Estimating California mussel (Mytilus californianus) size from hinge fragments: a methodological application in historical ecology Breana Campbell * , Todd J. Braje Department of Anthropology, San Diego State University, 5500 Campanile Dr., San Diego, CA 92182-6040, USA article info Article history: Received 6 November 2014 Received in revised form 5 February 2015 Accepted 7 February 2015 Available online 16 February 2015 Keywords: Shellfish size Human impacts Predation pressure Size estimates Experimental archaeology abstract Whole shell measurements from archaeological sites increasingly have become an important component of historical ecological research in coastal zones around the world. Size variation in California mussels (Mytilus californianus), one of the dominant taxa in North American Pacific Coast shell midden assem- blages, have been used to assess human impacts and natural climate change on intertidal resources over millennia. Archaeological samples are typically limited due to taphonomic processes, however, which can cause fractionation and leave precious few whole shells for measurement. Various methods have been proposed to estimate the total shell length of fragmented California mussels, but these have proved largely unreliable. Using modern samples from southern California, we developed a series of regression formulae to estimate whole California mussel sizes from hinge fragments. This allometric approach proved statistically reliable and can be applied during field or laboratory work. Here, we present our methodology to build regression formulae and our validating tests using modern samples from San Diego, California, and an archaeological sample from San Miguel Island, California. © 2015 Elsevier Ltd. All rights reserved. 1. Introduction In recent decades, interest has grown in the application of archaeological data to modern environmental issues in order to enhance our understanding of the consequences of human decision-making through time (e.g., Bal ee and Erikson, 2006; Butzer, 1982; Crumley, 1994; Fitzpatrick and Keegan, 2007; Johnson et al., 2005; Kirch, 1982; Lepofsky, 2009; Lyman, 2006; Redman, 1999, 2005; Rick and Erlandson, 2008; Rick and Lockwood, 2013; Swetnam et al., 1999; Wolverton and Lyman, 2012). Archaeological shellfish sizes have become an important method for reconstructing past environments and assessing human impacts on near shore ecosystems (e.g., Bailey and Milner, 2008; Campbell, 2008; de Boer et al., 2000; Erlandson et al., 2011a; Faulkner, 2009; Giovas et al., 2010, 2013; Jerardino, 1997, 2010; Lasiak, 1991; Mannino and Thomas, 2001; Milner et al., 2007; Morrison and Cochrane, 2008; Morrison and Hunt, 2007; Stager and Chen, 1996; Stiner et al., 1999), harvesting strategies (e.g., Alvarez-Fern andez et al., 2011; Jones and Richman, 1995; Whitaker, 2008), and mobility patterns (e.g., Allen, 2012; Mannino and Thomas, 2002). Heavy predation by humans and other large predators, as well as climatic variation including fluctuations in sea surface temperatures, can reduce the mean sizes of many shellfish populations. Reduction in resource availability can result in the innovation of new technologies to procure previously unattainable resources and changes in diet and foraging strategies, making shellfish size fluctuations an important component of under- standing past human behavior. Changes in California mussel (Mytilus californianus) size through time have been a particularly important proxy for natural fluctua- tions and anthropogenic impacts on near shore intertidal com- munities on North America's Pacific Coast (Braje, 2010; Braje et al., 2007, 2012; Erlandson et al., 2008; Jazwa et al., 2012; Rick, 2007). Archaeologists are often limited, however, by taphonomic pro- cesses that cause California mussel shells to fragment (see Claassen, 1998). Two primary methods have been employed to deal with this issue. First, some studies, particularly in locations and at sites with excellent preservation, have focused on the measurement of whole shells (Braje, 2010; Erlandson et al., 2008; Rick, 2007). When large samples from well-preserved deposits are not available, archaeol- ogists have employed a template to estimate whole shell length using the hinge portion of California mussel shells (e.g., Braje et al., 2007; Jazwa et al., 2012; White, 1989). This template allows the researcher to group mussels into size classes with 1 cm variation * Corresponding author. E-mail addresses: [email protected] (B. Campbell), [email protected] (T.J. Braje). Contents lists available at ScienceDirect Journal of Archaeological Science journal homepage: http://www.elsevier.com/locate/jas http://dx.doi.org/10.1016/j.jas.2015.02.007 0305-4403/© 2015 Elsevier Ltd. All rights reserved. Journal of Archaeological Science 58 (2015) 167e174

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Journal of Archaeological Science 58 (2015) 167e174
Contents lists avai
Journal of Archaeological Science
journal homepage: http : / /www.elsevier .com/locate/ jas
Estimating California mussel (Mytilus californianus) size from hingefragments: a methodological application in historical ecology
Breana Campbell*, Todd J. BrajeDepartment of Anthropology, San Diego State University, 5500 Campanile Dr., San Diego, CA 92182-6040, USA
a r t i c l e i n f o
Article history:Received 6 November 2014Received in revised form5 February 2015Accepted 7 February 2015Available online 16 February 2015
Keywords:Shellfish sizeHuman impactsPredation pressureSize estimatesExperimental archaeology
* Corresponding author.E-mail addresses: [email protected] (B. Camp
(T.J. Braje).
http://dx.doi.org/10.1016/j.jas.2015.02.0070305-4403/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
Whole shell measurements from archaeological sites increasingly have become an important componentof historical ecological research in coastal zones around the world. Size variation in California mussels(Mytilus californianus), one of the dominant taxa in North American Pacific Coast shell midden assem-blages, have been used to assess human impacts and natural climate change on intertidal resources overmillennia. Archaeological samples are typically limited due to taphonomic processes, however, whichcan cause fractionation and leave precious few whole shells for measurement. Various methods havebeen proposed to estimate the total shell length of fragmented California mussels, but these have provedlargely unreliable. Using modern samples from southern California, we developed a series of regressionformulae to estimate whole California mussel sizes from hinge fragments. This allometric approachproved statistically reliable and can be applied during field or laboratory work. Here, we present ourmethodology to build regression formulae and our validating tests using modern samples from SanDiego, California, and an archaeological sample from San Miguel Island, California.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
In recent decades, interest has grown in the application ofarchaeological data to modern environmental issues in order toenhance our understanding of the consequences of humandecision-making through time (e.g., Bal�ee and Erikson, 2006;Butzer, 1982; Crumley, 1994; Fitzpatrick and Keegan, 2007;Johnson et al., 2005; Kirch, 1982; Lepofsky, 2009; Lyman, 2006;Redman, 1999, 2005; Rick and Erlandson, 2008; Rick andLockwood, 2013; Swetnam et al., 1999; Wolverton and Lyman,2012). Archaeological shellfish sizes have become an importantmethod for reconstructing past environments and assessing humanimpacts on near shore ecosystems (e.g., Bailey and Milner, 2008;Campbell, 2008; de Boer et al., 2000; Erlandson et al., 2011a;Faulkner, 2009; Giovas et al., 2010, 2013; Jerardino, 1997, 2010;Lasiak, 1991; Mannino and Thomas, 2001; Milner et al., 2007;Morrison and Cochrane, 2008; Morrison and Hunt, 2007; Stagerand Chen, 1996; Stiner et al., 1999), harvesting strategies (e.g.,Alvarez-Fern�andez et al., 2011; Jones and Richman,1995; Whitaker,2008), and mobility patterns (e.g., Allen, 2012; Mannino and
bell), [email protected]
Thomas, 2002). Heavy predation by humans and other largepredators, as well as climatic variation including fluctuations in seasurface temperatures, can reduce the mean sizes of many shellfishpopulations. Reduction in resource availability can result in theinnovation of new technologies to procure previously unattainableresources and changes in diet and foraging strategies, makingshellfish size fluctuations an important component of under-standing past human behavior.
