurons are typically driven by very strong monosynapticAMPA/NMDA excitatory inputs from a single retinal gan-glion cell [75]. The response to this monosynaptic inputdepends strongly on the membrane potential at which thepost-synaptic cell is held. Hyperpolarization of the TCmembrane results in de-inactivation of the low-thresholdcalcium spike [27], which is a potent trigger for actionpotentials burst discharge. This is illustrated in Fig. 1B,where single shock electrical stimulation of retinal axonsin the optic tract evoked a single short latency actionpotential in the post-synaptic TC neuron at �61 mV.Hyperpolarization of the TC neuron to �63 mV was suffi-cient to de-inactivate the low-threshold calcium current,producing a slow calcium spike in response to the sameoptic tract stimulation and at �66 mV the TC neuron fireda short burst of action potentials superimposed on the cal-cium potential.
25–28 Hz trains of retinal input stimulations were thenapplied to the slice during various states of membranepolarization. Fig. 1C shows the generation of a spindlewave during which a short duration train of OT stimula-tion is applied. In the resting condition no action potentialwas produced during OT stimulation suggesting that thepattern of inhibitions associated with spindles and thehyperpolarized membrane potential at which this oscilla-tions appear were acting together to prevent the detectionof the EPSPs. In order to dissociate the influence of thevoltage from that of the synaptic inhibitions, the cell wasdepolarized at a level where individual EPSPs would reli-ably trigger a spike without any contribution of the slowcalcium spike (Fig. 1D). In these conditions, at least halfof the input EPSPs remained undetected (they did not pro-duced any spike). In a second related experimental proto-col, we replaced the retinal axon stimulation by theinjection of square pulses of depolarizing current of suffi-cient amplitude in order to trigger a single action potentialper pulse. This experiment revealed that during spindlewaves, action potentials were specifically suppressed byinhibitions (Fig. 2) (n = 3). It is only when the pulseoccurred after sufficient time had passed following the peakof an inhibition or during a rebound calcium spike, that aspike could be produced (Fig. 2D). These experiments sug-gest that inhibitions associated with spindle generationwere responsible for the gating of input signals.
This hypothesis was confirmed in subsequent experi-ments where the influence of the PGN was removed eitherpharmacologically using GABAA receptor antagonist orsurgically by a cut between PGN and LGNd. By physicallyremoving the feedback pathway, the occurrence of feed-back inhibition and the generation of spindles was pre-vented. In the resulting ‘‘quiescent’’ activity condition,trains of electrical OT stimulation resulted in the reliabletransmission of the signal (Fig. 1E; n = 3). Bath applicationof Bicuculline, a GABAA-receptor antagonist, in an intact
spindle generating network, blocked the generation of fastfeedback inhibition [77,4] and this reinstated a more reli-able transfer of the input signal (Fig. 3). In experimentswhere stimulations were applied in trains of long durationand high frequency, the generation of spikes vanished atthe end of the train (Fig. 3C) presumably due to the recruit-ment of slower GABAB-receptor activation [77] and/orsynaptic depression [9].
Next we asked whether the presence of the low-thresh-old calcium current in thalamocortical cells could compen-sate for the influence of inhibitions and help the cell to fireaction potentials in response to synaptic inputs. We foundthat such a boosting effect of the EPSPs detection by theLTS could only occur during restricted time windows andtherefore was not available at all times during the spindle.This observation was made under the protocol shown inFig. 4: OT stimulations were induced after increasingdelays following a spontaneous burst within a spindle. Thisexperiment shows that a period of 200 ms following arebound calcium spike is necessary to recover 50% of theresponse to a retinal input. Before this delay the cellremains barely responsive to the input (Fig. 4A). The delaynecessary to recover 50% responsiveness can be explained
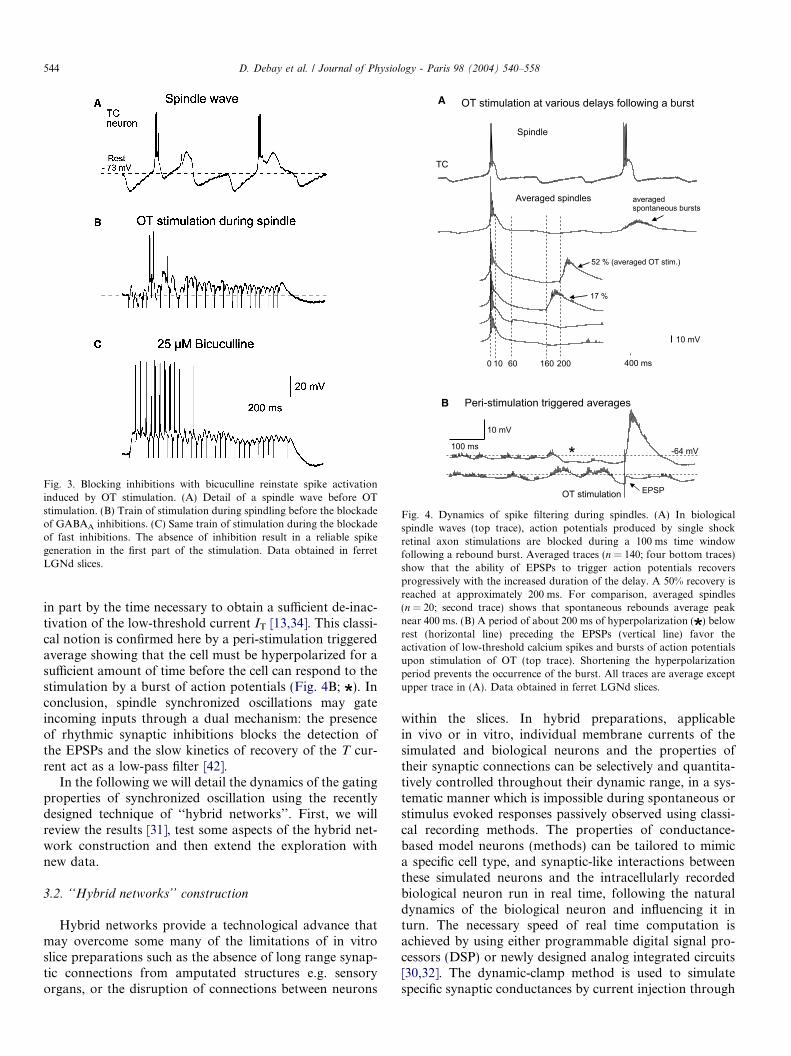
Fig. 3. Blocking inhibitions with bicuculline reinstate spike activationinduced by OT stimulation. (A) Detail of a spindle wave before OTstimulation. (B) Train of stimulation during spindling before the blockadeof GABAA inhibitions. (C) Same train of stimulation during the blockadeof fast inhibitions. The absence of inhibition result in a reliable spikegeneration in the first part of the stimulation. Data obtained in ferretLGNd slices.
A
B
Fig. 4. Dynamics of spike filtering during spindles. (A) In biologicalspindle waves (top trace), action potentials produced by single shockretinal axon stimulations are blocked during a 100 ms time windowfollowing a rebound burst. Averaged traces (n = 140; four bottom traces)show that the ability of EPSPs to trigger action potentials recoversprogressively with the increased duration of the delay. A 50% recovery isreached at approximately 200 ms. For comparison, averaged spindles(n = 20; second trace) shows that spontaneous rebounds average peaknear 400 ms. (B) A period of about 200 ms of hyperpolarization ( ) belowrest (horizontal line) preceding the EPSPs (vertical line) favor theactivation of low-threshold calcium spikes and bursts of action potentialsupon stimulation of OT (top trace). Shortening the hyperpolarizationperiod prevents the occurrence of the burst. All traces are average exceptupper trace in (A). Data obtained in ferret LGNd slices.
544 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
in part by the time necessary to obtain a sufficient de-inac-tivation of the low-threshold current IT [13,34]. This classi-cal notion is confirmed here by a peri-stimulation triggeredaverage showing that the cell must be hyperpolarized for asufficient amount of time before the cell can respond to thestimulation by a burst of action potentials (Fig. 4B; ). Inconclusion, spindle synchronized oscillations may gateincoming inputs through a dual mechanism: the presenceof rhythmic synaptic inhibitions blocks the detection ofthe EPSPs and the slow kinetics of recovery of the T cur-rent act as a low-pass filter [42].
In the following we will detail the dynamics of the gatingproperties of synchronized oscillation using the recentlydesigned technique of ‘‘hybrid networks’’. First, we willreview the results [31], test some aspects of the hybrid net-work construction and then extend the exploration withnew data.
3.2. ‘‘Hybrid networks’’ construction
Hybrid networks provide a technological advance thatmay overcome some many of the limitations of in vitroslice preparations such as the absence of long range synap-tic connections from amputated structures e.g. sensoryorgans, or the disruption of connections between neurons
within the slices. In hybrid preparations, applicablein vivo or in vitro, individual membrane currents of thesimulated and biological neurons and the properties oftheir synaptic connections can be selectively and quantita-tively controlled throughout their dynamic range, in a sys-tematic manner which is impossible during spontaneous orstimulus evoked responses passively observed using classi-cal recording methods. The properties of conductance-based model neurons (methods) can be tailored to mimica specific cell type, and synaptic-like interactions betweenthese simulated neurons and the intracellularly recordedbiological neuron run in real time, following the naturaldynamics of the biological neuron and influencing it inturn. The necessary speed of real time computation isachieved by using either programmable digital signal pro-cessors (DSP) or newly designed analog integrated circuits[30,32]. The dynamic-clamp method is used to simulatespecific synaptic conductances by current injection through
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 545
the intracellular recording pipette [52,54,55]. An importantspecificity of the present system is the modification ofNEURON as the core software for real time simulations(see Section 2).
A particularity of the synapses that form the intratha-lamic loop is the relative efficiency with which a singlePGN cell is able to trigger a rebound action potentialsresponse in the post-synaptic cell (Fig. 5B; upper trace)[28,41]. A post-hyperpolarization rebound burst of actionpotentials evoked in a single TC neuron following the
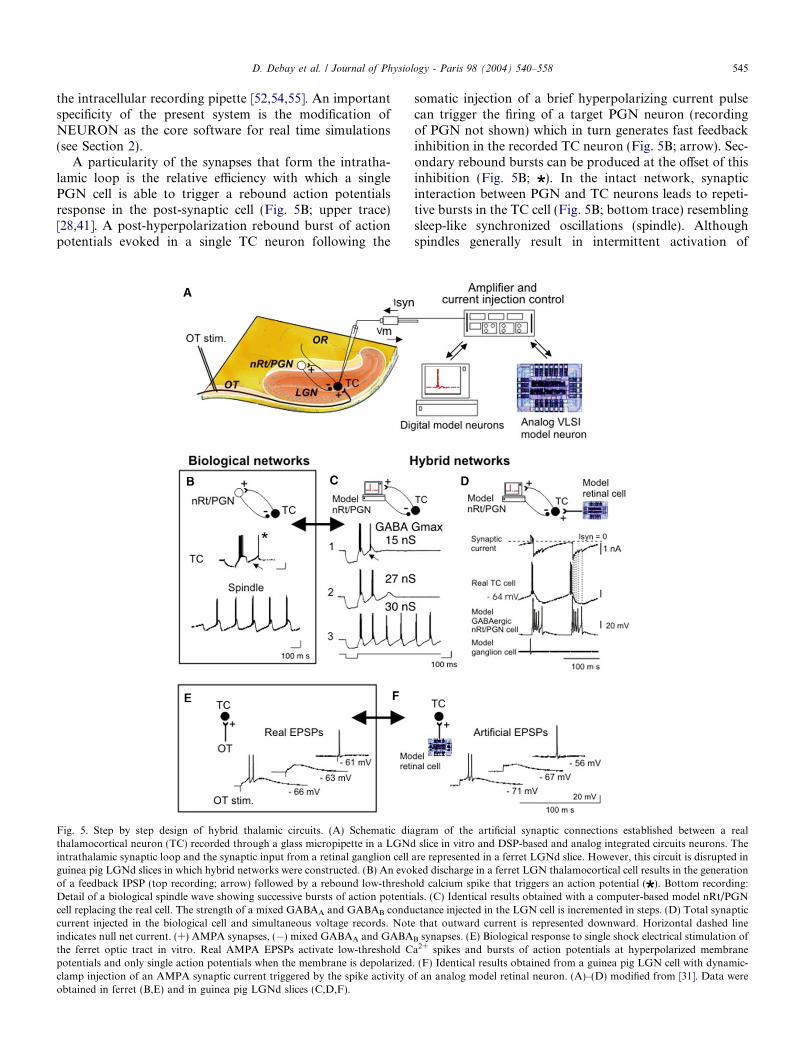
Fig. 5. Step by step design of hybrid thalamic circuits. (A) Schematic diathalamocortical neuron (TC) recorded through a glass micropipette in a LGNdintrathalamic synaptic loop and the synaptic input from a retinal ganglion cellguinea pig LGNd slices in which hybrid networks were constructed. (B) An evoof a feedback IPSP (top recording; arrow) followed by a rebound low-threshoDetail of a biological spindle wave showing successive bursts of action potenticell replacing the real cell. The strength of a mixed GABAA and GABAB conducurrent injected in the biological cell and simultaneous voltage records. Noteindicates null net current. (+) AMPA synapses, (�) mixed GABAA and GABAthe ferret optic tract in vitro. Real AMPA EPSPs activate low-threshold Cpotentials and only single action potentials when the membrane is depolarizedclamp injection of an AMPA synaptic current triggered by the spike activity oobtained in ferret (B,E) and in guinea pig LGNd slices (C,D,F).
