BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofit publishers, academic institutions, research libraries, and research funders in the common goal of maximizing access to critical research. Evaluation of Potential Versus Realized Primary Infection of Gypsy Moth (Lepidoptera: Lymantriidae) by Entomophaga Maimaiga (Zygomycetes: Entomophthorales) Author(s): Nathan W. Siegert, Deborah G. Mccullough, Micheal M. Wheeler, and Ann E. Hajek Source: Environmental Entomology, 41(5):1115-1124. 2012. Published By: Entomological Society of America DOI: http://dx.doi.org/10.1603/EN12137 URL: http://www.bioone.org/doi/full/10.1603/EN12137 BioOne (www.bioone.org ) is a nonprofit, online aggregation of core research in the biological, ecological, and environmental sciences. BioOne provides a sustainable online platform for over 170 journals and books published by nonprofit societies, associations, museums, institutions, and presses. Your use of this PDF, the BioOne Web site, and all posted and associated content indicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/ terms_of_use . Usage of BioOne content is strictly limited to personal, educational, and non-commercial use. Commercial inquiries or rights and permissions requests should be directed to the individual publisher as copyright holder.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

BioOne sees sustainable scholarly publishing as an inherently collaborative enterprise connecting authors, nonprofitpublishers, academic institutions, research libraries, and research funders in the common goal of maximizing access tocritical research.
Evaluation of Potential Versus Realized Primary Infection ofGypsy Moth (Lepidoptera: Lymantriidae) by EntomophagaMaimaiga (Zygomycetes: Entomophthorales)Author(s): Nathan W. Siegert, Deborah G. Mccullough, Micheal M. Wheeler, andAnn E. HajekSource: Environmental Entomology, 41(5):1115-1124. 2012.Published By: Entomological Society of AmericaDOI: http://dx.doi.org/10.1603/EN12137URL: http://www.bioone.org/doi/full/10.1603/EN12137
BioOne (www.bioone.org) is a nonprofit, online aggregation of core research in thebiological, ecological, and environmental sciences. BioOne provides a sustainable onlineplatform for over 170 journals and books published by nonprofit societies, associations,museums, institutions, and presses.
Your use of this PDF, the BioOne Web site, and all posted and associated contentindicates your acceptance of BioOne’s Terms of Use, available at www.bioone.org/page/terms_of_use.
Usage of BioOne content is strictly limited to personal, educational, and non-commercialuse. Commercial inquiries or rights and permissions requests should be directed to theindividual publisher as copyright holder.

BIOLOGICAL CONTROL-MICROBIALS
Evaluation of Potential Versus Realized Primary Infection of GypsyMoth (Lepidoptera: Lymantriidae) by Entomophaga maimaiga
(Zygomycetes: Entomophthorales)
NATHAN W. SIEGERT,1,2 DEBORAH G. MCCULLOUGH,1,3 MICHEAL M. WHEELER,4,5
AND ANN E. HAJEK4
Environ. Entomol. 41(5): 1115Ð1124 (2012); DOI: http://dx.doi.org/10.1603/EN12137
ABSTRACT The fungal entomopathogenEntomophagamaimaigahas provided important biologicalcontrol of gypsy moth, Lymantria dispar (L.), since the Þrst epizootics occurred in the northeasternUnited States in 1989. Epizootics are initiated by germination of soil-borne resting spores, which arehighly sensitive to spring temperature and moisture. We compared gypsy moth infection by E.maimaiga in 33 oak stands in Michigan with infection under optimal laboratory conditions from 1999to 2001 to assess differences between potential and realized efÞcacy of E. maimaiga. Field bioassayswere conducted by exposing laboratory-reared, fourth-instar gypsy moth to soil at the base of oak treesfor 4 d. Additional larvae were similarly exposed to soil collected from the Þeld plots in laboratorybioassays with temperature, humidity, and moisture levels optimal for fungal germination. Overall E.maimaiga infection ranged from means of 3.2Ð29.8% in the Þeld compared with 20.9Ð59.7% in thelaboratory during three Þeld seasons. Resting spore density in soil and gypsy moth egg mass densitywere signiÞcant predictors of Þeld infections in two of the 3 yr, whereas resting spore density was asigniÞcant predictor of laboratory infections each year. Other variables that signiÞcantly predictedlaboratory infections in one of the 3 yr included egg mass density, canopy cover, and soil pH. Inlaboratory bioassays, soil pH and E. maimaiga resting spore density were positively associated withincreasing E. maimaiga infection rates of gypsy moth larvae.
KEY WORDS entomopathogen, Entomophaga maimaiga, fungal epizootic, gypsy moth, soil pH
The fungal pathogen Entomophaga maimaiga Hum-ber, Shimazu and Soper (Zygomycetes: Entomoph-thorales) has been responsible for signiÞcant declinesin gypsy moth, Lymantria dispar (L.) (Lepidoptera:Lymantriidae), defoliation in the northeastern UnitedStates since 1989 (Andreadis and Weseloh 1990, Hajeket al. 1990, Hajek 1999). Its compatibility with othernatural enemies, including the gypsy moth nucleopo-lyhedrovirus (NPV) (Malakar et al. 1999b), and highhost speciÞcity (Hajek et al. 1996a, 2000a, 2004a) makeE. maimaiga a desirable biological control agent formanaging gypsy moth. Consequently, in addition tonatural spread, E. maimaiga has been introduced intomany areas with established gypsy moth populations(Smitley et al. 1995, Hajek et al. 1996b, Hajek andTobin 2011).
Gypsy moth larvae become infected when E. mai-maiga spores adhere to the cuticle, then gain entry tothe host by using a combination of mechanical pres-sure and enzymatic degradation (Hajek 1999).E.mai-maiga resting spores (i.e., azygospores) overwinter inthe soil, with the greatest density of resting sporesoccurring in theorganic layerof soil at thebaseof trees(Hajek et al. 1998a). Germination of soil-borne E.maimaiga resting spores in spring is closely synchro-nized with gypsy moth phenology (Hajek and Hum-ber 1997, Weseloh and Andreadis 1997). These sporesinfect early-instar gypsy moth as they move betweenthe tree canopy and the ground (i.e., primary infec-tion) (Hajek 2001). Resting spore germination, how-ever, is highly sensitive to spring temperature andmoisture conditions (Hajek and Humber 1997, Siegertet al. 2008). When infected early-instar larvae die, E.maimaiga produces external conidiophores on gypsymoth cadavers that discharge conidia that infect mid-to late-instar gypsy moth (i.e., secondary infection)(Hajek 1997a). Infected late-instar gypsy moth cadav-ers principally produce resting spores and usually arefound attached to lower tree trunks by their prolegswith their heads oriented downwards (Hajek andSoper 1991, Hajek et al. 1998b). These cadavers dropto the base of the tree, decompose, and release resting
1 Department of Entomology, Michigan State University, East Lan-sing, MI 48824.
2 Corresponding author: Nathan W. Siegert, US Forest Service,Northeastern Area State and Private Forestry, Forest Health Protec-tion, 271 Mast Rd., Durham, NH 03824 (e-mail: [email protected]).
3 Department of Forestry, Michigan State University, East Lansing,MI 48824.
4 Department of Entomology, Cornell University, Ithaca, NY 14853.5 Current address: USDAÐARS, Biological integrated pest manage-
ment (IPM) Research Unit, Ithaca, NY 14853.
0046-225X/12/1115Ð1124$04.00/0 � 2012 Entomological Society of America
spores that remain in the soil until the next spring. Aportion of the E. maimaiga resting spores, however,remain dormant and can remain viable for 10 yr (We-seloh and Andreadis 2002) or even longer in the soil(Hajek et al. 2000b).
