Estradiol decreases the orexigenic effect of melanin-concentrating hormone in ovariectomized rats Michelina M. Messina a,b , Gretha Boersma a,b , J. Michael Overton a,c , Lisa A. Eckel a,b, ⁎ a Program in Neuroscience, Florida State University, Tallahassee, FL, USA b Department of Psychology, Florida State University, Tallahassee, FL 32306-1270, USA c Biomedical Sciences, Florida State University, Tallahassee, FL, USA Received 17 November 2005; received in revised form 28 April 2006; accepted 1 May 2006 Abstract Estradiol exerts an inhibitory effect on food intake via interactions with anorexigenic peptides, like cholecystokinin, that function to decrease meal size. It is currently unknown whether estradiol also interacts with orexigenic compounds implicated in the physiological control of food intake. Thus, the primary goal of this study was to determine whether estradiol decreases the orexigenic effect of melanin-concentrating hormone (MCH), a neuropeptide that, like estradiol, appears to influence food intake by selectively affecting the controls of meal size. Food and water intake were monitored following lateral ventricular (icv) infusions of 5 μg MCH or saline vehicle in oil- and estradiol-treated ovariectomized rats. MCH increased food intake throughout the first 4 h of the dark phase in oil-treated rats, but only for the last 2 h of the same 4-h interval in estradiol-treated rats. As a result, the orexigenic effect of MCH was significantly lower in estradiol-treated rats, relative to oil-treated rats. During this interval of MCH-stimulated feeding, a prandial increase in water intake was not observed in either oil- or estradiol-treated rats. We conclude that estradiol decreases the orexigenic effect of MCH in ovariectomized rats. © 2006 Elsevier Inc. All rights reserved. Keywords: Estrogen; Meal size; Neuropeptide; Water intake; Food intake 1. Introduction The ovarian hormone estradiol is involved in the physiological control of food intake in female rats. For example, ovariectomy induces hyperphagia [20,36]; however, this behavioral response is abolished by estradiol treatment alone [1]. In addition, the preovulatory rise in estradiol secretion is associated with a decrease in food intake during estrus in ovarian-intact, cycling rats [8,33]. Together, these studies demonstrate that estradiol exerts an in- hibitory effect on food intake. Because changes in food intake are mediated by changes in meal size and/or meal number [32], the feeding patterns of female rats have been examined to better understand the mechanism underlying the anorexigenic effect of estradiol. Such analyses reveal that estradiol inhibits food intake by decreasing meal size, not meal number (reviewed in [6]). Rather than acting alone to decrease meal size, estradiol interacts with other anorexigenic compounds that function to decrease meal size. For example, the inhibitory effect of cholecystokinin on meal size is increased by estradiol treatment in ovariectomized rats [3,12,37], and by the preovulatory rise in estradiol secretion in cycling rats [7]. At present, similar studies are necessary to investigate whether estradiol also interacts with orexigenic compounds, particularly those that function to increase meal size. One putative candidate is melanin-concentrating hormone (MCH), an orexigenic peptide synthesized primarily in the lateral hypothalamus (LH) and zona incerta (ZI) [31]. In rodents, acute intracerebroventricular (icv) administration of MCH increases food intake [4,5,25,27,35], and chronic icv administration of MCH or an MCH-1 receptor agonist promotes hyperphagia, weight gain, and lipogenesis ([5,28], but see [27]). In addition, MCH deficiency in mice, induced by targeted deletion of the gene that encodes prepro-MCH, is associated with hypophagia, reduced body adiposity, and increased energy expenditure [16,29]. Finally, Physiology & Behavior 88 (2006) 523 – 528 ⁎ Corresponding author. Department of Psychology, Florida State University, Tallahassee, FL 32306-1270, USA. Tel.: +1 850 644 3480; fax: +1 850 644 7739. E-mail address: [email protected] (L.A. Eckel). 0031-9384/$ - see front matter © 2006 Elsevier Inc. All rights reserved. doi:10.1016/j.physbeh.2006.05.002

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

8 (2006) 523–528
Physiology & Behavior 8Estradiol decreases the orexigenic effect of melanin-concentratinghormone in ovariectomized rats
Michelina M. Messina a,b, Gretha Boersma a,b, J. Michael Overton a,c, Lisa A. Eckel a,b,⁎
a Program in Neuroscience, Florida State University, Tallahassee, FL, USAb Department of Psychology, Florida State University, Tallahassee, FL 32306-1270, USA
c Biomedical Sciences, Florida State University, Tallahassee, FL, USA
Received 17 November 2005; received in revised form 28 April 2006; accepted 1 May 2006
Abstract
Estradiol exerts an inhibitory effect on food intake via interactions with anorexigenic peptides, like cholecystokinin, that function to decreasemeal size. It is currently unknown whether estradiol also interacts with orexigenic compounds implicated in the physiological control of foodintake. Thus, the primary goal of this study was to determine whether estradiol decreases the orexigenic effect of melanin-concentrating hormone(MCH), a neuropeptide that, like estradiol, appears to influence food intake by selectively affecting the controls of meal size. Food and waterintake were monitored following lateral ventricular (icv) infusions of 5 μg MCH or saline vehicle in oil- and estradiol-treated ovariectomized rats.MCH increased food intake throughout the first 4 h of the dark phase in oil-treated rats, but only for the last 2 h of the same 4-h interval inestradiol-treated rats. As a result, the orexigenic effect of MCH was significantly lower in estradiol-treated rats, relative to oil-treated rats. Duringthis interval of MCH-stimulated feeding, a prandial increase in water intake was not observed in either oil- or estradiol-treated rats. We concludethat estradiol decreases the orexigenic effect of MCH in ovariectomized rats.© 2006 Elsevier Inc. All rights reserved.
