REGULAR ARTICLE Estradiol effects on intracellular Ca 2+ homeostasis in bovine brain-derived endothelial cells Matteo Suman & Marta Giacomello & Livio Corain & Cristina Ballarin & Stefano Montelli & Bruno Cozzi & Antonella Peruffo Received: 24 October 2011 / Accepted: 31 May 2012 / Published online: 20 July 2012 # Springer-Verlag 2012 Abstract Estrogens diversely affect various physiological processes by genomic or non-genomic mechanisms, in both excitable and non-excitable cells. Additional to the trophic effects of estrogens promoting cell growth and differentiation, recent experimental evidence highlights their involvement in the regulation of intracellular Ca 2+ homeostasis. The effects of estrogens on excitable cells are well documented. However, these steroids also influence numerous physiological events in non-excitable cells, such as fibroblasts or vascular endothelial cells. We have focused our attention on an immortalized endothelial-like cell line derived from fetal bovine cerebellum. Estradiol (E 2 ) effects on intracellular Ca 2+ homeostasis were tested by varying the exposure time to the hormone (8, 24, 48 h). Calcium measurements were performed with genetically encoded Ca 2+ probes (Cameleons) targeted to the main subcellular compartments involved in intracellular Ca 2 + homeostasis (cytosol, endoplasmic reticulum, mitochondria). Mitochondrial Ca 2+ uptake significantly decreased after 48-h exposure to E 2 , whereas cytosolic and endoplasmic reticulum responses were unaffected. The effect of E 2 on mitochondrial Ca 2+ handling was blocked by ICI 182,780, a pure estrogen receptor antagonist, suggesting that the effect was estrogen-receptor–mediated. To evaluate whether the decrease of Ca 2+ uptake affected mitochondrial membrane potential (ΔΨm), cells were monitored in the presence of tetra-methyl-rhodamine-methylester; no significant changes were seen between cells treated with E 2 and controls. To investigate a mechanism of action, we assessed the possibile involvement of the permeability transition pore (PTP), an inner mitochondrial membrane channel influencing energy metabolism and cell viability. We treated cells with CyclosporinA (CsA), which binds to the matrix chaperone cyclophilin-D and regulates PTP opening. CsA reversed the effects of a 48-h treatment with E 2 , suggesting a possible transcriptional modulation of proteins involved in the mitochondrial permeability transition process. Keywords Estradiol . Endothelial cells . Mitochondria . Ca 2+ homeostasis . Bovine Introduction The concentration of calcium [Ca 2+ ] in a cell is managed by specific intracellular organelles, such as the endoplasmic reticulum (ER) and mitochondria. In the lumen of ER, the [Ca 2+ ] depends mainly on the balanced activity of the inositol triphosphate (IP 3 ) receptor (IP 3 R) and of the sarco-endoplasmic reticulum Ca 2+ ATPase (SERCA), which drives the ion from the cytoplasm into the organelle (Berridge et al. 2003). The storage of Ca 2+ in the ER is regulated by Ca 2+ -binding proteins that act as buffers. Stimulation of cells by agonists coupled to IP 3 production M. Suman : M. Giacomello : C. Ballarin : S. Montelli : B. Cozzi : A. Peruffo (*) Department of Comparative Biomedicine and Food Science, University of Padova, Viale dell’Università 16, 35020 Legnaro–Agripolis (PD), Italy e-mail: [email protected] L. Corain Department of Management and Engineering, University of Padova, Via S. Nicola 3, 36100 Vicenza (VI), Italy Present Address: M. Giacomello Venetian Institute of Molecular Medicine, Via Orus 2, 35129 Padova (PD), Italy Cell Tissue Res (2012) 350:109–118 DOI 10.1007/s00441-012-1460-2

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

REGULAR ARTICLE
Estradiol effects on intracellular Ca2+ homeostasis in bovinebrain-derived endothelial cells
Matteo Suman & Marta Giacomello & Livio Corain &
Cristina Ballarin & Stefano Montelli & Bruno Cozzi &Antonella Peruffo
Received: 24 October 2011 /Accepted: 31 May 2012 /Published online: 20 July 2012# Springer-Verlag 2012
Abstract Estrogens diversely affect various physiologicalprocesses by genomic or non-genomic mechanisms, in bothexcitable and non-excitable cells. Additional to the trophiceffects of estrogens promoting cell growth and differentiation,recent experimental evidence highlights their involvement inthe regulation of intracellular Ca2+ homeostasis. The effects ofestrogens on excitable cells are well documented. However,these steroids also influence numerous physiological events innon-excitable cells, such as fibroblasts or vascular endothelialcells. We have focused our attention on an immortalizedendothelial-like cell line derived from fetal bovine cerebellum.Estradiol (E2) effects on intracellular Ca2+ homeostasis weretested by varying the exposure time to the hormone(8, 24, 48 h). Calcium measurements were performedwith genetically encoded Ca2+ probes (Cameleons) targeted tothemain subcellular compartments involved in intracellular Ca2+ homeostasis (cytosol, endoplasmic reticulum, mitochondria).Mitochondrial Ca2+ uptake significantly decreased after
48-h exposure to E2, whereas cytosolic and endoplasmicreticulum responses were unaffected. The effect of E2
on mitochondrial Ca2+ handling was blocked by ICI 182,780,a pure estrogen receptor antagonist, suggesting that the effectwas estrogen-receptor–mediated. To evaluate whetherthe decrease of Ca2+ uptake affected mitochondrialmembrane potential (ΔΨm), cells were monitored inthe presence of tetra-methyl-rhodamine-methylester; nosignificant changes were seen between cells treated withE2 and controls. To investigate a mechanism of action,we assessed the possibile involvement of the permeabilitytransition pore (PTP), an inner mitochondrial membranechannel influencing energy metabolism and cell viability. Wetreated cells with CyclosporinA (CsA), which binds to thematrix chaperone cyclophilin-D and regulates PTP opening.CsA reversed the effects of a 48-h treatment with E2,suggesting a possible transcriptional modulation of proteinsinvolved in the mitochondrial permeability transition process.
Keywords Estradiol . Endothelial cells . Mitochondria .
Ca2+ homeostasis . Bovine
Introduction
The concentration of calcium [Ca2+] in a cell is managed byspecific intracellular organelles, such as the endoplasmicreticulum (ER) and mitochondria. In the lumen of ER, the[Ca2+] depends mainly on the balanced activity of theinositol triphosphate (IP3) receptor (IP3R) and of thesarco-endoplasmic reticulum Ca2+ ATPase (SERCA),which drives the ion from the cytoplasm into the organelle(Berridge et al. 2003). The storage of Ca2+ in the ER isregulated by Ca2+-binding proteins that act as buffers.Stimulation of cells by agonists coupled to IP3 production
M. Suman :M. Giacomello : C. Ballarin : S. Montelli : B. Cozzi :A. Peruffo (*)Department of Comparative Biomedicine and Food Science,University of Padova,Viale dell’Università 16,35020 Legnaro–Agripolis (PD), Italye-mail: [email protected]
L. CorainDepartment of Management and Engineering,University of Padova,Via S. Nicola 3,36100 Vicenza (VI), Italy
Present Address:M. GiacomelloVenetian Institute of Molecular Medicine,Via Orus 2,35129 Padova (PD), Italy
Cell Tissue Res (2012) 350:109–118DOI 10.1007/s00441-012-1460-2

causes Ca2+ release into the cytosol through the IP3R.Subsequently, the released Ca2+ can exit from the cellby the calcium extrusion systems of the plasma membrane,can be stored back into the ER, or can enter into themitochondrial matrix through the mitochondrial Ca2+
uniporter, whose molecular identity has recently beendefined (De Stefani et al. 2011). The Ca2+ is extrudedby mitochondria through cation exchangers, namely, theNa+/Ca2+ exchanger (NCX) and H+/Ca2+ exchanger andthrough the transient opening of the permeability transitionpore (PTP). The opening of this inner mitochondrialmembrane channel leads to the permeabilization of theinner membrane, a phenomenon known as mitochondrialpermeability transition (MPT). The MPT causes the release ofcations, the loss of mitochondrial transmembrane potentialand eventually outer membrane damage (Rasola et al. 2010).
