Entwicklung von Fermentationsprozessen zur Produktion rekombinanter Antikörperfragmente in Pichia pastoris und Nicotiana tabacum. Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des akademischen Grades eines Doktors der Naturwissenschaften genehmigte Dissertation von Diplom-Biologe Stephan Hellwig aus Birkesdorf jetzt Düren Berichter: Universitätsprofessor Dr. rer. nat. F. Kreuzaler Universitätsprofessor Dr. Ing. W. Hartmeier Tag der mündlichen Prüfung: 23. August 2000

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Entwicklung vonFermentationsprozessen zur Produktion
rekombinanter Antikörperfragmentein Pichia pastoris
und Nicotiana tabacum.
Von der Fakultät für Mathematik, Informatik und Naturwissenschaften der
Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des
akademischen Grades eines Doktors der Naturwissenschaften genehmigte
Dissertation
von
Diplom-Biologe
Stephan Hellwigaus Birkesdorf jetzt Düren
Berichter: Universitätsprofessor Dr. rer. nat. F. Kreuzaler
Universitätsprofessor Dr. Ing. W. Hartmeier
Tag der mündlichen Prüfung: 23. August 2000


i
1 Einleitung ............................................................................................................................................1
1.1 Antikörper und Antikörperfragmente...............................................................................................1
1.1.1 Rolle, Aufbau und Struktur von Antikörpern ..........................................................................1
1.1.2 Modifizierte Antikörper und Antikörperfragmente .................................................................2
1.1.3 Bedeutung rekombinanter Antikörper und Antikörperfragmente ...........................................4
1.2 Expressionssysteme für rekombinante Proteine...............................................................................4
1.2.1 Produktion rekombinanter Proteine in Bakterien ....................................................................5
1.2.2 Produktion rekombinanter Proteine in tierischen Zellkulturen................................................6
1.2.3 Produktion rekombinanter Proteine in pflanzlichen Expressionssystemen.............................6
1.2.4 Produktion rekombinanter Proteine in Hefen ........................................................................12
1.3 Problemstellung..............................................................................................................................17
2 Material und Methoden ...................................................................................................................20
2.1 Material ..........................................................................................................................................20
2.1.1 Chemikalien und Verbrauchsmaterialien ..............................................................................20
2.1.2 Verwendete Medien, Puffer und Lösungen ...........................................................................20
2.1.3 Enzyme und Reaktionskits ....................................................................................................20
2.1.4 Antikörper..............................................................................................................................20
2.1.5 Geräte, Apparaturen und Zubehör .........................................................................................21
2.1.6 Fermenter...............................................................................................................................22
2.1.6.1 Fermenter für die Pflanzensuspensionskultur ...................................................................22
2.1.6.2 Fermenter für die Kultivierung von Pichia pastoris .........................................................23
2.1.7 Verwendete Zellinien ............................................................................................................24
2.1.7.1 Zellinien von Nicotiana tabacum cv Bright-Yellow-2 (BY-2) .........................................24
2.1.7.2 Zellinien von Pichia pastoris ............................................................................................24
2.2 Molekularbiologische Methoden....................................................................................................25
2.2.1 PCR zur Bestimmung des Methanolverwertungs-Phänotyps von Pichia pastoris................25
2.2.2 Analytische Agarosegelelektrophorese .................................................................................26
2.3 Proteinchemische, naßchemische und immunologische Methoden ...............................................26
2.3.1 Herstellung von Proteinextrakten aus BY-2 Zellen...............................................................26
2.3.2 Bestimmung des Gesamtproteingehaltes von BY-2 Zellen...................................................27
2.3.3 Bestimmung des Phosphat-Gehalts in Fermentationsüberständen ........................................27
2.3.4 Bestimmung von Glukose, Fruktose und Saccharose in Fermentationsüberständen ............28
2.3.5 SDS-PAA Gelelektrophorese (SDS-PAGE) und Coomassie Brilliant Blue Färbetechnik ...28
2.3.5.1 Gelelektrophorese .............................................................................................................28
2.3.5.2 Coomassie Brilliant Blue Färbung ....................................................................................28
2.3.6 Western Blot ..........................................................................................................................29
2.3.7 ELISA....................................................................................................................................30
2.3.7.1 ELISA zum Nachweis des biscFv2429 in Pflanzensuspensionskultur .............................31
2.3.7.2 ELISA zum Nachweis von CEA-NA3 in Pichia pastoris – Überständen ........................32
2.3.7.3 ELISA zum Nachweis von scFv4813 in Pichia pastoris – Überständen..........................32
2.3.8 Konzentrationsbestimmung des biscFv2429 mit Hilfe der Oberflächenplasmonresonanz ...33
2.3.9 Konzentrationsbestimmung des scFv4813 ............................................................................33

ii
2.3.9.1 Konzentrationsbestimmung von scFv4813 mit Hilfe der Oberflächenplasmonresonanz 33
2.3.9.2 Photometrische Konzentrationsbestimmung .................................................................... 34
2.3.9.3 Konzentrationsbestimmung durch Bradford-Färbung...................................................... 34
2.3.10 Reinigung von scFv4813 aus Pichia-Fermentationsüberstand............................................. 35
2.3.10.1Metall-Affinitätschromatographie.................................................................................... 35
2.3.10.2Kationenaustauscherchromatographie.............................................................................. 36
2.3.10.3Aufkonzentrierung des scFv4813..................................................................................... 36
2.3.10.4Gelchromatographie ......................................................................................................... 37
2.4 Kultivierung von Nicotiana tabacum cv BY-2.............................................................................. 37
2.4.1 Stammhaltung....................................................................................................................... 37
2.4.2 Vorkultur .............................................................................................................................. 37
2.4.3 Bestimmung von Frischgewicht, Trockengewicht und Zellanteil in BY-2 Kulturen ........... 38
2.4.4 Fermentation von Nicotiana tabacum cv BY-2 .................................................................... 38
2.4.4.1 Fermentationen im 5-Liter Maßstab................................................................................. 38
2.4.4.2 Fermentationen im 30-Liter Maßstab............................................................................... 38
2.5 Kultivierung von Pichia pastoris .................................................................................................. 39
2.5.1 Kryokulturen......................................................................................................................... 39
2.5.2 Vorkulturen........................................................................................................................... 40
2.5.3 Bestimmung von optischer Dichte, Frisch- und Trockengewicht ........................................ 40
2.5.4 Expression im Schüttelkolben .............................................................................................. 41
2.5.5 Fermentation von Pichia pastoris......................................................................................... 41
2.5.5.1 Vorbereitung und erste Phase der Fermentation .............................................................. 41
2.5.5.2 Glyzerin-Zulaufverfahren................................................................................................. 42
2.5.5.3 Methanol-Zulaufverfahren ............................................................................................... 43
2.5.5.4 Induktion mit Echtzeit-Methanol-Kontrolle..................................................................... 44
3 Ergebnisse und Diskussion.............................................................................................................. 46
3.1 Entwicklung eines Fermentationsprotokolls für Nicotiana tabacum cv BY-2.............................. 46
3.1.1 Wachstumskinetik von Nicotiana tabacum cv BY-2 im Rührreaktor .................................. 46
3.1.2 Nährstoffaufnahmekinetik .................................................................................................... 49
3.1.3 Kinetik der Expression des biscFv2429................................................................................ 50
3.1.4 Versuche zur Stimulation des CaMV 35SS Promotor........................................................... 52
3.2 Produktion von scFv4813 in Nicotiana tabacum cv BY-2............................................................ 54
3.2.1 Produktion von scFv 4813 im 5-Liter Maßstab .................................................................... 54
3.2.2 Maßstabsvergrößerung in den 30-Liter Maßstab.................................................................. 55
3.3 Entwicklung eines Fermentationsprotokolls für Pichia pastoris................................................... 57
3.3.1 Wachstumskinetik und Medienoptimierung im Schüttelkolben........................................... 57
3.3.2 Bestimmung des Methanolverwertungs-Phänotpys des Expressionsklons .......................... 59
3.3.3 Methanol-Toleranz von Pichia pastoris im Schüttelkolben ................................................. 60
3.3.4 Wachstumskinetik im Fermenter und Optimierung der Fermentationsstrategie .................. 61
3.3.5 Maßstabsvergrößerung in den 30-Liter Maßstab.................................................................. 64
3.3.6 Optimierung der Methanol-Zufuhrsteuerung........................................................................ 65
3.3.6.1 Messung der Methanolkonzentration bei manueller Steuerung der Zufuhrrate ............... 66
3.3.6.2 Steuerung der Methanolzufuhr über die Gelöstsauerstoffkonzentration .......................... 68

iii
3.3.6.3 Methanolanalytik und –steuerung in Echtzeit ...................................................................69
3.3.6.4 Glyzerinsupplementierung während der Induktionsphase ................................................70
3.4 Produktion von scFv4813 in Pichia pastoris .................................................................................75
3.4.1 Produktion von scFv4813 im 5-Liter Maßstab......................................................................75
3.4.2 Produktion des scFv4813 im 30-Liter Maßstab.....................................................................76
3.5 Reinigung von scFv4813 im Gramm-Maßstab ..............................................................................77
3.5.1 Zellabtrennung.......................................................................................................................77
3.5.2 Metall-Affinitätschromatographie .........................................................................................77
3.5.3 Ionenaustauscherchromatographie ........................................................................................79
3.5.4 Gelchromatographie ..............................................................................................................80
3.5.5 Konzentrationsbestimmung des scFv4813 ............................................................................81
3.5.6 Stabilität und Lagerung von scFv4813..................................................................................82
4 Abschlußdiskussion und Ausblick ..................................................................................................84
4.1 Produktion des scFv4813 durch Fermentation von Nicotiana tabacum cv BY-2..........................84
4.2 Produktion des scFv4813 durch Fermentation von Pichia pastoris...............................................86
4.2.1 Fermentationsstrategie...........................................................................................................86
4.2.1.1 Medium .............................................................................................................................86
4.2.1.2 Glyzerin-Zufuhrphase .......................................................................................................87
4.2.1.3 Methanol-Zufuhrphase ......................................................................................................88
4.2.1.4 Vergleich mit anderen in Pichia pastoris exprimierten scFvs ..........................................90
4.3 Reinigung und Lagerung des in Pichia produzierten scFv4813 ....................................................91
4.4 Vergleich der Expressionssysteme unter Einbeziehung ökonomischer Gesichtspunkte................92
5 Zusammenfassung ............................................................................................................................95
6 Literatur ............................................................................................................................................97


Einleitung
1
1 Einleitung
1.1 Antikörper und Antikörperfragmente
1.1.1 Rolle, Aufbau und Struktur von Antikörpern
Der zentrale Vorgang in der Immunreaktion eines Vertebraten auf eine Infektion ist die
Erkennung des Erregers als Fremdstoff. Diese Erkennung erfolgt auf molekularer Ebene durch
die Bindung von Antikörpern an Antigene. Antikörper gehören zur Familie der Immunglobuline
(Ig) und werden als Hauptbestandteil der humoralen Immunantwort von Vertebraten durch B-
Lymphozyten gebildet, die jeweils nur Antikörper einer einzigen Spezifität auf ihrer Oberfläche
tragen. Der Kontakt eines B-Lymphozyten mit "seinem" Antigen löst die Proliferation und
Differenzierung dieser Zelle aus. Die im Rahmen dieser Reaktion entstehenden Plasmazellen
sezernieren nun eine große Zahl löslicher Antikörper der entsprechenden Spezifität in das Blut
und die lymphatischen Organe. Die Inaktivierung der Erreger erfolgt durch Komplexierung und
Markierung der Antigene, die wiederum weitere Komponenten des zellulären Immunsystems
aktiviert.
Die Vielfalt der Antikörper und die dadurch erreichbare Spezifität der Erkennungsreaktion
wird auf genetischer Ebene erreicht, indem die Gensequenzen, die für die Erkennungsstelle eines
Antikörpers kodieren, bei der Reifung der B-Lymphozyten aus einigen hundert Gensegmenten
zufällig zusammengesetzt werden.
Man unterscheidet 5 verschiedene Klassen oder Isotypen von Immunglobulinen, die sich
durch verschiedene schwere Ketten auszeichnen: IgA, IgG, IgM, IgD und IgE. Die einfachste
und im Plasma am stärksten vertretene Klasse ist das IgG. IgG Antikörper setzen sich aus vier
Polypeptidketten zusammen, zwei identischen leichten Ketten und zwei identischen schweren
Ketten. Die einzelnen Ketten sind modular aus ca. 110 Aminosäuren enthaltenden Domänen
aufgebaut, wobei die leichten Ketten jeweils zwei und die schweren Ketten jeweils vier solcher
Domänen enthalten (Schiffer et al., 1973). Die N-terminalen Domänen der Ketten enthalten
jeweils drei hypervariable Regionen (complementarity determining regions, CDRs) und werden
als variable Domänen (VL bzw. VH, entsprechend ihrer Zugehörigkeit zur leichten bzw.
schweren Kette) bezeichnet (Kabat et al., 1990). Die CDRs jeweils einer leichten und einer
schweren Kette bilden die eigentliche Antigenerkennungsstelle (Paratop) (Jones et al., 1986;
Verhoeyen et al., 1988). Abb. 1-1 zeigt den Aufbau eines IgG-Antikörpers aus den vier Ketten

Einleitung
2
sowie die Disulfidbrücken, die die schwere und leichte Kette und die beiden schweren Ketten
verbinden, und die Glykosylierungsstellen.
1.1.2 Modifizierte Antikörper und Antikörperfragmente
Mit der Einführung der Hybridoma-Technologie (Köhler und Milstein, 1975) die es erlaubt,
B-Lymphozyten zu immortalisieren und zu klonieren, wurde es möglich, gezielt größere Mengen
monoklonaler Antikörper mit definierter Spezifität zu produzieren. Damit gewannen Antikörper
rasch an Bedeutung für Nachweisverfahren in der Molekularbiologie und Biochemie sowie für
diagnostische und therapeutische Anwendung in der Medizin. Antikörper können, ohne ihre
Affinität oder Spezifität für das Antigen zu verlieren, kovalent an andere Moleküle gebunden
werden, wodurch eine ganze Generation von Antikörper-vermittelten Nachweisverfahren
entstanden ist. Beispiele dafür sind die Kopplung von Enzymen wie alkalische Phosphatase
(AP), an Fluoreszenzfarbstoffe oder andere Stoffe, die qualitative oder quantitative
Nachweisverfahren ermöglichen. Im therapeutischen Bereich lassen sich durch Gabe von
spezifischen Antikörpern Giftstoffe in der Blutbahn neutralisieren oder es lassen sich an einen
Antikörper gekoppelte Effektoren in die räumliche Nähe von z. B. Krebszellen dirigieren
(Colcher et al., 1990; Wong et al., 1995). Dabei ist die Größe eines Vollängen-IgG und seine
Eigenschaft, zwei Bindungsstellen für ein Epitop zu besitzen, teilweise überflüssig oder sogar
hinderlich, so daß kleinere, monovalente Fragmente interessante Alternativen darstellen.
IgG-Antikörper lassen sich proteolytisch in mehrere funktionelle Untereinheiten zerlegen
(Eyk et al., 1967), beispielsweise in die Fc und F(ab)'2–Fragmente (Parham, 1986), Fab-
Fragmente (Porter, 1959) oder Fv-Fragmente (Inbar et al., 1972; Hochman et al., 1973). Abb.
1-1 zeigt die von IgG abgeleiteten Antikörperfragmente.

Einleitung
3
S SS S
COOH
Antigen-Bindungsstelle
VariableDomänen
CDRs
KonstanteDomänen
IgG-Antikörper
Glykosylierungs-Stelle
"Linker"-Peptide
"diabody""bi-scFv"
Fv-FragmentFab-Fragment
S SS S
F(ab)'2-Fragment Fc-Fragment
"Linker"-peptid
"scFv"-Fragment
"Linker"-Peptid
"Linker"-Peptid
Abb. 1-1: IgG-Antikörper und abgeleitete Fragmente: Fc: Kristallisierbares Fragment; Fab:Antigen-bindendes Fragment; F(ab)'2: kovalent verbundenes Dimer des Fab, bivalent; Fv:Fragment der variablen Region, scFv: Einzelketten-Fragment der variablen Region; bi-scFv:bivalentes, bispezifisches Einzelkettenkonstrukt; diabody: aus zwei aggregierten Einzelkettenbestehendes, bivalentes, bispezifisches Konstrukt.
Das kleinste proteolytisch herstellbare antigenbindende Fragment, das Fv-Fragment, ist in
niedrigen Konzentrationen relativ instabil. Mit molekularbiologischen Methoden ist es aber
möglich durch Hinzufügen eines, 10-30 Aminosäuren langen Verbindungspeptids ("Linker") auf
genetischer Ebene ein Konstrukt herzustellen, das ein aus nur einer Kette bestehendes
Antikörperfragment kodiert, das bei geringer Größe (ca. 250 Aminosäuren) gute Spezifität und
Affinität für sein Epitop aufweist (Bird et al., 1988; Huston et al., 1988; Winter und Milstein,
1991). Diese scFv-Fragmente (Abb. 1-1) sind im Vergleich zu den etwa gleich großen Fv-
Fragmenten stabiler und lassen sich molekularbiologisch gut handhaben. Weiterentwicklungen
der scFvs sind die ebenfalls auf genetischer Ebene erzeugten bivalenten und bispezifschen scFvs
(Mallender und Voss, 1994; Schumann, 1996; Fischer et al., 1999e), "minibodies" (Hu et al.,
1996) oder "diabodies" (Perisic et al., 1994; Adams et al., 1998; Atwell et al., 1999) (Abb. 1-1).
Auch Fusionsprodukte von Antikörperfragmenten mit z. B. Enzymen oder Toxinen lassen sich

Einleitung
4
heute auf genetischem Weg herstellen (Tai et al., 1990; Boleti et al., 1995; Harper et al., 1997;
Luo et al., 1998). Eine Verbesserung der Stabilität von scFvs und "diabodies" läßt sich über das
Einfügen von Cysteinresten auf genetischer Ebene erreichen, die Produkte können dann über
Disulfidbrücken quervernetzt werden (FitzGerald et al., 1997; Govardhan, 1999).
1.1.3 Bedeutung rekombinanter Antikörper und Antikörperfragmente
Polyklonale Seren und monoklonale Antikörper sowie die daraus abgeleiteten Fragmente und
Fusionsproteine (1.1.2) sind die Grundlage zahlreicher proteinchemischer und
molekularbiologischer Nachweisverfahren, beispielsweise für den "Enzyme-linked
immunosorbent assay" (ELISA) oder die Immunfärbung zur Detektion geblotteter Proteine bzw.
Nukleinsäuren (z. B. DIG-Markierung).
Daneben werden rekombinante Antikörperfragmente zunehmend für therapeutische
Applikationen genutzt, beispielsweise ein Fab-Fragment (Abb. 1-1), das bei Myasthenia-gravis-
Patienten die Acetylcholinrezeptoren vor dem Angriff körpereigener Antikörper schützt (Graus
et al., 1997), scFv-Toxin Fusionsproteine (Reiter et al., 1996) oder bispezifische Antikörper, die
einerseits bestimmte Tumorantigene binden und mit der zweiten Spezifität Killerzellen des
Immunsystems rekrutieren und aktivieren (Holliger et al., 1996; Helfrich et al., 1998;
Kipriyanov et al., 1998; Arndt et al., 1999). Die Vorteile von Antikörperfragmenten liegen bei
solchen Anwendungen in der besseren Durchdringung solider Tumore im Vergleich zu
Vollängenantikörpern. Dabei haben scFvs im Vergleich zu Fab-Fragmenten den Vorteil, sich bei
guter Tumorerkennung nicht in den Nieren anzureichern (Colcher et al., 1990). Die Stabilität
von scFvs in humanem Plasma ist ausreichend um einen therapeutischen Effekt zu erzielen, sie
läßt sich aber auch durch weitere Modifikationen verbessern (FitzGerald et al., 1997; Kipriyanov
et al., 1999).
1.2 Expressionssysteme für rekombinante Proteine
Zur Expression rekombinanter Proteine stehen verschiedene Expressionssysteme zur
Auswahl. Das Spektrum reicht von Bakterien über einfache Eukaryonten bis hin zu Pflanzen-,
Insekten- oder Säugerzellkulturen. Bei der Entscheidung für ein bestimmtes System spielt im
Labormaßstab häufig die erwartete Expressionsrate eine entscheidende Rolle, daneben Herkunft,
Eigenschaften und Komplexität des zu exprimierenden Proteins sowie die Verfügbarkeit
bestimmter Technologien.
Bei der Prozeßentwicklung für den industriellen Maßstab stehen andere Fragestellungen im
Vordergrund. Neben der Expressionsrate gehen z. B. der Jahresbedarf, Kosten für Medien und

Einleitung
5
Aufarbeitung/Reinigung des Produktes, die Patentsituation, die Durchführbarkeit aller
Arbeitsschritte im großen Maßstab und die Erfüllung entsprechender Zulassungsanforderungen
in die Überlegungen ein, um das günstigste, sicherste, am besten kontrollierbare und am
schnellsten zu einem marktfähigen Produkt führende Expressionssystem ist.
1.2.1 Produktion rekombinanter Proteine in Bakterien
Das gramnegative Bakterium Escherichia coli stellt den wichtigsten bakteriellen
Wirtsorganismus für die Expression von rekombinanten Proteinen dar. Dahinter steht eine große
Zahl von erfolgreichen Expressionen bis in den industriellen Maßstab und entsprechende
Erfahrung in der genetischen Manipulation und Kultivierung dieses Organismus (Baneyx, 1999).
Die genetische Information wird häufig auf autonom replizierten Plasmiden mit relaxierter
Kontrolle der Kopienzahl in die Bakterien eingebracht. Das führt unter Umständen bei
Kultivierung ohne Selektionsdruck zum Verlust der Plasmide. Außerdem ist die Größe der in
Plasmiden einsetzbaren Fremd-DNA nach oben begrenzt. Solche Probleme lassen sich durch
Verwendung von integrativen Vektoren bzw. durch Verwendung von auxotrophen
Wirtsstämmen mit Komplementation des defekten Gens auf dem Plasmid umgehen (Baneyx,
1999).
Ein vor allem bei der Verwendung starker Promotoren auftretendes Problem hängt mit der
engen Kopplung von Transkription und Translation bei Prokaryonten zusammen: Die
Fremdproteine liegen zwar in großer Menge, aber oftmals nicht in löslicher Form, sondern als
sogenannte Einschlußkörper ("inclusion bodies") vor. In diesem Fall wird die häufig mit großen
Ausbeuteverlusten und Zeit- und Kostenaufwand verbundene Rückfaltung zum löslichen Protein
notwendig.
Die Ausbildung von Disulfidbrücken erfolgt in E. coli infolge des reduzierenden Milieus des
Cytoplasmas und des Vorhandenseins von Thioredoxinen nicht oder nicht in der gewünschten
Weise. Ebensowenig kann E. coli posttranslationale Glykosylierungen ausführen.
Trotz dieser Probleme und Hindernisse stellt die Expression einfacher rekombinanter
Proteine in E. coli einen für industrielle Anwendungen und Jahresproduktionsmengen bis
1000 kg gangbaren Weg dar, was aber zu großen Teil darin begründet liegt, daß die Prozesse und
Arbeitsschritte im industriellen Maßstab bekannt und in regulatorischer Hinsicht abschätzbar
sind (Dr. Minuth, Bayer AG, pers. Mitteilung).

Einleitung
6
1.2.2 Produktion rekombinanter Proteine in tierischen Zellkulturen
Insekten- und Säugetierzellkulturen spielen vor allem für die Produktion von glykosylierten
Proteinen eine Rolle, da diese Systeme die größte Wahrscheinlichkeit für die korrekte
Glykosylierung von heterolog exprimierten Säugetierproteinen bieten (Altmann et al., 1999;
Kost und Condreay, 1999). Für die häufig verwendeten Zellinien wie z. B. Sf9 (Spodoptera
frugiperda) oder CHO ("Chinese hamster ovary") existieren Protokolle, die unter Verwendung
viraler Vektoren oder Plasmide die Expression rekombinanter Proteine im Maßstab bis ca. 1000 l
mit Ausbeuten von 1-20 mg·l-1 erlauben (Wurm und Bernard, 1999). Damit eignen sich tierische
Zellkulturen für Produkte, deren Jahresbedarf 200 kg nicht übersteigt (Dr. Minuth, Bayer AG,
pers. Mitteilung). Die Gefahr einer Kontamination des Produktes durch Humanpathogene ist
allerdings bei tierischen Zellkulturen besonders hoch und erfordert daher zusätzlichen Aufwand
bei der Reinigung des Produktes. Dadurch und durch die aufwendigere Technik und die teureren
Medien liegen die Produktionskosten für ein rekombinantes Protein in tierischer Zellkultur um
den Faktor 2-20 höher als bei mikrobiellen Expressionssystemen.
1.2.3 Produktion rekombinanter Proteine in pflanzlichen Expressionssystemen
Die Transformation von Pflanzen erfolgt hauptsächlich mit Hilfe von Agrobacterium
tumefaciens durch rekombinante Ti-Plasmide oder über ballistische Transformation mit Hilfe
von "particle guns". Als Promotoren werden häufig der 35S-Promoter des "Cauliflower Mosaic
Virus" (CaMV), davon abgeleitete Sequenzen oder andere pflanzliche Promotoren verwendet.
Mittlerweile sind organspezifische Promotoren bekannt, die beispielsweise die gezielte
Expression eines heterologen Proteins in Samen (Fiedler und Conrad, 1995; Parmenter et al.,
1995; Cramer et al., 1999) oder in Kartoffelknollen (Mason et al., 1992; Mason und Arntzen,
1995; Richter et al., 1996; Fiedler et al., 1997; Arakawa et al., 1999) erlauben. Dazu kommt die
Möglichkeit, Proteine in bestimmten Zellkompartimenten anzureichern, z. B. im Cytosol
(Schillberg et al., 1999, Spiegel, 1999 #218), in der Vakuole (Dombrowski und Raikhel, 1996;
Hoppmann, 2000), im endoplasmatischen Retikulum (ER) (Denecke et al., 1992; Vitale et al.,
1993, Spiegel, 1999 #218; Artsaenko et al., 1995), in den Chloroplasten (Staub und Maliga,
1992) oder in den Apoplasten zu sekretieren (Schillberg et al., 1999; Ziegler et al., 2000). Häufig
zeigt sich, daß unabhängig vom Promotor die Akkumulation im ER besonders hohe Ausbeuten
ergibt (Fiedler et al., 1997).
Die Hauptvorteile von Pflanzen als Expressionssystem zur Produktion rekombinanter
Proteine liegen in der preisgünstigen Kultivierung von Pflanzen im Gewächshaus- oder
Freilandanbau und den praktisch unbegrenzten Möglichkeiten der Maßstabsvergrößerung der

Einleitung
7
Produktion (Ma und Hein, 1995b; Herbers und Sonnewald, 1999; Hood und Jilka, 1999; Fischer
et al., 1999c; Fischer et al., 1999d). In der Kultivierung, Ernte und Aufarbeitung selbst größter
Mengen Biomasse kann auf die im Nahrungsmittelbereich etablierte Prozessierungstechnologie
zurückgegriffen werden (Kusnadi et al., 1998), so daß in den letzten Jahren der Begriff
"Molecular Farming" geprägt wurde. Die Expression rekombinanter Proteine in Pflanzen hat
mehrere Ziele z. B. die Veränderung der Eigenschaften von Pflanzen oder Pflanzenteilen oder
die Produktion von industriellen Proteinen, Impfstoffen und Antikörpern.
Der Begriff "Industrielle Proteine" beschreibt Proteine, die in großen Mengen, aber nicht für
therapeutische Anwendungen benötigt werden. Dazu zählen beispielsweise Invertasen,
Amylasen, Proteasen, Phytasen und Zellulasen, aber auch nicht-enzymatische Proteine: (Pen et
al., 1992) zeigten die Expression einer bakteriellen Amylase in Tabaksamen und die direkte
Anwendbarkeit der transgenen Samen für die Stärkeverflüssigung. Als weitere Beispiele können
die Expression einer Phytase in Nicotiana tabacum (Tabak) (Verwoerd et al., 1995), in Glycine
max (Soja) (Denbow et al., 1998), und einer β−Glucanase in Hordeum vulgare (Gerste) (Jensen
et al., 1996). Die Expression von Avidin und von β-Glucuronidase (aus E. coli) in Zea mays
(Mais) (Kusnadi et al., 1998) wird bereits kommerziell.
Bei der Produktion therapeutisch eingesetzter Proteine spielen neben den ökonomischen
Vorteilen der Produktion in Pflanzen zwei Punkte eine wesentliche Rolle: Das Vorhandensein
eines komplexen eukaryontischen Proteinsyntheseapparates, der dem von tierischen Zellen in
vieler Hinsicht ähnlich ist, und das Nichtvorhandensein von Animal- oder Humanpathogenen,
das die Aufarbeitung und Qualitätskontrolle der rekombinanten Produkte erheblich einfacher und
günstiger macht (Herbers und Sonnewald, 1999; Fischer et al., 1999d).
Für die Produktion von Impfstoffen, die über den Verdauungstrakt aufgenommen werden,
bietet sich die Produktion in eßbaren Pflanzen an (Mason und Arntzen, 1995). Eßbare Impfstoffe
("edible vaccines") haben den Vorteil, daß ohne großen Aufwand hinsichtlich der Reinigung
oder Formulierung, ein zur Verabreichung geeignetes Produkt entsteht. Dadurch ließen sich
ausgesprochen preisgünstige Impfpräparate realisieren. Verschiedene virale Antigene wurden
bereits in Pflanzen exprimiert und auf ihre Wirksamkeit bei Tieren getestet (Mason et al., 1996;
Yusibov et al., 1997; Modelska et al., 1998), in der Humanmedizin befinden sich in Kartoffeln
und Mais produzierte bakterielle Antigene, z. B. das Choleratoxin (Arakawa et al., 1999) und das
hitzelabile E. coli Enterotoxin (Mason et al., 1998) sowie ein in Lupine und Salat (Lactuca
sativa) exprimiertes Hepatitis B-Oberflächenantigen (Mason et al., 1992; Kapusta et al., 1999) in
präklinischen bzw. klinischen Phase-I Studien (Hood und Jilka, 1999).