Keywords: Estrogen; Meal size; Neuropeptide; Water intake; Food intake
1. Introduction
The ovarian hormone estradiol is involved in the physiologicalcontrol of food intake in female rats. For example, ovariectomyinduces hyperphagia [20,36]; however, this behavioral response isabolished by estradiol treatment alone [1]. In addition, thepreovulatory rise in estradiol secretion is associated with a decreasein food intake during estrus in ovarian-intact, cycling rats [8,33].Together, these studies demonstrate that estradiol exerts an in-hibitory effect on food intake. Because changes in food intake aremediated by changes in meal size and/or meal number [32], thefeeding patterns of female rats have been examined to betterunderstand the mechanism underlying the anorexigenic effect ofestradiol. Such analyses reveal that estradiol inhibits food intake by
⁎ Corresponding author. Department of Psychology, Florida State University,Tallahassee, FL 32306-1270, USA. Tel.: +1 850 644 3480; fax: +1 850 644 7739.
E-mail address: [email protected] (L.A. Eckel).
0031-9384/$ - see front matter © 2006 Elsevier Inc. All rights reserved.doi:10.1016/j.physbeh.2006.05.002
decreasing meal size, not meal number (reviewed in [6]). Ratherthan acting alone to decrease meal size, estradiol interacts withother anorexigenic compounds that function to decrease meal size.For example, the inhibitory effect of cholecystokinin onmeal size isincreased by estradiol treatment in ovariectomized rats [3,12,37],and by the preovulatory rise in estradiol secretion in cycling rats [7].At present, similar studies are necessary to investigate whetherestradiol also interacts with orexigenic compounds, particularlythose that function to increase meal size.
One putative candidate is melanin-concentrating hormone(MCH), an orexigenic peptide synthesized primarily in the lateralhypothalamus (LH) and zona incerta (ZI) [31]. In rodents, acuteintracerebroventricular (icv) administration of MCH increasesfood intake [4,5,25,27,35], and chronic icv administration ofMCH or an MCH-1 receptor agonist promotes hyperphagia,weight gain, and lipogenesis ([5,28], but see [27]). In addition,MCH deficiency in mice, induced by targeted deletion of the genethat encodes prepro-MCH, is associatedwith hypophagia, reducedbody adiposity, and increased energy expenditure [16,29]. Finally,

524 M.M. Messina et al. / Physiology & Behavior 88 (2006) 523–528
transgenic mice that overexpress prepro-MCH display hyperpha-gia when fed a high fat diet, and develop obesity and insulinresistance when fed chow diets [19]. Currently, indirect evidencesuggests that MCH, like estradiol, influences food intake byselectively affecting the controls of meal size. A detailed analysisof the feeding patterns of diet-induced obese rats revealed that thehypophagia induced by T-226296, a potent and selective MCH-1receptor antagonist, is mediated by a selective decrease in mealsize, not meal number [17]. In as far as this action of an MCH-1receptor antagonist reflects the effects of endogenous MCH, thisfinding suggests that MCH influences feeding by selectivelyaffecting the controls of meal size.
Emerging evidence suggests that MCH gene expression is re-gulated by an estrogen-sensitive mechanism. For example, a phy-siological regimen of estradiol treatment decreased prepro-MCHmRNA expression in the medial ZI of ovariectomized rats [22]. Inaddition, the increase in lateral hypothalamic MCHmRNA expre-ssion induced by negative energy balance is blocked in hypere-strogenemic male rats implanted with either estrogen-secretingleydig tumor cells or pellets containing pharmacological concen-trations of estradiol [21,23]. Because estrogen receptors (ERs) areexpressed at high levels within the LH and ZI [30], estradiol mayact locally within these brain regions to inhibit the activity ofMCHneurons. In addition, estradiol may also regulate MCH signaling,given that ERs are expressed within many of the brain regionscontainingMCH-1 receptors [14,30]. Taken together, these studiesraise the possibility that the anorexigenic effect of estradiol mayinvolve a selective inhibition of MCH activity. To investigate thishypothesis, we examined whether estradiol decreases MCH-in-duced feeding in ovariectomized rats.
2. Methods
2.1. Animals and housing
Sixteen female Long-Evans rats (Charles River BreedingLaboratory, Raleigh, NC), weighing between 240 and 270 g atstudy onset, were housed individually in custom-designed Ple-xiglas cages equipped with feeding niches that provided access topowdered chow (Purina 5001). Food and tap water were freelyavailable, except as otherwise noted. The roomwas maintained at20±2 °C with a 12:12 light/dark cycle (dark onset=1300 h). Awhite noise generator was run at moderate intensity to mask ex-traneous noise. Animal usage and all procedures were in strictcompliancewith the guidelines of theNational Institutes ofHealthand approved by the Florida State University Institutional AnimalCare and Use Committee.
