Publication of the International Union Against Cancer * Publication de I Union Internationale Contre le Cancer Int. J. Cancer: 56,522-528 (1994) 0 1994 Wiley-Liss, Inc. erbB-2 EXPRESSION IN ESTROGEN-RECEPTOR-POSITIVE BREAST-TUMOR CELLS IS REGULATED BY GROWTH-MODULATORY REAGENTS Daniela TAVERNA’, Susanna ANTONIOTTI~, Piera MAGGIORA~, Claudio DATI~, Michele DE BORTOL? and Nancy E. HYNES’J ’Friedrich Miescher Institute, P. 0. Box 2543, CH 4002 Basel, Switzerland; and 2Laboratoiy of Molecular Cell Biology, Department of Animal Biology, University of Turin, Via Acc. Albertina 17, 10123 Turin, Italy. it has previously been shown that, in the estrogen-receptor- positive breast-tumor cell lines T47D and ZR75. I, the erbB-2 protein and mRNA content are controlled negatively and positively by, respectively, estrogens and anti-estrogens. Since estrogens have a positive effect on cell proliferation, while anti-estrogens inhibit cell growth, the results suggested that there may be an inverse correlation between growth and erbB-2 expression. We have now examined this matter further. The effect of various growth-modulatory agents including estro- gen (E2), progesterone (Pg), retinoic acid (RA), epidermal growth factor (EGF), insulin (Ins), prolactin (Prl). I2-0- tetradecanolyl-phorbol-13-acetate (TPA) and dibutyryl-3‘:5’- cyclic-AMP (CAMP) on c-erbB-2 promoter activity, RNA and protein expression have been examined. The growth stimula- tors E2 and EGF both reduced the level of erbB-2 protein. However, while E2 clearly repressederbB-2 transcription, in the case of EGF, neither mRNA nor transcription were decreased. Of the agents which inhibit the growth of T47D and ZR75.1 cells-Pg, Prl, CAMP, RA and TPA-only Pg and CAMP caused an increase in the erbB-2 protein level. Pg and CAMP positively influenced c-erbB-2 promoter activity and RNA amount. TPA and RA also increased promoter activity but neither erbB-2 mRNA nor protein level was enhanced. The erbB-2 protein expression in cultures of T47D and ZR75.1 cells at different densities was also analyzed. Both the level of erbB-2 protein and c-erbB-2 promoter activity rose markedly in confluent cultures, suggesting a transcriptional mechanism of control. In conclusion, the data suggest that the effects of various agents on erbB-2 expression are complex and cannot be explained simply as reflecting the growth state of the cells. o 1994 Wiley-Liss, Inc. The c-erbB-2 gene encodes a member of the receptor tyrosine kinase family (Coussens et al., 1985). The gene is amplified in approximately 30% of primary human breast canccrs (Slamon et al., 1987) leading to over-expression of the erbB-2 protein. It has also been observed that high levels of the protein are present in cells which have no apparent gene alteration, suggesting that other mechanisms exist to control the amount of erbB-2 RNA and protein (Kraus et al., 1987). Since high levels of protein generally correlate with poor patient prognosis (Hynes, 1993 and references therein), a thorough understanding of the control of erbB-2 expression is important. Treatment of ER+ breast-tumor cells with estrogens, which enhances the growth of the cells, causes a decrease in erbB-2 RNA and protein expression (Dati et af., 1990). In contrast, treatment of the cells with the anti-estrogen tamoxifen leads to growth inhibition and an increase in erbB-2 RNA and protein (Antoniotti et al., 1992). This may represent an important point in selecting anti-estrogenic adjuvant therapies wherever amplification of erbB-2 accompanies ER-positivity (Wright et ul., 1992). In addition, we have previously shown that in rapidly growing cultures of normal mammary epithelial cells the level of erbB-2 protein is much reduced compared to confluent, “rcsting” cultures (Kornilova et al., 1992). These data sug- gested that there might be a correlation between the growth state of the cells and the level of erbB-2. Thus, we examined the effects of various agents and cell density on growth and on erbB-2 expression of the ER+ breast-tumor cell lines T47D and ZR75.1. c-erbB-2 promoter activity was analyzed and the erbB-2-specific RNA and protein were quantified. The effects of these agents on erbB-2 expression were complex and cannot be interpreted simply as reflecting the growth state of the cells. MATERIAL AND METHODS Cell culture and reagents The T47D and ZR75.1 human breast cancer cells were grown in complete medium (CM) consisting of DMEM, 510% FCS and 5 pg/ml Ins (Sigma, St. Louis, MO). For experiments in which a low proliferative activity was needed, cells were switched to stripped medium (SM) devoid of estrogenic activity, consisting of essential minimum Eagle’s medium without phenol red (which has estrogenic activity), supplemented with 5% FCS prc-absorbed on Dextran-coated charcoal (Dati et al., 1990). Culturing of T47D and ZR75.1 cells in this medium for 2-4 days reduced cell proliferation to a minimum. For the experiments where a complete absence of hormone was preferred, cells were switched from CM to serum-free medium (SFM) which contains DMEM + 1 mgiml BSA or 0.5 mgiml fetuin (Sigma). Culturing of T47D or ZR75.1 in this medium for 24 hr reduced the proliferation rate to a level comparable to that obtained within 3-4 days in SM. All media contained 2 mM L-glutamine and 50 pgiml gentami- cin. Cell proliferation was tested on “starved” cells as follows: 2 x lo5 cells/3.5 cm culture dish were plated out in CM, switched 24 hr later either to SFM plus 0.5 mg/ml fetuin or to SM and kept, respectively, for 1 or 4 days before addition of the various reagents. The cell number was determined 2-4 days later. The following growth-modulatory agents were tested: E2 (17 P-estradiol), Pg (progestin ORG 2058, Organon, Oss, The Netherlands), human or ovine Prl, Ins, EGF (mouse, from submaxillary glands), CAMP (dibutyryl-3’:5’-cyclic- AMP), TPA (12-0-tetradecanoyl-phorbol-13-acetate) and RA. Immunoblot analysis Cells were washed with cold PBS, collected and centrifuged at 2,OOOg for 2 min. The pellet was lysed in a buffer containing: 20 mM Tris pH 7.4, 0.1 M NaCI, 5 mM MgCI2, 1% Nonidet P40, 0.5% sodium deoxycholate, 0.1 mM 2-mercaptoethanol and 2 IU/ml Trasylol (Baxter, Duedingen, Switzerland). De- bris were removed by centrifugation at 800 g for 20 min, then 75 pg of protein extracts were separated by 8% SDS-PAGE, electroblotted onto nitrocellulose or PVDF membranes (Im- whom correspondence and reprint requests should be ad- dressed. Fax: 0041 61 6963835. Abbreviations: E2, estrogen; Pg, progesterone; hPrl, human prolac- tin; oPrl, ovine prolactin; EGF, epidermal growth factor; Ins, insulin; CAMP, dibutyryl-3‘:Sf-cyclic-AMP; TPA, 12-0-tetradecanoyl-phorbol- 13-acetate; RA, retinoic acid; FCS, fetal calf serum; CM, complete medium; SM, stripped medium; SFM, serum-free medium; SDS- PAGE, sodium dodecylsulfate-polycrylamide gel electrophoresis; GAPDH, glyceraldehyde phosphate dehydrogenase; ER+, estrogen- receptor positive; ECM, extracellular matrix. Received: June 18, 1993 and in revised form October 25, 1993.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Publication of the International Union Against Cancer * Publication de I Union Internationale Contre le Cancer Int. J. Cancer: 56,522-528 (1994) 0 1994 Wiley-Liss, Inc.