Changes in California mussel (Mytilus californianus) size throughtime have been a particularly important proxy for natural fluctua-tions and anthropogenic impacts on near shore intertidal com-munities on North America's Pacific Coast (Braje, 2010; Braje et al.,2007, 2012; Erlandson et al., 2008; Jazwa et al., 2012; Rick, 2007).Archaeologists are often limited, however, by taphonomic pro-cesses that cause California mussel shells to fragment (see Claassen,1998). Two primary methods have been employed to deal with thisissue. First, some studies, particularly in locations and at sites withexcellent preservation, have focused on the measurement of wholeshells (Braje, 2010; Erlandson et al., 2008; Rick, 2007). When largesamples from well-preserved deposits are not available, archaeol-ogists have employed a template to estimate whole shell lengthusing the hinge portion of California mussel shells (e.g., Braje et al.,2007; Jazwa et al., 2012; White, 1989). This template allows theresearcher to group mussels into size classes with 1 cm variation
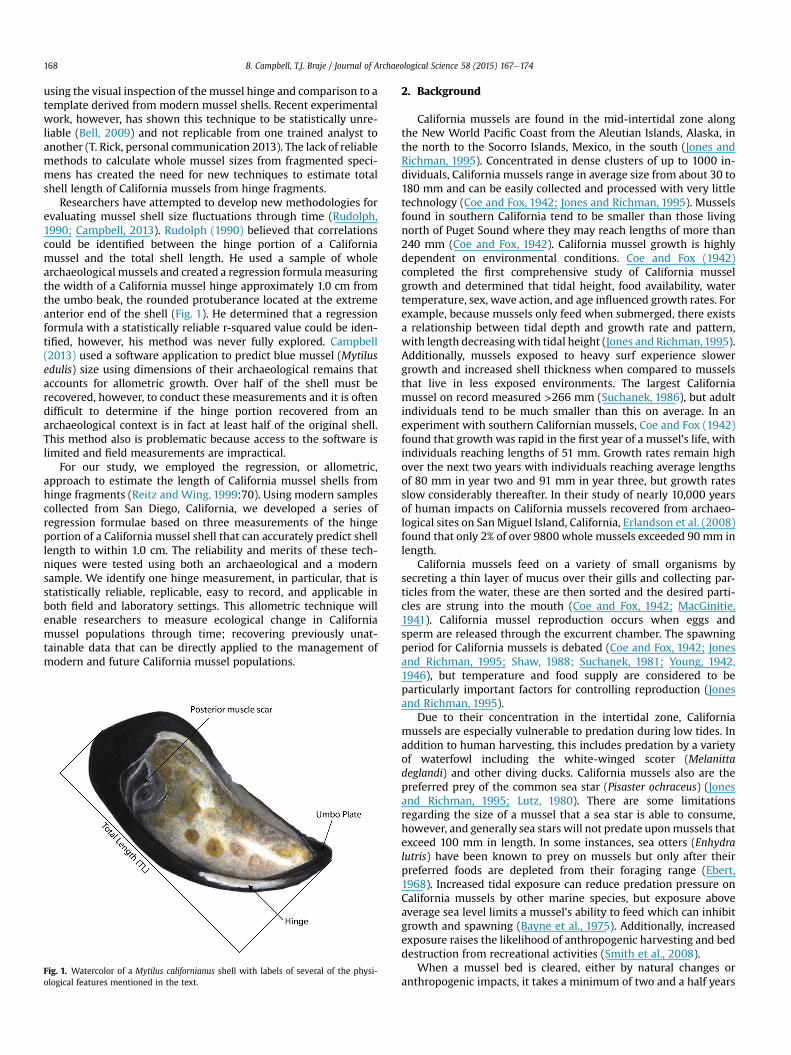
B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174168
using the visual inspection of the mussel hinge and comparison to atemplate derived from modern mussel shells. Recent experimentalwork, however, has shown this technique to be statistically unre-liable (Bell, 2009) and not replicable from one trained analyst toanother (T. Rick, personal communication 2013). The lack of reliablemethods to calculate whole mussel sizes from fragmented speci-mens has created the need for new techniques to estimate totalshell length of California mussels from hinge fragments.
Researchers have attempted to develop new methodologies forevaluating mussel shell size fluctuations through time (Rudolph,1990; Campbell, 2013). Rudolph (1990) believed that correlationscould be identified between the hinge portion of a Californiamussel and the total shell length. He used a sample of wholearchaeological mussels and created a regression formulameasuringthe width of a California mussel hinge approximately 1.0 cm fromthe umbo beak, the rounded protuberance located at the extremeanterior end of the shell (Fig. 1). He determined that a regressionformula with a statistically reliable r-squared value could be iden-tified, however, his method was never fully explored. Campbell(2013) used a software application to predict blue mussel (Mytilusedulis) size using dimensions of their archaeological remains thataccounts for allometric growth. Over half of the shell must berecovered, however, to conduct these measurements and it is oftendifficult to determine if the hinge portion recovered from anarchaeological context is in fact at least half of the original shell.This method also is problematic because access to the software islimited and field measurements are impractical.
For our study, we employed the regression, or allometric,approach to estimate the length of California mussel shells fromhinge fragments (Reitz and Wing, 1999:70). Using modern samplescollected from San Diego, California, we developed a series ofregression formulae based on three measurements of the hingeportion of a California mussel shell that can accurately predict shelllength to within 1.0 cm. The reliability and merits of these tech-niques were tested using both an archaeological and a modernsample. We identify one hinge measurement, in particular, that isstatistically reliable, replicable, easy to record, and applicable inboth field and laboratory settings. This allometric technique willenable researchers to measure ecological change in Californiamussel populations through time; recovering previously unat-tainable data that can be directly applied to the management ofmodern and future California mussel populations.
Fig. 1. Watercolor of a Mytilus californianus shell with labels of several of the physi-ological features mentioned in the text.