2.2. Surgery
Rats were anesthetized with intraperitoneal injections of amixture of ketamine (50 mg/ml; Ketaset, Fort Dodge, IA) andxylazine (4.5 mg/ml; Rompun, Mobay, Shawnee, KS) and thenbilaterally ovariectomized using an intra-abdominal approach.Immediately following this procedure, rats were implanted with26-gauge stainless steel guide cannulas that targeted the rightlateral ventricle. Stereotaxic coordinates (RC=−0.3 mm, LM=
−1.1 mm, DV=−3.5 mm from bregma) were based on the ratatlas of Paxinos and Watson [24]. After 7 days of post-operativerecovery, cannula placement was verified by monitoring the rats'water intake for 20 min following icv infusions of 50 ng ofangiotensin II (Sigma-Aldrich, St. Louis,MO) delivered in 5 μl ofsaline vehicle over 1 min. Only those rats that consumed at least5 ml of water in 20 min were included in the subsequent studies.
2.3. Experiment 1: dark-phase food and water intake
The goal of this experiment was to examine the effects of MCHon dark-phase food and water intake in ovariectomized rats withand without estradiol treatment. We choose to examine waterintake, in addition to food intake, because MCH has been reportedto exert a dipsogenic effect in male rats [4]. Using a within-subjectsdesign, ovariectomized rats (n=10) received 2 cycles of 4 μg 17-β-estradiol-3-benzoate (estradiol; Sigma-Aldrich, St. Louis, MO) in0.1 ml sesame oil vehicle (Sigma-Aldrich, St. Louis, MO), and2 cycles of 0.1 ml oil alone. Intrascapular subcutaneous (sc)injections of estradiol or oil were administered on two consecutivedays (i.e., days 1 and 2) of each replacement cycle. This regimen ofestradiol treatment was chosen because it mimics the changes inestradiol secretion observed across the 4-day estrous cycle ofovarian-intact rats [1], and it decreases food intake on day 4, the daythat models estrus [1]. Accordingly, the effects of MCH or vehicletreatment on food and water intake were examined on day 4. Onthese test days, food and water were removed from the rats' cages30min prior to dark onset. Rats then received icv infusions of either5 μgMCH (BachemBioscience Inc., King of Prussia, PA) in 2.5 μlsaline vehicle or 2.5 μl vehicle alone at a rate of 2.5 μl/min. Thedose ofMCHwas chosen because it produces a reliable increase infood intake in male rats [4,27,35]. Food and water were returned tothe rats' cages at dark onset and then food and water intake weremonitored hourly for 4 h post-infusion and again at 20 h post-infusion. Following each feeding/drinking test, rats were given aminimum of 4 days prior to beginning the next cycle of estradiol oroil treatment. Hormone replacement (estradiol and oil) and drugtreatment (MCH and saline) were counterbalanced across treatmentcycles.