somatic injection of a brief hyperpolarizing current pulsecan trigger the firing of a target PGN neuron (recordingof PGN not shown) which in turn generates fast feedbackinhibition in the recorded TC neuron (Fig. 5B; arrow). Sec-ondary rebound bursts can be produced at the offset of thisinhibition (Fig. 5B; ). In the intact network, synapticinteraction between PGN and TC neurons leads to repeti-tive bursts in the TC cell (Fig. 5B; bottom trace) resemblingsleep-like synchronized oscillations (spindle). Althoughspindles generally result in intermittent activation of
gram of the artificial synaptic connections established between a realslice in vitro and DSP-based and analog integrated circuits neurons. The
are represented in a ferret LGNd slice. However, this circuit is disrupted inked discharge in a ferret LGN thalamocortical cell results in the generationld calcium spike that triggers an action potential ( ). Bottom recording:als. (C) Identical results obtained with a computer-based model nRt/PGNctance injected in the LGN cell is incremented in steps. (D) Total synapticthat outward current is represented downward. Horizontal dashed line
B synapses. (E) Biological response to single shock electrical stimulation ofa2+ spikes and bursts of action potentials at hyperpolarized membrane. (F) Identical results obtained from a guinea pig LGN cell with dynamic-f an analog model retinal neuron. (A)–(D) modified from [31]. Data were

546 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
low-threshold calcium spikes and associated bursts ofaction potentials, in the hybrid circuit, rhythmic IPSPs acti-vate a rebound burst in the real cell at each cycle of spindleoscillation. Such periods of systematic rebound burstingare observed on occasion in the self-generating spindlingferret thalamic slice.
These basic properties can be reproduced in the hybridcircuit which allows full control of the conductances modu-lating synaptic strength (Fig. 5C) [31]. Progressive increaseof the maximal conductance (Gmax) of the nRt/PGN-to-TCGABAergic synapse led to an increasing probability ofrebound burst generation in the TC neuron (Fig. 5C). Sus-tained oscillations, resembling spindle waves, were obtainedfor a critical threshold value of GABA Gmax of approxi-mately 30 nS that showed little variation between recordedTC cells [31]. This remarkable constancy in the injected cur-rent value across hybrid circuit constructions using physi-cally different micropipettes and cells argues for thereliability of the technique. GABAergic inhibition can bemediated via both GABAA and GABAB receptor subtypes.The hybrid approach allows systematic screening of a con-ductance parameter space (here GABAA Gmax versusGABAB Gmax), and it was found that the optimal modelrepresentation reproducing intraspindle frequency andduration of natural spindles, was obtained when GABAA
and GABAB conductances were respectively 96% and 4%.This ratio is consistent with previous findings indicatingthat GABAA inhibition dominates during spindle waves[77,4,67]. Detailed traces, expanded in Fig. 5D, show howthe total synaptic current injected during a typical ‘‘hybridspindle’’ driven by the firing of a model nRt/PGN celland an analog retinal cell is integrated as compound IPSPsand EPSPs by the biological membrane of the TC cell.
The hybrid method was then used to reconstruct twoversions of a canonical intrathalamic circuit where the bio-logical neuron was replaced by the model neurons and viceversa. If the resulting inverted configurations would result
Fig. 6. Validation of the hybrid method using different circuit configurations. (biological nRt/PGN interneuron recorded intracellularly. The two cell circuitthe TC cell is real and the nRt/PGN is artificial results in a similar patternhyperpolarization in the TC cell. Data obtained in guinea pig LGNd slices.
in the emergence of similar global behaviors, then it wouldconfirm the accuracy of the method and the models. In thefirst circuit (Fig. 6A), a model thalamocortical (TC) neuronwas connected to a real perigeniculate inhibitory interneu-ron (nRt/PGN) and in the second (Fig. 6B), a real TC neu-ron was connected to a model nRt/PGN neuron. The twohybrid circuits had similar behaviour. The relative indepen-dence of the results from which neuron of the circuit isrecorded and which one is modeled, suggest that thismethod introduce a limited bias. This invariance of the pat-tern of discharge is a key validation protocol for the hybridmethod.
Taking another perspective, this procedure could applyto the refinement of theoretical models. Iterative simula-tions of reciprocal circuit configurations could be per-formed while exploring key points in the parameter spaceof the models until a realistic activity emerges. In this man-ner, the different biological cells types, involved in the mir-ror hybrid circuits, would serve as �teachers� to guide theadjustment of models neurons.
3.3. The gating of input signals explored in ‘‘hybridnetworks’’
The reconstruction of the retino-thalamic hybrid circuitwas then completed with the addition of either a biological(OT stimulation) or a modeled (dynamic clamp) retinalinput. Fig. 7 illustrates the response of a biological TC cellto the stimulation of the optic tract in a ferret slice. In thisexample the TC neuron is reciprocally connected with amodeled nRt–PGN neuron forming a hybrid circuit thatcan generate spindle oscillations as described above. Thestimulation of the optic tract at regular intervals could trig-ger spindles depending mainly upon the readiness of thebiological TC cell (see below); the readiness depends uponthe completion of a refractory period that limits the rate ofspindle occurrence [3]. Similarly to what was shown in the
A) A model TC neuron capable of endogenous oscillations interacts with aproduces spontaneous spindles. (B) Reciprocal circuit configuration whereof spindle oscillation. Here the circuit activity is initiated by a transient
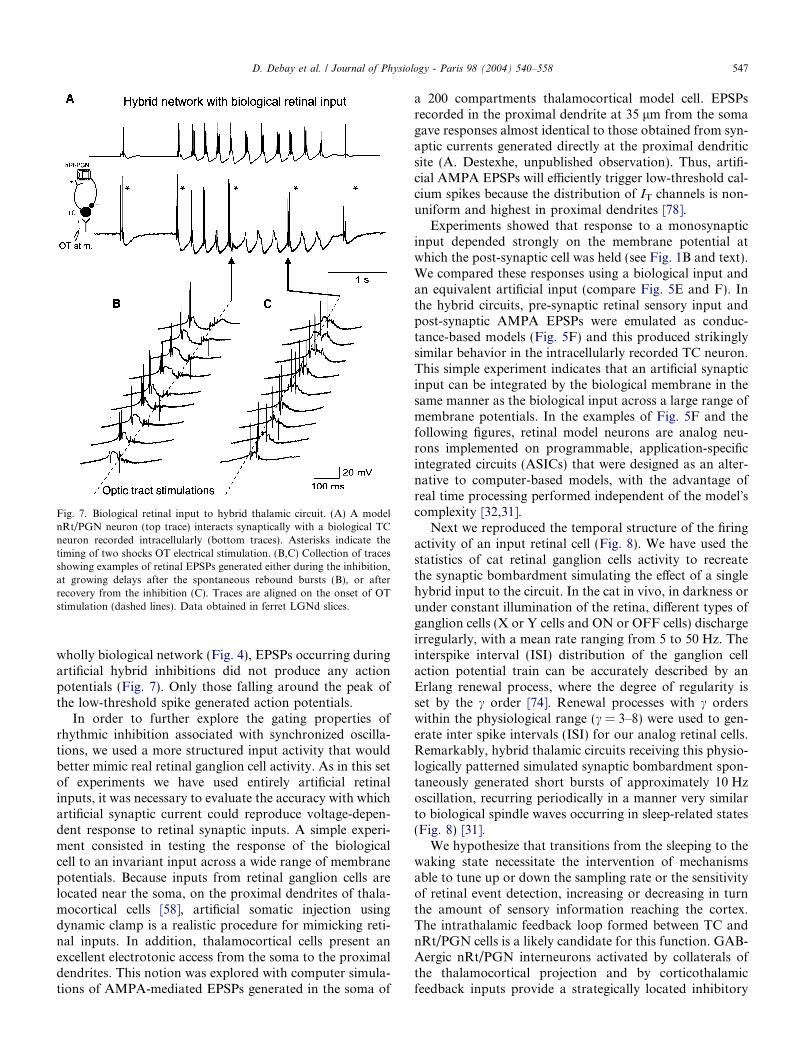
Fig. 7. Biological retinal input to hybrid thalamic circuit. (A) A modelnRt/PGN neuron (top trace) interacts synaptically with a biological TCneuron recorded intracellularly (bottom traces). Asterisks indicate thetiming of two shocks OT electrical stimulation. (B,C) Collection of tracesshowing examples of retinal EPSPs generated either during the inhibition,at growing delays after the spontaneous rebound bursts (B), or afterrecovery from the inhibition (C). Traces are aligned on the onset of OTstimulation (dashed lines). Data obtained in ferret LGNd slices.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 547
wholly biological network (Fig. 4), EPSPs occurring duringartificial hybrid inhibitions did not produce any actionpotentials (Fig. 7). Only those falling around the peak ofthe low-threshold spike generated action potentials.
In order to further explore the gating properties ofrhythmic inhibition associated with synchronized oscilla-tions, we used a more structured input activity that wouldbetter mimic real retinal ganglion cell activity. As in this setof experiments we have used entirely artificial retinalinputs, it was necessary to evaluate the accuracy with whichartificial synaptic current could reproduce voltage-depen-dent response to retinal synaptic inputs. A simple experi-ment consisted in testing the response of the biologicalcell to an invariant input across a wide range of membranepotentials. Because inputs from retinal ganglion cells arelocated near the soma, on the proximal dendrites of thala-mocortical cells [58], artificial somatic injection usingdynamic clamp is a realistic procedure for mimicking reti-nal inputs. In addition, thalamocortical cells present anexcellent electrotonic access from the soma to the proximaldendrites. This notion was explored with computer simula-tions of AMPA-mediated EPSPs generated in the soma of
a 200 compartments thalamocortical model cell. EPSPsrecorded in the proximal dendrite at 35 lm from the somagave responses almost identical to those obtained from syn-aptic currents generated directly at the proximal dendriticsite (A. Destexhe, unpublished observation). Thus, artifi-cial AMPA EPSPs will efficiently trigger low-threshold cal-cium spikes because the distribution of IT channels is non-uniform and highest in proximal dendrites [78].