Estradiol (E2) controls numerous cellular processes,including cell growth and differentiation, by modulatingCa2+ homeostasis (McCarthy 2008). E2 can influenceCa2+ handling by two different molecular mechanisms:a genomic action ("slow mechanism"), i.e., the bindingof E2 to either of the estrogen receptor isoforms resulting inthe regulation of gene transcription and a non-genomic action(“rapid mechanism”) generally initiated at the plasmamembrane by E2 and promoting the activation of signaltransduction pathways (Vasudevan and Pfaff 2008). Severalstudies have highlighted the capability of E2 to induce orrepress the expression of a variety of Ca2+ channels. Gu etal. (2001) have reported that E2 increases the expression oftwo subunits of the L-type voltage-dependent Ca2+
channels (L-VOCCs). Another study by Ritchie (2008)has demonstrated an increased density of L-VOCCsfollowing E2 treatment. To improve our understanding of theeffect of E2 on intracellular Ca2+ homeostasis, we have usedan in vitro model based on an immortalized endothelial cellline derived from fetal bovine cerebellum.
The effects of estrogens have been mainly investigated inexcitable cells. However, recent studies have demonstratedthat these steroids also act on non-excitable cells, includingendothelial cells and dermal fibroblasts (Arnal et al. 2010;Salazar-Colocho et al. 2008). Recent findings suggest thatthe endothelium and vascular smooth muscle are controlledby circulating steroid hormones and by steroids synthesizedlocally. Increasing evidence indicates that E2 exerts itsprotective effects through mitochondrial mechanisms incerebral endothelial cells (Guo et al. 2010). Other experimentaldata show that E2 has significant effects on cell migration inhuman skin fibroblasts (Stevenson et al. 2009).
To evaluate whether intracellular Ca2+ handling issignificantly affected by E2 treatment, we have performedexperiments on endothelial cells treated with variousconcentration of E2 (1, 10 and 100 nM) for variousincubation times (8, 24 and 48 h). We have monitored
Ca2+ oscillations in the main intracellular compartmentsinvolved in Ca2+ handling, namely, the cytosol, ER andmitochondria, by means of genetically encoded, specificallytargeted Ca2+ probes called Cameleons (Palmer et al. 2006).To confirm that the effect of E2 is mediated by estrogenreceptor isoforms, we have further tested its effect in thepresence of the estrogen receptor antagonist ICI 182,780.
We have also checked whether intracellular Ca2+
handling induces mitochondrial membrane potential(ΔΨm) variation. Such variation is critical for maintainingthe physiological function of the respiratory chain and forgenerating ATP and a significant loss of ΔΨm renders cellsdepleted of energy leading to their demise (Joshi andBakowska 2011). Therefore, in order to check whetherΔΨmis affected by E2 treatment, cells have been monitored in thepresence of tetra-methyl-rhodamine-methylester (TMRM).
Finally, to investigate a possible involvement of the PTP,an inner mitochondrial membrane channel (see above) thatinfluences energy metabolism and cell viability, cell cultureshave been treated with CyclosporinA (CsA), a specificinhibitor of the PTP (Rasola et al. 2010).
Materials and methods
Cell cultures
Primary cell cultures were derived from fetal bovinecerebellum. Male bovine fetuses at 4 months wereobtained at nearby commercial abattoirs upon accidentalslaughtering of pregnant cows. Animals were treatedaccording to the European Community Council Directive(86/609/EEC) concerning animal welfare during thecommercial slaughtering process and were constantlymonitored under mandatory official veterinary medicalcare. Age was determined by crown-rump length basedon commonly accepted reference tables (McGeady et al.2006). The use of bovine fetal cells is a reliable tool forstudying the molecular mechanisms of Ca2+ regulationand thus follows the joint declaration of the American,European and Japanese Societies for Neuroscience thatendorses replacement of laboratory animals whenever avaluable scientific alternative model is available.
Primary cell culture was performed following anestablished laboratory protocol (Peruffo et al. 2004).To obtain a stable cell line, cells were immortalizedfollowing the protocol by Takenouchi et al. (2007). Atday 2 in culture, cells were transfected with pSV3neoplasmid (LGC Promochem, Teddington, UK) by usingthe cationic lipid Lipofectamine 2000 (Invitrogen, Carlsbad,Calif., USA). Subsequently, a selection medium was addedand resistant cells were selected with the antibiotic G418(400 μg/ml; Gibco, Life Techologies BRL).
110 Cell Tissue Res (2012) 350:109–118

Cell line characterization
Staining analyses were performed following an establishedlaboratory protocol (Peruffo et al. 2008). The immortalized cellline was characterized immunocytochemically with antibodiesagainst neural and endothelial markers: anti-neurofilament-200(Sigma–Aldrich, dilution 1:40), anti-glial fibrillary acidicprotein (GFAP; DakoCytomation, Glostrup, Denmark,dilution 1:200), anti-integrin αM (Santa Cruz Biotechnology,Santa Cruz, Calif., USA, dilution 1:200), anti-vimentin(Sigma–Aldrich, dilution 1:100), anti-von Willebrand factor(vWF; Sigma–Aldrich, dilution 1:250) and anti-endothelialNO synthase (e-NOS; Abcam, dilution 1:250). Cells werethen incubated with secondary antibodies against IgG-tetramethylrhodamine-isothiocyanate (DakoCytomation,dilution 1:100) and IgG-fluorescein isothiocynate (SantaCruz Biotechnology, dilution 1:100). Immunostained cultureswere observed under a Leika TCS confocal microscope.Negative controls were performed by substituting primaryantibodies with bovine serum albumin in phosphate-bufferedsaline.
Fluorescence-imaging experiments
Endothelial-like cells were grown in DMEM-F12 containing10% fetal calf serum, L-glutamine (2 mM), penicillin (30 μg/ml), streptomycin (50 μg/ml) and G418 (400 μg/ml), at37 °C in a humidified atmosphere containing 5% CO2.Cells were transfected with genetically encoded Ca2+
probes (Cameleons) targeted to the main subcellularcompartments involved in intracellular Ca2+ homeostasis.The cDNA used for transfection coded for the Ca2+ probesD3cpv (cytosol), ERD1 (ER) and 4mtD3cpv (mitochondria;kindly provided by Prof. R.Y. Tsien, University of San Diego,USA; Palmer et al. 2004, 2006). Transfection with Cameleonswas performed at 70-80% cell confluence by using the Ca2+-phosphate technique in the presence of 2 μg DNA.
Experiments were carried out in an open-topped chamberthermostatically controlled at 37 °C on a fluorescencemicroscope system (Nikon TE2000-E stage) fitted withan immersion oil objective (CFl Plan Apochromat TIRF60X, N.A. 1.45). Cells were maintained in an extracellularphysiological solution containing 135 mM NaCl, 5 mM KCl,0.4 mM KH2PO4, 1 mM MgCl2, 5.5 mM glucose, 20 mMHEPES (pH 7.4 at 37°C) and were challenged either with ATPor with cyclopiazonic acid (CPA). FRET recordings wereobtained through a Beamsplitter (OES, Padua; emission filtersHQ 470/30M for CFP [cyano fluorescent protein] and HQ535/30M for yellow fluorescent protein [YFP]) and a dichroicmirror 505 LD. Images were collected by a back-thinnedcharge-coupled device camera (MicroMax 512 BFT,Princeton Instruments, USA) with a 300-ms exposuretime.
