This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Estrogen and androgen receptors:regulators of fuel homeostasis andemerging targets for diabetesand obesityFranck Mauvais-Jarvis
Department of Medicine, Division of Endocrinology, Metabolism and Molecular Medicine, and Comprehensive Center
on Obesity, Feinberg School of Medicine, Northwestern University, Chicago, IL 60611, USA
Because of increasing life expectancy, the contributionof age-related estrogen or androgen deficiency to obesi-ty and type 2 diabetes will become a new therapeuticchallenge. This review integrates current concepts onthe mechanisms through which estrogen receptors(ERs) and androgen receptor (AR) regulate energy ho-meostasis in rodents and humans. In females, estrogenmaintains energy homeostasis via ERa and ERb, bysuppressing energy intake and lipogenesis, enhancingenergy expenditure, and ameliorating insulin secretionand sensitivity. In males, testosterone is converted toestrogen and maintains fuel homeostasis via ERs andAR, which share related functions to suppress adiposetissue accumulation and improve insulin sensitivity. Wesuggest that ERs and AR could be potential targets in theprevention of age-related metabolic disorders.
Contribution of sex hormones to metabolic diseasesIncreased food supply and decreased physical activity haveresulted in a worldwide epidemic of obesity. As a conse-quence of these environmental changes, the incidence oftype 2 diabetes (T2D) is on the rise [1]. In addition, adisorder involving increased visceral adipose tissue, hy-perlipidemia, insulin resistance, and hypertension termedmetabolic syndrome, has emerged [2].
There is a concerted interaction between sex/reproduc-tion and energy metabolism [3]. First, extreme conditionsof disrupted energy balance such as obesity at one end ofthe spectrum, or anorexia leading to cachexia at the other,both negatively affect fertility. Second, there are funda-mental aspects of energy metabolism that are regulateddifferently in males and females [4]. To cite one crucialexample, female mammals, who bear the burden of gesta-tion and lactation, have been favorably altered duringevolution to resist loss of body energy stores during pro-longed periods of food scarcity, and therefore they depositadipose tissue in the lower subcutaneous area, with lowerlipolytic activity. Conversely, males deposit adipose tissuein visceral areas, with greater lipolytic activity, to be ableto mobilize energy stores promptly for muscle activity.
It is believed that the circulating gonadal hormones,specifically androgen and estrogen, control these sex dif-
ferences in energy balance between the onset of pubertyand menopause. Because of the dramatic increase in lifeexpectancy, women will spend the second half of their life,after menopause, in estrogen deficiency, which predisposesto metabolic syndrome and T2D [5]. Men will also spend asignificant part of their life in age-related androgen defi-ciency. Although no clear relationship exists between thegradual loss of testosterone production and T2D, androgendeficiency clearly predisposes men to the metabolic syn-drome [6]. Therefore, the contribution of sex hormonedeficiency to metabolic diseases will become a new thera-peutic challenge in the 21st century. Understanding howestrogen and androgen contribute to fuel homeostasis viatheir receptors promises crucial therapeutic applications.
In this review, we integrate current concepts on the roleof estrogen and androgen and their receptors in regulatingenergy homeostasis in male and female rodents andhumans. We also discuss how the estrogen receptors(ERs) and androgen receptor (AR) are important targetsfor age-related metabolic disorders.
The Estrogen receptorsMechanism of ERs action
In healthy premenopausal women, 17b-estradiol (E2) isproduced by the ovaries by the aromatization of andro-stenedione to estrone, followed by conversion to E2. Inthese women, E2 functions as a circulating hormone thatacts on distant target tissues. However, in women aftermenopause (when the ovaries fail to produce E2) and inmen, E2 is produced in extragonadal sites, mainly adiposetissue, bone, vessels and brain by local tissue aromatiza-tion from circulating testosterone [7]. Therefore, in malesand females, testosterone should be considered a circulat-ing prohormone that is locally converted to either E2 actingon ERs, or to 5a-dihydrotestosterone (DHT), the mainligand of the AR. Although DHT cannot be aromatizedto estrogen, the situation is complicated by the fact thatDHT can still be converted to a ‘second estrogen’, 5a-androstane-3b,17b-diol, which acts on ERs [8].
The ER exists in two main forms, ERa and ERb, whichhave multiple isoforms and exhibit distinct tissue expres-sion patterns and functions [9]. In the classic ER signalingpathway, E2-activated ERs binds as a homodimer to the
Review
Corresponding author: Mauvais-Jarvis, F. ([email protected]).
24 1043-2760/$ – see front matter � 2010 Elsevier Ltd. All rights reserved. doi:10.1016/j.tem.2010.10.002 Trends in Endocrinology and Metabolism, January 2011, Vol. 22, No. 1

Author's personal copy
estrogen response element (ERE) in target promoters orindirectly to an activator protein (AP)-1 or Sp-1 responseelement through association with other transcription fac-tors, such as Fos/Jun, which tether the activated ERs toDNA [9]. This classic, ‘genomic’ mechanism typicallyoccurs within hours, leading to upregulation or downregu-lation of gene transcription. E2 can also activate rapidsignals, acting within minutes or seconds via extranuclearand membrane-associated forms of ERs and the G protein-coupled (GP)ER, leading to activation of ion channels andprotein kinases [10]. Although reproductive functions aremostly mediated via classic nuclear ERs acting as ligand-activated transcription factors, a large component of ERsactions related to energy metabolism also involves extra-nuclear ERs, indirectly modulating gene expression oracting independently of nuclear events [11].
ERs control of energy intake and expenditure
The documented anti-obesity effects of E2 in vivo arecentrally mediated. Surprisingly, the major models of es-trogen deficiency and resistance do not exhibit hyperpha-gia. Thus, mice of both sexes lacking the aromataseenzyme, which cannot synthesize E2, develop obesitybut show no hyperphagia or reduced energy expenditure.Rather, they exhibit reduced spontaneous physical activityand a decrease in lean body mass [12]. Similarly, ERa
deficiency in male and female mice causes obesity withouthyperphagia but with decreased energy expenditure[13,14]. Clegg et al. made the observation that in maleand female rats, E2 enhances the ability of centrallyadministered leptin to suppress food intake [15]. This‘leptinomimetic’ function of E2 is best observed in leptindeficient (ob/ob) and leptin resistant (db/db) mice of bothsexes, in which E2 decreases food intake and increasesenergy expenditure, resulting in a reduction in bodyweight[16]. The anorectic function of exogenously administeredE2 is present in female wild type mice and is lost in femaleERa-deficient mice, demonstrating that ERa activation isanorexigenic [17]. Thus, loss of ERa action produces apredominant decrease in energy expenditure, whereasincreasing ERa signaling by raising serum E2 concentra-tions both suppresses energy intake and increases energyexpenditure. E2 also suppresses food intake through ERb,as the anorectic effect of intracerebroventricular injectionof E2 is blocked by co-administration of ERb antisenseoligodeoxynucleotides to female rats [18].
The precise anatomic site in the brain of ERs suppres-sion of body weight is still unknown. The arcuate nucleus(ARC) is a key hypothalamic area for mediating leptininhibition of food intake. It contains first order, leptinresponsive, anorexigenic pro-opiomelanocortin (POMC)neurons and orexigenic neuropeptide Y/agouti-related pep-tide (NPY/AgRP) neurons. Tamas Horvath and coworkersshowed that E2 triggers excitatory inputs to POMC neu-rons in the ARC of female rodents [16]. However, E2 doesnot seem to suppress food intake via action on ERs in ARCPOMC neurons. In fact, it was recently suggested that theanorexigenic function of E2 in female mice is mediated viaa decrease in hypothalamic orexigenic NPY and AgRP, butis independent of ER action on ARC NPY and AgRPneurons because these cells do not express ERa [19]. Most
importantly, E2, acting via ERa in nucleus tractus solitar-ius neurons of the brainstem, is sufficient to inhibit feedingin female rats, suggesting that the anorectic function of E2could originate in the hindbrain [20]. E2 stimulation ofenergy expenditure could involve both ERa and ERb, as
silencing of ERa in the hypothalamus of female rodentsreduces energy expenditure and produces obesity withouthyperphagia [21], whereas administration of an ERb-se-lective agonist to female mice fed a high fat diet (HFD)increases expression of thermogenic uncoupling protein-1in brown adipose tissue and reduces obesity [22]. Evidencepoints toward a site of action in the ventromedial nucleusof the hypothalamus (VMH) for ERa, as silencing of ERa inthe VMH reduces energy expenditure [21].