erbB-2 EXPRESSION IN ESTROGEN-RECEPTOR-POSITIVE BREAST-TUMOR CELLS IS REGULATED BY GROWTH-MODULATORY REAGENTS Daniela TAVERNA’, Susanna ANTONIOTTI~, Piera MAGGIORA~, Claudio DATI~, Michele DE BORTOL? and Nancy E. HYNES’J ’Friedrich Miescher Institute, P. 0. Box 2543, CH 4002 Basel, Switzerland; and 2Laboratoiy of Molecular Cell Biology, Department of Animal Biology, University of Turin, Via Acc. Albertina 17, 10123 Turin, Italy.
it has previously been shown that, in the estrogen-receptor- positive breast-tumor cell lines T47D and ZR75. I, the erbB-2 protein and mRNA content are controlled negatively and positively by, respectively, estrogens and anti-estrogens. Since estrogens have a positive effect on cell proliferation, while anti-estrogens inhibit cell growth, the results suggested that there may be an inverse correlation between growth and erbB-2 expression. We have now examined this matter further. The effect of various growth-modulatory agents including estro- gen (E2), progesterone (Pg), retinoic acid (RA), epidermal growth factor (EGF), insulin (Ins), prolactin (Prl). I2-0- tetradecanolyl-phorbol- 13-acetate (TPA) and dibutyryl-3‘:5’- cyclic-AMP (CAMP) on c-erbB-2 promoter activity, RNA and protein expression have been examined. The growth stimula- tors E2 and EGF both reduced the level of erbB-2 protein. However, while E2 clearly repressed erbB-2 transcription, in the case of EGF, neither mRNA nor transcription were decreased. Of the agents which inhibit the growth of T47D and ZR75.1 cells-Pg, Prl, CAMP, RA and TPA-only Pg and CAMP caused an increase in the erbB-2 protein level. Pg and CAMP positively influenced c-erbB-2 promoter activity and RNA amount. TPA and RA also increased promoter activity but neither erbB-2 mRNA nor protein level was enhanced. The erbB-2 protein expression in cultures of T47D and ZR75.1 cells at different densities was also analyzed. Both the level of erbB-2 protein and c-erbB-2 promoter activity rose markedly in confluent cultures, suggesting a transcriptional mechanism of control. In conclusion, the data suggest that the effects of various agents on erbB-2 expression are complex and cannot be explained simply as reflecting the growth state of the cells. o 1994 Wiley-Liss, Inc.
The c-erbB-2 gene encodes a member of the receptor tyrosine kinase family (Coussens et al., 1985). The gene is amplified in approximately 30% of primary human breast canccrs (Slamon et al., 1987) leading to over-expression of the erbB-2 protein. It has also been observed that high levels of the protein are present in cells which have no apparent gene alteration, suggesting that other mechanisms exist to control the amount of erbB-2 RNA and protein (Kraus et al., 1987). Since high levels of protein generally correlate with poor patient prognosis (Hynes, 1993 and references therein), a thorough understanding of the control of erbB-2 expression is important.
Treatment of ER+ breast-tumor cells with estrogens, which enhances the growth of the cells, causes a decrease in erbB-2 RNA and protein expression (Dati et af., 1990). In contrast, treatment of the cells with the anti-estrogen tamoxifen leads to growth inhibition and an increase in erbB-2 RNA and protein (Antoniotti et al., 1992). This may represent an important point in selecting anti-estrogenic adjuvant therapies wherever amplification of erbB-2 accompanies ER-positivity (Wright et ul., 1992). In addition, we have previously shown that in rapidly growing cultures of normal mammary epithelial cells the level of erbB-2 protein is much reduced compared to confluent, “rcsting” cultures (Kornilova et al., 1992). These data sug- gested that there might be a correlation between the growth state of the cells and the level of erbB-2. Thus, we examined the effects of various agents and cell density on growth and on erbB-2 expression of the ER+ breast-tumor cell lines T47D and ZR75.1. c-erbB-2 promoter activity was analyzed and the
erbB-2-specific RNA and protein were quantified. The effects of these agents on erbB-2 expression were complex and cannot be interpreted simply as reflecting the growth state of the cells.
MATERIAL AND METHODS Cell culture and reagents
The T47D and ZR75.1 human breast cancer cells were grown in complete medium (CM) consisting of DMEM, 5 1 0 % FCS and 5 pg/ml Ins (Sigma, St. Louis, MO). For experiments in which a low proliferative activity was needed, cells were switched to stripped medium (SM) devoid of estrogenic activity, consisting of essential minimum Eagle’s medium without phenol red (which has estrogenic activity), supplemented with 5% FCS prc-absorbed on Dextran-coated charcoal (Dati et al., 1990). Culturing of T47D and ZR75.1 cells in this medium for 2-4 days reduced cell proliferation to a minimum. For the experiments where a complete absence of hormone was preferred, cells were switched from CM to serum-free medium (SFM) which contains DMEM + 1 mgiml BSA or 0.5 mgiml fetuin (Sigma). Culturing of T47D or ZR75.1 in this medium for 24 hr reduced the proliferation rate to a level comparable to that obtained within 3-4 days in SM. All media contained 2 mM L-glutamine and 50 pgiml gentami- cin. Cell proliferation was tested on “starved” cells as follows: 2 x lo5 cells/3.5 cm culture dish were plated out in CM, switched 24 hr later either to SFM plus 0.5 mg/ml fetuin or to SM and kept, respectively, for 1 or 4 days before addition of the various reagents. The cell number was determined 2-4 days later. The following growth-modulatory agents were tested: E2 (17 P-estradiol), Pg (progestin ORG 2058, Organon, Oss, The Netherlands), human or ovine Prl, Ins, EGF (mouse, from submaxillary glands), CAMP (dibutyryl-3’:5’-cyclic- AMP), TPA (12-0-tetradecanoyl-phorbol-13-acetate) and RA.