The ratio values (R) were obtained as follows: R ¼Fcpv � Fbgcpv
� �= Fcfp � Fbgcfp
� �, where Fcpv, Fcfp are the fluo-
rescence emission of the two fluorophores and Fbgcpv, Fbgcfprepresent background values for cpV (circularly permutedVenus) and CFP, respectively (Palmer et al. 2006). The datawere normalized to ΔR/R0 values, where R0 is the fluores-cence emission ratio at time 0 (i.e., the basal Ca2+ level beforeATP stimulation) and ΔR is the increase in fluorescenceemission ratio at any point. Analysis of the data was per-formed with NIS-elements AR software (Nikon Instuments,Japan).
To test the presence of ΔΨm variations, cells were bathedat room temperature in the dark with a physiological solutionsupplemented with 20 nM TMRM and 1 mM Ca2+. Tomeasure changes in ΔΨm, cells were stimulated with 5 μMtri-fluorocarbonyl-cyanide-phenylhydrazone (FCCP). Live-imaging experiments lasted 4 min and images were collectedevery 2 s with a 0.5-s exposure time. The differences betweenthe average basal level of TMRM fluorescence intensity value(i.e., before applying FCCP stimulation) and the averagefluorescence intensity values reached at plateau (i.e.,after FCCP addition) were calculated and corrected forbackground. Data were normalized to ΔR/R0 values,where R0 is the fluorescence intensity at time 0 (i.e.,the basal fluorescence intensity level) and ΔR is thedecrease in fluorescence intensity at any point. Analysisof these data was performed with HCImage software(Hamamatsu, Japan).
Exposure of immortalized cell line to E2
All experiments were performed 48 h after transfection withthe cDNA coding for Cameleons. In the cells transfectedwith Ca2+ probes D3cpv (cytosol) and ERD1 (ER), E2
(Sigma-Aldrich, Milan, Italy) was added to the culturemedium for 8, 24, or 48 h at a final concentration of10 nM. In the cells transfected with the Ca2+ probe4mtD3cpv (mitochondria), E2 was added to the culturemedium for 8 and 24 h at a final concentration of10 nM and for 48 h at a final concentration of 1, 10and 100 nM. To achieve 48-h cell exposure, E2 was added atthe time of transfection. For 24-h exposure, E2 was added 24 hafter transfection. Finally, to achieve 8-h exposure, E2 wasadded 8 h before the fluorescence-imaging experiment (40 hafter transfection).
In one set of experiments, cells were treated for 48 h withE2 (10 nM) and ICI 182,780 (10 nM; Sigma-Aldrich) beforethe fluorescence-imaging experiment. To test the presenceof any differences in ΔΨm between control and samplestreated with E2 (10 nM) for 48 h, cells were monitored in thepresence of the TMRM probe. Finally, in another series ofexperiments, cells treated for 48 h with E2 (10 nM) wereincubated for 30 min with CsA (Sigma-Aldrich) at a final
Cell Tissue Res (2012) 350:109–118 111
concentration of 0.8 μM before fluorescence-imagingexperiment.
Statistical analysis
All data are representative of at least seven independentexperiments. Values are expressed as means±SEM. Thestatistical analysis was conducted by using nonparametricpermutation-based one-way analysis of variance (ANOVA)and post-hoc pairwise comparisons (Basso et al. 2009;Pesarin and Salmaso 2010), implemented by the softwareNPC Test R10. Permutation tests were applied instead oftraditional parametric ANOVA and t-tests, because theyrepresent an exact inferential approach (valid for any finitesample size) and allow relaxation of the assumption of thenormality of response variables (or random errors).
Results
Cell line characterization
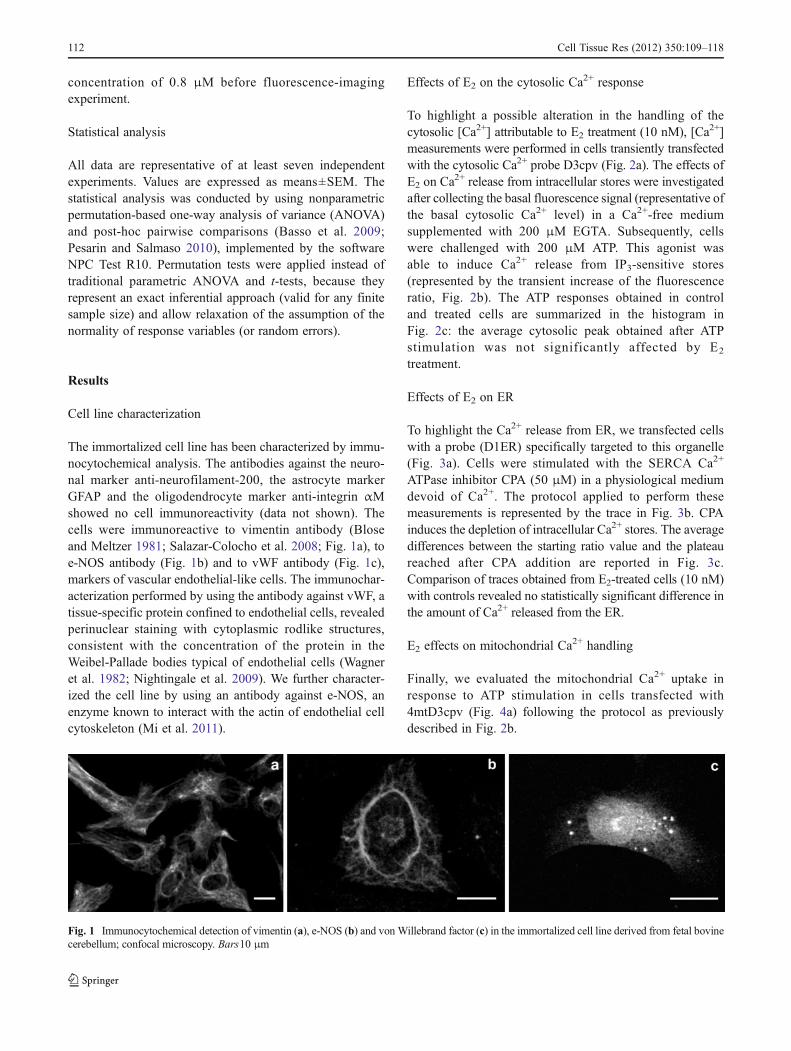
The immortalized cell line has been characterized by immu-nocytochemical analysis. The antibodies against the neuro-nal marker anti-neurofilament-200, the astrocyte markerGFAP and the oligodendrocyte marker anti-integrin αMshowed no cell immunoreactivity (data not shown). Thecells were immunoreactive to vimentin antibody (Bloseand Meltzer 1981; Salazar-Colocho et al. 2008; Fig. 1a), toe-NOS antibody (Fig. 1b) and to vWF antibody (Fig. 1c),markers of vascular endothelial-like cells. The immunochar-acterization performed by using the antibody against vWF, atissue-specific protein confined to endothelial cells, revealedperinuclear staining with cytoplasmic rodlike structures,consistent with the concentration of the protein in theWeibel-Pallade bodies typical of endothelial cells (Wagneret al. 1982; Nightingale et al. 2009). We further character-ized the cell line by using an antibody against e-NOS, anenzyme known to interact with the actin of endothelial cellcytoskeleton (Mi et al. 2011).