Experiments showed that response to a monosynapticinput depended strongly on the membrane potential atwhich the post-synaptic cell was held (see Fig. 1B and text).We compared these responses using a biological input andan equivalent artificial input (compare Fig. 5E and F). Inthe hybrid circuits, pre-synaptic retinal sensory input andpost-synaptic AMPA EPSPs were emulated as conduc-tance-based models (Fig. 5F) and this produced strikinglysimilar behavior in the intracellularly recorded TC neuron.This simple experiment indicates that an artificial synapticinput can be integrated by the biological membrane in thesame manner as the biological input across a large range ofmembrane potentials. In the examples of Fig. 5F and thefollowing figures, retinal model neurons are analog neu-rons implemented on programmable, application-specificintegrated circuits (ASICs) that were designed as an alter-native to computer-based models, with the advantage ofreal time processing performed independent of the model�scomplexity [32,31].
Next we reproduced the temporal structure of the firingactivity of an input retinal cell (Fig. 8). We have used thestatistics of cat retinal ganglion cells activity to recreatethe synaptic bombardment simulating the effect of a singlehybrid input to the circuit. In the cat in vivo, in darkness orunder constant illumination of the retina, different types ofganglion cells (X or Y cells and ON or OFF cells) dischargeirregularly, with a mean rate ranging from 5 to 50 Hz. Theinterspike interval (ISI) distribution of the ganglion cellaction potential train can be accurately described by anErlang renewal process, where the degree of regularity isset by the c order [74]. Renewal processes with c orderswithin the physiological range (c = 3–8) were used to gen-erate inter spike intervals (ISI) for our analog retinal cells.Remarkably, hybrid thalamic circuits receiving this physio-logically patterned simulated synaptic bombardment spon-taneously generated short bursts of approximately 10 Hzoscillation, recurring periodically in a manner very similarto biological spindle waves occurring in sleep-related states(Fig. 8) [31].
We hypothesize that transitions from the sleeping to thewaking state necessitate the intervention of mechanismsable to tune up or down the sampling rate or the sensitivityof retinal event detection, increasing or decreasing in turnthe amount of sensory information reaching the cortex.The intrathalamic feedback loop formed between TC andnRt/PGN cells is a likely candidate for this function. GAB-Aergic nRt/PGN interneurons activated by collaterals ofthe thalamocortical projection and by corticothalamicfeedback inputs provide a strategically located inhibitory
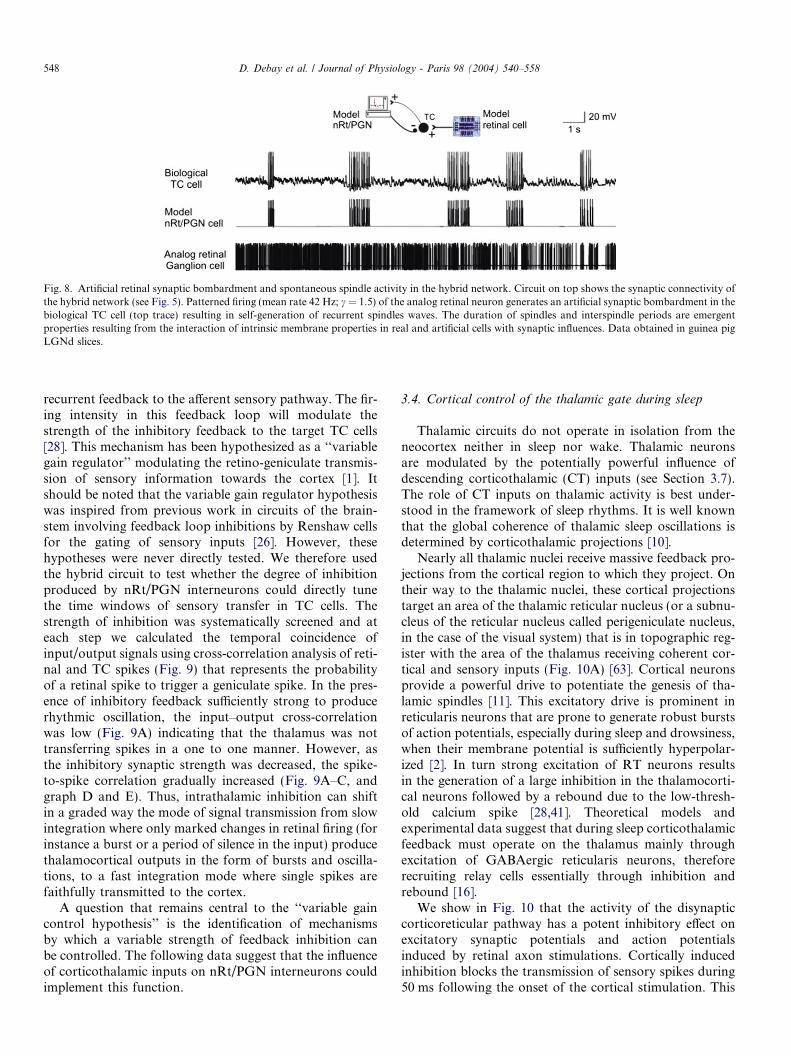
Fig. 8. Artificial retinal synaptic bombardment and spontaneous spindle activity in the hybrid network. Circuit on top shows the synaptic connectivity ofthe hybrid network (see Fig. 5). Patterned firing (mean rate 42 Hz; c = 1.5) of the analog retinal neuron generates an artificial synaptic bombardment in thebiological TC cell (top trace) resulting in self-generation of recurrent spindles waves. The duration of spindles and interspindle periods are emergentproperties resulting from the interaction of intrinsic membrane properties in real and artificial cells with synaptic influences. Data obtained in guinea pigLGNd slices.
548 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
recurrent feedback to the afferent sensory pathway. The fir-ing intensity in this feedback loop will modulate thestrength of the inhibitory feedback to the target TC cells[28]. This mechanism has been hypothesized as a ‘‘variablegain regulator’’ modulating the retino-geniculate transmis-sion of sensory information towards the cortex [1]. Itshould be noted that the variable gain regulator hypothesiswas inspired from previous work in circuits of the brain-stem involving feedback loop inhibitions by Renshaw cellsfor the gating of sensory inputs [26]. However, thesehypotheses were never directly tested. We therefore usedthe hybrid circuit to test whether the degree of inhibitionproduced by nRt/PGN interneurons could directly tunethe time windows of sensory transfer in TC cells. Thestrength of inhibition was systematically screened and ateach step we calculated the temporal coincidence ofinput/output signals using cross-correlation analysis of reti-nal and TC spikes (Fig. 9) that represents the probabilityof a retinal spike to trigger a geniculate spike. In the pres-ence of inhibitory feedback sufficiently strong to producerhythmic oscillation, the input–output cross-correlationwas low (Fig. 9A) indicating that the thalamus was nottransferring spikes in a one to one manner. However, asthe inhibitory synaptic strength was decreased, the spike-to-spike correlation gradually increased (Fig. 9A–C, andgraph D and E). Thus, intrathalamic inhibition can shiftin a graded way the mode of signal transmission from slowintegration where only marked changes in retinal firing (forinstance a burst or a period of silence in the input) producethalamocortical outputs in the form of bursts and oscilla-tions, to a fast integration mode where single spikes arefaithfully transmitted to the cortex.
A question that remains central to the ‘‘variable gaincontrol hypothesis’’ is the identification of mechanismsby which a variable strength of feedback inhibition canbe controlled. The following data suggest that the influenceof corticothalamic inputs on nRt/PGN interneurons couldimplement this function.
3.4. Cortical control of the thalamic gate during sleep
Thalamic circuits do not operate in isolation from theneocortex neither in sleep nor wake. Thalamic neuronsare modulated by the potentially powerful influence ofdescending corticothalamic (CT) inputs (see Section 3.7).The role of CT inputs on thalamic activity is best under-stood in the framework of sleep rhythms. It is well knownthat the global coherence of thalamic sleep oscillations isdetermined by corticothalamic projections [10].
Nearly all thalamic nuclei receive massive feedback pro-jections from the cortical region to which they project. Ontheir way to the thalamic nuclei, these cortical projectionstarget an area of the thalamic reticular nucleus (or a subnu-cleus of the reticular nucleus called perigeniculate nucleus,in the case of the visual system) that is in topographic reg-ister with the area of the thalamus receiving coherent cor-tical and sensory inputs (Fig. 10A) [63]. Cortical neuronsprovide a powerful drive to potentiate the genesis of tha-lamic spindles [11]. This excitatory drive is prominent inreticularis neurons that are prone to generate robust burstsof action potentials, especially during sleep and drowsiness,when their membrane potential is sufficiently hyperpolar-ized [2]. In turn strong excitation of RT neurons resultsin the generation of a large inhibition in the thalamocorti-cal neurons followed by a rebound due to the low-thresh-old calcium spike [28,41]. Theoretical models andexperimental data suggest that during sleep corticothalamicfeedback must operate on the thalamus mainly throughexcitation of GABAergic reticularis neurons, thereforerecruiting relay cells essentially through inhibition andrebound [16].