2.4. Experiment 2: light-phase water intake
In Experiment 2, we examined whether MCH stimulates anincrease in water intake, independent of food intake, similar tothat observed previously in male rats [4]. Using a within-sub-jects design, the effects of MCH and saline vehicle on light-phase water intake was assessed in ovariectomized rats (n=6)that received 2 cycles of oil treatment, as described in Ex-periment 1. We did not include an estradiol-treated conditionbased on our findings in Experiment 1, which revealed thatestradiol decreased the orexigenic effect of MCH and could,therefore, exert a similar effect on water intake. On test days(day 4), food and water were removed from the rats' cages 6 hprior to dark onset. One hour later, rats received infusions ofeither 5 μg MCH in 2.5 μl saline vehicle or 2.5 μl vehicle alone.Beginning 30 min following MCH or saline infusions, water,but not food, was returned to the rats' cages. Light-phase water
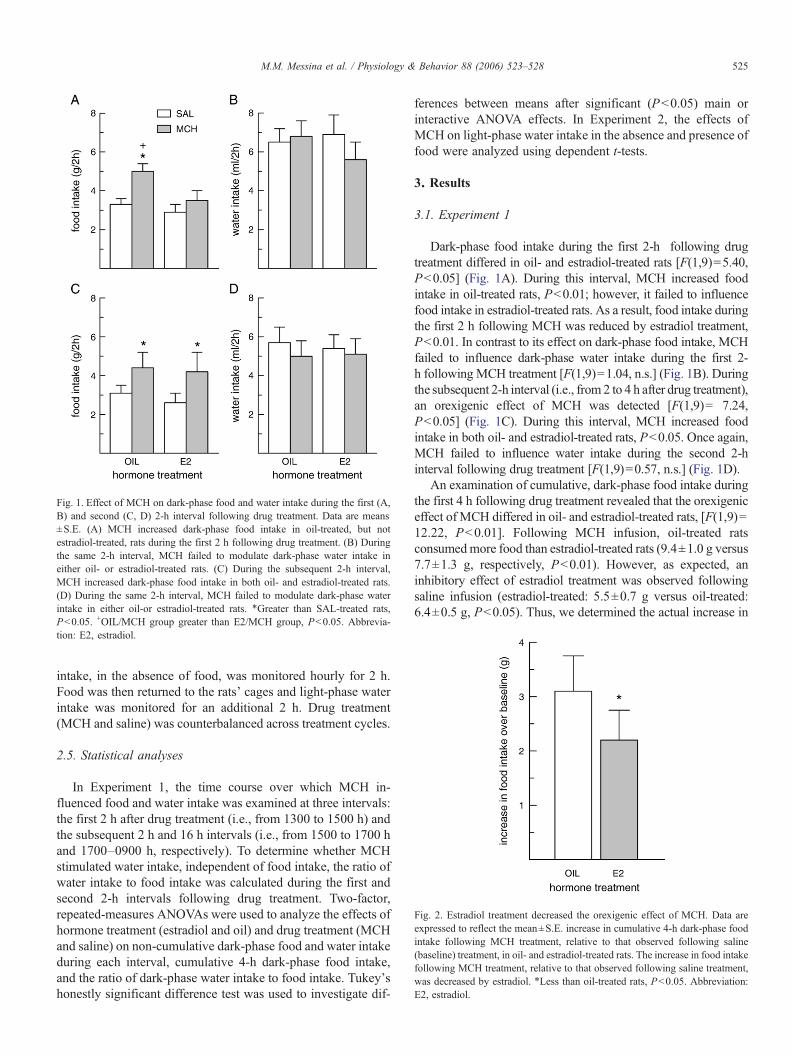
Fig. 1. Effect of MCH on dark-phase food and water intake during the first (A,B) and second (C, D) 2-h interval following drug treatment. Data are means±S.E. (A) MCH increased dark-phase food intake in oil-treated, but notestradiol-treated, rats during the first 2 h following drug treatment. (B) Duringthe same 2-h interval, MCH failed to modulate dark-phase water intake ineither oil- or estradiol-treated rats. (C) During the subsequent 2-h interval,MCH increased dark-phase food intake in both oil- and estradiol-treated rats.(D) During the same 2-h interval, MCH failed to modulate dark-phase waterintake in either oil-or estradiol-treated rats. ⁎Greater than SAL-treated rats,Pb0.05. +OIL/MCH group greater than E2/MCH group, Pb0.05. Abbrevia-tion: E2, estradiol.
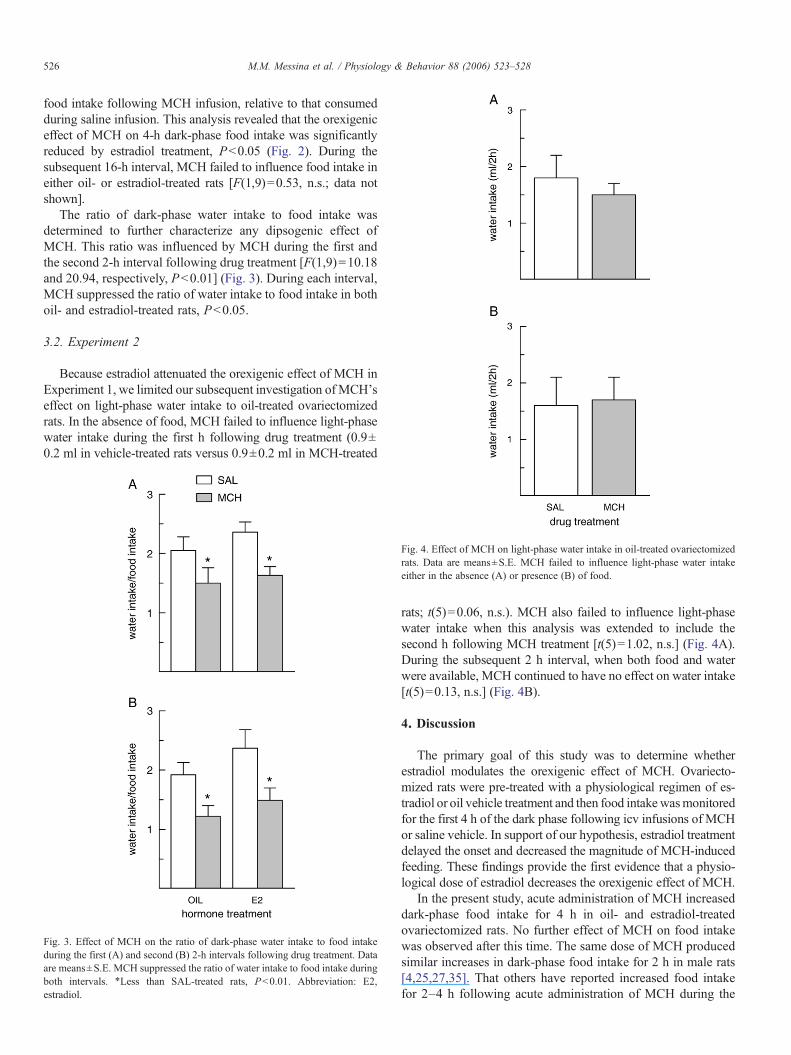
Fig. 2. Estradiol treatment decreased the orexigenic effect of MCH. Data areexpressed to reflect the mean±S.E. increase in cumulative 4-h dark-phase foodintake following MCH treatment, relative to that observed following saline(baseline) treatment, in oil- and estradiol-treated rats. The increase in food intakefollowing MCH treatment, relative to that observed following saline treatment,was decreased by estradiol. ⁎Less than oil-treated rats, Pb0.05. Abbreviation:E2, estradiol.