Unlike ERa and ERb, the role of GPER in body weightregulation still needs validation. In one study, female micelacking GPER developed obesity [23], but the obesityphenotype was observed in only one of the four GPERmutant mouse lines studied [24].
The signaling mechanisms of ERs actions in hypotha-lamic neurons are not fully elucidated, but available evi-dence suggests that they involve extranuclear ERs. First,E2 triggers a rapid increase in excitatory inputs to POMCneurons in the ARC in vivo, consistent with rapid, extra-nuclear or membrane initiated actions [16], and according-ly E2 can suppress NPY in clonal hypothalamic neuronsvia a membrane form of ERa [25]. Second, an E2-respon-sive, Gq-coupled membrane receptor (Gq-mER) is involvedin mediating the anorectic effects of E2 on food intake andbody temperature in hypoestrogenic female rodents[26,27].
ERs suppress lipogenesis in white adipose tissue and
liver
E2 also suppresses white adipose tissue (WAT) accumula-tion by decreasing fatty acid and triglyceride synthesis orlipogenesis. Greenberg and coworkers showed that in vivotreatment with E2 reduces adipocyte size in ovariectomizedfemale mice by reducing fatty acid uptake (downregulationof lipoprotein lipase) and lipogenesis (downregulation ofacetyl-coAcarboxylateand fattyacidsynthase), and increas-ing catecholamine-stimulated lipolysis [28]. Similarly, E2suppresses lipogenic genes and triglyceride accumulation inWAT and liver in HFD fed [29] and leptin resistant femalemice [30]. Interestingly, this effect is reproducedby selectiveagonists of ERb, but not ERa [22,31].
ERs are expressed in adipocytes and hepatocytes of bothsexes, and extensive evidence demonstrates that E2 hasdirect effects on cultured adipocytes, with the overall effectof inhibiting adipogenesis and lipogenesis [32]. Thus, theE2 effects described above could result from ER action inperipheral tissues. Nevertheless, the exact contribution ofthe anti-lipogenic effects of E2 in vivo resulting from directER action in WAT and liver, or from central ER actionaffecting adipose and liver via the autonomous system, isstill unknown. Although the overall effect of E2 is todecrease WAT accumulation, E2 favors subcutaneousWAT accumulation via central [15] and peripheral [32]mechanisms in both sexes.
ERb is anti-lipogenic and anti-adipogenic. ERb deficien-cy favorsWAT accumulation in femalemice during high fat
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
25

Author's personal copy
feeding by increasing peroxisome proliferator-activatedreceptor (PPAR)g signaling in WAT, thus demonstratingthat ERb acts directly on adipocytes in vivo and is anegative regulator of PPARg [33]. In addition, ERb selec-tive ligands show PPARg antagonistic actions in adipo-cytes mediated through a mechanism involving ERb
competing with PPARg for PPAR-g coactivator 1a [22].
ERs improve insulin sensitivity
E2, at physiological concentrations, favors insulin sensi-tivity, and E2 deficiency and/or resistance provokes insulinresistance. Perhaps the best evidence is that men lackingin E2 production because of mutations in the aromatasegene or men harboring E2 resistance because of geneticERa deficiency develop insulin resistance and/or glucoseintolerance [34,35]. Accordingly, male and female micewith E2 deficiency or E2 resistance caused by eliminationof the aromatase or ERa genes develop insulin resistance[12,13].
The cause of insulin resistance induced by E2 deficiencyor resistance is probably multifactorial. In one study,female mice lacking ERa did not show insulin resistancein skeletal muscle, but exhibited decreased insulin sup-pression of hepatic glucose production (HGP) during aeuglycemic hyperinsulinemic clamp under anesthesia,suggesting that ERa deficiency provokes hepatic insulinresistance [36]. Andrea Hevener and coworkers, however,Ribas et al. reported that ERa deficient female mice accu-mulate proinflammatory lipid intermediates in skeletalmuscle, leading to marked muscle insulin resistance,and that these mice when conscious under euglycemichyperinsulinemic clamp conditions, exhibited minoralterations in liver insulin sensitivity [14]. In addition,decreased expression of the insulin sensitive glucose trans-porter (GLUT)4 is observed in skeletal muscle of maleERa-deficient mice; this might contribute to the muscleinsulin resistance observed in these mice, as GLUT4 isessential to insulin-sensitive glucose transport in skeletalmuscle and WAT [37]. E2 treatment improves insulinresistance in female mice fed a HFD [29,38] and in obesefemale mice with genetic leptin resistance [30], through apathway at least partially dependent on ERa [31,38]. E2treatment also reduces HFD-induced insulin resistance inskeletal muscle by 50% during euglycemic hyperinsuline-mic clamp, in an ERa-dependent manner [38]. However, asdiscussed, E2 also suppresses lipogenesis and steatosis inliver of HFD-fed [29] and leptin resistant mice [30] sug-gesting that it protects from insulin resistance by prevent-ing ectopic lipid accumulation (lipotoxicity).
In summary, ERa deficiency decreases GLUT4 expres-sion in skeletal muscle, and impairs lipid homeostasis inskeletal muscle and liver of rodents, thus decreasingthe ability of insulin to suppress HGP and to promoteskeletal muscle glucose utilization. Accordingly, activationof ERa during HFD and genetic leptin resistance improvesinsulin resistance induced by ectopic lipid accumulation inskeletal muscle [29–31,38]. However, the effect of ERa inmediating insulin sensitivity via central mechanismsremains to be determined. In the absence of ERa signaling,ERb could promote insulin resistance in skeletal muscle.Ovariectomy in hyperestrogenic female ERa-deficientmice
(which suppresses E2 action through ERb), improves glu-cose tolerance and insulin sensitivity [39], and adminis-tration of an ERb selective agonist in male E2 deficientArKO mice decreases skeletal muscle GLUT4 expression[37]. Accordingly, administration of tamoxifen, acting as anERb antagonist in male ERa-deficient mice, increasedGLUT4 expression and improved insulin sensitivity [40].Interestingly, ERs modulate GLUT4 expression in WATand skeletal muscle in a tissue-specific manner. WhereasERb-mediated repression of GLUT4 predominates in skel-etal muscle, ERa-mediated induction of GLUT4 predomi-nates in WAT [40].
Finally, recent evidence indicates that ERb deficiencyprotects against diet induced insulin resistance in malemice by increasing PPARg signaling in adipose tissue,which indirectly improves skeletal muscle insulin actionby promoting lipid accumulation away from muscle andinto adipose tissue [33].
The physiological and genetic evidence indicates thatE2 and ERa favor insulin sensitivity in rodents andhumans of both sexes when E2 concentrations stay withina tight physiological window. Conversely, high doses ofestrogens provoke insulin resistance [41,42]. In fact, tworecent studies have reported that in postmenopausal wom-en, higher plasma levels of E2 (associated with highertestosterone levels) were strongly and prospectively relat-ed to an increased risk of developing T2D [43,44].
ERs favor b cell function and survival
The beneficial effect of estrogens on b cell function inhumans and rodents has been reviewed recently [11]. Wewill focus on the most important and recent developments.
There are gender dimorphisms in rodentmodels of b cellfailure that have helped identify the function of ERs in b
cells. A classic sexually dimorphic model of T2D is thetransgenic mouse overexpressing human islet amyloidpolypeptide (hIAPP) in pancreatic b cells. The hIAPP isa classic late b cell injury in T2D. Kahn and co-workersinitially reported that overexpression of hIAPP in isletspredisposes mice to the development of islet amyloid andhyperglycemia, with a strong male predominance [45].This led to the paradigm that suppression of E2 productionby ovariectomy enhanced islet amyloid formation in femalemice [46] and that conversely, E2 treatment preventsamyloid formation and b cell failure in males [47].