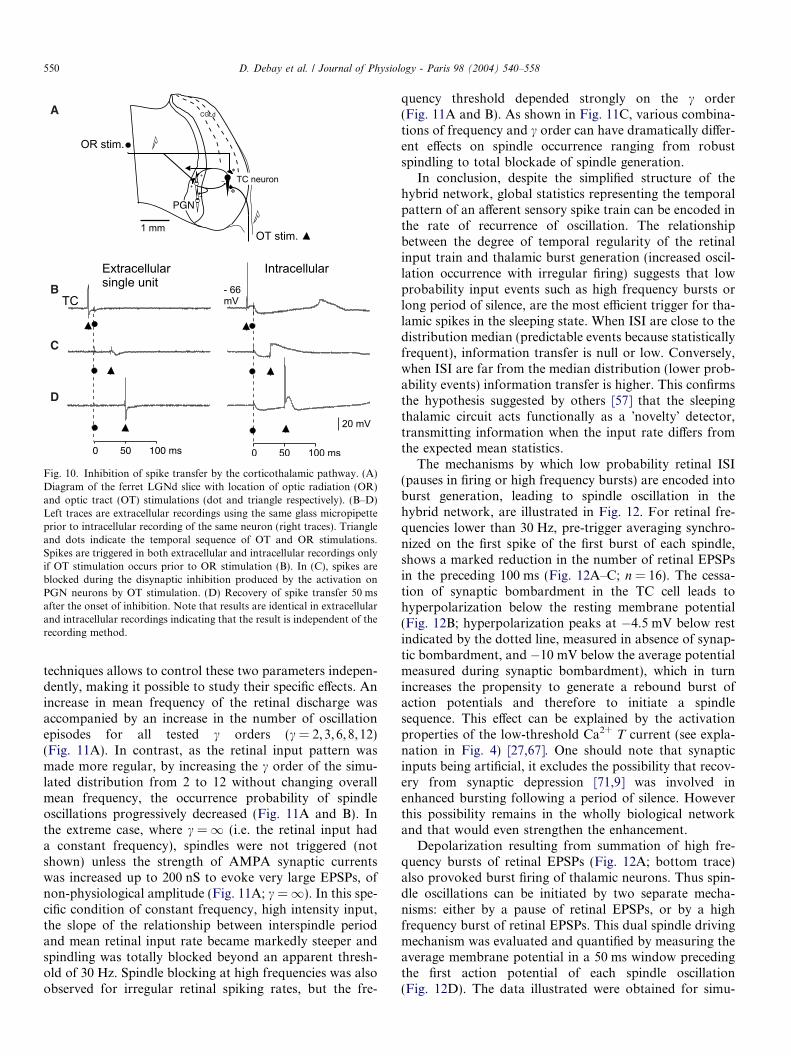
We show in Fig. 10 that the activity of the disynapticcorticoreticular pathway has a potent inhibitory effect onexcitatory synaptic potentials and action potentialsinduced by retinal axon stimulations. Cortically inducedinhibition blocks the transmission of sensory spikes during50 ms following the onset of the cortical stimulation. This
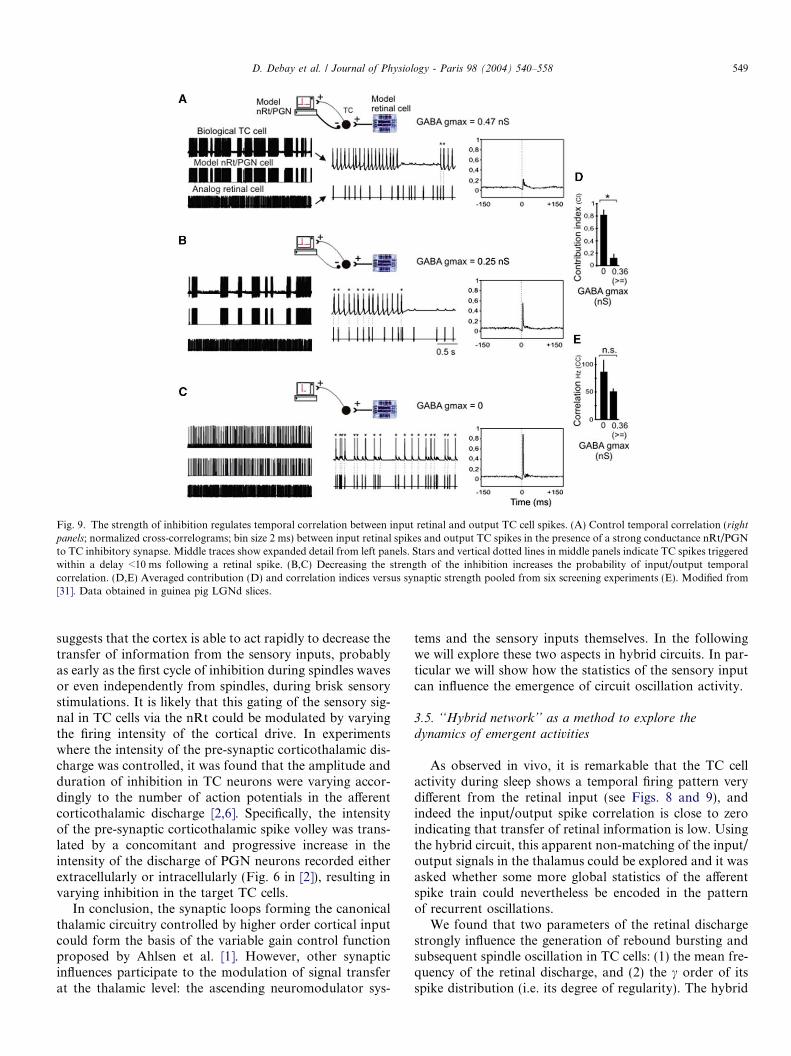
Fig. 9. The strength of inhibition regulates temporal correlation between input retinal and output TC cell spikes. (A) Control temporal correlation (rightpanels; normalized cross-correlograms; bin size 2 ms) between input retinal spikes and output TC spikes in the presence of a strong conductance nRt/PGNto TC inhibitory synapse. Middle traces show expanded detail from left panels. Stars and vertical dotted lines in middle panels indicate TC spikes triggeredwithin a delay <10 ms following a retinal spike. (B,C) Decreasing the strength of the inhibition increases the probability of input/output temporalcorrelation. (D,E) Averaged contribution (D) and correlation indices versus synaptic strength pooled from six screening experiments (E). Modified from[31]. Data obtained in guinea pig LGNd slices.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 549
suggests that the cortex is able to act rapidly to decrease thetransfer of information from the sensory inputs, probablyas early as the first cycle of inhibition during spindles wavesor even independently from spindles, during brisk sensorystimulations. It is likely that this gating of the sensory sig-nal in TC cells via the nRt could be modulated by varyingthe firing intensity of the cortical drive. In experimentswhere the intensity of the pre-synaptic corticothalamic dis-charge was controlled, it was found that the amplitude andduration of inhibition in TC neurons were varying accor-dingly to the number of action potentials in the afferentcorticothalamic discharge [2,6]. Specifically, the intensityof the pre-synaptic corticothalamic spike volley was trans-lated by a concomitant and progressive increase in theintensity of the discharge of PGN neurons recorded eitherextracellularly or intracellularly (Fig. 6 in [2]), resulting invarying inhibition in the target TC cells.
In conclusion, the synaptic loops forming the canonicalthalamic circuitry controlled by higher order cortical inputcould form the basis of the variable gain control functionproposed by Ahlsen et al. [1]. However, other synapticinfluences participate to the modulation of signal transferat the thalamic level: the ascending neuromodulator sys-
tems and the sensory inputs themselves. In the followingwe will explore these two aspects in hybrid circuits. In par-ticular we will show how the statistics of the sensory inputcan influence the emergence of circuit oscillation activity.
3.5. ‘‘Hybrid network’’ as a method to explore the
dynamics of emergent activities
As observed in vivo, it is remarkable that the TC cellactivity during sleep shows a temporal firing pattern verydifferent from the retinal input (see Figs. 8 and 9), andindeed the input/output spike correlation is close to zeroindicating that transfer of retinal information is low. Usingthe hybrid circuit, this apparent non-matching of the input/output signals in the thalamus could be explored and it wasasked whether some more global statistics of the afferentspike train could nevertheless be encoded in the patternof recurrent oscillations.
We found that two parameters of the retinal dischargestrongly influence the generation of rebound bursting andsubsequent spindle oscillation in TC cells: (1) the mean fre-quency of the retinal discharge, and (2) the c order of itsspike distribution (i.e. its degree of regularity). The hybrid
D
C
B
A
Fig. 10. Inhibition of spike transfer by the corticothalamic pathway. (A)Diagram of the ferret LGNd slice with location of optic radiation (OR)and optic tract (OT) stimulations (dot and triangle respectively). (B–D)Left traces are extracellular recordings using the same glass micropipetteprior to intracellular recording of the same neuron (right traces). Triangleand dots indicate the temporal sequence of OT and OR stimulations.Spikes are triggered in both extracellular and intracellular recordings onlyif OT stimulation occurs prior to OR stimulation (B). In (C), spikes areblocked during the disynaptic inhibition produced by the activation onPGN neurons by OT stimulation. (D) Recovery of spike transfer 50 msafter the onset of inhibition. Note that results are identical in extracellularand intracellular recordings indicating that the result is independent of therecording method.
550 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
techniques allows to control these two parameters indepen-dently, making it possible to study their specific effects. Anincrease in mean frequency of the retinal discharge wasaccompanied by an increase in the number of oscillationepisodes for all tested c orders (c = 2,3,6,8,12)(Fig. 11A). In contrast, as the retinal input pattern wasmade more regular, by increasing the c order of the simu-lated distribution from 2 to 12 without changing overallmean frequency, the occurrence probability of spindleoscillations progressively decreased (Fig. 11A and B). Inthe extreme case, where c = 1 (i.e. the retinal input hada constant frequency), spindles were not triggered (notshown) unless the strength of AMPA synaptic currentswas increased up to 200 nS to evoke very large EPSPs, ofnon-physiological amplitude (Fig. 11A; c =1). In this spe-cific condition of constant frequency, high intensity input,the slope of the relationship between interspindle periodand mean retinal input rate became markedly steeper andspindling was totally blocked beyond an apparent thresh-old of 30 Hz. Spindle blocking at high frequencies was alsoobserved for irregular retinal spiking rates, but the fre-
quency threshold depended strongly on the c order(Fig. 11A and B). As shown in Fig. 11C, various combina-tions of frequency and c order can have dramatically differ-ent effects on spindle occurrence ranging from robustspindling to total blockade of spindle generation.
In conclusion, despite the simplified structure of thehybrid network, global statistics representing the temporalpattern of an afferent sensory spike train can be encoded inthe rate of recurrence of oscillation. The relationshipbetween the degree of temporal regularity of the retinalinput train and thalamic burst generation (increased oscil-lation occurrence with irregular firing) suggests that lowprobability input events such as high frequency bursts orlong period of silence, are the most efficient trigger for tha-lamic spikes in the sleeping state. When ISI are close to thedistribution median (predictable events because statisticallyfrequent), information transfer is null or low. Conversely,when ISI are far from the median distribution (lower prob-ability events) information transfer is higher. This confirmsthe hypothesis suggested by others [57] that the sleepingthalamic circuit acts functionally as a �novelty� detector,transmitting information when the input rate differs fromthe expected mean statistics.
The mechanisms by which low probability retinal ISI(pauses in firing or high frequency bursts) are encoded intoburst generation, leading to spindle oscillation in thehybrid network, are illustrated in Fig. 12. For retinal fre-quencies lower than 30 Hz, pre-trigger averaging synchro-nized on the first spike of the first burst of each spindle,shows a marked reduction in the number of retinal EPSPsin the preceding 100 ms (Fig. 12A–C; n = 16). The cessa-tion of synaptic bombardment in the TC cell leads tohyperpolarization below the resting membrane potential(Fig. 12B; hyperpolarization peaks at �4.5 mV below restindicated by the dotted line, measured in absence of synap-tic bombardment, and �10 mV below the average potentialmeasured during synaptic bombardment), which in turnincreases the propensity to generate a rebound burst ofaction potentials and therefore to initiate a spindlesequence. This effect can be explained by the activationproperties of the low-threshold Ca2+ T current (see expla-nation in Fig. 4) [27,67]. One should note that synapticinputs being artificial, it excludes the possibility that recov-ery from synaptic depression [71,9] was involved inenhanced bursting following a period of silence. Howeverthis possibility remains in the wholly biological networkand that would even strengthen the enhancement.