Effects of E2 on the cytosolic Ca2+ response
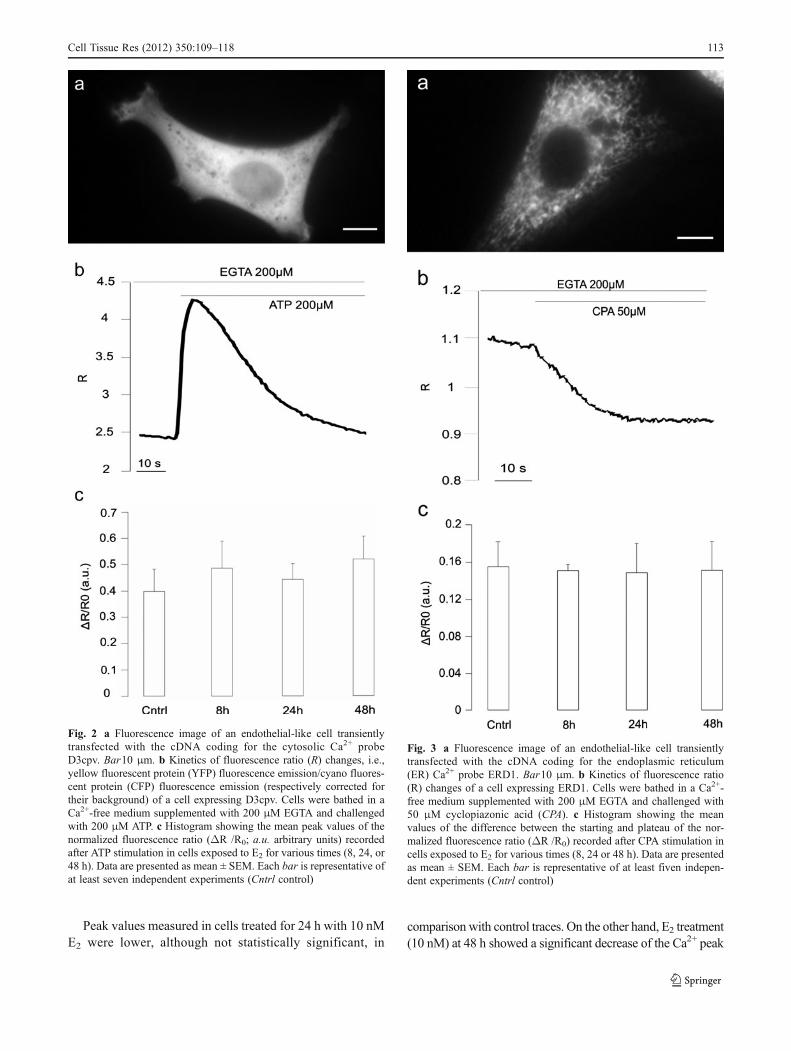
To highlight a possible alteration in the handling of thecytosolic [Ca2+] attributable to E2 treatment (10 nM), [Ca2+]measurements were performed in cells transiently transfectedwith the cytosolic Ca2+ probe D3cpv (Fig. 2a). The effects ofE2 on Ca
2+ release from intracellular stores were investigatedafter collecting the basal fluorescence signal (representative ofthe basal cytosolic Ca2+ level) in a Ca2+-free mediumsupplemented with 200 μM EGTA. Subsequently, cellswere challenged with 200 μM ATP. This agonist wasable to induce Ca2+ release from IP3-sensitive stores(represented by the transient increase of the fluorescenceratio, Fig. 2b). The ATP responses obtained in controland treated cells are summarized in the histogram inFig. 2c: the average cytosolic peak obtained after ATPstimulation was not significantly affected by E2
treatment.
Effects of E2 on ER
To highlight the Ca2+ release from ER, we transfected cellswith a probe (D1ER) specifically targeted to this organelle(Fig. 3a). Cells were stimulated with the SERCA Ca2+
ATPase inhibitor CPA (50 μM) in a physiological mediumdevoid of Ca2+. The protocol applied to perform thesemeasurements is represented by the trace in Fig. 3b. CPAinduces the depletion of intracellular Ca2+ stores. The averagedifferences between the starting ratio value and the plateaureached after CPA addition are reported in Fig. 3c.Comparison of traces obtained from E2-treated cells (10 nM)with controls revealed no statistically significant difference inthe amount of Ca2+ released from the ER.
E2 effects on mitochondrial Ca2+ handling
Finally, we evaluated the mitochondrial Ca2+ uptake inresponse to ATP stimulation in cells transfected with4mtD3cpv (Fig. 4a) following the protocol as previouslydescribed in Fig. 2b.
Fig. 1 Immunocytochemical detection of vimentin (a), e-NOS (b) and von Willebrand factor (c) in the immortalized cell line derived from fetal bovinecerebellum; confocal microscopy. Bars10 μm
112 Cell Tissue Res (2012) 350:109–118
Peak values measured in cells treated for 24 h with 10 nME2 were lower, although not statistically significant, in
comparison with control traces. On the other hand, E2 treatment(10 nM) at 48 h showed a significant decrease of the Ca2+ peak
Fig. 2 a Fluorescence image of an endothelial-like cell transientlytransfected with the cDNA coding for the cytosolic Ca2+ probeD3cpv. Bar10 μm. b Kinetics of fluorescence ratio (R) changes, i.e.,yellow fluorescent protein (YFP) fluorescence emission/cyano fluores-cent protein (CFP) fluorescence emission (respectively corrected fortheir background) of a cell expressing D3cpv. Cells were bathed in aCa2+-free medium supplemented with 200 μM EGTA and challengedwith 200 μM ATP. c Histogram showing the mean peak values of thenormalized fluorescence ratio (ΔR /R0; a.u. arbitrary units) recordedafter ATP stimulation in cells exposed to E2 for various times (8, 24, or48 h). Data are presented as mean ± SEM. Each bar is representative ofat least seven independent experiments (Cntrl control)