To study the antiapoptotic action of E2 on islets in vivo,we used the mouse model of b cell injury induced bystreptozotocin (STZ) and showed that circulating E2 actsas a protective hormone, preventing b cell apoptosis in vivoin both sexes and at physiological concentrations [48]. ERa
and ERb are expressed in rodent and human b cells in bothsexes, and exhibit a predominant extranuclear localization[48–50]. E2-activated ERa prevents islet apoptosis inmales and females via an ERE-independent pathway[50]. This is mediated via activation of extranuclearERs, with a predominant ERa effect [50]. Although theprecise signaling pathways are still under investigation, itappears that ERa and ERb prevent apoptosis via distinctpathways, independently of gene transcription or de novoprotein synthesis, suggesting that this cytoprotection hap-pens independently of nuclear events [50,51].
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
26

Author's personal copy
GPER is present in b cells, and in one study, GPER-deficient mice displayed altered insulin release from iso-lated islets stimulated with pharmacological concentra-tions of E2. In that study, impaired glucose-stimulatedinsulin secretion (GSIS) was observed in GPER-deficientmice, but the mice were chronically treated with E2 andinsulin sensitivity was not assessed [52]. However, we didnot observe any alteration in GSIS in another GPER-deficient mouse [50]. Thus, the role of GPER in GSISremains controversial.
Elimination of GPER predisposes to STZ-induced isletapoptosis in female but not in male mice [50]. We andothers have observed that pharmacological activation ofGPER by the agonist, G1, prevents oxidative stress andcytokine- induced apoptosis in cultured mouse and humanislets [50,53]. G1 has recently been shown to induce theexpression and activation of a small (36 kDa) ERa isoformlacking transcriptional activity and mediating rapid estro-gen signaling, suggesting that GPER signals as an inducerof ERa36 [54]. However, the observation that G1 cytopro-tection is lost in cultured GPER deficient islets furthersupports the functional significance of GPER itself in isletsurvival [50]. Recently, Nadal and co-workers reportedthat E2 activation of ERb enhances GSIS in cultured isletsby suppressing the ATP sensitive potassium channelthrough effects on the membrane atrial natriuretic peptidereceptor [55]. This finding shed new light on the role of ERb
in islet function.The Zucker diabetic fatty (ZDF) rat is a classic model of
T2D, and a classic example of sex dimorphism. Male ZDFrats develop pancreatic b cell failure to compensate forinsulin resistance leading to overt T2D [56]. Fifteen yearsago Roger Unger reported that b cell failure in male ZDFrats is secondary to islet triglyceride accumulation leadingto b cell apoptosis, and the concept of ‘lipotoxicity’ was born[57]. b cell failure occurs almost exclusively in male ZDFrats, whereas female ZDF rats remain normoglycemic [57].Interestingly, the islet triglyceride content in the adultZDF female is 70% lower than that of males [57] suggestingthat E2 prevents islet lipid accumulation. Indeed, werecently reported that E2 treatment of male ZDF ratssuppresses islet lipogenesis and prevents b cell failure,probably via ERa action in islets [58].
ERa is also important for insulin biosynthesis. We andothers have shown that exposure to physiological concen-trations of E2 increases b cell insulin gene expression andinsulin content via an extranuclear ERa-dependent mech-anism, involving Src and ERK kinases [49,59] and anincrease in neuroD-1 binding to the insulin promoter[59]. Thus, the elevated E2 concentration during pregnan-cy might participate in islet adaptation to the increasedmetabolic demand by enhancing insulin biosynthesis andrelease via ERa and ERb [49,55,59].
In summary, E2 at physiological concentrationsincreases insulin production and protects the pancreaticb cells against the major b cell injuries encountered indiabetes, such as lipotoxicity, hIAPP, oxidative stress andapoptosis.
Most ERa actions that control body weight, insulin sen-sitivity, and b cell biology [12–15,32,34,35,37,40,48,50] arepresent in both sexes, demonstrating that testosterone
aromatization in E2 acting on ERa is important to energyhomeostasis in males. However, the role of ERb has beenstudied in both sexes for b cell survival only [50], and foreither females for body weight regulation [18,22,33] ormales for insulin sensitivity [37,40]. Therefore, it is assumedthat ERa and ERb share similar metabolic function in bothsexes. Figures 1 and 2 summarize these actions in womenand men, respectively.
The androgen receptorMechanism of AR action
Androgens influence gene transcription through the acti-vation of the AR, a ligand-activated transcription factorthat subsequently binds as a homodimer with specific DNAmotifs in its target genes [60]. These DNA motifs, calledandrogen response elements (AREs), can be categorized asclassic AREs, which are recognized by glucocorticoid orprogesterone receptors, and AR-specific AREs, which dis-play selectivity for the AR [61]. As in the case of estrogens,over the past two decades evidence has accumulated toimplicate rapid responses to androgens, dependent orindependent of the AR [62].
AR prevent visceral fat accumulation in males
Testosterone deprivation in men contributes to the devel-opment of metabolic syndrome. There is an inverse rela-tionship between total serum testosterone, and the amountof visceral adipose tissue in the metabolic syndrome [63].This is observed in age related hypogonadism [64], inher-ited testosterone deficiency [65], and androgen deprivationduring treatment of prostate cancer [66]. Accordingly, inmen, high testosterone levels are linked to insulin sensi-tivity [67]. Evidence discussed in the ERs section demon-strates that aromatization of testosterone to E2 is crucialto energy homeostasis in males, suggesting that testoster-one acts as a prohormone in men to provide E2 for tissueenergy homeostasis. Indeed, orchidectomizedmale rodentstreated with either testosterone or E2 remain lean, al-though those treated with the pure androgen DHT (that is,not aromatized to E2), develop obesity [68].
Several lines of evidence demonstrate that testosteronehas anti-obesity actions that are mediated via AR. First,men with genetic androgen resistance linked to CAGrepeats in the AR gene, which decreases AR-mediatedgene transcription, have elevated visceral fat [69]. Second,male mice lacking AR develop late onset visceral obesitywith increased lipogenesis in WAT and liver [70,71]. Fur-thermore, AR is involved in adiponectin biology. Adiponec-tin levels are high in hypogonadal men and reduced bytestosterone therapy [72]. Testosterone infusion alsodecreases adiponectin in mice [73], an effect that is at leastpartially mediated via AR, as adiponectin is increased inAR-deficient mice [70]. Whether AR suppression of adipo-nectin reflects increased adiponectin sensitivity or de-creased adipocyte number remains to be determined.
The suppressing effect of testosterone on WAT mass inmales might be indirectly mediated via AR signaling inskeletal muscle. Several lines of evidence support thisscenario. First, in vitro, testosterone promotes the commit-ment of pluripotent mesenchymal stem cells into myogeniclineage while inhibiting the adipogenic lineage via an
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
27
Author's personal copy
AR-dependentmechanism [74]mediated via non-canonicalWnt signaling [75]. This androgenic anabolism involves aninduction of IGF1, leading to nuclear accumulation of b-catenin, a pro-myogenic, anti-adipogenic stem cell regula-tory factor [76]. Accordingly, selective overexpression of ARin muscle cells of transgenic male rats increases lean masswith hypertrophy of type IIb fibers, increasing oxidativemetabolism, and decreasing adipocyte size and WAT mass[77]. Conversely, and consistent with thismodel, malemicelacking AR in adipose tissue are not obese. These mice
show increased WAT production of leptin without leptinresistance [78]. Thus, activation of AR in skeletal musclemight indirectly decrease WAT mass by increasing muscleoxidative metabolism or through the release of a circulat-ing factor.
The action of AR in skeletal muscle promotes insulin
sensitivity in males
Apart from increasing visceral WAT, the mechanism ofinsulin resistance induced by AR deficiency probably
[()TD$FIG]
Skeletal muscle↑ Insulin sensitivity↑ GLUT4↑ Lipid oxidation
CNS↓ Food intake↑ Energy expenditure↓ Obesity
Pancreatic islets↑ Survival↑ Insulin biosynthesis↑ GSIS↓ Lipogenesis
Liver↑ Insulin sensitivity↓ HGP↓ Lipogenesis?