2. Background
California mussels are found in the mid-intertidal zone alongthe New World Pacific Coast from the Aleutian Islands, Alaska, inthe north to the Socorro Islands, Mexico, in the south (Jones andRichman, 1995). Concentrated in dense clusters of up to 1000 in-dividuals, California mussels range in average size from about 30 to180 mm and can be easily collected and processed with very littletechnology (Coe and Fox, 1942; Jones and Richman, 1995). Musselsfound in southern California tend to be smaller than those livingnorth of Puget Sound where they may reach lengths of more than240 mm (Coe and Fox, 1942). California mussel growth is highlydependent on environmental conditions. Coe and Fox (1942)completed the first comprehensive study of California musselgrowth and determined that tidal height, food availability, watertemperature, sex, wave action, and age influenced growth rates. Forexample, because mussels only feed when submerged, there existsa relationship between tidal depth and growth rate and pattern,with length decreasingwith tidal height (Jones and Richman,1995).Additionally, mussels exposed to heavy surf experience slowergrowth and increased shell thickness when compared to musselsthat live in less exposed environments. The largest Californiamussel on record measured >266 mm (Suchanek, 1986), but adultindividuals tend to be much smaller than this on average. In anexperiment with southern Californian mussels, Coe and Fox (1942)found that growth was rapid in the first year of a mussel's life, withindividuals reaching lengths of 51 mm. Growth rates remain highover the next two years with individuals reaching average lengthsof 80 mm in year two and 91 mm in year three, but growth ratesslow considerably thereafter. In their study of nearly 10,000 yearsof human impacts on California mussels recovered from archaeo-logical sites on San Miguel Island, California, Erlandson et al. (2008)found that only 2% of over 9800 whole mussels exceeded 90 mm inlength.
California mussels feed on a variety of small organisms bysecreting a thin layer of mucus over their gills and collecting par-ticles from the water, these are then sorted and the desired parti-cles are strung into the mouth (Coe and Fox, 1942; MacGinitie,1941). California mussel reproduction occurs when eggs andsperm are released through the excurrent chamber. The spawningperiod for California mussels is debated (Coe and Fox, 1942; Jonesand Richman, 1995; Shaw, 1988; Suchanek, 1981; Young, 1942,1946), but temperature and food supply are considered to beparticularly important factors for controlling reproduction (Jonesand Richman, 1995).
Due to their concentration in the intertidal zone, Californiamussels are especially vulnerable to predation during low tides. Inaddition to human harvesting, this includes predation by a varietyof waterfowl including the white-winged scoter (Melanittadeglandi) and other diving ducks. California mussels also are thepreferred prey of the common sea star (Pisaster ochraceus) (Jonesand Richman, 1995; Lutz, 1980). There are some limitationsregarding the size of a mussel that a sea star is able to consume,however, and generally sea stars will not predate uponmussels thatexceed 100 mm in length. In some instances, sea otters (Enhydralutris) have been known to prey on mussels but only after theirpreferred foods are depleted from their foraging range (Ebert,1968). Increased tidal exposure can reduce predation pressure onCalifornia mussels by other marine species, but exposure aboveaverage sea level limits a mussel's ability to feed which can inhibitgrowth and spawning (Bayne et al., 1975). Additionally, increasedexposure raises the likelihood of anthropogenic harvesting and beddestruction from recreational activities (Smith et al., 2008).
When a mussel bed is cleared, either by natural changes oranthropogenic impacts, it takes a minimum of two and a half years

B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174 169
to start the regeneration process (Jones and Richman, 1995). Oncemussels begin to repopulate an area it typically takes between 8and 35 years to return to its pre-disturbance state. Suchanek (1979)has suggested, however, this process, which is highly sensitive toenvironmental conditions, can take up to 100 years. Once repo-pulated, a relatively undisturbed population can survive for 50e100years (Suchanek, 1981).
California mussels have been an important food and tool sourcefor ancient and historical occupants of the North American PacificCoast for at least 12,000 years (e.g., Erlandson et al., 2011b). On theNorthern Channel Islands (Fig. 2), located between 20 and 44 kmfrom the mainland coast of California, data from archaeologicalsites have long been used to explore the relationship betweenhumans and intertidal shellfish communities (e.g., Braje et al., 2007,2012; Erlandson et al., 2008, 2011a; Jazwa et al., 2012). Due to theease of collection en masse as well as their extensive distributionaround the islands, Braje et al. (2007:740) ranked Californiamusselsas the top prey choice for marine foragers living on the ChannelIslands. Today, California mussel harvesting is mostly recreational;a ten pound (in shell) collection limit requiring a one-day fishinglicense is the only legislation regarding the harvest of musselsoutside of marine protected areas.
3. Materials and methods
In November 2012, Campbell collected a robust sample ofmodern California mussels (n¼ 135) from a single rocky intertidalhabitat in San Diego, California. Prior to collection, arbitrary classeswere established to ensure adequate total shell length variationduring the collection process: Class 1 (n ¼ 55, 0e38 mm), Class 2(n ¼ 50, 38.1e68 mm), and Class 3 (n ¼ 30, 68.1e86.32 mm). Eachmussel was measured and classified in the field, and transported toSan Diego State University's Environmental Anthropology andArchaeology Laboratory for further analysis.
The viscera of each mussel were discarded and broken musselshells were removed from the sample. Each valve was placed inexterior view with the umbo located at the distal end, the place-ment of the hinge (either on the left or right side of the shell) wasused to determine the left from the right valve. The total length ofthe right and left bivalves of eachmussel wasmeasuredwith digitalcalipers, recorded, and shells were cataloged and stored. The rightvalve of each mussel was used to record three different hinge
Fig. 2. Location map of the Santa Barbara Channel region, the No
measurements: umbowidth, umbo length, and umbo height. Umbowidth was measured where the hinge plate is at its greatest width.Umbo length was recorded bymeasuring from the tip of the beak tothe interior end of the hinge plate. Umbo height was measured5mm from the tip of the beak and the calipers were placedwith theteeth perpendicular to the shell (see inset photographs in Fig. 3).Each measurement was recorded three times and the results wereaveraged.
We then employed a regression method to build formulae thatcan predict whole California mussel size from hingemeasurements.The regression approach provides an accurate method for esti-mating the original size of whole shells from fragments and in-volves the determination of the relationship between twovariables, commonly accomplished using the least-squaresregression method (Johnson and Bhattacharyya, 1992). For twovariables under consideration, x and y are plotted against oneanother and the least-squaresmethod determines “the equation forthe line that minimizes the sum of squares of the vertical distancesfrom the data points to the regression line” (Orchard, 2001:67, alsosee Casteel, 1976; Ricker, 1973). Common for estimating whole shelllength, the linear or exponential relationship is expressed with amathematical formula that describes the relationship between thetwo variables being compared. In Microsoft Excel, regressionformulae were built using the averaged measurements to deter-mine correlations in total mussel shell length and umbo mea-surements. The umbo width, length, and height measurementswere assigned the x-variable; total mussel shell length wasassigned the y-variable for each equation.
Two tests were conducted to determine the validity of theregression formulae. First, a sample of the modernmussels (n¼ 30)used to identify the regression formulae were selected by the au-thors; 10 mussels were chosen randomly from each of the previ-ously established shell classes to ensure a variety in sizes tested.Using a hammer, the mussels were broken, leaving only the hingeportions intact. A graduate student was trained to measure umbowidth, length, and height using a sample of whole mussel shellsand then instructed to record these measurements on the frag-mented mussels, repeating the measurement three times. The re-sults were averaged, applied to the respective regression formulae,and total shell length for each mussel was estimated. A one-wayanalysis of variance (ANOVA), a procedure that analyzes whetherthe differences between more than two groups are statistically
rthern Channel Islands, San Miguel Island, and CA-SMI-232.