The potential effectiveness of E. maimaiga as a re-liable biological control agent, however, may be lim-ited by climatic conditions in some regions of theUnited States where gypsy moth is already establishedas well as in some areas with suitable forest types thatgypsy moth has not yet invaded (Siegert et al. 2009).Gypsy moth outbreaks are not necessarily suppressedconsistently in areas whereE.maimaiga is known to beestablished (Siegert et al. 2009) and there is a need toidentify and understand the factors that drive its prev-alence from year to year. Several manipulative exper-iments have demonstratedE.maimaiga infection ratesmay be affected by moisture availability (Hajek andRoberts 1991, Weseloh and Andreadis 1992, Hajek etal. 1996b) and extensive E. maimaiga epizootics havebeen associated with above average rainfall (Andrea-dis and Weseloh 1990, Hajek 1999, Webb et al. 1999).
The goal of this study was to assess differencesbetween potential and realized efÞcacy of the E. mai-maiga entomopathogen. Our Þrst objective was to
compare primary infection of gypsy moth larvae by E.maimaiga under Þeld conditions in Michigan withprimary infection levels under laboratory conditionsthat were optimal for fungal germination. We hypoth-esized that E. maimaiga infection levels would belower in Þeld bioassays than in laboratory bioassaysbecause of the inherent variability of meteorologicalconditions experienced by the climate-sensitive rest-ing spores in the Þeld. Our second objective was toexamine the association of several site-related factors,including percentage of canopy cover, total basal area,soil pH, gypsy moth egg mass density, and restingspore density in the soil, with E. maimaiga infectionunder Þeld conditions and optimal laboratory condi-tions.
Materials and Methods
Study Sites and Field Measurements. Thirty-twooak-dominated stands, each �10 ha in size, were se-lected in Michigan in 1999 (Fig. 1). All stands had atleast one documentedE.maimaiga epizootic between1993 and 1998 (Buss 1997; D.G.M., unpublished data).An additional stand that experienced a large-scale E.maimaiga epizootic in 1999 (N.W.S., unpublished
Fig. 1. Location of E. maimaiga Þeld sites (denoted by circle with dot) in Michigan, 1999Ð2001.
1116 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 5

data) was included in the study in 2000 and 2001.Stands were located on public land (Huron-ManisteeNational Forest, Au Sable State Forest, Lake SuperiorState Forest, Pere Marquette State Forest and Mich-igan State University, W.K. Kellogg Experimental For-est) and were separated by at least 5 km. Within eachstand, we established a plot center and selected adominant oak tree at 10, 25, and 50 m along transectsin each cardinal direction from the plot center (i.e., 12sample trees per plot). Overall, sample trees (n� 396)had a mean � SE diameter at breast height of 38.4 �0.6 cm.
Stands were characterized by a dominant mixed oak(Quercus spp.) overstory and well-drained soils witha thin organic layer. Understory vegetation rangedfrom sparse to moderately dense and primarily con-sisted of mixed oak, witch-hazel (Hamamelis virgini-anaL.), red maple (Acer rubrumL.), white pine (Pinusstrobus L.), black cherry (Prunus serotina Ehrh.), sas-safras [Sassafras albidum (Nutt.) Nees], and service-berry[Amelanchierarborea(Michaux f.)Fern.](Sieg-ert 2004). Ground ßora tended to be moderately denseand most commonly included bracken fern [Pteridiumaquilinum (L.) Kuhn], grasses and sedges (Gramineaeand Cyperaceae), red maple and mixed oak regener-ation, and low sweet blueberry (Vaccinium angustifo-lium Aiton) (Siegert 2004).
Several site-related characteristics that could po-tentially inßuence E. maimaiga infection of gypsymoth larvae in Þeld or laboratory bioassays were mea-sured at the 33 Þeld plots, including percentage ofcanopy cover, total basal area, soil pH, gypsy moth eggmass density, and E. maimaiga resting spore density.Percentage of canopy cover was measured in the fourcardinal directions at each plot center with a concavespherical densiometer (Lemmon Forest Densiom-eters, Bartlesville, OK) and was averaged per plot foranalyses. Total basal area was estimated with a 10-factor wedge prism at the center of each plot. Soil pHwas measured with a handheld pH meter (WTW Mea-surement Systems, Inc., Ft. Myers, FL) from homog-enized soil collected from the top 2Ð3 cm of the north-ern and southern aspects at the bases of the 12 sampletrees in each plot. Density of the gypsy moth popu-lation in each plot was quantiÞed annually by aver-aging counts of egg masses in two to four 0.01-haÞxed-radius plots (Kolodny-Hirsch 1986). In addition,we assessed whetherE.maimaiga infection in Þeld andlaboratory bioassays varied with trends in gypsy mothpopulations at each site. Gypsy moth populations werecategorized as generally decreasing, constant, increas-ing or variable (i.e., decreasing from 1999 to 2000 thenincreasing again in 2001), from 1999 to 2001 at the 33plots.
Soil from the plots was collected each year and usedfor laboratory bioassays andE.maimaiga resting sporequantiÞcation (see Laboratory Bioassays section). Asoil sample, �85 cm3, was collected from the upper 2Ð3cm at the base of each sample tree, where the highestlevels of resting spores occur (Hajek et al. 1998a),using a modiÞed bulb planter. Soil samples were col-lected as cages of gypsy moth larvae were placed at the
bases of the sample trees for Þeld bioassays (see be-low). To avoid inadvertently transportingE.maimaigaresting spores between Þeld plots, disposable nonlatexgloves (Medline Industries, Inc., Mundelein, IL) andboot covers (McKesson General Medical Corpora-tion, Richmond, VA) were used in each plot. Allequipment used in the plots was sterilized with 95%ethanol and thoroughly rinsed with distilled waterbetween plots. Tools used to collect soil samples wereadditionally sterilized and rinsed between sampleswithin plots. In 1999, soil samples were composited bycardinal direction for each plot (e.g., four merged soilsamples per plot) and securely stored in plastic re-sealable bags. In 2000 and 2001, soil samples werecollected from the northern and southern aspects atthe base of each sample tree and merged by aspect(e.g., two composite soil samples per plot). The ho-mogenized soil samples were transported from theÞeld to the laboratory in coolers with ice packs andstored at 5�C to inhibit E. maimaiga germination be-fore laboratory bioassays and quantiÞcation of restingspores.E.maimaiga resting spores were quantiÞed for each
composite soil sample in 1999 and 2000 to estimate theamount of fungal inoculum present at each plot. Wet-sieving of the soil, followed by density-gradient cen-trifugation by using Percoll (Sigma-Aldrich Company,St. Louis, MO), and microscopy were used to quantifythe number of E. maimaiga resting spores per gram ofdry soil (Hajek and Wheeler 1994). Although restingspores of Entomophthoralean species cannot be dis-tinguished morphologically, we assumed those pres-ent in our samples were predominately E. maimaigagiven that our sites experienced E. maimaiga epizoot-ics within 5 yr of the study. Counts of E. maimaigaresting spores in soil were not conducted in 2001because of the limited availability of resources to com-plete the labor-intensive sample processing.Field Bioassays.To assessE.maimaiga infection rates
under Þeld conditions, we conducted 4-d Þeld bioassayswith freshly molted, fourth-instar gypsy moth. Gypsymoth egg masses were obtained from USDAÐAPHIS,Otis Air National Guard Base, MA, and larvae werereared in May and June each year on artiÞcial diet(OÕDell et al. 1985) at the USDAÐAPHISÐPPQ Bio-logical Control Laboratory, Niles, MI. Larval devel-opment was staggered to ensure sufÞcient numbers offreshly molted fourth instars (�4,000 larvae per day in1999and�2,000 larvaeperday in2000and2001)couldbe collected each morning for the duration of the Þeldbioassays. In total, 62,400 gypsy moth larvae were usedin the Þeld bioassays during this 3-yr study, including30,720 larvae in 1999 and 15,840 larvae in both 2000 and2001.