Immunoblot analysis Cells were washed with cold PBS, collected and centrifuged
at 2,OOOg for 2 min. The pellet was lysed in a buffer containing: 20 mM Tris pH 7.4, 0.1 M NaCI, 5 mM MgCI2, 1% Nonidet P40, 0.5% sodium deoxycholate, 0.1 mM 2-mercaptoethanol and 2 IU/ml Trasylol (Baxter, Duedingen, Switzerland). De- bris were removed by centrifugation at 800 g for 20 min, then 75 pg of protein extracts were separated by 8% SDS-PAGE, electroblotted onto nitrocellulose or PVDF membranes (Im-
whom correspondence and reprint requests should be ad- dressed. Fax: 0041 61 6963835.
Abbreviations: E2, estrogen; Pg, progesterone; hPrl, human prolac- tin; oPrl, ovine prolactin; EGF, epidermal growth factor; Ins, insulin; CAMP, dibutyryl-3‘:Sf-cyclic-AMP; TPA, 12-0-tetradecanoyl-phorbol- 13-acetate; RA, retinoic acid; FCS, fetal calf serum; CM, complete medium; SM, stripped medium; SFM, serum-free medium; SDS- PAGE, sodium dodecylsulfate-polycrylamide gel electrophoresis; GAPDH, glyceraldehyde phosphate dehydrogenase; ER+, estrogen- receptor positive; ECM, extracellular matrix.
Received: June 18, 1993 and in revised form October 25, 1993.

TRANSCRIPTIONAL REGULATION OF c-erbB-2 523
mobilon, Millipore, Bedford, MA) and the erbB-2 protein was detected using 21N antiserum followed by treatment with '2sI-labeled protein A (Hynes et al., 1989).
RNA extraction and Northem-blot analysis Total RNA was extracted from cell pellets by the guanidine-
lithium chloride or acidic phenol procedures (Dati et al., 1990; Chomczynski and Sacchi, 1987). Aliquots of 20 p,g were separated on 1.2% agarose-formaldehyde gels and transferred to a Hybond-N membrane (Amersham, Aylesbury, UK) then the c-erbB-2 specific transcripts were detected with a 1.1-kb Bam HI cDNA fragment from pCER204 (Antoniotti et al., 1992). RNA loading was controlled by ethidium bromide staining of the gel and by rehybridizing the filters either to a p-actin or to a GAPDH probe (Dati et al., 1990; Antoniotti et al., 1992).
Construction of the c-erbB-2 promoter-CA Tplasmid For a functional analysis of the c-erbB-2 promoter, 1216 bp
5' of the transcription start site of the c-erbB-2 gene (Hudson ef al., 1990) were amplified from genomic DNA by polymerase chain reaction (PCR) technology and cloned into the vector pBLCAT3. The forward primer (5' AGAACTGCCAAATT- TAGCAC 3') corresponded to the sequence -1391 to -1372 5' of the c-erbB-2 translation start site and possessed an additional Xba I sequence at its 5' end. The reverse primer (5' GAATGAAGTTGTGAAGCTGAGATT 3') corresponded to the sequence -199 to -175 5' of the c-erbB-2 translation start site and contained additional Hind I11 and Xho-I restriction sites at thc 3' end. Template DNA was from HBLlOO normal human breast cells. The PCR amplification was performed for 30 cycles of 1 rnin at 94"C, 2 rnin at 55°C and 1 rnin at 72"C, followed by an extension step at 72°C for 5 min. In order to rule out effects arising from PCR-induced mutations, 3 indepen- dent reactions were performed. The 3 amplified products were gel-purified, cleaved with Xba I and Xho I, ligated into the Xba-I - Xho-I-digested pBLCAT3 to yield the plasmids perhB-2CAT1, -2 and -3.
Transfections To obtain stable transfectants, the calcium phosphate pre-
cipitation technique was used. T47D cells (1-2 x lo6) were transfected in CM with 0.5 pg of pSV2neo and 10 pg of the promoter-CAT expression plasmids. The plasmids pRSVCAT, pSV2CAT and perbB-2CAT1, -2 and -3 were used. The transfected cells were selected in CM containing 800 kg/ml G418, then 20-30 resistant colonies were pooled and treated with various agents and CAT assays were performed. For transient transfections 1 X 106 T47D cells were transfected with 10 pg of the promoter-CAT plasmids using the DEAE- dextran technique followed by a DMSO shock.
CAT assays Extracts used to determine CAT activity were obtained from
pellets of T47D cells prepared by centrifugation in cold PBS at 1,200 g for 2 min at 4°C. Pellets were resuspended in 0.25 M Tris-HCI pH 7.8, then frozen in liquid nitrogen and thawed at 40°C 3 times. Following centrifugation at 13,000 g for 5 min, the supernatant was heated for 10 rnin at 60°C. Denatured protein was pelleted at 10,000 g for 5 rnin and samples of the supernatant were taken for a determination of the protein concentration. CAT enzyme activity was determined as nmolesi min/mg of protein.
RESULTS Growth of ER+ breast-tumor cells in the presence of various growth-modulatory agents
An attempt was made to correlate the growth state of the ER+ breast-tumor cell lines T47D and ZR75.1 with the expression of erbB-2. The cells were treated with hormones
and peptide factors known to modulate growth and differentia- tion of normal breast cells and breast tumor cells. In addition to E2, the agents tested included RA, the steroid hormone Pg, the peptide factors Ins and EGF, which bind receptor tyrosine kinases, Prl which activates a cytokine receptor, finally TPA, and CAMP, both activators of cytoplasmic ser/thr kinases. The results are shown in Figure 1. In panel a the cells were grown in complete medium (CM) and switched to hormone-deprived medium (SM) 4 days before treatment was begun. The results show that cell growth was enhanced by E2 and EGF, moder- ately stimulated by Ins and inhibited by Pg and Prl. In panel b the cells were switched to SFM 1 day before treatment was begun. The results show that CAMP, TPA and RA, agents which have been shown to inhibit growth of other breast-tumor cell lines (Bacus et al., 1990) caused a decrease in T47D cell growth. Similar results were obtained with ZR75.1 cells (data not shown).
erbB-2protein and mRNA level in breast-cancer cells treated with growth-modulatory agents
The level of erbB-2 protein in the breast-tumor cells treated with E2, Pg, Prl, EGF and Ins was determined by a protein blotting analysis using the 21N erbB-2-specific serum. The results are shown in Figure 2a and b for, respectively, T47D and ZR75.1 cells. Compared to control untreated cells, treat- ment with E2 and EGF led to a reduction in the level of erbB-2 protein. Pg-treated cells contained elevated amounts of erbB-2. Prl and Ins treatment did not affect erbB-2 protein expression in either cell line. The level of erbB-2 protein in, respectively, T47D and ZR75.1 cells treated with RA, TPA and CAMP is shown in Figure 2, d. RA had no significant effect on the level of erbB-2 protein after 2 days of treatment (Fig. 2) but after longer times the level decreased slightly (data not shown). TPA treatment led to an electrophoretic mobility shift in the erbB-2 protein and lowered its amount in ZR75.1 cells. In both cell lines there was a dose-dependent increase in the erbB-2 protein level following CAMP treatment.
