Differential activities of Sonic hedgehog mediated by Gli transcription factors define distinct neuronal subtypes in the dorsal thalamus Kazue Hashimoto-Torii a,b,c , Jun Motoyama d , Chi-Chung Hui e , Atsushi Kuroiwa b , Masato Nakafuku c , Kenji Shimamura a,c, * a Division of Morphogenesis, Department of Embryogenesis, Institute of Molecular Embryology and Genetics, Kumamoto University, Honjo 2-2-1, Kumamoto 860-0811, Japan b Division of Biological Science Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8602, Japan c Department of Neurobiology, Graduate School of Medicine, University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, Japan d Molecular Neuropathology Group, Brain Science Institute, RIKEN, Wako, Saitama 351-0198, Japan e Program in Developmental Biology and Division of Endocrinology, Research Institute, The Hospital for Sick Children, 555 University Avenue, Toronto, Ont., Canada M5G 1X8 Received 27 June 2003; received in revised form 27 August 2003; accepted 28 August 2003 Abstract The dorsal thalamus (DT) is a pivotal region in the vertebrate brain that relays inputs from the peripheral sensory organs to higher cognitive centers. It consists of clusters of neurons with relevant functions, called brain nuclei. However, the mechanisms underlying development of the DT, including specification of the neuronal subtypes and morphogenesis of the nuclear structures, remain largely unknown. As a first step to this end, we focused on two transcription factors Sox14 and Gbx2 that are expressed in the specific brain nuclei in the chick DT. The onset of their expression was found in distinct populations of the postmitotic cells in the prosomere 2, which was regulated by the differential activities of Sonic hedgehog (Shh) in a manner consistent with the action as a morphogen. Furthermore, both gain- and loss-of-function results strongly suggest that such distinct inductive activities are mediated selectively by different Gli factors. These results suggest that cooperation of the differential expression of Gli factors and the activity gradient of Shh signaling generates the distinct thalamic neurons at the specific locations. q 2003 Elsevier Ireland Ltd. All rights reserved. Keywords: Dorsal thalamus; Shh; Sox14; Gbx2; Gli1; Gli2; Diencephalon; Patterning; Brain nuclei; Morphogen 1. Introduction A number of studies have established that the embryonic central nervous system (CNS) is regionalized as Cartesian grids through the actions of anteroposterior and dorsoventral patterning mechanisms (reviewed by Lumsden and Krumlauf, 1996; Rubenstein et al., 1998). Proliferative progenitor cells at the different locations of the CNS produce distinct sets of neurons that constitute various brain tissues. For instance, studies of the spinal cord and telencephalon have revealed that molecularly distinct domains of progenitor cells generate specific neuronal subtypes that contribute to nearby as well as distant tissues (reviewed by Jessell and Sanes, 2000; Corbin et al., 2001). Patterning of the early neuroepithelial fields is in part achieved by the actions of inductive signals emanating from the localized sources, so that cells with distinct properties arise in a spatially organized manner with respect to the signaling centers (Agarwala et al., 2001). There is evidence that an inductive signal regulates the expression of distinct sets of transcription factors depending on its concentration. For instance, a secreted glycoprotein Sonic hedgehog (Shh) has been demonstrated to induce floor plate properties at a high concentration and progres- sively more dorsal molecular properties at lower concen- trations in vitro (Roelink et al., 1995; Ericson et al., 1997). 0925-4773/$ - see front matter q 2003 Elsevier Ireland Ltd. All rights reserved. doi:10.1016/j.mod.2003.09.001 Mechanisms of Development 120 (2003) 1097–1111 www.elsevier.com/locate/modo * Corresponding author. Address: Division of Morphogenesis, Department of Embryogenesis, Institute of Molecular Embryology and Genetics, Kumamoto University, Honjo 2-2-1, Kumamoto 860-0811, Japan. Tel.: þ 81-96-373-6583; fax: þ81-96-373-6586. E-mail address: [email protected] (K. Shimamura).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Differential activities of Sonic hedgehog mediated by Gli transcription
factors define distinct neuronal subtypes in the dorsal thalamus
Kazue Hashimoto-Toriia,b,c, Jun Motoyamad, Chi-Chung Huie, Atsushi Kuroiwab,Masato Nakafukuc, Kenji Shimamuraa,c,*
aDivision of Morphogenesis, Department of Embryogenesis, Institute of Molecular Embryology and Genetics,
Kumamoto University, Honjo 2-2-1, Kumamoto 860-0811, JapanbDivision of Biological Science Graduate School of Science, Nagoya University, Furo-cho, Chikusa-ku, Nagoya 464-8602, Japan
cDepartment of Neurobiology, Graduate School of Medicine, University of Tokyo, 7-3-1 Hongo, Bunkyo-ku, Tokyo 113-0033, JapandMolecular Neuropathology Group, Brain Science Institute, RIKEN, Wako, Saitama 351-0198, Japan
eProgram in Developmental Biology and Division of Endocrinology, Research Institute, The Hospital for Sick Children,
555 University Avenue, Toronto, Ont., Canada M5G 1X8
Received 27 June 2003; received in revised form 27 August 2003; accepted 28 August 2003
Abstract
The dorsal thalamus (DT) is a pivotal region in the vertebrate brain that relays inputs from the peripheral sensory organs to higher cognitive
centers. It consists of clusters of neurons with relevant functions, called brain nuclei. However, the mechanisms underlying development of
the DT, including specification of the neuronal subtypes and morphogenesis of the nuclear structures, remain largely unknown. As a first step
to this end, we focused on two transcription factors Sox14 and Gbx2 that are expressed in the specific brain nuclei in the chick DT. The onset
of their expression was found in distinct populations of the postmitotic cells in the prosomere 2, which was regulated by the differential
activities of Sonic hedgehog (Shh) in a manner consistent with the action as a morphogen. Furthermore, both gain- and loss-of-function
results strongly suggest that such distinct inductive activities are mediated selectively by different Gli factors. These results suggest that
cooperation of the differential expression of Gli factors and the activity gradient of Shh signaling generates the distinct thalamic neurons at
the specific locations.
q 2003 Elsevier Ireland Ltd. All rights reserved.
Keywords: Dorsal thalamus; Shh; Sox14; Gbx2; Gli1; Gli2; Diencephalon; Patterning; Brain nuclei; Morphogen
1. Introduction
A number of studies have established that the
embryonic central nervous system (CNS) is regionalized
as Cartesian grids through the actions of anteroposterior
and dorsoventral patterning mechanisms (reviewed by
Lumsden and Krumlauf, 1996; Rubenstein et al., 1998).
Proliferative progenitor cells at the different locations of the
CNS produce distinct sets of neurons that constitute various
brain tissues. For instance, studies of the spinal cord and
telencephalon have revealed that molecularly distinct
domains of progenitor cells generate specific neuronal
subtypes that contribute to nearby as well as distant tissues
(reviewed by Jessell and Sanes, 2000; Corbin et al., 2001).
Patterning of the early neuroepithelial fields is in part
achieved by the actions of inductive signals emanating from
the localized sources, so that cells with distinct properties
arise in a spatially organized manner with respect to the
signaling centers (Agarwala et al., 2001).
There is evidence that an inductive signal regulates the
expression of distinct sets of transcription factors depending
on its concentration. For instance, a secreted glycoprotein
Sonic hedgehog (Shh) has been demonstrated to induce
floor plate properties at a high concentration and progres-
sively more dorsal molecular properties at lower concen-
trations in vitro (Roelink et al., 1995; Ericson et al., 1997).
0925-4773/$ - see front matter q 2003 Elsevier Ireland Ltd. All rights reserved.
doi:10.1016/j.mod.2003.09.001
Mechanisms of Development 120 (2003) 1097–1111
www.elsevier.com/locate/modo
* Corresponding author. Address: Division of Morphogenesis,
Department of Embryogenesis, Institute of Molecular Embryology and
Genetics, Kumamoto University, Honjo 2-2-1, Kumamoto 860-0811,
Japan. Tel.: þ81-96-373-6583; fax: þ81-96-373-6586.
E-mail address: [email protected] (K.
Shimamura).

Factors involved in the Hedgehog (Hh) signaling cascade
have been identified and turned out to be evolutionally
conserved among diverse organisms (reviewed by
Nybakken and Perrimon, 2002). However, the molecular
mechanism underlying the differential activities of Shh are
not yet fully understood. The Gli zinc finger transcription
factor is believed to mediate most of the transcriptional
outputs of Hh signaling (reviewed by Koebernick and
Pieler, 2002; Ruiz i Altaba et al., 2002). While a single Gli
factor Cubitus interruptus (Ci) is absolutely required for Hh
signaling in Drosophila (Methot and Basler, 2001; reviewed
by Aza-Blanc and Kornberg, 1999), vertebrates have three
Ci homologues, Gli1, Gli2 and Gli3, which exhibit different
expression patterns in the developing embryos and exert
distinct functions in some biological situations (Hui et al.,
1994; Ruiz i Altaba, 1998; Aza-Blanc et al., 2000; Persson
et al., 2002; Karlstrom et al., 2003). While models have
been proposed that the graded actions of Hh signaling are
attributed to the activities of several different states of Ci
(Wang and Holmgren, 2000; Methot and Basler, 1999),
exactly how these vertebrate Gli factors are involved in the
execution of the morphogen-like feature of Shh has not yet
been elucidated.
The dorsal thalamus (DT) is a pivotal forebrain structure
that functions as the major relay center connecting the
peripheral sensory organs and cerebral cortex (Jones, 1985,
1998). The DT is partitioned into many clusters of neurons
with relevant functions, called nuclei, which can be
distinguished histologically by features such as the cellular
morphology, differential cell density, and the trajectories of
axon bundles that often demarcate the outlines of the brain
nuclei. Each DT nucleus projects to a specific location
mainly in the telencephalon, and receives inputs from
specific targets. Therefore, the spatial organization of each
DT nucleus is crucial for establishing this precise topo-
graphical relationship to fulfill its functions as a relay
center. Developmentally, the DT is believed to be derived
from a single embryonic subdivision, the alar plate of
prosomere 2 (p2) (Puelles and Rubenstein, 1993). Since the
morphological characteristics that define each DT nucleus
become evident only late in development, the mechanisms
underlying the early stages of its development remain
largely unknown. Several attempts to address this issue have
been made recently (Redies et al., 2000; Yoon et al., 2000;
Nakagawa and O’Leary, 2001; Martınez-de-la-Torre et al.,
2002). For instance, Nakagawa and O’Leary have reported
the nested expression of transcription factors in the
developing mouse thalamus (Nakagawa and O’Leary,
2001). The distinct but overlapping expression patterns of
these regulatory genes often correlated with the histologi-
cally defined borders of the nuclei. Through ontogenic
studies of the expression patterns, they have suggested that
neurons constituting each thalamic nucleus are born with
distinct molecular properties early in development.