Fig. 3 a Fluorescence image of an endothelial-like cell transientlytransfected with the cDNA coding for the endoplasmic reticulum(ER) Ca2+ probe ERD1. Bar10 μm. b Kinetics of fluorescence ratio(R) changes of a cell expressing ERD1. Cells were bathed in a Ca2+-free medium supplemented with 200 μM EGTA and challenged with50 μM cyclopiazonic acid (CPA). c Histogram showing the meanvalues of the difference between the starting and plateau of the nor-malized fluorescence ratio (ΔR /R0) recorded after CPA stimulation incells exposed to E2 for various times (8, 24 or 48 h). Data are presentedas mean ± SEM. Each bar is representative of at least fiven indepen-dent experiments (Cntrl control)
Cell Tissue Res (2012) 350:109–118 113
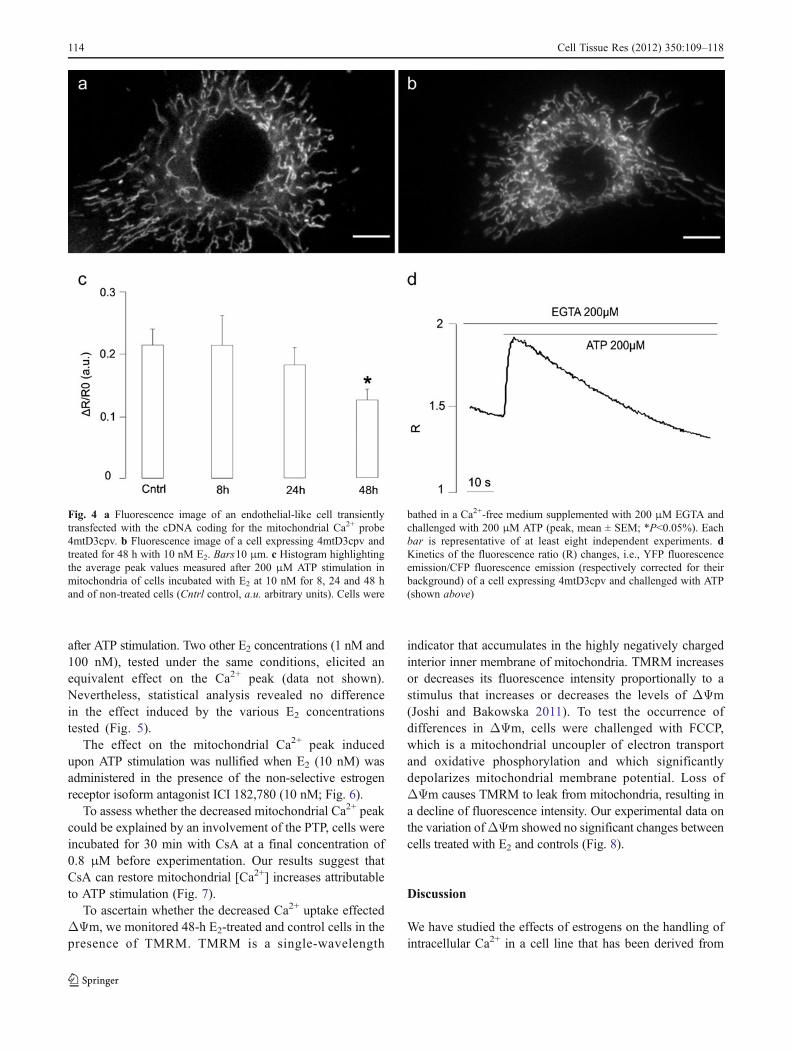
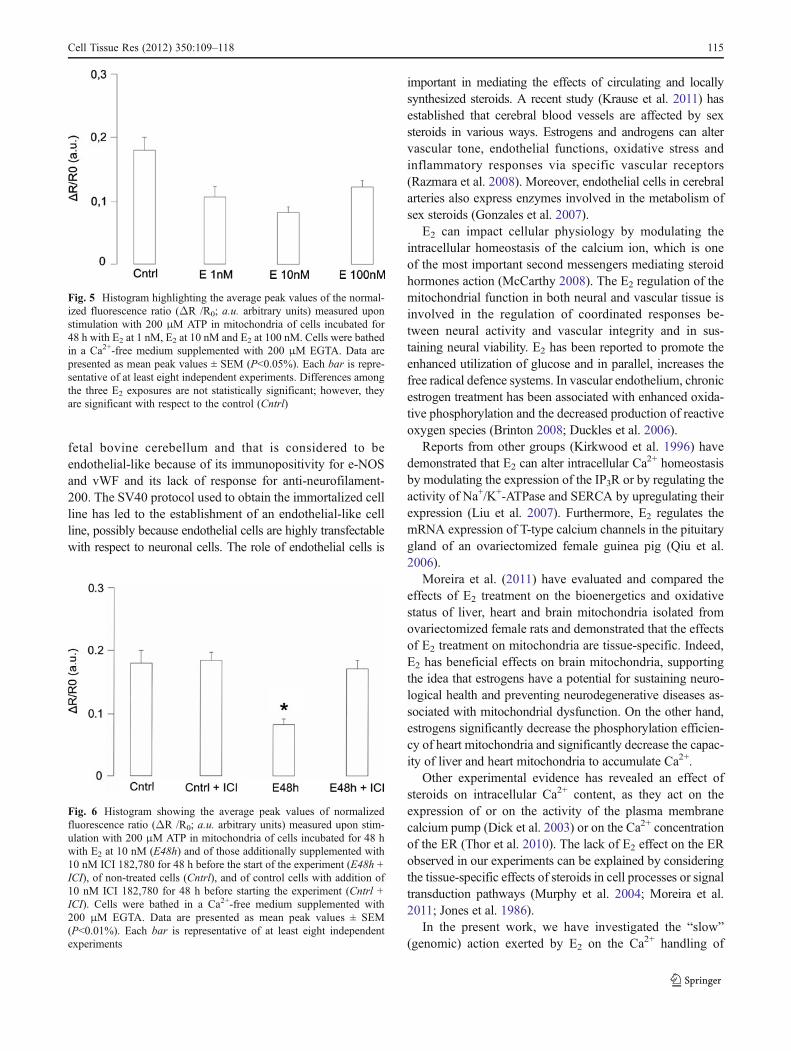
after ATP stimulation. Two other E2 concentrations (1 nM and100 nM), tested under the same conditions, elicited anequivalent effect on the Ca2+ peak (data not shown).Nevertheless, statistical analysis revealed no differencein the effect induced by the various E2 concentrationstested (Fig. 5).
The effect on the mitochondrial Ca2+ peak inducedupon ATP stimulation was nullified when E2 (10 nM) wasadministered in the presence of the non-selective estrogenreceptor isoform antagonist ICI 182,780 (10 nM; Fig. 6).
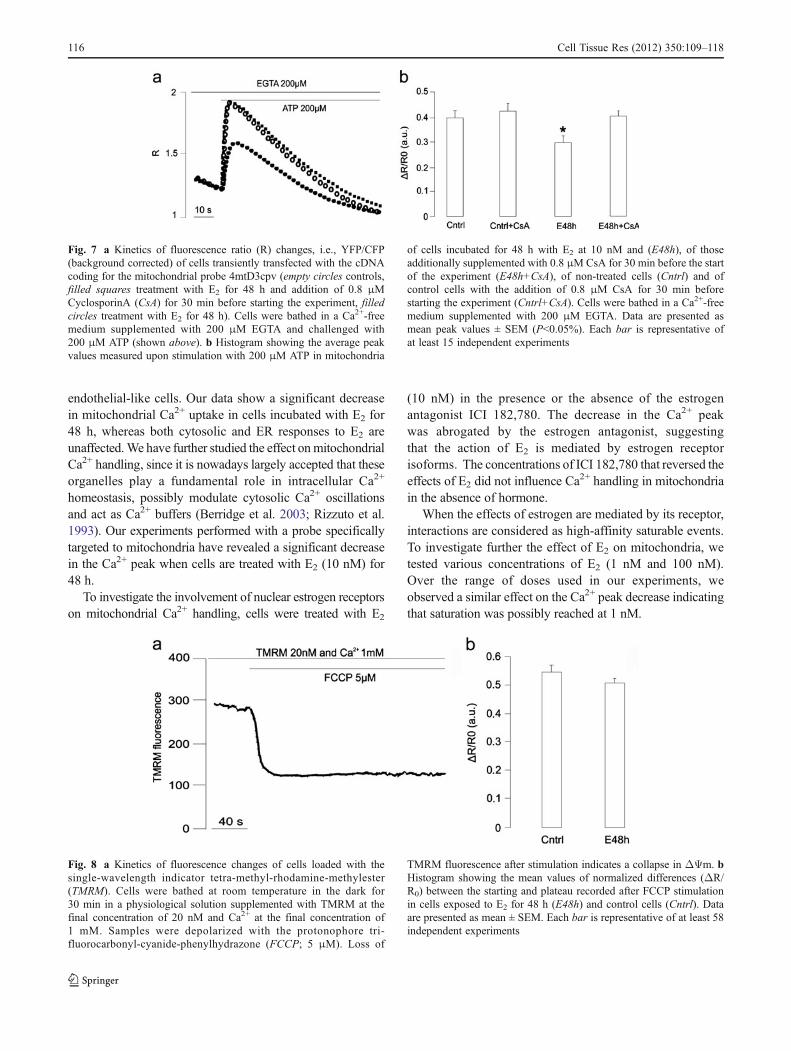
To assess whether the decreased mitochondrial Ca2+ peakcould be explained by an involvement of the PTP, cells wereincubated for 30 min with CsA at a final concentration of0.8 μM before experimentation. Our results suggest thatCsA can restore mitochondrial [Ca2+] increases attributableto ATP stimulation (Fig. 7).
To ascertain whether the decreased Ca2+ uptake effectedΔΨm, we monitored 48-h E2-treated and control cells in thepresence of TMRM. TMRM is a single-wavelength
indicator that accumulates in the highly negatively chargedinterior inner membrane of mitochondria. TMRM increasesor decreases its fluorescence intensity proportionally to astimulus that increases or decreases the levels of ΔΨm(Joshi and Bakowska 2011). To test the occurrence ofdifferences in ΔΨm, cells were challenged with FCCP,which is a mitochondrial uncoupler of electron transportand oxidative phosphorylation and which significantlydepolarizes mitochondrial membrane potential. Loss ofΔΨm causes TMRM to leak from mitochondria, resulting ina decline of fluorescence intensity. Our experimental data onthe variation ofΔΨm showed no significant changes betweencells treated with E2 and controls (Fig. 8).