525M.M. Messina et al. / Physiology & Behavior 88 (2006) 523–528
intake, in the absence of food, was monitored hourly for 2 h.Food was then returned to the rats' cages and light-phase waterintake was monitored for an additional 2 h. Drug treatment(MCH and saline) was counterbalanced across treatment cycles.
2.5. Statistical analyses
In Experiment 1, the time course over which MCH in-fluenced food and water intake was examined at three intervals:the first 2 h after drug treatment (i.e., from 1300 to 1500 h) andthe subsequent 2 h and 16 h intervals (i.e., from 1500 to 1700 hand 1700–0900 h, respectively). To determine whether MCHstimulated water intake, independent of food intake, the ratio ofwater intake to food intake was calculated during the first andsecond 2-h intervals following drug treatment. Two-factor,repeated-measures ANOVAs were used to analyze the effects ofhormone treatment (estradiol and oil) and drug treatment (MCHand saline) on non-cumulative dark-phase food and water intakeduring each interval, cumulative 4-h dark-phase food intake,and the ratio of dark-phase water intake to food intake. Tukey'shonestly significant difference test was used to investigate dif-
ferences between means after significant (Pb0.05) main orinteractive ANOVA effects. In Experiment 2, the effects ofMCH on light-phase water intake in the absence and presence offood were analyzed using dependent t-tests.
3. Results
3.1. Experiment 1
Dark-phase food intake during the first 2-h following drugtreatment differed in oil- and estradiol-treated rats [F(1,9)=5.40,Pb0.05] (Fig. 1A). During this interval, MCH increased foodintake in oil-treated rats, Pb0.01; however, it failed to influencefood intake in estradiol-treated rats. As a result, food intake duringthe first 2 h following MCH was reduced by estradiol treatment,Pb0.01. In contrast to its effect on dark-phase food intake, MCHfailed to influence dark-phase water intake during the first 2-h following MCH treatment [F(1,9)=1.04, n.s.] (Fig. 1B). Duringthe subsequent 2-h interval (i.e., from 2 to 4 h after drug treatment),an orexigenic effect of MCH was detected [F(1,9)= 7.24,Pb0.05] (Fig. 1C). During this interval, MCH increased foodintake in both oil- and estradiol-treated rats, Pb0.05. Once again,MCH failed to influence water intake during the second 2-hinterval following drug treatment [F(1,9)=0.57, n.s.] (Fig. 1D).
An examination of cumulative, dark-phase food intake duringthe first 4 h following drug treatment revealed that the orexigeniceffect of MCH differed in oil- and estradiol-treated rats, [F(1,9)=12.22, Pb0.01]. Following MCH infusion, oil-treated ratsconsumedmore food than estradiol-treated rats (9.4±1.0 g versus7.7±1.3 g, respectively, Pb0.01). However, as expected, aninhibitory effect of estradiol treatment was observed followingsaline infusion (estradiol-treated: 5.5±0.7 g versus oil-treated:6.4±0.5 g, Pb0.05). Thus, we determined the actual increase in
526 M.M. Messina et al. / Physiology & Behavior 88 (2006) 523–528
food intake following MCH infusion, relative to that consumedduring saline infusion. This analysis revealed that the orexigeniceffect of MCH on 4-h dark-phase food intake was significantlyreduced by estradiol treatment, Pb0.05 (Fig. 2). During thesubsequent 16-h interval, MCH failed to influence food intake ineither oil- or estradiol-treated rats [F(1,9)=0.53, n.s.; data notshown].
The ratio of dark-phase water intake to food intake wasdetermined to further characterize any dipsogenic effect ofMCH. This ratio was influenced by MCH during the first andthe second 2-h interval following drug treatment [F(1,9)=10.18and 20.94, respectively, Pb0.01] (Fig. 3). During each interval,MCH suppressed the ratio of water intake to food intake in bothoil- and estradiol-treated rats, Pb0.05.
3.2. Experiment 2
Because estradiol attenuated the orexigenic effect of MCH inExperiment 1, we limited our subsequent investigation ofMCH'seffect on light-phase water intake to oil-treated ovariectomizedrats. In the absence of food, MCH failed to influence light-phasewater intake during the first h following drug treatment (0.9±0.2 ml in vehicle-treated rats versus 0.9±0.2 ml in MCH-treated
Fig. 3. Effect of MCH on the ratio of dark-phase water intake to food intakeduring the first (A) and second (B) 2-h intervals following drug treatment. Dataare means±S.E. MCH suppressed the ratio of water intake to food intake duringboth intervals. ⁎Less than SAL-treated rats, Pb0.01. Abbreviation: E2,estradiol.
Fig. 4. Effect of MCH on light-phase water intake in oil-treated ovariectomizedrats. Data are means±S.E. MCH failed to influence light-phase water intakeeither in the absence (A) or presence (B) of food.
rats; t(5)=0.06, n.s.). MCH also failed to influence light-phasewater intake when this analysis was extended to include thesecond h following MCH treatment [t(5)=1.02, n.s.] (Fig. 4A).During the subsequent 2 h interval, when both food and waterwere available, MCH continued to have no effect on water intake[t(5)=0.13, n.s.] (Fig. 4B).