Einleitung
8
Die Produktion von Antikörpern in Pflanzen (Hiatt et al., 1989; Düring et al., 1990; Fischer
et al., 1999c) ist neben der Vermittlung von Resistenzen durch die Herstellung von Antikörpern
zur passiven Immunisierung motiviert (Hiatt und Ma, 1992; Ma und Hein, 1995b; Fischer et al.,
1999d). Die besonderen Vorteile der pflanzlichen Expressionsysteme liegen dabei in der
korrekten Expression komplexer Antikörper, wie anhand von auf genetischer Basis erzeugten
Fv-Antikörpern (Benvenuto et al., 1991), scFv-Antikörpern (Fecker et al., 1996; Fiedler et al.,
1997; Schillberg et al., 1999), bispezifischen Antikörpern (Fischer et al., 1999e), Fab-
Antikörpern (De Wilde et al., 1998; De Neve et al., 1999), Vollängen IgG Antikörpern (Hiatt et
al., 1989; Schillberg et al., 1999; Fischer et al., 1999f), sekretorischen IgA-Antikörpern (Ma et
al., 1994; Ma et al., 1995a) und Antikörper-Fusionsproteinen (Spiegel et al., 1999) gezeigt
wurde. Ein Grund dafür ist die offensichtliche strukturelle und funktionelle Homologie
pflanzlicher Chaperone zum BiP ("Binding Protein") aus Säugetieren (Denecke et al., 1991;
Fontes et al., 1991). Die Einsetzbarkeit solcher in Pflanzen erzeugter Antikörper wurde anhand
eines in Tabak produzierten murinen Antikörpers gegen ein Streptococcus mutans
Adhäsionsprotein bereits gezeigt: Der rekombinante Antikörper war im Mund von Probanden
mehrere Tage stabil und verhinderte die Kolonisierung dentaler Oberflächen durch den
Karieserreger (Ma et al., 1998). Ein in Glycine max produzierter Antikörper gegen Herpes
simplex verhinderte im Mausmodell die Bildung von Läsionen und die Ausbreitung des Virus
(Zeitlin et al., 1998), die Einsatzfähigkeit eines in Nicotiana tabacum produzierten Antikörpers
gegen ein Darmkrebs-assoziiertes Oberflächengantigen wird noch untersucht (Verch et al.,
1998). Vaquero et al. beschrieben die Herstellung und Charakterisierung eines therapeutisch
relevanten, chimären Vollängen-Antikörpers und eines davon abgeleiteten scFv in Nicotiana
tabacum durch transiente Expression in Blattmaterial (Vaquero et al., 1999). Die Eignung von
Reis, Reis-Suspensionskulturen und Weizen zur Produktion therapeutisch relevanter
Antikörperfragmente ist ebenfalls beschrieben (Torres et al., 1999; Stoger et al., 2000).
Neben intakten Pflanzen spielen Pflanzensuspensionskulturen für die Expression
rekombinanter Proteine eine Rolle. Pflanzenteile, typischerweise kreisförmige Ausstiche aus
Blättern ("leaf disks"), werden oberflächensterilisiert und auf für pflanzliche Gewebekulturen
geeigneten festen, Nährböden ausgelegt. Nach Einstellung bestimmter Pflanzenhormonkonzen-
trationen entstehen aus den Blattscheiben undifferenziert wachsende Zellklumpen, sog. "Kalli".
Das Wachstum erfolgt dabei heterotroph, als Kohlenstoff- und Energiequelle werden z. B.
Saccharose, Maltose oder Glukose in 1-4prozentiger Ausgangskonzentration verwendet.
Pflanzensuspensionskulturen werden erzeugt, indem solche Kalli in Flüssigmedien überführt und
unter Schütteln inkubiert werden (Scragg, 1997). Die Zellen teilen sich weiter, und aus den 3-10

Einleitung
9
mm großen Kalli entstehen unter Einwirkung von Scherkräften Suspensionen von Aggregaten
von ca. 0.1 bis 3 mm, die aus ca. 2-100 Zellen bestehen (Taticek et al., 1991; Scragg, 1997). Die
Größe dieser Aggregate ist art- und zellinienspezifisch und kann sich mit fortgesetzter
Subkultivierung in Flüssigkultur hin zu kleineren Aggregaten entwickeln. Die einzelnen Zellen
weisen sphärische bis langgestreckte Formen mit Durchmessern von bis zu 50 µm und Längen
bis ca. 300 µm auf.
Das Wachstum von Pflanzenzellkulturen ist im Vergleich zu mikrobiellen Systemen
langsam. Bei Generationszeiten oberhalb von 24 h sind entsprechend lange Kultivierungszeiten
von ca. 7-21 Tagen erforderlich. Durch die starke Wasseraufnahme und Vakuolisierung der
Pflanzenzellen gegen Ende der Wachstumsphase werden hohe Zelldichten bis ca. 300 g·l-1
(Frischgewicht) erreicht, das entpricht einem Feststoffgehalt um 50 %. Aufgrund des langsamen
Wachstums liegt der trockengewichtsspezifische Sauerstoffbedarf von Pflanzensuspensions-
kulturen bei etwa 0,2 bis 0,6 mmol·g-1·h-1, das entspricht bei Trockenmassekonzentrationen von
maximal 10-20 g·l-1 einer Sauerstoffübertragungsrate von maximal 12 mmol·l-1·h-1 (Singh und
Curtis, 1995). Das ist im Vergleich zu den für mikrobielle Fermentationen notwendigen
Sauerstoffübertragungsraten von bis zu 90 mmol·l-1·h-1 (Taticek et al., 1991) wenig, und
ermöglicht die Prozessführung mit Begasungsraten unter 0,3 l·l-1·min-1 und bei niedrigen
Rührerdrehzahlen.
Im Schüttelkolben stellen diese Rahmenbedingungen kein Problem dar, bei der
Maßstabsvergrößerung in den Fermenter ist es aber nötig, diesen gegenüber Mikroorganismen
stark abweichenden Anforderungen Rechnung zu tragen. Der niedrige Sauerstoffbedarf macht
das Zerschlagen eingetragener Luftblasen zur Oberflächenvergrößerung unnötig. Das kommt der
Rücksichtnahme auf die infolge der Größe möglicherweise bestehende Scherempfindlichkeit der
Zellen entgegen und führt dazu, daß nur selten die in der Mikrobiologie üblichen Turbinenrührer
eingesetzt werden. Statt dessen kommen großflächige Blattrührer oder ähnliche Systeme mit
vergleichsweise niedrigen Drehzahlen zum Einsatz, oder die Durchmischung wird durch
alternative Reaktorbauweisen erreicht. Tabelle 1-1 gibt einen Überblick über die verwendeten
für Pflanzensuspensionskulturen verwendeten Bioreaktortypen:

Einleitung
10
Tabelle 1-1: Beispiele für mit Pflanzensuspensionskulturen verwendete Reaktortypen
Organismus Reaktortyp Maßstab [l] LiteraturstelleDigitalis lanata Rührkessel, Gitterrührer 27 (Spieler et al., 1985)
Echinacea purpurea,Rauwolfia serpentina
Rührkesselkaskade,INTERMIG -Rührer
75000 (Rittershaus et al., 1989)
Dioscorea deltoidea,Catharanthus roseus
Rührkessel, Rushton-Scheibenblattrührer
14 (Drapeau et al., 1986)
Catharanthus roseus Rührkessel, Helixrührer 11 (Jolicoeur et al., 1990)
Catharanthus roseus Rührkessel, Rushton-Scheibenblattrührer
7,5 (Leckie et al., 1991)
Lithospermumerythrorhizon
Drehtrommelreaktor 1 (Tanaka et al., 1983)
Digitalis lanata Airlift Schlaufenreaktor 20 (Spieler et al., 1985)
Eschscholtzia californica Airlift Schlaufenreaktor 2,3 (Byun und Pedersen, 1994)
Mittlerweile hat man erkannt, daß die Annahme, Pflanzensuspensionskulturen seien
scherkraftempfindlich, nicht auf alle Kultivare zutrifft. Vielmehr sind die meisten Zellinien trotz
der Größe der Zellen und Aggregate schertolerant (Taticek et al., 1991; Doran, 1993).
Problematischer als die Scherkraftempfindlichkeit ist die Durchmischung der Kulturen, die
gegen Ende einer Fermentation einen hohen Feststoffanteil und entsprechend hohe apparente
Viskositäten aufweisen. Im Falle von Rührreaktoren erfordert dies den Einsatz relativ
großflächiger, langsam drehender Rührer (Hooker et al., 1990; Kim et al., 1991; Scragg, 1991;
Scragg, 1992; Doran, 1993; Singh und Curtis, 1995). Die Steuerung der Belüftungsrate von
pflanzlichen Zellkulturen ist insofern kritisch, als die Löslichkeit des Pflanzenhormons CO2 bei
zu hoher Belüftungsrate verringert wird, was zu schlechterem Wachstum führt (Smart und
Fowler, 1981; Ducos und Pareilleux, 1986).
Das ursprüngliche Interesse an Pflanzensuspensionskulturen lag weniger in der Expression
rekombinanter Proteine als in der Mikropropagation von Nutzpflanzen (aus Suspensionskulturen
können Kalli und daraus wiederum intakte Pflanzen regeneriert werden) und der Gewinnung von
Sekundärmetaboliten und Aromen (Dörnenburg und Knorr, 1995; Scragg, 1997). Einige
Beispiele für die Produktion von Sekundärmetaboliten in Pflanzensuspensionskulturen sind in
Tabelle 1-2 genannt.

Einleitung
11
Tabelle 1-2: Beispiele für die Produktion von Sekundärmetaboliten in Pflanzenzellkulturen
Organismus Produkt Anwendungsbereich Literaturstelle
Catharanthus roseus Ajmalicin Therapeutikum(Steroidvorstufe)
(Drapeau et al., 1986)
Dioscorea deltoidea Diosgenin Therapeutikum(Bluthochdruck)
(Drapeau et al., 1986)
Eschscholtzia californica Benzophenanthridin Therapeutikum(Zahnmedizin)
(Byun und Pedersen, 1994)
Fragaria ananassa Anthocyanin Farbstoff (Sato et al., 1996)
Taxus spec. Taxol (Paclitaxel) Therapeutikum(Cytostatikum)
(Ketchum et al., 1999)
Mittlerweile setzt sich die Erkenntnis durch, daß Pflanzensuspensionskulturen auch ein
attraktives Expressionssystem für komplexe rekombinante Proteine, insbesondere für
pharmazeutische Anwendungen sein können. Die Gründe dafür sind neben den bereits
angeführten allgemeinen Vorteilen pflanzlicher Expressionssysteme die Möglichkeit, große
Mengen an Biomasse in Fermentern schnell, unter exakt reproduzierbaren Bedingungen sowie
unter Einhaltung strenger Qualitätsmaßstäbe (GMP) produzieren zu können. Diese Erkenntnis
und das zunehmende Interesse an der Expression rekombinanter Proteine in
Pflanzensuspensionskulturen spiegelt sich in der neueren Literatur wieder (Tabelle 1-3).
Tabelle 1-3: Beispiele für die Produktion von rekombinanten Proteinen in Pflanzenzellkulturen
Organismus Produkt Anwendungsbereich Literaturstelle
Glycine max Phytase Viehfutterverbesserung(Phosphatverfügbarkeit)
(Li et al., 1997)
Oryza japonica hAAT Therapeutikum (Elastasehemmer) (Terashima et al., 1999)
Nicotiana tabacum mGM-CSF Therapeutikum (Cytokin) (Lee et al., 1997)
Nicotiana tabacum cv BY-2 GUS Reporterenzym (Shinmyo et al., 1998)
Nicotiana tabacum cv BY-2 Invertase Reporterenzym (des Molles et al., 1999)
Nicotiana tabacum scFv Modellantikörper (Firek et al., 1993)
Nicotiana tabacum rAk Modellantikörper (Magnuson et al., 1996)
Nicotiana tabacum cv BY-2 rAb Modellantikörper (Sharp und Doran, 1999)
Nicotiana tabacum cv BY-2 biscFv Virusresistenz in N. tabacum (Fischer et al., 1999e)
Nicotiana tabacum cv SR-1 rAK Virusresistenz in N. tabacum (Fischer et al., 1999f)
Nicotiana tabacum Ricin Therapeutikum (Sehnke und Ferl, 1999)
Nicotiana tabacum Bryodin Therapeutikum
(Immunotoxin, RIP)
(Francisco et al., 1997)
Eine besondere Rolle kommt den Suspensionskulturen von Nicotiana tabacum zu,
insbesondere der seit 1972 als Suspensionskultur geführten Zellinie BY-2. Dieses Nikotin-freie
Kultivar wächst mit einer Generationszeit von unter 24 h relativ schnell und mit hoher
Synchronizität der Zellteilung. Zudem bildet BY-2 nur kleine Aggregate von ca. 1 mm

Einleitung
12
Durchmesser, so daß verfahrenstechnisch einfach zu behandelnde, feine Suspensionen vorliegen
(Nagata et al., 1992). Obwohl BY-2 Zellen sich nicht mehr zu intakten Pflanzen regenerieren
lassen, machen diese Vorteile sowie die leichte Transformierbarkeit durch einfache Co-
Kultivierung mit rekombinanten Agrobakterien (Fischer et al., 1999e) diese Zellinie zu einem
geeigneten Objekt für die weitere Entwicklung von Applikationen der Pflanzensuspensions-
kultur.
Wie in intakten Pflanzen zeigte sich auch in Suspensionskulturen, daß die Retention der
rekombinanten Proteine im ER über die C-terminalen Signalpeptide H/R/KDEL die höchsten
Ausbeuten erbringen (Fischer et al., 1999a; Doran, 2000). Die erreichten Expressionsraten sind
allerdings mit 0,2 bis 2 % des Gesamtproteins noch unbefriedigend. Promotorstudien zeigen, daß
z. B. in Suspensionskulturen von Nicotiana tabacum der 35S Promotor aus CaMV oder der
davon abgeleitete stärkere 35SS (Kay et al., 1987) übertroffen werden kann (Ni et al., 1995;
Fiedler et al., 1997; Shinmyo et al., 1998).
Pflanzen und pflanzliche Suspensionskulturen stellen damit insbesondere für die Produktion
komplexer oder glykosylierter Proteine (Lerouge et al., 1998; Doran, 2000) eine interessante
Alternative dar, die durch die attraktiven Perspektiven hinsichtlich der Maßstabsvergrößerung
zusätzliche Vorteile aufweist. Die mit Pflanzen bisher gezeigten spezifischen Ausbeuten
rekombinanter Proteine bedürfen allerdings für den industriellen Einsatz noch der Verbesserung.
1.2.4 Produktion rekombinanter Proteine in Hefen
Hefen bieten als Wirtsorganismen für die Überexpression rekombinanter Proteine
wesentliche Vorteile: Die Kultivierung von Hefen ist in der Regel einfach. Sie stellen im
Vergleich zu Zellinien höherer Organismen geringe Ansprüche an das Medium und die
Kultivierungsbedingungen. Hefen lassen sich im wesentlichen in Rührkessel-Bioreaktoren mit
Standardausstattung kultivieren. Bei der Maßstabsvergrößerung von Produktionsprozessen kann
auf jahrzehntelange Erfahrungen aus der Nahrungsmittelindustrie zurückgegriffen werden.
Einzellige Organismen lassen sich leicht klonieren und erlauben daher, mit wohldefinierten
Genpools zu arbeiten. Zudem sind Hefen genetisch gut untersucht und Protokolle zu ihrer
Transformation etabliert. Wesentliche Fortschritte in der Aufklärung der genetischen Abläufe in
Eukaryonten wurden an Saccharomyces cerevisiae gemacht (Buckholz und Gleeson, 1991).
Neben S. cerevisiae haben sich aber auch andere Hefen als Expressionssysteme etabliert. Tabelle
1-4 nennt wesentliche Beispiele:

Einleitung
13
Tabelle 1-4: Beispiele für die Verwendung von Hefe als Expressionssysteme
Spezies Promotor Literaturstelle
Saccharomyces cerevisiae GALx (Galaktose-induziert)GAPDH (konstitutiv)
(Clare und Romanos, 1995; Gellissen undHollenberg, 1997; Muller et al., 1998)
Schizosaccharomyces pombe NMT1 (konstitutiv) (Lu et al., 1997)
Pichia pastoris AOX1 (Methanol-induziert)GAP (konstitutiv)
(Sreekrishna et al., 1997; Higgins und Cregg,1998)
Pichia stipitis XYL1 (Xylose-induziert) (Piontek et al., 1998)
Pichia methanolica AUG1 (Methanol-induziert) (Raymond et al., 1998)
Hansenula polymorpha MOX (Methanol-induziert) (Gellissen und Hollenberg, 1997)
Kluyveromyces lactis LAC4 (Lactose-induziert) (Swinkels et al., 1993)
Schwanniomyces occidentalis AMY1 (Stärke-induziert) (Piontek et al., 1998)
Candida boidinii AOD1 (Methanol-induziert) (Sakai et al., 1996)
Yarrowia lipolytica TEF (konstitutiv), XPR2(Pepton-induziert)
(Park et al., 1997; Muller et al., 1998)
verändert, nach (Lin Cereghino und Cregg, 1999)
Mit den Vorteilen der einfachen Handhabbarkeit eines Einzeller-Systems verbindet sich bei
Hefen das Vorhandensein eines eukaryontischen Proteinsyntheseapparates. Hefen können die
notwendigen Schritte zur korrekten posttranslationalen Modifizierung von Proteinen höherer
Organismen durchführen. Dazu gehören proteolytische Prozessierung, Faltung, Bildung von
Disulfidbrücken und Glykosylierung (Luo et al., 1995; Eckart und Bussineau, 1996; Ikegaya et
al., 1997; Miele et al., 1997; Lin Cereghino und Cregg, 1999; Markaryan et al., 1999).
Die methylotrophe Hefe Pichia pastoris hat in den letzten Jahren besondere Aufmerksamkeit
als Expressionssystem erhalten. Das ursprüngliche Interesse der Forschung an Pichia richtete
sich auf die Produktion von Einzellerprotein auf der Basis von Methanol (Higgins und Cregg,
1998). Die Entdeckung des extrem starken, methanol-abhängigen AOX1-Promotors führte dann
zur Entwicklung des Pichia pastoris Expressionssystems, das von Invitrogen (Carlsbad, CA,
USA) kommerziell erhältlich ist. Dieser Promotor steuert in Pichia die Expression des ersten
Enzyms in der Methanol-Verwertungskette, der Alkoholoxidase1, kurz AOX1 (EC 1.1.3.13).
Dieses Enzym katalysiert in besonderen Organellen, den Peroxisomen die Oxidation von
Methanol zu Formaldehyd. FAD dient dabei als Cofaktor, das anfallende toxische
Wasserstoffperoxid wird durch Katalase entgiftet. Das ebenfalls toxische Produkt Formaldehyd
wird über die Kopplung an Glutathion "entschärft" und weiter verwertet (Ellis et al., 1985).
Die Expressionsrate der Alkoholoxidase1 in Pichia ist extrem hoch: Beim Wachstum auf
Methanol als einziger Kohlenstoffquelle hat AOX1 einen Anteil von 30% am gesamten löslichen
Protein (Couderc und Baratti, 1980). Neben der Alkoholoxidase1 existiert die Alkoholoxidase2,
die zwar weitgehend homolog ist, aber aufgrund eines schwächeren Promotors in geringerer

Einleitung
14
Menge exprimiert wird. Sie ermöglicht beim Ausfall des AOX1-Gens trotzdem eine - wenn auch
verlangsamte – Verwertung von Methanol (Ellis et al., 1985; Cregg et al., 1989).
Das Pichia-Expressionssystem basiert im wesentlichen auf der Verwendung des AOX1-
Promotors, auch wenn mittlerweile einige andere Promotoren untersucht worden sind (Tschopp
et al., 1987; Waterham et al., 1997). Dabei werden im Gegensatz zum häufig verwendeten
autonom replizierten 2µ-Plasmid bei Saccharomyces cerevisiae integrative Vektoren verwendet.
Die Klonierungsschritte und Amplifikation der Plasmide werden in E. coli durchgeführt.
Als Selektionsmarker dient in den meisten Fällen das Gen der Histidinol-Dehydrogenase
(HIS4), das im Wirtsstamm GS115 mutiert ist, so daß eine Histidin-Auxotrophie entsteht, die
durch Integration des Vektors komplementiert wird (Higgins und Cregg, 1998). Das kanr- Gen
aus Tn903, das in P. pastoris Resistenz gegen das Antibiotikum G418 vermittelt, dient in einigen
Vektoren zur Selektion auf die Integration mehrerer Expressionskassetten. Eine direkte Selektion
der Transformanten auf G418-Resistenz ist allerdings nicht möglich. Zusätzlich wurden in den
letzten Jahren Vektoren entwickelt, die auf der Vermittlung einer Resistenz gegen das
Antibiotikum Zeocin basieren sowie Vektoren, die den konstitutiven GAP Promotor verwenden.
Üblicherweise existieren die Vektoren in Ausführungen mit und ohne Sekretionssignal, wobei in
erster Linie das Sekretionssignal des S. cerevisiae alpha-Paarungsfaktors verwendet wird. Einen
Überblick über die kommerziell erhältlichen, gebräuchlichen Vektoren gibt Tabelle 1-5.
Tabelle 1-5: Gebräuchliche Pichia-Expressionsvektoren
Bezeichnung Selektions-marker
Promotor Bemerkungen
pPIC3(K) his4 AOX1 Intrazelluläre Expression. K: Multi-copy-Selektion überG418-Resistenz möglich.
pPIC9(K) his4 AOX1 Enthält α-Faktor Sekretionssignal aus S. cerevisiae. K:Multi-copy-Selektion über G418-Resistenz möglich.
pPICZ(α) bler AOX1 Kleiner als pPIC3/pPIC9-Vektoren. Mit und ohne α-Faktor Sekretionssignal. Direkte Selektion und Multi-copy-Selektion über Zeocin-Resistenz.
pGAPZ(α) bler GAP Konstitutive Expression. Kleiner als pPIC3/pPIC9-Vektoren. Mit und ohne α-Faktor Sekretionssignal.Direkte und Multi-copy-Selektion über Zeocin-Resistenz.

Einleitung
15
Die Charakteristika der kommerziell erhältlichen Wirtsstämme sind in Tabelle 1-6
wiedergegeben:
Tabelle 1-6: Pichia-Wirtsstämme
Bezeichnung Genotyp Bemerkungen
GS115 his4 Standard-Wirtsstamm. Vektoren der pHIL, pPIC3- und pPIC9
Familien
SMD1168 his4, pep4 Protease-defizienter Wirtsstamm. Vektoren wie oben. Durch
Eliminierung von pep4 fallen weitere Proteasen aus, die durch pep4
aktiviert werden.
X33(Y-11430)
Wildtyp Vektoren der pPICz-Familie (Selektion auf Zeocin-Resistenz)
KM71 his4, aox1::arg4 Langsame Verwertung von Methanol
Mit dem Pichia-Expressionssystem sind in den letzten Jahren mehr als 300 Proteine
überexprimiert worden. Die Herkunft der exprimierten Gene umfaßt dabei das gesamte
Spektrum der belebten Welt und die Art der exprimierten Proteine reicht von komplexen
Hepatitis-B Oberflächenantigenen (Cregg et al., 1987) über kleine Peptidhormone (Clare et al.,
1991b) bis hin zu industriellen Enzymen (Juge et al., 1996). Eine Reihe von Proteinen konnte
dabei im Grammbereich (bezogen auf 1 Liter Kulturvolumen) exprimiert werden. Tabelle 1-7
faßt die bisher besten Expressionsergebnisse zusammen.
Die in Tabelle 1-7 genannten Beispiele stellen die Obergrenze der bisher erzielten
Expressionsraten dar. Für das Gros der in Pichia exprimierten Proteine werden dagegen
Expressionsraten im Bereich um 50 mg/L beschrieben. Am unteren Ende der Skala ist die
Expression einer Reihe von Proteinen im Bereich <1 mg·l-1 dokumentiert (Ohsawa et al., 1995;
Weiss et al., 1995; Pflanz et al., 1999), daneben existieren Fälle, in denen gar keine Expression
nachgewiesen werden konnte, obwohl die Transkription im Northern-Blot nachweisbar war, so
z. B. bei einem Konstrukt, das die VH und CH1 Domänen eines humanen Antikörpers umfaßt (M.
Stassen, Universität Maastricht, persönliche Mitteilung).
Hinsichtlich ihrer qualitativen Eigenschaften unterscheiden sich viele in Pichia
überexprimierte Proteine nicht von den entsprechenden nativen Proteinen (Fierobe et al., 1997;
Nilsen et al., 1997; Murphy et al., 1998; Han und Lei, 1999; Sun et al., 1999). Bei einigen
Proteinen verursachen Pichia-eigene Proteasen Probleme durch Degradation des Produkts noch
während der Expression. Zum Teil ließen sich diese Probleme durch Expression bei niedrigem
pH (Scorer et al., 1993; Werten et al., 1999), Zusatz von Kasein-Totalhydrolysat (Clare et al.,
1991b; Clare et al., 1998; Werten et al., 1999) oder Verwendung des proteasedefizienten

Einleitung
16
Wirtsstammes SMD1168 (siehe Tabelle 1-6) (Brankamp et al., 1995; White et al., 1995)
vermeiden, zum Teil waren solche Maßnahmen aber auch erfolglos (Juge et al., 1996; Henry et
al., 1997; Holmquist et al., 1997).
Tabelle 1-7: Mit dem Pichia-Expressionssystem erzielte Ergebnisse
intrazellulär/
sekretiert
Produkt
(Herkunftsorganismus)
Expressionslevel
[g·l-1]
Literaturstelle
I Tetanustoxin Fragment C
(Clostridium tetani)
12 (Clare et al., 1991a)
I Hydroxynitril-Lyase
(Hevea brasiliensis)
22 (Hasslacher et al., 1997)
I Hüllprotein (HIV) 1,25 (Scorer et al., 1993)
I Pertactin (Bordatella pertussis) 3,0 (Romanos et al., 1991)
I Tumor-Nekrosefaktor (human) 10 (Sreekrishna et al., 1989)
S α-Amylase (Bacillus licheniformis) 2,5 (Paifer et al., 1994)
S Koagulationshemmer
(Ornithodoros moubata)
1,7 (Laroche et al., 1994)
S Bm86 Antigen (Boophilus microplus) 1,5 (Rodriguez et al., 1994)
S Invertase (Saccharomyces cerevisiae) 2,3 (Tschopp et al., 1987)
S Kunitz Proteaseinhibitor (human) 1,0 (Van Nostrand et al., 1994)
S Synthetische Gelatine (Mus spec.) 14,8 (Werten et al., 1999)
S scFv (α-CEA) 1,2 (Freyre et al., 2000)
Auch bezüglich der Glykosylierung sind erhebliche Unterschiede zwischen verschiedenen
Proteinen gefunden worden: Pichia erkennt O- und N-Glykosylierungstellen (Heimo et al.,
1997) und ist in der Lage, Glykosylierungen durchzuführen, die denen der nativen Proteine sehr
ähnlich sind (Holmquist et al., 1997; Murphy et al., 1998; Han und Lei, 1999; Sun et al., 1999).
Die in Pichia festgestellten Glykosylierungen waren in einigen Fällen unterschiedlich von den in
Saccharomyces beobachteten (Miele et al., 1997), da Pichia scheint weniger zur
Hyperglykosylierung zu neigen scheint. In einigen Fällen wurde aber auch Hyperglykosylierung
beobachtet (Scorer et al., 1993; Hellwig et al., 1999), die zum Teil dazu führte, daß das
rekombinante Protein von Antikörpern, die gegen das native Gegenstück erzeugt wurden, nicht
erkannt wurde (Clare et al., 1998). Es ist in solchen Fällen möglich, die Glykosylierungstellen
auf genetischer Ebene durch ortsgerichtete Mutagenese auszuschalten (Munshi et al., 1997).
Die Fermentation von P. pastoris ist insofern einfach, als das der Organismus prinzipiell
keine besonderen Ansprüche an die Ausstattung des Reaktors stellt und auf billigen

Einleitung
17
synthetischen Medien wächst (Zhu et al., 1995; Stratton et al., 1998). Mit Glyzerin oder
Methanol als Kohlenstoffquelle lassen sich im Zulaufverfahren extrem hohe Zelldichten von bis
zu 450 g Frischgewicht pro Liter Kulturvolumen erreichen (Siegel und Brierley, 1989; Clare und
Romanos, 1995; Kim et al., 1997).
Die Fermentationsstrategie folgt in der Regel den im Handbuch der Fa. Invitrogen gegebenen
Vorschlägen: Zunächst wird im ansatzweisen Verfahren mit Glyzerin als Kohlenstoffquelle
fermentiert, bis die vorgelegte Menge Glyzerin verbraucht ist, was am rapiden Ansteigen der
Gelöstsauerstoffkonzentration erkennbar wird. Zu diesem Zeitpunkt wird Glyzerin im
Zulaufverfahren in limitierender Rate zugefüttert, entweder für eine bestimmte Zeit oder bis eine
bestimmte Zelldichte erreicht ist. Anschließend wird das Zulaufverfahren zur Induktion auf
Methanol als einzige Kohlenstoffquelle umgestellt. Die Zufuhrrate von Methanol wird dabei
anhand von Erfahrungswerten langsam gesteigert. Um zu verhindern, daß Methanol zu toxischen
Konzentrationen akkumuliert, wird die Methanolzufuhr in regelmäßigen Abständen gestoppt und
die Zeit bestimmt, die bis zum Ansteigen der Gelöstsauerstoffkonzentration verstreicht. Liegt
diese Zeit über einem Wert von 30-60 s, wird die Methanolzufuhrrate anschließend gesenkt. Mit
dieser Strategie lassen sich Zufuhrraten, die in etwa im Bereich der kombinierten Verbrauchs-
und Verdunstungsraten liegen, realisieren. Abweichend von diesen Fermentationsstrategien sind
vereinzelt Pichia-Fermentationen im kontinuierlichen Verfahren (Ellis et al., 1985; Digan et al.,
1989; Sreekrishna et al., 1989) und Fermentationen im Zufuhrverfahren mit Mischungen von
Glyzerin und Methanol in verschiedenen Verhältnissen beschrieben (Sreekrishna et al., 1989;
Loewen et al., 1997; Sreekrishna et al., 1997; Katakura et al., 1998).
Diese Eigenschaften charakterisieren das Pichia Expressionssystem als ein System, mit dem
sich bestimmte Proteine speziell bei Einsatz etwas aufwendigerer Fermentationstechnik durchaus
in großen Mengen produzieren lassen. Das Problem bei der Erzeugung und Selektion von
Expressorklonen für bestimmte Proteine liegt offensichtlich aufgrund proteinspezifischer
Eigenschaften auf posttranskriptionaler Ebene.
1.3 Problemstellung
Antikörper und Antikörperfragmente spielen schon heute in den medizinischen und
biologischen Wissenschaftsbereichen eine wichtige Rolle als molekulare Werkzeuge, deren
Bedeutung der der Restriktionsenzyme gleichkommt. Zudem rücken Fortschritte in der
Humanisierung von Maus-Antikörpern, der Herstellung von Antikörperbibliotheken und der
Kopplung von Antikörpern mit den verschiedensten Effektoren Antikörper-basierende