WAT↓ Total WAT mass↓ Adipogenesis↓ FFA uptake↓ Lipogenesis↑ Lipolysis↑ SC distribution
E2 E2
E2 E2
E2E2
E2
E2 E2
Direct E2 effectson tissues
E2 effects mediated via the autonomousnervous system
ERα
CNS↓ Food intake↑ Energy expenditure↓ Obesity
Pancreatic islets↑ Survival↑ GSIS
Liver↓ TG accumulation
WAT↓ PPARγ action↓ Adipogenesis↓ Lipogenesis↓ Total WAT mass
ERβ
Skeletal muscle↓ Insulin sensitivity↓ GLUT4
E2 E2
E2 E2
E2E2
E2 E2
Key:
TRENDS in Endocrinology & Metabolism
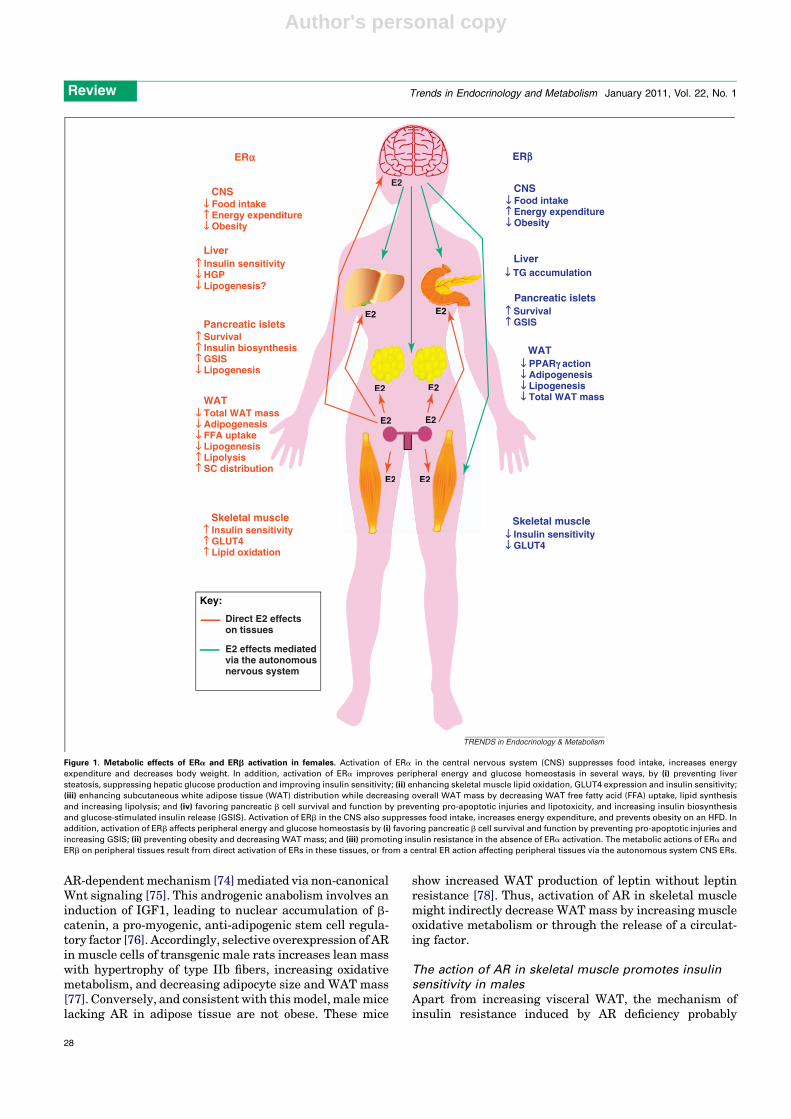
Figure 1. Metabolic effects of ERa and ERb activation in females. Activation of ERa in the central nervous system (CNS) suppresses food intake, increases energy
expenditure and decreases body weight. In addition, activation of ERa improves peripheral energy and glucose homeostasis in several ways, by (i) preventing liver
steatosis, suppressing hepatic glucose production and improving insulin sensitivity; (ii) enhancing skeletal muscle lipid oxidation, GLUT4 expression and insulin sensitivity;
(iii) enhancing subcutaneous white adipose tissue (WAT) distribution while decreasing overall WAT mass by decreasing WAT free fatty acid (FFA) uptake, lipid synthesis
and increasing lipolysis; and (iv) favoring pancreatic b cell survival and function by preventing pro-apoptotic injuries and lipotoxicity, and increasing insulin biosynthesis
and glucose-stimulated insulin release (GSIS). Activation of ERb in the CNS also suppresses food intake, increases energy expenditure, and prevents obesity on an HFD. In
addition, activation of ERb affects peripheral energy and glucose homeostasis by (i) favoring pancreatic b cell survival and function by preventing pro-apoptotic injuries and
increasing GSIS; (ii) preventing obesity and decreasing WAT mass; and (iii) promoting insulin resistance in the absence of ERa activation. The metabolic actions of ERa and
ERb on peripheral tissues result from direct activation of ERs in these tissues, or from a central ER action affecting peripheral tissues via the autonomous system CNS ERs.
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
28
Author's personal copy
involves a decrease in the transcription factor PGC1a inskeletal muscle. Indeed, PGC1a stimulates mitochondrialbiogenesis and skeletal muscle oxidative fibers, and is thusa molecular marker of muscle insulin sensitivity. A de-crease in PGC1a expression in skeletal muscle of patientswith T2D is associated with insulin resistance [79]. Simi-larly, in men, low testosterone levels are associated withlow PGC1a expression levels in muscle [67], and AR-defi-cient mice have low levels of PGC1a in tissues [70]. Thus,testosterone deficiency promotes insulin resistance at least
partially via an AR-dependent mechanism, involving adecrease in PGC1a-mediated oxidative and insulin sensi-tive muscle fibers, and increased visceral WAT and liversteatosis.
Central AR actions favor energy homeostasis in males
AR are expressed in the brain more abundantly in males[80]. Male AR deficient mice develop obesity without hyper-phagia, but with reduced locomotor activity and energyexpenditure associatedwith decreased brown adipose tissue
[()TD$FIG]
TT T
T T
CNS↓ Food intake↑ Energy expenditure↓ Obesity
Pancreatic islets↑ Survival↑ GSIS
Liver↓ TG accumulation
WAT↓ PPARγ action↓ Adipogenesis↓ Lipogenesis↓ Total WAT mass
ERβ
Skeletal muscle↓ Insulin sensitivity↓ GLUT4
Direct E2 effects
E2 effects mediated via the autonomousnervous system
Key:
Skeletal muscle↑ Insulin sensitivity↑ GLUT4↑ Lipid oxidation
CNS↓ Food intake↑ Energy expenditure↓ Obesity
Pancreatic islets↑ Survival↑ Insulin biosynthesis↑ GSIS↓ Lipogenesis
Liver↑ Insulin sensitivity↓ HGP↓ Lipogenesis?