4. Discussion
The primary goal of this study was to determine whetherestradiol modulates the orexigenic effect of MCH. Ovariecto-mized rats were pre-treated with a physiological regimen of es-tradiol or oil vehicle treatment and then food intakewasmonitoredfor the first 4 h of the dark phase following icv infusions of MCHor saline vehicle. In support of our hypothesis, estradiol treatmentdelayed the onset and decreased the magnitude of MCH-inducedfeeding. These findings provide the first evidence that a physio-logical dose of estradiol decreases the orexigenic effect of MCH.
In the present study, acute administration of MCH increaseddark-phase food intake for 4 h in oil- and estradiol-treatedovariectomized rats. No further effect of MCH on food intakewas observed after this time. The same dose of MCH producedsimilar increases in dark-phase food intake for 2 h in male rats[4,25,27,35]. That others have reported increased food intakefor 2–4 h following acute administration of MCH during the

527M.M. Messina et al. / Physiology & Behavior 88 (2006) 523–528
early to mid-light cycle [4,5,27] provides compelling evidencethat MCH is a potent, short-term feeding stimulus with similaractions during both phases of the lighting cycle. A novel findinghere is that estradiol decreased the orexigenic effect of MCH.That is, the increase in food intake by MCH, relative to thatconsumed following vehicle treatment, a calculation that con-trols for estradiol's inhibitory effect on baseline food intake,was significantly reduced in estradiol-treated rats, relative to oil-treated rats (Fig. 2). These findings provide the first evidencethat a physiological dose of estradiol is sufficient to decreaseMCH-induced feeding. Importantly, rats received a regimen ofestradiol treatment that mimics the changes in estradiol sec-retion in cycling female rats [1]. This raises the possibility thatthe estrous decrease in food intake, which is well characterizedin cycling female rats, may be mediated, at least in part, by adecrease in MCH neurotransmission.
Additional studies are necessary to elucidate the mechanismby which estradiol interacts with MCH to decrease food intake infemale rats. One possibility is that estradiol may decrease theactivity of MCH neurons within the LH and/or ZI. As a steroidhormone, estradiol can modulate gene expression after couplingwith nuclear ERproteins locatedwithinmultiple tissues and organsystems containing estrogen-responsive genes. That ERs havebeen localized within the LH and ZI [30], makes MCH neuronspossible targets of estradiol action. In support of this hypothesis,physiological doses of estradiol have been shown to decreaseMCH mRNA expression in the medial ZI of ovariectomized rats[22] and the LH of obese male mice [21]. However, pharmaco-logical doses of estradiol failed to alter MCH mRNA expressionin the LH of male rats [23]. Taken together, these studies suggestthat physiological, but not pharmacological, doses of estradiol candecrease the expression of MCH mRNAwithin the LH/ZI com-plex of male and female rodents. Thus, estradiol may act on apopulation of cells within the LH/ZI complex to suppress tran-scription of the gene that encodes prepro-MCH and, thereby, de-crease the orexigenic effect ofMCH.Additional studies involvinga selective MCH-1 receptor antagonist are necessary to establishwhether the anorexigenic effect of estradiol involves inhibition ofthe synthesis or release of endogenous MCH. A second mecha-nism involves estradiol modulation of the number and/or bindingaffinity of postsynaptic MCH-1 receptors. Because MCH-1 re-ceptors and ERs have been localizedwithinmultiple brain regionsimplicated in the control of food intake [14,30], it will beimportant to determinewhether cells containingMCH-1 receptorsalso contain ERs and, thereby, could serve as targets of estradiolaction.
Previously, MCH was reported to increase water intake in-dependent of food intake in male rats [4]. On the basis of thisstudy, we examined water intake, in addition to food intake,following infusion of MCH and saline vehicle at the onset of thedark phase. Because rats are prandial drinkers, the ratio of waterintake to food intake was also examined. Following saline in-fusions, both oil- and estradiol-treated rats consumed abouttwice as much water than food. This is consistent with previousstudies involving both male and female rats (e.g., [2,8,13]).While several groups have shown that estradiol treatment inovariectomized rats decreases stimulated water intake following
water deprivation or administration of angiotensin [10,15,18],few have demonstrated an inhibitory effect of estradiol onspontaneous water intake (see [18]). Thus, it is not surprisingthat estradiol did not modulate water intake in the present study.Although dark-phase food intake was increased by MCH, aconcomitant increase in water intake was not observed in eitheroil- or estradiol-treated rats. The selective feeding response re-sulted in a lower ratio of water intake to food intake followingMCH infusion, relative to that observed following saline infu-sion. This finding is consistent with a previous study in whichacute administration of another orexigenic peptide, agouti-re-lated protein (AgRP), induced a transient increase in dark-phasefood intake, without influencing water intake [4]. These find-ings suggest that, under certain conditions, both MCH andAgRP stimulate feeding without inducing a prandial-related in-crease in water intake.