Einleitung
18
Therapieformen in den Mittelpunkt des Interesses. Es kann mit Sicherheit angenommen werden,
daß die Zahl der Anwendungen ebenso wie der Maßstab und die Qualität, in dem rekombinante
Antikörper benötigt werden, in den nächsten Jahren noch steigen wird. Damit wird auch die
Erforschung preiswerter, sicherer und zuverlässiger Expressionssysteme erforderlich.
Im Rahmen dieser Arbeit galt es, einen single-chain-Fv Antikörper mit diagnostischer
Relevanz, der beispielhaft für die eingangs erläuterte Entwicklung steht, im Maßstab von 500 mg
zu produzieren und zu reinigen. Dazu sollten als Expressionssysteme die Hefe P. pastoris und
die N. tabacum cv BY-2 Zellkulturlinie verwendet und verglichen werden. Für beide Systeme
sollten Fermentationsstrategien entwickelt, die Prozesse auf Ausbeute und Wachstum untersucht
und optimiert und im 30-l Maßstab durchgeführt werden, um Aussagen über die Eignung dieser
Expressionssysteme im größeren Maßstab zu erhalten. Die Rahmenbedingungen sollten die
Produktion unter den Bedingungen der Guten Herstellungspraxis (GMP) zulassen, um die
Ergebnisse auf therapeutisch relevante Antikörperfragmente übertragen zu können. Die beiden
Expressionssysteme sollten hinsichtlich der volumetrischen Produktivität der Prozesse, des
erforderlichen Aufwands und ökonomischer Aspekte in verschiedenen Produktionsmaßstäben
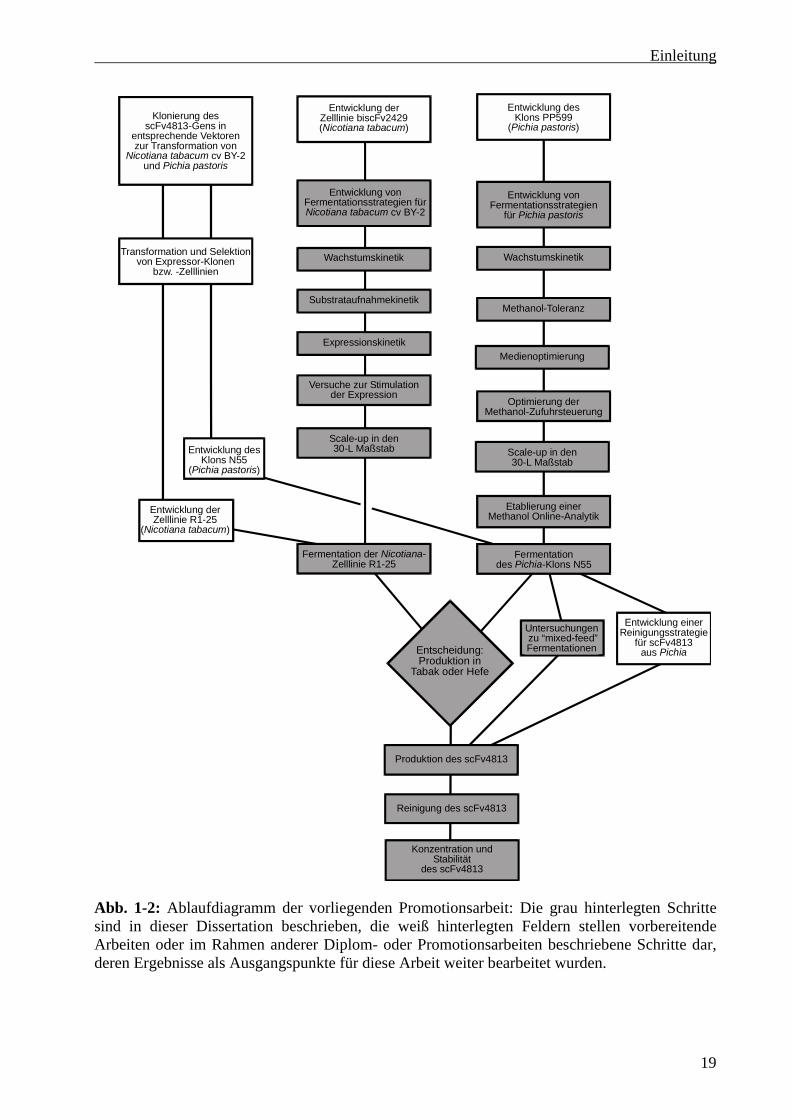
vergleichend beurteilt werden. Abb. 1-2 zeigt den Ablauf der Promotionsarbeit.
Einleitung
19
Konzentration undStabilität
des scFv4813
Fermentation der Nicotiana-Zelllinie R1-25
Fermentationdes Pichia-Klons N55
Etablierung einerMethanol Online-Analytik
Scale-up in den30-L Maßstab
Versuche zur Stimulationder Expression
Expressionskinetik
Substrataufnahmekinetik
Wachstumskinetik
Entwicklung vonFermentationsstrategien fürNicotiana tabacum cv BY-2
Entwicklung vonFermentationsstrategien
für Pichia pastoris
Wachstumskinetik
Methanol-Toleranz
Medienoptimierung
Optimierung derMethanol-Zufuhrsteuerung
Scale-up in den30-L Maßstab
Transformation und Selektionvon Expressor-Klonen
bzw. -Zelllinien
Klonierung desscFv4813-Gens in
entsprechende Vektorenzur Transformation von
Nicotiana tabacum cv BY-2und Pichia pastoris
Entwicklung derZelllinie biscFv2429(Nicotiana tabacum)
��������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������������
Entwicklung desKlons PP599
(Pichia pastoris)
Entscheidung:Produktion in
Tabak oder Hefe
Untersuchungen zu “mixed-feed”Fermentationen
Entwicklung einerReinigungsstrategie
für scFv4813aus Pichia
Entwicklung derZelllinie R1-25
(Nicotiana tabacum)
Entwicklung desKlons N55
(Pichia pastoris)
Reinigung des scFv4813
Produktion des scFv4813
Abb. 1-2: Ablaufdiagramm der vorliegenden Promotionsarbeit: Die grau hinterlegten Schrittesind in dieser Dissertation beschrieben, die weiß hinterlegten Feldern stellen vorbereitendeArbeiten oder im Rahmen anderer Diplom- oder Promotionsarbeiten beschriebene Schritte dar,deren Ergebnisse als Ausgangspunkte für diese Arbeit weiter bearbeitet wurden.

Material und Methoden
20
2 Material und Methoden
2.1 Material
2.1.1 Chemikalien und Verbrauchsmaterialien
Alle Chemikalien und Fertiglösungen wurden mindestens mit dem Reinheitsgrad p.a.
verwendet und von folgenden Firmen bezogen: Serva (Heidelberg), Sigma (Deisenhofen),
BioRad (München), Boehringer (Mannheim), Pharmacia (Freiburg), GibcoBRL (Eggenstein),
Merck (Darmstadt), Fluka (Neu-Ulm), Roth (Karlsruhe), Novex (San Diego) und New England
Biolabs (Schwalbach).
Verbrauchsmaterialien stammten von Millipore (Eschborn), Eppendorf (Hamburg),
Whatman (Maidstone, England), USB/Amersham (Bad Homburg), Serva (Heidelberg), Schott
Glaswerke (Mainz), Zeiss (Oberkochen), Greiner (Solingen) und Roth (Karlsruhe).
2.1.2 Verwendete Medien, Puffer und Lösungen
Standard-Lösungen, Puffer und Medien wurden nach (Sambrook et al., 1989) oder (Ausubel
et al., 1994) mit entionisiertem Wasser hergestellt. Der pH-Wert der Lösungen wurde, wenn
nicht anders erwähnt, mit HCl bzw. NaOH eingestellt. Medien, Medienkonzentrate und –zusätze
wurden durch Autoklavieren (25 min, 121° C) oder durch Sterilfiltration (0.2 µm) sterilisiert.
2.1.3 Enzyme und Reaktionskits
Falls nicht anders angegeben wurden alle Enzyme und Kits inkl. Reaktionspuffer von der
Firma Boehringer (Mannheim) bezogen und nach Herstellerangaben eingesetzt.
2.1.4 Antikörper
Im Rahmen dieser Arbeit wurden für ELISA (2.3.7) oder Western Blot (2.3.6) die folgenden
monoklonalen Antikörper bzw. polyklonalen Seren verwendet:
• C-α-TMV: Polyklonale Huhn-anti-TMV Ak (Tabak-Mosaikvirus) aus Eigelb (Monecke,
RWTH-Aachen)
• Fab24: Fab-Fragmente des mAk24 (Fischer, 1990)
• R-α−id24: Polyklonales Kaninchen-anti-Idiotyp mAk 24 Serum (Fischer, 1990)

Material und Methoden
21
• mAk T84.66 Monoklonaler Mausantikörper gegen CEA (City of Hope, Duarte, CA)
• M-α-His: Monoklonaler Mausantikörper gegen das His6-Epitop (Qiagen, Hilden)
• GAMAP: Polyklonales, AP-konjugiertes Ziege-anti-Maus Serum (Dianova, Hamburg)
• GARAP: Polyklonales, AP-konjugiertes Ziege-anti-Kaninchen Serum, (Dianova,
Hamburg)
• M-α-KDEL: Monoklonaler Mausantikörper mit Spezifität für das Grp78 Epitop (Biomol,
Hamburg)
2.1.5 Geräte, Apparaturen und Zubehör
• Reinraumwerkbank (BABCOCK-BS; Bad Hersfeld)
• Inkubationsschüttler Innova 4830 (New Brunswick Scientific, Nürtingen)
• Dialux 20 Mikroskop, Objektiv NPL Fluotar 40/0,70 NPT, Okular Periplan 12,5(Leitz,
Wetzlar), Photoautomat MP55 (Wild, Köln)
• Photometer: Zweistrahl-Spektrophotometer Uvikon 930 (Kontron, München),
Quarzglasküvetten
• PCR-Thermocycler: Primus 96 (MWG-Biotech, Ebersberg)
• Fotodokumentation mit UV-Transilluminator: Wellenlänge 302nm (Herolab, Wiesloch)
• Gelelektrophorese-Apparaturen: Elektrophoresekammern, Gelträger, Kämme, Netzteile,
MiniProtean II- und Transblot-Zelle, Vakuumgeltrockner (BioRad, München)
• Ultraschallgeber "Labsonic U" (B. Braun Biotech, Melsungen)
• Rührzellen-Druckkonzentratoren (Millipore, Eschborn)
• Fotoscanner Arcus II (Agfa, Köln)
• BIACORE 2000, CM5-Sensorchip, NTA-Sensorchip (Biosensor, Uppsala, Schweden)
• Multikanalphotometer SpectraMAX 340 (Molecular Devices, Sunnyvale, CA)
• Zentrifugen: RC-5B/RC-5C (Sorvall, Bad Homburg), Biofuge A (Heraeus, Osterode),
Eppendorfzentrifuge (Eppendorf, Hamburg), Avanti 30 und Avanti J25 (Beckman, Fullerton,
CA, USA)
• Durchflußzentrifuge: CARR Powerfuge P6 (Carr, Ranschbach)
• Econo-Proteinreinigungsanlage (BioRad, München)
• Streamline 25-Säule mit "Streamline Chelating Medium" (Pharmacia, Freiburg)
• Äkta Purifier Proteinreinigungsanlage (Pharmacia, Freiburg)

Material und Methoden
22
2.1.6 Fermenter
2.1.6.1 Fermenter für die Pflanzensuspensionskultur
Für die Kultivierung der Nicotiana tabacum BY-2 Zellinien wurden drei verschiedene
Rührreaktoren aus den Bioclave-, Biobench- und Biopilot-Baureihen der Firma Applikon
(Schiedam, Niederlande) verwendet. Alle Kessel wiesen einen Schlankheitsgrad von 2:1
(Füllhöhe:Duchmesser) bezogen auf das Arbeitsvolumen auf. Die Durchmischung erfolgte in
allen Fällen von oben durch jeweils einen dreiblättrigen Schrägblattrührer. Alle Reaktoren
wurden ohne Schikanen betrieben. Die Kenndaten der Reaktoren sind in Tabelle 2-1
zusammengefaßt.
Tabelle 2-1: Kenndaten der Rührreaktoren für die Pflanzensuspensionskultur
Bezeichnung 5-l Bioclave
5-l Biobench
30-l Biopilot
Arbeitsvolumen 5 l 30 l
Durchmesser 162 mm 265 mm
Füllhöhe 320 cm 530 mm
Rührerart
(Anzahl)
3-blättr.
Schrägblattrührer
(1)
3-blättr.
Schrägblattrührer
(1)
Rührer-Durchmesser 60 mm 140 mm
Rührerdrehzahl 130 U·min-1 100 U·min-1
Abb. 2-1 zeigt maßstabsgerecht die Geometrie der Biobench 7 und des Biopilot 40
Rührkessel. Im folgenden werden die Bioreaktoren einheitlich entsprechend den
Arbeitsvolumina als 5-l Reaktor (Biobench 7) und 30-l Reaktor (Biopilot 40) bezeichnet.

Material und Methoden
23
Abb. 2-1: Geometrie und Durchmischungsverhältnisse der für Nicotiana tabacum cv BY-2(links) bzw. Pichia pastoris (rechts) verwendeten 5-l bzw. 30-l Rührkessel.
Die Meßwertumwandlung, -anzeige und ggf. Kontrolle von Temperatur, Belüftungsrate,
Rührerdrehzahl, pH-Wert und Antischaumzugabe wurde von den Applikon Steuereinheiten
ADI 1030, ADI 1040 bzw. ADI 1060 in Verbindung mit den Basiseinheiten durchgeführt. Die
Meßwertaufzeichnung erfolgte über die BioXpert Software (Applikon) in den Versionen 1.10 bis
2.21 in Intervallen von 1-5 Minuten.
2.1.6.2 Fermenter für die Kultivierung von Pichia pastoris
Für die Kultivierung von Pichia pastoris wurden 5-l Biobench und 30-l Biopilot Reaktoren
verwendet, die bis auf die eingesetzten Rührer und den Einbau von Schikanen den in der
Pflanzensuspensionskultur verwendeten Reaktoren baugleich waren (siehe Tabelle 2-1, Abb.
2-1). Es kamen jeweils drei 6-blättrige Rushton Scheibenblattrührer von 63 mm (5-l Biobench)
bzw. 83 mm (30-l Biopilot) Durchmesser zum Einsatz. Der verwendete Motor war stärker
dimensioniert und erlaubte Drehzahlen von 1000 U·min-1 (5-l Biobench) bzw. 500 U·min-1 (30-l
Biopilot).

Material und Methoden
24
2.1.7 Verwendete Zellinien
2.1.7.1 Zellinien von Nicotiana tabacum cv Bright-Yellow-2 (BY-2)
Im Rahmen dieser Arbeit wurden zwei verschiedene Suspensionskulturen von Nicotiana
tabacum cv BY-2 verwendet: Die als biscFv2429KDEL bezeichnete Zellinie exprimiert einen
aus den monoklonalen Antikörpern "24" und "29" gegen TMV konstruierten bispezifischen
single-chain Antikörper biscFv2429 unter der Kontrolle eines modifizierten CaMV 35SS
Promotors. Das C-terminale KDEL Signalpeptid führt zur Retardierung des Proteins im
endoplasmatischen Retikulum (ER) der Zellen. Die Herstellung des Expressionskonstrukts und
der Zellinie wurde beschrieben (Schumann, 1996).
Die als R1-25 bezeichnete Zellinie exprimiert das scFv-Fragment des FSH-spezifischen mAk
4813 (FSH, "follicle stimulating hormone" unter Kontrolle eines synthetischen Promotors aus
Regulationssequenzen der Octopin-Synthase und Mannopin-Synthase-Gene (Ni et al., 1995).
Auch in diesem Fall wurde die C-terminale KDEL-Sequenz zur Akkumulation des scFv4813 im
ER verwendet. Das scFv4813-Konstrukt wurde freundlicherweise von Dr. Paul van der Logt
(Unilever, Bedford, GB) zur Verfügung gestellt, von Dr. Olga Artsaenko (Institut für Biologie I,
RWTH-Aachen) in den Pflanzenexpressionsvektor pSSH1 im Rahmen eines Industrieprojekts
umkloniert und anschließend zur Transformation von Agrobacterium tumefaciens verwendet.
Aus der Co-Kultivierung rekombinanter A. tumefaciens mit Nicotiana tabacum cv BY-2 Wildtyp
ging die rekombinante Tabak-Zellinie R1-25 hervor (Raven, 1999).
2.1.7.2 Zellinien von Pichia pastoris
Im Rahmen dieser Arbeit wurden drei verschiedene Expressorklone von Pichia pastoris
verwendet: Alle drei Klone sind Konstrukte, die auf dem pPIC9K Vektor basieren (Tabelle 1-5;
(White et al., 1995)).
Der Klon PP3299 wurde freundlicherweise von Dr. Paul van der Logt (Unilever Research
Laboratories, Bedford, UK) zur Verfügung gestellt. Dieser Klon exprimiert das scFv-Fragment
des mAk 3299 (Maus-anti-hCG, human chorionic gonadotrophin), das nach Induktion im
Medium in Konzentrationen von ca. 100 mg·l-1 akkumuliert. Dieser Klon wurde phänotypisch als
MutS-Klon charakterisiert (pers. Mitteilung, Dr. P. van der Logt, Unilever). Anhand dieses Klons
wurde im Rahmen der Maßstabsvergrößerung in den 30-l Maßstab das Auftreten proteolytischer
Effekte bei mangelnder Durchmischung gezeigt (3.3.5, Abb. 3-15).
Der Klon PP599 exprimiert das Fusionsprotein CEA N-A3, das aus zwei Domänen des
humanen "Carcinoembryonic antigen" (CEA) in sekretierter, glykosylierter Form ins Medium.

Material und Methoden
25
Die Klonierung des Gens sowie die Erzeugung und Charakterisierung dieses Mut+-
Expressorklons sind beschrieben (Robin, 1998; You et al., 1998; Hellwig et al., 1999).
Der Klon N55 exprimiert das scFv-Fragment des mAk4813 (2.1.7.1). Das Protein wird mit
Hilfe der im Vektor pPIC9K enthaltenen prepro-"leader"-Sequenz des α-Paarungsfaktors von
S. cerevisiae sekretiert und besitzt eine C-terminale his6-Sequenz zur Detektion und Reinigung
(Aminosäuren 253 bis 258, s. u.). Die Erzeugung und Charakterisierung dieses Klons ist
beschrieben (Raven, 1999). Die Aminosäuresequenz dieses Antikörpers lautet:
1 11 21 31 41 51
| | | | | |
1 MAEVKLQESG GGLVKPGGSL KLSCAASGFT FSSYVMSWVR QTPEKRLEWV ATISSGGSYT 60
61 YYPDSVKGRF TISRDNAKNT LYLQMSSLRS EDTAMYYCAR QRVDGYSSYW YFDVWGQGTT 120
121 VTVSSGGGGS GGGGSGGGGS VDIELTQSPK FMSTSVGDRV SITCKASQNV GTAVAWYQQK 180
181 PGQSPKLLIY SASNRYSGVP DRFAGSGSGT DFTLTISNMQ SEDLADYFCQ QYSTSPYTFG 240
241 GGTKLELKRA AAHHHHHH
Die theoretische Masse beträgt demnach 27,8 kDa.
2.2 Molekularbiologische Methoden
Alle gentechnologischen Arbeiten wurden nach S1-Richtlinien durchgeführt und waren
durch das Regierungspräsidium des Landes NRW sowie vom BGA genehmigt
[AZ 521-K-1-8/98: AI3-04/1/0866/88 (S1)].
2.2.1 PCR zur Bestimmung des Methanolverwertungs-Phänotyps von Pichia pastoris
Die Bestimmung des "Methanol utilization"-Phänotyps wurde mit Hilfe der PCR (Saiki et
al., 1985) unter Abwandlung des Protokolls von Linder durchgeführt (Linder et al., 1996): 10 µl
einer 1:10 verdünnten, 16 h alten Pichia-Vorkultur wurden mit 5 µl einer frisch angesetzten
Lyticase-Lösung (Sigma, 5 U·µl-1) versetzt. Der Ansatz wurde 20 min bei 28° C inkubiert und
anschließend bei –20° C tiefgefroren und wieder aufgetaut, um die Zellen aufzuschließen.
Zelltrümmer wurden abzentrifugiert und 5 µl des Überstandes wurden in 200-µL silikonisierten
"PCR-strips" direkt zur PCR eingesetzt (s. u.). Zur Vermeidung von Verdunstung wurden die
Ansätze mit 50 µl Mineralöl überschichtet. Die PCR wurde nach dem folgenden Schema im
Thermocycler (MWG Biotech) durchgeführt.
10 µl des PCR-Ansatzes wurden mit Hilfe eines 1,2 %igen Agarosegels analysiert (2.2.2).

Material und Methoden
26
PCR-Ansatz:1-10 ng Matrizen-DNA (Pichia Zellaufschluß)5-10 pmol 5’-Primer (5'-AOX, s. u.)5-10 pmol 3'-Primer (3'-AOX, s. u.)
1,5 mM MgCl2
0.25 mM dNTP mix5 % (v/v) DMSO
10 % (v/v) 10-fach Taq DNA-Polymerase-Puffer2 U Taq-Polymerase
ad 50 µl H2O
PCR Primer (MWG)5'-AOX: 5'-GAC TGG TTC CAA TTG ACA AGC3'-AOX: 5'-GGC AAA TGG CAT TCT GAC ATC
PCR-Schema:5 min 95° C (Template-Denaturierung)
30 Wiederholungen:1 min 95° C (Strangtrennung)1 min 54° C (Primer-Anlagerung)1 min 72° C (Kettenverlängerung)
3 min 72° C
2.2.2 Analytische Agarosegelelektrophorese
Die Auftrennung der in der PCR amplifizierten DNA-Fragmente erfolgte in 1,2%igen (w/v)
Agarosegelen bei einer Feldstärke von 4-5 V/cm2. Als Größen- und Konzentrationsstandard
wurde PstI geschnittene DNA des λ-Phagen verwendet. Die Herstellung der Gele und die
Durchführung der Elektrophorese erfolgte in 1xTBE-Puffer unter Zusatz von 0,1 µg/ml
Ethidiumbromid (Sambrook et al., 1989). Dokumentiert wurden die Agarosegele mit dem
digitalen Photodokumentationssystem von Herolab bei einer Wellenlänge von 302 nm.
1xTBE-Puffer90 mM TRIS90 mM B(OH)3
2,5 mM EDTA
2.3 Proteinchemische, naßchemische und immunologische Methoden
2.3.1 Herstellung von Proteinextrakten aus BY-2 Zellen
Zur Analyse der intrazellulär lokalisierten Antikörperfragmente biscFv2429 und scFv4813
wurden Gesamtproteinextrakte hergestellt. Dazu wurde die Fermentationsbrühe zentrifugiert
(4300 g, 5 min), der Überstand dekantiert, das Zellvolumen bestimmt und mit dem gleichen
Volumen Proteinextraktionspuffer resuspendiert. Ein Aliquot von 4 ml wurde in ein frisches
Röhrchen überführt und mit Hilfe eines Ultraschallgebers (Braun) 60 halbsekündigen

Material und Methoden
27
Ultraschallimpulsen, unterbrochen von halbsekündigen Pausen, ausgesetzt. Die Leistungsabgabe
des Geräts betrug 60 W. Anschließend wurden Trümmer von Zellwänden und Organellen
abzentrifugiert (4300 g, 5 min) und der Überstand bis zur Analyse bei –20° C gelagert.
Proteinextraktionspuffer, pH 8,0200 mM TRIS
5 mM EDTA1 mM DTT
150 mM NaCl0,02 % (w/v) NaN3
0,1 % (v/v) Tween 20
2.3.2 Bestimmung des Gesamtproteingehaltes von BY-2 Zellen
Zur Bestimmung des Gesamtproteingehaltes von BY-2 Zellextrakt (2.3.1) wurde der
"BioRad Protein assay" (BioRad, München) verwendet, der auf der Bradford-Färbung beruht
(Bradford, 1976).
50 µl eines 1:100 verdünnten BY-2 Zellextrakts wurden in Mikrotiterplatten pipettiert und
mit 160 µL der 1:4 verdünnten Färbelösung versetzt. Nach einer Inkubationszeit von 5-10
Minuten wurde die Absorption bei 595 nm bestimmt. Der Mittelwert der Dreifachansätze wurde
mit einer Kalibrationskurve von RSA Fraktion V (Merck) über den Bereich von 5-50 µg/ml
korreliert und darüber die Proteinkonzentration ermittelt.
2.3.3 Bestimmung des Phosphat-Gehalts in Fermentationsüberständen
Zur Bestimmung des Phosphat-Gehalts in Überständen von N. tabacum cv BY-2
Fermentationen wurde eine nach DIN 38405 entwickelte, naßchemische Methode verwendet.
Phosphat bildet in Gegenwart von Ascorbinsäure und Antimon mit Molybdat-Ionen einen blauen
Komplex ("Phosphormolybdän-Blau"), dessen Bildung bei 680 nm verfolgt werden kann. 100 µl
des zu messenden Überstandes wurden mit 200 µl Ascorbinsäure-Lösung und 400 µl Molybdat-
Reagenz versetzt und mit entionisiertem Wasser auf 10 ml aufgefüllt. Nach exakt 15 Minuten
wurde die Absorption bei 680 nm bestimmt und mit einer Kalibrationskurve verglichen.

Material und Methoden
28
Ascorbinsäurelösung0,57 M Ascorbinsäure
Molybdat-Reagenzlösung21 mM (NH4)Mo7O24
2,1 mM K(SbO)C4H4O6
25 % (v/v) H2SO4
2.3.4 Bestimmung von Glukose, Fruktose und Saccharose in Fermentationsüberständen
Die Bestimmung von Glukose, Fruktose und Saccharose in Fermentationsüberständen von N.
tabacum cv BY-2 erfolgte mit Hilfe des Enzymkits "Sucrose, D-Glucose, D-Fructose"
(Boehringer, Mannheim) nach Angaben des Herstellers. Der Test beruht auf der photometrischen
Messung der Abnahme der Extinktion bei 340 nm infolge enzymatischer, NADPH-abhängiger
Umsetzung der Saccharide.
2.3.5 SDS-PAA Gelelektrophorese (SDS-PAGE) und Coomassie Brilliant BlueFärbetechnik
2.3.5.1 Gelelektrophorese
Die denaturierende SDS-PAGE zur Trennung von Proteinen gemäß ihrer molekularen Größe
wurde in einem diskontinuierlichen Puffersystem nach Laemmli durchgeführt (Sammelgel:
T=4%, C=2,7%, pH 6,8; Trenngel: T=12%, C=2,7%, pH 8,8) (Laemmli, 1970). Die Gele wurden
in der „Mini Protean II“-Apparatur (BioRad) nach Herstellerangaben hergestellt. Die in den
Proben enthaltenen Proteine wurden durch Erhitzen mit 10x SDS-PAGE-Probenpuffer
(5min/95°C) solubilisiert und anschließend bei konstanter Spannung von 150 V im Gel getrennt.
10x SDS-PAGE-Probenpuffer62,5 mM TRIS
10 % (v/v) Glyzerin2 % (w/v) SDS5 % (v/v) ß-Mercaptoethanol
0,05 % (w/v) Bromphenolblau
Elektrophorese-Laufpuffer, pH 8.325 mM TRIS
192 mM Glyzin0,1 % (w/v) SDS
2.3.5.2 Coomassie Brilliant Blue Färbung
Die Visualisierung elektrophoretisch aufgetrennter Proteine im Gel erfolgte nach Hersteller-
angaben durch Anfärbung mit Coomassie Brilliant Blue G-250 Färbelösung (ca. 1 h bei RT).
Zum Entfärben wurde das Gel in Entfärbelösung zwei mal kurz erhitzt (95° C). Anschließend
wurde das Gel zwischen zwei Cellophanfolien getrocknet und lichtgeschützt gelagert. Als

Material und Methoden
29
Größenstandard für alle Coomassie-gefärbten Proteingele diente Mark 12 (Novex). Dieser
Größenstandard enthält Proteine der Molekulargewichte 200, 115, 98, 66,3, 55,4, 36,5, 31,0,
21,5 und 15,4 kDa.
Coomassie-Färbelösung0,25 % (w/v) Coomassie Brilliant Blue G250
50 % (v/v) Methanol9 % (v/v) Essigsäure2 % (w/v) tri-Chlor-Essigsäure
Coomassie Entfärbelösung5 % (v/v) Methanol
7,5 % (v/v) Essigsäure
2.3.6 Western Blot
Zum Nachweis von scFvs im Nanogrammbereich über die C-terminalen his6 bzw. KDEL
Peptide wurden die reduzierten und denaturierten Proteine nach gelelektrophoretischer
Auftrennung (2.3.5) auf Nitrozellulose (Amersham) transferiert (Towbin et al., 1979). Die
entsprechende Proteinbande wurde auf der Nitrozellulosemembran nach Zugabe von
spezifischen Antikörpern durch die Bindung eines mit alkalischer Phosphatase (AP) konjugierten
sekundären anti-Antikörpers mit nachfolgendem Substratumsatz sichtbar gemacht (Burnette,
1981). Dies ermöglicht die eindeutige Identifizierung des scFvs unter den anderen in der Probe
vorhandenen Proteinen und eine Gößenabschätzung. Gleichzeitig lassen sich Degradationen am
Auftreten zusätzlicher, kleinerer Banden erkennen.
Als Größenmaßstab diente in allen Western Blots eine Mischung aus vorgefärbten Proteinen
von mit den Molekulargewichten 150, 62, 47,5, 32,5, 25 und 16,5 kDa (New England Biolabs).
Zum Blockieren freier Bindungsstellen auf der Nitrozellulosemembran vor der Inkubation mit
sekundären Antikörpern diente in allen Fällen "Marvel" Skim Milk Powder (0,01 % Fett), in
2 %iger (w/v) Lösung (1 h bei RT). Als sekundäre Antikörper kamen M-α-his (Qiagen) in der
Konzentration 100 ng·ml-1 bzw. M-α-KDEL (Biomol) in der Konzentration 1,5 ng·ml-1 zum
Einsatz (2.1.4). In beiden Fällen wurde Fc-spezifischer GAMAP (Dianova) in einer Konzentration
von 120 ng·ml-1 als Anti-Antikörper verwendet (2.1.4). Als Substrat zur Anfärbung gebundener
AP-markierter Antikörper diente die NBT/BCIP Fertigsubstratlösung (GibcoBRL), die nach
Herstellerangaben eingesetzt wurde.