In order to elucidate the developmental mechanism of the
DT, we took a similar approach, finding molecular
differences in the DT nuclei and the DT rudiment of the
chick. We provide evidence that Shh emanating from the
basal plate of the p2 specifies two distinct neuronal subtypes
in the DT rudiment in a concentration-dependent manner.
Moreover, these graded actions of Shh are likely to be
mediated by Gli1 and Gli2 selectively, which has not been
reported in other biological situations in which Hh signaling
is involved. These results provide a significant insight into
the molecular mechanism underlying the morphogen-like
property of Shh signaling.
2. Results
2.1. Several DT nuclei are distinguished by expression
of transcription factors
In order to obtain molecular clues to study the DT
development, we have searched for molecules expressed in
subsets of DT nuclei. Among the list, we found that Sox14,
Gbx2 and Sox2 are expressed in restricted regions of the DT
at Hamburger and Hamilton’s stage 42 (HH42) when each
DT nucleus can be identified histologically (Fig. 1A,D).
Most of their expression correlated well with the topogra-
phical organization of the DT nuclei, in that the borders of
their expression domains often coincide with the boundaries
of the nuclei defined morphologically. Expression of Sox14
was restricted to the perirotundic area and the prospective
interstitial nucleus of the optic tract (arrowheads in Fig. 1B).
Gbx2 was expressed in many DT nuclei, as reported in mice
and chick (Miyashita-Lin et al., 1999; Nakagawa and
O’Leary, 2001; Martınez-de-la-Torre et al., 2002). For
instance, the nucleus rotundus, the largest nucleus in the
chick DT, was labeled by the prominent expression of Gbx2
(Fig. 1C), which was surrounded by the Sox14-positive
perirotundic area (Fig. 1B). The subrotundic nucleus and
subhabenular region also showed strong Gbx2 expression,
whereas the dorsal anterior nucleus and dorsolateral anterior
nucleus exhibited lower expression levels (Fig. 1C). Sox2
identified a different set of nuclei, such as nucleus ovoidalis
and the dorsointermediate posterior nucleus, both of which
also showed weak Gbx2 expression (arrowheads in Fig. 1E;
data not shown).
We next asked when such molecular heterogeneity
emerges during development. At HH22, when the thin
mantle layer of postmitotic neurons is formed in the DT (see
Fig. 1M), the expression patterns were already distinct in the
whole-mount specimens (Fig. 1F–H). Sox14 and Gbx2
expression is first detectable at about the same time (HH17)
in a region adjacent to the basal plate of p2 (Fig. 1I–K);
Sox14 expression started in a line of cells arrayed parallel to
the dorsal limit of the basal plate (Fig. 1I), whereas Gbx2-
positive cells emerged somewhat sporadically in a small
area with a extension dorsally (Fig. 1J). Double labeling
showed that there was no overlap between the expression of
these markers (Fig. 1K). As development proceeds,
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111098

the Gbx2 expression occupied a large quadrilateral area in
the DT rudiment whose anterior and ventral edges were
fringed by the hinge-shaped Sox14-positive domain
(Fig. 1L). Close examination of the sectioned material
revealed that they were expressed in a mutually exclusive
pattern, such that cells expressing either of them are
clustered right next to each other (Fig. 1N).
These observations provide evidence that some early
postmitotic neurons in the DT primordium are already
molecularly distinct, although lineage relationship between
these populations at the different stages remains unclear at
this moment.
2.2. Tissues adjacent to the DT rudiment are required
for the expression of Sox14 and Gbx2
To understand how this early molecular heterogeneity is
created in the rudiment of DT, we first asked whether the DT
Fig. 1. Expression of the transcription factors in the DT. (A–C) Coronal sections of HH42 embryos at the level of the rostral DT stained for thionin (A), Sox14
(B) and Gbx2 (C). (D,E) Coronal sections of HH42 embryos at the level of the caudal DT stained for thionin (D) and Sox2 (E). Sox14 is expressed in the cells
(arrowheads in B) that surround the nucleus rotundus where Gbx2-positive cells are evenly distributed (C). A high magnification of the Sox14-expressing
region is shown in the inset in (B). (F–H) HH22 dissected brain whole-mount in situ hybridized for Sox14 (F), Gbx2 (G) and Sox2(H). Note that Sox2 is
expressed strongly in the mantle layer of the DT anlagen (arrows in H), and weakly in the ventricular layer of the entire brain. Dark staining in the
telencephalon is background. For all the whole-mount specimens, the anterior is to the right and the dorsal is to the top. (I,J) HH17 DT rudiments stained for
Sox14 (I) and Gbx2 (J) showing the onset of their expression (arrowheads). Faint darkening in a broad area of p2 is background (J). (K,L) Two color in situ
hybridization for Sox14 (blue) and Gbx2(brown) of the whole-mount DT rudiments at HH17 (K) and HH22 (L). (M,N) Coronal sections of the DT anlagen at
HH20 stained for NeuN (M) and Sox14 (blue in N) and Gbx2 (orange in N). Note that Gbx2-expressing cells are adjacent to, but segregated from the small
domain of Sox14 expression in the layer of postmitotic neurons (arrowhead in K,L,N). For (M) and (N), the third ventricle is located at the right side of the
panels and the dorsal is to the top. Bars, 0.5 mm for (A)–(L); 0.05 mm for (M) and (N). ApR, perirotundic area; DA, dorsal anterior nucleus; DIP,
dorsointermediate posterior nucleus; DLA, dorsolateral anterior nucleus; DLL, dorsolateral lateral nucleus; DLP, Dorsolateral posterior nucleus; DMP,
dorsomedial posterior nucleus; (H), habenula; ITO, prospective interstitial nucleus of the optic tract; Me, mesencephalon; Ov, nucleus ovoidalis; PM, nucleus
paramedianus internus; p1, prosomere 1; p2, prosomere 2; Rt, rotundus nucleus; SH, subhabenular nucleus; SPC, Parvocellular superficial nucleus; SRt,
subrotundic nucleus; Te, telencephalon; VT, ventral thalamus; ZLI, zona limitans intrathalamica.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1099

rudiment acquires this spatial pattern intrinsically or
extrinsically. Previous study has shown that the molecular
regionality in the isolated mouse diencephalic explants
cultured in vitro was remarkably stable (Echevarria et al.,
2001). Fragments of the neural tube that contained the DT
rudiment and some flanking tissues that include the basal
plate, a posterior part of p3, and an anterior part of p1, were
isolated from HH8–12 embryos before the onset of Sox14
and Gbx2 expression (type A in Fig. 2A). After being
cultured in vitro for 72 h, the characteristic patterns of
Sox14 and Gbx2 expression were detected (Fig. 2B), which
were comparable to those in the normal embryo at the
corresponding stages (see Fig. 1L). By contrast, when those
flanking tissues, the boundaries of which are easily
discernible under a dissection microscope, were eliminated
prior to the culture (type B in Fig. 2A), neither Sox14 nor
Gbx2 expression was detected (Fig. 2C). Presence of the
flanking tissues was monitored in part by Shh which is
expressed in the basal plate and the boundary that
demarcates the anterior edge of DT (Fig. 2D,E). The results
of the explant studies are summarized in Table 1. Unlike
Sox14 or Gbx2, generation of Sox2-positive neurons did not
appear to require these tissues (n ¼ 14=14; Fig. 2F,G).
Expression of Tuj1 and MAP2, pan-neuronal markers, was
indistinguishable between these different preparations,
suggesting that neurogenesis in general was not affected
by the removal of the flanking tissues (Fig. 2H,I; data not
shown). No significant difference in cell death was detected
by TUNEL analysis (Fig. 2J,K).
These results suggest that the spatial information is
not intrinsic to the DT rudiment but dependent on the
environmental cues around it. Shh is expressed in the basal
plate of p2 at the time of their onset, and also in the anterior
boundary of the DT, zona limitans intrathalamica (ZLI),
after HH19 (data not shown; Martı et al., 1995; Larsen et al.,
2001; Zeltser et al., 2001). In fact, there is a tight correlation
between expression of Shh and that of Sox14 and Gbx2 in
the cultured explants (see Table 1). These results prompted
us to examine a role of Shh in the pattern formation of the
DT rudiment.
Fig. 2. Tissues adjacent to the DT rudiment are required for the expression
of Sox14 and Gbx2. (A) Experimental paradigms for dissection and culture
of the brain explants isolated from HH8–12 embryos. Cultured explants
which had been prepared as type A (B,D,F,H,J) or type B (C,E,G,I,K) were
fixed after 72 h of culture and stained for Sox14 (blue in B,C), Gbx2 (orange
in B,C), Shh (D,E), Sox2 (F,G) and Tuj1 (H,I), or examined for apoptosis
using TUNEL (J,K). The pineal body is developed in the dorsal tip of the
explant, which is indicative of p2 (asterisk in B–G,J,K). Bar, 0.25 mm.
Table 1
Summary of the explant studies
Age Shh þ Sox14 þ Gbx2 þ Gli1 þ Gli2 þ
Type A
HH12 6/6 20/20 20/20 15/15 15/15
HH11 32/32 28/28 24/24 2/2 2/2
HH10 18/18 14/14 13/13 ND ND
HH9 10/10 6/6 4/4 ND ND
HH8 23/23 13/13 16/16 4/4 4/4
Type B
HH12 0/8 0/20 0/20 12/12 12/12
HH11 0/28 0/26 0/16 14/14 14/14
HH10 0/13 0/15 0/14 ND ND
HH9 1/12 0/10 2/16 6/6 6/6
HH8 4/20 4/32 2/8 ND ND
The number of DT explants positive for the markers listed above which
were dissected from various stages of embryos shown in the left column.