Timing of Þeld bioassays corresponded to the typ-ical occurrence of fourth-instar gypsy moth in wildpopulations (i.e., early to mid-June in Michigan) tosimulate the timing of naturally occurringE.maimaigainfections. Field bioassays were conducted from 2June to 14 June 1999, 5 June to 17 June 2000, and 6 Juneto 18 June 2001. In each plot, cages containing 20fourth-instar gypsy moth were placed at the base of
October 2012 SIEGERT ET AL.: FIELD VS. LABORATORY INFECTION BY E. maimaiga 1117

the sample trees. Cages were 15 by 20 cm in size andmade of 6- by 7-mesh/cm2 aluminum screening(Hajek and Humber 1997), and were sterilized be-tween uses with 95% ethanol. Two �15-g pieces ofhigh wheat germ artiÞcial diet (OÕDell et al. 1985),sufÞcient to last the duration of a 4-d Þeld bioassay,were placed in each cage. Leaves were brushed awayfrom the base of each sample tree, so cages could beplace directly on the soil surface. There were fourcages per tree in the cardinal directions in 1999 andtwo cages per tree on the northern and southernaspects in 2000 and 2001.
After 4 d in the Þeld, cages of larvae were collected,individually placed into plastic bags to prevent cross-contamination during transport, and returned to theUSDAÐAPHISÐPPQ Biological Control Laboratory.Larvae were removed from the cages and reared in-dividually in 50-ml cups on artiÞcial diet by usingstandard protocols for assessing fungal infections (Pa-pierok and Hajek 1997). Larvae were reared at 20�Cand a photoperiod of 14:10 (L:D) h for 10 d andchecked daily for mortality. Any larvae that died werechecked daily for 3 d for presence or absence of E.maimaiga conidiophores. If conidiophores were pres-ent, then cadavers were transferred to cold storage(4�C and dark). If conidiophores were not present,then cadavers were kept at 20�C in the dark for anadditional 7 d to allow for resting spore maturationbefore being transferred to cold storage. Gypsy mothcadavers were subsequently dissected and examinedwith microscopy to determine whether E. maimaigaresting spores were present. Because of the more rapidmortality from E. maimaiga than NPV after simulta-neous infection (Hajek 1997b; Malakar et al. 1999a, b),larval cadavers found to be co-infected with E. mai-maiga resting spores and NPV polyhedral inclusionbodies were assumed to have died from E. maimaigainfection. Infection rates for E. maimaiga were calcu-lated as the percentage of larvae infected by E. mai-maiga out of the total number of larvae examined (i.e.,total number of cadavers processed plus the numberof larvae that survived to pupation). Quality checks toassess possible laboratory contamination with E. mai-maiga were conducted annually; �300 additionalgypsy moth larvae not exposed to resting spore-ladensoil were reared to pupation without fungal infectionin the laboratory each year.Laboratory Bioassays. Bioassays to measure E. mai-maiga infection under optimal conditions were con-ducted in the laboratory by using soil collected at thebeginning of the Þeld bioassays from the bases of thesample trees in each stand. Homogenized soil sampleswere shipped overnight to Cornell University, Ithaca,NY, in coolers with ice packs to prevent potentialgermination of the E. maimaiga resting spores. AtCornell University, 35 g of soil from each Þeld bioassayplot was placed in a lidded polypropylene cup (4.5 cmin height by 10.5 cm in diameter) and distilled waterwas added so that soil was very moist but not saturated.Ten freshly molted, fourth-instar gypsy moth (rearedon artiÞcial diet from egg masses obtained fromUSDAÐAPHIS, Otis Air National Guard Base, MA)
were placed in the cups and reared at 15�C and aphotoperiod of 14:10 (L:D) h for 4 d (Hajek et al.2004a). These conditions are optimal for E. maimaigaresting spore germination (Shimazu and Soper 1986,Shimazu 1987). A subset of data from 15 of the 33 plotswas analyzed previously by Hajek et al. (2004b) tomeasure whether laboratory bioassays accurately es-timated E. maimaiga resting spore density in soil sam-ples. In the current study, 9,120 gypsy moth larvae intotal were used in the laboratory bioassays from 1999to 2001 (3,840 in 1999; 2,640 in both 2000 and 2001). In1999, 30 gypsy moth larvae per aspect were exposed tosoil collected from each plot (120 larvae per plot; 3,840total larvae). In 2000 and 2001, 40 larvae per aspectwere exposed to soil from each of the 33 plots used forthe Þeld bioassays (80 larvae per plot each year; 2,640total larvae each year). Larvae then were moved tofresh diet and reared to detect fungal infections asdescribed in the Field Bioassays section.Statistical Analysis. Pearson correlation coefÞcients
were calculated to assess the linear relationshipsamong the number ofE.maimaiga resting spores in thesoil for plots between years, percentage of canopycover, total basal area, soil pH, and gypsy moth eggmass densities. Linear regression was used to analyzerelationships between E. maimaiga resting sporecounts and soil pH, E. maimaiga infection under op-timal conditions and soil pH, and E. maimaiga restingspore counts andE.maimaiga infection under optimalconditions. A backward-stepping multiple regressionanalysis was used to analyze effects of site-relatedvariables on the levels ofE.maimaiga infection in Þeldand laboratory bioassays for all plots each year. Pre-dictor variables were removed individually from thefull regression model if their Þt in the model exceeded� � 0.15. Site-related variables used in analyses in-cluded percentage of canopy cover, total basal area,soil pH, gypsy moth egg mass densities, and restingspore densities in the soil. For 2001 analyses, we usedthe average resting spore density from 1999 and 2000.
Analysis of variance (ANOVA) was used to examinewhether site-related factors, including percentage ofcanopy cover, total basal area, soil pH, and average E.maimaiga resting spore density in the soil affectedannual E. maimaiga infection rates under Þeld andlaboratory conditions. ANOVA additionally was ap-plied to assess whether site-related variables variedwith trends in gypsy moth populations. Average Þeldinfection rate was grouped into four categories (i.e.,5Ð10, 10Ð15, 15Ð20, and 20Ð25%) for analysis. Differ-ences in E. maimaiga resting spore counts and infec-tion rates in Þeld and laboratory bioassays amongaspects also were analyzed for each year by usingANOVA. When ANOVA results were signiÞcant, com-parisons among means were made using FischerÕs leastsigniÞcant difference (LSD). Differences in annual E.maimaiga infection levels between laboratory andÞeld bioassays were tested for using the Wilcoxonsigned-rank test (Sokal and Rohlf 1995). All analyseswere conducted using SYSTAT software (SYSTAT2000) at the 0.05 level of signiÞcance.
1118 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 5

SpecimenDeposition.Voucher specimens of gypsymoth larvae used in this project were deposited in theA.J. Cook Arthropod Research Collection, Depart-ment of Entomology, Michigan State University, EastLansing, MI (Siegert 2004). Voucher specimens of E.maimaiga isolates collected from Þeld plots were de-posited in theCollectionofEntomopathogenicFungalCultures,USDAAgriculturalResearchService, Ithaca,NY (Siegert 2004).
Results
Field Plot Characteristics, Gypsy Moth Egg MassDensities, and E. maimaiga Resting Spore Densities.Overall, canopy cover was 90.7 � 0.7% (mean � SE)at the 33 Þeld plots (range � 82.6Ð96.4%) (Table 1).Total basal area, which was dominated by overstoryoaks, ranged from 14 to 30 m2/ha with a mean of 23.8 �0.7 m2/ha (Table 1). Soil pH was strongly acidic at allplots, ranging from 4.1 to 5.4 with an overall mean pHof 4.6 � 0.1 (Table 1).