In Experiment 2, we further tested the dipsogenic effect ofMCHbymonitoring light-phase water intake first in the absence and thenin the presence of food. Because estradiol decreased the orexigeniceffect ofMCH inExperiment 1, only oil-treated ratswere examinedin order tomaximize our chances of detecting a dipsogenic effect ofMCH. Acute administration of MCH failed to increase light-phasewater intake, regardless of food availability. Thus, both our light-and dark-phase tests failed to reveal a dipsogenic effect of MCH inovariectomized rats. Despite virtually identical protocols, our fin-dings are not consistent with a previous report that MCH increaseddark-phase water intake in the presence of food, as well as light-phase water intake in the absence of food, in male rats [4]. That wewere unable to replicate these findings suggests that the dipsogeniceffect of MCH is limited to male rats. However, this hypothesis isweakened somewhat by a recent report by Shearman et al. [28] inwhich neither anMCH receptor agonist (Merck Compound A) noran MCH receptor antagonist (Merck Compound B) influencedwater intake in male rats [28]. Our discrepant findings may also berelated to the route of MCH administration. Previously, MCH wasinfused into the third ventricle, not the lateral ventricle, as in ourstudy. Thus, a dipsogenic effect of MCH may be more readilyapparent whenMCH is infused into the third ventricle, which lies incloser proximity to brain areas implicated in the control of fluidbalance. Additional research is necessary to determine the extent towhich MCH participates in the regulation of fluid balance in maleand female rats.
In summary, we have provided the first evidence that a phy-siological dose of estradiol decreases the orexigenic effect of MCHin ovariectomized rats. Estradiol is also known to decrease foodintake by increasing the strength of anorexigenic compounds thatfunction to limit meal size including cholecystokinin, fenfluramine,and glucagon [3,7,9,11,12,26,37]. Thus, the hypophagic effect ofestradiol appears to be mediated by its ability to interact with mul-tiple hormone, neuropeptide, and neurotransmitter systems thatcontrol food intake. While the extent to which each individualcompound contributes to the hypophagic effect of estradiol is notknown, there is some evidence that a lack of any one compoundmay not alter the hypophagic effect of estradiol. For example, inMCH-deficient male mice, estradiol induced a transient decrease infood intake that was limited to the first day of a chronic (7-day)estradiol treatment regimen [34]. Although these data suggest that

528 M.M. Messina et al. / Physiology & Behavior 88 (2006) 523–528
MCH-deficient mice maintain some short-term sensitivity to theanorexigenic effect of estradiol, additional studies are required todetermine the extent to which the anorexigenic effect of estradiolcan be expressed independent of MCH signaling. A secondaryfinding in the present study was the failure of MCH to elicit adipsogenic response in either oil- or estradiol-treated ova-riectomized rats. This is not consistent with a recent report thatMCH can stimulate water intake, independent of food intake, inmale rats [4]. We conclude, therefore, that female rats may be lesssensitive than male rats to the orexigenic and dipsogenic effects ofMCH. Additional research is necessary to elucidate the relativecontribution of MCH to the control of food intake and water ba-lance in male and female rats.
Acknowledgements
We gratefully acknowledge H.M. Rivera for excellent surgicalassistance and S. Norrbin and J. Moore for their assistance in datacollection. This work was supported by a National Institute ofMental Health Grant MH-63932 (L.A. Eckel).
References
[1] Asarian L, Geary N. Cyclic estradiol treatment normalizes body weightand restores physiological patterns of spontaneous feeding and sexualreceptivity in ovariectomized rats. Horm Behav 2002;42:461–71.
[2] Brown KJ, Grunberg NE. Effects of environmental conditions on foodconsumption in female and male rats. Physiol Behav 1996;60:221–6.
[3] Butera PC, Bradway DM, Cataldo NJ. Modulation of the satiety effect ofcholecystokinin by estradiol. Physiol Behav 1993;53:1235–8.
[4] CleggDJ, Air EL, Benoit SC, Sakai RS, Seeley RJ,Woods SC. Intraventricularmelanin-concentrating hormone stimulates water intake independent of foodintake. Am J Physiol 2003;284:R494–9.
[5] Della-Zuana O, Presse F, Ortola C, Duhault J, Nahon JL, Levens N. Acuteand chronic administration ofmelanin-concentrating hormone enhances foodintake and body weight inWistar and Sprague–Dawley rats. Int J Obes RelatMetab Disord 2002;26:1289–95.
[6] Eckel LA. Estradiol: an indirect control of meal size. Physiol Behav2004;82:35–41.
[7] Eckel LA, Geary N. Endogenous cholecystokinin's satiating action increasesduring estrus in female rats. Peptides 1999;20:451–6.
[8] Eckel LA,Houpt TA,GearyN. Spontaneousmeal patterns in female rats withand without access to running wheels. Physiol Behav 2000;70:397–405.
[9] Eckel LA, Rivera HM, Atchley DPD. The anorectic effect of fenfluramineis influenced by sex and stage of the estrous cycle in rats. Am J Physiol2005;288:R1486–91.
[10] Findlay ALR, Fitzsimons JT, Kucharczyk J. Dependence of spontaneousand angiotensin-induced drinking in the rat upon the oestrous cycle andovarian hormones. J Endocrinol 1979;82:215–25.
[11] Geary N, Asarian L. Estradiol increases glucagon's satiating potency inovariectomized rats. Am J Physiol 2001;281:R1290–4.
[12] Geary N, Trace D, McEwen B, Smith GP. Cyclic estradiol replacementincreases the satiety effect of CCK-8 in ovariectomized rats. Physiol Behav1994;56:281–9.
[13] Grill HJ, Ginsberg AB, Seeley RJ, Kaplan JM. Brainstem application ofmelanocortin receptor ligands produces long-lasting effects on feeding andbody weight. J Neurosci 1998;18:10128–35.