Material und Methoden
30
Transferpuffer25 mM TRIS
192 mM Glyzin20 % (v/v) Methanol
2.3.7 ELISA
Der ELISA (Engvall und Perlmann, 1971) stellt eine äußerst sensitive Methode zum
Nachweis der Funktionalität von Antikörpern und Antikörperfragmenten dar. Darüber hinaus
zeichnet er sich durch hohe Selektivität aus. Im Rahmen dieser Arbeit erfolgte der Nachweis der
Antikörperfragmente mit dem Verfahren des direkten oder indirekten ELISA (Fischer, 1990).
Alle ELISA Puffer wurden nach (Clark und Adams, 1977) hergestellt. In allen ELISAs kamen
M129B "high binding" Polystyrol-Mikrotiterplatten (Greiner) zum Einsatz.
"Coating"- Puffer (CP), pH 9,615 mM Na2CO3
25 mM NaHCO3
0,02 % (w/v) NaN3
Glutardialdehyd-Bindungspuffer (GBP), pH 5,0100 mM K2HPO4
Phosphatpuffer (PBS), pH 7,410 % (v/v) 10xPBS
Physiologische Salzlösung (PS)0,85 % (w/v) NaCl
Probenpuffer (PP), pH 6,010 % (v/v) 10 x PBS
2 % (w/v) PVP (20-30 kDa)0,2 % (w/v) NaN3
0,05 % (w/v) Tween 20
Proteinextraktionspuffer (EP), pH 7,50,2 M 1 M Tris-HCl
0,05 M EDTA0,02 % (w/v) NaN3
0,1 % (v/v) Tween 201 mM PMSF
Enzymmarkierte Antikörper-Puffer (EMAP), pH 7,410 % (v/v) 10 x PBS
2 % (w/v) PVP (20-30 kDa)0,2 % (w/v) BSA
0,02 % (w/v) NaN3
0,05 % (v/v) Tween 20
Substratpuffer (SP), pH 9,810 % (v/v) di-Ethanolamin
1 mM MgCl2
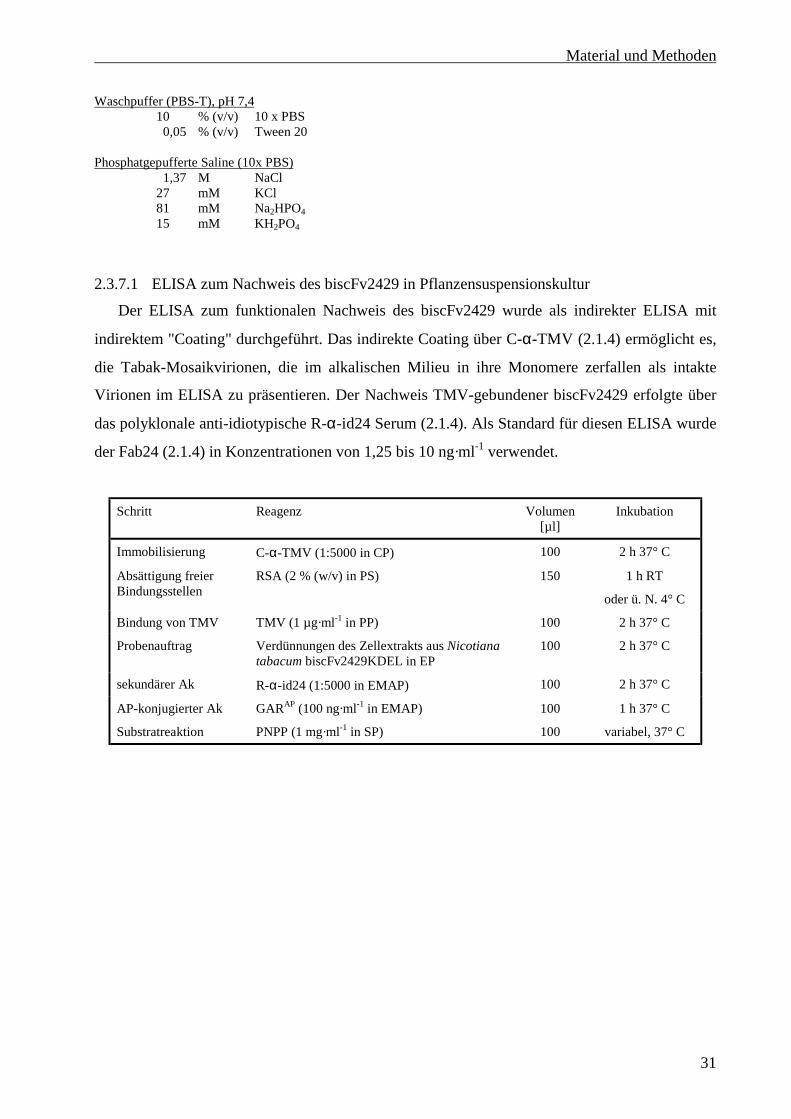
Material und Methoden
31
Waschpuffer (PBS-T), pH 7,410 % (v/v) 10 x PBS
0,05 % (v/v) Tween 20
Phosphatgepufferte Saline (10x PBS)1,37 M NaCl
27 mM KCl81 mM Na2HPO4
15 mM KH2PO4
2.3.7.1 ELISA zum Nachweis des biscFv2429 in Pflanzensuspensionskultur
Der ELISA zum funktionalen Nachweis des biscFv2429 wurde als indirekter ELISA mit
indirektem "Coating" durchgeführt. Das indirekte Coating über C-α-TMV (2.1.4) ermöglicht es,
die Tabak-Mosaikvirionen, die im alkalischen Milieu in ihre Monomere zerfallen als intakte
Virionen im ELISA zu präsentieren. Der Nachweis TMV-gebundener biscFv2429 erfolgte über
das polyklonale anti-idiotypische R-α-id24 Serum (2.1.4). Als Standard für diesen ELISA wurde
der Fab24 (2.1.4) in Konzentrationen von 1,25 bis 10 ng·ml-1 verwendet.
Schritt Reagenz Volumen[µl]
Inkubation
Immobilisierung C-α-TMV (1:5000 in CP) 100 2 h 37° C
Absättigung freierBindungsstellen
RSA (2 % (w/v) in PS) 150 1 h RT
oder ü. N. 4° C
Bindung von TMV TMV (1 µg·ml-1 in PP) 100 2 h 37° C
Probenauftrag Verdünnungen des Zellextrakts aus Nicotianatabacum biscFv2429KDEL in EP
100 2 h 37° C
sekundärer Ak R-α-id24 (1:5000 in EMAP) 100 2 h 37° C
AP-konjugierter Ak GARAP (100 ng·ml-1 in EMAP) 100 1 h 37° C
Substratreaktion PNPP (1 mg·ml-1 in SP) 100 variabel, 37° C
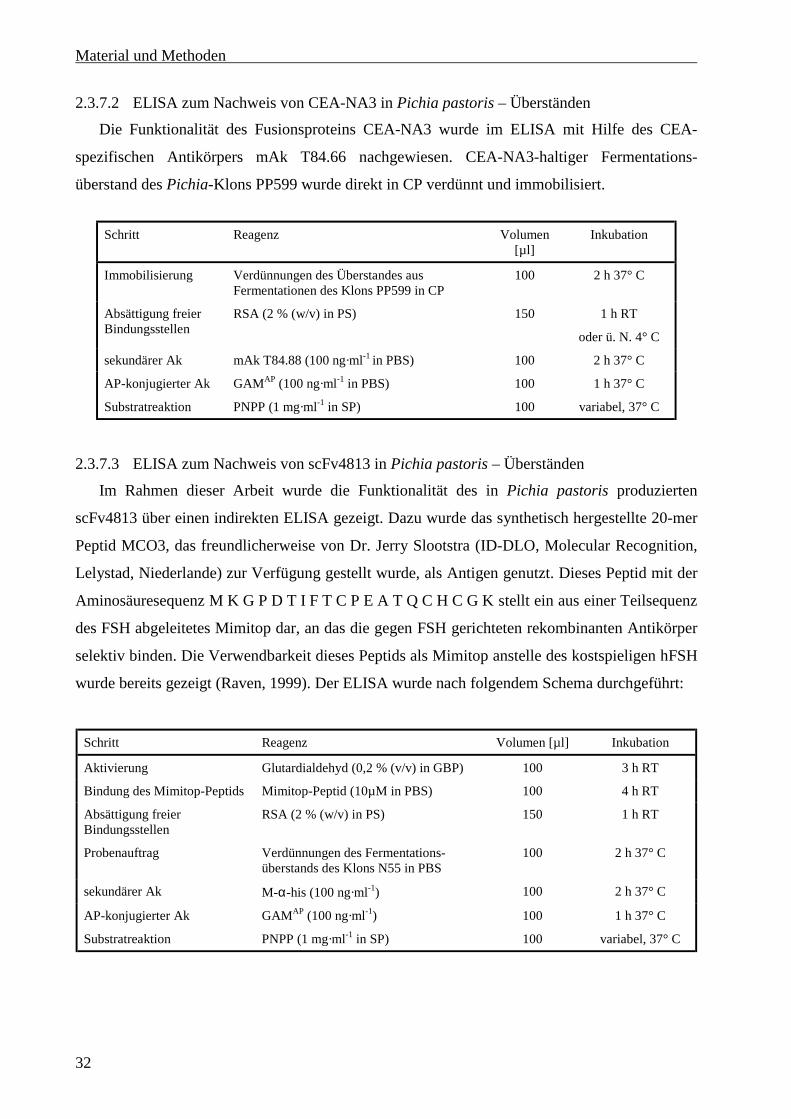
Material und Methoden
32
2.3.7.2 ELISA zum Nachweis von CEA-NA3 in Pichia pastoris – Überständen
Die Funktionalität des Fusionsproteins CEA-NA3 wurde im ELISA mit Hilfe des CEA-
spezifischen Antikörpers mAk T84.66 nachgewiesen. CEA-NA3-haltiger Fermentations-
überstand des Pichia-Klons PP599 wurde direkt in CP verdünnt und immobilisiert.
Schritt Reagenz Volumen[µl]
Inkubation
Immobilisierung Verdünnungen des Überstandes ausFermentationen des Klons PP599 in CP
100 2 h 37° C
Absättigung freierBindungsstellen
RSA (2 % (w/v) in PS) 150 1 h RT
oder ü. N. 4° C
sekundärer Ak mAk T84.88 (100 ng·ml-1 in PBS) 100 2 h 37° C
AP-konjugierter Ak GAMAP (100 ng·ml-1 in PBS) 100 1 h 37° C
Substratreaktion PNPP (1 mg·ml-1 in SP) 100 variabel, 37° C
2.3.7.3 ELISA zum Nachweis von scFv4813 in Pichia pastoris – Überständen
Im Rahmen dieser Arbeit wurde die Funktionalität des in Pichia pastoris produzierten
scFv4813 über einen indirekten ELISA gezeigt. Dazu wurde das synthetisch hergestellte 20-mer
Peptid MCO3, das freundlicherweise von Dr. Jerry Slootstra (ID-DLO, Molecular Recognition,
Lelystad, Niederlande) zur Verfügung gestellt wurde, als Antigen genutzt. Dieses Peptid mit der
Aminosäuresequenz M K G P D T I F T C P E A T Q C H C G K stellt ein aus einer Teilsequenz
des FSH abgeleitetes Mimitop dar, an das die gegen FSH gerichteten rekombinanten Antikörper
selektiv binden. Die Verwendbarkeit dieses Peptids als Mimitop anstelle des kostspieligen hFSH
wurde bereits gezeigt (Raven, 1999). Der ELISA wurde nach folgendem Schema durchgeführt:
Schritt Reagenz Volumen [µl] Inkubation
Aktivierung Glutardialdehyd (0,2 % (v/v) in GBP) 100 3 h RT
Bindung des Mimitop-Peptids Mimitop-Peptid (10µM in PBS) 100 4 h RT
Absättigung freierBindungsstellen
RSA (2 % (w/v) in PS) 150 1 h RT
Probenauftrag Verdünnungen des Fermentations-überstands des Klons N55 in PBS
100 2 h 37° C
sekundärer Ak M-α-his (100 ng·ml-1) 100 2 h 37° C
AP-konjugierter Ak GAMAP (100 ng·ml-1) 100 1 h 37° C
Substratreaktion PNPP (1 mg·ml-1 in SP) 100 variabel, 37° C

Material und Methoden
33
2.3.8 Konzentrationsbestimmung des biscFv2429 mit Hilfe der Oberflächenplasmon-resonanz
Die Konzentrationsbestimmung des biscFv2429 (Fischer et al., 1999e) wurde mit Hilfe einer
BIACORE® 2000 (BioSensor) durchgeführt. Dabei wird die Bindung eines Analyten an einen
Liganden, der in einer Flußzelle immobilisiert ist, in Echtzeit gemessen. Das Ausgangssignal
wird in der Einheit RU ("Response units") angegeben und ist direkt proportional zur Masse des
gebunden Analyten. Das Meßprinzip und seine Anwendung für quantitative und kinetische
Messungen ist mehrfach beschrieben (Jonsson et al., 1991; Stenberg et al., 1991; Malmqvist und
Karlsson, 1997; Sack, 1998; Malmqvist, 1999).
Die Messung der Konzentration des biscFv2429 im Extrakt aus BY-2 Zellen (2.3.1) erfolgte
aufgrund der Bindung dieses Fragments an intakte Tabak-Mosaikvirionen (TMV), der in der
Meßzelle immobilisiert war. Die Oberfläche der Flußzelle wurde vor jeder Probe regeneriert und
die Probenvorbereitung und –verdünnung erfolgte stets nach dem gleichen Protokoll (s. u.). Als
Standard wurde das in etwa gleich große Fab-Fragment des mAk 24 gewählt, das ebenfalls eine
Bindungsstelle für TMV aufweist und daher ähnliche Meßergebnisse ergeben sollte. Es standen
auf 1 µg·ml-1 und 0.5 µg·ml-1 eingestellte Standardkonzentrationen zur Verfügung.
Die Proben wurden zentrifugiert (10000 g/2 min/4° C), um die Zelltrümmer abzutrennen und
der Überstand mit löslichen Bestandteilen wurde 1:4 in Laufpuffer verdünnt. Ein Probenzyklus
sah wie folgt aus:
Regeneration: 10 µl Regenerationspuffer (30 µL·min-1)
Binden: 20 µl Probe (5µl·min-1)
Laufpuffer, pH 7,410 mM HEPES
0,15 M NaCl0,5 mM EDTA
0,005 % (v/v) Tween 20
Regenerationspuffer30 mM HCl
2.3.9 Konzentrationsbestimmung des scFv4813
2.3.9.1 Konzentrationsbestimmung von scFv4813 mit Hilfe der Oberflächenplasmonresonanz
Die Konzentrationsbestimmung his6-markierter Proteine in Pichia-Kulturüberständen wurde
analog zu 2.3.8 mit Hilfe der Oberflächenplasmonresonanz durchgeführt. Im Falle des scFv4813

Material und Methoden
34
erfolgte die Messung aufgrund der Bindung des scFv4813 über den carboxyterminalen his6-Tag
an eine Nickel-NTA ("Nitrilo-triacetic acid") Oberfläche. Die NTA-Oberfläche der Flußzelle
wurde vor jeder Probe regeneriert und die Probenvorbereitung und –verdünnung erfolgte stets
nach dem gleichen Protokoll. Unter diesen Bedingungen entsprechen 1000 RU Konzentration
des his6-markierten Analyten etwa 10 µg·ml-1.
Die Proben wurden zentrifugiert (10000g/2min/4°C) um die Zelltrümmer abzutrennen, der
Überstand mit den löslichen Bestandteilen wurde 1:20 in Laufpuffer verdünnt.
Regeneration: 5 µl Regenerationspuffer (5µl·min-1)
Beladen mit Nickel: 10 µl Ladepuffer (5µl·min-1)
Binden: 20 µl Probe (5µl·min-1)
Laufpuffer, pH 7,410 mM HEPES
0,15 M NaCl0,005 % (v/v) Tween 20
Ladepuffer, pH 7,4:500 µM NiCl2
10 mM HEPES0,15 M NaCl
0,005 % (v/v) Tween 20
Regenerationspuffer pH 8,3:10 mM HEPES
0,15 M NaCl0,35 M EDTA
0,005 % (v/v) Tween 20
2.3.9.2 Photometrische Konzentrationsbestimmung
Die photometrische Bestimmung der Konzentration von gereinigtem scFv4813 (2.3.10) aus
Pichia pastoris (2.3.10) erfolgte bei 280 nm in PBS. Der theoretische millimolare
Extinktionskoeffizient wurde aus der Aminosäuresequenz mit Hilfe des im Internet vom "Swiss
Institute of Bioinformatics" (SIB) zur Verfügung gestellten Programms "ProtParam" ermittelt
(http://www.expasy.ch/tools/protparam.html (Stand 4/2000)).
Der theoretische millimolare Extinktionskoeffizient bei 280 nm ist 49170, demnach
entspricht eine E280 von 1 einer scFv-Konzentration von 0,56 mg·ml-1.
2.3.9.3 Konzentrationsbestimmung durch Bradford-Färbung
Die Konzentration von gereinigtem scFv4813 aus Pichia pastoris (2.3.10) wurde zusätzlich
durch Anfärbung mit dem Bradford-Assay (BioRad, München, siehe 2.3.2) ermittelt. 50 µl des

Material und Methoden
35
gereinigten Proteins wurden in verschiedenen Verdünnungen in Mikrotiterplatten vorgelegt.
Dazu wurden 160 µl einer 1:4 oder 1:10 verdünnten Lösung der Färbereagenz gegeben. Die
Extinktion bei 595 nm wurde nach 15minütiger Inkubation bei Raumtemperatur gemessen. Als
Standard diente RSA Fraktion V (Merck), das zwischen 5 und 50 µg·ml-1 eine lineare
Korrelation mit der Extinktion aufweist.
2.3.10 Reinigung von scFv4813 aus Pichia-Fermentationsüberstand
2.3.10.1 Metall-Affinitätschromatographie
Bei der Metall-Affinitätschromatographie wird ein zweiwertiges Kation, typischerweise Ni2+
oder Cu2+ durch einen kovalent an eine Matrix gebundenen Chelatbildner, z. B. NTA (Nitrilo-tri-
acetic acid) oder IDA (Imino-diacetic acid) gebunden. Mit der mobilen Phase aufgetragene
andere Chelatbildner, z. B. his6-markierte Proteine, können durch weitere Komplexierung des
Kations selektiv gebunden werden. Die Elution erfolgt durch starke, lösliche Chelatbildner wie
EDTA oder Imidazol. Selektive "Fang"-Schritte bzw. Affinitätschromatographie eignen sich
besonders als erster Schritt in einer Aufarbeitungskaskade, da mit der selektiven Bindung eine
effiziente Aufkonzentrierung des Produkts verbunden ist.
Für den scFv4813 aus Pichia wurde Ni-IDA Affinitätschromatographie in Kombination mit
der "expanded bed"-Technologie verwendet. Dabei werden die Partikel der Säulenmatrix
innerhalb der Säule von unten angeströmt und fluidisiert. Das Bett expandiert abhängig von der
Dichte, der Flußrate und dem Partikelgehalt der mobilen Phase um das drei- bis sechsfache, es
bleibt aber durch entsprechenden Schlankheitsgrad des Säulenkörpers und geeignete
Zulaufgeometrie bei einer idealen Verdrängungsströmung mit relativ enger
Verweilzeitverteilung. Dieses Verfahren ermöglicht es, stark partikelhaltige Suspensionen
aufzutragen, ohne daß es zum Verblocken der Säule kommt. Bei der Elution wird die Säule in
umgekehrter Richtung betrieben, so daß das Bett- und damit das Elutionsvolumen möglichst
gering ist.
Zum Einsatz kamen eine "Streamline 50"-Säule (Pharmacia) mit 75 ml "Streamline
Chelating" Matrix (Pharmacia). Die Matrix wurde zum Laden mit Nickel, Auftrag von
Fermentationsüberstand und Waschen (Puffer und Lösungen s. u.) im expandierten Zustand bei
einem Säulenvolumen (SV) von ca. 300 ml betrieben. Zur Elution wurde die Flußrichtung
umgekehrt, und der Stempel der Säule bis an die Obergrenze des nun gepackten Bettes
herabgefahren (SV=75 ml). Die Elution des scFv4813 wurde durch Messung der Absorption bei
280 nm in einer entsprechenden Fließzelle (BioRad) überwacht und der Eluatstrom von Hand

Material und Methoden
36
fraktioniert. Während der Elution wurde der Eluatstrom durch ein in Serie geschaltetes
Hohlfaserdialysemodul in den Laufpuffer für die anschließende Kationenaustauscher-
chromatographie umgepuffert (2.3.10.2).
Äquilibrierungspuffer, pH 7,41 M NaCl2,7 mM KCl8,1 mM Na2HPO4
1,5 mM KH2PO4
Nickel-Ladepuffer100 mM NiSO4
Elutionspuffer, pH 7,4250 mM Imidazol
1 M NaCl2,7 mM KCl8,1 mM Na2HPO4
1,5 mM KH2PO4
Dialysepuffer, pH 5.050 mM NaCH3COO
2.3.10.2 Kationenaustauscherchromatographie
Zur weiteren Reinigung des scFv4813 wurde wegen des hohen isoelektrischen Punktes des
Proteins ein Kationenaustauscher (Poros 50 HS) verwendet. Die Säule mit einem Bettvolumen
von 8 ml war in ein "Äkta Purifier" Reinigungssystem (Pharmacia) integriert, über dessen
Pumpen und Meßzellen die Beladung und Elution der Säule sowie die Fraktionierung des Eluats
gesteuert wurden. Die Säule wurde mit Laufpuffer äquilibriert und mit dem in Laufpuffer
dialysierten Eluat der Ni-Affinitätschromatographie beladen. Die Elution erfolgte mit einem
stufenlosen 0-100 % Gradienten in Elutionspuffer.
Laufpuffer, pH 5.050 mM NaCH3COO
Elutionspuffer, pH 5.050 mM NaCH3COO
1 M NaCl
2.3.10.3 Aufkonzentrierung des scFv4813
Zur Vorbereitung auf die Gelchromatographie wurde das Eluat der Kationenaustauscher-
chromatographie (2.3.10.2) durch Druckfiltration konzentriert. Dazu diente eine Ultrafiltrations-
Rührzelle (Millipore), die mit einer Celluloseacetatmembran mit 10 kDa Ausschlußgrenze

Material und Methoden
37
ausgestattet war. Die Zelle wurde mit 6 bar Stickstoff überlagert. Das Fassungsvermögen der
Rührzelle betrug max. 70 ml, die auf ein Minimum von etwa 5 ml konzentriert werden konnten.
2.3.10.4 Gelchromatographie
Die Gelchromatographie wurde als letzter Schritt in der Reinigung des scFv4813 eingesetzt.
Als Säulenfüllung wurden 450 ml Sephacryl S100 HR (Pharmacia) verwendet. Die Säule wurde
bei 4° C betrieben. Probenauftrag (aufkonzentriertes Eluat aus 2.3.10.3) und Elution mit
Gelfiltrations-Laufpuffer wurden mit dem "Äkta Explorer" Reinigungssystem (Pharmacia)
durchgeführt, das auch zur Detektion des Produkts und Fraktionierung des Eluats anhand der
Extinktionsmessung bei 280 nm verwendet wurde.
Laufpuffer pH, 7,55 mM TRIS2 mM DTT
0,5 mM EDTA
2.4 Kultivierung von Nicotiana tabacum cv BY-2
2.4.1 Stammhaltung
Die verwendeten Zellinien von Nicotiana tabacum cv BY-2 wurden als Suspensionskulturen
durch wöchentliches Überimpfen kultiviert. Dazu wurden 50 ml BY-2 Medium in einem
250-ml Erlenmeyerkolben mit 5% (v/v) einer 7 Tage alten Kultur inokuliert und bei 100 U·min-1
und bei 26° C im Dunkeln inkubiert.
BY-2 – Medium, pH 5,80,443 % (v/v) MSMO (Sigma)
1 mg·l-1 Thiamin HCl0,2 mg·l-1 2.4-D
200 mg·l-1 KH2PO4
100 mg·l-1 myo-Inosite3 % (w/v) Sucrose
2.4.2 Vorkultur
Zur Anzucht von Vorkulturen wurden die entsprechenden Zellinien in 250 ml BY-2 Medium
(2.4.1) in 1-l Erlenmeyerkolben 5%ig (v/v) überimpft und für 5 Tage bei 26° C und 100 U·min-1
kultiviert. Die Vorkulturen wurden einzeln mikroskopisch auf Kontaminationen überprüft, bevor
sie vereinigt und zur Inokulation einer Fermentation verwendet wurden.

Material und Methoden
38
2.4.3 Bestimmung von Frischgewicht, Trockengewicht und Zellanteil in BY-2 Kulturen
Zur Bestimmung des Zellanteils und des Frischgewichts wurden 10 ml einer BY-2 Kultur in
einem graduierten 14-ml Zentrifugenröhrchen (Polypropylen) bei 4300 g für 5 min in einem
Ausschwingrotor zentrifugiert. Danach wurde der Feststoffanteil ("Packed Cell Volume", PCV)
an der Graduierung des Röhrchens abgelesen. Die Genauigkeit dieser Methode liegt bei +/- 2%.
Der Überstand wurde dekantiert, die Zellen mit entionisiertem Wasser resuspendiert, mit Hilfe
einer Wasserstrahlpumpe auf einen 50-mm Cellulosefilter (Whatman) übertragen, und einmal
gewaschen. Anschließend wurden die Zellen quantitativ auf vorgewogene Aluminium-
Wägeschälchen übertragen und das Frischgewicht ermittelt. Die Genauigkeit dieser Methode
liegt bei Biomassekonzentrationen bis 50 g·l-1 bei +/- 3 % und bei Biomassekonzentrationen
zwischen 50 und 250 g·l-1 bei +/- 2 %.
Anschließend wurde die Biomasse bei 60° C bis zur Gewichtskonstanz (24 h) getrocknet und
das Trockengewicht mit einer Genauigkeit von +/- 5 % bestimmt.
2.4.4 Fermentation von Nicotiana tabacum cv BY-2
2.4.4.1 Fermentationen im 5-Liter Maßstab
Zur Vorbereitung der Fermentation wurde die pH-Elektrode des Fermenters kalibriert. 4 Liter
BY-2 Medium (2.4.1) wurden mit einigen Tropfen Antischaummittel (Roth) versetzt und
zusammen mit dem Reaktor sterilisiert (121° C, 30 min). Nach dem Abkühlen wurde die
Temperatur auf 26° C geregelt und die Sauerstoffelektrode bei einer konstanten Belüftungsrate
von 0,1 l·l-1·min-1 und einer Rührerdrehzahl von 130 U·min-1 auf 100 % Sauerstoffsättigung
kalibriert. Temperatur und Rührerdrehzahl wurden während der Fermentation konstant gehalten.
Der pH-Wert wurde gemessen, aber nicht geregelt. Die Luftzufuhr wurde über die
Gelöstsauerstoffkonzentration (dO2) derart gesteuert, daß die Luftzufuhr bei dO2-Werten über
20 % gestoppt wurde. Die Belüftungsrate bei offenem Ventil lag zu Beginn der Fermentation bei
0,1 l·l-1·min-1 und wurde manuell erhöht, wenn diese Rate bei konstant offener Luftzufuhr nicht
ausreichte, um 20 % dO2 zu halten, bis zu einer maximalen Belüftungsrate von 0,3 l·l-1·min-1. Im
Falle der Verwendung der Bioclave-Glasreaktoren wurde – sofern nicht anders angegeben – der
Reaktor während der Fermentation mit lichtdichter Folie abgedunkelt. Die Inokulation erfolgte
mit 900-1000 ml Vorkultur (2.4.2), die ein PCV von ca. 50 % (v/v) aufwies.
2.4.4.2 Fermentationen im 30-Liter Maßstab
Für Fermentationen im 30-l Maßstab wurden nach der Kalibration der pH-Elektrode 26 Liter
BY-2 Medium (2.4.1) gemeinsam mit einigen Tropfen Antischaummittel im Reaktor sterilisiert

Material und Methoden
39
(121° C, 30 min). Nach dem Abkühlen auf 26° C wurde die Sauerstoffelektrode bei einer
konstanten Belüftungsrate von 0,2 l·l-1·min-1 und einer Rührerdrehzahl von 80 U·min-1 auf 100 %
Sauerstoffsättigung kalibriert. Temperatur und Rührerdrehzahl blieben während der
Fermentation konstant. Der pH-Wert wurde gemessen, aber nicht geregelt. Die Luftzufuhr wurde
über die Gelöstsauerstoffkonzentration (dO2) derart gesteuert, daß die Luftzufuhr bei dO2-
Werten über 20 % gestoppt wurde. Die Belüftungsrate bei offenem Ventil lag zu Beginn der
Fermentation bei 0,2 l·l-1·min-1 und wurde manuell erhöht, wenn diese Rate bei konstant offener
Luftzufuhr nicht ausreichte, um 20 % dO2 zu halten, bis zu einer maximalen Belüftungsrate von
0,4 l·l-1·min-1. Als Inokulum diente die Fermentationsbrühe einer 5-l Fermentation (2.4.4.1) mit
einem PCV von 40-50 % (v/v). Dazu wurden der 5-l Reaktor und der 30-l Reaktor über einen
Silikonschlauch (ID 8 mm) miteinander verbunden, die Verbindung dampfsterilisiert und das
Inokulum durch Beaufschlagung des Animpffermenters mit 1 bar Überdruck steril in den 30-l
Kessel überführt.
2.5 Kultivierung von Pichia pastoris
2.5.1 Kryokulturen
Von allen Pichia-Klonen wurden Stammkulturen in Form von Kryokulturen angelegt. Ein
durch Quadrantenaustrich eines Stammes auf YPD-Agar vereinzelter rekombinanter Klon wurde
in 10 ml YPD-Flüssigmedium in einem 50-ml "Falcon"-Reaktionsgefäß (Greiner, Solingen)
inokuliert. Das Kulturröhrchen wurde unter aeroben Bedingungen bei 30° C unter starkem
Schütteln (250 U·min-1) inkubiert, bis eine Optische Dichte bei einer Wellenlänge von 600 nm
(OD600) von 1,5-2 erreicht war.
Die Kultur wurde nach mikroskopischer Kontrolle auf Kontaminationen durch Zugabe von
sterilem, 60%igem (v/v) Glyzerin auf eine Endkonzentration von 15 % (v/v) Glyzerin eingestellt
und in sterilen 1,5-ml-Eppendorfröhrchen (Greiner, Solingen) in 100-µl-Aliquots in flüssigem
Stickstoff eingefroren. Die Stammbank wurde anschließend bei –80° C gelagert. Die
Kryokulturen wurden in regelmäßigen Abständen durch Ausstrich auf YPD-Agar auf Sterilität
und Lebendzellzahl überprüft.

