Eur. J. Biochem. 262, 781–789 (1999) q FEBS 1999 Enzymatic action of human glandular kallikrein 2 (hK2) Substrate specificity and regulation by Zn 2+ and extracellular protease inhibitors Janita Lo ¨ vgren 1,2 , Kalervo Airas 3 and Hans Lilja 1 1 Department of Laboratory Medicine, Division of Clinical Chemistry, Lund University, Malmo ¨, Sweden; 2 Departments of Biotechnology and 3 Biochemistry, University of Turku, Finland Human glandular kallikrein 2 (hK2) is a serine protease expressed by the prostate gland with 80% identity in primary structure to prostate-specific antigen (PSA). Recently, hK2 was shown to activate the zymogen form of PSA (proPSA) in vitro and is likely to be the physiological activator of PSA in the prostate. hK2 is also able to activate urokinase and effectively cleave fibronectin. We studied the substrate specificity of hK2 and regulation of its activity by zinc and extracellular protease inhibitors present in the prostate and seminal plasma. The enzymatic activity and substrate specificity was studied by determining hK2 cleavage sites in the major gel proteins in semen, semenogelin I and II, and by measuring hydrolysis of various tripeptide aminomethylcoumarin substrates. HK2 cleaves substrates C-terminal of single or double arginines. Basic amino acids were also occasionally found at several other positions N-terminal of the cleavage site. Therefore, the substrate specificity of hK2 fits in well with that of a processor of protein precursors. Possible regulation mechanisms were studied by testing the ability of Zn 2+ and different protease inhibitors to inhibit hK2 by kinetic measurements. Inhibitory constants were determined for the most effective inhibitors PCI and Zn 2+ . The high affinity of PCI for hK2 (k ass = 2.0 10 5 m 21 ·s 21 ) and the high concentrations of PCI (4 mm) and hK2 (0.2 mm) in seminal plasma make hK2 a very likely physiological target protease for PCI. hK2 is inhibited by Zn 2+ at micromolar concentrations well below the 9 mm zinc concentration found in the prostate. The enzymatic activity of hK2 is likely to be reversibly regulated by Zn 2+ in prostatic fluid. This regulation may be impaired in CAP and advanced metastatic cancer resulting in lack of control of the hK2 activity and a need for other means of control. Keywords: kallikrein; serpin; zinc; substrate specificity; prostate. The glandular kallikreins are serine proteases encoded by a conserved multigene family with a high number of members in different species [1]. The human glandular kallikrein gene family consists of three members; tissue kallikrein (hK1), human glandular kallikrein 2 (hK2), and prostate-specific antigen (PSA or hK3). In general, the glandular kallikreins are involved in processing polypeptide precursors into their bioactive forms and have attracted interest as they liberate kinin from kininogen and release murine epidermal growth factor (EGF) and nerve growth factor (NGF) from their respective precursors [2]. However, interspecies comparisons are complicated by the finding that the human glandular kallikreins have evolved from a common ancestor subsequent to the separation of the murine and human lineages, indicating that there is no direct human equivalent to the murine enzymes [3]. Little is known about the physiological functions of the human glandular kallikreins. hK2 was the last of the three members of the human glandular kallikrein gene family to be identified in 1987 [4]. Both PSA and hK2 are expressed abundantly in the prostate and the levels of hK2 found in seminal fluid are about 1% of those of PSA (0.3–5 mg·mL 21 ). Both proteins are expressed at considerably lower levels in the endometrium, breast, and salivary glands, and presence of the transcripts has been demonstrated by RT-PCR in the pituitary [3,5]. The two proteins are secreted into amniotic fluid, colostrum, milk and saliva. The secretion of hK2 is stimulated by androgens, progestins, glucocorticoids, and mineralocorticoids and is regulated differently in the breast tumour cell line T47-D and the prostate cancer cell line LNCaP [6]. hK2 has attracted interest as a potential marker for prostate cancer due to its 80% sequence identity with PSA [4]. hK2 crossreacts with some monoclonal anti-PSA antibodies and can therefore affect the results of PSA measurements [7–9]. This information has been used in the development of immunoassays specific for PSA and hK2, respectively [10], and recently it has been shown that measurement of hK2 in addition to free and total PSA in serum enables greater separation of men with benign prostatic hyperplasia (BPH) from those with carcinoma of the prostate (CAP) [11,12]. hK2 has been purified and characterized from seminal plasma where most of the protein was found in complex with protein C inhibitor (PCI) [14]. It also forms complexes with several plasma protease inhibitors such as a2-antiplasmin, a1-antichymotrypsin, antithrombin III, and a2-macroglobulin [15]. The substrate specificity of hK2 has been studied and compared to that of hK1 using serpin derived peptide substrates. Correspondence to J. Lo ¨vgren, Department of Biotechnology, University of Turku, Tykisto ¨katu 6, 20520 Turku, Finland. Fax: +358 2 3338050. E-mail: [email protected] Abbreviations: hK2, Human glandular kallikrein 2; PSA, prostate-specific antigen; proPSA, zymogen form of PSA; EGF, epidermal growth factor; NGF, nerve growth factor; BPH, benign prostatic hyperplasia; CAP, carcinoma of the prostate; PCI, protein C inhibitor; Sg, semenogelin; uPA, urokinase; AMC, 7-amino-4-methylcoumarin; serpin, serine proteinase inhibitor. (Received 19 January 1999, accepted 24 March 1999)

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Eur. J. Biochem. 262, 781±789 (1999) q FEBS 1999
Enzymatic action of human glandular kallikrein 2 (hK2)Substrate specificity and regulation by Zn2+ and extracellular protease inhibitors
Janita LoÈvgren1,2, Kalervo Airas3 and Hans Lilja1
1Department of Laboratory Medicine, Division of Clinical Chemistry, Lund University, MalmoÈ, Sweden; 2Departments of Biotechnology and3Biochemistry, University of Turku, Finland
Human glandular kallikrein 2 (hK2) is a serine protease expressed by the prostate gland with 80% identity in
primary structure to prostate-specific antigen (PSA). Recently, hK2 was shown to activate the zymogen form of PSA
(proPSA) in vitro and is likely to be the physiological activator of PSA in the prostate. hK2 is also able to activate
urokinase and effectively cleave fibronectin. We studied the substrate specificity of hK2 and regulation of its activity
by zinc and extracellular protease inhibitors present in the prostate and seminal plasma. The enzymatic activity and
substrate specificity was studied by determining hK2 cleavage sites in the major gel proteins in semen, semenogelin
I and II, and by measuring hydrolysis of various tripeptide aminomethylcoumarin substrates. HK2 cleaves substrates
C-terminal of single or double arginines. Basic amino acids were also occasionally found at several other positions
N-terminal of the cleavage site. Therefore, the substrate specificity of hK2 fits in well with that of a processor of
protein precursors. Possible regulation mechanisms were studied by testing the ability of Zn2+ and different protease
inhibitors to inhibit hK2 by kinetic measurements. Inhibitory constants were determined for the most effective
inhibitors PCI and Zn2+. The high affinity of PCI for hK2 (kass = 2.0 � 105 m21´s21) and the high concentrations of
PCI (4 mm) and hK2 (0.2 mm) in seminal plasma make hK2 a very likely physiological target protease for PCI. hK2
is inhibited by Zn2+ at micromolar concentrations well below the 9 mm zinc concentration found in the prostate. The
enzymatic activity of hK2 is likely to be reversibly regulated by Zn2+ in prostatic fluid. This regulation may be
impaired in CAP and advanced metastatic cancer resulting in lack of control of the hK2 activity and a need for other
means of control.