[14] Hervieu GJ, Cluderay JE, Harrison D, Meakin J, Maycox P, Nasir S, et al.The distribution of the mRNA and protein products of the melanin-con-centrating hormone (MCH) receptor gene, slc-1, in the central nervoussystem of the rat. Eur J Neurosci 2000;12:1194–216.
[15] Kisley LR, Sakai RR, Ma LY, Fluharty SJ. Ovarian steroid regulation ofangiotensin II-inducedwater intake in the rat. Am J Physiol 1999;276:R90–6.
[16] Kokkotou E, Jeon JY, Wang X, Marino FE, Carlson M, Trombly DJ, et al.Mice with MCH ablation resist diet-induced obesity through strain-specificmechanisms. Am J Physiol 2005;289:R117–24.
[17] Kowalski TJ, Farley C, Cohen-Williams ME, Varty G, Spar BD. Melanin-concentrating hormone-1 receptor antagonism decreases feeding by redu-cing meal size. Eur J Pharmacol 2004;497:41–7.
[18] Krause EG, Curtis KS, Davis LM, Stowe JR, Contreras RJ. Estrogen influencesstimulated water intake by ovariectomized female rats. Physiol Behav2003;79:267–74.
[19] Ludwig DS, Tritos NA,Mastaitis JW, Kulkarni R, Kokkotou E, Elmquist J,et al. Melanin-concentrating hormone overexpression in transgenic miceleads to obesity and insulin resistance. J Clin Invest 2001;107:379–86.
[20] McElroy JF, Wade GN. Short- and long-term effects of ovariectomy onfood intake, body weight, carcass composition, and brown adipose tissuein rats. Physiol Behav 1987;39:361–5.
[21] Morton GJ, Mystkowski P, Matsumoto AM, Schwartz MW. Increasedhypothalamicmelanin concentrating hormone gene expression during energyrestriction involves a melanocortin-independent, estrogen sensitive mecha-nism. Peptides 2004;25:667–74.
[22] Murray JF, Baker BI, Levy A, Wilson CA. The influence of gonadal steroids onpre-promelanin-concentratinghormonemRNAin female rats. JNeuroendocrinol2000;12:53–9.
[23] Mystkowski P, Seeley RJ, Hahn TM, Baskin DG, Havel PJ, MatsumotoAM, et al. Hypothalamic melanin-concentrating hormone and estrogen-induced weight loss. J Neurosci 2000;20:8637–42.
[24] Paxinos G, Watson C. The rat brain in stereotaxic coordinates. Fourthedition. San Diego, CA: Academic Press; 1998.
[25] Qu D, Ludwig DS, Gammeltoft S, Piper M, Pelleymounter MA, Cullen MJ,et al. A role for melanin-concentrating hormone in the central regulation offeeding behaviour. Nature 1996;380:243–7.
[26] Rivera HM, Eckel LA. The anorectic effect of fenfluramine is increased byestradiol treatment in ovariectomized rats. Physiol Behav 2005;86:331–7.
[27] Rossi M, Choi SJ, O'Shea D, Miyoshi T, Ghatei MA, Bloom SR. Melanin-concentrating hormone acutely stimulates feeding; but chronic adminis-tration has no effect on body weight. Endocrinology 1997;138:351–5.
[28] Shearman LP, Camacho RE, Sloan-Stribling D, Zhou D, Bednarek MA,Hreniuk DL, et al. Chronic MCH-1 receptor modulation alters appetite,body weight and adiposity in rats. Eur J Pharmacol 2003;475:37–47.
[29] Shimada M, Tritos NA, Lowell BB, Flier JS, Maratos-Flier E. Mice lackingmelanin-concentrating hormone are hypophagic and lean. Nature1998;396:670–4.
[30] Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution of estrogenreceptor-a and -b mRNA in the rat central nervous system. J Comp Neurol1997;388:507–25.
[31] Skofitsch G, Jacobowitz DM, Zamir N. Immunohistochemical localization ofa melanin-concentrating-like hormone in the rat brain. Brain Res Bull1985;15:635–49.
[32] Smith GP. The direct and indirect controls of meal size. Neurosci BiobehavRev 1996;20:41–6.
[33] Tarttelin MF, Gorski RA. Variations in food and water intake in the normaland acyclic female rat. Physiol Behav 1971;7:847–52.
[34] Tritos NA, Segal-Lieberman G, Vezeridis PS, Maratos-Flier E. Estradiol-induced anorexia is independent of leptin and melanin-concentrating hormone.Obesity Res 2004;12:716–24.
[35] Tritos NA, Vicent D, Gillette J, Ludwig DS, Flier ES, Maratos-Flier E.Functional interactions between melanin-concentrating hormone; neuropep-tide Y; and anorectic neuropeptides in the rat hypothalamus. Diabetes1998;47:1687–92.
[36] WadeGN. Some effects of ovarian hormones on food intake and bodyweightin female rats. J Comp Physiol Psychol 1975;88:183–93.
[37] Wager-Srdar SA,GannonM, Levine AS. The effect of cholecystokinin on foodintake in gonadectomized and intact rats: the influence of sex hormones. PhysiolBehav 1987;40:25–8.
Related Documents