Material und Methoden
40
YPD-Flüssigmedium2 % (w/v) Hefeextrakt1 % (w/v) Pepton2 % (w/v) Glukose
YPD-Nährbodenentspricht YPD-Flüssigmedium mit 2 % (w/v) Agar
2.5.2 Vorkulturen
Der zu fermentierende Klon von Pichia pastoris wurde aus einer Kryokultur auf YPD-
Nährboden (2.5.1) ausgestrichen und 2 Tage bei 30° C inkubiert. Von diesem Ausstrich wurden
Vorkulturen in jeweils 10 ml BMGY oder YPG Vorkulturmedium in 50-ml Falcon-Gefäßen
angelegt und unter aeroben Verhältnissen inkubiert (30° C, 250 U·min-1, 24 h). Diese
Vorkulturen wurden verwendet, um jeweils 200 ml BMGY oder YPG in 1000-ml
Erlenmeyerkolben (mit Schikanen) anzuimpfen, die unter den gleichen Bedingungen für 16-24 h
inkubiert wurden. Die Vorkulturen wurden einzeln mikroskopisch auf Kontaminationen
überprüft, bevor sie vereinigt und zur Inokulation verwendet wurden.
BMGY Vorkulturmedium2 % (w/v) Hefeextrakt1 % (w/v) Pepton
1,37 % (w/v) Yeast Nitrogen Base, ohne Aminosäuren1 % (v/v) Glyzerin
100 mM KH2PO4
YPG Vorkulturmedium2 % (w/v) Hefeextrakt1 % (w/v) Pepton1 % (v/v) Glyzerin
2.5.3 Bestimmung von optischer Dichte, Frisch- und Trockengewicht
Im unteren Zelldichtebereich wurde die optische Dichte bei einer Wellenlänge von 600 nm
(OD600) von Pichia-Kulturen als Maß für die Biomassekonzentration herangezogen. Dazu
wurden die Kulturen im Verhältnis 1:20 in PBS (2.3.7) verdünnt und die OD600 gegen PBS
bestimmt. Die so erhaltenen Werte korrelieren bis zu einer OD600 von ca. 25 linear mit dem
Frischgewicht. In Hochzelldichtekulturen über 50 g·l-1 ergibt diese Methode aufgrund der
nichtlinearen Korrelation bei höheren Verdünnungsstufen ungenaue Werte.
Zur Bestimmung des Frischgewichts von Pichia-Kulturen wurden 2 x 1500 µl der jeweiligen
Kultur in zwei vorgewogene Eppendorf-Röhrchen pipettiert und die Zellen bei 4000 g für 2 min.
zentrifugiert. Der Überstand wurde dekantiert. Anschließend wurden die Röhrchen gewogen und

Material und Methoden
41
das Frischgewicht ermittelt. Mit dieser Methode läßt sich eine Genauigkeit von +/- 3 g·l-1
erzielen, was für den Hochzelldichtebereich ausreichend ist.
Zur Bestimmung des Trockengewichts wurden 10 ml einer Kultur bei 3400 g für 5 min. in
vorgewogenen 10-ml Zentrifugenröhrchen (Polypropylen) abzentrifugiert, der Überstand wurde
dekantiert und die Röhrchen wurden bei 60° C für 24 h getrocknet. Anschließend wurde die
Biotrockenmasse gravimetrisch bestimmt. Für Pichia pastoris ergab sich ein konstantes
Verhältnis von 1:4 – 1:3,8, d. h. ein Trockenanteil von 25-26 % am Frischgewicht.
2.5.4 Expression im Schüttelkolben
Zur Bestimmung der Methanol-Toleranz wurden jeweils 40 ml einer YPG-Vorkultur (2.5.2)
von Pichia pastoris Klon N55 (2.1.7.2) bei einer OD600 von 24 unter sterilen Bedingungen in 50-
ml "Falcon"-Gefäßen abzentrifugiert (1500 g, 5 min), der Überstand dekantiert und das Pellet in
10 ml YPM Induktionsmedium aufgenommen. Methanol wurde bis zur jeweiligen
Endkonzentration zugegeben und die Falcon-Röhrchen unter aeroben Bedingungen für 36 h
inkubiert (250 U·min-1, 30° C). Zu diesem Zeitpunkt wurden die Zellen zentrifugiert (5000 g,
5 min) und die Konzentration des scFv4813 per Oberflächenplasmonresonanz (2.3.9.1) im
Überstand bestimmt.
YPM Induktionsmedium2 % (w/v) Hefeextrakt1 % (w/v) Pepton
0,5-6,5 % (v/v) Methanol
2.5.5 Fermentation von Pichia pastoris
2.5.5.1 Vorbereitung und erste Phase der Fermentation
Das Pichia-Fermentationsmedium ohne die Spurenelementelösung wurde nach der
Kalibrierung der pH-Elektrode eingefüllt und zusammen mit dem Fermenter sterilisiert. Das
jeweils verwendete Medium und die Glyzerin-Ausgangskonzentration sind im Ergebnisteil
genannt. Anschließend wurden Zugabeflaschen für Antischaummittel und 28%ige
Ammoniaklösung steril angeschlossen und der pH-Wert wurde durch Zugabe von Ammoniak
auf 6,0 eingestellt. Ammoniak dient neben der pH-Kontrolle auch als einzige Stickstoffquelle
des Fermentationsmediums. Die sterilfiltrierten Ptm1-Spurenelemente und einige Tropfen
Antischaummittel wurden zugegeben, bevor die Sauerstoffelektrode bei Belüftungs- und
Rührwerkseinstellungen identisch mit denen während der Fermentation auf 100 %
Sauerstoffsättigung kalibriert wurde. Die Belüftungsrate wurde – falls nicht anders angegeben –

Material und Methoden
42
auf 2 l·l-1·min-1 (Preßluft) eingestellt. Bei Fermentationen im 5-l Reaktor (2.1.6) wurde der
Rührer auf konstant 1000 U·min-1 eingestellt, im 30-l Reaktor (2.1.6) auf 500 U·min-1. Alle
Fermentationen wurden bei einer Temperatur von 30° C durchgeführt.
Der erste Teil der Fermentation bestand in allen Fällen aus der Biomassebildung im
ansatzweisen Verfahren auf der Basis von Glyzerin als Kohlenstoffquelle ("glycerol batch
phase"). Der pH-Wert wurde durch automatische Zugabe von Ammoniaklösung konstant bei 6.0
gehalten. Während dieser Phase sinkt die Gelöstsauerstoffkonzentration (dO2) mit zunehmender
Biomasse ab, um dann plötzlich anzusteigen. Dieser Anstieg ("dO2 spike") signalisiert den
völligen Verbrauch des vorgelegten Glyzerins und das Ende der ersten Fermentationsphase.
Pichia Fermentationsmedium A (Invitrogen)2,2 mM CaSO4
18,8 mM MgSO4
22,6 mM KOH32,2 mM K2SO4
0,84 % (v/v) Phosphorsäure (85 %)4 % (v/v) Glyzerin
0,4 % (v/v) Ptm1 Spurenelementelösung (Invitrogen)
Pichia Fermentationsmedium B (verändert)1,1 mM CaSO4
9,4 mM MgSO4
11,3 mM KOH16,1 mM K2SO4
0,4 % (v/v) Phosphorsäure (85 %)5 % (v/v) Glyzerin
0,4 % (v/v) Ptm1 Spurenelementelösung (Invitrogen)
Ptm1 Spurenelementelösung (Invitrogen)230 mM FeSO4
24 mM CuSO4
19,9 mM MnSO4
2,10 mM CoCl2
0,53 mM NaI0,83 mM Na2MoO4
0,32 mM B(OH)3
0,15 mM ZnCl2
0,82 mM Biotin
2.5.5.2 Glyzerin-Zulaufverfahren
In einigen Fällen wurde nach Abschluß der Biomassebildung im ansatzweisen Verfahren
(2.5.5.1) eine zusätzliche Phase der Biomassebildung im limitierten Glyzerin-Zulaufverfahren
("limited glycerol fed-batch") durchgeführt, um vor Beginn der Induktion eine höhere
Biomassekonzentration zu erreichen. Einige Autoren halten diese Phase C-limitierten

Material und Methoden
43
Wachstums für mit einer Derepression des AOX1-Promotors verbunden und damit der
darauffolgenden Induktionsphase förderlich (Stratton et al., 1998).
Die Zufuhr der Ptm1-supplementierten Glyzerinlösung erfolgte intervallartig mit fester
Zufuhrrate, die im Ergebnisteil jeweils angegeben ist. Diese Art der Zugabe führt bei
limitierender Zufuhrrate zu stark schwankenden dO2-Werten.
Glyzerin-Zulaufmedium50 % (v/v) Glyzerin
1,2 % (v/v) Ptm1-Spurenelementelösung (2.5.5.1)
2.5.5.3 Methanol-Zulaufverfahren
Zur Induktion der Expression des scFv4813 wurde im Methanol-Zulaufverfahren fermentiert:
Nach Abschluß der Biomassebildung auf Basis von Glyzerin und vollständiger Erschöpfung des
Glyzerins wurden die Zellen auf die Methanolverwertung adaptiert. Zwischen der Erschöpfung
des Glyzerins und dem Beginn der Adaption lagen zum Teil technisch bedingte, bis zu 6-
stündige Phasen ohne Kohlenstoffquelle. Zur Induktion wurde mit Spurenelementelösung
supplementiertes Methanol automatisch gesteuert intervallartig zugeführt.
Methanol-Zulaufmedium98,8 % (v/v) Methanol
1,2 % (v/v) Ptm1-Spurenelementelösung (2.5.5.1)
Induktion mit manuell gewählter Zufuhrrate
Im einfachsten Fall wurde die Zufuhrrate manuell vorgewählt und es erfolgte keine
Rückkopplung auf den Regler (Abb. 2-2 A). Die Gelöstsauerstoffkonzentration (dO2) gab dabei
indirekte Hinweise auf die Methanolkonzentration: bei zu niedriger Zufuhrrate liegt der dO2-
Wert nahe 100 %, bei zu hoher Zufuhrrate liegt der dO2-Wert nahe 0, dann besteht die Gefahr
der Akkumulation von Methanol.
Induktion mit dO2-kontrollierter Zufuhrrate
Die Induktion mit dO2-gesteuerter Methanolzufuhr basiert auf der schnellen Reaktion des
dO2-Wertes auf die Erschöpfung der Kohlenstoffquelle. Die Steuereinheit ADI 1030 wurde dazu
so programmiert, daß bei Überschreiten eines dO2-Sollwertes die Methanolzufuhrpumpe
aktiviert wurde (Abb. 2-2 B). Bei dieser Art der Zufuhrsteuerung schwankt die
Methanolkonzentration um 0 %, da der dO2-Wert schon bei niedrigen Methanolkonzentrationen

Material und Methoden
44
unter den Sollwert absinkt. In dieser Konfiguration kann der dO2-Istwert nicht mehr zur
Steuerung der Belüftungsrate verwendet werden.
BA
dO2
const.
dO2
C
MeOH
dO2
Abb. 2-2: Die verschiedenen Regelungsstrategien zur Steuerung der Methanol-Zufuhrrate. A:Manuelle Vorwahl einer konstanten Rate ohne Rückkopplung. B: Automatische Steuerung derMethanolzufuhr abhängig von der Gelöstsauerstoffkonzentration. C: Automatische Steuerungder Methanolzufuhr durch Echtzeit-Methanol-Analytik.
2.5.5.4 Induktion mit Echtzeit-Methanol-Kontrolle
Um Methanolkonzentrationen in einem Bereich oberhalb von 0 % regeln zu können, wurde
eine Echtzeit-Methanol-Analytik aufgebaut (Abb. 2-2 C). Das System basiert auf dem
kommerziell erhältlichen Alkoholsensor (ALKOSENS II) und der dazugehörigen
Regelungseinheit (ACETOMAT) der Fa. Heinrich Frings GmbH & Co KG (Bonn) und wird in
der industriellen Essigsäurefermentation eingesetzt. Der in-situ sterilisierbare Sensor besteht aus
einer Membran, durch die flüchtige Alkohole aus der Fermentationsbrühe pervaporieren. Auf der
Rückseite der Membran transportiert ein konstanter Luftstrom die Moleküle zu einem beheizten,
dotierten SnO2 Halbleitersensor, an dem durch die Oxidation der Alkohole eine Verringerung
des Widerstandes bewirkt wird (Abb. 2-3).
In der Regelungseinheit wird der gemessene Widerstand mit einer nichtlinearen
Kalibrationskurve korreliert und digital sowie analog als Methanolkonzentration ausgegeben und
mit einem Sollwert verglichen. Abhängig vom Sollwert können interne Relais angesteuert
werden. Im vorliegenden Fall wurde der ACETOMAT so konfiguriert, daß eine externe
Zufuhrpumpe bei Unterschreiten des Sollwertes eine bestimmte Menge Methanol, die
0,08 % (v/v) entsprach, förderte.

Material und Methoden
45
���������������������������������������
����������������������������������������������������������
����������������������
����������������������������
TrägergasHalbleiter-Gassensor
Selektive Membran
Abb. 2-3: Funktionsprinzip des Methanolsensors: Flüchtige Alkohole pervaporieren durch dieselektive Membran und werden von einem Trägergasstrom zum Gassensor transportiert.
Der Analogausgang wurde über einen A/D-Wandler in das Prozessleitsystem BioXpert
eingespeist und so die Methanolkonzentration synchron mit den übrigen Prozessdaten erfaßt.

Ergebnisse und Diskussion
46
3 Ergebnisse und Diskussion
Vorangegangene Arbeiten am Institut für Biologie I der RWTH Aachen hatten bereits die
Expression von Antikörperfragmenten in pflanzlichen Suspensionskulturen (Drossard, 1996;
Schumann, 1996; Drossard et al., 1997; Sack, 1998; Raven, 1999) behandelt. Die Eignung dieses
Expressionssystem zur Produktion rekombinanter Antikörper hatte sich dabei erwiesen. Die
Zellinie Nicotiana tabacum cv BY-2 erwies sich als genetisch stabil und erbrachte bei gutem
Wachstumsverhalten reproduzierbare Produktausbeuten. Es hatte sich gezeigt, daß die Sekretion
von Antikörperfragmenten in den Apoplasten mit Hilfe entsprechender „Leader“-Peptide
möglich ist, die Akkumulation im ER mit Hilfe des KDEL Retentionssignals aber höhere
Ausbeuten erbringt.
Das Pichia pastoris Expressionssystems war am Institut für Biologie I als alternatives
Expressionssystem für die Produktion sekretierter, rekombinanter Proteine etabliert worden. Die
erfolgreiche Transformation, Selektion und Expression eines humanen Glykoproteins und eines
murinen scFv-Fragments sind in den jeweiligen Arbeiten von Robin und Raven beschrieben
(Robin, 1998; Raven, 1999).
Im Rahmen der vorliegenden Promotionsarbeit stand aufbauend auf den vorgenannten
Ergebnissen die Entwicklung und Optimierung von Fermentationsstrategien für die
Suspensionskultur von Nicotiana tabacum cv BY-2 sowie Pichia pastoris im Vordergrund. Der
diagnostisch relevante murine scFv4813 sollte im Maßstab bis 500 mg produziert werden.
3.1 Entwicklung eines Fermentationsprotokolls für Nicotiana tabacum cvBY-2
Die Entwicklung eines Fermentationsprotokolls für Nicotiana tabacum cv BY-2 erfolgte
anhand einer im Institut für Biologie I bereits bestehenden transgenen Zellinie, die einen biscFv
(Abb. 1-1) produzierte (Schumann, 1996), da zu Beginn dieser Promotionsarbeit die
Entwicklung einer Zellinie zur Produktion des scFv4813 noch nicht abgeschlossen war (Raven,
1999).
3.1.1 Wachstumskinetik von Nicotiana tabacum cv BY-2 im Rührreaktor
Initiale Versuche mit 5 % bzw. 10 % (v/v) Inokulumgröße hatten ergeben, daß für das
Anwachsen der BY-2 Zellinie biscFv2429 im Fermenter ein Inokulum in der Größenordnung
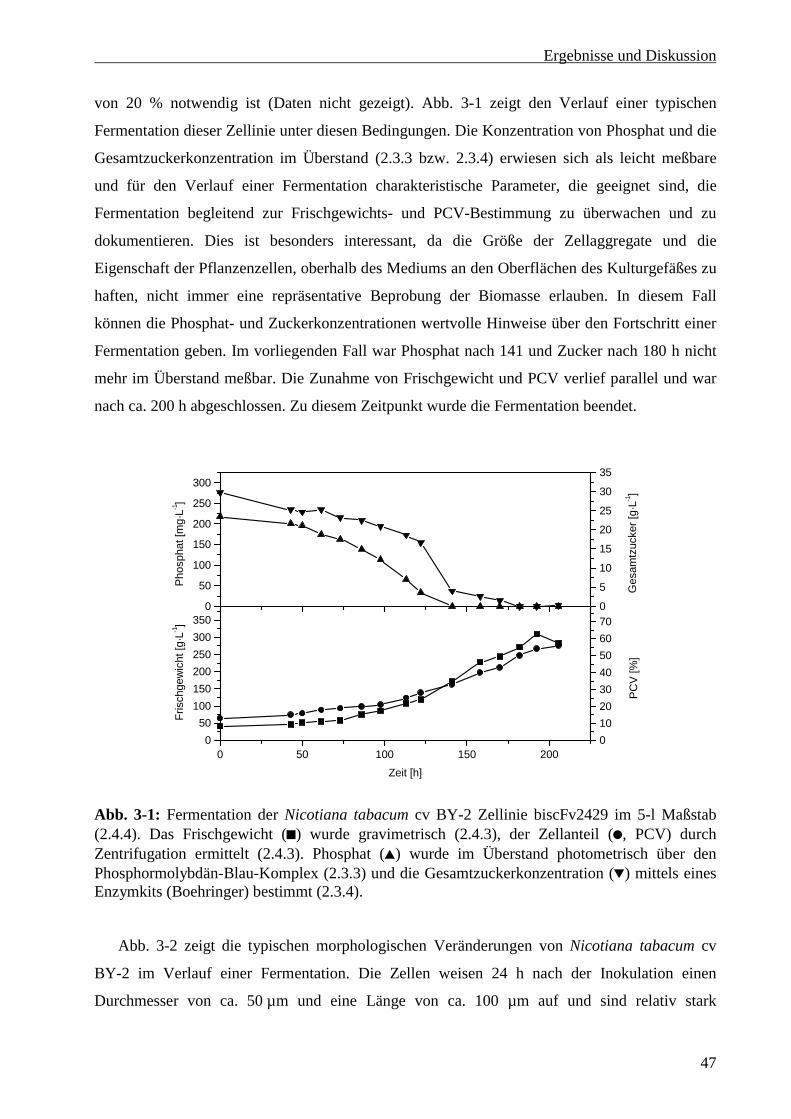
Ergebnisse und Diskussion
47
von 20 % notwendig ist (Daten nicht gezeigt). Abb. 3-1 zeigt den Verlauf einer typischen
Fermentation dieser Zellinie unter diesen Bedingungen. Die Konzentration von Phosphat und die
Gesamtzuckerkonzentration im Überstand (2.3.3 bzw. 2.3.4) erwiesen sich als leicht meßbare
und für den Verlauf einer Fermentation charakteristische Parameter, die geeignet sind, die
Fermentation begleitend zur Frischgewichts- und PCV-Bestimmung zu überwachen und zu
dokumentieren. Dies ist besonders interessant, da die Größe der Zellaggregate und die
Eigenschaft der Pflanzenzellen, oberhalb des Mediums an den Oberflächen des Kulturgefäßes zu
haften, nicht immer eine repräsentative Beprobung der Biomasse erlauben. In diesem Fall
können die Phosphat- und Zuckerkonzentrationen wertvolle Hinweise über den Fortschritt einer
Fermentation geben. Im vorliegenden Fall war Phosphat nach 141 und Zucker nach 180 h nicht
mehr im Überstand meßbar. Die Zunahme von Frischgewicht und PCV verlief parallel und war
nach ca. 200 h abgeschlossen. Zu diesem Zeitpunkt wurde die Fermentation beendet.
0 50 100 150 2000
50
100
150
200
250
300
350
Fris
chge
wic
ht [g
·L-1]
Zeit [h]
0
50
100
150
200
250
300
Pho
spha
t [m
g·L-1
]
0
10
20
30
40
50
60
70P
CV
[%]
0
5
10
15
20
25
30
35
Ges
amtz
ucke
r [g
·L-1]
Abb. 3-1: Fermentation der Nicotiana tabacum cv BY-2 Zellinie biscFv2429 im 5-l Maßstab(2.4.4). Das Frischgewicht (!) wurde gravimetrisch (2.4.3), der Zellanteil (", PCV) durchZentrifugation ermittelt (2.4.3). Phosphat (#) wurde im Überstand photometrisch über denPhosphormolybdän-Blau-Komplex (2.3.3) und die Gesamtzuckerkonzentration ($) mittels einesEnzymkits (Boehringer) bestimmt (2.3.4).
Abb. 3-2 zeigt die typischen morphologischen Veränderungen von Nicotiana tabacum cv
BY-2 im Verlauf einer Fermentation. Die Zellen weisen 24 h nach der Inokulation einen
Durchmesser von ca. 50 µm und eine Länge von ca. 100 µm auf und sind relativ stark

Ergebnisse und Diskussion
48
vakuolisiert. Nach ca. 100 h haben die Zellen mehrere Teilungszyklen vollzogen, wobei eine
Teilung jeweils senkrecht auf die letzte Teilungsebene erfolgt. Diese Teilungen erfolgen unter
nur geringer Volumenzunahme, so daß sich in erster Linie die Zellzahl, weniger das PCV erhöht.
In diesem Zustand sind die Zellen kubisch bis kugelig mit einem Durchmesser von ca. 25 µm.
Die Zellen bilden Aggregate von ca. 8-80 Zellen. Gegen Ende der Fermentation weisen die
Zellen nach erfolgtem Streckungswachstum wieder eine starke Vakuolisierung und eine Länge
von 100 bis 150 µm auf. Die Vakuolisierung und Streckung ist an eine starke Zunahme des PCV
sowie an ein Auseinanderbrechen der Aggregate gebunden. Da die Zellen einer BY-2 Kultur ein
synchrones Wachstumsverhalten aufweisen (Nagata et al., 1992), stellt die lichtmikroskopische
Untersuchung ebenso wie die Bestimmung des Phosphat- und Gesamtzuckergehaltes eine
geeignete Methode dar, dem Verlauf der Fermentation neben der Bestimmung von PCV und
Frischgewicht zu folgen.
Abb. 3-2: Morphologie von Nicotiana tabacum cv BY-2 nach 24 h (A), 100 h (B) und 180 h (C)im Verlauf einer Fermentation. Die Proben wurden aus einer Fermentation der ZelliniebiscFv2429 (2.1.7.1) nach Standardbedingungen (2.4.4) gezogen. Die Hellfeld-Aufnahmen(Leitz Dialux 20, Wild Photoautomat MP55) wurden bei 400facher Vergrößerung angefertigt.Die Skalen wurden durch Abfotografieren der Skalierungen auf entsprechenden Objektträgern(Leitz) mit der gleichen Objektiv/Okular Kombination erstellt.
Die Eigenschaft der Zellen, zunächst Nährstoffe aus dem Medium aufzunehmen, sich zu
teilen und erst anschließend durch Wasseraufnahme, Vakuolisierung und Streckungswachstum
die Größe der individuellen Zellen wieder zu erhöhen, führt zu einem nicht-synchronen Verlauf
der Frisch- und Trockengewichtsentwicklung. Während sich Frischgewicht und PCV parallel
entwickeln, steigt das Trockengewicht zunächst stärker an, um nach der vollständigen Aufnahme
der Nährstoffe konstant zu bleiben, während PCV und Frischgewicht noch stark ansteigen
(Daten nicht gezeigt). Der Trockengewichtsanteil von Nicotiana tabacum BY-2 Biomasse
schwankt daher zwischen 4 % und 7,5 % des Frischgewichts.
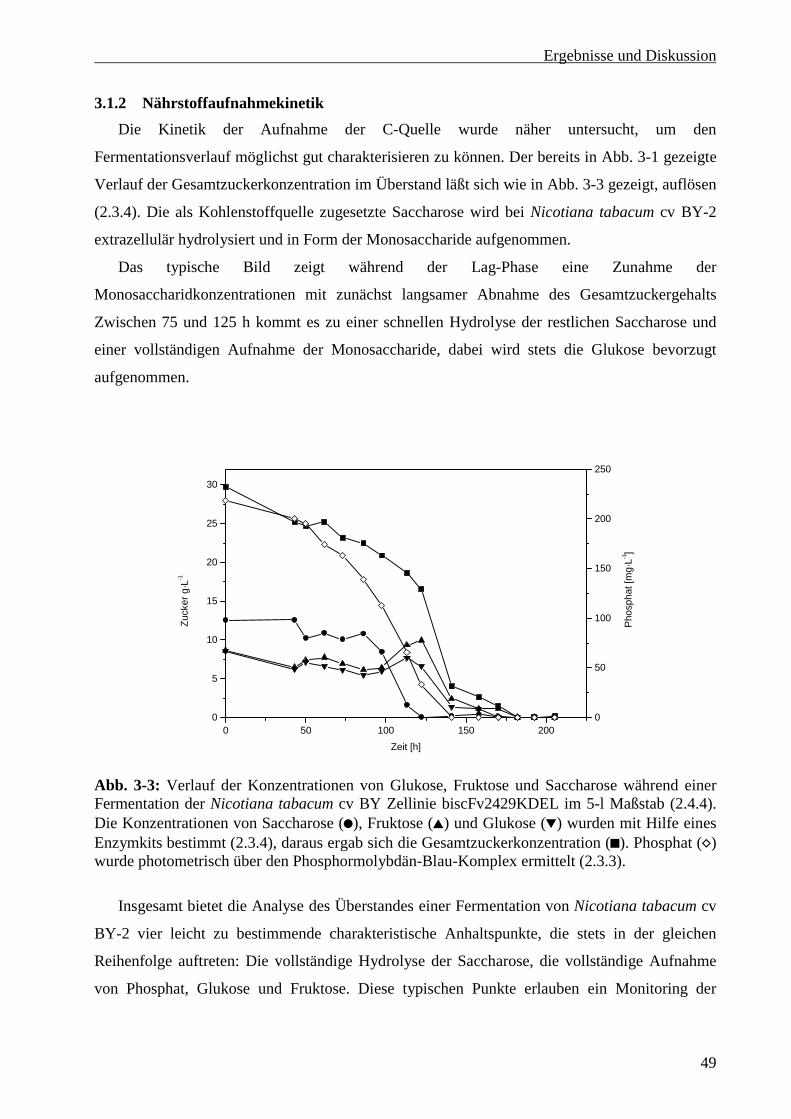
Ergebnisse und Diskussion
49
3.1.2 Nährstoffaufnahmekinetik
Die Kinetik der Aufnahme der C-Quelle wurde näher untersucht, um den
Fermentationsverlauf möglichst gut charakterisieren zu können. Der bereits in Abb. 3-1 gezeigte
Verlauf der Gesamtzuckerkonzentration im Überstand läßt sich wie in Abb. 3-3 gezeigt, auflösen
(2.3.4). Die als Kohlenstoffquelle zugesetzte Saccharose wird bei Nicotiana tabacum cv BY-2
extrazellulär hydrolysiert und in Form der Monosaccharide aufgenommen.
Das typische Bild zeigt während der Lag-Phase eine Zunahme der
Monosaccharidkonzentrationen mit zunächst langsamer Abnahme des Gesamtzuckergehalts
Zwischen 75 und 125 h kommt es zu einer schnellen Hydrolyse der restlichen Saccharose und
einer vollständigen Aufnahme der Monosaccharide, dabei wird stets die Glukose bevorzugt
aufgenommen.
0 50 100 150 2000
5
10
15
20
25
30
Zuc
ker
g·L-1
Zeit [h]
0
50
100
150
200
250
Pho
spha
t [m
g·L-1
]
Abb. 3-3: Verlauf der Konzentrationen von Glukose, Fruktose und Saccharose während einerFermentation der Nicotiana tabacum cv BY Zellinie biscFv2429KDEL im 5-l Maßstab (2.4.4).Die Konzentrationen von Saccharose ("), Fruktose (#) und Glukose ($) wurden mit Hilfe einesEnzymkits bestimmt (2.3.4), daraus ergab sich die Gesamtzuckerkonzentration (!). Phosphat (%)wurde photometrisch über den Phosphormolybdän-Blau-Komplex ermittelt (2.3.3).
Insgesamt bietet die Analyse des Überstandes einer Fermentation von Nicotiana tabacum cv
BY-2 vier leicht zu bestimmende charakteristische Anhaltspunkte, die stets in der gleichen
Reihenfolge auftreten: Die vollständige Hydrolyse der Saccharose, die vollständige Aufnahme
von Phosphat, Glukose und Fruktose. Diese typischen Punkte erlauben ein Monitoring der

Ergebnisse und Diskussion
50
Fermentation auch ohne zuverlässige Bestimmung der Biomassekonzentration und eine – mit
Hilfe entsprechender Analytik in Echtzeit durchführbare – Dokumentation einer solchen
Fermentation.
3.1.3 Kinetik der Expression des biscFv2429
Die Expression des biscFv2429 im Verlauf der oben im Detail analysierten Fermentation ist
in Abb. 3-4 gezeigt. Die Konzentration des Antikörperfragments wurde in diesem Fall per
ELISA (2.3.7.1) gemessen, wobei das Fab24-Fragment des mAk24 (2.1.4) als Standard
verwendet wurde. Sie lag über den Verlauf der Fermentation im Zellextrakt (2.3.1) zwischen
1163 und 2261 ng·ml-1, dabei lagen die Werte zu Beginn und zum Ende der Fermentation
tendenziell niedriger. Die mit Hilfe der Bradford-Färbung (2.3.2) gemessene Konzentration an
Gesamtprotein zeigte die gleiche Tendenz, mit den Extrema 508 und 2357 µg·ml-1 war die
Schwankungsbreite jedoch größer. Bei der Bestimmung des gesamtlöslichen Proteins wurde
RSA Fraktion V als Standard verwendet. Da die Empfindlichkeit der Reaktion von Proteinen mit
Bradford-Färbereagenz von Protein zu Protein unterschiedlich ist und die Anfärbung von RSA
bekanntermaßen relativ empfindlich ist, unterliegt die Bestimmung des gesamtlöslichen Proteins
nach dieser Methode einem nicht näher quantifizierbaren Fehler – die tatsächlichen Werte
dürften höher liegen. Das Verhältnis der Gesamtprotein-Konzentrationen der einzelnen Proben
zueinander bleibt davon unberührt.
Entsprechend dem tendenziell gleichen Verlauf von biscFv2429 und Gesamtprotein ergibt
sich die ebenfalls in Abb. 3-4 gezeigte Expressionsrate, die in etwa konstant um den Mittelwert
von ca. 0,125 % lag. Die Tatsache, daß die Konzentration an biscFv2429 außerhalb der Phasen
intensiver metabolischer Aktivität, wenn die Gesamtproteinkonzentration niedrig ist, kaum
absinkt, entspricht den Erwartungen für die Expression eines rekombinanten Proteins über einen
konstitutiven Promotor und dessen Akkumulation im ER.