A Northern-blot analysis on total RNA isolated from ZR75.1 cells was performed to determine if the observed variations in the amount of erbB-2 protein reflect changes in RNA content. The results are shown in Figure 3. Panel (a)
A
Cell number (x 1000) - _. 800 ,
700
600
500
400
300
200
C P r l EGF €2 Pg Ins
B
Cell number (x 1000) 500
400
300
200
C EGF cAMP TPA RA
FIGURE 1 - Effects of different treatments on breast-cancer cell proliferation. T47D cells (2 X lo5) were plated out in CM and switched 24 hr later either to SM (a) or to SFM (b). After 1 (b) or 4 (a) days the cells were treated with Ins (10 pg/rnl), E2 (10 nM 17-p estradiol), EGF (100 ngiml), Pg (10 nM progestin ORG2058), hPrl (2.50 ngiml), cAMP (1 mM), TPA (100 ngiml) or RA (10 pM). The control cells were left untreated (C). The cells were counted at day 3 (a) or 4 (b) of treatment. Each column represents the average cell number of 2 wells and the standard deviation is indicated.

5 24 TAVERNA ETAL.
FIGURE 2 - Western-blot analysis of the erbB-2 protein in breast-cancer cells. Sparse cultures of T47D cells (u, c) or ZR 75.1 cells (b, d) were grown for 1 (c, d) or 4 (a, h) days In SM (a, b, d) or SFM (cf, then treated for 2 or 4 days with the same medium containing El; (10 nM 17-@-estradiol), Pg (10 nM progestin ORG2058), hPrl(250 ngiml), EGF (100 ng/ml), Ins (10 mg/ml), cAMP (0.1 or 1 mM), TPA (10 or 100 ngiml) and RA (10 FM). Control cells were left untreated (C). Cell lysates were prepared, separated on 8% SDS-PAGE and electroblotted, and the erbB-2 protein (p185) was detected using the 21N antiserum followed by (lZsI) protein-A treatment. Each lane contains equal amounts of protein extracts. The lane marked SKBR3 contains protein extract from SKBR3 cells which over-express the erbB-2 protein and are here used as a control for the 21N serum.
FIGURE 3 - Northern-blot analysis of the erbB-2 protein in breast-cancer cells. Sparse cultures of ZR 75.1 cells (u, b) were grown for 3 days in SM then treated for 2 days with the same medium containing E2 (10 nM 17-@-estradiol), Pg (10 nM proges- tin ORG2058), hPrl (250 ngiml), EGF (100 ngiml), Ins (10 mgiml), cAMP (0.1 or 1 mM), TPA (100 ng/ml) and RA (10 pM). Control cells were left untreated (C). Total RNA was prepared and 20 p,g were fractionated on a denaturing formaldehyde gel, blotted on a nylon membrane and hybridized to a 32P-labeled c-erbB-2-specific probe. The lower panel shows the results of hybridizing the same filter with a p-actin (a) or a GAPDH (b) probe to control for RNA loading.
lane 2 shows that the level of erbB-2 mRNA decreased following E2 treatment and increased aftcr Pg treatment of the cells. EGF, Ins or Prl treatment did not affect the level of erbB-2 specific mRNA (lanes 3-6). To control for R N A
loading, the same filter was hybridized with a p-actin probe. Similar results were obtained using the T47D cells (data not shown). Figure 3b shows the erbB-2 specific mRNA level in ZR75.1 cells treated with CAMP, TPA and RA. CAMP-treated ZR75.1 cells contained elevated levels of erbB-2 specific mRNA and protein (Fig. 2). Neither TPA nor R A had any significant effect upon the erbB-2 mRNA level.
Construction and expression of a c-erbB-2 promoter-CA T construct
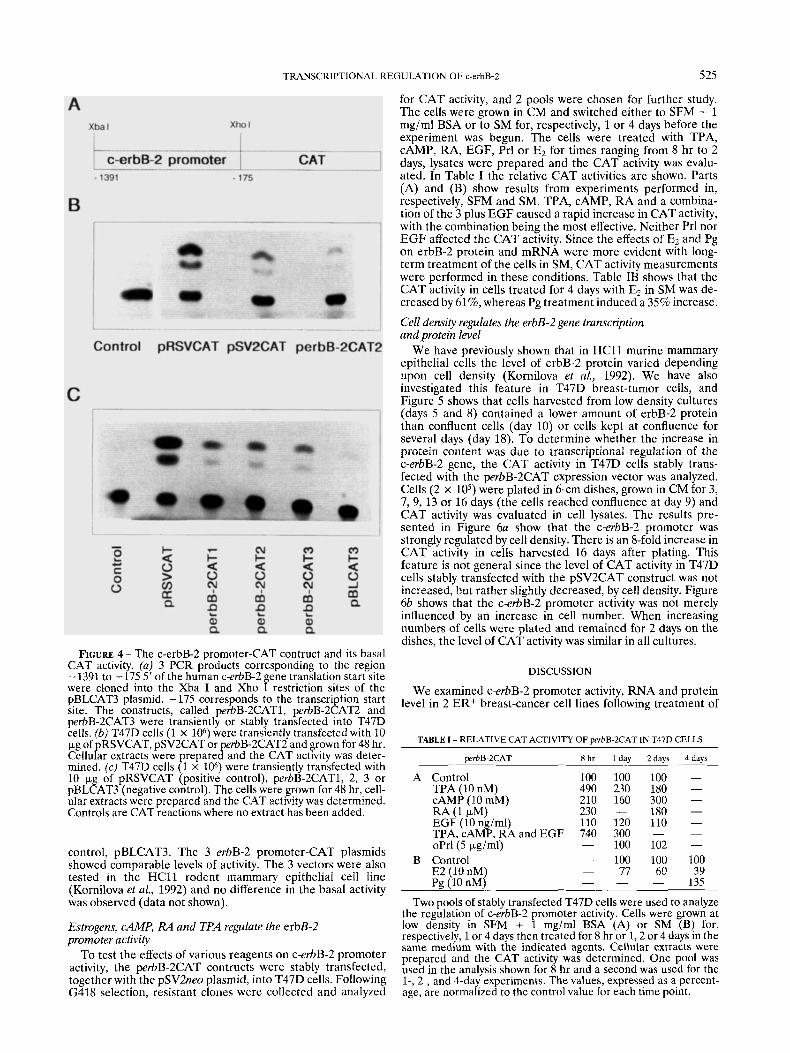
In order to analyze c-erbB-2 promoter activity, a 1216-bp fragment corresponding to the 5’ promoter region of the c-erbB-2 gene was obtained by PCR technology. This fragment corresponds to the sequence - 1391 to - 175 upstream of the erbB-2 translation start site, as described by Hudson et al. (1990). -175 corresponds to the transcription start site. The amplified products from 3 individual PCR reactions were inserted into the pBLCAT3 plasmid as shown in Figure 4u. Thc c-erbB-2 promoter activity of the perbB-2CAT plasmids, denoted 1-2-3, was assayed following transient transfection into the T47D cells. As controls the pRSVCAT and the pSV2CAT were also transfected. Forty-eight hours later, the cells were lysed and the basal activity of the different promot- ers was analyzed. Figure 4b shows that the CAT activity was low in the perbB-2CAT2-containing cells compared to the activity from the cells transfected with the pRSVCAT and the pSVCAT, suggesting that the erbB-2 promoter is weaker than the other 2 promoters.