WAT↓ Total WAT mass↓ Adipogenesis↓ FFA uptake↓ Lipogenesis↑ Lipolysis↑ SC distribution
ERβ
TRENDS in Endocrinology & Metabolism
TT T
T T
E2
E2
E2
E2E2
E2
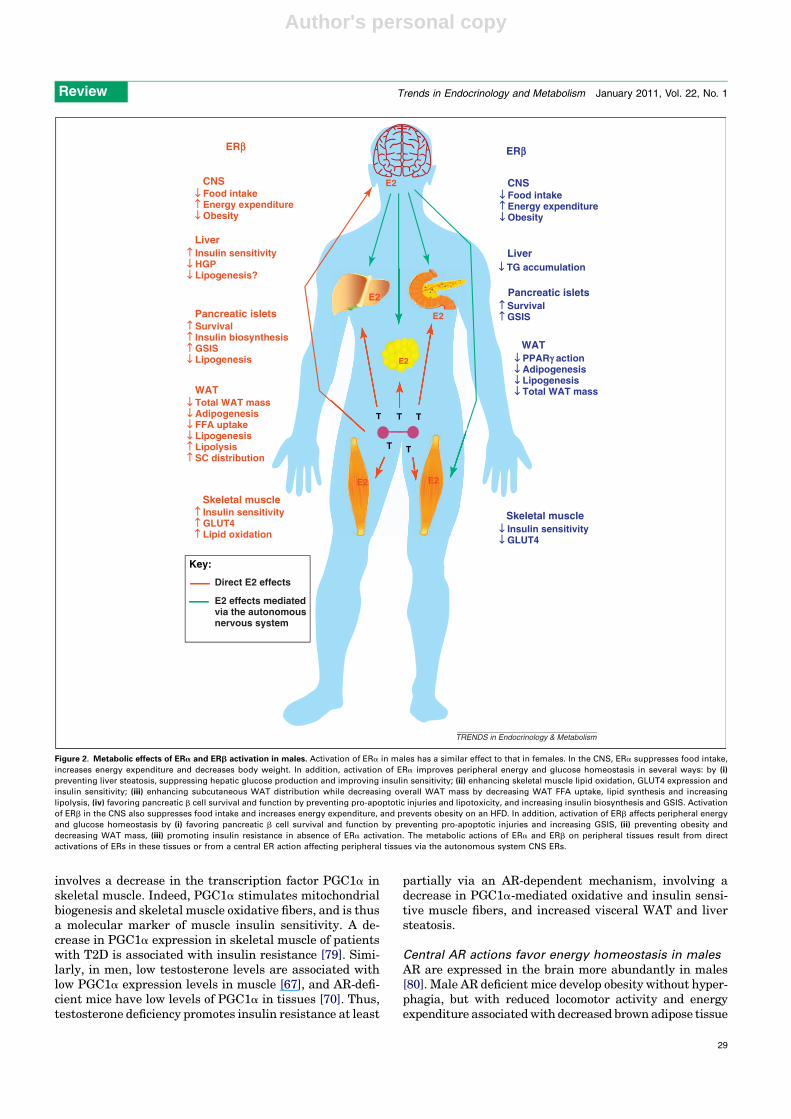
Figure 2. Metabolic effects of ERa and ERb activation in males. Activation of ERa in males has a similar effect to that in females. In the CNS, ERa suppresses food intake,
increases energy expenditure and decreases body weight. In addition, activation of ERa improves peripheral energy and glucose homeostasis in several ways: by (i)
preventing liver steatosis, suppressing hepatic glucose production and improving insulin sensitivity; (ii) enhancing skeletal muscle lipid oxidation, GLUT4 expression and
insulin sensitivity; (iii) enhancing subcutaneous WAT distribution while decreasing overall WAT mass by decreasing WAT FFA uptake, lipid synthesis and increasing
lipolysis, (iv) favoring pancreatic b cell survival and function by preventing pro-apoptotic injuries and lipotoxicity, and increasing insulin biosynthesis and GSIS. Activation
of ERb in the CNS also suppresses food intake and increases energy expenditure, and prevents obesity on an HFD. In addition, activation of ERb affects peripheral energy
and glucose homeostasis by (i) favoring pancreatic b cell survival and function by preventing pro-apoptotic injuries and increasing GSIS, (ii) preventing obesity and
decreasing WAT mass, (iii) promoting insulin resistance in absence of ERa activation. The metabolic actions of ERa and ERb on peripheral tissues result from direct
activations of ERs in these tissues or from a central ER action affecting peripheral tissues via the autonomous system CNS ERs.
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
29
Author's personal copy
thermogenesis [70]. AR suppress lipogenesis inmales, andmale AR deficient mice exhibit unsuppressed lipogenesisin muscle and liver [70,71]. AR also function in the malehypothalamus to favor central leptin signaling. AR-defi-cient male mice exhibit a failure of leptin to promoteSTAT3 nuclear localization in ARC neurons, and to sup-press food intake and reduce body weight even before theonset of overt obesity [80]. In summary, in males, AR areinvolved in the control of WAT mass via central andperipheral effects.
AR and b cells in males
Early studies reported that testosterone accelerates hy-perglycemic decompensation via an AR-dependent mecha-nism in male mouse models of insulin-deficient diabetes inwhich b cell destruction is induced by STZ [81,82]. Recent-ly, however, it was reported that testosterone protectsagainst early apoptotic damage induced by STZ in malerat pancreas via an AR dependent mechanism [83,84]. Inthe later study, however, the effect of testosterone ondiabetes incidence was not reported, therefore the role ofAR in male b cell survival and function still needs clarifi-cation. Figure 3 summarizes the effects of AR on energyhomeostasis in men.
Role of AR in females
The role of AR in female energy metabolism is not wellcharacterized. Although AR deficiency is reported to haveno effect on body weight in female mice [85], women withcomplete androgen insensitivity syndrome have increasedtotal fat mass compared with both female and male agematched control subjects [86]. Therefore, further studiesare needed to determine the role of AR in female energymetabolism.
Although the consequence of AR deficiency in females innot well studied, the association between hyperandrogeni-city and diabetes in women has been known for almost acentury [87]. It has been postulated that excess androgenprovokes insulin resistance. In women, hyperandrogenismis a risk factor for metabolic syndrome, independently ofobesity and insulin resistance [88]. Furthermore, testos-terone infusion in healthy women decreases insulin-stimulated, whole body glucose uptake [89]. The role ofexcess testosterone in promoting skeletal muscle insulinresistance with fiber type switch has also been confirmedfrom studies in female rodents [90].
Hyperandrogenemia is also associated with pancreaticb cell dysfunction [91–93]. In some studies of women withpolycystic ovary syndrome (PCOS), b cell dysfunction isclosely associated with the degree of androgenicity, inde-pendent of insulin resistance, raising the possibility thatexcess testosterone might predispose to secondary b cellfailure [92,93]. Consistent with this hypothesis, in mice,testosterone accelerates the hyperglycemic decompensa-tion in experimental models of insulin-dependent diabetesin which b cell destruction is induced by oxidative stress orinflammation [81,94]. In addition, hyperandrogenemia inwomen with PCOS is accompanied by systemic oxidativestress [95], and excess testosterone in female mice similar-ly provokes systemic oxidative stress via an AR-dependentmechanism [94]. We previously showed that in the pres-
ence of a prior b cell injury, excess testosterone predisposesfemale mice to b cell failure via an AR-dependent mecha-nism [94], which could involve an AR present in b cells [96].Thus, excess AR activation in b cells might participate inthe b cell dysfunction observed in women with androgenexcess.
Despite accumulated evidence that testosterone excessalters metabolism in females, it is not clear whether tes-tosterone excess initiates metabolic abnormalities or per-petuates them. Indeed, treatment with AR antagonists orsuppression of ovarian androgen production with GnRHanalogues in hyperandrogenic women does not alwaysimprove insulin resistance [97], thereby suggesting thatexcess androgen in women might not be instrumental inthe metabolic abnormalities but rather an aggravatingfactor. Further studies in this area are needed.
[()TD$FIG]
T
Direct Key:
DHT effects
E2 DHT effects mediated via the autonomous nervoussystem
AR
T
DHT
T
DHT
Skeletal muscle↑ Insulin sensitivity↑ PGC-1α↑ Lipid oxidation
CNS↓ Food intake↑ Energy expenditure
Pancreatic islets↑ survival?