Keywords: kallikrein; serpin; zinc; substrate specificity; prostate.
The glandular kallikreins are serine proteases encoded by aconserved multigene family with a high number of members indifferent species [1]. The human glandular kallikrein genefamily consists of three members; tissue kallikrein (hK1),human glandular kallikrein 2 (hK2), and prostate-specificantigen (PSA or hK3). In general, the glandular kallikreins areinvolved in processing polypeptide precursors into theirbioactive forms and have attracted interest as they liberatekinin from kininogen and release murine epidermal growthfactor (EGF) and nerve growth factor (NGF) from theirrespective precursors [2]. However, interspecies comparisonsare complicated by the finding that the human glandularkallikreins have evolved from a common ancestor subsequentto the separation of the murine and human lineages, indicatingthat there is no direct human equivalent to the murine enzymes[3]. Little is known about the physiological functions of thehuman glandular kallikreins.
hK2 was the last of the three members of the human glandularkallikrein gene family to be identified in 1987 [4]. Both PSAand hK2 are expressed abundantly in the prostate and the levelsof hK2 found in seminal fluid are about 1% of those of PSA(0.3±5 mg´mL21). Both proteins are expressed at considerablylower levels in the endometrium, breast, and salivary glands, andpresence of the transcripts has been demonstrated by RT-PCR inthe pituitary [3,5]. The two proteins are secreted into amnioticfluid, colostrum, milk and saliva. The secretion of hK2 isstimulated by androgens, progestins, glucocorticoids, andmineralocorticoids and is regulated differently in the breasttumour cell line T47-D and the prostate cancer cell line LNCaP[6]. hK2 has attracted interest as a potential marker for prostatecancer due to its 80% sequence identity with PSA [4]. hK2crossreacts with some monoclonal anti-PSA antibodies and cantherefore affect the results of PSA measurements [7±9]. Thisinformation has been used in the development of immunoassaysspecific for PSA and hK2, respectively [10], and recently it hasbeen shown that measurement of hK2 in addition to free andtotal PSA in serum enables greater separation of men withbenign prostatic hyperplasia (BPH) from those with carcinomaof the prostate (CAP) [11,12].
hK2 has been purified and characterized from seminalplasma where most of the protein was found in complexwith protein C inhibitor (PCI) [14]. It also forms complexeswith several plasma protease inhibitors such as a2-antiplasmin,a1-antichymotrypsin, antithrombin III, and a2-macroglobulin[15]. The substrate specificity of hK2 has been studied andcompared to that of hK1 using serpin derived peptide substrates.
Correspondence to J. LoÈvgren, Department of Biotechnology, University of
Turku, TykistoÈkatu 6, 20520 Turku, Finland. Fax: +358 2 3338050.
E-mail: [email protected]
Abbreviations: hK2, Human glandular kallikrein 2; PSA, prostate-specific
antigen; proPSA, zymogen form of PSA; EGF, epidermal growth factor;
NGF, nerve growth factor; BPH, benign prostatic hyperplasia; CAP,
carcinoma of the prostate; PCI, protein C inhibitor; Sg, semenogelin; uPA,
urokinase; AMC, 7-amino-4-methylcoumarin; serpin, serine proteinase
inhibitor.
(Received 19 January 1999, accepted 24 March 1999)

782 J. LoÈvgren et al. (Eur. J. Biochem. 262) q FEBS 1999
Both enzymes manifest trypsin-like substrate specificity thoughhK2 may accommodate small and positively charged residues atposition P2. Furthermore, hK2 requires an arginyl residue atposition P1 while hK1 usually requires a large hydrophobicresidue at P2 and arginine or lysine at P1 [16]. In exception to allglandular kallikreins, PSA manifests restricted chymotrypsin-like activity and cleaves substrates on the C-terminal side oftyrosine, histidine, leucine and glutamine residues [17±20]. PSAcleaves the insulin like growth factor binding protein andparathyroid related peptide in vitro, but the physiologicalsignificance of these actions is unclear [21,22]. In semen, PSAis responsible for dissolution of the immediately forming gelstructure by cleaving the gel forming proteins, semenogelin Iand II (Sg I and II) [23], which are also referred to as spermmotility inhibitors [24]. The multiple cleavages result in therelease of progressively motile sperm. Recently several potentialhK2 substrates have been identified. HK2, like PSA, has thecapacity to cleave Sg I, Sg II, and fibronectin [25,26]. In theprostate it may be autoactivated and activate the zymogen formsof PSA (proPSA) and urokinase (uPA) [26±30]. hK2 maytherefore be of importance in the progression of prostate cancerand play a role in generating the difference in the ratio of freeand total PSA, which is currently used to differentiate betweenCAP and BPH [31].
Elucidation of the substrate specificity and regulation of theenzymatic activity of hK2 is crucial for the understanding of itsphysiological significance. This study was performed to furtherinvestigate the substrate specificity of hK2 using variouspeptides and Sg I and Sg II as substrates, to study possiblephysiological regulation mechanisms in the prostate and seminalfluid, and to identify inhibitors that may be used duringproduction and purification of hK2.
MATERIALS AND METHODS
Reagents
Recombinant hK2 was produced and purified as describedpreviously [26]. Sg I and II were isolated from human semen asdescribed previously [32]. The tripeptide aminomethylcoumarin(AMC) substrates Boc-Phe-Ser-Arg-AMC, Boc-Gln-Gly-Arg-AMC, H-Pro-Phe-Arg-AMC, Boc-Val-Pro-Arg-AMC, H-D-Val-Leu-Lys-AMC, Tos-Gly-Pro-Arg-AMC, Tos-Gly-Pro-Lys-AMC, Z-Leu-Leu-Arg-AMC, Z-Val-Val-Arg-AMC, Z-Ala-Arg-Arg-AMC, and H-Arg-Gln-Arg-Arg-AMC were fromBachem (Bubendorf, Switzerland). The heptapeptide substratesMu-Ala-Pro-Val-Leu-Ile-Leu-Ser-Arg-AMC and Mu-Val-Pro-Leu-Ile-Gln-Ser-Arg-AMC corresponding to the pro peptidesof PSA and hK2 were from Enzyme Systems Products(Livermore, CA, USA). ACT was purified from human bloodplasma as described earlier [17]. PCI was provided by Prof.Johan Stenflo (MalmoÈ University Hospital, MalmoÈ, Sweden),and SLPI, and PSTI by Prof. Kjell Ohlsson (MalmoÈ UniversityHospital). Benzamidine hydrochloride was from Amrescow
(Solon, OH, USA), leupeptin and antipain were from ICNBiomedicals (Costa Mesa, CA, USA), Aprotinin was fromSigma (St Louis, MO, USA), and PPACK from Calbiochem (LaJolla, CA, USA).
Determination of hK2 cleavage sites in Sg I and II
Purified Sg I and II (40 mg), was incubated with hK2 (8 mg) in50 mm Tris pH 7.5, 0.1 m NaCl and 0.15 m urea at 37 8C for4 h. The fragments generated were purified by reverse phaseHPLC using a C-8 column. Elution was achieved with a 0±30%
(0.25% min21) linear acetonitrile gradient and fractions corres-ponding to individual peaks were collected. The N-terminalsequences of the individual peaks were determined byautomated N-terminal sequencing with an Applied Biosystems470 A gas-phase sequencer.
pH dependence of the enzymatic action of hK2
The pH dependence of hK2 was determined using a universalbuffer composed of 29 mm diethylbarbituric acid, 29 mm citricacid, 29 mm KH2PO4, 29 mm boric acid, 0.1 m NaCl and 0.2%BSA. The buffering range is pH 2.4±11.8. The rate of thecleavage of the substrate I-1295 (100 mm) by 1.6 pmol hK2was followed for 20 min at pH 5, 5.5, 6, 6.5, 7, 7.5, 8, 8.5, 9,9.5, and 10.