In order to exclude variations in activity between the 3 perbB-2CAT constructs due to PCR-induced mutations, we compared the activity of the 3 independent plasmids in 2 different sets of transiently transfected T47D cells. Figure 4c shows the CAT activity of perbB-2CAT1, -2 and -3 in compari- son to the positive control, pRSVCAT and the negative
TRANSCRIPTIONAL REGULATION OF c-erbB-2 525
for CAT activity, and 2 pools were chosen for further study. The cells were grown in CM and switched either to SFM + 1 mg/ml BSA or to SM for, respectively, 1 or 4 days before the experiment was begun. The cells were treated with TPA, CAMP, RA, EGF, Prl o r E2 for times ranging from 8 hr to 2 days, lysates were prepared and the CAT activity was evalu- ated. In Table I the relative CAT activities are shown. Parts (A) and (B) show results from experiments performed in, respectively, SFM and SM. TPA, CAMP, RA and a combina- tion of the 3 plus EGF caused a rapid increase in CAT activity, with the combination being the most effective. Neither Prl nor E G F affected the CAT activity. Since the effects of E2 and Pg on erbB-2 protein and mRNA were more evident with long- term treatment of the cells in SM, CAT activity measurements were performed in these conditions. Table IB shows that the CAT activity in cells treated for 4 days with E2 in SM was de- creased by 61%, whereas Pg treatment induced a 35% increase.
Cell density regulates the erbB-2 gene transcription and protein level
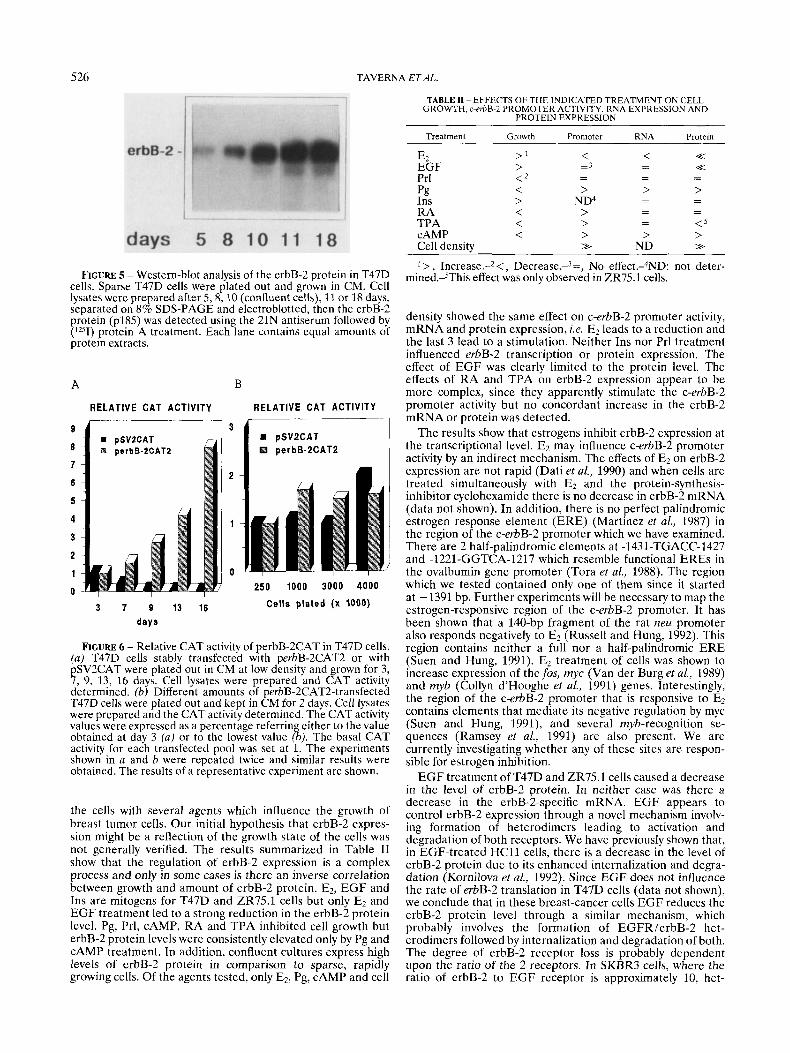
We have previously shown that in H C l l murine mammary epithelial cells the level of erbB-2 protein varied depending upon cell density (Kornilova et al., 1992). We have also investigated this feature in T47D breast-tumor cells, and Figure 5 shows that cells harvested from low-density cultures (days 5 and 8) contained a lower amount of erbB-2 protein than confluent cells (day 10) or cells kept at confluence for several days (day 18). To determine whether the increase in protein content was due to transcriptional regulation of the c-erbB-2 gene, the CAT activity in T47D cells stably trans- fected with the perbB-2CAT expression vector was analyzed. Cells (2 x lo5) were plated in 6-cm dishes, grown in CM for 3, 7 ,9 , 13 or 16 days (the cells reached confluence at day 9) and CAT activity was evaluated in cell lysates. The results pre- sented in Figure 6a show that the c-erbB-2 promoter was strongly regulated by cell density. There is an 8-fold increase in CAT activity in cells harvested 16 days after plating. This feature is not general since the level of CAT activity in T47D cells stably transfected with the pSV2CAT construct was not increased, but rather slightly decreased, by cell density. Figure 6b shows that the c-erbB-2 promoter activity was not merely influenced by an increase in cell number. When increasing numbers of cells were plated and remained for 2 days on the dishes, the level of CAT activity was similar in all cultures.