Denominators represent the total number of explants examined. Type A and
B explants are depicted in Fig. 2A.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111100

2.3. Shh induces both Sox14 and Gbx2 in the DT
We first examined whether Shh is capable of inducing the
markers that we identified. Approximately same size of the
type B explants (Fig. 2A) dissected from HH12 embryos
were cultured with various concentrations of recombinant
Shh-N protein (Roelink et al., 1995). At 50 nM, only Gbx2
expression was detected in the explants (Fig. 3B). As the
concentration increased, Sox14 was induced progressively,
while Gbx2 expression appeared reduced (Fig. 3C,D). These
results are represented quantitatively in Fig. 3E, which is
based on 82 specimens that were single stained for either
Sox14 or Gbx2 with the same chromatic substrate. The
number of Tuj1-positive cells in the Shh-treated explants
was comparable to the control (data not shown), which
suggests that the augmentation of these markers expressed
in the postmitotic neurons was not simply due to promotion
of generic neurogenesis. We therefore concluded that Shh
altered the neuronal fates of the cells in the explants.
To address further whether the differential induction of
Sox14 and Gbx2 is indeed caused by a direct action of Shh
signal, we took advantage of the transmembrane protein
Smoothened which transduces the Hh signals (Alcedo et al.,
1996; van den Heuvel and Ingham, 1996). It has been shown
that a constitutively active form of Smoothened (SmoM2)
recapitulated the actions of Shh in a cell-autonomous
manner (Xie et al., 1998; Hynes et al., 2000). The type B
explants (see Fig. 2A) from HH12 embryos were micro-
electroporated with different concentrations of the SmoM2
plasmid using a fine glass needle (Ø ¼ 0.03 mm; Fig. 3H,I;
Haas et al., 2001), so that varying amounts of the
constitutively active Smoothened protein would be pro-
duced. The methodological validity was provided using a
Gfp construct (Fig. 3J,K). For strict comparison, approxi-
mately same locations within p2 were targeted for
electroporation (dorsal 3/4; Fig. 3J,K). As a result, only
Gbx2 was induced by the low level of SmoM2 expression
(2.5 mg/ml; n ¼ 17=21; Fig. 3N,O), whereas Sox14 was
predominantly induced instead by the high level of SmoM2
expression apparently in a cell-autonomous manner
(6.0 mg/ml; n ¼ 11=11; Fig. 3P,Q). Under this experimental
condition, any sign of Shh induction was detected by either
doses of SmoM2 (n ¼ 0=19; data not shown). A similar, but
less clear dose-dependency was also observed with the Shh
plasmid (data not shown). These results indicate that the
graded activities of Shh signaling can selectively induce
Sox14 and Gbx2 without a relay signal. On the other hand,
we did not see any significant change in Sox2 expression in
the DT upon Shh misexpression (n ¼ 12=12; data not
shown), suggesting that some neuronal populations in the
DT are independent of Shh signaling.
Next, we examined whether Shh is indeed required for
the induction of Sox14 and Gbx2 by using a function-
blocking antibody (Ericson et al., 1996; Gunhaga et al.,
2000). The DT rudiments with flanking tissues (type A
explant; Fig. 2A) in which both Sox14 and Gbx2 would be
expressed (see Fig. 2B) were dissected from HH12 embryos,
and cultured in the presence of anti-Shh monoclonal
antibody (5E1; 15 mg/ml). As a consequence, the expression
of Sox14 and Gbx2 was absent or severely reduced in those
explants (n ¼ 8=12; Fig. 4A,B). No significant change in
either the expression of Tuj1 (Fig. 4C,D) or TUNEL
staining (Fig. 4E,F) was observed, suggesting that the
concentration of the antibody used in this assay did not
affect generic neuronal differentiation or apoptosis. We also
estimated the time window for the requirement of Shh by
12 h-pulse-incubation with the antibody at the different time
point during the culture (Fig. 4G). The antibody treatment
during the first 24 h was sufficient to block Sox14 and Gbx2
expression, whereas the later treatment was much less
effective (Fig. 4G). Neuronal differentiation did not take
place markedly until after 24 h of culture (Fig. 4H–J), and
the onset of Sox14 and Gbx2 expression was about after 36 h
of culture (data not shown). Collectively, these results
suggest that Shh is required by the mitotic progenitor cells
in the DT, but to a much less extent by the postmitotic
neurons to maintain these molecular properties.
Thus, this series of experiments demonstrated that Shh
presumably produced by the adjacent tissues is necessary
and sufficient for the proper expression of Sox14 and Gbx2
in the DT.
2.4. The distinct activities of Shh signaling are mediated
by different Gli factors
As Shh signaling is mediated by Gli zinc finger proteins
(Lee et al., 1997; Hynes et al., 1997; Sasaki et al., 1997;
reviewed by Ingham and McMahon, 2001), we hypo-
thesized that these Gli factors may be differently involved
in the induction of Sox14 and Gbx2 in the DT. In support of
this idea, the expression of Gli1 and Gli2 appeared
complimentary in the DT rudiment at HH15 just before
the generation of Sox14- and Gbx2-expressing cells: Gli1
expression is restricted to a band just dorsal to the alar/basal
boundary, whereas Gli2 is expressed dorsal to the Gli1-
expressing domain (Fig. 5A). Soon after this stage (HH18),
the Gli1 expression expands dorsally retaining the highest
intensity towards the alar/basal boundary, and thus the
Gli1- and Gli2-expressing domains became overlapping
(Fig. 5B–E). Meanwhile, Sox14-positive postmitotic cells
emerged at the ventral edge of Gli1-expressing domain
(Fig. 5D), whereas Gbx2-positive cells were distributed over
the Gli2-expressing progenitor zone (Fig. 5E). The third
member, Gli3, was expressed in the most dorsal areas of the
DT, which fit neither the Sox14 nor Gbx2 territory at these
stages (Fig. 5F).
First, we tried to reveal potential differences in Gli1 and
Gli2 simply by misexpression. The pretectum or p1 was
chosen for this assay, because it turned out that the DT
rudiment or p2 did not respond to the change of Gli
expression for unknown reason (data not shown). We have
found that p1 similarly responded to activation of the Hh
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1101

Fig. 3. Shh is necessary and sufficient for the induction of Sox14 and Gbx2. (A–D,F,G) Explants of the DT rudiment without the flanking tissues isolated from
HH12 embryos were cultured in vitro for 72 h (A–D) or 48 h (F,G) with 0 (A,F), 50 (B), 300 (C) or 1250 nM (D,G) of recombinant Shh-N, and then hybridized
for Sox14 (dark blue in A–D) and Gbx2 (brown in A–D), or Gli1 (blue in F, brown in G) and Gli2 (brown in F, blue in G). (E) Quantitative representation of the
induction analysis. The same numbers explants for each treatment [n ¼ 8 (0 nM); n ¼ 11 (150 nM); n ¼ 10 (300 nM); n ¼ 12 (1250 nM)] were stained
individually for Sox14 or Gbx2 with the same chromatic substrate. Proportions of the Sox14- or Gbx2-expressing domains over the explants are represented by
the orange and green bars, respectively. (H) Schematic illustration depicting the microelectroporation for the type B explant. A fine glass capillary filled with
various concentrations of DNA was used for the cathode (I), and electroporation was made focally at the equivalent sites (dorsal 3/4 level) in the type B
explants. As examples, explants electroporated with 2.5 mg/ml (J) or 6.0 mg/ml (K) of the Gfp plasmid were shown. (L–Q0) Type B explants that had been
electroporated with control vector (L–M0), 2.5 mg/ml (N–O0), or 6.0 mg/ml (P–Q0) of constitutively active Smoothened (SmoM2) were cultured for 72 h and
then stained for Sox14 (L,N,P) and Gbx2 (M,O,Q). Locations of exogenous gene expression are visualized by GFP fluorescence (L0,M0,N0,O0,P0,Q0). Total
concentration of DNA was adjusted equally (6.0 mg/ml) for the all experiments. Bars, 0.25 mm for (A)–(D), (F), (G), (J), (K); 0.1 mm for (I), (L)–(Q0).
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111102

signaling (i.e. Shh, SmoM2) to express Sox14 and Gbx2 as
p2 did in the previous assays up to HH14 (Sox14 by Shh,
n ¼ 24=34; Gbx2 by Shh, n ¼ 35=38; Sox14 by SmoM2,
n ¼ 10=13; Gbx2 by SmoM2, n ¼ 13=13; data not shown),
consistent with the notion that the pretectum and DT
progressively segregate by HH16 in the chick (Larsen et al.,
2001). Again, the focal microelectroporation technique for
explants was employed to achieve precise control of the
location and the concentration of DNA. When Gli1 was
electroporated, Sox14 but not Gbx2 was induced, although
the location of this induction was restricted to the subregion
of the pretectm (n ¼ 2=9; Fig. 5I,J). Conversely, electro-
poration of Gli2 into the equivalent sites resulted in
induction of Gbx2 instead of Sox14 (n ¼ 8=12; Fig. 5K,L),
even when the concentration of the Gli2 plasmid was raised
up to four fold (data not shown). In the normal p1, however,
neither Gbx2 nor Sox14 are expressed in a similar way as in
the DT despite this competence and the presence of Shh in
the basal plate of p1 (see Fig. 1F,G). Yet, the potential
difference of Gli1 and Gli2 revealed here could be relevant
to the DT patterning, at least in the context of gene
regulation.
To address further whether Gli1 and Gli2 indeed play a
role in the expression of Sox14 and Gbx2, respectively, the
Gli mutant mice were analyzed. While single mutants for
Gli2 showed indistinguishable expression of Gbx2 and
Sox14 in the DT rudiment from the wild-type (Fig. 6A,B),
the Gli2; Gli3 double mutant mice exhibited a drastic
reduction of Gbx2 expression in the DT, but not of Sox14 at
11.5 days post coitum (dpc) (n ¼ 3=3; Fig. 6C). In these
animals, Shh expression in the ventral CNS was severely
down-regulated (compare Fig. 6D,E). Yet, Shh expression
in the presumptive ZLI was clearly detectable (Fig. 6E).