Depolarization resulting from summation of high fre-quency bursts of retinal EPSPs (Fig. 12A; bottom trace)also provoked burst firing of thalamic neurons. Thus spin-dle oscillations can be initiated by two separate mecha-nisms: either by a pause of retinal EPSPs, or by a highfrequency burst of retinal EPSPs. This dual spindle drivingmechanism was evaluated and quantified by measuring theaverage membrane potential in a 50 ms window precedingthe first action potential of each spindle oscillation(Fig. 12D). The data illustrated were obtained for simu-
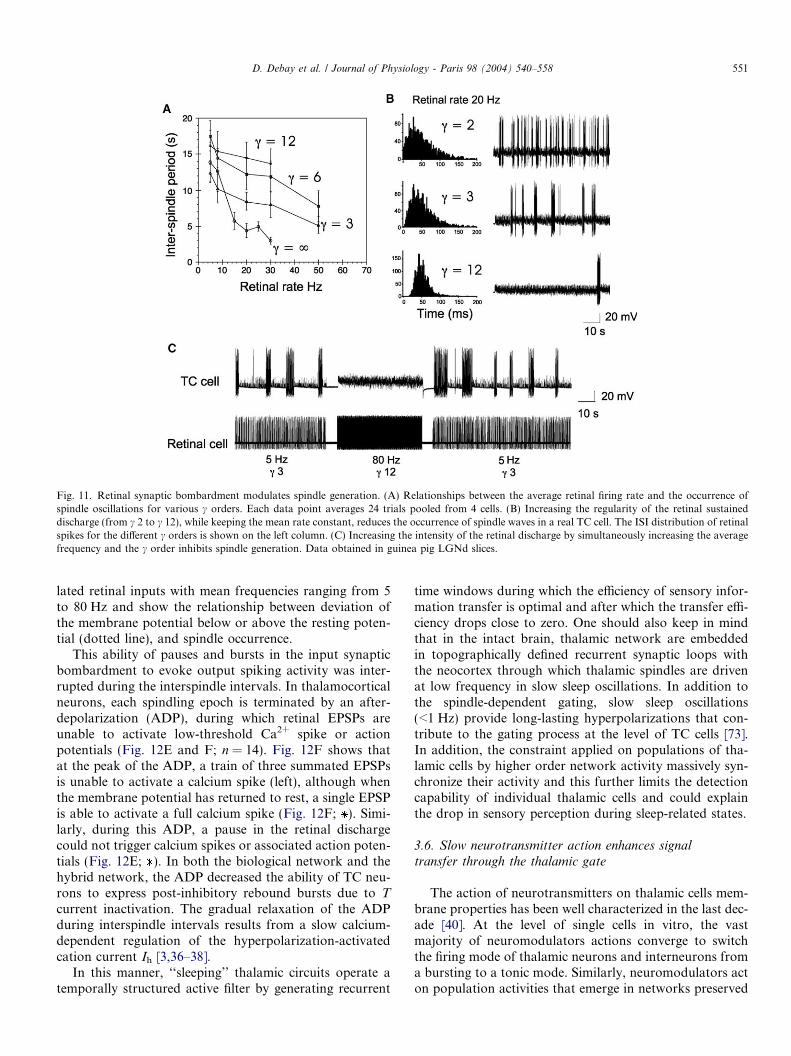
Fig. 11. Retinal synaptic bombardment modulates spindle generation. (A) Relationships between the average retinal firing rate and the occurrence ofspindle oscillations for various c orders. Each data point averages 24 trials pooled from 4 cells. (B) Increasing the regularity of the retinal sustaineddischarge (from c 2 to c 12), while keeping the mean rate constant, reduces the occurrence of spindle waves in a real TC cell. The ISI distribution of retinalspikes for the different c orders is shown on the left column. (C) Increasing the intensity of the retinal discharge by simultaneously increasing the averagefrequency and the c order inhibits spindle generation. Data obtained in guinea pig LGNd slices.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 551
lated retinal inputs with mean frequencies ranging from 5to 80 Hz and show the relationship between deviation ofthe membrane potential below or above the resting poten-tial (dotted line), and spindle occurrence.
This ability of pauses and bursts in the input synapticbombardment to evoke output spiking activity was inter-rupted during the interspindle intervals. In thalamocorticalneurons, each spindling epoch is terminated by an after-depolarization (ADP), during which retinal EPSPs areunable to activate low-threshold Ca2+ spike or actionpotentials (Fig. 12E and F; n = 14). Fig. 12F shows thatat the peak of the ADP, a train of three summated EPSPsis unable to activate a calcium spike (left), although whenthe membrane potential has returned to rest, a single EPSPis able to activate a full calcium spike (Fig. 12F; ). Simi-larly, during this ADP, a pause in the retinal dischargecould not trigger calcium spikes or associated action poten-tials (Fig. 12E; ). In both the biological network and thehybrid network, the ADP decreased the ability of TC neu-rons to express post-inhibitory rebound bursts due to T
current inactivation. The gradual relaxation of the ADPduring interspindle intervals results from a slow calcium-dependent regulation of the hyperpolarization-activatedcation current Ih [3,36–38].
In this manner, ‘‘sleeping’’ thalamic circuits operate atemporally structured active filter by generating recurrent
time windows during which the efficiency of sensory infor-mation transfer is optimal and after which the transfer effi-ciency drops close to zero. One should also keep in mindthat in the intact brain, thalamic network are embeddedin topographically defined recurrent synaptic loops withthe neocortex through which thalamic spindles are drivenat low frequency in slow sleep oscillations. In addition tothe spindle-dependent gating, slow sleep oscillations(<1 Hz) provide long-lasting hyperpolarizations that con-tribute to the gating process at the level of TC cells [73].In addition, the constraint applied on populations of tha-lamic cells by higher order network activity massively syn-chronize their activity and this further limits the detectioncapability of individual thalamic cells and could explainthe drop in sensory perception during sleep-related states.
3.6. Slow neurotransmitter action enhances signal
transfer through the thalamic gate
The action of neurotransmitters on thalamic cells mem-brane properties has been well characterized in the last dec-ade [40]. At the level of single cells in vitro, the vastmajority of neuromodulators actions converge to switchthe firing mode of thalamic neurons and interneurons froma bursting to a tonic mode. Similarly, neuromodulators acton population activities that emerge in networks preserved
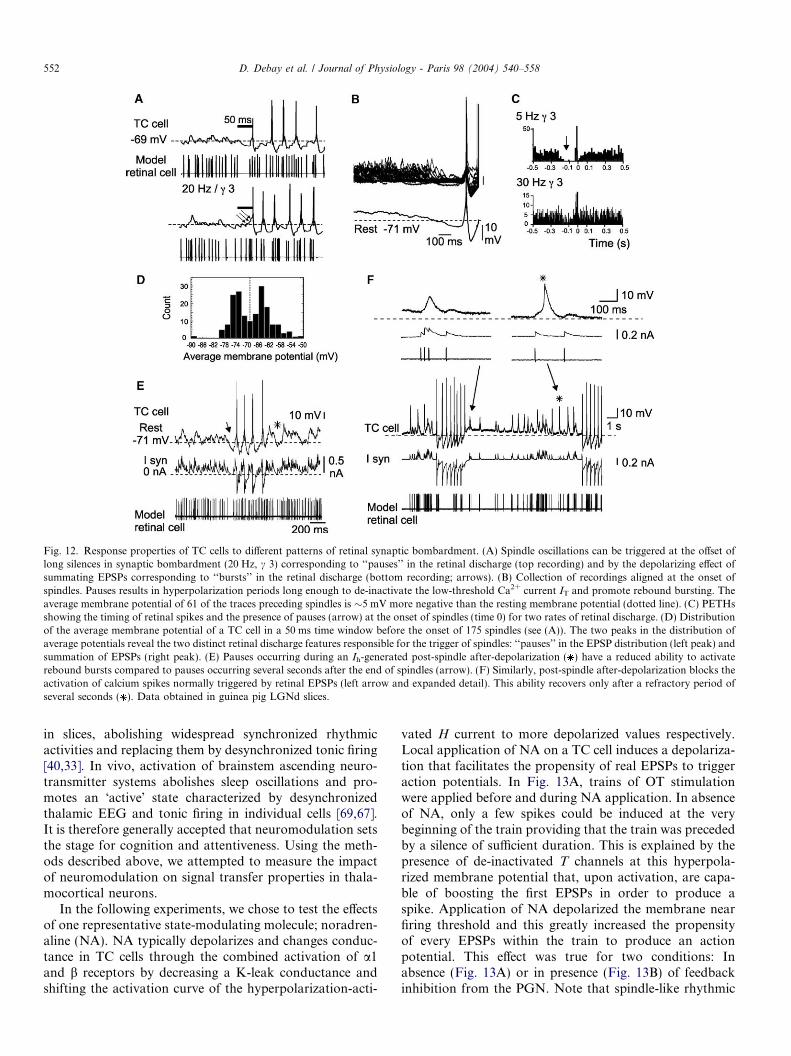
Fig. 12. Response properties of TC cells to different patterns of retinal synaptic bombardment. (A) Spindle oscillations can be triggered at the offset oflong silences in synaptic bombardment (20 Hz, c 3) corresponding to ‘‘pauses’’ in the retinal discharge (top recording) and by the depolarizing effect ofsummating EPSPs corresponding to ‘‘bursts’’ in the retinal discharge (bottom recording; arrows). (B) Collection of recordings aligned at the onset ofspindles. Pauses results in hyperpolarization periods long enough to de-inactivate the low-threshold Ca2+ current IT and promote rebound bursting. Theaverage membrane potential of 61 of the traces preceding spindles is �5 mV more negative than the resting membrane potential (dotted line). (C) PETHsshowing the timing of retinal spikes and the presence of pauses (arrow) at the onset of spindles (time 0) for two rates of retinal discharge. (D) Distributionof the average membrane potential of a TC cell in a 50 ms time window before the onset of 175 spindles (see (A)). The two peaks in the distribution ofaverage potentials reveal the two distinct retinal discharge features responsible for the trigger of spindles: ‘‘pauses’’ in the EPSP distribution (left peak) andsummation of EPSPs (right peak). (E) Pauses occurring during an Ih-generated post-spindle after-depolarization ( ) have a reduced ability to activaterebound bursts compared to pauses occurring several seconds after the end of spindles (arrow). (F) Similarly, post-spindle after-depolarization blocks theactivation of calcium spikes normally triggered by retinal EPSPs (left arrow and expanded detail). This ability recovers only after a refractory period ofseveral seconds ( ). Data obtained in guinea pig LGNd slices.
552 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
in slices, abolishing widespread synchronized rhythmicactivities and replacing them by desynchronized tonic firing[40,33]. In vivo, activation of brainstem ascending neuro-transmitter systems abolishes sleep oscillations and pro-motes an �active� state characterized by desynchronizedthalamic EEG and tonic firing in individual cells [69,67].It is therefore generally accepted that neuromodulation setsthe stage for cognition and attentiveness. Using the meth-ods described above, we attempted to measure the impactof neuromodulation on signal transfer properties in thala-mocortical neurons.
In the following experiments, we chose to test the effectsof one representative state-modulating molecule; noradren-aline (NA). NA typically depolarizes and changes conduc-tance in TC cells through the combined activation of a1and b receptors by decreasing a K-leak conductance andshifting the activation curve of the hyperpolarization-acti-
vated H current to more depolarized values respectively.Local application of NA on a TC cell induces a depolariza-tion that facilitates the propensity of real EPSPs to triggeraction potentials. In Fig. 13A, trains of OT stimulationwere applied before and during NA application. In absenceof NA, only a few spikes could be induced at the verybeginning of the train providing that the train was precededby a silence of sufficient duration. This is explained by thepresence of de-inactivated T channels at this hyperpola-rized membrane potential that, upon activation, are capa-ble of boosting the first EPSPs in order to produce aspike. Application of NA depolarized the membrane nearfiring threshold and this greatly increased the propensityof every EPSPs within the train to produce an actionpotential. This effect was true for two conditions: Inabsence (Fig. 13A) or in presence (Fig. 13B) of feedbackinhibition from the PGN. Note that spindle-like rhythmic
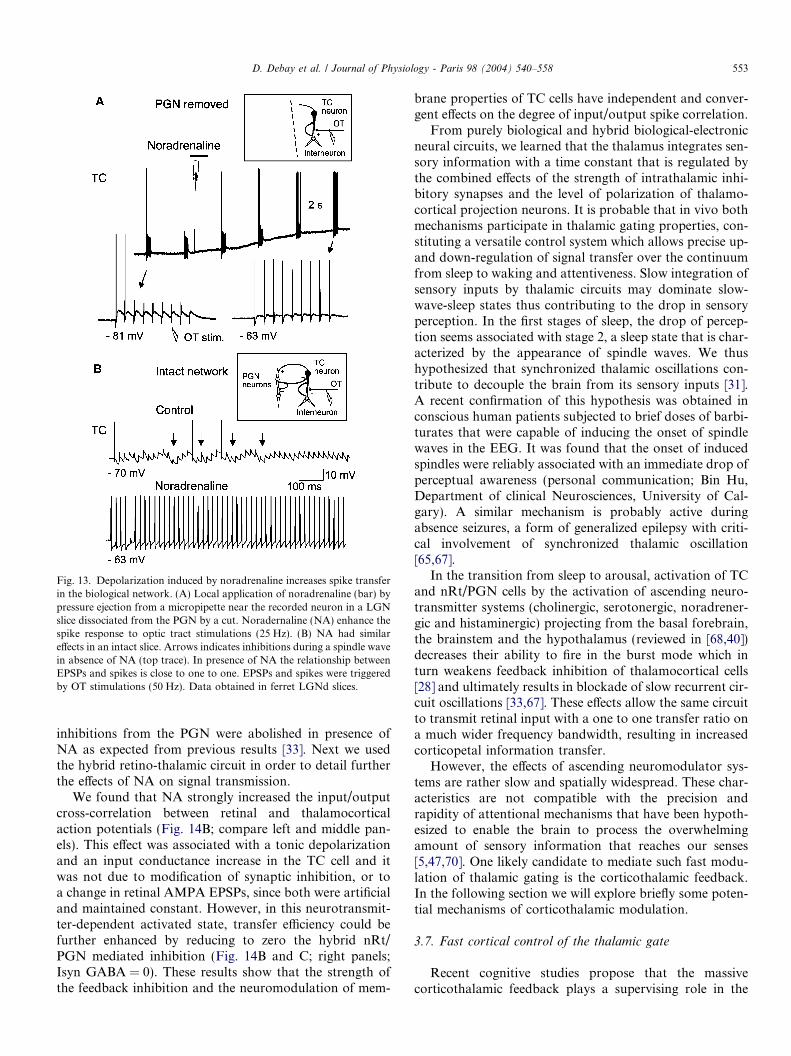
Fig. 13. Depolarization induced by noradrenaline increases spike transferin the biological network. (A) Local application of noradrenaline (bar) bypressure ejection from a micropipette near the recorded neuron in a LGNslice dissociated from the PGN by a cut. Noradernaline (NA) enhance thespike response to optic tract stimulations (25 Hz). (B) NA had similareffects in an intact slice. Arrows indicates inhibitions during a spindle wavein absence of NA (top trace). In presence of NA the relationship betweenEPSPs and spikes is close to one to one. EPSPs and spikes were triggeredby OT stimulations (50 Hz). Data obtained in ferret LGNd slices.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 553
inhibitions from the PGN were abolished in presence ofNA as expected from previous results [33]. Next we usedthe hybrid retino-thalamic circuit in order to detail furtherthe effects of NA on signal transmission.