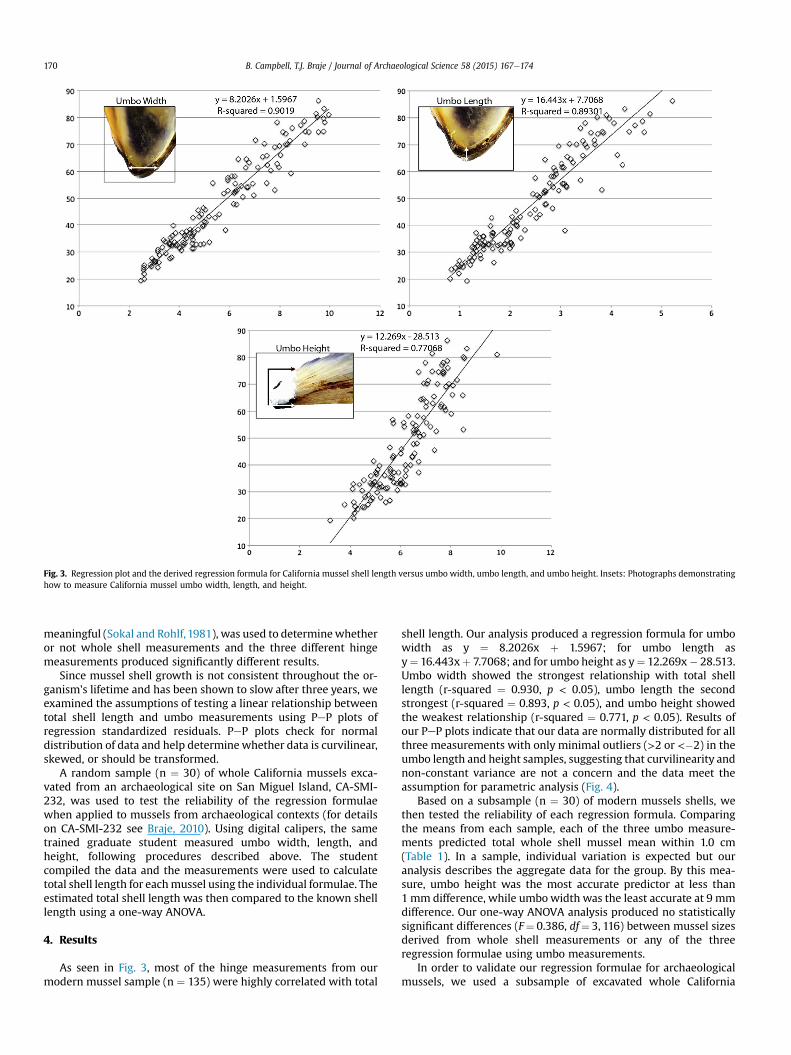
Fig. 3. Regression plot and the derived regression formula for California mussel shell length versus umbo width, umbo length, and umbo height. Insets: Photographs demonstratinghow to measure California mussel umbo width, length, and height.
B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174170
meaningful (Sokal and Rohlf, 1981), was used to determinewhetheror not whole shell measurements and the three different hingemeasurements produced significantly different results.
Since mussel shell growth is not consistent throughout the or-ganism's lifetime and has been shown to slow after three years, weexamined the assumptions of testing a linear relationship betweentotal shell length and umbo measurements using PeP plots ofregression standardized residuals. PeP plots check for normaldistribution of data and help determine whether data is curvilinear,skewed, or should be transformed.
A random sample (n ¼ 30) of whole California mussels exca-vated from an archaeological site on San Miguel Island, CA-SMI-232, was used to test the reliability of the regression formulaewhen applied to mussels from archaeological contexts (for detailson CA-SMI-232 see Braje, 2010). Using digital calipers, the sametrained graduate student measured umbo width, length, andheight, following procedures described above. The studentcompiled the data and the measurements were used to calculatetotal shell length for eachmussel using the individual formulae. Theestimated total shell length was then compared to the known shelllength using a one-way ANOVA.
4. Results
As seen in Fig. 3, most of the hinge measurements from ourmodern mussel sample (n ¼ 135) were highly correlated with total
shell length. Our analysis produced a regression formula for umbowidth as y ¼ 8.2026x þ 1.5967; for umbo length asy¼ 16.443xþ 7.7068; and for umbo height as y¼ 12.269x� 28.513.Umbo width showed the strongest relationship with total shelllength (r-squared ¼ 0.930, p < 0.05), umbo length the secondstrongest (r-squared ¼ 0.893, p < 0.05), and umbo height showedthe weakest relationship (r-squared ¼ 0.771, p < 0.05). Results ofour PeP plots indicate that our data are normally distributed for allthree measurements with only minimal outliers (>2 or <�2) in theumbo length and height samples, suggesting that curvilinearity andnon-constant variance are not a concern and the data meet theassumption for parametric analysis (Fig. 4).
Based on a subsample (n ¼ 30) of modern mussels shells, wethen tested the reliability of each regression formula. Comparingthe means from each sample, each of the three umbo measure-ments predicted total whole shell mussel mean within 1.0 cm(Table 1). In a sample, individual variation is expected but ouranalysis describes the aggregate data for the group. By this mea-sure, umbo height was the most accurate predictor at less than1mm difference, while umbowidth was the least accurate at 9 mmdifference. Our one-way ANOVA analysis produced no statisticallysignificant differences (F¼ 0.386, df¼ 3, 116) between mussel sizesderived from whole shell measurements or any of the threeregression formulae using umbo measurements.