Moreover LacZ expression driven by the Patched1 promoter
was also detected in the prospective DT and ventral
thalamus (Fig. 6G), suggesting that Hh signaling was still
occurring in the absence of Gli2 and Gli3. While it has been
shown that Gli3 has an antagonizing activity against Hh
signaling (Ruiz i Altaba, 1999; Litingtung and Chiang,
2000), Gli3 has also been implicated in compensating for
loss of Gli2 activities (Mo et al., 1997; Motoyama et al.,
1998). Gli3 single mutants themselves had no defect in the
Gbx2 and Sox14 expression in the DT (data not shown).
Although the previous reports have identified no obvious
abnormality in the ventral patterning of the spinal cord in
the Gli1 mutants (Matise et al., 1998; Park et al., 2000; Bai
et al., 2002), the Sox14 expression in the DT, but not in the
pretectum, was absent or severely reduced at 11.5 dpc
(Fig. 7A,B). In contrast, the Gbx2 expression in the DT was
indistinguishable from the wild-type or heterozygotes
(Fig. 7C,D). No alteration in Shh or Patched1-LacZ
expression has been detected in the Gli1 mutants (data not
shown; Fig. 7E,F). We then examined whether Shh at any
dose can induce Sox14 in the absence of Gli1. The DT
explants equivalent to the type A in the chick were prepared
from 9.5 dpc Gli1 mutants and cultured in the presence of
various concentrations of recombinant Shh-N for 3 days. As
much as 1800 nM of Shh-N did not induce Sox14 in the
Gli1 2/2 explants (data not shown; Fig. 7G), whereas Sox14
was constantly induced in the wild type and Gli1 þ/2
explants with Shh-N above 600 nM (data not shown). When
Gli1 was electroporated to the Gli1-deficient explants,
Fig. 4. Explants of the DT rudiment with the flanking tissues dissected form
HH12 embryos cultured with normal mouse IgG (A,C,E) or anti-Shh
antibodies (B,D,F) for 48 h (C,D) and 72 h (A,B,E,F) were stained for
Sox14 (blue in A,B), Gbx2 (red in A,B) and Tuj1 (C,D), or examined for
apoptosis using TUNEL (E,F). Both Sox14 and Gbx2 expressions were
greatly diminished by anti-Shh antibodies without significant alterations in
neuronal differentiation or apoptosis. (G) Temporal change of the effect of
anti-Shh antibody on Sox14 and Gbx2 expression. The antibody was added
to the culture media for 12 h at different time points during the culture (72 h
in total) as indicated in the horizontal scale. Pixels of Sox14- (red for control
IgG, magenta for anti-Shh) and Gbx2-positive areas (turquoise for control
IgG, purple for anti-Shh) in the explants were calculated by NIH image
software. (H–J) HH12 type A explants at 12 (H), 18 (I), and 24 h (J) of
culture stained for Tuj1. Bar, 0.25 mm.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1103

Sox14 was induced where the exogenous gene was
expressed (Fig. 7I), although we found that a considerable
dose of Shh-N (.600 nM) was needed to for this induction
(Fig. 7G). Importantly, this was not achieved by exogenous
Gli2, except to a very low extent only in the presence of very
high dose of Shh-N (.1250 nM), indicating overt pre-
cedence of Gli1 for the Sox14 induction in the DT (Fig. 7G).
2.5. Mutual repression between Sox14 and Gbx2
While combination of the differential Gli expression and
the graded distribution of Shh protein could theoretically
define the precise positions of the distinct neuronal
subtypes, the complementary expression of Sox14 and
Gbx2 (see Fig. 1N) suggested that there may be mutual
repression between these transcription factors. In fact, when
Gbx2 was electroporated in the prospective Sox14 territory
at HH12, the Sox14 expression was suppressed (n ¼ 7=7;
Fig. 8B). Conversely, forced expression of Sox14 resulted in
suppression of Gbx2 apparently in a cell-autonomous
manner (n ¼ 8=8; Fig. 8D). Importantly, this cross regu-
lation is specific, as neither of them affected the expression
of Sox2 (for Sox14, n ¼ 8=8; Fig. 8F; data not shown for
Gbx2, n ¼ 12=12). On the other hand, we found that the
exogenous Gbx2 can induce endogenous Gbx2 (Fig. 8G). At
this stage (HH12 , ), exogenous Gbx2 did not induce Fgf8
(n ¼ 16=16; data not shown) as has been reported for the
earlier stages (Millet et al., 1999; Katahira et al., 2000).
Moreover, co-electroporation of Gbx2 and the dominant-
negative form of Fgfr3(Amaya et al., 1993; Kobayashi et al.,
2002) did not perturb this induction (n ¼ 4=4; data not
shown). Therefore it is unlikely that this auto-induction was
due to the cross-regulatory loop involved in the mid-
hindbrain development (Garda et al., 2001). No such auto-
induction, however, was observed for Sox14 (Fig. 8H).
These results suggest that the initial pattern created by
the diffusible signal is then consolidated by the transcrip-
tional regulations of the target genes in the DT, as reported
Fig. 5. Gli1 and Gli2 can selectively induce Sox14 and Gbx2, respectively. (A) A coronal section of the DT at HH15 double stained for Gli1 (blue) and Gli2
(orange). (B,C) HH18 chick brains whole-mount stained for Gli1 (B) and Gli2 (C). (D,E,F) Adjacent transverse sections of the DT at HH19 double stained for
Gli1 (blue) and Sox14 (orange, arrowhead) (D), for Gli2 (blue) and Gbx2 (orange) (E), or for Gli3 (purple) and Gbx2 (orange) (F). (F) is slightly more dorsal
than (D),(E). P1 explants dissected from HH12 embryos that had been electroporated with control vector (G–H0), mGli1 (I–J0) and mGli2 (K–L0) were cultured
for 72 h and stained for Sox14 (G,I,K) and Gbx2 (H,J,L). Locations of the exogenous gene expression are visualized by GFP fluorescence (G0,H0,I0,J0,K0,L0).
Bars, 0.1 mm for (A),(D)–(K0); 0.4 mm for (B),(C).
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111104

for many other developmental situations (reviewed by
Jessell, 2000).
3. Discussion
In this paper, we have shown that three regulatory genes,
Gbx2, Sox14 and Sox2 are expressed in the specific DT
nuclei (Fig. 1). Gbx2 encodes a homeodomain transcription
factor essential for the development of many DT nuclei
(Miyashita-Lin et al., 1999). Sox2 and Sox14, which belong
to subgroups B1 and B2 of the Sox family, respectively,
encode a group of proteins that carry a HMG DNA-binding
domain (Uchikawa et al., 1999; Hargrave et al., 2000;
reviewed by Kamachi et al., 2000). Detailed descriptions of
their expression in relation to the histogenesis of DT will be
published elsewhere (Hashimoto-Torii et al., in prepara-
tion). Here we will discuss the patterning mechanisms
operating in the early development of the DT with particular
regard to the Hh signaling pathway.
3.1. Differential involvement of the Gli factors in Hh
signaling
While Shh has been shown to act as a morphogen
inducing distinct genes in a concentration dependent
manner (Roelink et al., 1995; Ericson et al., 1997), the
underlying mechanisms are not yet fully understood. Our
present findings on the induction of Sox14 and Gbx2 by Shh
and Gli factors provide an important insight into this issue.
Namely, the high dose of Shh signaling and exogenous Gli1
led to Sox14 induction, whereas the low dose of Shh
signaling and Gli2 led to Gbx2 expression. This raises the
possibility that the high and low doses of Shh signals are
selectively mediated by Gli1 and Gli2, respectively. While
all Gli proteins recognize a consensus DNA sequence
(Kinzler and Vogelstein, 1990; Vortkamp et al., 1995), both
gain- and loss-of-function studies have revealed that Gli
factors differ in the property of Hh target gene regulation
(Ruiz i Altaba, 1998, 1999; Persson et al., 2002; Karlstrom
et al., 2003). For instance, it has been shown that Gli1 can
induce FoxA4 (Hnf3b), a floor plate marker, and Nkx2.1 in
the frog embryos, whereas Gli2 can induce motor neuron
marker HB9 instead (Ruiz i Altaba, 1998). Our in vitro
result that exogenous Gli2 was not able to induce Sox14
efficiently in the Gli1-deficient explants even in the
presence of very high dose of Shh-N would support this
possibility. Although it appears that Gli3 is also required for
inducing Gbx2 from the double mutant phenotype
(Fig. 6B,C), we think that it is unlikely that Gli3 plays a
role in the onset of Gbx2 expression in the normal situation,
simply because it is not expressed in the ventral but dorsal
domain of DT (Fig. 5F, Grove et al., 1998).
Curiously, however, no obvious defect has been reported
in the spinal cord of the Gli1-deficient animals (Matise et al.,
1998; Park et al., 2000; Bai et al., 2002). A conceivable
explanation is that other Gli factors, most likely to be Gli2,
may compensate for Gli1 functions in some situations. In
support of this notion, the Gli1; Gli2 double mutants exhibit
more severe abnormalities in the Hh-dependent processes
than either single mutant (Park et al., 2000). Alternatively,
for spatial patterning, the assigned functions to each Gli
factor may vary among the different regions of the CNS.
Even the morphogen-like action of Shh may also be variably
responsible for the spatial patterning among the different
regions of the CNS. For instance, Agarwala et al. (2001)
have recently provided in vivo evidence that an ectopic
focal source of Shh is solely sufficient for generating the
exquisite spatial pattern in the ventral midbrain as predicted
by the hypothesis that Shh acts as a morphogen. However,
Fig. 6. Gli2 and Gli3 are essential for Gbx2 expression, but not for Sox14.