Discussion
We have studied the effects of estrogens on the handling ofintracellular Ca2+ in a cell line that has been derived from
Fig. 4 a Fluorescence image of an endothelial-like cell transientlytransfected with the cDNA coding for the mitochondrial Ca2+ probe4mtD3cpv. b Fluorescence image of a cell expressing 4mtD3cpv andtreated for 48 h with 10 nM E2. Bars10 μm. c Histogram highlightingthe average peak values measured after 200 μM ATP stimulation inmitochondria of cells incubated with E2 at 10 nM for 8, 24 and 48 hand of non-treated cells (Cntrl control, a.u. arbitrary units). Cells were
bathed in a Ca2+-free medium supplemented with 200 μM EGTA andchallenged with 200 μM ATP (peak, mean ± SEM; *P<0.05%). Eachbar is representative of at least eight independent experiments. dKinetics of the fluorescence ratio (R) changes, i.e., YFP fluorescenceemission/CFP fluorescence emission (respectively corrected for theirbackground) of a cell expressing 4mtD3cpv and challenged with ATP(shown above)
114 Cell Tissue Res (2012) 350:109–118
fetal bovine cerebellum and that is considered to beendothelial-like because of its immunopositivity for e-NOSand vWF and its lack of response for anti-neurofilament-200. The SV40 protocol used to obtain the immortalized cellline has led to the establishment of an endothelial-like cellline, possibly because endothelial cells are highly transfectablewith respect to neuronal cells. The role of endothelial cells is
important in mediating the effects of circulating and locallysynthesized steroids. A recent study (Krause et al. 2011) hasestablished that cerebral blood vessels are affected by sexsteroids in various ways. Estrogens and androgens can altervascular tone, endothelial functions, oxidative stress andinflammatory responses via specific vascular receptors(Razmara et al. 2008). Moreover, endothelial cells in cerebralarteries also express enzymes involved in the metabolism ofsex steroids (Gonzales et al. 2007).
E2 can impact cellular physiology by modulating theintracellular homeostasis of the calcium ion, which is oneof the most important second messengers mediating steroidhormones action (McCarthy 2008). The E2 regulation of themitochondrial function in both neural and vascular tissue isinvolved in the regulation of coordinated responses be-tween neural activity and vascular integrity and in sus-taining neural viability. E2 has been reported to promote theenhanced utilization of glucose and in parallel, increases thefree radical defence systems. In vascular endothelium, chronicestrogen treatment has been associated with enhanced oxida-tive phosphorylation and the decreased production of reactiveoxygen species (Brinton 2008; Duckles et al. 2006).
Reports from other groups (Kirkwood et al. 1996) havedemonstrated that E2 can alter intracellular Ca2+ homeostasisby modulating the expression of the IP3R or by regulating theactivity of Na+/K+-ATPase and SERCA by upregulating theirexpression (Liu et al. 2007). Furthermore, E2 regulates themRNA expression of T-type calcium channels in the pituitarygland of an ovariectomized female guinea pig (Qiu et al.2006).
Moreira et al. (2011) have evaluated and compared theeffects of E2 treatment on the bioenergetics and oxidativestatus of liver, heart and brain mitochondria isolated fromovariectomized female rats and demonstrated that the effectsof E2 treatment on mitochondria are tissue-specific. Indeed,E2 has beneficial effects on brain mitochondria, supportingthe idea that estrogens have a potential for sustaining neuro-logical health and preventing neurodegenerative diseases as-sociated with mitochondrial dysfunction. On the other hand,estrogens significantly decrease the phosphorylation efficien-cy of heart mitochondria and significantly decrease the capac-ity of liver and heart mitochondria to accumulate Ca2+.
Other experimental evidence has revealed an effect ofsteroids on intracellular Ca2+ content, as they act on theexpression of or on the activity of the plasma membranecalcium pump (Dick et al. 2003) or on the Ca2+ concentrationof the ER (Thor et al. 2010). The lack of E2 effect on the ERobserved in our experiments can be explained by consideringthe tissue-specific effects of steroids in cell processes or signaltransduction pathways (Murphy et al. 2004; Moreira et al.2011; Jones et al. 1986).
In the present work, we have investigated the “slow”(genomic) action exerted by E2 on the Ca2+ handling of
Fig. 5 Histogram highlighting the average peak values of the normal-ized fluorescence ratio (ΔR /R0; a.u. arbitrary units) measured uponstimulation with 200 μM ATP in mitochondria of cells incubated for48 h with E2 at 1 nM, E2 at 10 nM and E2 at 100 nM. Cells were bathedin a Ca2+-free medium supplemented with 200 μM EGTA. Data arepresented as mean peak values ± SEM (P<0.05%). Each bar is repre-sentative of at least eight independent experiments. Differences amongthe three E2 exposures are not statistically significant; however, theyare significant with respect to the control (Cntrl)
Fig. 6 Histogram showing the average peak values of normalizedfluorescence ratio (ΔR /R0; a.u. arbitrary units) measured upon stim-ulation with 200 μM ATP in mitochondria of cells incubated for 48 hwith E2 at 10 nM (E48h) and of those additionally supplemented with10 nM ICI 182,780 for 48 h before the start of the experiment (E48h +ICI), of non-treated cells (Cntrl), and of control cells with addition of10 nM ICI 182,780 for 48 h before starting the experiment (Cntrl +ICI). Cells were bathed in a Ca2+-free medium supplemented with200 μM EGTA. Data are presented as mean peak values ± SEM(P<0.01%). Each bar is representative of at least eight independentexperiments
Cell Tissue Res (2012) 350:109–118 115
endothelial-like cells. Our data show a significant decreasein mitochondrial Ca2+ uptake in cells incubated with E2 for48 h, whereas both cytosolic and ER responses to E2 areunaffected.We have further studied the effect onmitochondrialCa2+ handling, since it is nowadays largely accepted that theseorganelles play a fundamental role in intracellular Ca2+
homeostasis, possibly modulate cytosolic Ca2+ oscillationsand act as Ca2+ buffers (Berridge et al. 2003; Rizzuto et al.1993). Our experiments performed with a probe specificallytargeted to mitochondria have revealed a significant decreasein the Ca2+ peak when cells are treated with E2 (10 nM) for48 h.
To investigate the involvement of nuclear estrogen receptorson mitochondrial Ca2+ handling, cells were treated with E2
(10 nM) in the presence or the absence of the estrogenantagonist ICI 182,780. The decrease in the Ca2+ peakwas abrogated by the estrogen antagonist, suggestingthat the action of E2 is mediated by estrogen receptorisoforms. The concentrations of ICI 182,780 that reversed theeffects of E2 did not influence Ca
2+ handling in mitochondriain the absence of hormone.
When the effects of estrogen are mediated by its receptor,interactions are considered as high-affinity saturable events.To investigate further the effect of E2 on mitochondria, wetested various concentrations of E2 (1 nM and 100 nM).Over the range of doses used in our experiments, weobserved a similar effect on the Ca2+ peak decrease indicatingthat saturation was possibly reached at 1 nM.