Gypsy moth egg mass densities were highly variableamong plots each year (Table 1) and overall were attheir highest levels in 1999 with a mean of 1,116 � 497egg masses per hectare. Egg mass densities decreasedto 95 � 33 egg masses per hectare in 2000 beforeincreasing slightly to 197 � 86 egg masses per hectarein 2001. The percentage of plots with �100 egg masses
per hectare was 43.8% in 1999, before increasing to78.8 and 75.8% in 2000 and 2001, respectively. Gypsymoth egg mass densities at the Þeld plots generallydecreased (46.9% of plots) or remained relatively con-stant (34.4% of plots) from 1999 to 2001. Egg massdensities between 1999 and 2001 increased at onlythree plots during this study and were variable at threeother plots, decreasing from 1999 to 2000 before in-creasing again in 2001 (Table 1).
Mean � SE density of E. maimaiga resting spores insoil samples collected from our plots was 154 � 12 and232 � 30 spores per gram dry soil in 1999 and 2000,respectively. Differences in E. maimaiga resting sporedensity among aspects were not signiÞcant (P� 0.05)in soil samples collected from our Þeld plots in eitheryear (Table 2). Spore density in 1999 and 2000 wassigniÞcantly correlated (y� 0.531x� 128.1; r2 � 0.18;P� 0.05). Density of resting spores was highly variableamong plots within each year, ranging from 21 to 623spores per g dry soil in 1999 and 33Ð932 spores per gdry soil in 2000.
Soil-borne E. maimaiga resting spore densities in1999 and 2000 were examined in relation to site char-acteristics, including percentage canopy cover, totalbasal area, soil pH, and gypsy moth egg mass density(Table 1). Only soil pH was signiÞcantly correlated(P � 0.05) with resting spore density in both years.Overall, plots with higher soil pH (i.e., less acidic)
Table 1. Location and characteristics of oak stands in Michigan used for E. maimaiga field sites, 1999–2001
Plot no. CountyCanopy
cover (%)Total basal
area (m2/ha)Soil pH
Gypsy moth egg mass density(no./ha)
1999 2000 2001
1 Chippewa 96.4 30 4.6 0 0 02 Grand Traverse 91.2 28 5.1 0 0 1003 Manistee 96.1 25 5.0 1,050 50 04 Manistee 84.7 14 5.1 600 250 505 Mason 90.9 18 5.0 3,600 150 5506 Mason 95.8 25 5.2 1,350 0 07 Mason 94.5 25 4.8 100 50 08 Oceana 95.6 25 4.3 0 0 09 Oceana 93.0 25 4.7 50 0 010 Oceana 90.9 30 4.5 300 50 011 Lake 86.7 16 4.2 7,900 400 1,55012 Lake 90.6 28 4.4 700 850 2,40013 Lake 94.5 28 4.7 450 600 40014 Lake 91.9 25 4.4 0 100 015 Lake 94.0 23 4.5 100 0 016 Newaygo 90.4 25 4.5 50 0 017 Newaygo 88.6 25 4.7 200 0 5018 Newaygo 93.0 25 4.2 450 0 019 Newaygo 87.8 25 4.3 50 0 020 Newaygo 90.6 21 4.2 400 0 021 Newaygo 95.6 28 4.1 200 0 5022 Newaygo 93.5 21 4.7 250 0 10023 Newaygo 91.2 25 4.4 0 0 024 Newaygo 90.4 18 4.8 1,150 0 025 Crawford 82.6 18 4.1 0 100 35026 Kalkaska 93.5 25 5.4 0 50 15027 Kalkaska 84.1 28 5.1 1,300 250 20028 Roscommon 89.6 23 4.3 200 0 029 Roscommon 89.6 23 4.3 1,100 0 030 Clare 87.3 21 4.3 0 0 031 Arenac 80.0 23 4.1 14,150 200 50032 Kalamazoo 90.5 25 4.8 0 0 033 Clare 89.3 21 5.2 n/a 50 50
October 2012 SIEGERT ET AL.: FIELD VS. LABORATORY INFECTION BY E. maimaiga 1119
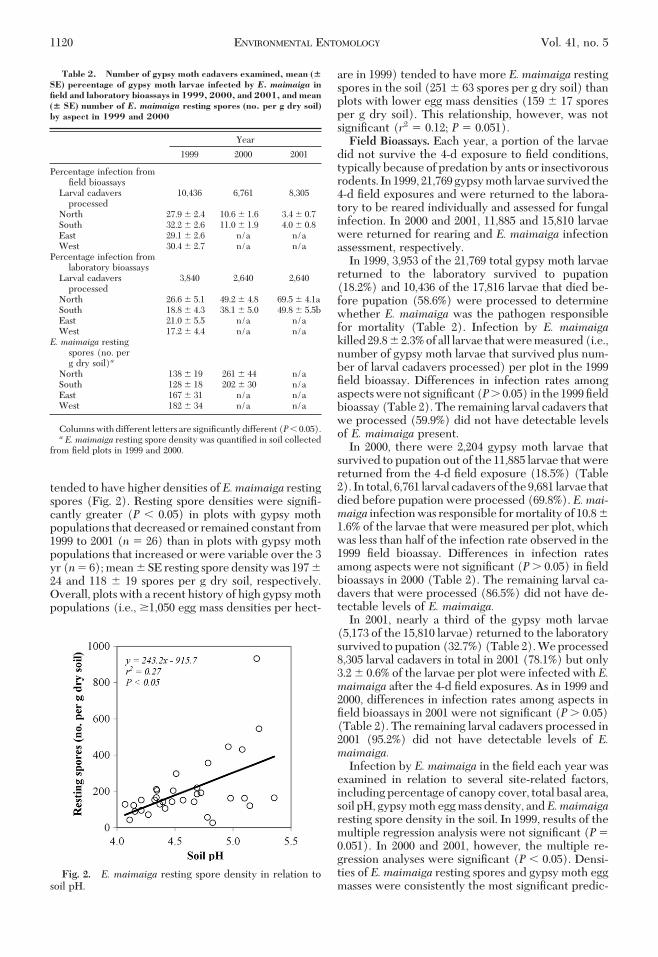
tended to have higher densities ofE.maimaiga restingspores (Fig. 2). Resting spore densities were signiÞ-cantly greater (P � 0.05) in plots with gypsy mothpopulations that decreased or remained constant from1999 to 2001 (n � 26) than in plots with gypsy mothpopulations that increased or were variable over the 3yr (n� 6); mean � SE resting spore density was 197 �24 and 118 � 19 spores per g dry soil, respectively.Overall, plots with a recent history of high gypsy mothpopulations (i.e., �1,050 egg mass densities per hect-
are in 1999) tended to have more E. maimaiga restingspores in the soil (251 � 63 spores per g dry soil) thanplots with lower egg mass densities (159 � 17 sporesper g dry soil). This relationship, however, was notsigniÞcant (r2 � 0.12; P � 0.051).Field Bioassays. Each year, a portion of the larvae
did not survive the 4-d exposure to Þeld conditions,typically because of predation by ants or insectivorousrodents. In 1999, 21,769 gypsy moth larvae survived the4-d Þeld exposures and were returned to the labora-tory to be reared individually and assessed for fungalinfection. In 2000 and 2001, 11,885 and 15,810 larvaewere returned for rearing and E. maimaiga infectionassessment, respectively.