(A–C) Lateral view of the DT of an 11.5 dpc wild-type A, Gli2 single (B)
and Gli2; Gli3 double mutant (C) double stained for Sox14 (blue) and Gbx2
(red). Note that Gbx2 expression is absent in the double mutant (C),
whereas Sox14 expression persists (arrow in C). (D,E) Head regions of a
9.5 dpc wild-type (D) and Gli2; Gli3 double homozygotes (E) in situ
hybridized for Shh. (F,G) Ptc1–LacZ expression revealed by X-gal staining
of the dissected brains from a 10.0 dpc Gli2 heterozygote (F) and Gli2; Gli3
double homozygote (G). Expression of Shh and Ptc1–LacZ persists around
the ZLI (arrow in D–G) in the double mutants. Bars, 0.5 mm.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1105
this has not been demonstrated in vivo in the spinal cord.
Regarding this, it may be noteworthy that the forebrain and
midbrain have a considerably larger alar plate area than the
spinal cord, which dramatically expands during this
patterning period. Consequently, the floor and basal plates
become located more distant from the roof plate, a tissue
also implicated in the dorsoventral patterning of the CNS.
Thus, the graded Shh signaling, together with differential
recruitment of Gli factors, may play more prominent roles in
establishing the dorsoventral patterning in these regions of
the CNS.
Gli genes are differentially expressed in the mitotic
progenitor cells of the chick DT (Fig. 5A–F), and therefore
could serve as a ‘pre-pattern’ for the action of Shh at the
neurogenic period when the Sox14- and Gbx2-positive
neurons are born. In fact, differential Gli expression
persisted in the type B explants from which the Shh
expressing regions had been excluded (see Fig. 3F), and the
induction of Sox14 by Shh-N appeared somewhat correlated
with the pattern of Gli1 expression (see Fig. 3C,F). This idea
would not necessarily be argued by the fact that Sox14 can
be induced by exogenous SmoM2 in the prospective Gbx2
territory (see Fig. 3D,P), because the expression of Gli
genes can also be regulated by Shh signaling. Transcrip-
tional activation of Gli1 by Shh has been shown previously
(Marigo et al., 1996; Grindley et al., 1997; Hynes et al.,
1997; Lee et al., 1997; Bai et al., 2002; Karlstrom et al.,
2003). We also found that a high dose of Shh or SmoM2
induced Gli1 and repressed Gli2, whereas a low dose
upregulated Gli2 expression (Fig. 3G; KH and KS,
unpublished).
Nevertheless, the Gli1-expressing domain appears con-
siderably broader than that of Sox14, even extending into the
Gbx2 domain (see Fig. 5D). Thus, the ‘Gli code’ does not
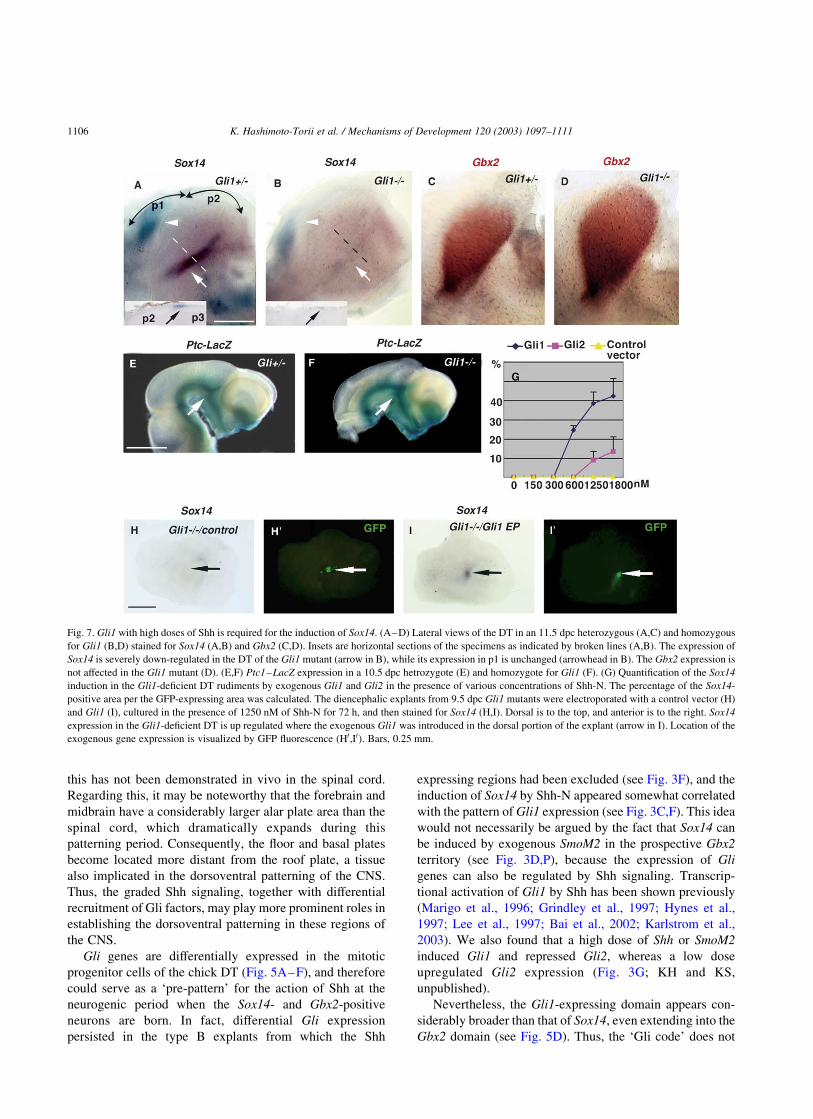
Fig. 7. Gli1 with high doses of Shh is required for the induction of Sox14. (A–D) Lateral views of the DT in an 11.5 dpc heterozygous (A,C) and homozygous
for Gli1 (B,D) stained for Sox14 (A,B) and Gbx2 (C,D). Insets are horizontal sections of the specimens as indicated by broken lines (A,B). The expression of
Sox14 is severely down-regulated in the DT of the Gli1 mutant (arrow in B), while its expression in p1 is unchanged (arrowhead in B). The Gbx2 expression is
not affected in the Gli1 mutant (D). (E,F) Ptc1–LacZ expression in a 10.5 dpc hetrozygote (E) and homozygote for Gli1 (F). (G) Quantification of the Sox14
induction in the Gli1-deficient DT rudiments by exogenous Gli1 and Gli2 in the presence of various concentrations of Shh-N. The percentage of the Sox14-
positive area per the GFP-expressing area was calculated. The diencephalic explants from 9.5 dpc Gli1 mutants were electroporated with a control vector (H)
and Gli1 (I), cultured in the presence of 1250 nM of Shh-N for 72 h, and then stained for Sox14 (H,I). Dorsal is to the top, and anterior is to the right. Sox14
expression in the Gli1-deficient DT is up regulated where the exogenous Gli1 was introduced in the dorsal portion of the explant (arrow in I). Location of the
exogenous gene expression is visualized by GFP fluorescence (H0,I0). Bars, 0.25 mm.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111106

seem to be solely responsible for defining the position of the
Sox14-positive neurons. Concerning this issue, it is inter-
esting that Gli1 and Gli2 may have different thresholds of
Shh dose to function. Although the previous studies have
reported that Gli1 activity is not post-translationally but
transcriptionally regulated by Hh signaling (Epstein et al.,
1996; Marigo et al., 1996; Dai et al., 1999), our in vitro
results suggest that Gli1 requires a high dose of Shh-N to
induces Sox14 (see Fig. 7G). In fact, Sox14 was not
expressed in the type B explants despite the persistent
expression of Gli1 (see Fig. 3A,F). This might explain why
the p2 alar plate was insensitive to the misexpression of
Gli1, and the Sox14 induction solely by exogenous Gli1 was
considerably inefficient in the p1 ðn ¼ 2=9Þ: Since Gbx2 was
induced by a low dose of Shh-N which is likely to be
mediated by Gli2, the lower dose of Shh may be sufficient to
activate Gli2 compared to Gli1. In Drosophila, Ci is the only
Gli factor that mediates Hh signaling (Orenic et al., 1990;
Von Ohlen and Hooper, 1997; Methot and Basler, 2001).
Previous studies have established that the Ci activities are
regulated at the multiple levels, such as proteolysis,
subcellular distribution, and activation, all of which are
dependent on Hh signaling (Aza-Blanc et al., 1997; Wang
and Holmgren, 2000; Methot and Basler, 1999, 2000). Thus
Gli1 and Gli2 may differ in the susceptibility to those Hh-
dependent regulations. Alternatively, the high dose of Shh
may indirectly allow Gli1 to induce Sox14, such as by
repressing antagonistic cues. For instance, it was shown that
Shh blocks Gli3 function to liberate Hh target genes from
repression (Lee et al, 1997; Ruiz i Altaba, 1998; Sasaki et al.,
1999; Von Mering and Basler, 1999; Aza-Blanc et al., 2000;
Litingtung and Chiang, 2000; Persson et al., 2002; Rallu
et al., 2002; Wijgerde et al., 2002). In the present case,
however, Gli3 does not appear to be the one, since Gli3 is
not expressed in the Sox14 territory, and there was in fact no
change in Sox14 expression in the Gli3 mutants. Members
of Bmp family expressed at the dorsal most portion of p2
(Furuta et al., 1997) could be good candidates, as it was
shown that Bmp signaling counteract with Shh signaling
(Liem et al., 2000).
Overall, the differential expression of Gli1 and Gli2
which possess different preference for the target genes, in
cooperation with the graded distribution of Shh protein,
perhaps play a role in assuring the precise birthplaces of the
distinct neuronal subtypes in the DT. The recruitment of
different Gli factors as an effector of Hh signaling must be a
key for further diversity and complexity of the patterns
generated by this signaling pathway during evolution as
discussed previously (Aza-Blanc et al., 2000; Persson et al.,
2002; Karlstrom et al., 2003).