FIGURE 4 - The c-erbB-2 promoter-CAT contruct and its basal CAT activity. (a) 3 PCR products corresponding to the region - 1391 to -175 5’ of the human c-erbB-2 gene translation start site were cloned into the Xba I and Xho I restriction sites of the pBLCAT3 plasmid. - 175 corresponds to the transcription start site. The constructs, called perbB-2CAT1, perbB-2CAT2 and perbB-2CAT3 were transiently or stably transfected into T47D cells. fb) T47D cells (1 x loh) were transiently transfected with 10 pg of pRSVCAT, pSV2CAT or perbB-2CAT2 and grown for 48 hr. Cellular extracts were repared and the CAT activity was deter- mined. (c) T47D cells 8 x 106) were transiently transfected with 10 pg of pRSVCAT (positive control), perbB-2CAT1, 2, 3 or pBLCAT3 (negative control). The cells were grown for 48 hr, cell- ular extracts were prepared and the CAT activity was determined. Controls are CAT reactions where no extract has been added.
control, pBLCAT3. The 3 erbB-2 promoter-CAT plasmids showed comparable levels of activity. The 3 vectors were also tested in the HCll rodent mammary epithelial cell line (Kornilova et al., 1992) and no difference in the basal activity was observed (data not shown).
Estrogens, CAMP, RA and TPA regulate the erbB-2 promoter activity
To test the effects of various reagents on c-erbB-2 promoter activity, the perbB-2CAT contructs were stably transfected, together with the pSV2neo plasmid, into T47D cells. Following G418 selection, resistant clones were collected and analyzed
DISCUSSION
We examined c-erbB-2 promoter activity, RNA and protein level in 2 ER+ breast-cancer cell lines following treatment of
TABLE I - RELATIVE CAT ACTIVITY OF perbB-2CAT IN T47D CELLS
wrbB-2CAT 8 h r 1 day 2davs 4days
Control TPA (10 nM) CAMP (10 mM) RA (1 uM)
100 100 100 - 490 230 180 - 210 160 300 - 230 - 180 -
EGF (io ng/ml) 110 120 110 - TPA,cAMP,RAandEGF 740 300 - - oPrl(5 pgiml) - 100 102 - Control - 100 100 100 E2 (10 nM) - 77 60 39
- 135 Pg (10 nM) _ - Two pools of stably transfected T47D cells were used to analyze
the regulation of c-erbB-2 promoter activity. Cells were grown at low density in SFM + 1 mg/ml BSA (A) or SM (B) for, respectively, 1 or 4 days then treated for 8 hr or 1 ,2 or 4 days in the same medium with the indicated agents. Cellular extracts were prepared and the CAT activity was determined. One pool was used in the analysis shown for 8 hr and a second was used for the 1-, 2-, and 4-day experiments. The values, expressed as a percent- age, are normalized to the control value for each time point.
526 TAVERNA ETAL.
TABLE I1 - EFFECTS OF THE INDICATED TREATMENT ON CELL GROWTH, c-erbB-2 PROMOTER ACTIVITY, RNA EXPRESSION AND
PROTEIN EXPRESSION
FIGURE 5 - Western-blot analysis of the erbB-2 protein in T47D cells. Sparse T47D cells were plated out and grown in CM. Cell lysates were prepared after 5,8 , 10 (confluent cells), 11 or 18 days, separated on 8% SDS-PAGE and electroblotted, then the erbB-2 protein (p185) was detected using the 21N antiserum followed by (lZ5I) protein A treatment. Each lane contains equal amounts of protein extracts.
A
RELATIVE CAT ACTIVITY
B
RELATIVE CAT ACTIVITY
3 7 9 13 16 days
D pSV2CAT perbB-2CAT2
3 1
250 1000 3000 4000 Cells plated (x 1000)
FIGURE 6 - Relative CAT activity of perbB-2CAT in T47D cells. (a) T47D cells stably transfected with perbB-2CAT2 or with pSV2CAT were plated out in CM at low density and grown for 3, 7, 9, 13, 16 days. Cell lysates were prepared and CAT activity determined. (b) Different amounts of perbB-2CAT2-transfected T47D cells were plated out and kept in CM for 2 days. Cell lysates were prepared and the CAT activity determined. The CAT activity values were expressed as a percentage referring either to the value obtained at day 3 (a) or to the lowest value (b). The basal CAT activity for each transfected pool was set at 1. The experiments shown in a and b were repeated twice and similar results were obtained. The results of a representative experiment are shown.
the cells with several agents which influence the growth of breast tumor cells. Our initial hypothesis that erbB-2 expres- sion might be a reflection of the growth state of the cells was not generally verified. The results summarized in Table I1 show that the regulation of erbB-2 expression is a complex process and only in some cases is there an inverse correlation between growth and amount of erbB-2 protein. E2, E G F and Ins are mitogens for T47D and ZR7.5.1 cells but only E2 and E G F treatment led to a strong reduction in the erbB-2 protein level. Pg, Prl, CAMP, R A and TPA inhibited cell growth but erbB-2 protein levels were consistently elevated only by Pg and cAMP treatment. In addition, confluent cultures express high levels of erbB-2 protein in comparison to sparse, rapidly growing cells. Of the agents tested, only E2, Pg, cAMP and cell
Treatment
EZ EGF Prl pg Ins RA TPA CAMP Cell density
Growth
>' > < 2 < > < < <
Promoter
< = 3
> ND4 > > > >>
- -
~~
Protein
<< <<
> - -
- - - - < 5 > >>
> , Increase.-2 <, Decrease.-.'=, No effect.sND: not deter- mined.-SThis effect was only observed in ZR75.1 cells.
density showed the same effect on c-erbB-2 promoter activity, mRNA and protein expression, i.e. EZ leads to a reduction and the last 3 lead to a stimulation. Neither Ins nor Prl treatment influenced erbB-2 transcription or protein expression. The effect of E G F was clearly limited to the protein level. The effects of R A and TPA on erbB-2 expression appear to be more complex, since they apparently stimulate the c-erbB-2 promoter activity but no concordant increase in the erbB-2 mRNA or protein was detected.
The results show that estrogens inhibit erbB-2 expression at the transcriptional level. E2 may influence c-erbB-2 promoter activity by an indirect mechanism. The effects of Ez on erbB-2 expression are not rapid (Dati et al., 1990) and when cells are treated simultaneously with E2 and the protein-synthesis- inhibitor cyclohexamide there is no decrease in erbB-2 mRNA (data not shown). In addition, there is no perfect palindromic estrogen response element (ERE) (Martinez et al., 1987) in the region of the c-erbB-2 promoter which we have examined. There are 2 half-palindromic elements at -1431-TGACC-1427 and -1221-GGTCA-1217 which resemble functional EREs in the ovalbumin gene promoter (Tora et a!., 1988). The region which we tested contained only one of them since it started at -1391 bp. Further experiments will be necessary to map thc estrogen-responsive region of the c-erbB-2 promoter. It has been shown that a 140-bp fragment of the rat m u promoter also responds negatively to E2 (Russell and Hung, 1992). This region contains neither a full nor a half-palindromic E R E (Suen and Hung, 1991). E2 treatment of cclls was shown to increase expression of thefos, myc (Van der Burg et al., 1989) and myb (Collyn d'Hooghe et al., 1991) genes. Interestingly, the region of the c-erbB-2 promoter that is responsive to E2 contains elements that mediate it5 negative regulation by myc (Suen and Hung, 1991), and several myh-recognition sc- quences (Ramsey et al., 1991) are a150 present. We are currently investigating whether any of these sites are respon- sible for estrogen inhibition.