We found that NA strongly increased the input/outputcross-correlation between retinal and thalamocorticalaction potentials (Fig. 14B; compare left and middle pan-els). This effect was associated with a tonic depolarizationand an input conductance increase in the TC cell and itwas not due to modification of synaptic inhibition, or toa change in retinal AMPA EPSPs, since both were artificialand maintained constant. However, in this neurotransmit-ter-dependent activated state, transfer efficiency could befurther enhanced by reducing to zero the hybrid nRt/PGN mediated inhibition (Fig. 14B and C; right panels;Isyn GABA = 0). These results show that the strength ofthe feedback inhibition and the neuromodulation of mem-
brane properties of TC cells have independent and conver-gent effects on the degree of input/output spike correlation.
From purely biological and hybrid biological-electronicneural circuits, we learned that the thalamus integrates sen-sory information with a time constant that is regulated bythe combined effects of the strength of intrathalamic inhi-bitory synapses and the level of polarization of thalamo-cortical projection neurons. It is probable that in vivo bothmechanisms participate in thalamic gating properties, con-stituting a versatile control system which allows precise up-and down-regulation of signal transfer over the continuumfrom sleep to waking and attentiveness. Slow integration ofsensory inputs by thalamic circuits may dominate slow-wave-sleep states thus contributing to the drop in sensoryperception. In the first stages of sleep, the drop of percep-tion seems associated with stage 2, a sleep state that is char-acterized by the appearance of spindle waves. We thushypothesized that synchronized thalamic oscillations con-tribute to decouple the brain from its sensory inputs [31].A recent confirmation of this hypothesis was obtained inconscious human patients subjected to brief doses of barbi-turates that were capable of inducing the onset of spindlewaves in the EEG. It was found that the onset of inducedspindles were reliably associated with an immediate drop ofperceptual awareness (personal communication; Bin Hu,Department of clinical Neurosciences, University of Cal-gary). A similar mechanism is probably active duringabsence seizures, a form of generalized epilepsy with criti-cal involvement of synchronized thalamic oscillation[65,67].
In the transition from sleep to arousal, activation of TCand nRt/PGN cells by the activation of ascending neuro-transmitter systems (cholinergic, serotonergic, noradrener-gic and histaminergic) projecting from the basal forebrain,the brainstem and the hypothalamus (reviewed in [68,40])decreases their ability to fire in the burst mode which inturn weakens feedback inhibition of thalamocortical cells[28] and ultimately results in blockade of slow recurrent cir-cuit oscillations [33,67]. These effects allow the same circuitto transmit retinal input with a one to one transfer ratio ona much wider frequency bandwidth, resulting in increasedcorticopetal information transfer.
However, the effects of ascending neuromodulator sys-tems are rather slow and spatially widespread. These char-acteristics are not compatible with the precision andrapidity of attentional mechanisms that have been hypoth-esized to enable the brain to process the overwhelmingamount of sensory information that reaches our senses[5,47,70]. One likely candidate to mediate such fast modu-lation of thalamic gating is the corticothalamic feedback.In the following section we will explore briefly some poten-tial mechanisms of corticothalamic modulation.
3.7. Fast cortical control of the thalamic gate
Recent cognitive studies propose that the massivecorticothalamic feedback plays a supervising role in the
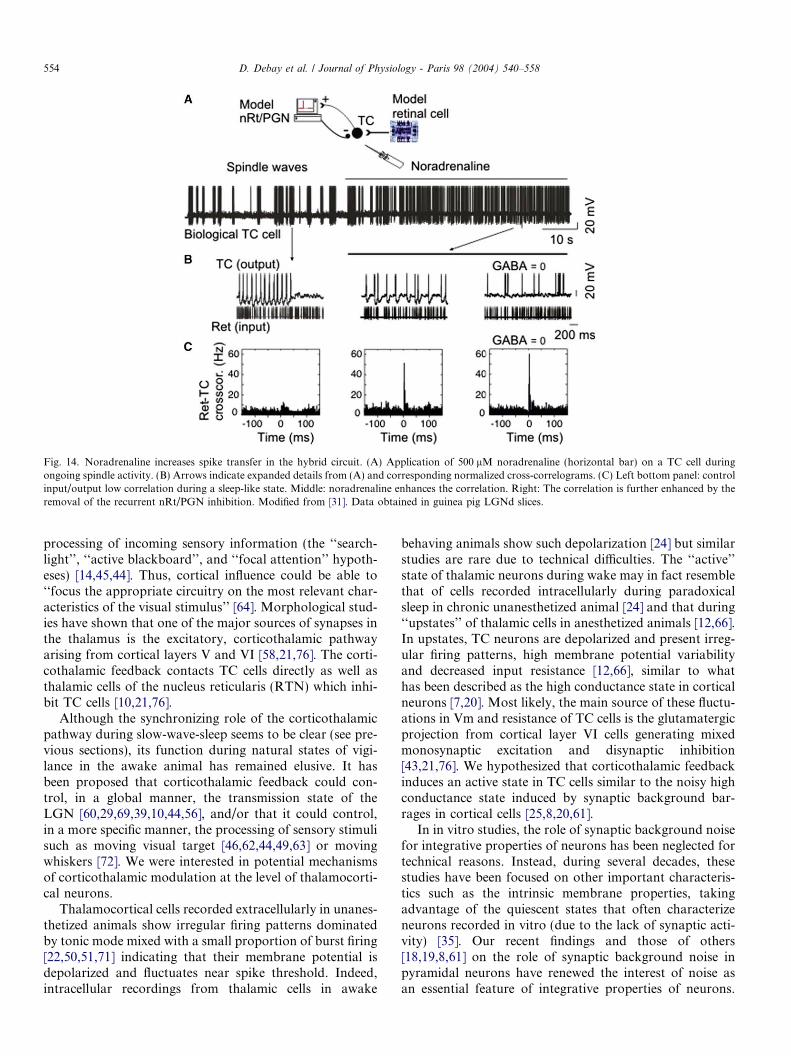
Fig. 14. Noradrenaline increases spike transfer in the hybrid circuit. (A) Application of 500 lM noradrenaline (horizontal bar) on a TC cell duringongoing spindle activity. (B) Arrows indicate expanded details from (A) and corresponding normalized cross-correlograms. (C) Left bottom panel: controlinput/output low correlation during a sleep-like state. Middle: noradrenaline enhances the correlation. Right: The correlation is further enhanced by theremoval of the recurrent nRt/PGN inhibition. Modified from [31]. Data obtained in guinea pig LGNd slices.
554 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
processing of incoming sensory information (the ‘‘search-light’’, ‘‘active blackboard’’, and ‘‘focal attention’’ hypoth-eses) [14,45,44]. Thus, cortical influence could be able to‘‘focus the appropriate circuitry on the most relevant char-acteristics of the visual stimulus’’ [64]. Morphological stud-ies have shown that one of the major sources of synapses inthe thalamus is the excitatory, corticothalamic pathwayarising from cortical layers V and VI [58,21,76]. The corti-cothalamic feedback contacts TC cells directly as well asthalamic cells of the nucleus reticularis (RTN) which inhi-bit TC cells [10,21,76].
Although the synchronizing role of the corticothalamicpathway during slow-wave-sleep seems to be clear (see pre-vious sections), its function during natural states of vigi-lance in the awake animal has remained elusive. It hasbeen proposed that corticothalamic feedback could con-trol, in a global manner, the transmission state of theLGN [60,29,69,39,10,44,56], and/or that it could control,in a more specific manner, the processing of sensory stimulisuch as moving visual target [46,62,44,49,63] or movingwhiskers [72]. We were interested in potential mechanismsof corticothalamic modulation at the level of thalamocorti-cal neurons.
Thalamocortical cells recorded extracellularly in unanes-thetized animals show irregular firing patterns dominatedby tonic mode mixed with a small proportion of burst firing[22,50,51,71] indicating that their membrane potential isdepolarized and fluctuates near spike threshold. Indeed,intracellular recordings from thalamic cells in awake
behaving animals show such depolarization [24] but similarstudies are rare due to technical difficulties. The ‘‘active’’state of thalamic neurons during wake may in fact resemblethat of cells recorded intracellularly during paradoxicalsleep in chronic unanesthetized animal [24] and that during‘‘upstates’’ of thalamic cells in anesthetized animals [12,66].In upstates, TC neurons are depolarized and present irreg-ular firing patterns, high membrane potential variabilityand decreased input resistance [12,66], similar to whathas been described as the high conductance state in corticalneurons [7,20]. Most likely, the main source of these fluctu-ations in Vm and resistance of TC cells is the glutamatergicprojection from cortical layer VI cells generating mixedmonosynaptic excitation and disynaptic inhibition[43,21,76]. We hypothesized that corticothalamic feedbackinduces an active state in TC cells similar to the noisy highconductance state induced by synaptic background bar-rages in cortical cells [25,8,20,61].
In in vitro studies, the role of synaptic background noisefor integrative properties of neurons has been neglected fortechnical reasons. Instead, during several decades, thesestudies have been focused on other important characteris-tics such as the intrinsic membrane properties, takingadvantage of the quiescent states that often characterizeneurons recorded in vitro (due to the lack of synaptic acti-vity) [35]. Our recent findings and those of others[18,19,8,61] on the role of synaptic background noise inpyramidal neurons have renewed the interest of noise asan essential feature of integrative properties of neurons.