3.2. ‘Ventral patterning’ in the alar plate
Our present finding that Gbx2 expression in the DT
rudiment is dependent on Shh signaling is somewhat
puzzling, as Gbx2 is expressed in and required for a large
region of the DT that is thought to be derived from the alar
plate (Fig. 4; Puelles and Rubenstein, 1993; Miyashita-Lin
et al., 1999). Studies in the spinal cord as well as other brain
territories have established that Shh is involved in the
ventral patterning of the CNS (the basal plate) and the alar
plate derivatives are thought to be negatively regulated by
Shh signaling (Ericson et al., 1996; Watanabe and
Nakamura, 2000). Furthermore, recent findings in the
developing telencephalon have led to the notion that
GABAergic and cholinergic interneurons are born in the
basal telencephalon presumably through the action of Shh,
whereas most glutamatergic neurons are not (reviewed by
Fig. 8. Cross-regulation between Gbx2 and Sox14. Oblique horizontal sections of HH21 embryos at the level of p2 which had been electroporated with a
control vector (A,C,E), mGbx2 (B), Sox14 (D,F) at HH12 were double stained for Sox14 (blue in A,B; brown in C,D) and cGbx2 (brown in A,B; blue in C,D)
which cross-hybridized with mGbx2 RNA, or single stained for Sox2 (E,F). Arrows indicate repression by the exogenous genes. (G,H) Dissected brains from
HH21 embryos electroporated at HH12 with mGbx2 (G) or Sox14 (H) hybridized with Gbx2- (G) or Sox14-30UTR probes (H). Exogenous Gbx2 induced
endogenous Gbx2 ectopically (arrowhead in G). Locations of exogenous gene expression are visualized by GFP fluorescence (F0,G0,H0). Bars, 0.4 mm.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1107

Wilson and Rubenstein, 2000; Corbin et al., 2001). Given
that the majority of DT neurons are glutamatergic
(Shigemoto et al., 1992), the dependence of Gbx2 on Shh
again does not make good sense. However, we found that
some Gbx2 expression, particularly at later stages, is
independent of Shh activity. For instance, DT rudiments
cultured in the presence of anti-Shh antibody did show
substantial Gbx2 expression when cultured for longer
periods (KH and KS, unpublished). Moreover, in Gli2;
Gli3 double mutant mice, Gbx2 expression was detectable
in the DT after 12.5 dpc (KH and KS, unpublished). These
findings raise the possibility that some Gbx2-positive cells
in the DT are independent of Shh activity, and these may
correspond to the glutamatergic DT neurons. Other
molecular markers that further distinguish subsets of cells
within the Gbx2-positive population (e.g. Sox2) will clarify
the precise requirement of Shh for the Gbx2-positive cells in
the DT. On the other hand, the notion for the generation of
GABAergic neurons in the telencephalon is consistent with
the fact that Sox14-positive cells in the DT, which are
dependent on Shh activity, are mostly GABAergic (Hashi-
moto-Torii et al., in preparation).
Thus, it appears that certain populations of DT neurons
are specified by Shh emanating from the basal plate, while
the DT itself is specified as the alar plate. This evokes an
idea that Shh expression, which spreads in the basal plate of
the anterior CNS (fore-midbrain region), may be respon-
sible for generating further cellular diversity within the alar
plate. In addition, it is conceivable that the ZLI where Shh
becomes expressed as development proceeds also contri-
butes to the further complexity in the DT as postulated
(Larsen et al., 2001; Zeltser et al., 2001). It has been shown
that the notochord plays a primary role in the ventral
specification of the spinal cord, which is then taken over by
the floor plate (Ericson et al., 1996). It could be that, in the
fore-midbrain region, signals including Shh emanating from
the axial mesoderm, the prechordal plate and notochord,
first specify the floor and basal plates, and subsequently Shh
released from the basal plate patterns the adjacent alar plate
cells. The initial Shh signaling from the axial mesoderm
might also set up the Gli patterns which serves as a pre-
pattern in the alar plate for the subsequent Shh signal from
the basal plate during the neurogenic periods as discussed
above.
4. Experimental procedures
4.1. Embryo manipulation
Embryological manipulations were performed in White
Leghorn chick embryos following standard protocols. Eggs
were incubated at 38 8C and the embryos were staged
according to Hamburger and Hamilton (Hamburger and
Hamilton, 1951).
4.2. Explant culture
In vitro culture of isolated brain fragments was carried
out as described previously (Shimamura and Takeichi,
1992), with slight modifications. Explants were isolated
from HH8-15 chick and 9.5–10 dpc mouse embryos and
cultured on the Nucleoporew filters (#110414; Costar)
floating on DMEM supplemented with 10% fetal bovine
serum (Nissui) in a CO2 incubator at 37 8C for 0.5–4 days.
No significant necrosis or obvious abnormalities in
morphogenesis were detected (see Figs. 2,4). For the Shh
titration experiments, recombinant mouse Shh-N (R and D
systems) was added to the culture media at 0–1800 nM. For
blocking of the Shh activity, monoclonal anti-Shh antibody
(5E1; DSHB, Iowa) was used (Ericson et al., 1996; Gunhaga
et al., 2000).
4.3. Electroporation
Full-length cDNA encoding mouse Gbx2 (a gift from Dr
G. Martin), chick Sox14 (a gift from Dr H. Kondoh), and a
constitutively active form of human Smoothened (SmoM2; a
gift from Dr A. Rosenthal) were inserted into the pCAGGS
vector (Tokui et al., 1997). A cDNA encoding the N-
terminal form of mouse Shh was inserted into the pEF-BOS
vector. Full-length mouse Gli1 and Gli2 cDNAs inserted
into the pCDNA3.1 vector were provided by Dr H. Sasaki
(Sasaki et al., 1999). Electroporation for HH8-14 chick
embryos was carried out in ovo as described (Funahashi
et al., 1999; Kobayashi et al., 2002). To achieve precise
control of DNA concentration and of transfection sites, a
method for microelectroporation using a microcapillary
electrode was employed for the explants (Haas et al., 2001).
Briefly, a pore of 0.03–4 mm diameter glass needle was
made from a glass capillary (Sutter, BF100-50-10) by a
Sutter P-97/IVF. A DNA solution was loaded into the tip of
glass needle and backfilled with mineral oil. A thin platinum
wire was inserted into the needle to function as a cathode.
For all the electroporation experiments, pCAGGS-GFP
plasmid (Momose et al., 1999) was mixed to 1 mg/ml, so
that the sites of exogenous gene expression can be
monitored under an epi-fluorescence dissecting microscope
(Leica MZFL3).
4.4. Mice
Mutant mouse strains, Ptc1 LacZ (Goodrich et al., 1997),
Gli1 zfd (Matise et al., 1998), Gli2 zfd (Mo et al., 1997; Ding
et al., 1998) and Gli3 XtJ (Johnson, 1967; Buscher et al.,
1997) were maintained in a mixed 129/Sv and CD1
background. Generation and analysis of Gli2; Gli3 double
mutants were performed as described (Mo et al., 1997).
To generate Gli2 zfd/zfd; Gli3 XtJ/XtJ; Ptc1 þ/LacZ mice, the
Gli2 þ/zfd; Gli3 þ/XtJ; Ptc1 þ/LacZ male mice were crossed
with Gli2 þ/zfd; Gli3 þ/XtJ female mice. Genotypes of mutant
embryos were determined by PCR using a standard
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111108

protocol. For genotyping the Gli1 mutant, GlidF
(50TTTGAAGGCTGTCGGAAGTCC30) and GlidR
(50TCATTGGAGTGGGTCCGATTC30) primers for ampli-
fication of the 165 bp fragment derived from the zinc finger
domain, as well as the standard primers for detection of Neo
cassette were used.
4.5. In situ hybridization
In situ hybridization of whole-mount specimens and
cryosectioned materials were performed as described
previously (Shimamura et al., 1995; Uchikawa et al.,
1999). Probes for mSox14 and cGli3 were obtained by
PCR. Probes for cSox2, cSox14, mGbx2, cGbx2, cShh,
cGli1, cGli2 and mShh were kind gifts from Drs H. Kondoh,
G. Martin, C. Tabin, and A. McMahon, respectively. Probes
for cSox14- and cGbx2-30 UTR were generated from the
Sac II–Apa I and HincII–Eco RI fragments, respectively.
For quantification of the labeled cells, NIH image software
was used as described previously (Ishihara et al., 2001).
Since cells expressing Sox14 or Gbx2 are distributed two-
dimensionally at the time of their emergence, pixels of the
labeled areas with an intensity above a given optical
thresholds were scored as the references for the number of
positive cells. When necessary, proportions of the labeled
areas to the defined fields of the explants were calculated as
an induction index.
4.6. Histostaining
Immunostaining of cryosections was performed as
described (Yoon et al., 2000). The antibodies used in this
study were monoclonal anti-Tuj1 (BABCO), anti-NeuN
(Chemicon), Texas Red-conjugated and HRP-conjugated
anti-mouse IgG (Jackson Lab.). Cell death was detected by
the TUNEL assay (Gavrieli et al., 1992; Wijsman et al.,
1993), using the Roche Kit as previously described (Teillet
et al., 1998). b-galactosidase staining for Ptc1 expression
was carried out as previously reported (Ding et al., 1998).
4.7. Nomenclature
The neuroanatomical nomenclature for chick brain used
in this study was adopted from previous literature (Redies
et al., 2000; Yoon et al., 2000; Puelles, 2001; Martınez-de-
la-Torre et al., 2002).
Acknowledgements
We are grateful to Drs Christoph Redies, Luis Puelles
and Loreta Medina for insightful discussions, Daisuke
Kobayashi for critical reading of the manuscript. We thank
Drs Alex Joyner, Jun Aruga, and Shigeru Makino for
providing the Gli1 KO mice, and Drs Masanori Uchikawa
and Hisato Kondoh for reagents and technical advice. This
work was supported by grants-in-aid from Ministry of
Education, Science, Sports and Culture of Japan. KH is the
recipient of a Research Fellowships for the Promotion of
Science for Young Scientists from the JSPS.
References
Agarwala, S., Sanders, T.A., Ragsdale, C.W., 2001. Sonic hedgehog control
of size and shape in midbrain pattern formation. Science 291,
2147–2150.