Liver↓ Lipogenesis
WAT↓ Adipogenesis↓ Visceral distribution↓ Lipogenesis↓ Adiponectin↓ Leptin
DHT
DHT DHT
DHT
TT
DHT
T
DHTDDHTD
TDHTT DDHTTTTTTTTTTTTTTTTTTTTTTTD
TRENDS in Endocrinology & Metabolism
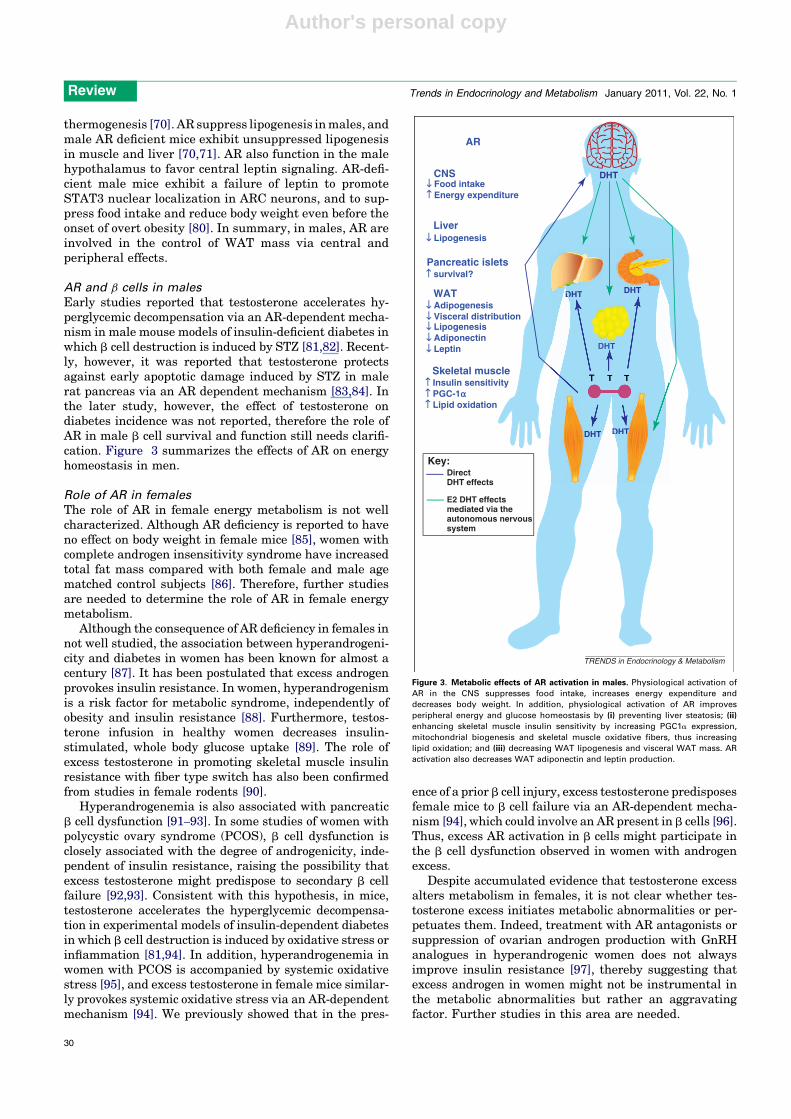
Figure 3. Metabolic effects of AR activation in males. Physiological activation of
AR in the CNS suppresses food intake, increases energy expenditure and
decreases body weight. In addition, physiological activation of AR improves
peripheral energy and glucose homeostasis by (i) preventing liver steatosis; (ii)
enhancing skeletal muscle insulin sensitivity by increasing PGC1a expression,
mitochondrial biogenesis and skeletal muscle oxidative fibers, thus increasing
lipid oxidation; and (iii) decreasing WAT lipogenesis and visceral WAT mass. AR
activation also decreases WAT adiponectin and leptin production.
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
30

Author's personal copy
Conclusions and future perspectivesE2 and testosterone are crucial hormonal signals main-taining energy homeostasis in both sexes, and the effect ofE2 treatment on obesity and the prevention of obesity anddiabetes is one of the most powerful observations of rodentphysiology. Although men have lower circulating E2 con-centrations than do premenopausal women, aromatizationof circulating testosterone to E2 in target metabolic tissuesequilibrates cellular E2 concentrations, and ER activationis similarly crucial in both sexes in promoting fuel homeo-stasis. Conversely, and probably reflecting the lower circu-lating and cellular testosterone andDHT concentrations infemales, AR activation is weak in females and thus the ARis less important. Indeed, if androgen concentrations in-crease in females to the level of males, this provokes excessAR activation leading to metabolic disturbances. Themechanism of this bidirectional modulation of metabolismby AR between males and females is unknown. Because ofthis sex-specific stoichoimetry of ER/AR activation, the ARis primarily a male drug target, whereas ERs are sex non-specific drug targets to improve metabolic diseases. Themajor obstacle to the development of ER and AR ligands totreat metabolic diseases is the concern about hormone-dependent cancer. Further studies are thus needed toidentify and develop new ligands that prevent diabetesand obesity that lack the mitogenic actions predisposing tohormone-dependent cancers. This can be achieved by tar-geting E2 or testosterone to the appropriate cells or devel-oping novel selective ER/AR modulators that retain thebeneficial effects of their ligand in selected tissues whilelacking mitogenic actions in reproductive organs.
AcknowledgementsThis work was supported by grants from National Institutes of Health(P50 HD044405, RO1 DK074970-01), the Juvenile Diabetes ResearchFoundation (1-2006-837), and the March of Dimes (6-FY07-678) and byNorthwestern University Institute for Women’s Health Research PioneerAward.
References1 Wild, S. et al. (2004) Global prevalence of diabetes: estimates for the
year 2000 and projections for 2030. Diabetes Care 27, 1047–10532 Alberti, K.G. et al. (2006) Metabolic syndrome—a new world-wide
definition. A Consensus Statement from the International DiabetesFederation. Diabet. Med. 23, 469–480
3 Hill, J.W. et al. (2008) Hypothalamic pathways linking energy balanceand reproduction. Am. J. Physiol. Endocrinol. Metab. 294, E827–
8324 Shi, H. et al. (2009) Sexual differences in the control of energy
homeostasis. Front Neuroendocrinol. 30, 396–4045 Carr MC (2003) The emergence of the metabolic syndrome with
menopause. J. Clin. Endocrinol. Metab. 88, 2404–24116 Zitzmann M (2009) Testosterone deficiency, insulin resistance and the
metabolic syndrome. Nat. Rev. Endocrinol. 5, 673–6817 Simpson, E.R. (2003) Sources of estrogen and their importance. J.