In 1999, 3,953 of the 21,769 total gypsy moth larvaereturned to the laboratory survived to pupation(18.2%) and 10,436 of the 17,816 larvae that died be-fore pupation (58.6%) were processed to determinewhether E. maimaiga was the pathogen responsiblefor mortality (Table 2). Infection by E. maimaigakilled29.8�2.3%ofall larvae thatweremeasured(i.e.,number of gypsy moth larvae that survived plus num-ber of larval cadavers processed) per plot in the 1999Þeld bioassay. Differences in infection rates amongaspects were not signiÞcant (P� 0.05) in the 1999 Þeldbioassay (Table 2). The remaining larval cadavers thatwe processed (59.9%) did not have detectable levelsof E. maimaiga present.
In 2000, there were 2,204 gypsy moth larvae thatsurvived to pupation out of the 11,885 larvae that werereturned from the 4-d Þeld exposure (18.5%) (Table2). In total, 6,761 larval cadaversof the9,681 larvae thatdied before pupation were processed (69.8%). E. mai-maiga infection was responsible for mortality of 10.8 �1.6% of the larvae that were measured per plot, whichwas less than half of the infection rate observed in the1999 Þeld bioassay. Differences in infection ratesamong aspects were not signiÞcant (P� 0.05) in Þeldbioassays in 2000 (Table 2). The remaining larval ca-davers that were processed (86.5%) did not have de-tectable levels of E. maimaiga.
In 2001, nearly a third of the gypsy moth larvae(5,173 of the 15,810 larvae) returned to the laboratorysurvived to pupation (32.7%) (Table 2). We processed8,305 larval cadavers in total in 2001 (78.1%) but only3.2 � 0.6% of the larvae per plot were infected with E.maimaiga after the 4-d Þeld exposures. As in 1999 and2000, differences in infection rates among aspects inÞeld bioassays in 2001 were not signiÞcant (P� 0.05)(Table 2). The remaining larval cadavers processed in2001 (95.2%) did not have detectable levels of E.maimaiga.
Infection by E. maimaiga in the Þeld each year wasexamined in relation to several site-related factors,including percentage of canopy cover, total basal area,soil pH, gypsy moth egg mass density, andE.maimaigaresting spore density in the soil. In 1999, results of themultiple regression analysis were not signiÞcant (P�0.051). In 2000 and 2001, however, the multiple re-gression analyses were signiÞcant (P � 0.05). Densi-ties of E. maimaiga resting spores and gypsy moth eggmasses were consistently the most signiÞcant predic-
Table 2. Number of gypsy moth cadavers examined, mean (�SE) percentage of gypsy moth larvae infected by E. maimaiga infield and laboratory bioassays in 1999, 2000, and 2001, and mean(� SE) number of E. maimaiga resting spores (no. per g dry soil)by aspect in 1999 and 2000
Year
1999 2000 2001
Percentage infection fromÞeld bioassays
Larval cadaversprocessed
10,436 6,761 8,305
North 27.9 � 2.4 10.6 � 1.6 3.4 � 0.7South 32.2 � 2.6 11.0 � 1.9 4.0 � 0.8East 29.1 � 2.6 n/a n/aWest 30.4 � 2.7 n/a n/a
Percentage infection fromlaboratory bioassays
Larval cadaversprocessed
3,840 2,640 2,640
North 26.6 � 5.1 49.2 � 4.8 69.5 � 4.1aSouth 18.8 � 4.3 38.1 � 5.0 49.8 � 5.5bEast 21.0 � 5.5 n/a n/aWest 17.2 � 4.4 n/a n/aE. maimaiga resting
spores (no. perg dry soil)a
North 138 � 19 261 � 44 n/aSouth 128 � 18 202 � 30 n/aEast 167 � 31 n/a n/aWest 182 � 34 n/a n/a
Columns with different letters are signiÞcantly different (P� 0.05).a E. maimaiga resting spore density was quantiÞed in soil collected
from Þeld plots in 1999 and 2000.
Fig. 2. E. maimaiga resting spore density in relation tosoil pH.
1120 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 5
tors of E. maimaiga Þeld infection in 2000 (Y �0.02[resting sporedensity]0.014[eggmassdensity]�7.5), (F2, 30 � 5.318, r2 � 0.26, P � 0.05) and in 2001(Y � 0.007[resting spore density] � 0.003[egg massdensity] � 1.06), (F2, 30 � 6.298, r2 � 0.30, P � 0.05).Results of ANOVA to assess whether site-related fac-tors varied with categories of Þeld infection rates andwhetherE.maimaiga Þeld infection varied with gypsymoth population trends were not signiÞcant (P �0.05).Laboratory Bioassays. In 1999, average E. maimaiga
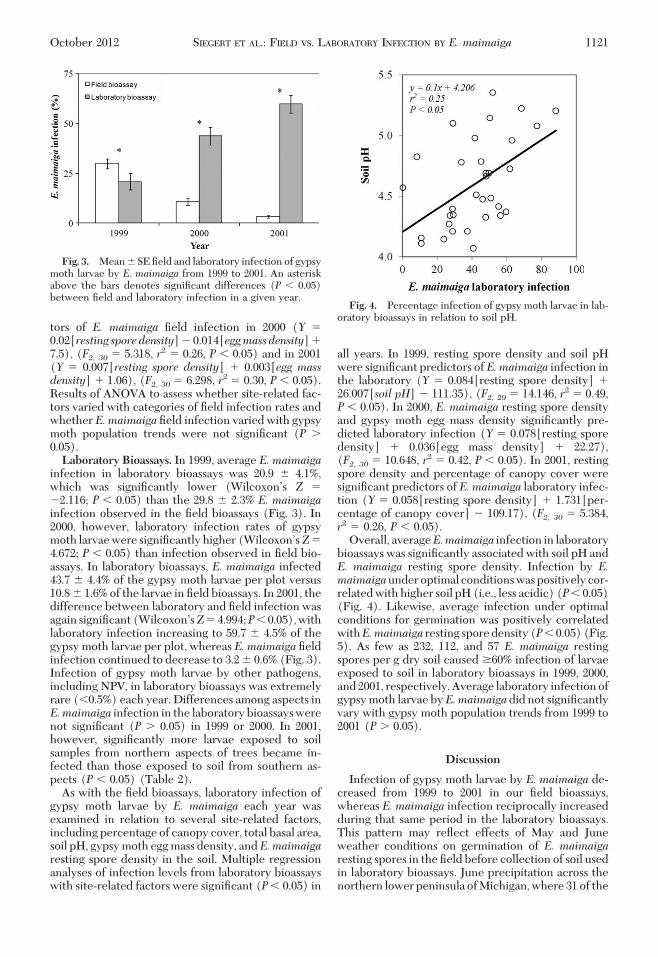
infection in laboratory bioassays was 20.9 � 4.1%,which was signiÞcantly lower (WilcoxonÕs Z �2.116; P � 0.05) than the 29.8 � 2.3% E. maimaigainfection observed in the Þeld bioassays (Fig. 3). In2000, however, laboratory infection rates of gypsymoth larvae were signiÞcantly higher (WilcoxonÕsZ�4.672; P � 0.05) than infection observed in Þeld bio-assays. In laboratory bioassays, E. maimaiga infected43.7 � 4.4% of the gypsy moth larvae per plot versus10.8 � 1.6% of the larvae in Þeld bioassays. In 2001, thedifference between laboratory and Þeld infection wasagain signiÞcant(WilcoxonÕsZ�4.994;P�0.05),withlaboratory infection increasing to 59.7 � 4.5% of thegypsy moth larvae per plot, whereas E. maimaiga Þeldinfection continued to decrease to 3.2 � 0.6% (Fig. 3).Infection of gypsy moth larvae by other pathogens,including NPV, in laboratory bioassays was extremelyrare (�0.5%) each year. Differences among aspects inE.maimaiga infection in the laboratory bioassays werenot signiÞcant (P � 0.05) in 1999 or 2000. In 2001,however, signiÞcantly more larvae exposed to soilsamples from northern aspects of trees became in-fected than those exposed to soil from southern as-pects (P � 0.05) (Table 2).