Determination of kinetic constants
The measurements were performed with a Fluoroscan II(Labsystems, Helsinki, Finland) using a 355-nm excitation filterand a 460-nm emission filter. The fluorescence of AMC (Sigma)was determined to be 700 FIU´nmol21 and this value was usedin the calculations of the rate of product formation. Unlessotherwise indicated all final analyses were performed in 200 mLof the buffer 50 mm Tris pH 7.5, 0.1 m NaCl, and 0.2% BSA at37 8C using 1.6 pmol hK2. BSA was added to the reactionmixture in order to minimize adsorption of the enzyme to thewalls of the microtiter wells. The amount of hK2 was quantifiedby a commercial PSA immunoassay (Prostatus, Wallac, Turku,Finland) with the Mabs H117 and H50 which recognize PSAand hK2 equally well [7]. Under the conditions described,1.6 pmol of the hK2 preparation cleaved 50 pmol´min21 of the100 mm substrate Pro-Phe-Arg-AMC. During the 20 minmeasurement time the consumption of the substrates is , 7%of their total amount and was not considered to affect thereaction rate. Initial analysis of the substrates were performedwith 3.2 pmol hK2 at a substrate concentration of 250 mm. TheKm value for the substrates cleaved by hK2 at these conditionswere determined using at least four substrate concentrationsranging from 0.2 � Km to 5 � Km. The Km and kcat values werecalculated from Lineweaver±Burk plots.
Inhibition of hK2
Activity of hK2 (1.6 pmol) was monitored using the substrateH-Pro-Phe-Arg-AMC (90 mm). Substrate hydrolysis was fol-lowed for 20 min starting directly or 10 min after mixing theenzyme with various inhibitors. Inhibition was evaluated bycomparison with enzyme-free controls.
Kinetic analysis of hK2 inhibition by zinc
The inhibition of hK2 by Zn2+ was studied using the substratesPro-Phe-Arg-AMC and Ala-Arg-Arg-AMC at concentrationsvarying from 9 to 180 mm and ZnCl2 concentrations rangingfrom 0.5 mm to 1 mm. Because BSA contains several bindingsites for Zn2+ it could not be used in the kinetics buffer duringthe zinc inhibition experiments. The binding of hK2 to themicrotiter well walls caused a constant decrease in the reactionrate, which was, however, similar at all zinc concentrations. Thevelocities were calculated from a five minute measurement timeafter mixing of the enzyme with the buffer containing substrateand zinc.

q FEBS 1999 Enzymatic action of human glandular kallikrein 2 (Eur. J. Biochem. 262) 783
Kinetic analysis of hK2 inhibition by PCI
The progress of the reaction of hK2 (8 nm final concentration)with the substrate Pro-Phe-Arg-AMC was monitored at twodifferent substrate concentrations without or with differentconcentrations of PCI (80, 40 or 16 nm final concentration). Thefluorescence measurements were started directly after mixingthe enzyme with the inhibitor. The inhibition of hK2 by PCIcould be described by the slow-binding inhibition mechanismpresented in Scheme 1, which has been used in analysing theinteraction of PCI with various serine proteases [33,34]. Thismechanism assumes that a reversible complex is formedbetween the proteinase and serine proteinase inhibitor (serpin).The issues justifying the use of the slow binding inhibitionmechanism despite the commonly held view that the serpin-proteinase complex is irreversible has been discussed in moredetail by Hermans et al. [33].
where E, S, P, and I represent the enzyme, substrate (peptidylAMC), product (AMC) and inhibitor (PCI), respectively; Km andkcat are Michaelis and catalytic constants for the enzymesubstrate interaction, and Ki is the inhibition constant which isequal to kdiss/kass. kass and kdiss are the association anddissociation rate constants for the enzyme inhibitor complex.For this mechanism, the progress curve of product formation isgiven by:
P � vst � �v0 2 vs�k 0
�1 2 e2kt�
where P is the amount of product at time t, k 0 is an apparent firstorder rate constant, and v0 and vs are the initial and steady-statevelocities, respectively. For the mechanism shown in Scheme 1,v0 will be independent of the inhibitor concentration, and vs andk 0 will vary with the inhibitor concentration according to thefollowing equations:
vs � v0
1� I / K 0i
k 0 � kdiss � k 0ass´I � k 0ass�K 0i � I�where K 0i and k 0ass are apparent constants that are related to thetrue constants by the expressions:
K i � K 0i/�1� S/Km�
k � k 0ass/�1� S/Km�The effect of heparin on the association rate of hK2 and PCI wasstudied using 40 nm PCI, 8 nm hK2, and heparin concentrationsranging from 1024 to 1027 m. The effect of the heparin on hK2activity was analysed by determining Km and kcat for thesubstrate at different heparin concentrations. Heparin slightlyincreased the Km of the substrate (data not shown). The increasedidn't have a significant effect on the calculation of theconstants.
RESULTS
pH optimum of the enzymatic action of hK2
The pH dependence of hK2 is shown in Fig. 1. All experimentswere performed close to the physiological pH of 7.5, which isvery close to the pH optimum of hK2.
HK2 cleavage positions in semenogelin I and II
Cleavage of either Sg I or Sg II with hK2 results in generationof a multitude of peptides. After partial separation of thepeptides by reversed phase HPLC on a C-8 column we obtainedsequences of four cleavage sites in Sg I and seven cleavage sites
Fig. 1. pH dependence of hK2 activity.
Fig. 2. hK2 cleavage sites in Sg I and II. Only the sequences surrounding
the cleavage sites are shown. The cleavage position is marked with an arrow.

784 J. LoÈvgren et al. (Eur. J. Biochem. 262) q FEBS 1999
in Sg II. The semenogelins contain three types of internalrepeats [35,36]. Most of the identified hK2 cleavage sites werelocated in different positions in these repeats. The position andsequence of the cleavage sites in Sg I and Sg II are shown inFig. 2. Three identical sites of cleavage in repeat type I, whichoccurs twice in Sg I and four times in Sg II, were identified atpositions 274 and 334 in Sg I and position 454 in Sg II. All butone of the cleavage sites contained arginine at position P1,except for one of the cleavages in semenogelin II, whichoccurred on the C-terminal side of a histidine. It is noteworthythat no cleavages occurred on the C-terminal side of a lysine.Five of the 11 cleavage sites determined were double basic, theamino acid at P2 being either arginine, lysine or histidine,indicating that hK2 may cleave substrates at both mono- anddi-basic sites. In one case P2 was occupied by phenylalaninewhich is found in the same position in PCI. In addition glycine,valine, serine, glutamine and aspartate were found at P2. In most
cleavage sites, P3 was occupied by a large group; in six of thecleavages it was glutamine or glutamate and in the others serine,histidine or lysine. In one case alanine was found at P3. Whenlooking at common motifs it can be seen that in seven casesserine was found in P6. Basic amino acids were found inaddition to positions P1 and P2 once in P5, twice in P3, P4, P6and P8, and four times in P7. On the carboxy terminal side of thecleavage site leucine was found five times in P1 and tyrosinefour times in position P3.