Steroid. Biochem. Mol. Biol. 86, 225–2308 Weihua, Z. et al. (2002) An endocrine pathway in the prostate, ERbeta,
AR, 5alpha-androstane-3beta,17beta-diol, and CYP7B1, regulatesprostate growth. Proc. Natl. Acad. Sci. U. S. A. 99, 13589–13594
9 Nilsson, S. et al. (2001)Mechanisms of estrogen action.Physiol. Rev. 81,1535–1565
10 Levin, E.R. (2009) Plasma membrane estrogen receptors. TrendsEndocrinol. Metab. 20, 477–482
11 Liu, S. and Mauvais-Jarvis, F. (2010) Minireview: Estrogenicprotection of beta-cell failure in metabolic diseases. Endocrinology151, 859–864
12 Jones, M.E. et al. (2000) Aromatase-deficient (ArKO) mice have aphenotype of increased adiposity. Proc. Natl. Acad. Sci. U. S. A. 97,12735–12740
13 Heine, P.A. et al. (2000) Increased adipose tissue in male and femaleestrogen receptor-alpha knockout mice. Proc. Natl. Acad. Sci. U. S. A.97, 12729–12734
14 Ribas, V. et al. (2009) Impaired Oxidative metabolism andinflammation are associated with insulin resistance in ER{alpha}deficient mice. Am. J. Physiol. Endocrinol. Metab. 298, E304–E319
15 Clegg, D.J. et al. (2006) Gonadal hormones determine sensitivity tocentral leptin and insulin. Diabetes 55, 978–987
16 Gao, Q. et al. (2007) Anorectic estrogen mimics leptin’s effect on therewiring of melanocortin cells and Stat3 signaling in obese animals.Nat. Med. 13, 89–94
17 Geary, N. et al. (2001) Deficits in E2-dependent control of feeding,weight gain, and cholecystokinin satiation in ER-alpha null mice.Endocrinology 142, 4751–4757
18 Liang, Y.Q. et al. (2002) Estrogen receptor beta is involved in theanorectic action of estrogen. Int. J. Obes. Relat. Metab. Disord. 26,1103–1109
19 Olofsson, L.E. et al. (2009) Functional requirement of AgRP and NPYneurons in ovarian cycle-dependent regulation of food intake. Proc.Natl. Acad. Sci. U. S. A. 106, 15932–15937
20 Thammacharoen, S. et al. (2008) Hindbrain administration of estradiolinhibits feeding and activates estrogen receptor-alpha-expressing cellsin the nucleus tractus solitarius of ovariectomized rats. Endocrinology149, 1609–1617
21 Musatov, S. et al. (2007) Silencing of estrogen receptor alpha in theventromedial nucleus of hypothalamus leads to metabolic syndrome.Proc. Natl. Acad. Sci. U. S. A. 104, 2501–2506
22 Yepuru, M. et al. (2010) Estrogen receptor-{beta} selective ligandsalleviate high-fat diet- and ovariectomy-induced obesity in mice. J.Biol. Chem. 285, 31292–31303
23 Haas, E. et al. (2009) Regulatory role of G protein-coupled estrogenreceptor for vascular function and obesity. Circ. Res. 104, 288–291
24 Langer, G. et al. (2010) A critical review of fundamental controversiesin the field of GPR30 research. Steroids 75, 603–610
25 Dhillon, S.S. and BelshamD.D. (2010) Estrogen inhibits NPY secretionthrough membrane-associated estrogen receptor (ER)-alpha in clonal,immortalized hypothalamic neurons. Int. J. Obes. (Lond.). Jun 15[Epub ahead of print]
26 Qiu, J. et al. (2006) A G-protein-coupled estrogen receptor is involved inhypothalamic control of energy homeostasis. J. Neurosci. 26, 5649–5655
27 Roepke, T.A. et al. (2010) Contribution of a membrane estrogenreceptor to the estrogenic regulation of body temperature andenergy homeostasis. Endocrinology 15, 4926–4937
28 D’Eon, T.M. et al. (2005) Estrogen regulation of adiposity and fuelpartitioning. Evidence of genomic and non-genomic regulation oflipogenic and oxidative pathways. J. Biol. Chem. 280, 35983–35991
29 Bryzgalova, G. et al. (2008) Mechanisms of antidiabetogenic and bodyweight-lowering effects of estrogen in high-fat diet-fed mice. Am. J.Physiol. Endocrinol. Metab. 295, E904–912
30 Gao, H. et al. (2006) Long-term administration of estradiol decreasesexpression of hepatic lipogenic genes and improves insulin sensitivityin ob/ob mice: a possible mechanism is through direct regulation ofsignal transducer and activator of transcription 3.Mol. Endocrinol. 20,1287–1299
31 Lundholm, L. et al. (2008) The estrogen receptor {alpha}-selectiveagonist propyl pyrazole triol improves glucose tolerance in ob/obmice; potential molecular mechanisms. J. Endocrinol. 199, 275–286
32 Cooke, P.S. and Naaz, A. (2004) Role of estrogens in adipocytedevelopment and function. Exp. Biol. Med. (Maywood) 229, 1127–1135
33 Foryst-Ludwig, A. et al. (2008) Metabolic actions of estrogen receptorbeta (ERbeta) are mediated by a negative cross-talk withPPARgamma. PLoS Genet. 4, e1000108
34 Morishima, A. et al. (1995) Aromatase deficiency in male and femalesiblings caused by a novel mutation and the physiological role ofestrogens. J. Clin. Endocrinol. Metab. 80, 3689–3698
35 Smith, E.P. et al. (1994) Estrogen resistance caused by a mutation inthe estrogen-receptor gene in a man. N. Engl. J. Med. 331, 1056–1061
36 Bryzgalova, G. et al. (2006) Evidence that oestrogen receptor-alphaplays an important role in the regulation of glucose homeostasis inmice: insulin sensitivity in the liver. Diabetologia 49, 588–597
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
31

Author's personal copy
37 Barros, R.P. et al. (2006) Muscle GLUT4 regulation by estrogenreceptors ERbeta and ERalpha. Proc. Natl. Acad. Sci. U. S. A. 103,1605–1608
38 Riant, E. et al. (2009) Estrogens protect against high-fat diet-inducedinsulin resistance and glucose intolerance in mice. Endocrinology 150,2109–2117
39 Naaz, A. et al. (2002) Effect of ovariectomy on adipose tissue of mice inthe absence of estrogen receptor alpha (ERalpha): a potential role forestrogen receptor beta (ERbeta). Horm. Metab. Res. 34, 758–763
40 Barros, R.P. et al. (2009) Participation of ERalpha and ERbeta inglucose homeostasis in skeletal muscle and white adipose tissue.Am. J. Physiol. Endocrinol. Metab. 297, E124–133
41 Barros, R.P. et al. (2006) Estrogen receptors: new players in diabetesmellitus. Trends Mol. Med. 12, 425–431
42 Nadal, A. et al. (2009) The pancreatic beta-cell as a target of estrogensand xenoestrogens: Implications for blood glucose homeostasis anddiabetes. Mol. Cell Endocrinol. 304, 63–68
43 Ding, E.L. et al. (2007) Plasma sex steroid hormones and risk ofdeveloping type 2 diabetes in women: a prospective study.Diabetologia 50, 2076–2084
44 Kalyani, R.R. et al. (2009) The association of endogenous sex hormones,adiposity, and insulin resistance with incident diabetes inpostmenopausal women. J. Clin. Endocrinol. Metab. 94, 4127–4135
45 Verchere, C.B. et al. (1996) Islet amyloid formation associated withhyperglycemia in transgenic mice with pancreatic beta cell expressionof human islet amyloid polypeptide. Proc. Natl. Acad. Sci. U. S. A. 93,3492–3496
46 Kahn, S.E. et al. (2000) Oophorectomy promotes islet amyloidformation in a transgenic mouse model of Type II diabetes.Diabetologia 43, 1309–1312
47 Geisler, J.G. et al. (2002) Estrogen can prevent or reverse obesity anddiabetes in mice expressing human islet amyloid polypeptide.Diabetes51, 2158–2169
48 Le May, C. et al. (2006) Estrogens protect pancreatic beta-cells fromapoptosis and prevent insulin-deficient diabetes mellitus in mice. Proc.Natl. Acad. Sci. U. S. A. 103, 9232–9237
49 Alonso-Magdalena, P. et al. (2008) Pancreatic insulin contentregulation by the estrogen receptor ER alpha. PLoS ONE 3, e2069
50 Liu, S. et al. (2009) Importance of extranuclear estrogen receptor-alphaand membrane G protein-coupled estrogen receptor in pancreatic isletsurvival. Diabetes 58, 2292–2302
51 Liu, S. and Mauvais-Jarvis, F. (2009) Rapid, nongenomic estrogenactions protect pancreatic islet survival. Islets 1, 273–275
52 Martensson, U.E. et al. (2009) Deletion of the G protein-coupledreceptor 30 impairs glucose tolerance, reduces bone growth,increases blood pressure, and eliminates estradiol-stimulatedinsulin release in female mice. Endocrinology 150, 687–698