As with the Þeld bioassays, laboratory infection ofgypsy moth larvae by E. maimaiga each year wasexamined in relation to several site-related factors,including percentage of canopy cover, total basal area,soil pH, gypsy moth egg mass density, andE.maimaigaresting spore density in the soil. Multiple regressionanalyses of infection levels from laboratory bioassayswith site-related factors were signiÞcant (P� 0.05) in
all years. In 1999, resting spore density and soil pHwere signiÞcant predictors ofE.maimaiga infection inthe laboratory (Y � 0.084[resting spore density] �26.007[soil pH] 111.35), (F2, 29 � 14.146, r2 � 0.49,P � 0.05). In 2000, E. maimaiga resting spore densityand gypsy moth egg mass density signiÞcantly pre-dicted laboratory infection (Y � 0.078[resting sporedensity] � 0.036[egg mass density] � 22.27),(F2, 30 � 10.648, r2 � 0.42, P � 0.05). In 2001, restingspore density and percentage of canopy cover weresigniÞcant predictors of E. maimaiga laboratory infec-tion (Y � 0.058[resting spore density] � 1.731[per-centage of canopy cover] 109.17), (F2, 30 � 5.384,r2 � 0.26, P � 0.05).
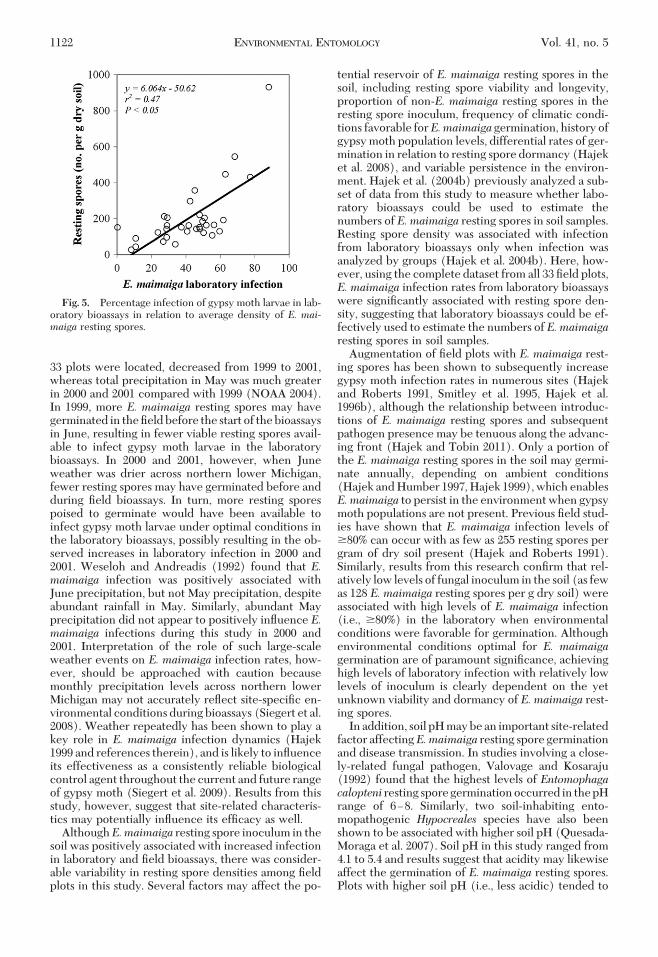
Overall, averageE.maimaiga infection in laboratorybioassays was signiÞcantly associated with soil pH andE. maimaiga resting spore density. Infection by E.maimaigaunderoptimal conditionswaspositivelycor-related with higher soil pH (i.e., less acidic) (P� 0.05)(Fig. 4). Likewise, average infection under optimalconditions for germination was positively correlatedwithE.maimaiga resting sporedensity(P�0.05)(Fig.5). As few as 232, 112, and 57 E. maimaiga restingspores per g dry soil caused �60% infection of larvaeexposed to soil in laboratory bioassays in 1999, 2000,and2001, respectively.Average laboratory infectionofgypsy moth larvae byE.maimaigadid not signiÞcantlyvary with gypsy moth population trends from 1999 to2001 (P � 0.05).
Discussion
Infection of gypsy moth larvae by E. maimaiga de-creased from 1999 to 2001 in our Þeld bioassays,whereas E. maimaiga infection reciprocally increasedduring that same period in the laboratory bioassays.This pattern may reßect effects of May and Juneweather conditions on germination of E. maimaigaresting spores in the Þeld before collection of soil usedin laboratory bioassays. June precipitation across thenorthern lower peninsula of Michigan, where 31 of the
Fig. 3. Mean � SE Þeld and laboratory infection of gypsymoth larvae by E. maimaiga from 1999 to 2001. An asteriskabove the bars denotes signiÞcant differences (P � 0.05)between Þeld and laboratory infection in a given year.
Fig. 4. Percentage infection of gypsy moth larvae in lab-oratory bioassays in relation to soil pH.
October 2012 SIEGERT ET AL.: FIELD VS. LABORATORY INFECTION BY E. maimaiga 1121
33 plots were located, decreased from 1999 to 2001,whereas total precipitation in May was much greaterin 2000 and 2001 compared with 1999 (NOAA 2004).In 1999, more E. maimaiga resting spores may havegerminated in the Þeld before the start of the bioassaysin June, resulting in fewer viable resting spores avail-able to infect gypsy moth larvae in the laboratorybioassays. In 2000 and 2001, however, when Juneweather was drier across northern lower Michigan,fewer resting spores may have germinated before andduring Þeld bioassays. In turn, more resting sporespoised to germinate would have been available toinfect gypsy moth larvae under optimal conditions inthe laboratory bioassays, possibly resulting in the ob-served increases in laboratory infection in 2000 and2001. Weseloh and Andreadis (1992) found that E.maimaiga infection was positively associated withJune precipitation, but not May precipitation, despiteabundant rainfall in May. Similarly, abundant Mayprecipitation did not appear to positively inßuence E.maimaiga infections during this study in 2000 and2001. Interpretation of the role of such large-scaleweather events on E. maimaiga infection rates, how-ever, should be approached with caution becausemonthly precipitation levels across northern lowerMichigan may not accurately reßect site-speciÞc en-vironmental conditions during bioassays (Siegert et al.2008). Weather repeatedly has been shown to play akey role in E. maimaiga infection dynamics (Hajek1999 and references therein), and is likely to inßuenceits effectiveness as a consistently reliable biologicalcontrol agent throughout the current and future rangeof gypsy moth (Siegert et al. 2009). Results from thisstudy, however, suggest that site-related characteris-tics may potentially inßuence its efÞcacy as well.
AlthoughE.maimaiga resting spore inoculum in thesoil was positively associated with increased infectionin laboratory and Þeld bioassays, there was consider-able variability in resting spore densities among Þeldplots in this study. Several factors may affect the po-
tential reservoir of E. maimaiga resting spores in thesoil, including resting spore viability and longevity,proportion of non-E. maimaiga resting spores in theresting spore inoculum, frequency of climatic condi-tions favorable forE.maimaiga germination, history ofgypsy moth population levels, differential rates of ger-mination in relation to resting spore dormancy (Hajeket al. 2008), and variable persistence in the environ-ment. Hajek et al. (2004b) previously analyzed a sub-set of data from this study to measure whether labo-ratory bioassays could be used to estimate thenumbers of E. maimaiga resting spores in soil samples.Resting spore density was associated with infectionfrom laboratory bioassays only when infection wasanalyzed by groups (Hajek et al. 2004b). Here, how-ever, using the complete dataset from all 33 Þeld plots,E. maimaiga infection rates from laboratory bioassayswere signiÞcantly associated with resting spore den-sity, suggesting that laboratory bioassays could be ef-fectively used to estimate the numbers of E. maimaigaresting spores in soil samples.