Hydrolysis of various peptide substrates by hK2
Substrates ending in either arginine or lysine were tested. Thekinetic constants for hydrolysis of the substrates by hK2 areshown in Table 1. The best substrate was the kallikrein substratePro-Phe-Arg-AMC having the highest kcat and kcat/Km values.The cathepsin B substrate Ala-Arg-Arg-AMC was also cleavedquite effectively, having a relatively high kcat value and a low Km
resulting in a four times lower kcat/Km value than that obtainedfor the kallikrein substrate Pro-Phe-Arg-AMC. However, nohydrolysis of Arg-Gln-Arg-Arg-AMC was detected. hK2additionally cleaved Val-Pro-Arg-AMC, and Leu-Leu-Arg-AMC, but with lower efficiency. As with the semenogelinshK2 also cleaves substrates with Arg at position P1 andpreferentially a large residue or another Arg at position P2. Noneof the substrates with lysine in the C-terminal position werecleaved. Surprisingly, hK2 did not cleave the heptapeptidesubstrates corresponding to the propeptides of PSA and hK2,which is remarkable in view of the fact that hK2 has previouslybeen shown to be able to activate the proteins by release of the
Fig. 3. Inhibition of hK2 by PCI. The cleavage of 100 mm Pro-Phe-Arg-
AMC by hK2 (8 nm) was followed in microtiter wells without (+) or with
16 nm (V), 40 nm (O), or 80 nm (*) PCI. The curves represent the fit of the
data to the equation describing the slow binding inhibition presented in
Scheme 1.
Table 3. Comparison of PCI interactions with different serine
proteases.
Enzyme
kass
(m21´s21)
Ki
(pm)
kass with heparin
(m21´s21)
hK2 (1�.98 ^ 0.26) � 105 , 25 (8�.7 ^ 0.1) � 105
APCa (8�.0 ^ 0.1) � 102 11900 ^ 600 (1�.47 ^ 0.03) � 104
thrombina (3�.31 ^ 0.02) � 104 1100 ^ 100 (2�.05 ^ 0.01) � 105
acrosina (2�.41 ^ 0.03) � 105 46 ^ 5 (5�.6 ^ 0.1) � 107
a The values for APC, thrombin and acrosin were taken from Hermans et al.
[33,34].
Table 1. Substrate hydrolysis by hK2. The substrates were used at a
concentration range of 0.2 � Km to 5 � Km and the constants calculated
from Lineweaver±Burk plots. Conditions were; buffer: 50 mm Tris pH 7.5,
0.1 m NaCl, 0.2% BSA; 37 8C; hK2 1.6 pmol; reaction volume 200 mL. NR,
no reaction detected.
Substrates
Km
(mm)
kcat
(min21)
kcat/Km
(mm21´min21)
Activitya
(%)
Pro-Phe-Arg-AMC 40 55 1�.375 100
Val-Pro-Arg-AMC 48 1�.6 0�.034 6
Gly-Pro-Arg-AMC NR
Gly-Pro-Lys-AMC NR
Leu-Leu-Arg-AMC 71 2�.4 0�.034 7
Val-Val-Arg-AMC NR
Val-Leu-Lys-AMC NR
Phe-Ser-Arg-AMC NR
Gln-Gly-Arg-AMC NR
Ala-Arg-Arg-AMC 20 7�.2 0�.360 33
Arg-Gln-Arg-Arg-AMC NR
a The hydrolytic activity of hK2 with 100 mm substrate in relation to the
hydrolytic activity of hK2 with 100 mM of the tissue kallikrein substrate
H-Pro-Phe-Arg-AMC.
Table 2. Effects of protease inhibitors on hK2 activity. Inhibitors, at
commonly used concentrations, and hK2 (8.3 nm) were mixed and
proteolysis of 90 mm H-Pro-Phe-Arg-AMC was followed up to 20 min.
The proteolytic activity is expressed in percentage of inhibitor-free control
after 10 min of incubation.
Inhibitor
Final concentration
(mm)
Activity
(%)
ZnCl2 200
100
1�.2
10
PCI 0�.08
0.016
0
50
PPACK 5 0
Benzamidine 20000
5000
8
27
Leupeptin 100 12
Antipain 500
100
10
33
Aprotinin 5
30
75
47
SLPI 4
0.08
35
92
PSTI 4
0.08
89
100
ACT 0�.8 100
q FEBS 1999 Enzymatic action of human glandular kallikrein 2 (Eur. J. Biochem. 262) 785
propeptide from the zymogen forms of these proteins [26,27,29].The lack of hydrolysis of the corresponding AMC-peptidesubstrates may be due to the high hydrophobicity of the peptide,which could result in inappropriate folding.
Effects of protease inhibitors on hK2
The effects of several protease inhibitors on the hydrolyticactivity of hK2 are shown in Table 2. None of the reversibleprotease inhibitors fully inhibited 8 nm hK2 when 90 mmsubstrate was used. hK2 was only weakly inhibited by thereversible peptide inhibitors leupeptin and antipain. The highestrecommended working concentration (100 mm) of the respectiveinhibitor was found to give 60% and 90% inhibition of hK2activity against the 90 mm peptide substrate. Aprotinin provednot to be a good hK2 inhibitor, and benzamidine is required atconcentrations above 20 mm for efficient inhibition. Theirreversible thrombin inhibitor PPACK inhibited hK2 rapidlywhen used at a 5-mm concentration. Therefore, PPACK can be
used to obtain fast irreversible inhibition of hK2. ZnCl2effectively inhibits hK2 but when used at high concentrationseasily causes precipitation of proteins. Of the protease inhibitorspresent in the prostate, PSTI and SLPI inhibited hK2 weakly andthis inhibition is probably not physiologically significant. ACThas been previously shown to form complexes with hK2 [15].This reaction is however, slow and no inhibition of the hK2activity by a 100-fold molar excess of ACT was detected duringthe 20 min measurement time.