53 Balhuizen, A. et al. (2010) Activation of G protein-coupled receptor 30modulates hormone secretion and counteracts cytokine-inducedapoptosis in pancreatic islets of female mice. Mol. Cell Endocrinol.320, 16–24
54 Kang, L. et al. (2010) Involvement of estrogen receptor variant ER-alpha36, not GPR30, in nongenomic estrogen signaling. Mol.Endocrinol. 24, 709–721
55 Soriano, S. et al. (2009) Rapid regulation of K(ATP) channel activity by17{beta}-estradiol in pancreatic {beta}-cells involves the estrogenreceptor {beta} and the atrial natriuretic peptide receptor. Mol.Endocrinol. 23, 1973–1982
56 Tokuyama, Y. et al. (1995) Evolution of beta-cell dysfunction in themale Zucker diabetic fatty rat. Diabetes 44, 1447–1457
57 Lee, Y. et al. (1994) Beta-cell lipotoxicity in the pathogenesis of non-insulin-dependent diabetes mellitus of obese rats: impairment inadipocyte-beta-cell relationships. Proc. Natl. Acad. Sci. U. S. A. 91,10878–10882
58 Tiano J, et al., (2010) The extranuclear estrogen receptora improvespancreatic islet lipid homeostasis. The Endocrine Society’s 92ndAnnual Meeting San Diego, CA June 19–22, 2010 OR18–6
59 Wong, W.P. et al. (2010) Extranuclear estrogen receptor-{alpha}stimulates NeuroD1 binding to the insulin promoter and favorsinsulin synthesis. Proc. Natl. Acad. Sci. U. S. A. 107, 13057–13062
60 Beato M (1989) Gene regulation by steroid hormones. Cell 56, 335–34461 Schoenmakers, E. et al. (2000) Differences in DNA binding
characteristics of the androgen and glucocorticoid receptors can
determine hormone-specific responses. J. Biol. Chem. 275, 12290–
1229762 Foradori, C.D. et al. (2008) Non-genomic actions of androgens. Front
Neuroendocrinol. 29, 169–18163 Khaw, K.T. and Barrett-Connor, E. (1992) Lower endogenous
androgens predict central adiposity in men. Ann. Epidemiol. 2, 675–
68264 Zitzmann, M. et al. (2006) Association of specific symptoms and
metabolic risks with serum testosterone in older men. J. Clin.Endocrinol. Metab. 91, 4335–4343
65 Bojesen, A. et al. (2006) The metabolic syndrome is frequent inKlinefelter’s syndrome and is associated with abdominal obesity andhypogonadism. Diabetes Care 29, 1591–1598
66 Basaria, S. et al. (2006) Hyperglycemia and insulin resistance in menwith prostate carcinoma who receive androgen-deprivation therapy.Cancer 106, 581–588
67 Pitteloud, N. et al. (2005) Relationship between testosterone levels,insulin sensitivity, and mitochondrial function in men. Diabetes Care28, 1636–1642
68 Moverare-Skrtic, S. et al. (2006) Dihydrotestosterone treatment resultsin obesity and altered lipid metabolism in orchidectomized mice.Obesity (Silver Spring) 14, 662–672
69 Zitzmann, M. et al. (2003) The CAG repeat polymorphism in theandrogen receptor gene modulates body fat mass and serumconcentrations of leptin and insulin in men. Diabetologia 46, 31–39
70 Fan, W. et al. (2005) Androgen receptor null male mice develop late-onset obesity caused by decreased energy expenditure and lipolyticactivity but show normal insulin sensitivity with high adiponectinsecretion. Diabetes 54, 1000–1008
71 Lin, H.Y. et al. (2005) Insulin and leptin resistance withhyperleptinemia in mice lacking androgen receptor. Diabetes 54,1717–1725
72 Lanfranco, F. et al. (2004) Serum adiponectin levels in hypogonadalmales: influence of testosterone replacement therapy. Clin. Endocrinol.(Oxf.) 60, 500–507
73 Nishizawa, H. et al. (2002) Androgens decrease plasma adiponectin,an insulin-sensitizing adipocyte-derived protein. Diabetes 51, 2734–
274174 Singh, R. et al. (2003) Androgens stimulate myogenic differentiation
and inhibit adipogenesis in C3H 10T1/2 pluripotent cells through anandrogen receptor-mediated pathway. Endocrinology 144, 5081–5088
75 Singh, R. et al. (2006) Testosterone inhibits adipogenic differentiationin 3T3-L1 cells: nuclear translocation of androgen receptor complexwith beta-catenin and T-cell factor 4 may bypass canonical Wntsignaling to down-regulate adipogenic transcription factors.Endocrinology 147, 141–154
76 Gentile, M.A. et al. (2010) Androgen-mediated improvement of bodycomposition and muscle function involves a novel early transcriptionalprogram including IGF1, mechano growth factor, and induction of{beta}-catenin. J. Mol. Endocrinol. 44, 55–73
77 Fernando, S.M. et al. (2010) Myocyte androgen receptors increasemetabolic rate and improve body composition by reducing fat mass.Endocrinology 151, 3125–3132
78 Yu, I.C. et al. (2008) Hyperleptinemia without obesity in male micelacking androgen receptor in adipose tissue. Endocrinology 149, 2361–
236879 Mootha, V.K. et al. (2003) PGC-1alpha-responsive genes involved in
oxidative phosphorylation are coordinately downregulated in humandiabetes. Nat. Genet. 34, 267–273
80 Fan, W. et al. (2008) Functional potentiation of leptin-signaltransducer and activator of transcription 3 signaling by theandrogen receptor. Endocrinology 149, 6028–6036
81 Maclaren, N.K. et al. (1980) Androgen sensitization of streptozotocin-induced diabetes in mice. Diabetes 29, 710–716
82 Paik, S.G. et al. (1982) Induction of insulin-dependent diabetes bystreptozotocin. Inhibition by estrogens and potentiation by androgens.Diabetes 31, 724–729
83 Morimoto, S. et al. (2005) Protective effect of testosterone on earlyapoptotic damage induced by streptozotocin in rat pancreas.J. Endocrinol. 187, 217–224
84 Palomar-Morales, M. et al. (2010) The protective effect of testosteroneon streptozotocin-induced apoptosis in beta cells is sex specific.Pancreas 39, 193–200
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
32

Author's personal copy
85 Sato, T. et al. (2003) Late onset of obesity in male androgen receptor-deficient (ARKO)mice.Biochem. Biophys. Res. Commun. 300, 167–171
86 Dati, E. et al. (2009) Body composition and metabolic profile in womenwith complete androgen insensitivity syndrome. Sex Dev. 3, 188–193
87 Achard, C. and Thiers, J. (1921) Le virilisme pilaire et son association al’insuffisance glycolytique (diabete des femmes a barbe). Bull. Acad.Natl. Med. Paris 86, 51–55
88 Coviello, A.D. et al. (2006) Adolescent girls with polycystic ovarysyndrome have an increased risk of the metabolic syndromeassociated with increasing androgen levels independent of obesityand insulin resistance. J. Clin. Endocrinol. Metab. 91, 492–497
89 Diamond, M.P. et al. (1998) Effects of methyltestosterone on insulinsecretion and sensitivity in women. J. Clin. Endocrinol. Metab. 83,4420–4425
90 Holmang, A. et al. (1990) Effects of testosterone on muscle insulinsensitivity and morphology in female rats. Am. J. Physiol. 259,E555–560
91 O’Meara, N.M. et al. (1993) Defects in beta-cell function in functionalovarian hyperandrogenism. J. Clin. Endocrinol. Metab. 76, 1241–1247
92 Holte, J. et al. (1994) Enhanced early insulin response to glucose inrelation to insulin resistance in women with polycystic ovary syndromeand normal glucose tolerance. J. Clin. Endocrinol. Metab. 78, 1052–
105893 Goodarzi, M.O. et al. (2005) beta-Cell function: a key pathological
determinant in polycystic ovary syndrome. J. Clin. Endocrinol.Metab. 90, 310–315
94 Liu, S. et al. (2010) Androgen excess produces systemic oxidativestress and predisposes to beta-cell failure in female mice. PLoS One5, e11302
95 Gonzalez, F. et al. (2006) Reactive oxygen species-induced oxidativestress in the development of insulin resistance and hyperandrogenismin polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 91, 336–340
96 Navarro, G. et al. (2010) Androgen excess in females predisposes toinsulin deficiency via AR in beta-cells. 92nd Annual Meeting of theEndocrine Society, San Diego, CA, 19–22 June 2010, PI-468
97 Corbould A (2008) Effects of androgens on insulin action in women: isandrogen excess a component of female metabolic syndrome? DiabetesMetab. Res. Rev. 24, 520–532
Review Trends in Endocrinology and Metabolism January 2011, Vol. 22, No. 1
33
Related Documents