In order to validate our regression formulae for archaeologicalmussels, we used a subsample of excavated whole California
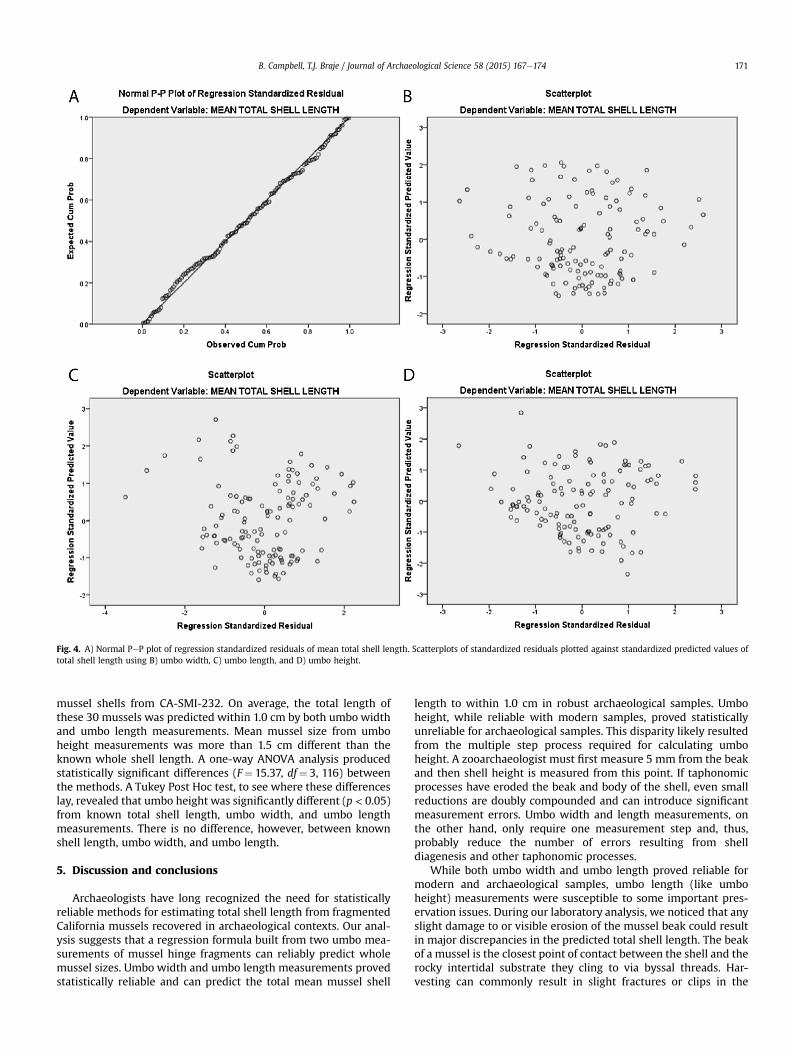
Fig. 4. A) Normal PeP plot of regression standardized residuals of mean total shell length. Scatterplots of standardized residuals plotted against standardized predicted values oftotal shell length using B) umbo width, C) umbo length, and D) umbo height.
B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174 171
mussel shells from CA-SMI-232. On average, the total length ofthese 30 mussels was predicted within 1.0 cm by both umbo widthand umbo length measurements. Mean mussel size from umboheight measurements was more than 1.5 cm different than theknown whole shell length. A one-way ANOVA analysis producedstatistically significant differences (F¼ 15.37, df¼ 3, 116) betweenthe methods. A Tukey Post Hoc test, to see where these differenceslay, revealed that umbo height was significantly different (p < 0.05)from known total shell length, umbo width, and umbo lengthmeasurements. There is no difference, however, between knownshell length, umbo width, and umbo length.
5. Discussion and conclusions
Archaeologists have long recognized the need for statisticallyreliable methods for estimating total shell length from fragmentedCalifornia mussels recovered in archaeological contexts. Our anal-ysis suggests that a regression formula built from two umbo mea-surements of mussel hinge fragments can reliably predict wholemussel sizes. Umbo width and umbo length measurements provedstatistically reliable and can predict the total mean mussel shell
length to within 1.0 cm in robust archaeological samples. Umboheight, while reliable with modern samples, proved statisticallyunreliable for archaeological samples. This disparity likely resultedfrom the multiple step process required for calculating umboheight. A zooarchaeologist must first measure 5 mm from the beakand then shell height is measured from this point. If taphonomicprocesses have eroded the beak and body of the shell, even smallreductions are doubly compounded and can introduce significantmeasurement errors. Umbo width and length measurements, onthe other hand, only require one measurement step and, thus,probably reduce the number of errors resulting from shelldiagenesis and other taphonomic processes.
While both umbo width and umbo length proved reliable formodern and archaeological samples, umbo length (like umboheight) measurements were susceptible to some important pres-ervation issues. During our laboratory analysis, we noticed that anyslight damage to or visible erosion of the mussel beak could resultin major discrepancies in the predicted total shell length. The beakof a mussel is the closest point of contact between the shell and therocky intertidal substrate they cling to via byssal threads. Har-vesting can commonly result in slight fractures or clips in the
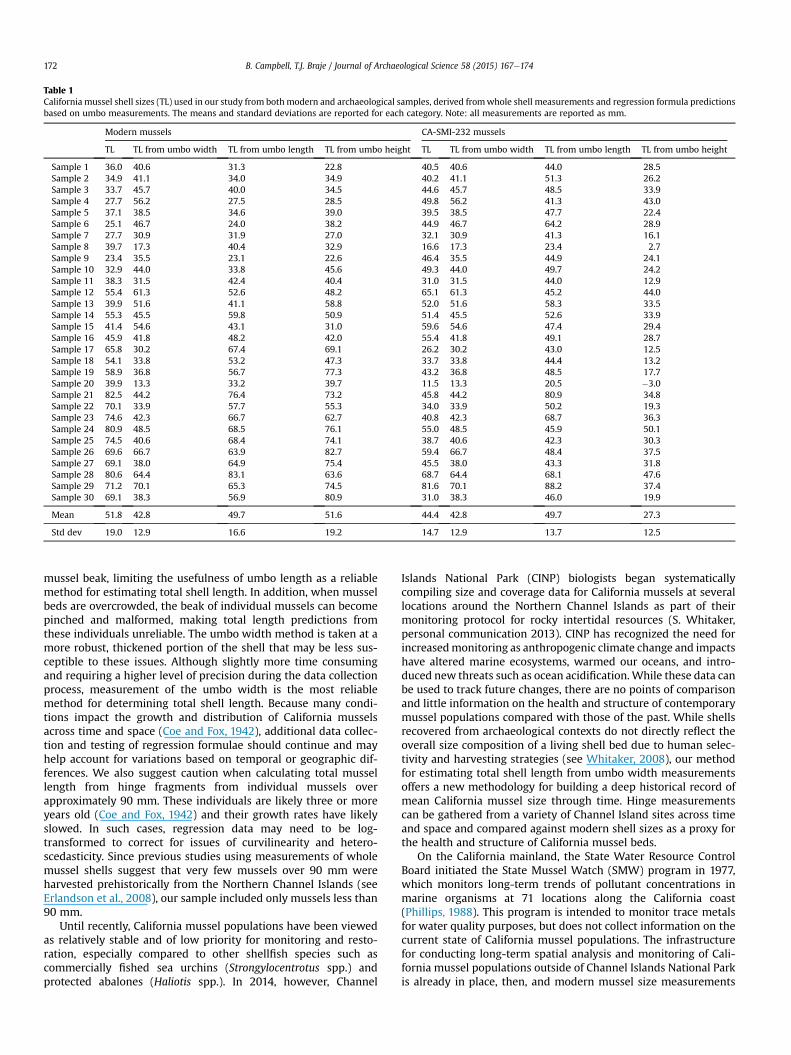
Table 1California mussel shell sizes (TL) used in our study from bothmodern and archaeological samples, derived fromwhole shell measurements and regression formula predictionsbased on umbo measurements. The means and standard deviations are reported for each category. Note: all measurements are reported as mm.