Kinetic analysis of hK2 inhibition by PCI
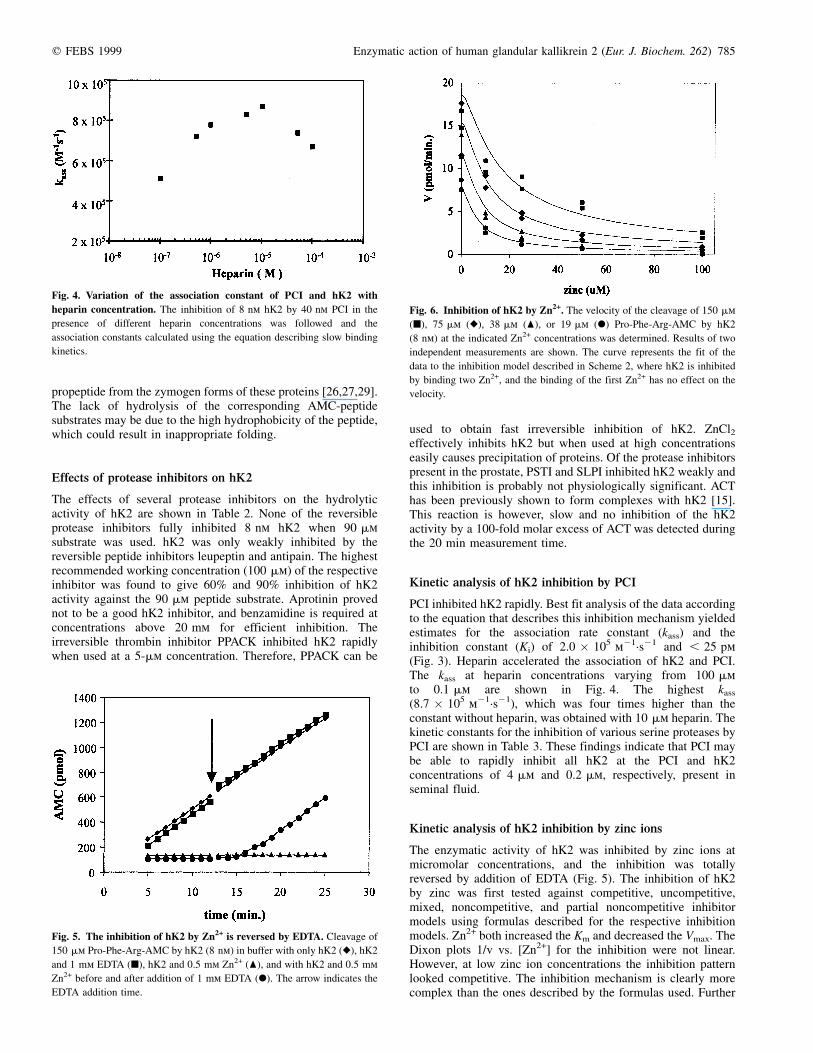
PCI inhibited hK2 rapidly. Best fit analysis of the data accordingto the equation that describes this inhibition mechanism yieldedestimates for the association rate constant (kass) and theinhibition constant (Ki) of 2.0 � 105 m21´s21 and , 25 pm(Fig. 3). Heparin accelerated the association of hK2 and PCI.The kass at heparin concentrations varying from 100 mmto 0.1 mm are shown in Fig. 4. The highest kass
(8.7 � 105 m21´s21), which was four times higher than theconstant without heparin, was obtained with 10 mm heparin. Thekinetic constants for the inhibition of various serine proteases byPCI are shown in Table 3. These findings indicate that PCI maybe able to rapidly inhibit all hK2 at the PCI and hK2concentrations of 4 mm and 0.2 mm, respectively, present inseminal fluid.
Kinetic analysis of hK2 inhibition by zinc ions
The enzymatic activity of hK2 was inhibited by zinc ions atmicromolar concentrations, and the inhibition was totallyreversed by addition of EDTA (Fig. 5). The inhibition of hK2by zinc was first tested against competitive, uncompetitive,mixed, noncompetitive, and partial noncompetitive inhibitormodels using formulas described for the respective inhibitionmodels. Zn2+ both increased the Km and decreased the Vmax. TheDixon plots 1/v vs. [Zn2+] for the inhibition were not linear.However, at low zinc ion concentrations the inhibition patternlooked competitive. The inhibition mechanism is clearly morecomplex than the ones described by the formulas used. Further
Fig. 4. Variation of the association constant of PCI and hK2 with
heparin concentration. The inhibition of 8 nm hK2 by 40 nm PCI in the
presence of different heparin concentrations was followed and the
association constants calculated using the equation describing slow binding
kinetics.
Fig. 5. The inhibition of hK2 by Zn2+ is reversed by EDTA. Cleavage of
150 mm Pro-Phe-Arg-AMC by hK2 (8 nm) in buffer with only hK2 (V), hK2
and 1 mm EDTA (B), hK2 and 0.5 mm Zn2+ (O), and with hK2 and 0.5 mm
Zn2+ before and after addition of 1 mm EDTA (X). The arrow indicates the
EDTA addition time.
Fig. 6. Inhibition of hK2 by Zn2+. The velocity of the cleavage of 150 mm
(B), 75 mm (V), 38 mm (O), or 19 mm (X) Pro-Phe-Arg-AMC by hK2
(8 nm) at the indicated Zn2+ concentrations was determined. Results of two
independent measurements are shown. The curve represents the fit of the
data to the inhibition model described in Scheme 2, where hK2 is inhibited
by binding two Zn2+, and the binding of the first Zn2+ has no effect on the
velocity.

786 J. LoÈvgren et al. (Eur. J. Biochem. 262) q FEBS 1999
analysis of the inhibition mechanism was performed by derivingthe rate equations for various more complex mechanisms andanalysing the data by least-squares best-fit systems. The possiblemechanism required two bound zinc ions, and is presented byScheme 2. In the best mechanism, the first bound zinc ion doesnot cause inhibition (k = k 0, or k 0 was even slightly higherthan k).
For this mechanism the rate of product formation (v) will begiven by the following equation
v � S
Ks
k � k 0Zn
K i1
� �e
where the concentration of free enzyme e is
e � etot
Zn
K i1
Zn
K i2
� Zn
Ki1
� 1
� �1 1
S
Ks
� �The equation fitted the experimental data quite satisfactorily ascan be seen in Fig. 6. The constant values wereKi1 = 4.6 ^ 3.9 mm, Ki2 = 3.2 ^ 0.7 mm and k = k 0. Best-fitanalyses were accomplished also for mechanisms involvinginactivating dimerizations of the hK2 molecules as zinc ionshave been shown to inhibit the mouse gamma-NGF and to becritical in the association of the mouse 7S NGF complex [37].The dimerizations were thought to occur either with a boundzinc ion in both hK2 molecules and/or with one zinc ion boundbetween the two hK2 molecules. These mechanisms did notresult in to a more optimal fit than those in Scheme 2. However,the difference between the two mechanism models was so smallthat the analysis could not decide whether this kind ofdimerzation occurs or not.
DISCUSSION
We examined the substrate specificity of hK2 by firstdetermining hK2 cleavage sites in Sg I and II. All but one ofthe cleavage sites had arginine at position P1, in one case Sg IIwas cleaved on the carboxy terminal side of a histidine. Five ofthe 11 cleavage sites determined were di-basic, the amino acid atP2 being either arginine, lysine or histidine. In the other casesthe amino acid at P2 was a large aliphatic or aromatic group, oran acidic amino acid. At one cleavage site P2 was glycine. Thisdata is in accordance with the data obtained by Bourgeois et al.using serpine derived peptide substrates indicating that hK2 canalso cleave substrates with small or basic amino acids in positionP2 [16]. Part of the differences in kallikrein substrate specificityhave been suggested to be dependent of the amino acids locatedin positions P3±P8 through an extended interaction sitecomposed of loop 4 of the kallikreins [38]. In addition to thelocation of basic amino acids at P1 and P2 in the hK2 cleavagesites in Sg I and Sg II, basic residues were additionally foundonce in P5, twice in P3, P4, P6 and P8, and four times in P7. Sixof the cleavage sites contained glutamine or glutamate at P3, andseven contained serine in P6. It is not yet known whether or nothK2 cleaves the semenogelins in vivo. However, the cleavage
sites identified so far from semenogelin fragments purified fromliquefied seminal plasma are those generated by PSA.
Four of the tested 13 peptide-AMC substrates were hydro-lysed by hK2, among which the kallikrein substrate Pro-Phe-Arg-AMC was the best with the highest kcat and kcat/Km values.The cathepsin B substrate Ala-Arg-Arg-AMC was also cleavedquite effectively while no hydrolysis of Arg-Gln-Arg-Arg-AMCwas detected. In addition hK2 hydrolysed Val-Pro-Arg-AMC,which is also a substrate for Kex2, thrombin, and trypsin, andLeu-Leu-Arg-AMC which corresponds to the amino acids in theprotease inhibitor leupeptin. The moderate inhibition of hK2 byleupeptine is consistent with the data obtained with the peptidesubstrate.