E G F treatment of T47D and ZR75.1 cells caused a decrease in the level of erbB-2 protein. In neither case was there a decrease in the erbB-2-specific mRNA. E G F appears to control erbB-2 expression through a novel mechanism involv- ing formation of heterodimers leading to activation and degradation of both receptors. We have previously shown that, in EGF-treated HCll cells, there is a decrease in the level of erbB-2 protein due to its enhanced internalization and degra- dation (Kornilova eta!., 1992). Since EGF does not influcnce the rate of erbB-2 translation in T47D cells (data not shown), we conclude that in these breast-cancer cells E G F reduces the erbB-2 protein level through a similar mechanism, which probably involves the formation of EGFR/erbB-2 het- erodimers followcd by internalization and degradation of both. The degree of erbB-2 receptor loss is probably dependent upon the ratio of the 2 receptors. In SKBR3 cells, where the ratio of erbB-2 to E G F receptor is approximately 10, het-

TRANSCRIPTIONAL REGULATION OF c-erbB-2 527
erodimers can be detected (Goldman et al., 1990) but we have not observed a decrease in erbB-2 receptor level (data not shown). In contrast, in cells with approximately equal numbers of both receptors the EGF-induced decrease in erbB-2 recep- tor is more evident (data not shown). This might explain the different extent of erbB-2 reduction in the 2 cell lines which we examined.
The addition of Pg, CAMP, RA and TPA to the T47D cells resulted in a decrease in cell growth. These agents increased the erbB-2 promoter activity to variable extents, but in the case of TPA and RA this was not accompanied by an increase in mRNA and protein levels. The 1.2-kb c-erbB-2 promoter fragment which we analyzed does not contain a classical Pg response element. Pg very often displays effects on breast cancer cells that are defined as anti-estrogenic and it is possible that Pg acts, with opposite effects, on the same factor(s) that mediate the estrogen effect on the c-erbB-2 promoter. Transcriptional activation of the c-erbB-2 promoter by RA, TPA and cAMP is consistent with results obtained in HeLa cells (Hudson et al., 1990). The additive effect of RA, TPA, cAMP and EGF in combination, previously observed in Hela cells (Hudson et al., 1990), was also observed in T47D cells. The promoter fragment which we have analyzed contains neither a cAMP response element (Montminy et al., 1986) nor a TPA response element (Angel et al., 1987). It is possible that cAMP and TPA act via the AP2 sites which are present in the c-erbB-2 promoter (Masayoshi et al., 1987). The c-erbB-2 promoter also displays no classical RA receptor response element (RARE) although direct repeats of an ERE-related half-palindrome, which are present only once in the erbB-2 promoter, have been reported to serve as a RARE (de-ThC et al., 1990). The effects of TPA and RA appear to be quite complex. Despite the fact that these agents increased the c-erbB-2 promoter activity, there was no concomitant increase in erbB-2 mRNA in TPA- and RA-treated ZR75.1 cells (shown here) or in TPA-treated MDA-MB235 or MCF7 cells (D. Fabbro, personal communication). The most straightfor- ward explanation is that there is an increase in the rate of degradation of erbB-2 mRNA following TPA or RA treat- ment. It has previously been shown that RA and PMA, like TPA, an activator of PKC, cause growth inhibition of 2 other breast-tumor cell lines. This is accompanied by a loss of cell-surface erbB-2, a transient increase in cytoplasmic protein but no effect on the overall quantity of erbB-2 protein (Bacus et al., 1990). RA did not markedly alter the amount of erbB-2 protein in T47D and ZR75.1 cells, but in ZR75.1 cells its level was reduced by TPA treatment.
The expression of erbB-2 protein was also strongly regulated by the culture density of T47D (shown here) and ZR75.1 (data not shown) cells, probably reflecting increased mRNA levels since the c-erbB-2 promoter activity was also strongly en- hanced. We have previously observed that the level of erbB-2 protein is elevated in confluent cultures of the normal mam- mary epithelial cell lines H C l l and 31E. The mechanism leading to elevated erbB-2 levels is different in these normal cells, since we observed an enhancement of erbB-2 protein stability and no changes in the amount of erbB-2 mRNA (Kornilova et al., 1992). The human c-erbB-2 promoter-CAT constructs were also tested in H e l l cells and cell density did not increase the CAT activity (not shown). We have shown that the increase in c-erbB-2 promoter activity observed in T47D cells is dependent on time of culture and not on cell number; therefore, we speculate that it is probably due to specific extracellular matrix (ECM) protein deposition. It has been observed that the p-casein and TGF-P promoters are also affected by ECM (Streuli et al., 1993; Schmidhauser et al., 1992). By using T47D cells transfected with the pSV2CAT construct, we proved that the increase in CAT activity is not a generalized occurrence. Indeed, we observed a slight decrease in CAT activity driven by the SV40. The reason for this decrease is unknown but may be due, for example, to a general decrease in CAT protein stability.
We have shown that erbB-2 expression is controlled at the transcriptional or post-transcriptional level by many factors, including growth regulators and cell density. c-erbB-2 gene amplification is one of the most consistent alterations found in human adenocarcinomas arising at sites including breast, ovary, lung, stomach and salivary gland. In addition, high levels of erbB-2 protein have been correlated with poor patient prognosis (Hynes, 1993 and references therein) making it important to understand the mechanisms which control expres- sion of this protein.
ACKNOWLEDGEMENTS
We thank Drs. D. Fabbro (Ciba-Geigy, Basel) and G. Merlo (Friedrich Miescher Institute) and Ms. B. Marte (Friedrich Miescher Institute) for their critical comments on the manu- script. The excellent technical assistance of Ms. D. Graus is acknowledged. D.T. is a recipient of a fellowship from the Swiss Cancer League. S.A. and P.M. are recipients of fellow- ships from the Associazione Italiana per la Ricerca sul Cancro (AIRC). This work was supported in part by a grant from the AIRC.