Fig. 7 a Kinetics of fluorescence ratio (R) changes, i.e., YFP/CFP(background corrected) of cells transiently transfected with the cDNAcoding for the mitochondrial probe 4mtD3cpv (empty circles controls,filled squares treatment with E2 for 48 h and addition of 0.8 μMCyclosporinA (CsA) for 30 min before starting the experiment, filledcircles treatment with E2 for 48 h). Cells were bathed in a Ca2+-freemedium supplemented with 200 μM EGTA and challenged with200 μM ATP (shown above). b Histogram showing the average peakvalues measured upon stimulation with 200 μM ATP in mitochondria
of cells incubated for 48 h with E2 at 10 nM and (E48h), of thoseadditionally supplemented with 0.8 μMCsA for 30 min before the startof the experiment (E48h+CsA), of non-treated cells (Cntrl) and ofcontrol cells with the addition of 0.8 μM CsA for 30 min beforestarting the experiment (Cntrl+CsA). Cells were bathed in a Ca2+-freemedium supplemented with 200 μM EGTA. Data are presented asmean peak values ± SEM (P<0.05%). Each bar is representative ofat least 15 independent experiments
Fig. 8 a Kinetics of fluorescence changes of cells loaded with thesingle-wavelength indicator tetra-methyl-rhodamine-methylester(TMRM). Cells were bathed at room temperature in the dark for30 min in a physiological solution supplemented with TMRM at thefinal concentration of 20 nM and Ca2+ at the final concentration of1 mM. Samples were depolarized with the protonophore tri-fluorocarbonyl-cyanide-phenylhydrazone (FCCP; 5 μM). Loss of
TMRM fluorescence after stimulation indicates a collapse in ΔΨm. bHistogram showing the mean values of normalized differences (ΔR/R0) between the starting and plateau recorded after FCCP stimulationin cells exposed to E2 for 48 h (E48h) and control cells (Cntrl). Dataare presented as mean ± SEM. Each bar is representative of at least 58independent experiments
116 Cell Tissue Res (2012) 350:109–118

Since the structure and distribution of mitochondria affecttheir function and their capability to handle intracellular Ca2+
(Rizzuto et al. 2009; Wang et al. 2003), we have consideredthe relationship between mitochondrial morphology or theirintracellular distribution and the variation of mitochondrialCa2+ uptake. Mitochondria are tubular structures largelyinterconnected inside the cell, they are located in closeproximity to the ER (Rizzuto et al. 2009) and theirmorphology is maintained through the action of manyproteins involved in the processes of mitochondrial fusion andfission (Dimmer and Scorrano 2006; Campello and Scorrano2010). The use of fluorescent Ca2+ probes targeted to themitochondria has allowed us to monitor both the morphologyand the distribution of mitochondria within the cell. Withinour experimental setting, E2 treatment causes no evidentrearrangement of the morphology and intracellular distributionof the mitochondria (see Fig. 4a, b). However, a more detailedimaging analysis is required to confirm this observation. Moroet al. (2010) have recently reported that estrogen deficiencyaffects the functional properties of mouse liver mitochondria,in particular by modulation of PTP.We have thus reasoned thatan alternative explanation for our results could be that of aninvolvement of PTP. Thus, we have used CsA, a specific PTPinhibitor, to verify this hypothesis. This molecule binds to thematrix chaperone cyclophilin D, a peptidyl-prolyl isomeraseknown to modulate the opening of the mitochondrial PTP(Rasola et al. 2010). In our experiments, CsA was able toreverse E2 effects suggesting a role of E2 in the modulationof the mitochondrial permeability transition process (MPTP).
As is well known,ΔΨm is crucial for maintaining cellularviability. Therefore, to improve our understanding of the waythat the decreased Ca2+ uptake, mediated by E2 through PTPmodulation, affects cell functions, we have measuredmitochondrial membrane potential with the TMRM probe.Our results reveal no statistically significantly changesbetween cells treated with E2 and controls (Fig. 8).Experimental evidence reported by Petronilli et al. (2001)demonstrates that, when the opening of PTP is transient, themitochondrial membrane potential is maintained. Indeed, datapresented by Dumas et al. (2009) show that PTP openingin intact cells may or may not lead to mitochondrialdepolarization, depending on the duration for which thePTP are open. Taken together, our data reveal that, in ourimmortalized endothelial-like cell line, the genomic effectexerted by E2 can affect PTP opening without causing acollapse in ΔΨm.
Our results are in agreement with the data obtained byMoreira et al. (2011) demonstrating a decreased capacity ofheart and liver mitochondria to accumulate Ca2+, because ofa higher susceptibility to PTP opening. Moreover, Moro etal. (2010) have found that estrogen deficiency inhibits theMPTP in isolated murine mitochondria and supplementationof mice with E2 restores this process.
In conclusion, our findings strongly suggest that E2 isinvolved in the modulation of PTP. Possibly, E2 genomicaction is associated with the expression of proteins thatmodulate PTP opening. E2 has been suggested to affect theexpression of numerous mitochondria-interacting proteins,regulators, or components of PTP, such as the antiapoptoticprotein Bcl-2 (Fan et al. 2008; Moro et al. 2010; Zhi et al.2007) and adenine nucleotide translocator (Too et al. 1999;Moro et al. 2010). Whether E2 modulation on PTP openinghas a bearing, in our experimental model, on the activationof pro-apoptotic pathways remains to be thoroughlyascertained. Further experiments are required to determinewhether and the manner in which the regulation of PTPopening by estrogens affects MPTP.
Acknowledgements We thank Dr. R.Y. Tsien for the cDNA of theCa2+ probes used to perfom the experiments described in this work. Wealso thank Dr. Maristella Giurisato and Dr. Michele Scorzeto (grantno. CPDR078721) for their scientific and technical assistance.
References
Arnal JF, Fontaine C, Billon-Galés A, Favre J, Laurell H, Lenfant F,Gourdy P (2010) Estrogen receptors and endothelium. ArteriosclerThromb Vasc Biol 30:1506–1512
Basso D, Pesarin F, Salmaso L, Solari A (2009) Permutation tests forstochastic ordering and ANOVA: theory and applications with R.Springer, Heidelberg
Berridge MJ, Bootman MD, Roderick HL (2003) Calcium signalling:dynamics, homeostasis and remodelling. Nat Rev Mol Cell Biol4:517–529
Blose SH, Meltzer DI (1981) Visualization of the 10-nm filamentvimentin rings in vascular endothelial cells in situ: close resem-blance to vimentin cytoskeletons found in monolayers in vitro.Exp Cell Res 135:299–309
Brinton RD (2008) Estrogen regulation of glucose metabolism andmitochondrial function: therapeutic implications for preventionof Alzheimer’s disease. Adv Drug Deliv Rev 60:1504–1511
Campello S, Scorrano L (2010) Mitochondrial shape changes:orchestrating cell pathophysiology. EMBO Rep 11:678–684
De Stefani D, Raffaello A, Teardo E, Szabò I, Rizzuto R (2011) Aforty-kilodalton protein of the inner membrane is the mitochon-drial calcium uniporter. Nature 476:336–340
Dick IM, Liu J, Glendenning P, Prince RL (2003) Estrogen andandrogen regulation of plasma membrane calcium pump activityin immortalized distal tubule kidney cells. Mol Cell Endocrinol212:11–18