The substrate specificity of hK2 fits well with that of aprocessor of polypeptide precursors. The majority of theprecursor cleavage sites are either dibasic (KR/RR), of thetype (RXK/RXXR), consist of single basic residues, or, veryrarely, chymotrypsin like recognition sites [39±42]. In addition,basic residues are often found on the amino-terminal side of thecleavage site. Many of the precursors are probable substrates ofthe subtilisin-like proprotein convertases PC1, PC2, furin, PC4,PACE4, PC6 or PC7. Furin has, for instance, the capacity toprocess b-NGF, which in the mouse is processed by a glandularkallikrein [43]. Whether or not PSA, with its chymotrypsin-likeactivity, and/or hK2 are also involved in processing polypeptideprecursors is an issue which deserves further investigation. hK2could conceivably process precursors with Pro-Arg sites, whichare not cleaved by furin related enzymes [41].
Several protease inhibitors such as ACT, SLPI, PSTI, and PCIare expressed in the prostate. SLPI and PCI are found in seminalfluid at concentrations of 20 mg´mL21, and 220 mg´mL21,respectively [44,45]. The molar SLPI concentrations are onlyabout fivefold higher than that of hK2 (6 mg´mL21). Because a100 � molar excess of SLPI was needed to inhibit half of thehK2 activity, SLPI can not be considered to be a significantinhibitor of hK2. PSTI was an even less efficient inhibitor ofhK2. hK2 has been shown to form complexes with ACTand low amounts of these complexes have also been foundin serum [46]. This complex formation is slow however,and ACT is not likely to be a physiologically importantinhibitor of hK2.
PCI has a very high affinity for hK2 (kass = 2.0 � 105 m21´s21)but in addition to hK2 PCI has been shown to react withAPC, uPA, tPA, plasma kallikrein, thrombin, factor Xa,factor XIa, acrosin, tissue kallikrein and PSA [47±50].Kinetics of the inhibition of tissue kallikrein, thrombin,protein C, and acrosin by PCI have been studied [33,34,51].Comparison of the results shows that hK2 and acrosin haveequally high kass with PCI, whereas the kass for APC andthrombin with PCI are 1000-fold and 10-fold lower,respectively. In general the inhibitory activity of PCI isstimulated by heparin and other glycosaminoglycans. The kass
of hK2 and PCI is increased fourfold by heparin. Heparinhas been shown to cause a similar increase in the associationrate of PCI and PSA [50], but has been shown to inhibit theassociation of PCI and tissue kallikrein which has anapparent kass of 2.3 � 104´m21s21 in the absence of heparin[51]. In the presence of heparin the kass of acrosin and PCIis 100-fold higher than that of hK2 and PCI. The kass forPCI and hK2 is however, still 10-fold higher than that of PCI andAPC and fourfold higher than that of PCI and thrombin in thepresence of heparin. It has been argued that PCI is likely to be akallikrein- or acrosin-binding protein due to its high affinity forthese proteases, and suggested that the function of PCI inseminal plasma could be to protect the female reproductive tract

q FEBS 1999 Enzymatic action of human glandular kallikrein 2 (Eur. J. Biochem. 262) 787
from unwanted degradation of proteins by acrosin. Anotheralternatives that the PCI acrosin interaction may represent aregulatory step in fertilization [34]. Our results indicate that PCImay be a physiological inhibitor of hK2 which may prefer tobind to hK2 in the presence of other serine proteases. The highaffinity of PCI for hK2 explains why most of the hK2 in seminalplasma, shortly after ejaculation, is found in complex with PCI.The half-life for the formation of the PCI-hK2 complex wascalculated to be 1 s at PCI and hK2 concentrations of 4 mm and0.2 mm, respectively, found in seminal plasma. PCI is found inall the body fluids which have been shown to contain hK2 [45].The majority of the PCI found in seminal plasma is contributedby the seminal vesicles, but PCI is expressed to some degreealso by the prostate [45]. In prostate cancer where the normalgland structure is disrupted PCI may be an important hK2inhibitor.
Normal and hyperplastic prostate tissues accumulate thehighest zinc levels in the body. The pituitary also contains highzinc concentrations with 4±5 mm zinc in its secretory granules[52] while the zinc concentration in the prostate and prostaticfluid is around 9 mm [53]. A major part of the zinc ions in theprostate and in prostatic fluid are bound to different proteins orcitrate, which is present at a 10-fold molar excess to zinc [54].The free zinc ion concentration is not known as it is difficult todetermine or predict, but is likely to constitute a small fractionof the total zinc amount. Zinc ions inhibit hK2 at micromolarlevels. A zinc binding site composed of the amino acids His91,101, and 233 has been suggested for PSA through molecularmodelling [55]. These amino acids correspond to His87, 95, and224 in the hK2 sequence. According to the experimental data theinhibition involves the binding of two zinc ions, the first ofwhich has little effect on the activity. The location of the secondsite is unknown. It may be generated by two adjacent hK2molecules. As the hK2 zinc binding constants are quite low Ki1
3±4 mm and Ki2 2±3 mm when compared to the total zincconcentrations of the prostate it is likely that zinc is an importantregulator of hK2 activity. The presented mechanism provides avery delicate means of controlling hK2 activity, allowing hK2activity against substrates for which hK2 has high affinity whileunspecific cleavage is prohibited. It is possible that binding ofthe first zinc ion even enhances hK2 activity against goodsubstrates. In seminal fluid the total zinc concentration isdecreased to a mean of 2 mm due to the mixing of prostatic fluidwith the seminal vesicle secretion. Additionally the major gelforming proteins of seminal plasma have been shown to bindzinc making the free zinc ion concentration in the seminal fluidlower than that in the prostate [56]. This may constitute a meansto reverse the regulation of hK2 by zinc ions, but hK2 is likely tobe rapidly inhibited by PCI as the half life for the association isonly a second. This is consistent with the finding of covalent andcomplete binding of hK2 to PCI within 10 minutes afterejaculation [25]. Therefore, the main function of hK2 couldmore likely be to activate precursor molecules in the prostateand prostatic fluid.
The zinc concentrations in normal prostate, BPH and CAPvary significantly due to the inability of the cancer cells toaccumulate and excrete zinc [57,58]. Also in metastatic cancerthe zinc amount is likely to be low. This could result in increasedhK2 activity and cleavage of proteins that are not normallysubstrates of hK2. Another alternative is that the expression ofextracellular protease inhibitors is increased and hK2 isirreversibly inhibited. Further investigation of the molecularforms of hK2 in the prostate and analysis of its actions in cellculture are needed for understanding its activity. HK2 couldaffect proteins involved in a diversity of functions including
growth control and cell to cell interactions and play an importantrole in the progression of prostate cancer.
ACKNOWLEDGEMENTS
This work was supported in part by grants from the Biomed 2 Program, Area
4.1.7 (contract number BMH4-CT96-0453); the Swedish Medical Research
Council (project number 7903); the Swedish Cancer Society (project number
3555); the Faculty of Medicine at Lund University; the Research Fund and
the Cancer Research Fund of the University Hospital, MalmoÈ; the Crafoord
Foundation; the Gunnar, Arvid, and Elisabeth Nilsson Foundation; and the
Fundacion Federico S. A.
REFERENCES
1. Clements, J.A. (1989) The glandular kallikrein family of enzymes:
tissue-specific expression and hormonal regulation. Endocrinol Rev.
10, 393±419.