REFERENCES
ANGEL, P., IMAGAWA, M., CHIU, R., STEIN, B., IMBRA, R.J., RAHMS- DORF, H.J., JONAT, C., HERRLICH, P. and KARIN, M., Phorbol ester- inducible genes contain a common cis element recognized by a TPA-modulated trans-acting factor. Cell, 49,729-739 (1987). ANTONIOrrI, S., MAGGIORA, P., DATI, C. and DE BORTOLI, M., Tamoxifen up-regulates c-erbB-2 expression in oestrogen-responsive breast cancer cells in vitro. Europ. J. Cancer, 28,318-321 (1992). BACUS, S.S., KIGUCHI, K., CHIN, D., KING, C.R. and HUBERMAN, E., Differentiation of cultured human breast cancer cells (AU-565 and MCF7) associated with loss of cell surface HER-2ineu antigen. Mol. Carcinogen.,3,350-362 (1990). CHOMCZYNSKI, P. and SACCHI, N., Single-step method of RNA isolation by the acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Biochem., 162,156-159 (1987). COLLYN-D’HOOGHE, M., VANDEWALLE, B., HORNEZ, L., LANTOINE, D., REVILLION, F., LEFEBVRE, J. and KERCKAERT, J.P., C-myc overexpres- sion, c-mil, c-myb expression in a breast tumor cell line. Effects of estrogen and antiestrogen. Anticancer Rex, 11,2175-2180 (1991). COUSSENS, L., YANG-FENG, T.L., LIAO, Y.-C., CHEN, E., GRAY, A,, MCGRATH, J., SEEBURG, P.H., LIBERMANN, T.A., SCHLESSINGER, J., FRANCE, U., LEVINSON, A. and ULLRICH, A., Tyrosine kinase recep-
tor with extensive homology to EGF receptor shares chromosomal location with neu oncogene. Science, 230,1132-1139 (1985). DATI, C., ANTONIOTTI, S., TAVERNA, D., PERROTEAU, I. and DE BORTOLI, M., Inhibition of c-erbB-2 oncogene expression by estrogens in human breast cancer cells. Oncogene, 5,1001-1006 (1990). DE-THE, H., VIVANCO-RUIZ, M.D.M., TIOLLAIS, P., STUNNENBERG, H. and DEJEAN, A,, Identification of a retinoic acid responsive element in the retinoic acid receptor b gene. Nature (Lond.), 343,177-180 (1990). GOLDMAN, R., BEN LEVY, R., PELES, E. and YARDEN, Y., Heterodimer- ization of the erbB-1 and erbB-2 receptors in human carcinoma cells: a mechanism for receptor transregulation. Biochemistry, 29, 11024- 11028 (1990). HUDSON, L.G., ERTI., A.P. and GILL, G.N., Structure and inducible regulation of the human c-erbB2ineu promoter. J. biol. Chem., 265, 4389-4393 (1990). HYNES, N.E., Amplification and overexpression of the erbB-2 gene in human tumors: its involvement in tumor development, significance as a prognostic factor, and potential as a target for cancer therapy. Semin. Cancer B i d , 4,19-26 (1993). HYNES, N.E., GERBER, H.A., SAURER, S. and GRONER, B., Overexpres- sion of the c-erbB-2 protein in human breast tumor cell lines. J. cell. Biochem., 39, 167-173 (1989).

528 TAVERNA ETAL.
KORNILOVA, E.S., TAVERNA, D., HOECK, W. and HYNES, N.E., Surface expression of erbB-2 protein is post-transcriptionally regulated in mammary epithelial cells by epidermal growth factor and by the culture density. Oncogene, 7,511-519 (1992). KRAUS, M.H., POPESCU, N.C., AMSBAUGH, S.C. and KING, C.R., Overexpression of the EGF receptor-related proto-oncogene erbB-2 in human mammary tumor cell lines by different molecular mechanisms.
MARTINEZ, E., GIVEL, F. and WAHLI, W., The estrogen-responsive element as an inducible enhancer: DNA sequence requirements and conversion to a glucocorticoid-responsive element. EMBO J., 6,
MASAYOSHI, I., CHIU, R. and KARIN, M., Transcription factor AP-2 mediates induction by two different signal-transduction pathways: protein kinase C and CAMP. Cell, 51,251-260 (1987). MONTMINY, M.R., SEVARINO, K.A., WAGNER, J.A., MANDEL, G. and GOODMAN, R.H., Identification of a cyclic AMP responsive element within the rat somatostatin gene. Proc. nat. Acad. Sci. (Wash.), 83,
RAMSEY, R.G., ISHII, S. and GONDA, T.J., Increase in specific DNA binding by carboxyl truncation suggests a mechanism for activation of Myb. Oncogene, 6,1875-1879 (1991). RUSSELL, K.S. and HUNG, M.-C., Transcriptional repression of the neu protooncogene by estrogen stimulates estrogen receptor. Cancer Rex, 52,6624-6629 (1992).
EMBOJ., 6,605-610 (1987).
3719-3727 (1987).
6682-6686 (1986).
SCHMIDHAUSER, C., CASPERSON, G.F., MYERS, C.A., SANZO, K.T., BOLTEN, S. and BISSELL, M.J., A novel transcriptional enhancer is involved in prolactin- and extracellular matrix-dependent regulation of b-casein. Mol. Biol. Cell, 3,699-709 (1992). SLAMON, D.J., CLARK, G.M., WONG, S.G., LEVIN, W.J., ULLRICH, A. and MCGUIRE, W.L., Human breast cancer: correlation of relapse and survival with amplification of the HER-2ineu oncogene. Sczence, 235, 177-182 (1987). STREULI, C.H., SCHMIDHAUSER, C., KOBRIN, M., BISSELL, M.J. and DERYNCK, R., Extracellular matrix regulates expression of the TGF-bl gene. J. Cell B i d , 120,253-260 (1993). SUEN, T.-C. and HUNG, M.-C., c-myc reverses neu-induced trans- formed morphology by transcriptional repression. Mol. Cell B i d , 11,
TORA, L., GAUB, M.P., MADER, S., DIERICH, A,, BELLARD, M. and CHAMBON, P., Cell-specific activity of a GGTCA half-palindromic oestrogen-responsive element in the chicken ovalbumin gene pro- moter. EMBO J., 7,3771-3778 (1988).
and VAN ZOELEN, E.J.J., Direct effects of estrogen on c-fos and c-myc protooncogene expression and cellular proliferation in human breast cancer cells. Mol. Cell. Endocrinol., 64,223-228 (1989). WRIGHT, C., NICHOLSON, S., ANGUS, B., SAINSBURY, J.R., FARNDON, J., CAIRNS, J. and HARRIS, A.L., Relationship between erbB-2 protein product expression and response to endocrine therapy in advanced breast cancer. Bnt. J. Cancer, 65,118-121 (1992)
354-362 (1991).
VAN DER BURG, B., VAN SEHM MILTENBURG, A.J.P., DE LAAT, S.W.
Related Documents