Dimmer KS, Scorrano L (2006) (De)constructing mitochondria: whatfor? Physiology 21:233–241
Duckles SP, Krause DN, Stirone C, Procaccio V (2006) Estrogen andmitochondria: a new paradigm for vascular protection? Mol Interv6:26–35
Dumas JF, Argaud L, Cottet-Rousselle C, Vial G, Gonzalez C, DetailleD, Leverve X, Fontaine E (2009) Effect of transient and permanentpermeability transition pore opening on NAD(P)H localization inintact cells. J Biol Chem 284:15117–15125
Fan L, Pandey SC, Cohen RS (2008) Estrogen affects levels of Bcl-2protein and mRNA in medial amygdala of ovariectomized rats. JNeurosci Res 86:3655–3664
Cell Tissue Res (2012) 350:109–118 117

Gonzales RJ, Ansar S, Duckles SP, Krause DN (2007) Androgenic/estro-genic balance in the male rat cerebral circulation: metabolic enzymesand sex steroid receptors. J Cereb Blood Flow Metab 27:1841–1852
Gu Y, Preston MR, Magnay J, El Haj AJ, Publicover SJ (2001)Hormonally-regulated expression of voltage-operated Ca2+
channels in osteocytic (MLO-Y4) cells. Biochem BiophysRes Commun 282:536–542
Guo J, Krause DN, Horne J, Weiss JH, Li X, Duckles SP (2010)Estrogen-receptor-mediated protection of cerebral endothelial cellviability and mitochondrial function after ischemic insult in vitro.J Cereb Blood Flow Metab 3:545–554
Jones KJ, McEwen BS, Pfaff DW (1986) Regional specificity inestradiol effects on [3H]uridine incorporation in rat brain. MolCell Endocrinol 45:57–63
Joshi DC, Bakowska JC (2011) Determination of mitochondrialmembrane potential and reactive oxygen species in live ratcortical neurons. J Vis Exp 51:2704, doi:10.3791/2704
Kirkwood KL, Dziak R, Bradford PG (1996) Inositol trisphosphatereceptor gene expression and hormonal regulation in osteoblast-like cell lines and primary osteoblastic cell cultures. J Bone MinerRes 11:1889–1896
Krause DN, Duckles SP, Gonzales RJ (2011) Local oestrogenic/androgenicbalance in the cerebral vasculature. Acta Physiol 203:181–186
Liu CG, Xu KQ, Xu X, Huang JJ, Xiao JC, Zhang JP, Song HP (2007)17β-Oestradiol regulates the expression of Na+/K+-ATPase β1-subunit, sarcoplasmic reticulum Ca2+-ATPase and carbonic anhy-drase IV in H9C2 cells. Clin Exp Pharmacol Physiol 34:998–1004
McCarthy MM (2008) Estradiol and the developing brain. Physiol Rev88:91–134
McGeady TA, Quinn PJ, FitzPatrick ES, Ryan MT (2006) Veterinaryembryology. Blackwell, Oxford
Mi Q, Chen N, Shaifta Y, Xie L, LuH, Liu Z, Chen Q, Hamid C, Becker S,Ji Y, Ferro A (2011) Activation of endothelial nitric oxide synthase isdependent on its interaction with globular actin in human umbilicalvein endothelial cells. J Mol Cell Cardiol 51:419–427
Moreira PI, Custódio JB, Nunes E, Oliveira PJ, Moreno A, Seiça R,Oliveira CR, Santos MS (2011) Mitochondria from distinct tis-sues are differently affected by 17β-estradiol and tamoxifen. JSteroid Biochem Mol Biol 123:8–16
Moro L, Arbini AA, Hsieh JT, Ford J, Simpson ER, Hajibeigi A, OzOK (2010) Aromatase deficiency inhibits the permeability transi-tion in mouse liver mitochondria. Endocrinology 151:1643–1652
Murphy HS, Sun Q, Murphy BA, Mo R, Huo J, Chen J, Chensue SW,Adams M, Richardson BC, Yung R (2004) Tissue-specific effectof estradiol on endothelial cell-dependent lymphocyte recruit-ment. Microvasc Res 68:273–285
Nightingale TD, Pattni K, Hume AN, Seabra MC, Cutler DF (2009)Rab27a and MyRIP regulate the amount and multimeric state ofvWF released from endothelial cells. Blood 113:5010–5018
Palmer A, Jin C, Reed JC, Tsien RY (2004) Bcl2-mediated alterationsin endoplasmic reticulum Ca2+ analyzed with an improved genet-ically encoded fluorescent sensor. Proc Natl Acad Sci USA101:17404–17409
Palmer AE, Giacomello M, Kortemme T, Hires SA, Lev-Ram V, BakerD, Tsien RY (2006) Ca2+ indicators based on computationallyredesigned calmodulin-peptide pairs. Chem Biol 13:521–530
Peruffo A, Massimino ML, Ballarin C, Carmignoto G, Rota A, Cozzi B(2004) Primary cultures from fetal bovine brain. Neuroreport15:1719–1722
Peruffo A, Buson G, Cozzi B, Ballarin C (2008) Primary cell culturesfrom fetal bovine hypothalamus and cerebral cortex: a reliablemodel to study P450Arom and alpha and beta estrogen receptorsin vitro. Neurosci Lett 434:83–87
Pesarin F, Salmaso L (2010) Permutation tests for complex data—theory and software. Wiley, Chichester
Petronilli V, Penzo D, Scorrano L, Bernardi P, Di Lisa F (2001) Themitochondrial permeability transition, release of cytochrome cand cell death. Correlation with the duration of pore openings insitu. J Biol Chem 276:12030–12034
Qiu J, Bosch MA, Jamali K, Xue C, Kelly MJ, Rønnekleiv OK (2006)Estrogen upregulates T-type calcium channels in the hypothalamusand pituitary. J Neurosci 43:11072–11082
Rasola A, Sciacovelli M, Pantic B, Bernardi P (2010) Signal transduc-tion to the permeability transition pore. FEBS Lett 584:1989–1996
Razmara A, Sunday L, Stirone C, Wang XB, Krause DN, Duckles SP,Procaccio V (2008) Mitochondrial effects of estrogen are mediatedby estrogen receptor alpha in brain endothelial cells. J PharmacolExp Ther 325:782–790
Ritchie AK (2008) Estrogen increases low voltage-activated calciumcurrent density in GH3 anterior pituitary cells. Endocrinology132:1621–1629
Rizzuto R, Brini M, Murgia M, Pozzan T (1993) Microdomains withhigh Ca2+ close to IP3-sensitive channels that are sensed byneighboring mitochondria. Science 262:744–747
Rizzuto R, Marchi S, Bonora M, Aguiari P, Bononi A, De Stefani D,Giorgi C, Leo S, Rimessi A, Siviero R, Zecchini E, Pinton P(2009) Ca(2+) transfer from the ER to mitochondria: when, howand why. Biochim Biophys Acta 1787:1342–1351
Salazar-Colocho P, Del Río J, Frechilla D (2008) Involvement of thevascular wall in regenerative processes after CA1 ischemicneuronal death. Int J Dev Neurosci 26:541–550
Stevenson S, Sharpe TD, Thronton J (2009) Effects of oestrogenagonists on human dermal fibroblasts in an in vitro woundingassay. Exp Dermatol 18:988–990
Takenouchi T, Iwamaru Y, Sato M, Yokoyama T, Shinagawa M, KitaniH (2007) Establishment and characterization of SV40 large Tantigen-immortalized cell lines derived from fetal bovine braintissues after prolonged cryopreservation. Cell Biol Int 31:57–64
Thor D, Uchizono JA, Lin-Cereghino GP, Rahimian R (2010) Theeffect of 17 beta-estradiol on intracellular calcium homeostasis inhuman endothelial cells. Eur J Pharmacol 630:92–99
Too CK, Giles A, Wilkinson M (1999) Estrogen stimulates expressionof adeninenucleotide translocator ANT1 messenger RNA infemale rat hearts. Mol Cell Endocrinol 150:161–167
Vasudevan N, Pfaff DW (2008) Non-genomic actions of estrogens andtheir interaction with genomic actions in the brain. Front Neuro-endocrinol 29:238–257
Wagner DD, Olmsted JB, Marder VJ (1982) Immunolocalization ofvon Willebrand protein in Weibel-Palade bodies of humanendothelial cells. J Cell Biol 95:355–360
Wang GJ, Jackson JG, Thayer SA (2003) Altered distribution ofmitochondria impairs calcium homeostasis in rat hippocampalneurons in culture. J Neurochem 87:85–94
Zhi X, Honda K, Sumi T, Yasui T, Nobeyama H, Yoshida H, Ishiko O(2007) Estradiol-17beta regulates vascular endothelial growthfactor and Bcl-2 expression in HHUA cells. Int J Oncol31:1333–1338
118 Cell Tissue Res (2012) 350:109–118
Related Documents