Ergebnisse und Diskussion
51
0 50 100 150 2000
500
1000
1500
2000
2500
Ges
amtlö
slic
hes
Pro
tein
[µg·
mL-1
]
Zeit [h]
0,00
0,05
0,10
0,15
0,20
0,25
0,30
Exp
ress
ions
rate
[%]
0
500
1000
1500
2000
2500
bisc
Fv2
429
[ng·
mL-1
]
Abb. 3-4: Analyse der intrazellulären Akkumulation des biscFv2429 während einerFermentation der Nicotiana tabacum cv BY-2 Zellinie biscFv2429KDEL im 5-l Maßstab (2.4.4).Die Konzentration des biscFv2429 (#) wurde per ELISA (2.3.7.1) bestimmt und zurKonzentration des gesamtlöslichen Proteins (2.3.2) im Zellextrakt (") in Beziehung gesetzt, umdie Expressionsrate in % des Gesamtproteins (&' zu bestimmen.
Das Verhältnis von Expressionsrate und volumetrischer Produktausbeute bei einem
intrazellulären Produkt verdeutlicht Abb. 3-5. Obwohl die Konzentration des biscFv2429 im
Zellextrakt zur Mitte der Fermentation am höchsten liegt (#, A), ist die Gesamtmenge des
Produktes gegen Ende der Fermentation am höchsten und korreliert im Verlauf der Fermentation
im wesentlichen mit dem PCV (B). Aus diesem Grund, sowie aufgrund des schwankenden
Trockengewichtsanteils (3.1.1) wird im folgenden das PCV als Maß für die
Biomassekonzentration verwendet.
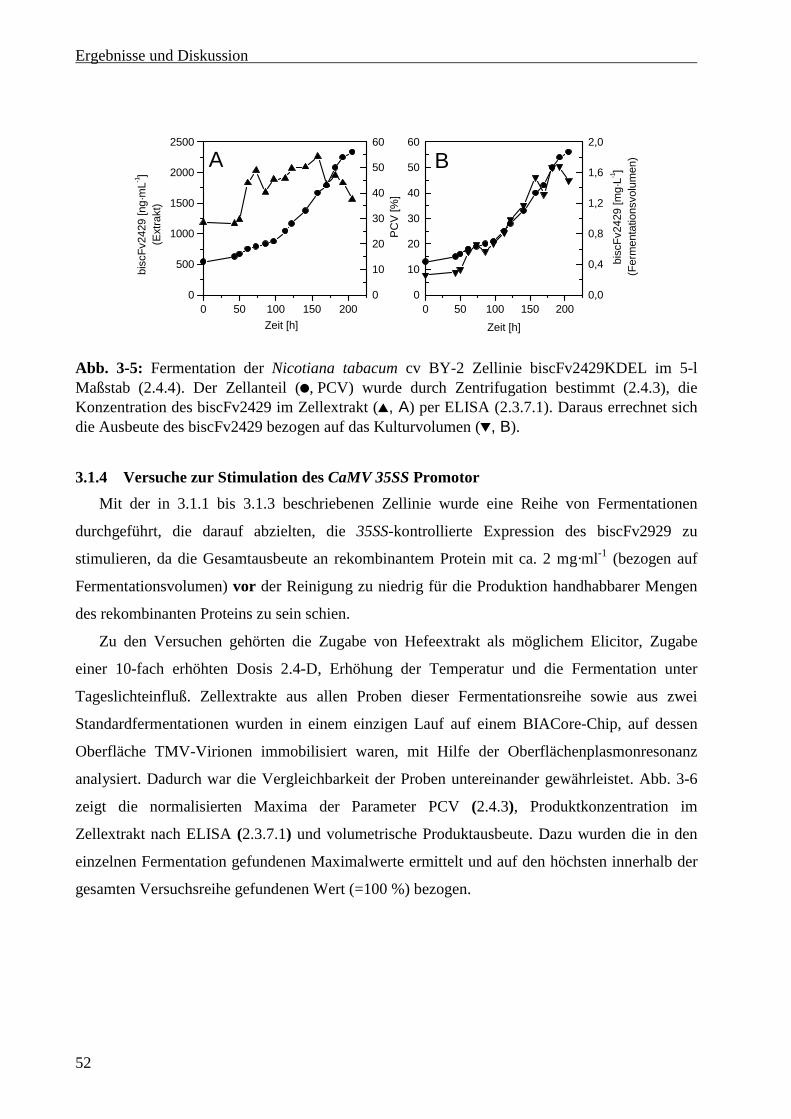
Ergebnisse und Diskussion
52
0 50 100 150 2000
500
1000
1500
2000
2500
BA
bi
scF
v242
9 [n
g·m
L-1]
(Ext
rakt
)
Zeit [h]
0 50 100 150 2000
10
20
30
40
50
60
PC
V [%
]
Zeit [h]
0
10
20
30
40
50
60
0,0
0,4
0,8
1,2
1,6
2,0
bisc
Fv2
429
[mg·
L-1]
(Fer
men
tatio
nsvo
lum
en)
Abb. 3-5: Fermentation der Nicotiana tabacum cv BY-2 Zellinie biscFv2429KDEL im 5-lMaßstab (2.4.4). Der Zellanteil (", PCV) wurde durch Zentrifugation bestimmt (2.4.3), dieKonzentration des biscFv2429 im Zellextrakt (#, A) per ELISA (2.3.7.1). Daraus errechnet sichdie Ausbeute des biscFv2429 bezogen auf das Kulturvolumen ($, B).
3.1.4 Versuche zur Stimulation des CaMV 35SS Promotor
Mit der in 3.1.1 bis 3.1.3 beschriebenen Zellinie wurde eine Reihe von Fermentationen
durchgeführt, die darauf abzielten, die 35SS-kontrollierte Expression des biscFv2929 zu
stimulieren, da die Gesamtausbeute an rekombinantem Protein mit ca. 2 mg·ml-1 (bezogen auf
Fermentationsvolumen) vor der Reinigung zu niedrig für die Produktion handhabbarer Mengen
des rekombinanten Proteins zu sein schien.
Zu den Versuchen gehörten die Zugabe von Hefeextrakt als möglichem Elicitor, Zugabe
einer 10-fach erhöhten Dosis 2.4-D, Erhöhung der Temperatur und die Fermentation unter
Tageslichteinfluß. Zellextrakte aus allen Proben dieser Fermentationsreihe sowie aus zwei
Standardfermentationen wurden in einem einzigen Lauf auf einem BIACore-Chip, auf dessen
Oberfläche TMV-Virionen immobilisiert waren, mit Hilfe der Oberflächenplasmonresonanz
analysiert. Dadurch war die Vergleichbarkeit der Proben untereinander gewährleistet. Abb. 3-6
zeigt die normalisierten Maxima der Parameter PCV (2.4.3), Produktkonzentration im
Zellextrakt nach ELISA (2.3.7.1) und volumetrische Produktausbeute. Dazu wurden die in den
einzelnen Fermentation gefundenen Maximalwerte ermittelt und auf den höchsten innerhalb der
gesamten Versuchsreihe gefundenen Wert (=100 %) bezogen.
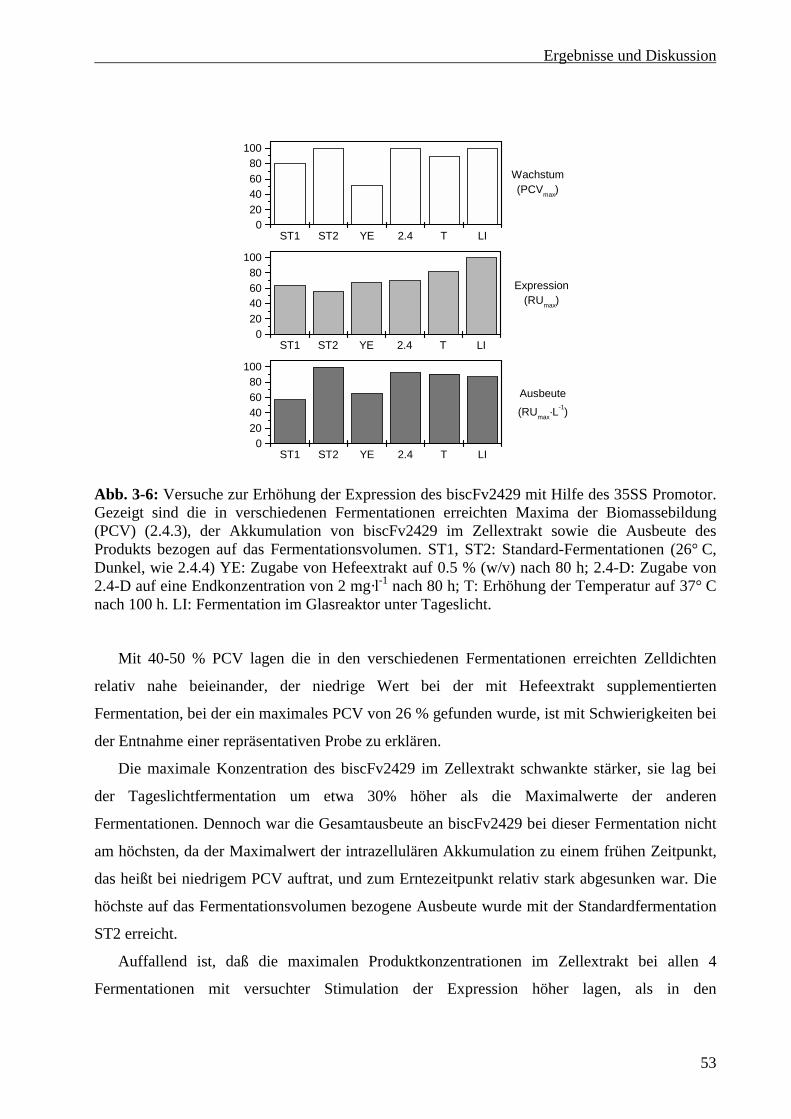
Ergebnisse und Diskussion
53
ST1 ST2 YE 2.4 T LI0
20
40
60
80100
ST1 ST2 YE 2.4 T LI0
20
40
60
80
100
Ausbeute
(RUmax
·L-1)
Wachstum(PCV
max)
Expression(RU
max)
ST1 ST2 YE 2.4 T LI0
20
4060
80
100
Abb. 3-6: Versuche zur Erhöhung der Expression des biscFv2429 mit Hilfe des 35SS Promotor.Gezeigt sind die in verschiedenen Fermentationen erreichten Maxima der Biomassebildung(PCV) (2.4.3), der Akkumulation von biscFv2429 im Zellextrakt sowie die Ausbeute desProdukts bezogen auf das Fermentationsvolumen. ST1, ST2: Standard-Fermentationen (26° C,Dunkel, wie 2.4.4) YE: Zugabe von Hefeextrakt auf 0.5 % (w/v) nach 80 h; 2.4-D: Zugabe von2.4-D auf eine Endkonzentration von 2 mg·l-1 nach 80 h; T: Erhöhung der Temperatur auf 37° Cnach 100 h. LI: Fermentation im Glasreaktor unter Tageslicht.
Mit 40-50 % PCV lagen die in den verschiedenen Fermentationen erreichten Zelldichten
relativ nahe beieinander, der niedrige Wert bei der mit Hefeextrakt supplementierten
Fermentation, bei der ein maximales PCV von 26 % gefunden wurde, ist mit Schwierigkeiten bei
der Entnahme einer repräsentativen Probe zu erklären.
Die maximale Konzentration des biscFv2429 im Zellextrakt schwankte stärker, sie lag bei
der Tageslichtfermentation um etwa 30% höher als die Maximalwerte der anderen
Fermentationen. Dennoch war die Gesamtausbeute an biscFv2429 bei dieser Fermentation nicht
am höchsten, da der Maximalwert der intrazellulären Akkumulation zu einem frühen Zeitpunkt,
das heißt bei niedrigem PCV auftrat, und zum Erntezeitpunkt relativ stark abgesunken war. Die
höchste auf das Fermentationsvolumen bezogene Ausbeute wurde mit der Standardfermentation
ST2 erreicht.
Auffallend ist, daß die maximalen Produktkonzentrationen im Zellextrakt bei allen 4
Fermentationen mit versuchter Stimulation der Expression höher lagen, als in den

Ergebnisse und Diskussion
54
Standardfermentationen. Daß die Gesamtausbeute dennoch bei einer Standardfermentation am
höchsten war, könnte daran liegen, daß die Expression zwar stimuliert wurde, dieser Effekt
jedoch von kurzer Dauer war. In diesem Fall könnte durch eine Verlagerung der exogenen
Stimulation zum Ende der Fermentation hin eine Verbesserung erzielt werden. Entscheidend für
die hohe volumetrische Produktivität bei der mit ST2 bezeichneten Fermentation war ein hohes
PCV zum Erntezeitpunkt in Verbindung mit einem relativ hohen Produktgehalt. Anhand der
durchgeführten Fermentationen ließ sich eine Stimulation der CaMV-35SS gesteuerten
Expression nicht zeigen.
Insgesamt waren die mit diesem System erreichten Konzentrationen des Produktes im
Zellextrakt immer noch niedrig, die höchste unter Verwendung dieses Promotors gefundene
Konzentration lag bei von 3,1 µg·ml-1, dieser Wert entspräche bei 55 % PCV, dem höchsten in
einer Fermentation gemessenen Wert, einer volumenbezogenen Ausbeute von 2,9 mg·l-1. Die
Reinigung rekombinanter Proteine aus dem komplexen Gesamtproteinextrakt von
Pflanzensuspensionskulturen ist aber mit Ausbeuteverlusten verbunden, so daß für die
Produktion handhabbarer Mengen des scFv4813 nach Alternativen gesucht wurde.
3.2 Produktion von scFv4813 in Nicotiana tabacum cv BY-2
3.2.1 Produktion von scFv 4813 im 5-Liter Maßstab
Aufgrund der mit dem CaMV 35SS erreichten Ergebnisse wurde für die Expressionskassette
des scFv4813 der (ocs)3mas Promotor gewählt, mit dem in Tabaksuspensionskulturen höhere
Expressionsraten erreichbar sind (2.1.7). Die Fermentation der Zellinie R1-25 zur Produktion des
scFv4813 in Pflanzensuspensionskultur verlief hinsichtlich der Wachstumskinetik entsprechend
den mit der Zellinie biscFv2429KDEL gemachten Erwartungen. Abb. 3-7 zeigt Biomassebildung
und Verlauf von pH-Wert und Gelöstsauerstoffkonzentration während einer Fermentation im 5-l
Reaktor.

Ergebnisse und Diskussion
55
00 48 96 144 1923
4
5
6
7
pH
Zeit [h]
5101520253035404550
PC
V [%
]
0
10
20
30
40
50
dO2
[%]
406080100120140160180200220
Fris
chge
wic
ht [g
·L-1]
Abb. 3-7: Fermentation der Zellinie R1-25 von Nicotiana tabacum cv BY-2 im 5-l Maßstab(2.4.4). Frischgewicht (!) wurde gravimetrisch bestimmt (2.4.3), der Zellanteil (", PCV) durchZentrifugation (2.4.3). Die Gelöstsauerstoffkonzentration dO2 (durchgezogene Linie) wurdedurch getaktete Belüftung auf 20 % konstant gehalten, die maximale Belüftungsrate war auf0.2 vvm begrenzt. Der pH-Wert (gestrichelte Linie) wurde gemessen, aber nicht geregelt.
Der Abfall der Gelöstsauerstoffkonzentration nach ca. 96 h ist auf eine zu niedrig eingestellte
maximale Belüftungsrate zurückzuführen: Bis ca. 88 h wurde die Luftzufuhr automatisch
getaktet um ein dO2 von 20% zu halten, danach stieg die Sauerstoffaufnahmerate der Zellen aber
weiter an, so daß die Belüftungsrate bei konstant offenem Ventil nicht ausreichte, 20 % zu
halten. Das Wachstumsverhalten der Zellen läßt keine Rückschlüsse zu, ob die kritische
Gelöstsauerstoffkonzentration, bei der das Wachstum der Zellen infolge Sauerstoffmangels
verlangsamt wird, unterschritten wurde. Mit einem PCV von 49 % und einem Frischgewicht von
215 g·l-1 wurden zum Erntezeitpunkt bei 190 h normale Werte erreicht.
3.2.2 Maßstabsvergrößerung in den 30-Liter Maßstab
Die im 5-l Reaktor angezogenen Zellen wurden direkt zur Inokulation des 30-l Reaktors
verwendet. Zum Transfer wurden die beiden Reaktoren steril miteinander verbunden und der 5-l
Reaktor mit 0,5 bar Überdruck beaufschlagt. Dadurch konnten eventuelle scherkraftbedingte
Zellschädigungen durch die Verwendung einer Pumpe vermieden werden. Der gesamte Inhalt
des 5-l Reaktors wurde in den 30-l Reaktor überführt, so daß die Größe des Inokulums ca. 16 %
(v/v) betrug. Abb. 3-8 zeigt den Verlauf der Fermentation im 30-l Maßstab.

Ergebnisse und Diskussion
56
00 24 48 72 96 120 144 1683
4
5
pH
Zeit [h]
0
10
20
30
40
PC
V [%
]
0
20
40
60
80
100
dO2
[%]
0
25
50
75
100
125
150
Fris
chge
wic
ht [g
·l-1]
Abb. 3-8: Fermentation der Zellinie R1-25 von Nicotiana tabacum cv BY-2 im 30-l Maßstab(2.4.4). Die Gelöstsauerstoffkonzentration dO2 (durchgezogene Linie) wurde durch getakteteBelüftung konstant auf 20 % gehalten. Das Frischgewicht (!) wurde gravimetrisch bestimmt(2.4.3), der Zellanteil (", PCV) durch Zentrifugation (2.4.3). Der pH-Wert (gestrichelte Linie)wurde gemessen, aber nicht geregelt.
Aufgrund starken Anhaftens der Zellen oberhalb der Flüssigkeitsoberfläche ("wall growth"),
möglicherweise auch aufgrund der ungünstigeren Lage des Probennahmeventils im 30-l Reaktor,
lagen die in den Proben erhaltenen PCV-Meßwerte deutlich unterhalb der Erwartungen. Der
Verlauf der Gesamtzuckerkonzentration (Ergebnisse nicht dargestellt) und auch die nach der
Ernte ausgewogene Zellmasse waren mit 8,2 kg Frischgewicht erwartungsgemäß. Für eine nicht
homogene Durchmischung des Reaktorinhalts während der letzten 48 h der Fermentation
sprechen auch die in diesem Zeitraum stark schwankenden Meßwerte der pH- und dO2-Sonden
(Abb. 3-8). Dieses Problem trat bei einigen Fermentation auf.
Die Analyse der Proben auf Expression des scFv4813 erfolgte durch Western-Blot mit Hilfe
eines monoklonalen Antikörpers mit Spezifität für das KDEL-Epitop (2.1.4). Zum Zeitpunkt der
Analyse dieser Proben waren weder ein gereinigter, eingestellter Standard des scFv4813, noch
humanes FSH zum Beschichten von ELISA-Platten oder Immobilisieren an einem BIACore
Chip (analog zu der Bestimmung des biscFv2429, 2.3.8) verfügbar, so daß diese Methode die
einzig mögliche zum Nachweis des scFv4813 war. Anstelle einer Quantifizierung konnte daher
lediglich eine Abschätzung der Konzentration erfolgen. Das Ergebnis ist in Abb. 3-9 dargestellt:

Ergebnisse und Diskussion
57
Abb. 3-9: Western Blot (2.3.5) von Gesamtproteinextrakten (2.3.1) aus Proben der Fermentation(2.4.4) der Zellinie R1-25 von Nicotiana tabacum cv BY-2. Die Proben wurden zu den überjeder Spur angegebenen Zeiten (in h) gezogen. 30 µl des Gesamtproteinextrakts wurden für jedeSpur aufgetragen. Die Detektion des scFv4813 (Pfeil) erfolgte über den M-α-KDEL Antikörper(100 ng·ml-1, 1 h, RT) und das GAM-AP Serum (120 ng·ml-1, 1h, RT) (2.1.4). Die Färbereaktionmit NBT-BCIP wurde nach 5 min durch Waschen in Wasser gestoppt.
Anhand der Stärke der Banden wurde die Konzentration des scFv4813 im
Gesamtproteinextrakt auf ca. 15 µg·ml-1 geschätzt. Die gesamte Zellmasse der Fermentation
hätte nach 2.3.2 ca. 22 Liter Proteinextrakt ergeben, also ca. 330 mg scFv4813, woraus sich eine
auf das Fermentationsvolumen bezogene Produktivität im Bereich von 1,6 mg·l-1·d-1 ergibt.
Verglichen mit den für den biscFv4813 (siehe 3.1.3-3.1.4) erhaltenen Werten (2-3 mg·l-1)
wurden also mit dem (ocs)3mas-Promotor etwa um den Faktor 4-5 gesteigerte Ausbeuten erzielt.
Dennoch stehen auch für die Zellinie R1-25 der notwendige Zellaufschluß und die erforderliche
Reinigung des Produkts aus dem komplexen Gesamtproteinextrakt, der Gewinnung des
rekombinanten Produktes im Gramm-Maßstab hinderlich gegenüber.
3.3 Entwicklung eines Fermentationsprotokolls für Pichia pastoris
3.3.1 Wachstumskinetik und Medienoptimierung im Schüttelkolben
Die Untersuchung des Wachstums von Pichia pastoris im Schüttelkolben wurde
durchgeführt, um einen Überblick über die auf verschiedenen Kohlenstoffquellen erzielbaren
Wachstumsraten und Biomasseerträge zu gewinnen. Untersucht wurden die im "Pichia
expression manual" (Invitrogen, Carlsbad, CA, Version D) vorgeschlagenen Medien YPD
(2.5.1) und BMGY (2.5.2). Zusätzlich wurde eine vereinfachte Version des Vorkulturmediums
BMGY, nämlich YPG (2.5.2) getestet, um die Frage zu beantworten, ob das teurere und
aufwendiger herzustellende BMGY ersetzt werden kann. Als Maß für das Wachstum wurde die

Ergebnisse und Diskussion
58
OD600 herangezogen (2.5.3), die im untersuchten Biomassebereich die höchste Genauigkeit
liefert. Die Mittelwerte aus jeweils zwei Messungen der beiden Ansätze eines jeden Mediums
wurden aufgetragen und mit Hilfe der Software Microcal Origin 5.0 ausgewertet (sigmoid fit).
Die Ergebnisse sind in Abb. 3-10 dargestellt.
0 4 8 12 16 20 240
5
10
15
20
25
30
BMGY Sigm. Fit BMGY YPD Sigm. Fit YPD YPG Sigm. Fit YPG
OD
600
Zeit [h]
Abb. 3-10: Wachstum von Pichia pastoris Klon N55 in verschiedenen Medien (2.5.1, 2.5.2) imSchüttelkolben. Die Kolben wurden zum Zeitpunkt t=0 jeweils 1%ig aus der gleichen Vorkultur(in BMGY) angeimpft. Zu verschiedenen Zeitpunkten wurden Proben gezogen und dasWachstum anhand der Bestimmung der Optischen Dichte (2.5.3) verfolgt.
Die Parameter der sigmoidalen Wachstumskurven (Tabelle 3-1) zeigen, daß die drei
verglichenen Medien hinsichtlich des Wachstumsverhaltens von Pichia gleichwertig sind. Die
Ergebnisse geben keine Auskunft darüber, ob die ermittelte maximale Wachstumsrate von 0,3 h-1
durch endogene Parameter (z. B. Replikationsgeschwindigkeit) oder exogene Parameter (z. B.
Sauerstoffeintrag im Schüttelkolben) bestimmt ist.
Tabelle 3-1: Wachstum des Pichia pastoris Klons N55 (2.1.7.2) im Schüttelkolben mitverschiedenen Medien:
Medium µmax
[h-1]OD600max FWmax
[g·l-1]Wendepunkt
[h]BMGY 0,31 27,2 61 13,6
YPD 0,30 28,9 65 13,4
YPG 0,31 28,3 63 13,6

Ergebnisse und Diskussion
59
3.3.2 Bestimmung des Methanolverwertungs-Phänotpys des Expressionsklons
Die Bestimmung des "Methanol utilization"-Phänotyps ist zur Charakterisierung eines Pichia
Expressionstammes unerläßlich, da dieser Einfluß auf das Wachstumsverhalten während der
Induktion hat. Abgesehen davon gibt es möglicherweise einen Einfluß auf die Expressionsraten
rekombinanter Proteine (Clare et al., 1991a; ; Romanos, 1995; Higgins und Cregg, 1998). Die im
Rahmen dieser Arbeit durchgeführte Bestimmung beruht auf dem Nachweis des AOX1-Gens mit
Hilfe spezifischer Primer für die 5' und 3' untranslatierten Regionen (2.2.1). Dieses Verfahren ist
schneller und zuverlässiger als die Bestimmung über die Wachstumsgeschwindigkeit von
Kolonien auf Nährböden mit Methanol als Kohlenstoffquelle.
Im Falle der Integration der zirkularisierten Expressionskassette über einen einzelnen
Cross-Over Vorgang in den 5' oder 3' AOX1-Regionen oder im HIS4-Gen (diese Sequenzen sind
in der Expressionskassette enthalten) wird die Methanolverwertungskapazität der Zelle nicht
beeinträchtigt. Die 5' und 3' Primer amplifizieren zwei DNA-Fragmente: Das native AOX1-Gen
und die Expressionskassette, die die gleichen flankierenden Regionen aufweist.
Wird das native AOX1-Gen durch die Expressionskassette infolge eines doppelten
Cross-Over Vorgangs an den flankierenden 5' und 3' Regionen ersetzt, so läßt sich durch die
entsprechenden Primer nur noch eine Bande in der Größe der Expressionskassette amplifizieren.
Die Bestimmung der Methanolverwertungsphänotypen ergab für den Pichia-Klon N55 das in
Abb. 3-11 gezeigte Resultat:
Abb. 3-11: Bestimmung des Methanolverwertungs-Phänotyps des Pichia Expressionsklons N55durch PCR (2.2.1). Die in der PCR mit spezifischen Primern für die Expressionskassette desscFv4813 und das native AOX1-Gen amplifizierten Banden wurden im Agarosegel (2.2.2)analysiert. Die Pfeile zeigen die Banden an, die dem intakten AOX1-Gen (2,3 kB), bzw. derscFv4813 Expressionskassette (1,3 kB) größenmäßig entsprechen.
Die Bande der von den 5'AOX und 3'AOX-Sequenzen flankierten Expressionskassette des
scFv4813 war deutlich erkennbar und wies die berechnete Größe von 1,3 kB auf. Die Bande für
das intakte AOX1-Gen war schwächer, was darauf zurückzuführen sein könnte, daß der
Expressorklon N55 im G418-Resistenztest über die Toleranz gegen erhöhte Konzentrationen des
Antibiotikums G418 (s. Einleitung) als Träger mehrerer Kopien der Expressionskassette
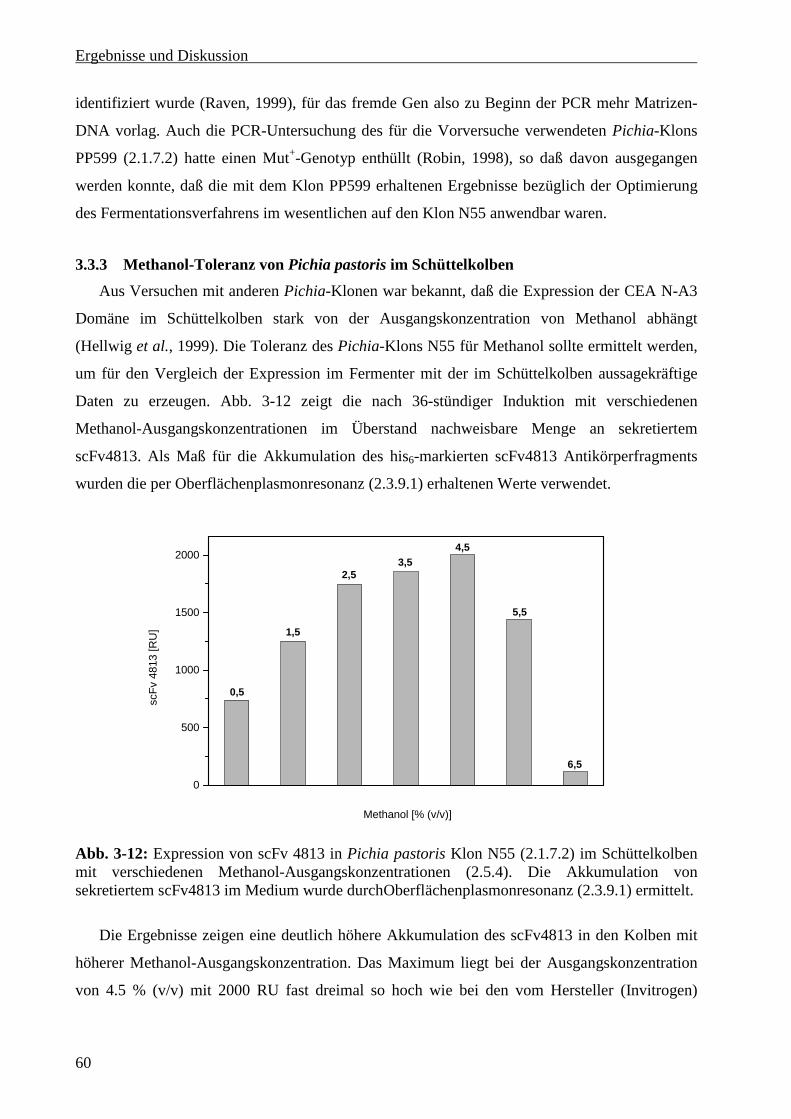
Ergebnisse und Diskussion
60
identifiziert wurde (Raven, 1999), für das fremde Gen also zu Beginn der PCR mehr Matrizen-
DNA vorlag. Auch die PCR-Untersuchung des für die Vorversuche verwendeten Pichia-Klons
PP599 (2.1.7.2) hatte einen Mut+-Genotyp enthüllt (Robin, 1998), so daß davon ausgegangen
werden konnte, daß die mit dem Klon PP599 erhaltenen Ergebnisse bezüglich der Optimierung
des Fermentationsverfahrens im wesentlichen auf den Klon N55 anwendbar waren.
3.3.3 Methanol-Toleranz von Pichia pastoris im Schüttelkolben
Aus Versuchen mit anderen Pichia-Klonen war bekannt, daß die Expression der CEA N-A3
Domäne im Schüttelkolben stark von der Ausgangskonzentration von Methanol abhängt
(Hellwig et al., 1999). Die Toleranz des Pichia-Klons N55 für Methanol sollte ermittelt werden,
um für den Vergleich der Expression im Fermenter mit der im Schüttelkolben aussagekräftige
Daten zu erzeugen. Abb. 3-12 zeigt die nach 36-stündiger Induktion mit verschiedenen
Methanol-Ausgangskonzentrationen im Überstand nachweisbare Menge an sekretiertem
scFv4813. Als Maß für die Akkumulation des his6-markierten scFv4813 Antikörperfragments
wurden die per Oberflächenplasmonresonanz (2.3.9.1) erhaltenen Werte verwendet.
0
500
1000
1500
2000
6,5
5,5
4,5
3,52,5
1,5
0,5
Methanol [% (v/v)]
scF
v 48
13 [R
U]
Abb. 3-12: Expression von scFv 4813 in Pichia pastoris Klon N55 (2.1.7.2) im Schüttelkolbenmit verschiedenen Methanol-Ausgangskonzentrationen (2.5.4). Die Akkumulation vonsekretiertem scFv4813 im Medium wurde durchOberflächenplasmonresonanz (2.3.9.1) ermittelt.
Die Ergebnisse zeigen eine deutlich höhere Akkumulation des scFv4813 in den Kolben mit
höherer Methanol-Ausgangskonzentration. Das Maximum liegt bei der Ausgangskonzentration
von 4.5 % (v/v) mit 2000 RU fast dreimal so hoch wie bei den vom Hersteller (Invitrogen)