Augmentation of Þeld plots with E. maimaiga rest-ing spores has been shown to subsequently increasegypsy moth infection rates in numerous sites (Hajekand Roberts 1991, Smitley et al. 1995, Hajek et al.1996b), although the relationship between introduc-tions of E. maimaiga resting spores and subsequentpathogen presence may be tenuous along the advanc-ing front (Hajek and Tobin 2011). Only a portion ofthe E. maimaiga resting spores in the soil may germi-nate annually, depending on ambient conditions(Hajek and Humber 1997, Hajek 1999), which enablesE.maimaiga to persist in the environment when gypsymoth populations are not present. Previous Þeld stud-ies have shown that E. maimaiga infection levels of�80% can occur with as few as 255 resting spores pergram of dry soil present (Hajek and Roberts 1991).Similarly, results from this research conÞrm that rel-atively low levels of fungal inoculum in the soil (as fewas 128 E. maimaiga resting spores per g dry soil) wereassociated with high levels of E. maimaiga infection(i.e., �80%) in the laboratory when environmentalconditions were favorable for germination. Althoughenvironmental conditions optimal for E. maimaigagermination are of paramount signiÞcance, achievinghigh levels of laboratory infection with relatively lowlevels of inoculum is clearly dependent on the yetunknown viability and dormancy of E. maimaiga rest-ing spores.
In addition, soil pH may be an important site-relatedfactor affectingE.maimaiga resting spore germinationand disease transmission. In studies involving a close-ly-related fungal pathogen, Valovage and Kosaraju(1992) found that the highest levels of Entomophagacalopteni resting sporegerminationoccurred in thepHrange of 6Ð8. Similarly, two soil-inhabiting ento-mopathogenic Hypocreales species have also beenshown to be associated with higher soil pH (Quesada-Moraga et al. 2007). Soil pH in this study ranged from4.1 to 5.4 and results suggest that acidity may likewiseaffect the germination of E. maimaiga resting spores.Plots with higher soil pH (i.e., less acidic) tended to
Fig. 5. Percentage infection of gypsy moth larvae in lab-oratory bioassays in relation to average density of E. mai-maiga resting spores.
1122 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 5

have higher densities of resting spores and, corre-spondingly, greater E. maimaiga infection under op-timal conditions for fungal germination. It is unclear,however, whether soil pH directly affected germina-tion of E. maimaiga resting spores or if results weremerely an artifact of higher resting spore density.More research is needed to conclusively measure theeffect of soil pH on E. maimaiga resting spore germi-nation and persistence.
Results of this 3-yr study indicate that efÞcacy of E.maimaiga as a biological control agent is strongly de-pendent on Þeld conditions favorable for germinationof resting spores and varies in relation to several site-related variables. Increased infection of gypsy mothlarvae in laboratory bioassays in years when Þeld in-fections were low suggests that environmental condi-tions were not optimal for resting spore germination.Whether this affects resting spore longevity in theÞeld remains to be tested. Future research includingexperimental manipulation is needed to further ex-plore the role and interaction of site-related factorssuch as previous gypsy moth density and resting sporedensity. Results will increase our ability to effectivelyuse the gypsy moth fungal pathogen E. maimaigawithin established gypsy moth populations and alongthe advancing front of gypsy moth spread as well as topredict its effectiveness in areas that will becomeinvaded in the future.
Acknowledgments
We gratefully acknowledge the assistance of DeborahGrooms, William Kauffman, and associates at the USDAÐAPHISÐPPQ Biological Control Laboratory, formerly ofNiles, MI, all of whom were of inestimable value during thisproject.Wealso thankMattDavenport,MelanieDePoy,TomEllis, Rachael Harris, Anne Henderson, and Abigail Sommers,all of Michigan State University (MSU), for their assistancewith numerous tasks. In addition, we thank Lyle Buss, of theUniversity of Florida, and Bob Heyd, Roger Mech, and FrankSapio, all of the Michigan Department of Natural Resources,for their assistance in locating the Þeld sites withE.maimaigain Michigan. We acknowledge Therese M. Poland, David E.Rothstein, and Suzanne M. Thiem, all of MSU, for theirhelpful comments during preparation of this manuscript.Funding for this research was provided by the Special Tech-nology Development Program, State and Private Forestry,USDA Forest Service, and the Michigan Department of Ag-riculture. This study was supported in part by MichiganAgricultural Experiment Station Project No. MICL01700.
References Cited
Andreadis, T. G., and R. M. Weseloh. 1990. Discovery ofEntomophaga maimaiga in North American gypsy moth,Lymantria dispar. Proc. Natl. Acad. Sci. U.S.A. 87: 2461Ð2465.
Buss, L. J. 1997. Evaluation of three egg mass survey meth-ods and two biological control agents for gypsy mothmanagement. M.S. thesis, Michigan State University, EastLansing.
Hajek, A. E. 1997a. Entomophaga maimaiga reproductiveoutput is determined by spore type initiating an infection.Mycol. Res. 101:971Ð974.
Hajek, A. E. 1997b. Fungal and viral epizootics in gypsymoth(Lepidoptera:Lymantriidae)populations incentralNew York. Biol. Control 10: 58Ð68.
Hajek, A. E. 1999. Pathology and epizootiology of Ento-mophaga maimaiga infections in forest Lepidoptera. Mi-crobiol. Mol. Biol. Rev. 63: 814Ð835.
Hajek, A. E. 2001. Larval behavior in Lymantria dispar in-creases risk of fungal infection. Oecologia 126: 285Ð291.
Hajek, A. E., and R. A. Humber. 1997. Formation and ger-mination of Entomophaga maimaiga azygospores. Can. J.Bot. 75: 1739Ð1747.
Hajek, A. E., and D. W. Roberts. 1991. Pathogen reservoirsas a biological control resource: introduction of Ento-mophaga maimaiga to North American gypsy moth, Ly-mantria dispar, populations. Biol. Control 1: 29Ð34.
Hajek, A. E., and R. S. Soper. 1991. Within-tree location ofgypsy moth, Lymantria dispar, larvae killed by Ento-mophaga maimaiga (Zygomycetes: Entomophthorales).J. Invertebr. Pathol. 58: 468Ð469.
Hajek, A. E., and P. C. Tobin. 2011. Introduced pathogensfollowthe invasion frontof a spreadingalienhost. J.Anim.Ecol. 80: 1217Ð1226.
Hajek, A. E., andM.M.Wheeler. 1994. Application of tech-niques for quantiÞcation of soil-borne Entomophtho-ralean resting spores. J. Invertebr. Pathol. 64: 71Ð73.
Hajek, A. E., R. A. Humber, J. S. Elkinton, R. May, S.R.A.Walsh, and J. C. Silver. 1990. Allozyme and restrictionfragment length polymorphism analyses conÞrm Ento-mophaga maimaiga responsible for 1989 epizootics inNorth American gypsy moth populations. Proc. Natl.Acad. Sci. U.S.A. 87: 6979Ð6982.
Hajek, A. E., L. Butler, S.R.A. Walsh, J. C. Silver, F. P. Hain,F. L. Hastings, T. M. O’Dell, and D. R. Smitley. 1996a.Host range of the gypsy moth (Lepidoptera: Lymantrii-dae) pathogen Entomophaga maimaiga (Zygomycetes:Entomophthorales) in the Þeld versus laboratory. Envi-ron. Entomol. 25: 709Ð721.
Hajek, A. E., J. S. Elkinton, and J. J. Witcosky. 1996b. In-troduction and spread of the fungal pathogen Ento-mophaga maimaiga (Zygomycetes: Entomophthorales)along the leading edge of gypsy moth (Lepidoptera:Lymantriidae) spread. Environ. Entomol. 25: 1235Ð1247.