Modern mussels CA-SMI-232 mussels
TL TL from umbo width TL from umbo length TL from umbo height TL TL from umbo width TL from umbo length TL from umbo height
Sample 1 36.0 40.6 31.3 22.8 40.5 40.6 44.0 28.5Sample 2 34.9 41.1 34.0 34.9 40.2 41.1 51.3 26.2Sample 3 33.7 45.7 40.0 34.5 44.6 45.7 48.5 33.9Sample 4 27.7 56.2 27.5 28.5 49.8 56.2 41.3 43.0Sample 5 37.1 38.5 34.6 39.0 39.5 38.5 47.7 22.4Sample 6 25.1 46.7 24.0 38.2 44.9 46.7 64.2 28.9Sample 7 27.7 30.9 31.9 27.0 32.1 30.9 41.3 16.1Sample 8 39.7 17.3 40.4 32.9 16.6 17.3 23.4 2.7Sample 9 23.4 35.5 23.1 22.6 46.4 35.5 44.9 24.1Sample 10 32.9 44.0 33.8 45.6 49.3 44.0 49.7 24.2Sample 11 38.3 31.5 42.4 40.4 31.0 31.5 44.0 12.9Sample 12 55.4 61.3 52.6 48.2 65.1 61.3 45.2 44.0Sample 13 39.9 51.6 41.1 58.8 52.0 51.6 58.3 33.5Sample 14 55.3 45.5 59.8 50.9 51.4 45.5 52.6 33.9Sample 15 41.4 54.6 43.1 31.0 59.6 54.6 47.4 29.4Sample 16 45.9 41.8 48.2 42.0 55.4 41.8 49.1 28.7Sample 17 65.8 30.2 67.4 69.1 26.2 30.2 43.0 12.5Sample 18 54.1 33.8 53.2 47.3 33.7 33.8 44.4 13.2Sample 19 58.9 36.8 56.7 77.3 43.2 36.8 48.5 17.7Sample 20 39.9 13.3 33.2 39.7 11.5 13.3 20.5 �3.0Sample 21 82.5 44.2 76.4 73.2 45.8 44.2 80.9 34.8Sample 22 70.1 33.9 57.7 55.3 34.0 33.9 50.2 19.3Sample 23 74.6 42.3 66.7 62.7 40.8 42.3 68.7 36.3Sample 24 80.9 48.5 68.5 76.1 55.0 48.5 45.9 50.1Sample 25 74.5 40.6 68.4 74.1 38.7 40.6 42.3 30.3Sample 26 69.6 66.7 63.9 82.7 59.4 66.7 48.4 37.5Sample 27 69.1 38.0 64.9 75.4 45.5 38.0 43.3 31.8Sample 28 80.6 64.4 83.1 63.6 68.7 64.4 68.1 47.6Sample 29 71.2 70.1 65.3 74.5 81.6 70.1 88.2 37.4Sample 30 69.1 38.3 56.9 80.9 31.0 38.3 46.0 19.9
Mean 51.8 42.8 49.7 51.6 44.4 42.8 49.7 27.3
Std dev 19.0 12.9 16.6 19.2 14.7 12.9 13.7 12.5
B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174172
mussel beak, limiting the usefulness of umbo length as a reliablemethod for estimating total shell length. In addition, when musselbeds are overcrowded, the beak of individual mussels can becomepinched and malformed, making total length predictions fromthese individuals unreliable. The umbo width method is taken at amore robust, thickened portion of the shell that may be less sus-ceptible to these issues. Although slightly more time consumingand requiring a higher level of precision during the data collectionprocess, measurement of the umbo width is the most reliablemethod for determining total shell length. Because many condi-tions impact the growth and distribution of California musselsacross time and space (Coe and Fox, 1942), additional data collec-tion and testing of regression formulae should continue and mayhelp account for variations based on temporal or geographic dif-ferences. We also suggest caution when calculating total mussellength from hinge fragments from individual mussels overapproximately 90 mm. These individuals are likely three or moreyears old (Coe and Fox, 1942) and their growth rates have likelyslowed. In such cases, regression data may need to be log-transformed to correct for issues of curvilinearity and hetero-scedasticity. Since previous studies using measurements of wholemussel shells suggest that very few mussels over 90 mm wereharvested prehistorically from the Northern Channel Islands (seeErlandson et al., 2008), our sample included only mussels less than90 mm.
Until recently, California mussel populations have been viewedas relatively stable and of low priority for monitoring and resto-ration, especially compared to other shellfish species such ascommercially fished sea urchins (Strongylocentrotus spp.) andprotected abalones (Haliotis spp.). In 2014, however, Channel
Islands National Park (CINP) biologists began systematicallycompiling size and coverage data for California mussels at severallocations around the Northern Channel Islands as part of theirmonitoring protocol for rocky intertidal resources (S. Whitaker,personal communication 2013). CINP has recognized the need forincreasedmonitoring as anthropogenic climate change and impactshave altered marine ecosystems, warmed our oceans, and intro-duced new threats such as ocean acidification.While these data canbe used to track future changes, there are no points of comparisonand little information on the health and structure of contemporarymussel populations compared with those of the past. While shellsrecovered from archaeological contexts do not directly reflect theoverall size composition of a living shell bed due to human selec-tivity and harvesting strategies (see Whitaker, 2008), our methodfor estimating total shell length from umbo width measurementsoffers a new methodology for building a deep historical record ofmean California mussel size through time. Hinge measurementscan be gathered from a variety of Channel Island sites across timeand space and compared against modern shell sizes as a proxy forthe health and structure of California mussel beds.
On the California mainland, the State Water Resource ControlBoard initiated the State Mussel Watch (SMW) program in 1977,which monitors long-term trends of pollutant concentrations inmarine organisms at 71 locations along the California coast(Phillips, 1988). This program is intended to monitor trace metalsfor water quality purposes, but does not collect information on thecurrent state of California mussel populations. The infrastructurefor conducting long-term spatial analysis and monitoring of Cali-fornia mussel populations outside of Channel Islands National Parkis already in place, then, and modern mussel size measurements

B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174 173
could be included as part of the annual study conducted by theSMW. These data could then be compared against mussel sizesfrom adjacent archaeological sites using umbo width measure-ments. Very quickly, a set of deep historical data would be availableto assess the health and structure of modern California mussel bedsacross the state.
Ultimately, our study offers an excellent opportunity to incor-porate deep historical perspectives in modernmarinemanagementprograms (Dayton et al., 1998; Jackson et al., 2001; Pauly, 1995;Pauly et al., 1998; Rick and Erlandson, 2008; Tegner and Dayton,2000). Using archaeological data to decipher the deeper ecolog-ical histories of specific areas and fisheries can provide valuableinformation regarding the structure of near shore ecosystems in thedistant past as well as the processes involved in the creation ofanthropogenic land- and seascapes during the Anthropocene.Building effective management and restoration plans requires in-formation about historical and ancient harvests and the establish-ment of reference points or baselines that account for overfishing,other anthropogenic disturbances, and natural climaticfluctuations.
Acknowledgments
Funding for this project was provided to Campbell by CaliforniaState University's COAST program and San Diego State University's(SDSU) Department of Anthropology Norton Allen ScholarshipFund. We thank Matthew Lauer, Brian Hentschel, and StephenSchellenberg for their comments and suggestions on early drafts ofthis manuscript. Special thanks to Sopagna Eap for statisticalanalysis consultation. Steve Campbell, Lucas Sanchez, Linda Bentz,Stephanie Duncan, Devin Chouinard, and several SDSU Environ-mental Anthropology and Archaeology Laboratory student internsvolunteered their time to assist with data collection. We thankthree anonymous reviewers and the editors at JAS for their helpwith the review and production of our manuscript.