Ergebnisse und Diskussion
61
verwendeten Ausgangskonzentrationen von 0.5 % (v/v). Dieses Ergebnis wurde durch die
Analyse der Proben durch SDS-PAGE und Coomassie Färbung bestätigt (Daten nicht gezeigt).
Die Tatsache, daß Pichia pastoris im Schüttelkolben Methanolkonzentrationen bis zu
4,5 % (v/v) toleriert, ist überraschend, da in der Literatur die Toxizitätsgrenze von Methanol bei
methylotrophen Hefen bei 0,5 bis 1 % (v/v) angesiedelt wird. Der Grund für diesen Effekt
könnte darin liegen, daß die Sauerstoffversorgung unter den gewählten Bedingungen (2.5.4)
unzureichend ist: Die Zelldichte liegt zu Beginn der Induktion bei ca. 250 g·l-1, so daß die
Gelöstsauerstoffkonzentration nahe 0 liegen dürfte. Möglicherweise erfolgt unter diesen
Bedingungen die Bildung des toxischen Formaldehyds im ersten Schritt der
Methanolverwertungskette so langsam, daß ein Effekt auf den Zellmetabolismus erst bei sehr
hohen Methanolkonzentrationen auftritt.
Dieses Ergebnis ist insofern relevant für die Fermentation von Pichia, als daß zumindest
unter sauerstofflimitierten Bedingungen von einer höheren Toleranz als 0,5 % (v/v) für Methanol
ausgegangen werden kann, was vereinfachte Zufütterungsstrategien und Zulaufgeometrien
erlaubt.
3.3.4 Wachstumskinetik im Fermenter und Optimierung der Fermentationsstrategie
Die "Standard"-Fermentationsstrategie für Pichia pastoris entsprechend dem Handbuch der
Fa. Invitrogen (Pichia Expression Manual, Invitrogen, Carlsbad, CA, Version D) ist in Abb. 3-13
anhand einer Fermentation des Klons PP599 demonstriert. Dieser Klon wurde verwendet, um die
Fermentationsstrategie von Pichia pastoris zu etablieren und zu optimieren, bevor ein
Expressorklon für den scFv4813 identifiziert war.

Ergebnisse und Diskussion
62
0 12 24 36 48 60 72 84 960
50
100
150
200
250
300
Fris
chge
wic
ht [g
·L-1
]
Zeit [h]
0
5
10
15
20
25
30
35
40
45
50
55
60
321
Gly
zerin
/Met
hano
l Zuf
uhrr
ate
[mL·
h-1]
Abb. 3-13: Fermentation von Pichia pastoris Klon PP599 im 5-l Maßstab nach demStandardprotokoll (2.5.5). Die Zufuhrraten von Glyzerin (gestrichelte Linie) bzw. Methanol(punktierte Linie) wurden manuell gewählt. Das Frischgewicht (!) wurde gravimetrischbestimmt (2.5.3). Die Pfeile geben die Dauer der Anwachsphase auf Glyzerin (1), derZwischenphase im Glyzerin-Zulaufverfahren (2) bzw. der Induktionsphase im Methanol-Zulaufverfahren (3) dar.
Die drei Abschnitte der Fermentation sind deutlich erkennbar. 4 l Fermentationsmedium "A"
(2.5.5.1) wurden mit 4 % (v/v) einer Vorkultur (2.5.2) des Klons PP599 beimpft. Innerhalb von
31 h wurde das gesamte eingesetzte Glyzerin verbraucht und ein Frischgewicht von 127 g·l-1
erreicht. Der zweite Abschnitt der Fermentation war durch eine 5 Stunden andauernde Glyzerin-
Zulaufphase definiert. Die Zufuhrrate der 50%igen Glyzerinlösung betrug 37 ml·h-1 und wurde
manuell gewählt. In dieser Phase erhöhte sich das Frischgewicht auf 214 g·l-1. In der dritten
Phase der Fermentation wurde die Expression des heterologen Proteins durch Zufuhr von
Methanol als einziger Kohlenstoffquelle bei einer manuell vorgegebenen Rate von 18 ml·h-1,
entsprechend ca. 4 ml·l-1·h-1 über einen Zeitraum von 46 h induziert. Das Wachstum war auf
Methanol deutlich langsamer, so daß das Frischgewicht während der Induktion plateauartig bei
ca. 225 g·l-1 verlief.
Das Standardverfahren wurde in mehreren Schritten hinsichtlich der Inokulumgröße, der
Salz- und Glyzerinkonzentrationen im Medium und der Fermentationsstrategie optimiert.
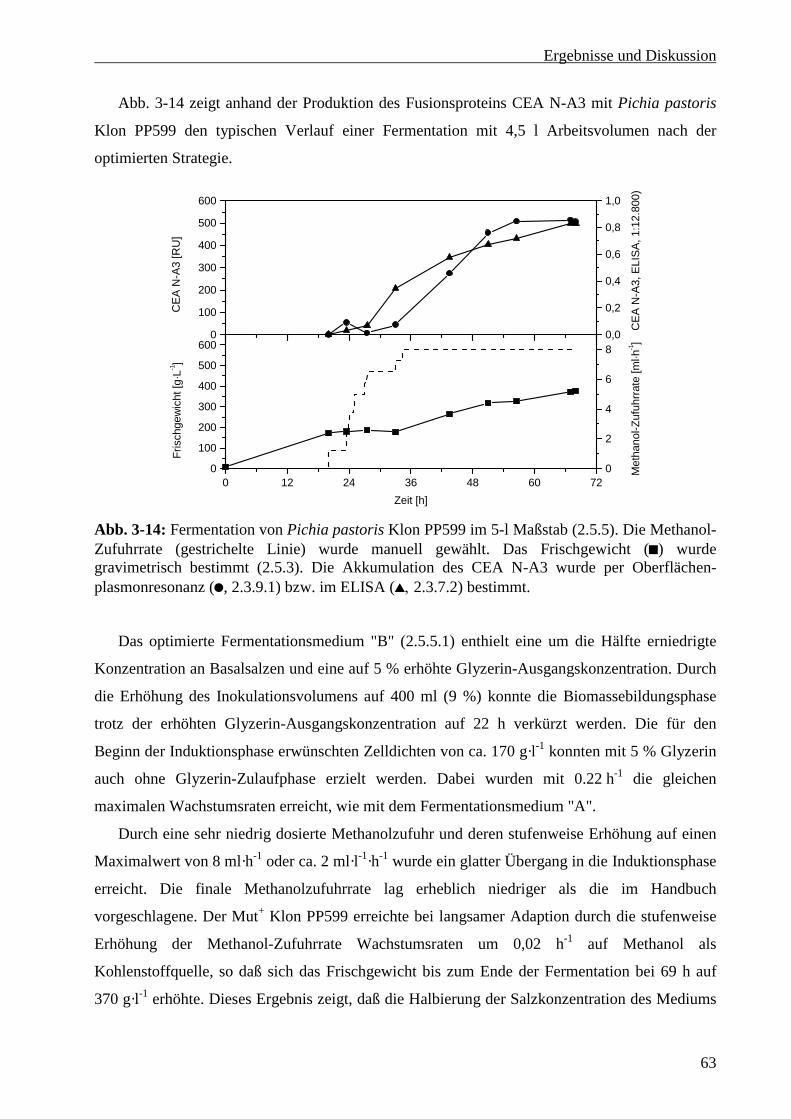
Ergebnisse und Diskussion
63
Abb. 3-14 zeigt anhand der Produktion des Fusionsproteins CEA N-A3 mit Pichia pastoris
Klon PP599 den typischen Verlauf einer Fermentation mit 4,5 l Arbeitsvolumen nach der
optimierten Strategie.
0 12 24 36 48 60 720
100
200
300
400
500
600
Fris
chge
wic
ht [g
·L-1]
Zeit [h]
0
100
200
300
400
500
600
CE
A N
-A3
[RU
]
0
2
4
6
8
Met
hano
l-Zuf
uhrr
ate
[ml·h
-1]
0,0
0,2
0,4
0,6
0,8
1,0
CE
A N
-A3,
ELI
SA
, 1:1
2.80
0)
Abb. 3-14: Fermentation von Pichia pastoris Klon PP599 im 5-l Maßstab (2.5.5). Die Methanol-Zufuhrrate (gestrichelte Linie) wurde manuell gewählt. Das Frischgewicht (!) wurdegravimetrisch bestimmt (2.5.3). Die Akkumulation des CEA N-A3 wurde per Oberflächen-plasmonresonanz (", 2.3.9.1) bzw. im ELISA (#()2.3.7.2) bestimmt.
Das optimierte Fermentationsmedium "B" (2.5.5.1) enthielt eine um die Hälfte erniedrigte
Konzentration an Basalsalzen und eine auf 5 % erhöhte Glyzerin-Ausgangskonzentration. Durch
die Erhöhung des Inokulationsvolumens auf 400 ml (9 %) konnte die Biomassebildungsphase
trotz der erhöhten Glyzerin-Ausgangskonzentration auf 22 h verkürzt werden. Die für den
Beginn der Induktionsphase erwünschten Zelldichten von ca. 170 g·l-1 konnten mit 5 % Glyzerin
auch ohne Glyzerin-Zulaufphase erzielt werden. Dabei wurden mit 0.22 h-1 die gleichen
maximalen Wachstumsraten erreicht, wie mit dem Fermentationsmedium "A".
Durch eine sehr niedrig dosierte Methanolzufuhr und deren stufenweise Erhöhung auf einen
Maximalwert von 8 ml·h-1 oder ca. 2 ml·l-1·h-1 wurde ein glatter Übergang in die Induktionsphase
erreicht. Die finale Methanolzufuhrrate lag erheblich niedriger als die im Handbuch
vorgeschlagene. Der Mut+ Klon PP599 erreichte bei langsamer Adaption durch die stufenweise
Erhöhung der Methanol-Zufuhrrate Wachstumsraten um 0,02 h-1 auf Methanol als
Kohlenstoffquelle, so daß sich das Frischgewicht bis zum Ende der Fermentation bei 69 h auf
370 g·l-1 erhöhte. Dieses Ergebnis zeigt, daß die Halbierung der Salzkonzentration des Mediums

Ergebnisse und Diskussion
64
sich selbst im extremen Hochzelldichtebereich nicht negativ auf die Wachstumsrate oder die
erreichbare Biomassekonzentration auswirkt.
Bei Verwendung des Fermentationsmediums "A" waren beim Einstellen des pH-Werts auf
neutral-basische Werte zu Beginn der Aufarbeitung stets größere Mengen Salze ausgefallen.
Dieser Effekt unterblieb mit dem salzreduzierten Fermentationsmedium "B".
Die Akkumulation des sekretierten CEA N-A3 im Überstand wurde mit Hilfe von der
Oberflächenplasmonresonanz (2.3.9.1) und des ELISA (2.3.7) bestimmt. Der Vergleich der
Werte zeigt, daß beide Methoden im wesentlichen gleiche, relative Ergebnisse bringen (Abb.
3-14). Dies bedeutet, daß die schnellere und wegen der hohen Kosten der für den ELISA
benötigten Antigene und sekundären Antikörper billigere Methode der Bestimmung über die
Oberflächenplasmonresonanz für die Standardanalytik einer Fermentationen verwendet werden
kann. Die Abschätzung der Konzentrationen war schwierig, da das vom Klon PP599 sekretierte
Fusionsprotein in Pichia stark und heterogen glykosyliert ist, und im PAA-Gel über den Bereich
von 45 bis 200 kDa "schmiert" (Hellwig et al., 1999).
Die mit Hilfe dieses Klons entwickelte Fermentationsstrategie in Bezug auf Inokulumgröße,
Medium und Adaption an Methanol im 5-l Maßstab konnte erfolgreich auf die Fermentation des
Klons N55 übertragen werden (3.4.1).
3.3.5 Maßstabsvergrößerung in den 30-Liter Maßstab
Die Maßstabsvergrößerung in den 30-l Maßstab erfolgte aufbauend auf den anhand des
Pichia-Klons PP599 gewonnenen Ergebnissen mit dem Klon N55. Die ersten Versuche wurden
im Biopilot 40 Reaktor ohne eingebaute Stromstörer gemacht (Abb. 2-1). In den Proben der
ersten Fermentationen ließ sich der scFv4813 weder über Oberflächenplasmonresonanz noch
über den ELISA nachweisen (Daten nicht gezeigt). Auch im Coomassie-gefärbten SDS-PAA-
Gel ließ sich kein scFv Fragment nachweisen, obwohl die Produktion im 5-l Reaktor erfolgreich
verlaufen war (A in Abb. 3-15). Wachstum und Sauerstoff-, Ammonium- sowie
Methanolverbrauch ließen keine direkten Schlüsse auf den Grund dafür zu (Daten nicht gezeigt).
Auch Zellen, die unter sterilen Bedingungen während der Induktionsphase aus dem 30-l
Fermenter geerntet und im Schüttelkolben mit BMMY-Medium induziert wurden, zeigten
normale Expression des scFv (Daten nicht gezeigt). Eine Fermentation des Klons 3299, der ein
anderes scFv Fragment sehr stark exprimiert (bis zu 200 mg·l-1 im Fermentationsüberstand),
zeigte deutliche Degradation dieses Fragments im 30-l Maßstab (B in Abb. 3-15).
Aus diesen beiden Befunden ließ sich folgern, daß es sich bei den enttäuschenden
Expressionsergebnissen im 30-l Reaktor nicht um ein Expressions- sondern um ein

Ergebnisse und Diskussion
65
Degradationsproblem handelte, daß durch chemisch-physikalische Faktoren der Kultivierung im
30-l Reaktor ausgelöst wurde.
Abb. 3-15: Coomassie-gefärbtes SDS PAA-Gel (2.3.5) von Proben aus verschiedenenFermentationen von Pichia pastoris im 30-l Reaktor vor dem Einbau von Stromstörern in denReaktor. In allen Fällen wurden 30 µl Überstand geladen. A: Klon N55 (2.1.7.2). 1-2:Fermentationen mit 30 l Arbeitsvolumen 3-5: Fermentation mit 18 l Arbeitsvolumen 6:Positivkontrolle aus dem 5-l Reaktor. B: Klon 3299 (2.1.7.2), 30 l Arbeitsvolumen. Die Pfeilezeigen die Banden des scFv4813 an (A, Spur 6), bzw. des scFv3299 (B) und dessenDegradationsprodukte.
In der Tat traten die Probleme nicht mehr auf, nachdem der 30-l Reaktor mit vier
Stromstörern nachgerüstet worden war. Die durch die Schikanen verbesserte Durchmischung hat
Einfluß auf zahlreiche Prozeßfaktoren: In schlecht durchmischten Zonen können Sauerstoff-
oder Nährstoffmangel oder lokale pH-Gradienten auftreten. Möglicherweise kommt es durch
einen solchen Faktor auch zur Expression von Proteasen. Die Fragen welcher dieser Faktoren die
Degradationsprobleme verursacht hatte und ob es sich um einen chemisch-physikalischen Faktor
oder um eine dadurch induzierte Expression extrazellulärer Proteasen handelte, wurden nicht
weiter verfolgt. SDS-PAA Gele, die die erfolgreiche Expression des scFv4813 im 30-l Reaktor
zeigen sind in Abb. 3-17 und Abb. 3-25 gezeigt.
3.3.6 Optimierung der Methanol-Zufuhrsteuerung
Die Standard-Ausstattung von Rührkesselreaktoren umfaßt üblicherweise Echtzeiterfassung
und -regelung von Temperatur, pH-Wert, Gelöstsauerstoff (dO2) und Füllhöhe, nicht aber eine
Methanol-Analytik. Aufgrund der Toxizität von Methanol ist es aber notwendig, bei dessen
Verwendung als Kohlenstoffquelle unterhalb einer bestimmten Grenze zu bleiben, die unter
anderem von der Sauerstoffverfügbarkeit abhängt. Auch wenn gezeigt werden konnte, daß
zumindest im Schüttelkolben die Methanoltoleranz erstaunlich hoch war, war eine Steuerung der

Ergebnisse und Diskussion
66
Methanolkonzentration in einem engeren Bereich wünschenswert. Entsprechende Versuche zur
Optimierung der Zufuhrsteuerung wurden sowohl im 5-l als auch im 30-l Reaktor durchgeführt.
3.3.6.1 Messung der Methanolkonzentration bei manueller Steuerung der Zufuhrrate
Zur Verdeutlichung der Problematik der Fermentationsführung mit manuell gesteuerter
Methanolzufuhr (entsprechend der in 3.3.4 geschilderten Strategie) wurde die
Methanolkonzentration im Verlauf einer solchen Fermentation gemessen und aufgezeichnet.
Dies geschah, nachdem die Technologie zur Methanolmessung in Echtzeit bereits etabliert war,
um Klarheit zu erhalten, welche Methanolkonzentrationen im Verlauf einer manuell gesteuerten
Fermentation überhaupt auftreten. Das Ergebnis ist in Abb. 3-16 dargestellt.
00 24 48 72 96 1200
100
200
300
400
Fris
chge
wic
ht [g
·L-1]
scF
v481
3 [R
U]
Zeit [h]
0,0
0,5
1,0
1,5
2,0
2,5
B
A
21
Met
hano
l-Zuf
uhrr
ate
[ml·l
-1·h
-1]
0,0
0,2
0,4
0,6
0,8
1,0
Met
hano
l [%
(v/
v)]
0
20
40
60
80
100
120
140
dO2
[%]
Abb. 3-16: Fermentation von Pichia pastoris Klon N55 im 30-l Maßstab mit Echtzeit Methanol-Analytik (2.5.5). Die Messung der Methanolkonzentration (A, durchgezogene Linie) erfolgte mitHilfe des Frings ALKOSENS II (2.5.5.4). Die Steuerung der Methanol-Zufuhrrate (A,gestrichelte Linie) erfolgte manuell aufgrund von Erfahrungswerten und anhand des Verlaufs derGelöstsauerstoffkonzentration. Die Gelöstsauerstoffkonzentration (B, durchgezogene Linie)wurde in Echtzeit erfaßt, Frischgewicht (!) gravimetrisch (2.5.3) und Akkumulation vonscFv 4813 (") per Oberflächenplasmonresonanz (2.3.9.1) aus den Proben. Die Pfeile geben dieDauer der Biomassebildungsphase (1) bzw. der Induktionsphase (2) an.
Nach der Anwachsphase von 35 h wurde bei einem Frischgewicht von 185 g·l-1 durch Zufuhr
von Methanol bei einer konstanten Rate von 1 ml·l-1·h-1 die Induktionsphase eingeleitet. Da die
Zellen zu diesem Zeitpunkt noch an Glyzerin adaptiert waren, akkumulierte Methanol. Mit
zunehmender Adaption der Zellen innerhalb der nächsten Stunden stieg der spezifische

Ergebnisse und Diskussion
67
Methanolverbrauch. Nach 42 h erreichte die Methanol-Verbrauchsrate den Betrag der Methanol-
Zufuhrrate und stieg darüber an, wodurch die Methanolkonzentration im Fermenter wieder
absank. Die Erschöpfung des während der Adaptionsphase akkumulierten Methanols wurde nach
59 h durch ein plötzliches Ansteigen der Gelöstsauerstoffkonzentration ("dO2-spike") angezeigt.
Dies war das Signal, die Methanolzufuhrrate auf 2 ml·l-1·h-1 zu erhöhen. Nach 4,5 h bei
2ml·l-1·h-1 wurde die Methanolzufuhr kurz gestoppt. Die Zeit bis zum Auftreten eines erneuten
dO2-spikes lag über 1 min, was darauf hinwies, daß die Methanol-Verbrauchsrate unter 2
ml·l-1·h-1 lag. Die Zufuhrrate wurde auf 1,2 ml·l-1·h-1 abgesenkt. Im weiteren Verlauf der
Induktionsphase wurde noch mehrfach die Zeit bestimmt, die nach dem Stop der Substratzufuhr
bis zum Auftreten des dO2-spikes vergeht (bei 87/89 h bzw. 107/111 h). Dieses Verfahren stellt
die einzige Möglichkeit dar, ohne Echtzeit-Methanolmessung die Methanolzufuhrrate auf den
Verbrauch abzustimmen.
Der Verlauf der Methanolkonzentration zeigt, daß bei diesem Verfahren die
Methanolkonzentration während der Induktionsphase zwischen 0 und 0,4 % (v/v) schwankte.
Das Frischgewicht nahm während der Induktionsphase nur von 182 auf 243 g·l-1 zu. Die
Konzentration von scFv4813 im Überstand nahm in der zweiten Hälfte der Induktionsphase
kaum noch zu, ein Befund der durch SDS-PAGE und Western Blot (Abb. 3-17) bestätigt wird.
Abb. 3-17: Analyse der Akkumulation von sekretiertem scFv4813 im Überstand der in Abb.3-16 gezeigten Fermentation des Klons N55. 30 µl zellfreier Überstand wurden durch SDS-PAGE (2.3.5) aufgetrennt und durch Coomassie-Färbung (2.3.5.2) nachgewiesen (links). Für denWestern Blot (2.3.6) der gleichen Proben wurden jeweils 10 µl aufgetrennt und nach demBlotten mit Hilfe des monoklonalen M-α-his6 (2.1.4) und des sekundären Antikörpers GAM-AP(2.1.4) detektiert (rechts). Die Pfeile zeigen die Banden an, deren Größe mit der für scFv4813berechneten Bande übereinstimmt. Die Proben wurden zu den über den jeweiligen Spurenstehenden Zeitpunkten gezogen. Pro Spur wurden 30 µl (Coomassie-Färbung) bzw. 10 µl(Western Blot) des zellfreien Überstandes aufgetragen.