Hajek, A. E., L. Bauer, M. L. McManus, and M. M.Wheeler.1998a. Distribution of resting spores of the Lymantriadispar pathogen Entomophaga maimaiga in soil and onbark. BioControl 43: 189Ð200.
Hajek, A. E., K. M. Tatman, P. H. Wanner, and M. M.Wheeler. 1998b. Location and persistence of cadaversof gypsy moth, Lymantria dispar, containing Ento-mophaga maimaiga azygospores. Mycologia 90: 754Ð760.
Hajek, A. E., L. Butler, J. K. Liebherr, and M. M. Wheeler.2000a. Risk of infection by the fungal pathogen Ento-mophaga maimaiga among Lepidoptera on the forestßoor. Environ. Entomol. 29: 645Ð650.
Hajek, A. E., M. Shimazu, and B. Knoblauch. 2000b. Isolat-ing a species of Entomophthorales using resting spore-bearing soil. J. Invertebr. Pathol. 75: 298Ð300.
Hajek,A.E., J. S. Strazanac,M.M.Wheeler,F.M.Vermeylen,andL. Butler. 2004a. Persistence of the fungal pathogenEntomophaga maimaiga and its impact on native Lyman-triidae. Biol. Control 30: 466Ð473.
Hajek, A. E., M. M. Wheeler, N. W. Siegert, and D. G.McCullough. 2004b. Using bioassays to estimate abun-dance of Entomophaga maimaiga resting spores in soil.J. Invertebr. Pathol. 86: 61Ð64.
Hajek, A. E., A. E. Burke, C. Nielsen, J. J. Hannam, and L. S.Bauer. 2008. Nondormancy in Entomophaga maimaiga
October 2012 SIEGERT ET AL.: FIELD VS. LABORATORY INFECTION BY E. maimaiga 1123

azygospores: effects of isolate and cold exposure. Myco-logia 100: 833Ð842.
Kolodny-Hirsch, D. M. 1986. Evaluation of methods forsampling gypsy moth (Lepidoptera: Lymantriidae) eggmass populations and development of sequential sam-pling plans. Environ. Entomol. 15: 122Ð127.
Malakar, R., J. S. Elkinton, S. D. Carroll, and V. D’Amico.1999a. Interactions between two gypsy moth (Lepidop-tera: Lymantriidae) pathogens: nucleopolyhedrovirusand Entomophaga maimaiga (Zygomycetes: Entomoph-thorales): Þeld studies and a simulation model. Biol. Con-trol 16: 189Ð198.
Malakar, R., J. S. Elkinton, A. E. Hajek, and J. P. Burand.1999b. Within-host interactions of Lymantria dispar(Lepidoptera: Lymantriidae) nucleopolyhedrosis virusand Entomophaga maimaiga (Zygomycetes: Entomoph-thorales). J. Invertebr. Pathol. 73: 91Ð100.
[NOAA] National Oceanic and Atmospheric Administra-tion. 2004. Time bias corrected divisional precipitationdata. (ftp://ftp.ncdc.noaa.gov/pub/data/cirs/drd964x.pcp.txt).
O’Dell, T. M., C. A. Butt, and A. W. Bridgeforth. 1985.Lymantria dispar, pp. 355Ð367. In P. Singh and R. Moore(eds.), Handbook of insect rearing, vol. 2. Elsevier, NewYork.
Papierok, B., and A. E. Hajek. 1997. Fungi: Entomoptho-rales, pp. 187Ð212. In L. Lacey (ed.), Manual of tech-niques in insect pathology. Academic, New York.
Quesada-Moraga, E., J. A. Navas-Cortes, E.A.A. Maranhao, A.Ortiz-Urquiza, and C. Santiago-Alverez. 2007. Factorsaffecting the occurrence and distribution of ento-mopathogenic fungi innatural andcultivated soils.Mycol.Res. 111: 947Ð966.
Shimazu,M. 1987. Effect of rearing humidity of host insectson the spore types of Entomophaga maimaiga Humber,Shimazu et Soper (Entomophthorales: Entomophtho-raceae). Appl. Entomol. Zool. 22: 394Ð397.
Shimazu, M., and R. S. Soper. 1986. Pathogenicity and spo-rulation of Entomophaga maimaiga Humber, Shimazu,Soper and Hajek (Zygomycetes: Entomophthorales) onlarvae of the gypsy moth, Lymantria dispar L. (Lepidop-tera: Lymantriidae). Appl. Entomol. Zool. 21: 589Ð596.
Siegert, N. W. 2004. Meteorological factors affecting thesuccess of the gypsy moth fungal pathogen Entomophagamaimaiga (Zygomycetes: Entomophthorales) in Michi-gan. Ph.D. dissertation, Michigan State University, EastLansing.
Siegert, N. W., D. G. McCullough, A. E. Hajek, and J. A.Andresen. 2008. Effect of microclimatic conditions onprimary transmission of the gypsy moth fungal pathogenEntomophaga maimaiga in Michigan. Gt. Lakes Entomol.41: 111Ð128.
Siegert, N.W.,D.G.McCullough, R. C. Venette, A. E.Hajek,and J. A. Andresen. 2009. Assessing the climatic poten-tial for epizootics of the gypsy moth fungal pathogenEntomophaga maimaiga in the north central UnitedStates. Can. J. For. Res. 39: 1958Ð1970.
Smitley, D. R., L. S. Bauer, A. E. Hajek, F. J. Sapio, and R. A.Humber. 1995. Introduction and establishment of Ento-mophaga maimaiga, a fungal pathogen of gypsy moth(Lepidoptera: Lymantriidae) in Michigan. Environ. En-tomol. 24: 1685Ð1695.
Sokal, R. R., and F. J. Rohlf. 1995. Biometry, 3rd ed. W.H.Freeman & Company, New York.
SYSTAT. 2000. SYSTAT, version 10. SPSS Inc., Chicago, IL.Valovage,W.D., and R. S. Kosaraju. 1992. Effects of pH and
buffer systems on resting spore germination of the grass-hopper (Orthoptera: Acrididae) pathogen, Entomophagacalopteni (�Entomophaga grylli, pathotype 2) (Ento-mophthorales: Entomophthoraceae). Environ. Entomol.21: 1202Ð1211.
Webb, R. E., G. B. White, K. W. Thorpe, and S. E. Talley.1999. Quantitative analysis of a pathogen-induced pre-mature collapse of a “leading edge” gypsy moth (Lepi-doptera: Lymantriidae) population in Virginia. J. Ento-mol. Sci. 34: 84Ð100.
Weseloh, R. M., and T. G. Andreadis. 1992. Mechanisms oftransmission of the gypsy moth (Lepidoptera: Lymant-riidae) fungus, Entomophaga maimaiga (Zygomycetes:Entomophthoraceae) and effects of site conditions on itsprevalence. Environ. Entomol. 21: 901Ð906.
Weseloh, R. M., and T. G. Andreadis. 1997. Persistence ofresting spores of Entomophaga maimaiga, a fungal patho-gen of the gypsy moth, Lymantria dispar. J. Invertebr.Pathol. 69: 195Ð196.
Weseloh, R. M., and T. G. Andreadis. 2002. Detecting thetiter in forest soils of spores of the gypsy moth (Lepi-doptera: Lymantriidae) fungal pathogen, Entomophagamaimaiga (Zygomycetes: Entomophthorales). Can. En-tomol. 134: 269Ð279.
Received 7 May 2012; accepted 11 July 2012.
1124 ENVIRONMENTAL ENTOMOLOGY Vol. 41, no. 5
Related Documents