2. Drinkwater, C.C., Evans, B.A. & Richards, R.I. (1988) Kallikreins,
kinins and growth factor biosynthesis. Trends Biochem. Sci. 13,
169±172.
3. LoÈvgren, J., Valtonen-AndreÂ, C., Marsal, K., Lilja, H. & Lundwall, AÊ .
(1999) Measurement of prostate-specific antigen and human glandular
kallikrein 2 in different body fluids. J. Androl. 20 (in press).
4. Schedlich, L.J., Bennetts, B.H. & Morris, B.J. (1987) Primary structure
of a human glandular kallikrein gene. DNA 6, 429±437.
5. Clements, J.A., Mukthar, A., Verity, K., Pullar, M., McNeill, P.,
Cummins, J. & Fuller, P.J. (1996) Kallikrein gene expression in human
pituitary tissue. Clin. Endcrinol. 44, 223±231.
6. Hsieh, M.-L., Charlesworth, C., Goodmanson, M., Zhang, S., Seay, T.,
Klee, G.G., Tindall, D.J. & Young, C.Y.F. (1997) Expression of human
prostate-specific glandular kallikrein protein (hK2) in the breast
camcer cell line T47-D. Cancer Res. 57, 897±899.
7. LoÈvgren, J., Piironen, T., OÈ vermo, C., Dowell, B., Karp, M., Pettersson,
K., Lilja, H. & Lundwall, AÊ . (1995) Production of recombinant PSA
and hK2 and analysis of their immunologic cross-reactivity. Biochem.
Bioph. Res. Co. 213, 888±895.
8. Piironen, T., Villoutreix, B.O., Becker, C., Hollingsworth, K., Vihinen,
M., Bridon, D., Qiu, X., Rapp, J., Dowell, B., LoÈvgren, T., Pettersson,
K. & Lilja, H. (1998) Determination and analysis of antigenic epitopes
of prostate-specific antigen (PSA) and human glandular kallikrein 2
(hK2) using synthetic peptides and computer modeling. Protein Sci. 7,
259±269.
9. Corey, E., Buhler, K.R. & Vessella, R.L. (1997) Cross-reactivity of ten
anti-prostate-specific antigen monoclonal antibodies with human
glandular kallikrein. Urology 50, 567±571.
10. Piironen, T., LoÈvgren, J., Karp, M., Eerola, R., Lundwall, AÊ ., Dowell,
B., LoÈvgren, T., Lilja, H. & Pettersson, K. (1996) Immunofluorometric
assay for the sensitive and specific measurement of human prostatic
glandular kallikrein (hK2) in serum. Clinical Chem. 42, 1034±1041.
11. Kwiatkowski, M., Recker, F., Piironen, T., Pettersson, K., Otto, T.,
Wernli, M. & Tscholl, R. (1998) In prostatism patients the ratio of
human glandular kallikrein to free PSA improves the discrimination
between prostate cancer and benign hyperplasia within the diagnostic
`gray zone' of total PSA 4±10 ng/ml. Urology 52, 360±365.
12. Recker, F., Kwiatkowski, M.K., Piironen, T., Pettersson, K., LuÈmmen,
G., Wernil, M., WlefelspuÈtz, J., Goepel, M., Huber, A. & Tscholl, R.
(1998) The importance of human glandular kallikrein and its
correlation with different prostate specific antigen serum forms in
the detection of prostate carcinoma. Cancer 83, 2540±2547.
13. Reference withdrawn.
14. Deperthes, D., Chapdelaine, P., Tremblay, R.R., Brunet, C., Berton, J.,
HeÂbert, J., Lazure, C. & DubeÂ, J.Y. (1995) Isolation of prostatic
kallikrein hK2, also known as hGK-1, in human seminal plasma.
Biochim. Biophys. Acta 1245, 311±316.
15. Frenette, G., Deperthes, D., Tremblay, R.R., Lazure, C. & DubeÂ,
J.Y. (1997) Purification of enzymatically active kallikrein hK2
from human seminal plasma. Biochim. Biophys. Acta 1334,
109±115.

788 J. LoÈvgren et al. (Eur. J. Biochem. 262) q FEBS 1999
16. Bourgeois, L., Brillard-Bourdet, M., Deperthes, D., Juliano, M.A.,
Juliano, L., Tremblay, R.R., DubeÂ, J.Y. & Gauthier, F. (1997) Serpin-
derived peptide substrates for investigating the substrate specificity
of human tissue kallikreins hK1 and hK2. J. Biol. Chem. 272,
29590±29595.
17. Christensson, A., Laurell, C.-B. & Lilja, H. (1990) Enzymatic activity of
prostate-specific antigen and it's reactions with extracellular serine
proteinase inhibitors. Eur. J. Biochem. 194, 755±763.
18. Malm, J., Hellman, J. & Lilja, H. (1997) Mapping of the unique enzyme
characteristics of PSA. J. Urology 157 (Suppl.), 345.
19. Denmeade, S.R., Lou, W., LoÈvgren, J., Malm, J., Lilja, H. & Isaacs, J.T.
(1997) Specific and efficient substrates for assaying the proteolytic
activity of prostate-specific antigen. Cancer Res. 57, 4924±4930.
20. Coombs, G.S., Bergstrom, R.C., Pellequer, J.-L., Baker, S.I., Navre, M.,
Smith, M.M., Tainer, J.A., Madison, E.L. & Corey, D.R. (1998)
Substrate specificity of prostate-specific antigen (PSA). Chem. Biol. 5,
475±488.
21. Cohen, P., Graves, H.C.B., Pheel, D.M., Kamarei, M., Giudice, L.C. &
Rosenfeld, R.G. (1992) Prostate-specific antigen (PSA) is an insulin-
like growth factor binding protein-3 protease found in seminal plasma.
J Clin. Endocr Metab. 75, 1046±1053.
22. Iwamura, M., Hellman, J., Cockett, A., Lilja, H. & Gershagen, S. (1996)
Alteration of the hormonal bioactivity of parathyroid hormone-related
protein (PTHrP) as a result of limited proteolysis by prostate-specific
antigen. Urology 48, 317±325.
23. Lilja, H. (1985) A kallikrein-like serine protease in prostatic fluid
cleaves the predominant seminal vesicle protein. J. Clin. Invest. 76,
1899±1903.
24. Robert, M. & Gagnon, C. (1996) Purification and characterization of the
active precursor of a human sperm motility inhibitor secreted by
the seminal vesicles: identity with semenogelin. Biol. Reprod. 55,
813±821.
25. Deperthes, D., Frenette, G., Brilliard-Bourdet, M., Bourgeous., L.,
Gauthier, F., Tremblay, R.R. & DubeÂ, J.Y. (1996) Potential involve-
ment of kallikrein hK2 in the hydrolysis of the human seminal vesicle
proteins after ejaculation. J. Androl. 17, 659±665.
26. LoÈvgren, J., Rajakoski, K., Karp, M., Lundwall, AÊ . & Lilja, H. (1997)
Activation of the zymogen form of prostate-specific antigen by human
glandular kallikrein 2. Biochem. Bioph. Res. Co. 238, 549±555.
27. Kumar, A., Mikolajczyk, S.D., Goel, A.S., Millar, L.S. & Saedi, M.S.
(1997) Expression of pro form of prostate-specific antigen by
mammalian cells and its conversion to mature, active form by
human kallikrein 2. Cancer Res. 57, 3111±3114.