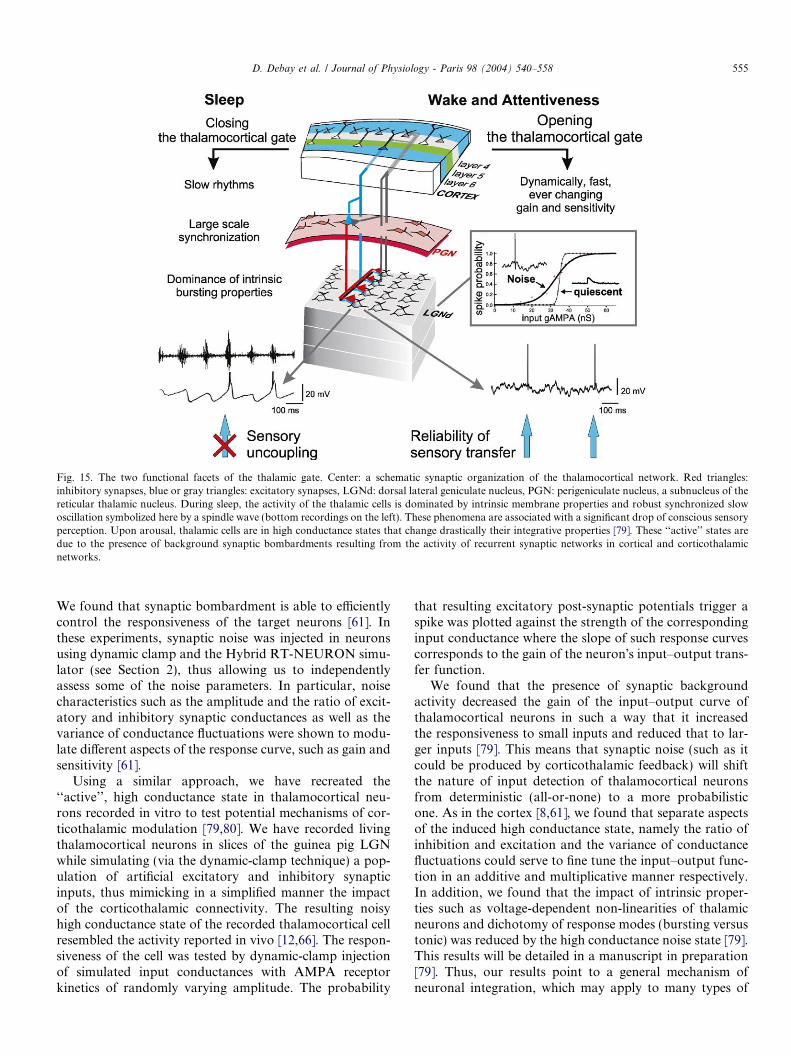
Fig. 15. The two functional facets of the thalamic gate. Center: a schematic synaptic organization of the thalamocortical network. Red triangles:inhibitory synapses, blue or gray triangles: excitatory synapses, LGNd: dorsal lateral geniculate nucleus, PGN: perigeniculate nucleus, a subnucleus of thereticular thalamic nucleus. During sleep, the activity of the thalamic cells is dominated by intrinsic membrane properties and robust synchronized slowoscillation symbolized here by a spindle wave (bottom recordings on the left). These phenomena are associated with a significant drop of conscious sensoryperception. Upon arousal, thalamic cells are in high conductance states that change drastically their integrative properties [79]. These ‘‘active’’ states aredue to the presence of background synaptic bombardments resulting from the activity of recurrent synaptic networks in cortical and corticothalamicnetworks.
D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 555
We found that synaptic bombardment is able to efficientlycontrol the responsiveness of the target neurons [61]. Inthese experiments, synaptic noise was injected in neuronsusing dynamic clamp and the Hybrid RT-NEURON simu-lator (see Section 2), thus allowing us to independentlyassess some of the noise parameters. In particular, noisecharacteristics such as the amplitude and the ratio of excit-atory and inhibitory synaptic conductances as well as thevariance of conductance fluctuations were shown to modu-late different aspects of the response curve, such as gain andsensitivity [61].
Using a similar approach, we have recreated the‘‘active’’, high conductance state in thalamocortical neu-rons recorded in vitro to test potential mechanisms of cor-ticothalamic modulation [79,80]. We have recorded livingthalamocortical neurons in slices of the guinea pig LGNwhile simulating (via the dynamic-clamp technique) a pop-ulation of artificial excitatory and inhibitory synapticinputs, thus mimicking in a simplified manner the impactof the corticothalamic connectivity. The resulting noisyhigh conductance state of the recorded thalamocortical cellresembled the activity reported in vivo [12,66]. The respon-siveness of the cell was tested by dynamic-clamp injectionof simulated input conductances with AMPA receptorkinetics of randomly varying amplitude. The probability
that resulting excitatory post-synaptic potentials trigger aspike was plotted against the strength of the correspondinginput conductance where the slope of such response curvescorresponds to the gain of the neuron�s input–output trans-fer function.
We found that the presence of synaptic backgroundactivity decreased the gain of the input–output curve ofthalamocortical neurons in such a way that it increasedthe responsiveness to small inputs and reduced that to lar-ger inputs [79]. This means that synaptic noise (such as itcould be produced by corticothalamic feedback) will shiftthe nature of input detection of thalamocortical neuronsfrom deterministic (all-or-none) to a more probabilisticone. As in the cortex [8,61], we found that separate aspectsof the induced high conductance state, namely the ratio ofinhibition and excitation and the variance of conductancefluctuations could serve to fine tune the input–output func-tion in an additive and multiplicative manner respectively.In addition, we found that the impact of intrinsic proper-ties such as voltage-dependent non-linearities of thalamicneurons and dichotomy of response modes (bursting versustonic) was reduced by the high conductance noise state [79].This results will be detailed in a manuscript in preparation[79]. Thus, our results point to a general mechanism ofneuronal integration, which may apply to many types of

556 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
neurons: the ‘‘noise mode’’, characterized by probabilisticinput–output transfer and reduced impact of intrinsicproperties. In particular, noise could be a mechanism bywhich corticothalamic feedback may modulate the tha-lamic relay of sensory information.
3.8. A coherent vision of the state-dependent thalamic
transfer function
In conclusion, the present work and that of others onthe transfer function of thalamocortical relay neurons leadsto a coherent vision based on the principles summarized inFig. 15. The decrease of conscious sensory perception atthe onset of sleep and during absence seizures may resultin part from a functional decoupling of the sensory flowduring slow synchronous oscillations reverberating in thal-amocortical networks. The genesis of these oscillations andtheir large-scale synchronization critically depends uponthe bursting properties of the thalamocortical cell mem-brane and the recurrency of synaptic connections withinthe thalamus and between thalamus and cortex.
Upon awakening, the function of top–down cortico-thalamic inputs could switch to precisely controlling thetransfer function of thalamocortical neurons. The demon-stration by hybrid experiments that the transmitting prop-erties of TC cells are dependent on the stochastic nature ofthe contextual synaptic feedback needs further confirma-tion in vivo but it already suggests that computationexerted by thalamic relay cells should not be entirelydefined on the basis of intrinsic conductances. Rather, tha-lamic operations could be under a graded control exertedby global network recurrency. Therefore, it is remarkablethat sleep is dominated by the impact of membrane prop-erties promoting intrinsic oscillation in thalamic cells, whilethis impact could be lessened upon awakening by the com-bined influences of ascending neuromodulatory systemsand that of background synaptic activity produced bydescending cortical inputs. The cortical control of thalamictransfer function by a tunable mixed excitatory and inhib-itory synaptic background activity presents several advan-tages over the modulation by slow neuromodulators: It isdynamic, fast and topographically precise.
Acknowledgements
This work was supported by CNRS, European Commis-sion (IST-2001-34712), Action Concertee Incitative neuro-sciences integratives et computationnelles and EU MarieCurie fellowship to J.W. We thank Gerard Sadoca andLoredana Focsaa for help with computation.
References
[1] G. Ahlsen, S. Lindstrom, F.S. Lo, Interaction between inhibitorypathways to principal cells in the lateral geniculate nucleus of the cat,Exp. Brain Res. 58 (1985) 134–143.
[2] T. Bal, D. Debay, A. Destexhe, Cortical feedback controls thefrequency and synchrony of oscillations in the visual thalamus, J.Neurosci. 20 (2000) 7478–7488.
[3] T. Bal, D.A. Mccormick, What stops synchronized thalamocorticaloscillations? Neuron 17 (1996) 297–308.
[4] T. Bal, M. Von Krosigk, D.A. Mccormick, Synaptic and membranemechanisms underlying synchronized oscillations in the ferret lateralgeniculate nucleus in vitro, J. Physiol. 483 (3) (1995) 641–663.
[5] H.B. Barlow, Critical limiting factors in the design of the eye andvisual cortex, Proc. Roy. Soc. Lond. Ser. B 212 (1981) 1–34.
[6] H. Blumenfeld, D.A. Mccormick, Corticothalamic inputs control thepattern of activity generated in thalamocortical networks, J. Neuro-sci. 20 (2000) 5153–5162.
[7] V. Bringuier, F. Chavane, L. Glaeser, Y. Fregnac, Horizontalpropagation of visual activity in the synaptic integration field of area17 neurons, Science 283 (1999) 695–699.
[8] F.S. Chance, L.F. Abbott, A.D. Reyes, Gain modulation frombackground synaptic input, Neuron 35 (2002) 773–782.
[9] C. Chen, D.M. Blitz, W.G. Regehr, Contributions of receptordesensitization and saturation to plasticity at the retinogeniculatesynapse, Neuron 33 (2002) 779–788.
[10] D. Contreras, A. Destexhe, T.J. Sejnowski, M. Steriade, Control ofspatiotemporal coherence of a thalamic oscillation by corticothalamicfeedback, Science 274 (1996) 771–774.
[11] D. Contreras, M. Steriade, Spindle oscillation in cats: the role ofcorticothalamic feedback in a thalamically generated rhythm, J.Physiol. 490 (1) (1996) 159–179.
[12] D. Contreras, I. Timofeev, M. Steriade, Mechanisms of long-lastinghyperpolarizations underlying slow sleep oscillations in cat cortico-thalamic networks, J. Physiol. 494 (1) (1996) 251–264.
[13] D.A. Coulter, J.R. Huguenard, D.A. Prince, Calcium currents in ratthalamocortical relay neurones: kinetic properties of the transient,low-threshold current, J. Physiol. 414 (1989) 587–604.
[14] F. Crick, Function of the thalamic reticular complex: the searchlighthypothesis, Proc. Natl. Acad. Sci. USA 81 (1984) 4586–4590.
[15] A. Destexhe, T. Bal, D.A. Mccormick, T.J. Sejnowski, Ionicmechanisms underlying synchronized oscillations and propagatingwaves in a model of ferret thalamic slices, J. Neurophysiol. 76 (1996)2049–2070.
[16] A. Destexhe, D. Contreras, M. Steriade, Mechanisms underlying thesynchronizing action of corticothalamic feedback through inhibitionof thalamic relay cells, J. Neurophysiol. 79 (1998) 999–1016.
[17] A. Destexhe, Z.F. Mainen, T.J. Sejnowski, Synthesis of models forexcitable membranes, synaptic transmission and neuromodulationusing a common kinetic formalism, J. Comput. Neurosci. 1 (1994)195–230.
[18] A. Destexhe, D. Pare, Impact of network activity on the integrativeproperties of neocortical pyramidal neurons in vivo, J. Neurophysiol.81 (1999) 1531–1547.
[19] A. Destexhe, M. Rudolph, J.M. Fellous, T.J. Sejnowski, Fluctuatingsynaptic conductances recreate in-vivo-like activity in neocorticalneurons, Neuroscience 107 (2001) 13–24.
[20] A. Destexhe, M. Rudolph, D. Pare, The high-conductance state ofneocortical neurons in vivo, Nat. Rev. Neurosci. 4 (2003) 739–751.