References
Allen, M.S., 2012. Molluscan foraging efficiency and patterns of mobility amongstforaging, agriculturalists: a case study from Northern New Zealand. J. Archaeol.Sci. 39, 295e307.
Alvarez-Fern�andez, E., Chauvin, A., Cubas, M., Arias, P., Onta~n�on, R., 2011. Molluscshell sizes in archaeological contexts in Northern Spain (13,200 to 2600 cal BC):new data from La Garma A and Los Gitanos (Cantabria). Archaeometry 53,963e985.
Bailey, G.N., Milner, N., 2008. Molluscan archives from European prehistory. In:Antczak, A., Cipriani, R. (Eds.), Early Human Impact on Megamolluscs, BAR In-ternational Series 1865. Archaeopress, Oxford, pp. 111e134.
Bal�ee, W., Erikson, C.L. (Eds.), 2006. Time and Complexity in Historical Ecology:Studies in the Neotropical Lowlands. Columbia University Press, New York.
Bayne, B.L., Gabbott, P.A., Widdows, J., 1975. Some effects of stress in the adult onthe eggs and larvae of Mytilus edulis L. J. Mar. Biol. Assoc. U. K. 55, 675e689.
Bell, A.M., 2009. On the Validity of Archaeological Shellfish Metrics in CoastalCalifornia. Department of Anthropology, California State University, Chico(Unpublished Masters thesis).
Braje, T.J., 2010. Modern Oceans, Ancient Sites: Archaeology and Marine Conser-vation on San Miguel Island, California. University of Utah Press, Salt Lake City.
Braje, T.J., Kennett, D.J., Erlandson, J.M., Culleton, B.J., 2007. Human impacts onnearshore shellfish taxa: a 7,000 year record from Santa Rosa Island, California.Am. Antiq. 72, 735e756.
Braje, T.J., Rick, T.C., Erlandson, J.M., 2012. A Trans-Holocene historical ecologicalrecord of shellfish harvesting on California's Northern Channel Islands. Quat.Int. 264, 109e120.
Butzer, K.W., 1982. Archaeology as Human Ecology. Cambridge University Press.Campbell, G.E., 2013. Size prediction in archaeomalacology: the Common Mussel,
Mytilus edulis L., as an example. Archaeol. Anthropol. Sci. 1e14.Campbell, G.E., 2008. Beyond means to meaning: using distributions of shell shapes
to reconstruct past collecting strategies. Environ. Archaeol. 13, 111e121.Casteel, R.W., 1976. Fish Remains in Archaeology and Paleo-environmental Studies.
Academic Press Inc., New York.Claassen, C., 1998. Shells. Cambridge University Press, Cambridge.
Coe, W.R., Fox, D.L., 1942. Biology of the California sea-mussel (Mytilus cal-ifornianus): influence of temperature, food supply, sex and age on the rate ofgrowth. J. Exp. Zool. 90, 1e30.
Crumley, C.L. (Ed.), 1994. Historical Ecology: Cultural Knowledge and ChangingLandscapes. School of American Research Press, Santa Fe, New Mexico.
Dayton, P.K., Tegner, M.J., Edwards, P.B., Riser, K.L., 1998. Sliding baselines, ghosts,and reduced expectations in kelp forest communities. Ecol. Appl. 8, 309e322.
de Boer, W.F., Pereira, T., Guissamulo, A., 2000. Comparing recent and abandonedshell middens to detect the impact of human exploitation on the intertidalecosystem. Aquat. Ecol. 34, 287e297.
Ebert, E., 1968. A food habits study of the southern sea otter, Enhydra lutris nereis.Calif. Fish Game 54, 33e42.
Erlandson, J.M., Braje, T.J., Rick, T.C., Jew, N.P., Kennett, D.J., Dwyer, N., Ainis, A.F.,Vellanoweth, R.L., Watts, J., 2011a. 10,000 years of human predation and sizechanges in the Owl Limpet (Lottia gigantea) on San Miguel Island, California.J. Archaeol. Sci. 38, 1127e1134.
Erlandson, J.M., Rick, T.C., Braje, T.B., Casperson, M., Culleton, B., Fulfrost, B.,Garcia, T., Guthrie, D.A., Jew, N.P., Kennett, D.J., Moss, M.L., Reeder, L., Skinner, C.,Watts, J., Willis, L., 2011b. Paleo-Indian seafaring, maritime technologies, andcoastal foraging on California's Channel Islands. Science 331, 1181e1185.
Erlandson, J.M., Rick, T.C., Braje, T.J., Steinberg, A., Vellanoweth, R.L., 2008. Humanimpacts on ancient shellfish: a 10,000 year record from San Miguel Island,California. J. Archaeol. Sci. 35, 2144e2152.
Faulkner, P., 2009. Focused, intense and long-term: evidence for Granular Ark(Anadara granosa) exploitation from Late Holocene shell mounds of Blue MudBay, Northern Australia. J. Archaeol. Sci. 36, 821e834.
Fitzpatrick, S.M., Keegan, W.F., 2007. Human impacts and adaptations in theCaribbean Islands: an historical ecology approach. Earth Environ. Sci. Trans. R.Soc. Edinb. 98, 29e45.
Giovas, C.M., Clark, M., Fitzpatrick, S.M., Stone, J., 2013. Intensifying collection andsize increase of the Tessellated Nerite Snail (Nerita tessellate) at the CoconutWalk site, Nevis, northern Lesser Antilles, AD 890e1440. J. Archaeol. Sci. 40,4024e4038.
Giovas, C.M., Fitzpatrick, S.F., Clark, M., Abed, M., 2010. Evidence for size increase inan exploited mollusc: Humped Conch (Strombus gibberulus) at Chelechol raOrrak, Palau from ca. 3000-0 BP. J. Archaeol. Sci. 37, 2788e2798.
Jackson, J.B.C., Kirby, M.X., Berger, W.H., Bjorndal, K.A., Botsford, L.W., Bourque, B.J.,Bradbury, R.H., Cooke, R., Erlandson, J.M., Estes, J.A., 2001. Historical overfishingand the recent collapse of coastal ecosystems. Science 293, 629e637.
Jazwa, C.S., Kennett, D.J., Hanson, D., 2012. Late Holocene subsistence change andmarine productivity on Western Santa Rosa Island, Alta California. Calif.Archaeol. 4, 69e98.
Jerardino, A., 1997. Changes in shellfish species composition and mean shell sizefrom a Late- Holocene record of the West Coast of South Africa. J. Archaeol. Sci.24, 1031e1044.
Jerardino, A., 2010. Large shell middens in Lamberts Bay, South Africa: a case ofhunter- gatherer resource intensification. J. Archaeol. Sci. 37, 2291e2302.
Johnson, R.A., Bhattacharyya, G.K., 1992. Statistics: Principles and Methods, seconded. John Wiley & Sons, Inc., New York.