28. Mikolajczyk, S.D., Millar, L.S., Marker, K.M., Grauer, L.S., Goel, A.,
Cass, M.M.J., Kumar, A. & Saedi, M.S. (1997) Ala217 is important
for the catalytic function and autoactivation of prostate-specific human
kallikrein 2. Eur. J. Biochem. 246, 440±446.
29. Takayama, T.K., Fujikawa, K. & Davie, E.W. (1997) Characteriza-
tion of the precursor of prostate-specific antigen. J. Biol. Chem.
272, 21582±21588.
30. Frenette, G., Tremblay, R.R., Lazure, C. & Dube, J.Y. (1997) Prostatic
kallikrein hK2 but not prostate-specific antigen (hK3) activates
single-chain urokinase-type plasminogen activator. Int. J. Cancer
71, 897±899.
31. Oesterling, J.E., Jacobsen, S.J., Klee, G.G., Pettresson, K., Piironen, T.,
Abrahamsson, P.-A., Stenman, U.-H., Dowell, B., LoÈvgren, T. & Lilja,
H. (1995) Free, complexed and total serum PSA: the establishment
of appropriate reference ranges for their concentratios and ratios.
J. Urology. 154, 1090±1095.
32. Malm, J., Hellman, J., Magnusson, H., Laurell, C.-B. & Lilja, H. (1996)
Isolation and characterization of the major gel proteins in human
semen, semenogelin I and semenogelin II. Eur. J. Biochem. 238,
48±53.
33. Hermans, J.M. & Stone, S.R. (1993) Interaction of activated protein C
with serpins. Biochem. J. 295, 239±245.
34. Hermans, J.M., Jones, R. & Stone, S.R. (1994) Rapid inhibition of
the sperm protease acrosin by protein C inhibitor. Biochemistry 33,
5440±5444.
35. Lilja, H., Abrahamsson, P.-A. & Lundwall, AÊ . (1989) Semenogelin,
the predominant protein in human semen. J Biol. Chem. 264,
1894±1900.
36. Lilja, H. & Lundwall, AÊ . (1992) Molecular cloning and epididymal and
seminal vesiculr transcripts encoding a semenogelin-related protein.
Proc. Natl Acad. Sci. USA. 89, 4559±4563.
37. Pattison, S.E. & Dunn, M.F. (1975) On the relationship of zinc ion to the
structure and function of the 7S nerve growth factor protein.
Biochemistry 14, 2733±2739.
38. Brillard-Bourdet, M., Moreau, T. & Gauthier, F. (1995) Substrate
specificity of tissue kallikreins: importance of an extended interaction
site. Biochim. Biophys. Acta 1246, 47±52.
39. Devi, L. (1991) Consensus sequence for processing of peptide
precursors at monobasic sites. FEBS Lett. 280, 189±194.
40. Steiner, D.F., Smeekens, S.P., Ohagi, S. & Chan, S.J. (1992) The new
enzymology of precursor processing endoproteases. J. Biol. Chem.
267, 23435±23438.
41. Nakayama, K., Watanabe, T., Nakagawa, T., Kim, W.-S., Nagahama, M.,
Hosaka, M., Hatsuzawa, K., Kondoh-Hashiba, K. & Murakami, K.
(1992) Consensus sequence for precursor processing at mono-arginyl
site. J. Biol. Chem. 267, 16335±16340.
42. Robichon, A. & Kuks, P. (1991) Proteolysis in rat hypothalamic
neurosecretory granules: characterization of an alpha-chymotrypsin-
like activity in the pathway of intracellular processing of prohor-
mones. Endocrinology 128, 1974±1980.
43. Bresnahan, P.A., Leduc, R., Thomas, L., Thorner, J., Gibson, H.L.,
Brake, A.J., Barr, P.J. & Thomas, G. (1990) Human fur gene encodes a
yeast KEX2-like endoprotease that cleaves prob-NGF in vivo. J. Cell.
Biol. 111, 2851±2859.
44. Ohlsson, K., Bjartell, A. & Lilja, H. (1995) Secretory leucocyte protease
inhibitor in the male genital tract: PSA-induced proteolytic processing
in human semen and tissue localization. J. Androl. 16, 64±73.
45. Laurell, M., Christensson, A., Abrahamsson, P.-A., Stenflo, J. & Lilja,
H. (1992) Protein C inhibitor in human body fluids. J. Clin. Invest. 89,
101±109.
46. Grauer, L.S., Finlay, J.A., Mikolajczyk, S.D., Pusateri, K.D. & Wolfert,
R.L. (1998) Detection of human glandular kallikrein, hK2, as its
precursor form and in complex with protease inhibitors in prostate
carcinoma serum. J. Androl. 19, 407±411.
47. Espana, F., EstelleÂs, A., FernaÂndez, P.J., Gilabert, J., SaÂnchez-
Cuenca, J. & Griffin, J. (1993) Evidence for the regulation of
urokinase and tissue type plasminogen activators by the serpin,
protein C inhibitor, in semen and blood plasma. Thromb.
Haemostasis 70, 989±994.
48. Zheng, X., Geiger, M., Ecke, S., Bielek, E., Donner, P., EberspaÈcher, U.,
Schleuning, W.-D. & Binder, B.R. (1994) Inhibition of acrosin by
protein C inhibitor and localization of protein C inhibitor to
spermatozoa. Am. J Physiol. 262, C466±C472.
49. Espana, F., Fink, E., Sanchez-Cuenca, J., Gilabert, J., Estelles, A.
& Witzgall, K. (1995) Complexes of tissue kallikrein with
protein C inhibitor in human semen and urine. Eur. J. Biochem.
234, 641±649.
50. Christensson, A. & Lilja, H. (1994) Complex formation between protein
C inhibitor and prostate-specific antigen in vitro and in human semen.
Eur. J. Biochem. 220, 45±53.
51. Ecke, S., Geiger, M., Resch, I., Jerabek, I., Sting, L., Maier, M., Binder,
B. & R. (1992) Inhibition of tissue kallikrein by protein C
inhibitor. J. Biol. Chem. 267, 7048±7052.
52. Cunningham, B.C., Mulkerrin, M.G. & Wells, J.A. (1991) Dimerization
of human growth hormone by zinc. Science 253, 545±253.
53. Kavanagh, J.P. (1985) Sodium, potassium, calcium, magnesium, zinc,
citrate and chloride content of human prostatic and seminal fluid.
J. Reprod. Fert. 75, 35±41.
54. Kavanagh, J.P. (1983) Zinc binding properties of human prostatic
tissue, prostatic secretion, and seminal fluid. J. Reprod. Fert. 68,
359±363.
55. Villoutreix, B.O., Getzoff, E.D. & Griffin, J.H. (1994) A structural
model for the prostate disease marker human prostate-specific antigen.
Protein Sci. 3, 2033±2044.

q FEBS 1999 Enzymatic action of human glandular kallikrein 2 (Eur. J. Biochem. 262) 789
56. Frenette, G., Tremblay, R.R. & Dube, J.Y. (1989) Zinc binding to major
human seminal coagulum proteins. Arch. Androl. 23, 155±163.
57. Gyorkey, F., Min, K.W., Huff, J.A. & Gyorkey, P. (1967) Zinc and
magnesium in human prostate gland: normal, hyperplastic, and
neoplastic. Cancer Res. 27, 1348±1353.
58. Feustel, A., Wennrich, R., Steininger, D. & Klauss, P. (1982) Zinc and
cadmium concentration in prostatic carcinoma of different histological
grading in comparison to normal prostate tissue and adenofibro-
myomatosis (BPH). Urol. Res. 10, 301±303.
Related Documents