Alcedo, J., Ayzenzon, M., Von Ohlen, T., Noll, M., Hooper, J.E., 1996. The
Drosophila smoothened gene encodes a seven-pass membrane protein, a
putative receptor for the hedgehog signal. Cell 86, 221–232.
Amaya, E., Stein, P.A., Musci, T.J., Kirschner, M.W., 1993. FGF signalling
in the early specification of mesoderm in Xenopus. Development 118,
477–487.
Aza-Blanc, P., Ramirez-Weber, F.A., Laget, M.P., Schwartz, C., Kornberg,
T.B., 1997. Proteolysis that is inhibited by hedgehog targets Cubitus
interruptus protein to the nucleus and converts it to a repressor. Cell 89,
1043–1053.
Aza-Blanc, P., Kornberg, T.B., 1999. Ci: a complex transducer of the
hedgehog signal. Trends Genet. 15, 458–462.
Aza-Blanc, P., Lin, H.Y., Ruiz i Altaba, A., Kornberg, T.B., 2000.
Expression of the vertebrate Gli proteins in Drosophila reveals a
distribution of activator and repressor activities. Development 127,
4293–4301.
Bai, C.B., Auerbach, W., Lee, J.S., Stephen, D., Joyner, A.L., 2002. Gli2,
but not Gli1, is required for initial Shh signaling and ectopic activation
of the Shh pathway. Development 129, 4753–4761.
Buscher, D., Bosse, B., Heymer, J., Ruther, U., 1997. Evidence for genetic
control of Sonic hedgehog by Gli3 in mouse limb development. Mech.
Dev. 62, 175–182.
Corbin, J.G., Nery, S., Fishell, G., 2001. Telencephalic cells take a tangent:
non-radial migration in the mammalian forebrain. Nat. Neurosci. 1,
1177–1182.
Dai, P., Akimaru, H., Tanaka, Y., Maekawa, T., Nakafuku, M., Ishii, S.,
1999. Sonic Hedgehog-induced activation of the Gli1 promoter is
mediated by GLI3. J. Biol. Chem. 274, 8143–8152.
Ding, Q., Motoyama, J., Gasca, S., Mo, R., Sasaki, H., Rossant, J., Hui,
C.C., 1998. Diminished Sonic hedgehog signaling and lack of floor
plate differentiation in Gli2 mutant mice. Development 125,
2533–2543.
Echevarria, D., Vieira, C., Martinez, S., 2001. Mammalian neural tube
grafting experiments: an in vitro system for mouse experimental
embryology. Int. J. Dev. Biol. 45, 895–902.
Epstein, D.J., Martı, E., Scott, M.P., McMahon, A.P., 1996. Antagonizing
cAMP-dependent protein kinase A in the dorsal CNS activates a
conserved Sonic hedgehog signaling pathway. Development 122,
2885–2894.
Ericson, J., Morton, S., Kawakami, A., Roelink, H., Jessell, T.M., 1996.
Two critical periods of Sonic Hedgehog signaling required for the
specification of motor neuron identity. Cell 87, 661–673.
Ericson, J., Rashbass, P., Schedl, A., Brenner-Morton, S., Kawakami, A.,
van Heyningen, V., Jessell, T.M., Briscoe, J., 1997. Pax6 controls
progenitor cell identity and neuronal fate in response to graded Shh
signaling. Cell 90, 169–180.
Funahashi, J., Okafuji, T., Ohuchi, H., Noji, S., Tanaka, H., Nakamura, H.,
1999. Role of Pax-5 in the regulation of a mid-hindbrain organizer’s
activity. Dev. Growth Differ. 41, 59–72.
Furuta, Y., Piston, D.W., Hogan, B.L., 1997. Bone morphogenetic proteins
(BMPs) as regulators of dorsal forebrain development. Development
124, 2203–2212.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1109

Garda, A.L., Echevarrıa, D., Martınez, S., 2001. Neuroepithelial co-
expression of Gbx2 and Otx2 precedes Fgf8 expression in the isthmic
organizer. Mech. Dev. 101, 111–118.
Gavrieli, Y., Sherman, Y., Ben-Sasson, S.A., 1992. Identification of
programmed cell death in situ via specific labeling of nuclear DNA
fragmentation. J. Cell. Biol. 119, 493–501.
Goodrich, L.V., Milenkovic, L., Higgins, K.M., Scott, M.P., 1997. Altered
neural cell fates and medulloblastoma in mouse patched mutants.
Science 277, 1109–1113.
Grindley, J.C., Bellusci, S., Perkins, D., Hogan, B.L., 1997. Evidence for
the involvement of the Gli gene family in embryonic mouse lung
development. Dev. Biol. 188, 337–348.
Grove, E.A., Tole, S., Limon, J., Yip, L., Ragsdale, C.W., 1998. The hem of
the embryonic cerebral cortex is defined by the expression of multiple
Wnt genes and is compromised in Gli3-deficient mice. Development
125, 2315–2325.
Gunhaga, L., Jessell, T.M., Edlund, T., 2000. Sonic hedgehog signaling at
gastrula stages specifies ventral telencephalic cells in the chick embryo.
Development 127, 3283–3293.
Haas, K., Sin, W.C., Javaherian, A., Li, Z., Cline, H.T., 2001. Single-cell
electroporation for gene transfer in vivo. Neuron 29, 583–591.
Hamburger, V., Hamilton, H.J., 1951. A series of normal stages in the
development of the chick embryo. J. Morphol. 94, 257.
Hargrave, M., Karunaratne, A., Cox, L., Wood, S., Koopman, P., Yamada,
T., 2000. The HMG box transcription factor gene Sox14 marks a novel
subset of ventral interneurons and is regulated by sonic hedgehog. Dev.
Biol. 219, 142–153.
Hui, C.C., Slusarski, D., Platt, K.A., Holmgren, R., Joyner, A.L., 1994.
Expression of three mouse homologs of the Drosophila segment
polarity gene cubitus interruptus, Gli, Gli-2, and Gli-3, in ectoderm- and
mesoderm-derived tissues suggests multiple roles during postimplanta-
tion development. Dev. Biol. 162, 402–413.
Hynes, M., Stone, D.M., Dowd, M., Pitts-Meek, S., Goddard, A., Gurney,
A., Rosenthal, A., 1997. Control of cell pattern in the neural tube by the
zinc finger transcription factor and oncogene Gli-1. Neuron 19, 15–26.
Hynes, M., Ye, W., Wang, K., Stone, D., Murone, M., Sauvage, F.,
Rosenthal, A., 2000. The seven-transmembrane receptor smoothened
cell-autonomously induces multiple ventral cell types. Nat. Neurosci. 3,
41–46.
Ingham, P.W., McMahon, A.P., 2001. Hedgehog signaling in animal
development: paradigms and principles. Genes Dev. 15, 3059–3087.
Ishihara, T., Araki, T., Sakuma, Y., 2001. Two distinct populations of
neurons expressing nitric oxide synthase mRNA in the female rat
preoptic area: site specific changes induced by sex steroids. J. Nippon
Med. Sch. 68, 328–334.
Jessell, T.M., 2000. Neuronal specification in the spinal cord: inductive
signals and transcriptional codes. Nat. Rev. Genet. 1, 20–29.
Jessell, T.M., Sanes, J.R., 2000. Development. The decade of the
developing brain. Curr. Opin. Neurobiol. 10, 599–611.
Johnson, D.R., 1967. Extra-toes: a new mutant gene causing multiple
abnormalities in the mouse. J. Embryol. Exp. Morphol. 175, 43–81.
Jones, E., 1985. The Thalamus, Plenum, New York.
Jones, E., 1998. A new view of specific and nonspecific thalamocortical
connections. Adv. Neurol. 77, 44–71.
Kamachi, Y., Uchikawa, M., Kondoh, H., 2000. Pairing SOX off: with
partners in the regulation of embryonic development. Trends Genet. 16,
182–187.
Karlstrom, R.O., Tyurina, O.V., Kawakami, A., Nishioka, N., Talbot, W.S.,
Sasaki, H., Schier, A.F., 2003. Genetic analysis of zebrafish gli1 and
gli2 reveals divergent requirements for gli genes in vertebrate
development. Development 130, 1549–1564.
Katahira, T., Sato, T., Sugiyama, S., Okafuji, T., Araki, I., Funahashi, J.,
Nakamura, H., 2000. Interaction between Otx2 and Gbx2 defines the
organizing center for the optic tectum. Mech.Dev. 91, 43–52.
Kinzler, K.W., Vogelstein, B., 1990. The GLI gene encodes a nuclear
protein which binds specific sequences in the human genome. Mol.
Cell. Biol. 10, 634–642.
Kobayashi, D., Kobayashi, M., Matsumoto, K., Ogura, T., Nakafuku, M.,
Shimamura, K., 2002. Early subdivisions in the neural plate define
distinct competence for inductive signals. Development 129, 83–93.
Koebernick, K., Pieler, T., 2002. Gli-type zinc finger proteins as bipotential
transducers of Hedgehog signaling. Differentiation 70, 69–76.
Larsen, C.W., Zeltser, L.M., Lumsden, A., 2001. Boundary formation and
compartition in the avian diencephalon. J. Neurosci. 21, 4699–4711.
Lee, J., Platt, K.A., Censullo, P., Ruiz i Altaba, A., 1997. Gli1 is a target of
Sonic hedgehog that induces ventral neural tube development.
Development 124, 2537–2552.
Liem, K.F. Jr., Jessell, T.M., Briscoe, J., 2000. Regulation of the neural
patterning activity of sonic hedgehog by secreted BMP inhibitors
expressed by notochord and somites. Development 127, 4855–4866.
Litingtung, Y., Chiang, C., 2000. Specification of ventral neuron types is
mediated by an antagonistic interaction between Shh and Gli3. Nat.
Neurosci. 3, 979–985.
Lumsden, A., Krumlauf, R., 1996. Patterning the vertebrate neuraxis.
Science 274, 1109–1115.
Marigo, V., Johnson, R.L., Vortkamp, A., Tabin, C.J., 1996. Sonic
hedgehog differentially regulates expression of GLI and GLI3 during
limb development. Dev. Biol. 180, 273–283.