[21] A. Erisir, S.C. Van Horn, S.M. Sherman, Relative numbers of corticaland brainstem inputs to the lateral geniculate nucleus, Proc. Natl.Acad. Sci. USA 94 (1997) 1517–1520.
[22] W. Guido, T. Weyand, Burst responses in thalamic relay cells of theawake behaving cat, J. Neurophysiol. 74 (1995) 1782–1786.
[23] M.L. Hines, N.T. Carnevale, The NEURON simulation environ-ment, Neural Comput. 9 (1997) 1179–1209.
[24] J.C. Hirsch, A. Fourment, M.E. Marc, Sleep-related variations ofmembrane potential in the lateral geniculate body relay neurons ofthe cat, Brain Res. 259 (1983) 308–312.
[25] N. Ho, A. Destexhe, Synaptic background activity enhances theresponsiveness of neocortical pyramidal neurons, J. Neurophysiol. 84(2000) 1488–1496.

D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558 557
[26] H. Hultborn, S. Lindstrom, H. Wigstrom, On the function ofrecurrent inhibition in the spinal cord, Exp. Brain Res. 37 (1979) 399–403.
[27] H. Jahnsen, R. Llinas, Electrophysiological properties of guinea pigthalamic neurones: an in vitro study, J. Physiol. (Lond.) 349 (1984)205–226.
[28] U. Kim, M.V. Sanchez-Vives, D.A. Mccormick, Functional dynamicsof GABAergic inhibition in the thalamus, Science 278 (1997) 130–134.
[29] C. Koch, The action of the corticofugal pathway on sensory thalamicnuclei: a hypothesis, Neuroscience 23 (1987) 399–406.
[30] G. Le Masson, S. Le Masson, M. Moulins, From conductances toneural network properties: analysis of simple circuits using the hybridnetwork method, Prog. Biophys. Mol. Biol. 64 (1995) 201–220.
[31] G. Le Masson, S. Renaud-Le Masson, D. Debay, T. Bal, Feedbackinhibition controls spike transfer in hybrid thalamic circuits, Nature417 (2002) 854–858.
[32] S. Le Masson, A. Laflaquiere, T. Bal, G. Le Masson, Analog circuitsfor modeling biological neural networks: design and applications,IEEE Trans. Biomed. Eng. 45 (1999) 638–645.
[33] K.H. Lee, D.A. Mccormick, Abolition of spindle oscillations byserotonin and norepinephrine in the ferret lateral geniculate andperigeniculate nuclei in vitro, Neuron 17 (1996) 309–321.
[34] N. Leresche, J. Hering, R.C. Lambert, Paradoxical potentiation ofneuronal T-type Ca2+ current by ATP at resting membrane potential,J. Neurosci. 24 (2004) 5592–5602.
[35] R.R. Llinas, The intrinsic electrophysiological properties of mamma-lian neurons: insights into central nervous system function, Science242 (1988) 1654–1664.
[36] A. Luthi, T. Bal, D.A. Mccormick, Periodicity of thalamic spindlewaves is abolished by ZD7288, a blocker of Ih, J. Neurophysiol. 79(1998) 3284–3289.
[37] A. Luthi, D.A. Mccormick, H-current: properties of a neuronal andnetwork pacemaker, Neuron 21 (1998) 9–12.
[38] A. Luthi, D.A. Mccormick, Periodicity of thalamic synchronizedoscillations: the role of Ca2+-mediated upregulation of Ih, Neuron 20(1998) 553–563.
[39] J.W. Mcclurkin, L.M. Optican, B.J. Richmond, Cortical feedbackincreases visual information transmitted by monkey parvocellularlateral geniculate nucleus neurons, Vis. Neurosci. 11 (1994) 601–617.
[40] D.A. Mccormick, Neurotransmitter actions in the thalamus andcerebral cortex and their role in neuromodulation of thalamocorticalactivity, Prog. Neurobiol. 39 (1992) 337–388.
[41] D.A. Mccormick, T. Bal, Sleep and arousal: thalamocortical mech-anisms, Ann. Rev. Neurosci. 20 (1997) 185–215.
[42] D.A. Mccormick, H.R. Feeser, Functional implications of burst firingand single spike activity in lateral geniculate relay neurons, Neuro-science 39 (1990) 103–113.
[43] D.A. Mccormick, M. Von Krosigk, Corticothalamic activationmodulates thalamic firing through glutamate ‘‘metabotropic’’ recep-tors, Proc. Natl. Acad. Sci. USA 89 (1992) 2774–2778.
[44] V.M. Montero, Amblyopia decreases activation of the corticogeni-culate pathway and visual thalamic reticularis in attentive rats: a�focal attention� hypothesis, Neuroscience 91 (1999) 805–817.
[45] D. Mumford, On the computational architecture of the neocortex. I.The role of the thalamo-cortical loop, Biol. Cyber. 65 (1991) 135–145.
[46] P.C. Murphy, A.M. Sillito, Corticofugal feedback influences thegeneration of length tuning in the visual pathway, Nature 329 (1987)727–729.
[47] M.I. Posner, S.E. Petersen, The attention system of the human brain,Ann. Rev. Neurosci. 13 (1990) 25–42.
[48] A.A. Prinz, L.F. Abbott, E. Marder, The dynamic clamp comes ofage, Trends Neurosci. 27 (2004) 218–224.
[49] A.W. Przybyszewski, J.P. Gaska, W. Foote, D.A. Pollen, Striatecortex increases contrast gain of macaque LGN neurons, Vis.Neurosci. 17 (2000) 485–494.
[50] E.J. Ramcharan, C.L. Cox, X.J. Zhan, S.M. Sherman, J.W. Gnadt,Cellular mechanisms underlying activity patterns in the monkeythalamus during visual behavior, J. Neurophysiol. 84 (2000) 1982–1987.
[51] E.J. Ramcharan, J.W. Gnadt, S.M. Sherman, Burst and tonic firing inthalamic cells of unanesthetized, behaving monkeys, Vis. Neurosci. 17(2000) 55–62.
[52] H.P. Robinson, N. Kawai, Injection of digitally synthesized synapticconductance transients to measure the integrative properties ofneurons, J. Neurosci. Methods 49 (1993) 157–165.
[53] H.E. Scharfman, S.M. Lu, W. Guido, P.R. Adams, S.M. Sherman,N-methyl-D-aspartate receptors contribute to excitatory postsynapticpotentials of cat lateral geniculate neurons recorded in thalamic slices,Proc. Natl. Acad. Sci. USA 87 (1990) 4548–4552.
[54] A.A. Sharp, M.B. O�neil, L.F. Abbott, E. Marder, The dynamicclamp: artificial conductances in biological neurons, Trends Neurosci.16 (1993) 389–394.
[55] A.A. Sharp, M.B. O�neil, L.F. Abbott, E. Marder, Dynamic clamp:computer-generated conductances in real neurons, J. Neurophysiol.69 (1993) 992–995.
[56] S.M. Sherman, Tonic and burst firing: dual modes of thalamocorticalrelay, Trends Neurosci. 24 (2001) 122–126.
[57] S.M. Sherman, A wake-up call from the thalamus, Nat. Neurosci. 4(2001) 344–346.
[58] S.M. Sherman, R.W. Guillery, Functional organization of thalamo-cortical relays, J. Neurophysiol. 76 (1996) 1367–1395.
[59] S.M. Sherman, R.W. Guillery, The role of the thalamus in the flow ofinformation to the cortex, Philos. Trans. Roy. Soc. Lond. B Biol. Sci.357 (2002) 1695–1708.
[60] S.M. Sherman, C. Koch, The control of retinogeniculate transmissionin the mammalian lateral geniculate nucleus, Exp. Brain Res. 63(1986) 1–20.
[61] Y. Shu, A.R. Hasenstaub, M. Badoual, T. Bal, D.A. Mccormick,Barrage of synaptic activity control the gain and sensitivity of corticalneurons, J. Neurosci. 23 (2003) 10388–10401.
[62] A.M. Sillito, J. Cudeiro, P.C. Murphy, Orientation sensitive elementsin the corticofugal influence on centre-surround interactions in thedorsal lateral geniculate nucleus, Exp. Brain Res. 93 (1993) 6–16.
[63] A.M. Sillito, H.E. Jones, Corticothalamic interactions in the transferof visual information, Philos. Trans. Roy. Soc. Lond. B Biol. Sci. 357(2002) 1739–1752.
[64] A.M. Sillito, H.E. Jones, G.L. Gerstein, D.C. West, Feature-linkedsynchronization of thalamic relay cell firing induced by feedback fromthe visual cortex, Nature 369 (1994) 479–482.
[65] O.C. Snead 3rd, Basic mechanisms of generalized absence seizures,Ann. Neurol. 37 (1995) 146–157.
[66] M. Steriade, Impact of network activities on neuronal properties incorticothalamic systems, J. Neurophysiol. 86 (2001) 1–39.
[67] M. Steriade, E.G. Jones, D.A. MccormickThalamus—Organisationand Function, vol. 1, Elsevier Science, 1997 (Chapters 5 and 7).
[68] M. Steriade, R. Mccarley, Brainstem Control of Wakefulness andSleep, Plenum press, New York, 1990.
[69] M. Steriade, D.A. Mccormick, T.J. Sejnowski, Thalamocorticaloscillations in the sleeping and aroused brain, Science 262 (1993)679–685.
[70] K. Suder, F. Worgotter, The control of low-level information flow inthe visual system, Rev. Neurosci. 11 (2000) 127–146.
[71] H.A. Swadlow, A.G. Gusev, The impact of �bursting� thalamicimpulses at a neocortical synapse, Nat. Neurosci. 4 (2001) 402–408.
[72] S. Temereanca, D.J. Simons, Functional topography of corticotha-lamic feedback enhances thalamic spatial response tuning in thesomatosensory whisker/barrel system, Neuron 41 (2004) 639–651.
[73] I. Timofeev, D. Contreras, M. Steriade, Synaptic responsiveness ofcortical and thalamic neurones during various phases of slow sleeposcillation in cat, J. Physiol. 494 (1) (1996) 265–278.
[74] J.B. Troy, J.G. Robson, Steady discharges of X and Y retinalganglion cells of cat under photopic illuminance, Vis. Neurosci. 9(1992) 535–553.

558 D. Debay et al. / Journal of Physiology - Paris 98 (2004) 540–558
[75] W.M. Usrey, R.C. Reid, Synchronous activity in the visual system,Ann. Rev. Physiol. 61 (1999) 435–456.
[76] S.C. Van Horn, A. Erisir, S.M. Sherman, Relative distribution ofsynapses in the A-laminae of the lateral geniculate nucleus of the cat,J. Comp. Neurol. 416 (2000) 509–520.
[77] M. Von Krosigk, T. Bal, D.A. Mccormick, Cellular mechanisms of asynchronized oscillation in the thalamus, Science 261 (1993) 361–364.
[78] S.R. Williams, G.J. Stuart, Action potential backpropagation andsomato-dendritic distribution of ion channels in thalamocorticalneurons, J. Neurosci. 20 (2000) 1307–1317.
[79] J. Wolfart, D. Debay, G. Le Masson, A. Destexhe, T. Bal, Synapticbackground activity controls spike transfer from thalamus to cortex,Nature Neurosci., in press.
[80] J. Wolfart, D. Debay, G. Le Masson, A. Destexhe, T. Bal, Synapticnoise modulates the gain and smoothes the non-linear response ofthalamocortical neurons, Soc. Neurosci. Abstr., New Orleans,USA.
[81] C.D. Yingling, J.E. Skinner, Gating of Thalamic Input to Cere-bral Cortex by Nucleus Reticularis Thalami, vol. 1, Karger, Basel,1977.
Related Documents