Johnson, C.D., Kohler, T.A., Cowan, J., 2005. Modeling historical ecology, thinkingabout contemporary systems. Am. Anthropol. 107, 96e107.
Jones, T.L., Richman, J.R., 1995. On mussels: Mytilus californianus as a prehistoricresource. North Am. Archaeol. 16, 33e58.
Kirch, P.V., 1982. The impact of the prehistoric Polynesians on the Hawaiianecosystem. Pac. Sci. 36, 1e14.
Lasiak, T., 1991. The susceptibility and/or resilience of rocky Littoral Molluscs tostock depletion by the indigenous coastal people of Transkei, Southern Africa.Biol. Conserv. 56, 245e264.
Lepofsky, D., 2009. The past, present, and future of traditional resource and envi-ronmental management. J. Ethnobiol. 29, 161e166.
Lutz, R.A. (Ed.), 1980. Mussel Culture and Harvest: a North American Perspective.Elsevier Scientific Publishing Company, Amsterdam.
Lyman, R.L., 2006. Paleozoology in the service of conservation biology. Evol.Anthropol. 15, 11e19.
MacGinitie, G.E., 1941. On the method of feeding of four pelecypods. Biol. Bull. 80,18e25.
Mannino, M.A., Thomas, K.D., 2001. Intensive Mesolithic exploitation of coastalresources? Evidence from a shell deposit on the Isle of Portland (SouthernEngland) for the impact of human foraging on populations of intertidal rockyshore molluscs. J. Archaeol. Sci. 28, 1101e1114.
Mannino, M.A., Thomas, K.D., 2002. Depletion of a resource? the impact of pre-historic human foraging on intertidal mollusc communities and its significancefor human settlement, mobility and dispersal. World Archaeol. 33, 452e474.
Milner, N., Barrett, J., Welsh, J., 2007. Marine resource intensification in Viking AgeEurope: the molluscan evidence from Quoygrew, Orkney. J. Archaeol. Sci. 34,1461e1472.
Morrison, A.E., Cochrane, E.E., 2008. Investigating shellfish deposition and land-scape history at the Natia Beach Site, Fiji. J. Archaeol. Sci. 35, 2387e2399.
Morrison, A.E., Hunt, T.L., 2007. Human impacts on the nearshore environment: anarchaeological case study from Kaua'i, Hawaiian Islands. Pac. Sci. 61, 325e345.
Orchard, T.J., 2001. The Role of Selected Fish Species in Aleut Paleodiet (UnpublishedMasters thesis). Department of Anthropology, University of Victoria.
Pauly, D., 1995. Anecdotes and the shifting baseline syndrome of fisheries. TrendsEcol. Evol. 10, 430.

B. Campbell, T.J. Braje / Journal of Archaeological Science 58 (2015) 167e174174
Pauly, D., Christensen, V., Dalsgaard, J., Froese, R., Torres, F., 1998. Fishing downmarine food webs. Science 279, 860e863.
Phillips, P.T., 1988. California State Mussel Watch, Ten Year Data Summary 1977-1987. State Water Resource Control Board, Sacramento.
Redman, C.L., 2005. Resilience theory in archaeology. Am. Anthropol. 107, 70e77.Redman, C.L., 1999. Human Impacts on Ancient Ecosystems. University of Arizona
Press, Tucson.Reitz, E.J., Wing, E.S., 1999. Zooarchaeology. Cambridge University Press.Rick, T.C., 2007. The Archaeology and Historical Ecology of Late Holocene San
Miguel Island. Cotsen Institute of Archaeology, University of California, LosAngeles.
Rick, T.C., Lockwood, R., 2013. Integrating paleobiology, archaeology, and history toinform biological conservation. Conserv. Biol. 27, 45e54.
Rick, T.C., Erlandson, J.M. (Eds.), 2008. Human Impacts in Ancient Marine Ecosys-tems: a Global Perspective. University of California Press, Berkeley.
Ricker, W.E., 1973. Linear regressions in fishery research. J. Fish. Res. Board Can. 30,409e434.
Rudolph, J.L., 1990. Size variation among molluscs from sites near the Lower SantaYnez River. Proc. Soc. Calif. Archaeol. 3, 261e274.
Shaw, W.N., 1988. Species Profiles: Life Histories and Environmental Requirementsof Coastal Fishes and Invertabrates (Pacific Southwest): California Sea Musseland Bay Mussel. U.S. Fish Wildlife Service Biological Report 82.
Smith, J.R., Fong, P., Ambrose, R.F., 2008. The impacts of human visitation on musselbed communities along the California coast: are regulatory marine reserveseffective in protecting these communities? Environ. Manag. 41, 599e612.
Sokal, R.R., Rohlf, F.J., 1981. Biometry. Freeman Press, San Francisco.Stager, J.C., Chen, V., 1996. Fossil evidence of shell length decline in Queen Conch
(Strombus gigas L.) at Middleton Cay, Turks and Caicos, British West Indies.Caribb. J. Sci. 32, 14e20.
Stiner, M.C., Munro, N.D., Surovell, T.A., Tchernov, E., Bar-Yosef, O., 1999. Paleolithicpopulation growth pulses evidenced by small animal exploitation. Science 283,190e194.
Suchanek, T.H., 1986. Mussels and their role in structuring rocky shore commu-nities. In: Moore, P.G., Seed, R. (Eds.), The Ecology of Rocky Coasts. ColumbiaUniversity Press, New York, pp. 70e96.
Suchanek, T.H., 1981. The role of disturbance in the evolution of life history stra-tegies in the intertidal mussels Mytilus edulis and Mytilus californianus. Oeco-logia 50, 143e152.
Suchanek, T.H., 1979. The Mytilus Californianus Community: Studies on theComposition, Structure, and Organization, and Dynamics of a Mussel Bed (Ph.D.dissertation). Department of Biology, University of Washington.
Swetnam, T.W., Allen, C.D., Betancourt, J.L., 1999. Applied historical ecology: usingthe past to manage for the future. Ecol. Appl. 9, 1189e1206.
Tegner, M.J., Dayton, P.K., 2000. Ecosystem effects of fishing in kelp forest com-munities. ICES J. Mar. Sci. J. du Conseil 57, 579e589.
Whitaker, A.R., 2008. Incipient aquaculture in prehistoric California? Long-termproductivity and sustainability vs. immediate returns for the harvest of ma-rine invertebrates. J. Archaeol. Sci. 35, 1114e1123.
White, G., 1989. A Report of Archaeological Investigations at Eleven Native Amer-ican Coastal Sites, MacKerricher State Park, Mendocino County, California(Unpublished report on file with California State Parks).
Wolverton, S., Lyman, R.L. (Eds.), 2012. Conservation Biology and AppliedZooarchaeology. University of Arizona Press, Tucson.
Young, R.T., 1946. Spawning and settling season of the mussel Mytilus californianus.Ecology 27, 354e363.
Young, R.T., 1942. Spawning season of the California Mussel, Mytilus californianus.Ecology 23, 490e492.
Related Documents