Martı, E., Takada, R., Bumcrot, D.A., Sasaki, H., McMahon, A.P., 1995.
Distribution of Sonic hedgehog peptides in the developing chick and
mouse embryo. Development 121, 2537–2547.
Martınez-de-la-Torre, M., Garda, A.L., Puelles, E., Puelles, L., 2002. Gbx2
expression in the late embryonic chick dorsal thalamus. Brain Res. Bull.
57, 435–438.
Matise, M.P., Epstein, D.J., Park, H.L., Platt, K.A., Joyner, A.L., 1998. Gli2
is required for induction of floor plate and adjacent cells, but not most
ventral neurons in the mouse central nervous system. Development 125,
2759–2770.
Methot, N., Basler, K., 1999. Hedgehog controls limb development by
regulating the activities of distinct transcriptional activator and
repressor forms of Cubitus interruptus. Cell 96, 819–831.
Methot, N., Basler, K., 2001. An absolute requirement for Cubitus
interruptus in Hedgehog signaling. Development 128, 733–742.
Millet, S., Campbell, K., Epstein, D.J., Losos, K., Harris, E., Joyner, A.L.,
1999. A role for Gbx2 in repression of Otx2 and positioning the mid/
hindbrain organizer. Nature 401, 161–164.
Miyashita-Lin, E.M., Hevner, R., Wassarman, K.M., Martinez, S.,
Rubenstein, J.L., 1999. Early neocortical regionalization in the absence
of thalamic innervation. Science 285, 906–909.
Mo, R., Freer, A.M., Zinyk, D., Crackower, M.A., Heng, H.H.Q., Chik,
K.W., et al., 1997. Specific and redundant functions of Gli2 and Gli3
zinc finger genes in skeletal patterning and development. Development
124, 113–123.
Momose, T., Tonegawa, A., Takeuchi, J., Ogawa, H., Umesono, K.,
Yasuda, K., 1999. Efficient targeting of gene expression in chick
embryos by microelectroporation. Dev. Growth Differ. 41, 335–344.
Motoyama, J., Liu, J., Mo, R., Ding, Q., Post, M., Hui, C.C., 1998. Essential
function of Gli2 and Gli3 in the formation of lung, trachea and
oesophagus. Nat. Genet. 20, 54–57.
Nakagawa, Y., O’Leary, D.D., 2001. Combinatorial expression patterns of
LIM-homeodomain and other regulatory genes parcellate developing
thalamus. J. Neurosci. 21, 2711–2725.
Nybakken, K., Perrimon, N., 2002. Hedgehog signal transduction: recent
findings. Curr. Opin. Genet. Dev. 12, 503–511.
Orenic, T.V., Slusarski, D.C., Kroll, K.L., Holmgren, R.A., 1990. Cloning
and characterization of the segment polarity gene cubitus interruptus
Dominant of Drosophila. Genes Dev. 4, 1053–1067.
Park, H.L., Bai, C., Platt, K.A., Matıse, M.P., Beeghly, A., Hui, C.C.,
Nakashima, M., Joyner, A.L., 2000. Mouse Gli1 mutants are viable but
have defects in SHH signaling in combination with a Gli2 mutation.
Development 127, 1593–1605.
Persson, M., Stamataki, D., te Welscher, P., Andersson, E., Bose, J., Ruther,
U., Ericson, J., Briscoe, J., 2002. Dorsal-ventral patterning of the spinal
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–11111110

cord requires Gli3 transcriptional repressor activity. Genes Dev. 16,
2865–2878.
Puelles, L., Rubenstein, J.L., 1993. Expression patterns of homeobox and
other putative regulatory genes in the embryonic mouse forebrain
suggest a neuromeric organization. Trends Neurosci. 16, 472–479.
Puelles, L., 2001. Thoughts on the development, structure and evolution of
the mammalian and avian telencephalic pallium. Philos. Trans. R. Soc.
Lond. B Biol. Sci. 356, 1583–1598.
Rallu, M., Machold, R., Gaiano, N., Corbin, J.G., McMahon, A.P., Fishell,
G., 2002. Dorsoventral patterning is established in the telencephalon of
mutants lacking both Gli3 and Hedgehog signaling. Development 129,
4963–4974.
Redies, C., Ast, M., Nakagawa, S., Takeichi, M., Martınez-de-la-Torre, M.,
Puelles, L., 2000. Morphologic fate of diencephalic prosomeres and
their subdivisions revealed by mapping cadherin expression. J. Comp.
Neurol. 421, 481–514.
Roelink, H., Porter, J.A., Chiang, C., Tanabe, Y., Chang, D.T., Beachy,
P.A., Jessell, T.M., 1995. Floor plate and motor neuron induction by
different concentrations of the amino-terminal cleavage product of
Sonic hedgehog autoproteolysis. Cell 81, 445.
Rubenstein, J.L., Shimamura, K., Martinez, S., Puelles, L., 1998.
Regionalization of the prosencephalic neural plate. Annu. Rev.
Neurosci. 21, 445–477.
Ruiz i Altaba, A., 1998. Combinatorial Gli gene function in floor plate and
neuronal inductions by Sonic hedgehog. Development 125,
2203–2212.
Ruiz i Altaba, A., 1999. Gli proteins encode context-dependent positive and
negative functions: implications for development and disease. Devel-
opment 126, 3205–3216.
Ruiz i Altaba, A., Palma, V., Dahmane, N., 2002. Hedgehog-Gli signalling
and the growth of the brain. Nat. Rev. Neurosci. 3, 24–33.
Sasaki, H., Hui, C., Nakafuku, M., Kondoh, H., 1997. A binding site for Gli
proteins is essential for HNF-3beta floor plate enhancer activity in
transgenics and can respond to Shh in vitro. Development 124,
1313–1322.
Sasaki, H., Nishizaki, Y., Hui, C., Nakafuku, M., Kondoh, H., 1999.
Regulation of Gli2 and Gli3 activities by an amino-terminal repression
domain: implication of Gli2 and Gli3 as primary mediators of Shh
signaling. Development 126, 3915–3924.
Shigemoto, R., Nakanishi, S., Mizuno, N., 1992. Distribution of the mRNA
for a metabotropic glutamate receptor (mGluR1) in the central nervous
system: an in situ hybridization study in adult and developing rat.
J. Comp. Neurol. 322, 121–135.
Shimamura, K., Takeichi, M., 1992. Local and transient expression of E-
cadherin involved in mouse embryonic brain morphogenesis. Devel-
opment 116, 1011–1019.
Shimamura, K., Hartigan, D.J., Martinez, S., Puelles, L., Rubenstein, J.L.,
1995. Longitudinal organization of the anterior neural plate and neural
tube. Development 121, 3923–3933.
Teillet, M., Watanabe, Y., Jeffs, P., Duprez, D., Lapointe, F., Le Douarin,
N.M., 1998. Sonic hedgehog is required for survival of both myogenic
and chondrogenic somitic lineages. Development 125, 2019–2030.
Tokui, M., Takei, I., Tashiro, F., Shimada, A., Kasuga, A., Ishii, M., Ishii,
T., Takatsu, K., Saruta, T., Miyazaki, J., 1997. Intramuscular injection
of expression plasmid DNA is an effective means of long-term systemic
delivery of interleukin-5. Biochem. Biophys. Res. Commun. 233,
527–531.
Uchikawa, M., Kamachi, Y., Kondoh, H., 1999. Two distinct subgroups of
Group B Sox genes for transcriptional activators and repressors: their
expression during embryonic organogenesis of the chicken. Mech. Dev.
84, 103–120.
van den Heuvel, M., Ingham, P.W., 1996. smoothened encodes a receptor-
like serpentine protein required for hedgehog signalling. Nature 382,
547–551.
Von Mering, C., Basler, K., 1999. Distinct and regulated activities of
human Gli proteins in Drosophila. Curr. Biol. 9, 1319–1322.
Von Ohlen, T., Hooper, J.E., 1997. Hedgehog signaling regulates
transcription through Gli/Ci binding sites in the wingless enhancer.
Mech. Dev. 68, 149–156.
Vortkamp, A., Gessler, M., Grzeschik, K.H., 1995. Identification of
optimized target sequences for the GLI3 zinc finger protein. DNA Cell.
Biol. 14, 629–634.
Wang, Q.T., Holmgren, R.A., 2000. Nuclear import of cubitus interruptus is
regulated by hedgehog via a mechanism distinct from Ci stabilization
and Ci activation. Development 127, 3131–3139.
Watanabe, Y., Nakamura, H., 2000. Control of chick tectum territory along
dorsoventral axis by Sonic hedgehog. Development 127, 1131–1140.
Wijgerde, M., McMahon, J.A., Rule, M., McMahon, A.P., 2002. A direct
requirement for Hedgehog signaling for normal specification of all
ventral progenitor domains in the presumptive mammalian spinal cord.
Genes Dev. 16, 2849–2864.
Wijsman, J.H., Jonker, R.R., Keijzer, R., van de Velde, C.J., Cornelisse,
C.J., van Dierendonck, J.H., 1993. A new method to detect apoptosis in
paraffin sections: in situ end-labeling of fragmented DNA. J. Histochem.
Cytochem. 41, 7–12.
Wilson, S.W., Rubenstein, J.L., 2000. Induction and dorsoventral
patterning of the telencephalon. Neuron 28, 641–651.
Xie, J., Murone, M., Luoh, S.M., Ryan, A., Gu, Q., Zhang, C., Bonifas,
J.M., Lam, C.W., Hynes, M., Goddard, A., et al., 1998. Activating
Smoothened mutations in sporadic basal-cell carcinoma. Nature 391,
90–92.
Yoon, M.S., Puelles, L., Redies, C., 2000. Formation of cadherin-
expressing brain nuclei in diencephalic alar plate divisions. J. Comp.
Neurol. 427, 461–480.
Zeltser, L.M., Larsen, C.W., Lumsden, A., 2001. A new developmental
compartment in the forebrain regulated by Lunatic fringe. Nat.
Neurosci. 4, 683–684.
K. Hashimoto-Torii et al. / Mechanisms of Development 120 (2003) 1097–1111 1111
Related Documents