Ergebnisse und Diskussion
68
Die Tatsache, daß im Western Blot außer der Bande des scFv4813 keine weiteren Banden
durch den anti-his6-Antikörper detektiert werden zeigt, daß keine proteolytischen Abbauprodukte
des scFv4813 im Medium vorliegen, ansonsten würden C-terminale kleinere Proteinbanden
detektiert werden. Außerdem weist dieses Ergebnis darauf hin, daß die schnelle Bestimmung der
im Überstand akkumulierten scFvs über den His6-tag mit der Oberflächenplasmonresonanz
(2.3.9.1) wahrscheinlich nicht durch andere Proteine im Überstand gestört wird bzw. es sich bei
der Gesamtheit des auf dem NTA-Chip gebundenen Proteins tatsächlich um scFv4813 handelt.
Insgesamt zeigt diese Fermentation, daß die manuelle Steuerung der Methanol-Zuflußrate
zwar möglich ist, aber zu stark schwankenden Methanolkonzentrationen führt. Eine solche
Fermentation ist nicht reproduzierbar, da eine Vielzahl von Parametern wie z. B. Frischgewicht,
Volumen, Belüftungsrate, Abluftkühlung etc. auf den Methanolverbrauch Auswirkungen haben,
der sich zudem aus der Summe von Methanol-Stoffwechsel und Methanolverlust über die Abluft
zusammensetzt. Die Methanolkonzentration stellt aber wegen der doppelten Funktion des
Methanols als einzige Kohlenstoffquelle und als Induktor einen wesentlichen Prozessparameter
dar. Zudem war dieses Ergebnis nur auf der Basis der aus vorangegangenen Fermentationen
gewonnenen Daten bezüglich des Methanolverbrauchs erzielbar. Beim Start der Methanolzufuhr
mit einer Rate von 4 ml·l-1·h-1, wie z. B. bei der in Abb. 3-13 gezeigten Fermentation wären bei
gleicher Dauer der Adaptionsphase wesentlich höhere Methanolkonzentrationen akkumuliert
worden.
3.3.6.2 Steuerung der Methanolzufuhr über die Gelöstsauerstoffkonzentration
Eine erste Verbesserung der Steuerung der Methanol-Zufuhrrate wurde erreicht, indem die
Methanol-Pumpe in Abhängigkeit von der Gelöstsauerstoffkonzentration durchgeführt wurde (B
in Abb. 2-2). Diese Strategie wurde in der in Abb. 3-18 gezeigten Fermentation im 5-l Maßstab
ab 53 h verfolgt (4), nachdem zuvor die Biomassebildungsphase auf Glyzerin (1), eine langsame
Adaption an Methanol (2) und eine Phase mit manuell gesteuerter Methanolzufuhr (3)
durchgeführt worden waren. Bei Überschreitung einer Gelöstsauerstoffkonzentration von 20 %
wurde die Methanol-Pumpe aktiviert. Die Stellgröße dieses Regelkreises wurde alle 15 s neu
berechnet. Dies entspricht der Zeit, die die Meßgröße dO2 unter den gegebenen Bedingungen bei
Erschöpfung der Kohlenstoffquelle benötigt, um ca. 20 % anzusteigen bzw. um bei erneuter
Dosierung von Methanol um diesen Wert abzufallen. Die Pumpintervalle waren so gewählt, daß
innerhalb von zwei Regelkreiszeiten maximal 0,05 % (v/v) Methanol gefördert werden konnten.
Die anhand der Einschaltzeiten der Methanol-Pumpe berechnete Förderrate lag in den ersten
18 h mit relativ konstanten 4,2 ml·min-1 nahe bei dem Wert, der sich in den manuell gesteuerten
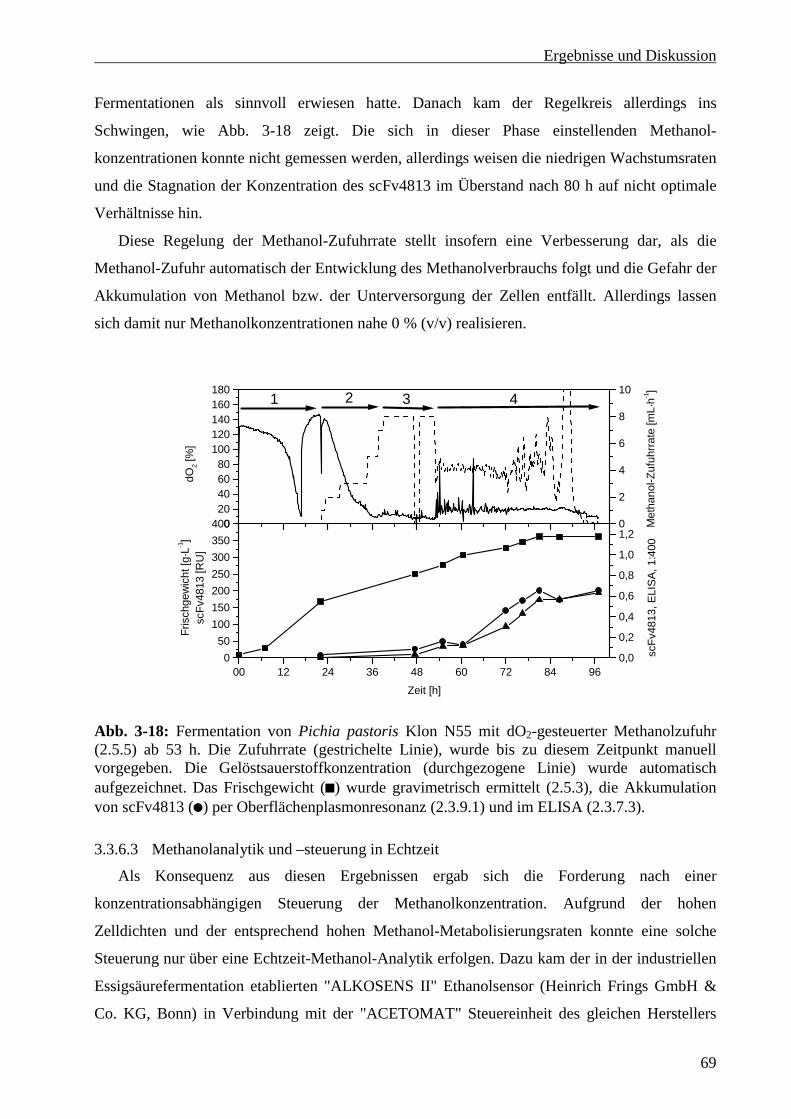
Ergebnisse und Diskussion
69
Fermentationen als sinnvoll erwiesen hatte. Danach kam der Regelkreis allerdings ins
Schwingen, wie Abb. 3-18 zeigt. Die sich in dieser Phase einstellenden Methanol-
konzentrationen konnte nicht gemessen werden, allerdings weisen die niedrigen Wachstumsraten
und die Stagnation der Konzentration des scFv4813 im Überstand nach 80 h auf nicht optimale
Verhältnisse hin.
Diese Regelung der Methanol-Zufuhrrate stellt insofern eine Verbesserung dar, als die
Methanol-Zufuhr automatisch der Entwicklung des Methanolverbrauchs folgt und die Gefahr der
Akkumulation von Methanol bzw. der Unterversorgung der Zellen entfällt. Allerdings lassen
sich damit nur Methanolkonzentrationen nahe 0 % (v/v) realisieren.
00 12 24 36 48 60 72 84 960
50
100
150
200
250
300
350
400
Fris
chge
wic
ht [g
·L-1]
scF
v481
3 [R
U]
Zeit [h]
020406080
100120140160180
3 41 2
dO2
[%]
0,0
0,2
0,4
0,6
0,8
1,0
1,2
scF
v481
3, E
LIS
A, 1
:400
0
2
4
6
8
10
Met
hano
l-Zuf
uhrr
ate
[mL·
h-1]
Abb. 3-18: Fermentation von Pichia pastoris Klon N55 mit dO2-gesteuerter Methanolzufuhr(2.5.5) ab 53 h. Die Zufuhrrate (gestrichelte Linie), wurde bis zu diesem Zeitpunkt manuellvorgegeben. Die Gelöstsauerstoffkonzentration (durchgezogene Linie) wurde automatischaufgezeichnet. Das Frischgewicht (!) wurde gravimetrisch ermittelt (2.5.3), die Akkumulationvon scFv4813 (") per Oberflächenplasmonresonanz (2.3.9.1) und im ELISA (2.3.7.3).
3.3.6.3 Methanolanalytik und –steuerung in Echtzeit
Als Konsequenz aus diesen Ergebnissen ergab sich die Forderung nach einer
konzentrationsabhängigen Steuerung der Methanolkonzentration. Aufgrund der hohen
Zelldichten und der entsprechend hohen Methanol-Metabolisierungsraten konnte eine solche
Steuerung nur über eine Echtzeit-Methanol-Analytik erfolgen. Dazu kam der in der industriellen
Essigsäurefermentation etablierten "ALKOSENS II" Ethanolsensor (Heinrich Frings GmbH &
Co. KG, Bonn) in Verbindung mit der "ACETOMAT" Steuereinheit des gleichen Herstellers
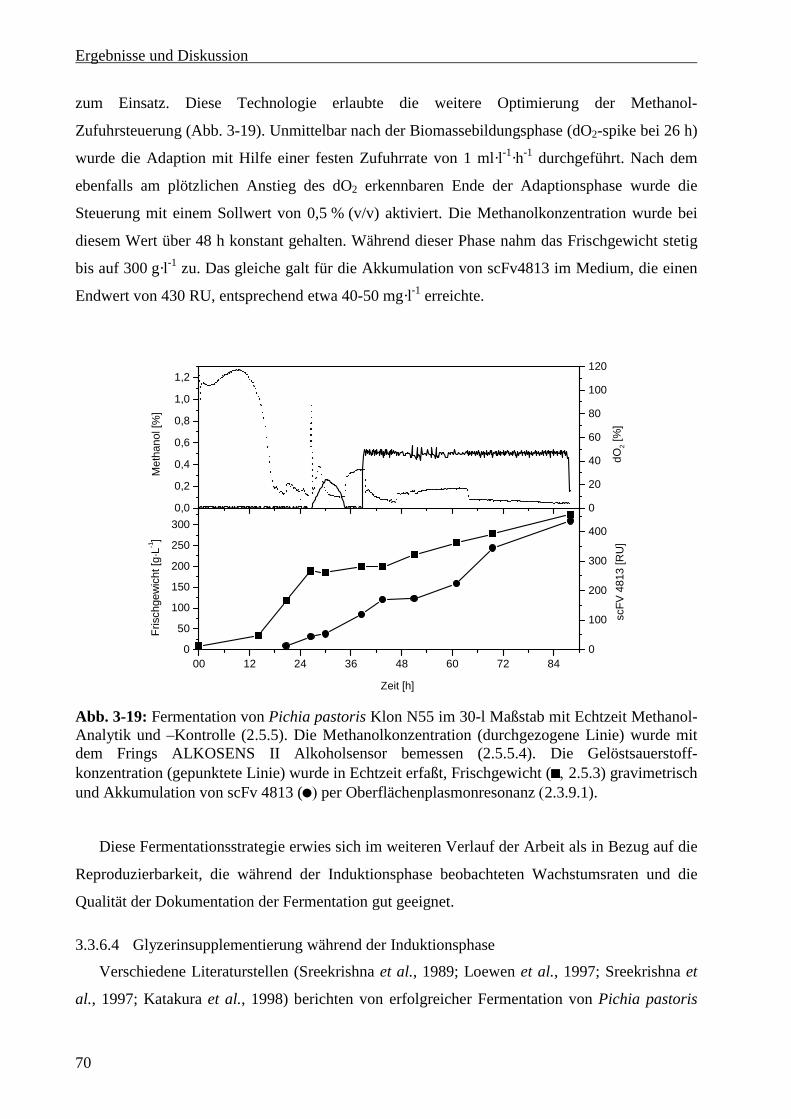
Ergebnisse und Diskussion
70
zum Einsatz. Diese Technologie erlaubte die weitere Optimierung der Methanol-
Zufuhrsteuerung (Abb. 3-19). Unmittelbar nach der Biomassebildungsphase (dO2-spike bei 26 h)
wurde die Adaption mit Hilfe einer festen Zufuhrrate von 1 ml·l-1·h-1 durchgeführt. Nach dem
ebenfalls am plötzlichen Anstieg des dO2 erkennbaren Ende der Adaptionsphase wurde die
Steuerung mit einem Sollwert von 0,5 % (v/v) aktiviert. Die Methanolkonzentration wurde bei
diesem Wert über 48 h konstant gehalten. Während dieser Phase nahm das Frischgewicht stetig
bis auf 300 g·l-1 zu. Das gleiche galt für die Akkumulation von scFv4813 im Medium, die einen
Endwert von 430 RU, entsprechend etwa 40-50 mg·l-1 erreichte.
00 12 24 36 48 60 72 840
50
100
150
200
250
300
Fris
chge
wic
ht [g
·L-1]
Zeit [h]
0,0
0,2
0,4
0,6
0,8
1,0
1,2
Met
hano
l [%
]
0
20
40
60
80
100
120
dO
2 [%
]
0
100
200
300
400
scF
V 4
813
[RU
]
Abb. 3-19: Fermentation von Pichia pastoris Klon N55 im 30-l Maßstab mit Echtzeit Methanol-Analytik und –Kontrolle (2.5.5). Die Methanolkonzentration (durchgezogene Linie) wurde mitdem Frings ALKOSENS II Alkoholsensor bemessen (2.5.5.4). Die Gelöstsauerstoff-konzentration (gepunktete Linie) wurde in Echtzeit erfaßt, Frischgewicht (!()2.5.3) gravimetrischund Akkumulation von scFv 4813 ("')per Oberflächenplasmonresonanz *2.3.9.1).
Diese Fermentationsstrategie erwies sich im weiteren Verlauf der Arbeit als in Bezug auf die
Reproduzierbarkeit, die während der Induktionsphase beobachteten Wachstumsraten und die
Qualität der Dokumentation der Fermentation gut geeignet.
3.3.6.4 Glyzerinsupplementierung während der Induktionsphase
Verschiedene Literaturstellen (Sreekrishna et al., 1989; Loewen et al., 1997; Sreekrishna et
al., 1997; Katakura et al., 1998) berichten von erfolgreicher Fermentation von Pichia pastoris
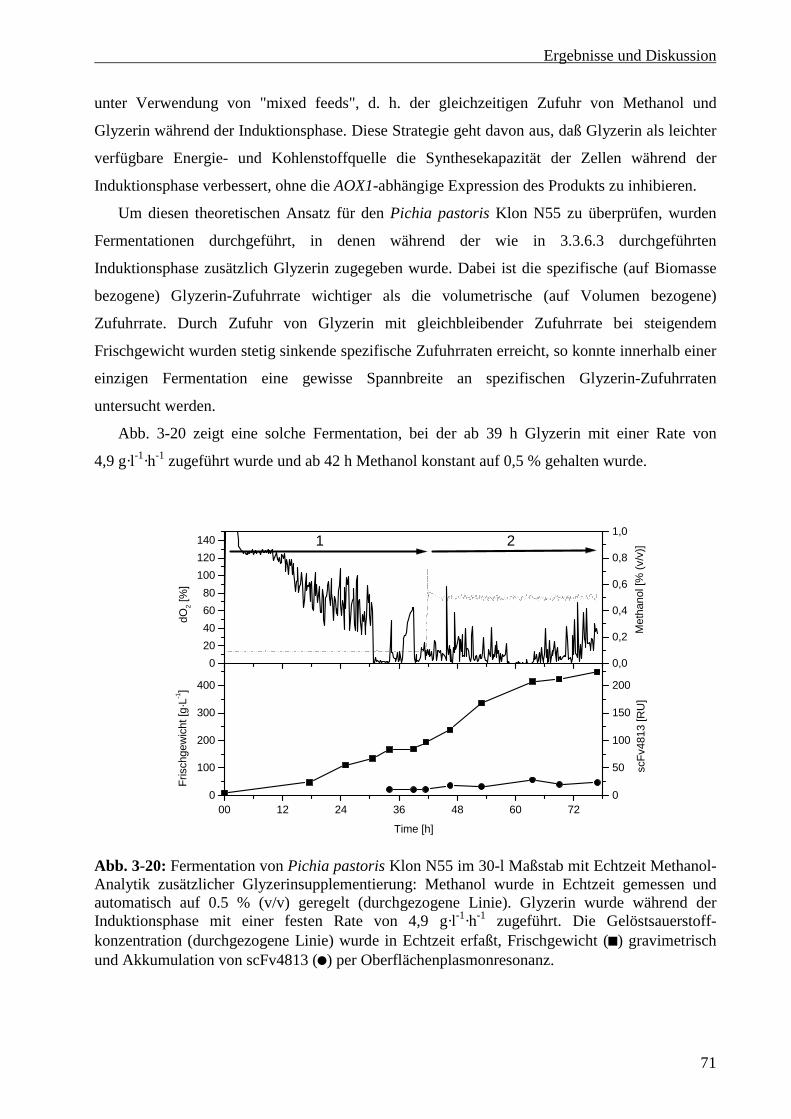
Ergebnisse und Diskussion
71
unter Verwendung von "mixed feeds", d. h. der gleichzeitigen Zufuhr von Methanol und
Glyzerin während der Induktionsphase. Diese Strategie geht davon aus, daß Glyzerin als leichter
verfügbare Energie- und Kohlenstoffquelle die Synthesekapazität der Zellen während der
Induktionsphase verbessert, ohne die AOX1-abhängige Expression des Produkts zu inhibieren.
Um diesen theoretischen Ansatz für den Pichia pastoris Klon N55 zu überprüfen, wurden
Fermentationen durchgeführt, in denen während der wie in 3.3.6.3 durchgeführten
Induktionsphase zusätzlich Glyzerin zugegeben wurde. Dabei ist die spezifische (auf Biomasse
bezogene) Glyzerin-Zufuhrrate wichtiger als die volumetrische (auf Volumen bezogene)
Zufuhrrate. Durch Zufuhr von Glyzerin mit gleichbleibender Zufuhrrate bei steigendem
Frischgewicht wurden stetig sinkende spezifische Zufuhrraten erreicht, so konnte innerhalb einer
einzigen Fermentation eine gewisse Spannbreite an spezifischen Glyzerin-Zufuhrraten
untersucht werden.
Abb. 3-20 zeigt eine solche Fermentation, bei der ab 39 h Glyzerin mit einer Rate von
4,9 g·l-1·h-1 zugeführt wurde und ab 42 h Methanol konstant auf 0,5 % gehalten wurde.
00 12 24 36 48 60 720
100
200
300
400
Fris
chge
wic
ht [g
·L-1]
Time [h]
0
20
40
60
80
100
120
140 21
dO2
[%]
0
50
100
150
200
scF
v481
3 [R
U]
0,0
0,2
0,4
0,6
0,8
1,0
Met
hano
l [%
(v/
v)]
Abb. 3-20: Fermentation von Pichia pastoris Klon N55 im 30-l Maßstab mit Echtzeit Methanol-Analytik zusätzlicher Glyzerinsupplementierung: Methanol wurde in Echtzeit gemessen undautomatisch auf 0.5 % (v/v) geregelt (durchgezogene Linie). Glyzerin wurde während derInduktionsphase mit einer festen Rate von 4,9 g·l-1·h-1 zugeführt. Die Gelöstsauerstoff-konzentration (durchgezogene Linie) wurde in Echtzeit erfaßt, Frischgewicht (!) gravimetrischund Akkumulation von scFv4813 (") per Oberflächenplasmonresonanz.
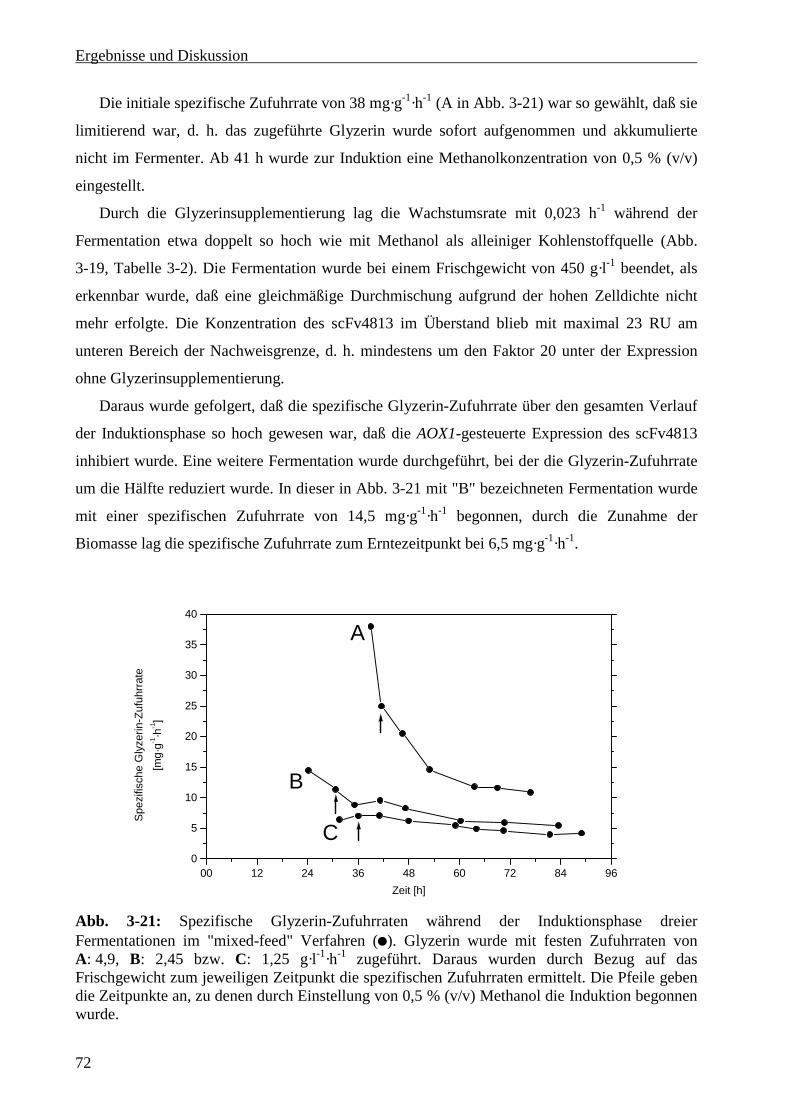
Ergebnisse und Diskussion
72
Die initiale spezifische Zufuhrrate von 38 mg·g-1·h-1 (A in Abb. 3-21) war so gewählt, daß sie
limitierend war, d. h. das zugeführte Glyzerin wurde sofort aufgenommen und akkumulierte
nicht im Fermenter. Ab 41 h wurde zur Induktion eine Methanolkonzentration von 0,5 % (v/v)
eingestellt.
Durch die Glyzerinsupplementierung lag die Wachstumsrate mit 0,023 h-1 während der
Fermentation etwa doppelt so hoch wie mit Methanol als alleiniger Kohlenstoffquelle (Abb.
3-19, Tabelle 3-2). Die Fermentation wurde bei einem Frischgewicht von 450 g·l-1 beendet, als
erkennbar wurde, daß eine gleichmäßige Durchmischung aufgrund der hohen Zelldichte nicht
mehr erfolgte. Die Konzentration des scFv4813 im Überstand blieb mit maximal 23 RU am
unteren Bereich der Nachweisgrenze, d. h. mindestens um den Faktor 20 unter der Expression
ohne Glyzerinsupplementierung.
Daraus wurde gefolgert, daß die spezifische Glyzerin-Zufuhrrate über den gesamten Verlauf
der Induktionsphase so hoch gewesen war, daß die AOX1-gesteuerte Expression des scFv4813
inhibiert wurde. Eine weitere Fermentation wurde durchgeführt, bei der die Glyzerin-Zufuhrrate
um die Hälfte reduziert wurde. In dieser in Abb. 3-21 mit "B" bezeichneten Fermentation wurde
mit einer spezifischen Zufuhrrate von 14,5 mg·g-1·h-1 begonnen, durch die Zunahme der
Biomasse lag die spezifische Zufuhrrate zum Erntezeitpunkt bei 6,5 mg·g-1·h-1.
00 12 24 36 48 60 72 84 960
5
10
15
20
25
30
35
40
C
B
A
Spe
zifis
che
Gly
zerin
-Zuf
uhrr
ate
[mg·
g-1·h
-1]
Zeit [h]
Abb. 3-21: Spezifische Glyzerin-Zufuhrraten während der Induktionsphase dreierFermentationen im "mixed-feed" Verfahren ("). Glyzerin wurde mit festen Zufuhrraten vonA: 4,9, B: 2,45 bzw. C: 1,25 g·l-1·h-1 zugeführt. Daraus wurden durch Bezug auf dasFrischgewicht zum jeweiligen Zeitpunkt die spezifischen Zufuhrraten ermittelt. Die Pfeile gebendie Zeitpunkte an, zu denen durch Einstellung von 0,5 % (v/v) Methanol die Induktion begonnenwurde.
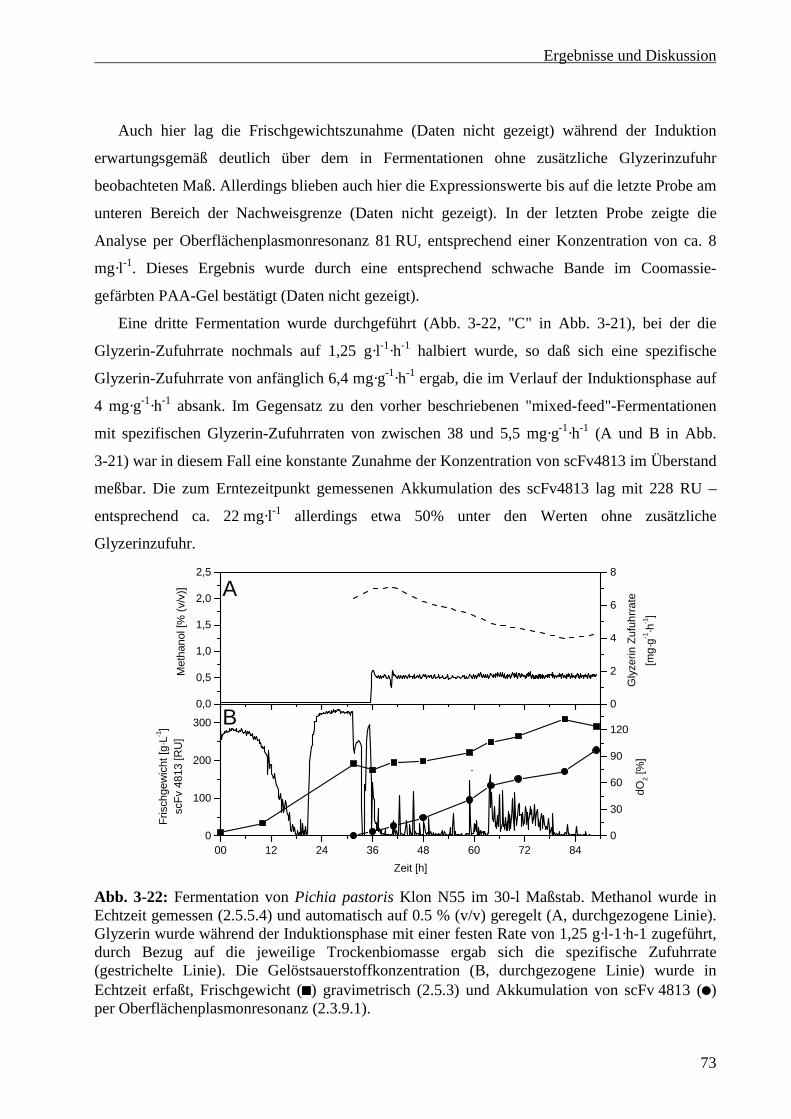
Ergebnisse und Diskussion
73
Auch hier lag die Frischgewichtszunahme (Daten nicht gezeigt) während der Induktion
erwartungsgemäß deutlich über dem in Fermentationen ohne zusätzliche Glyzerinzufuhr
beobachteten Maß. Allerdings blieben auch hier die Expressionswerte bis auf die letzte Probe am
unteren Bereich der Nachweisgrenze (Daten nicht gezeigt). In der letzten Probe zeigte die
Analyse per Oberflächenplasmonresonanz 81 RU, entsprechend einer Konzentration von ca. 8
mg·l-1. Dieses Ergebnis wurde durch eine entsprechend schwache Bande im Coomassie-
gefärbten PAA-Gel bestätigt (Daten nicht gezeigt).
Eine dritte Fermentation wurde durchgeführt (Abb. 3-22, "C" in Abb. 3-21), bei der die
Glyzerin-Zufuhrrate nochmals auf 1,25 g·l-1·h-1 halbiert wurde, so daß sich eine spezifische
Glyzerin-Zufuhrrate von anfänglich 6,4 mg·g-1·h-1 ergab, die im Verlauf der Induktionsphase auf
4 mg·g-1·h-1 absank. Im Gegensatz zu den vorher beschriebenen "mixed-feed"-Fermentationen
mit spezifischen Glyzerin-Zufuhrraten von zwischen 38 und 5,5 mg·g-1·h-1 (A und B in Abb.
3-21) war in diesem Fall eine konstante Zunahme der Konzentration von scFv4813 im Überstand
meßbar. Die zum Erntezeitpunkt gemessenen Akkumulation des scFv4813 lag mit 228 RU –
entsprechend ca. 22 mg·l-1 allerdings etwa 50% unter den Werten ohne zusätzliche
Glyzerinzufuhr.
00 12 24 36 48 60 72 840
100
200
300
Fris
chge
wic
ht [g
·L-1]
scF
v 48
13 [R
U]
Zeit [h]
0,0
0,5
1,0
1,5
2,0
2,5
Met
hano
l [%
(v/
v)]
0
2
4
6
8
Gly
zerin
Zuf
uhrr
ate
[mg·
g-1·h
-1]
0
30
60
90
120B
A
dO2
[%]
Abb. 3-22: Fermentation von Pichia pastoris Klon N55 im 30-l Maßstab. Methanol wurde inEchtzeit gemessen (2.5.5.4) und automatisch auf 0.5 % (v/v) geregelt (A, durchgezogene Linie).Glyzerin wurde während der Induktionsphase mit einer festen Rate von 1,25 g·l-1·h-1 zugeführt,durch Bezug auf die jeweilige Trockenbiomasse ergab sich die spezifische Zufuhrrate(gestrichelte Linie). Die Gelöstsauerstoffkonzentration (B, durchgezogene Linie) wurde inEchtzeit erfaßt, Frischgewicht (!) gravimetrisch (2.5.3) und Akkumulation von scFv 4813 (")per Oberflächenplasmonresonanz (2.3.9.1).
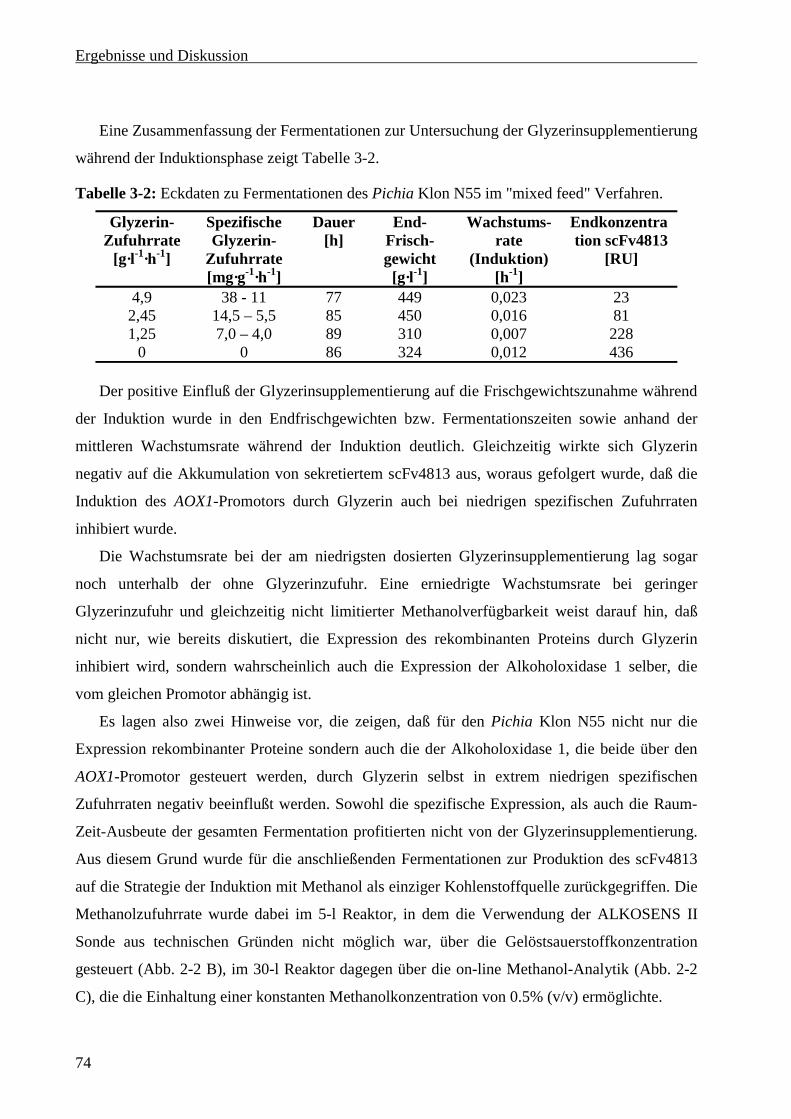
Ergebnisse und Diskussion
74
Eine Zusammenfassung der Fermentationen zur Untersuchung der Glyzerinsupplementierung
während der Induktionsphase zeigt Tabelle 3-2.
Tabelle 3-2: Eckdaten zu Fermentationen des Pichia Klon N55 im "mixed feed" Verfahren.
Glyzerin-Zufuhrrate
[g·l-1·h-1]
SpezifischeGlyzerin-
Zufuhrrate[mg·g-1·h-1]
Dauer[h]
End-Frisch-gewicht
[g·l-1]
Wachstums-rate
(Induktion)[h-1]
Endkonzentration scFv4813
[RU]
4,9 38 - 11 77 449 0,023 232,45 14,5 – 5,5 85 450 0,016 811,25 7,0 – 4,0 89 310 0,007 228
0 0 86 324 0,012 436
Der positive Einfluß der Glyzerinsupplementierung auf die Frischgewichtszunahme während
der Induktion wurde in den Endfrischgewichten bzw. Fermentationszeiten sowie anhand der
mittleren Wachstumsrate während der Induktion deutlich. Gleichzeitig wirkte sich Glyzerin
negativ auf die Akkumulation von sekretiertem scFv4813 aus, woraus gefolgert wurde, daß die
Induktion des AOX1-Promotors durch Glyzerin auch bei niedrigen spezifischen Zufuhrraten
inhibiert wurde.
Die Wachstumsrate bei der am niedrigsten dosierten Glyzerinsupplementierung lag sogar
noch unterhalb der ohne Glyzerinzufuhr. Eine erniedrigte Wachstumsrate bei geringer
Glyzerinzufuhr und gleichzeitig nicht limitierter Methanolverfügbarkeit weist darauf hin, daß
nicht nur, wie bereits diskutiert, die Expression des rekombinanten Proteins durch Glyzerin
inhibiert wird, sondern wahrscheinlich auch die Expression der Alkoholoxidase 1 selber, die
vom gleichen Promotor abhängig ist.
Es lagen also zwei Hinweise vor, die zeigen, daß für den Pichia Klon N55 nicht nur die
Expression rekombinanter Proteine sondern auch die der Alkoholoxidase 1, die beide über den
AOX1-Promotor gesteuert werden, durch Glyzerin selbst in extrem niedrigen spezifischen
Zufuhrraten negativ beeinflußt werden. Sowohl die spezifische Expression, als auch die Raum-
Zeit-Ausbeute der gesamten Fermentation profitierten nicht von der Glyzerinsupplementierung.
Aus diesem Grund wurde für die anschließenden Fermentationen zur Produktion des scFv4813
auf die Strategie der Induktion mit Methanol als einziger Kohlenstoffquelle zurückgegriffen. Die
Methanolzufuhrrate wurde dabei im 5-l Reaktor, in dem die Verwendung der ALKOSENS II
Sonde aus technischen Gründen nicht möglich war, über die Gelöstsauerstoffkonzentration
gesteuert (Abb. 2-2 B), im 30-l Reaktor dagegen über die on-line Methanol-Analytik (Abb. 2-2
C), die die Einhaltung einer konstanten Methanolkonzentration von 0.5% (v/v) ermöglichte.
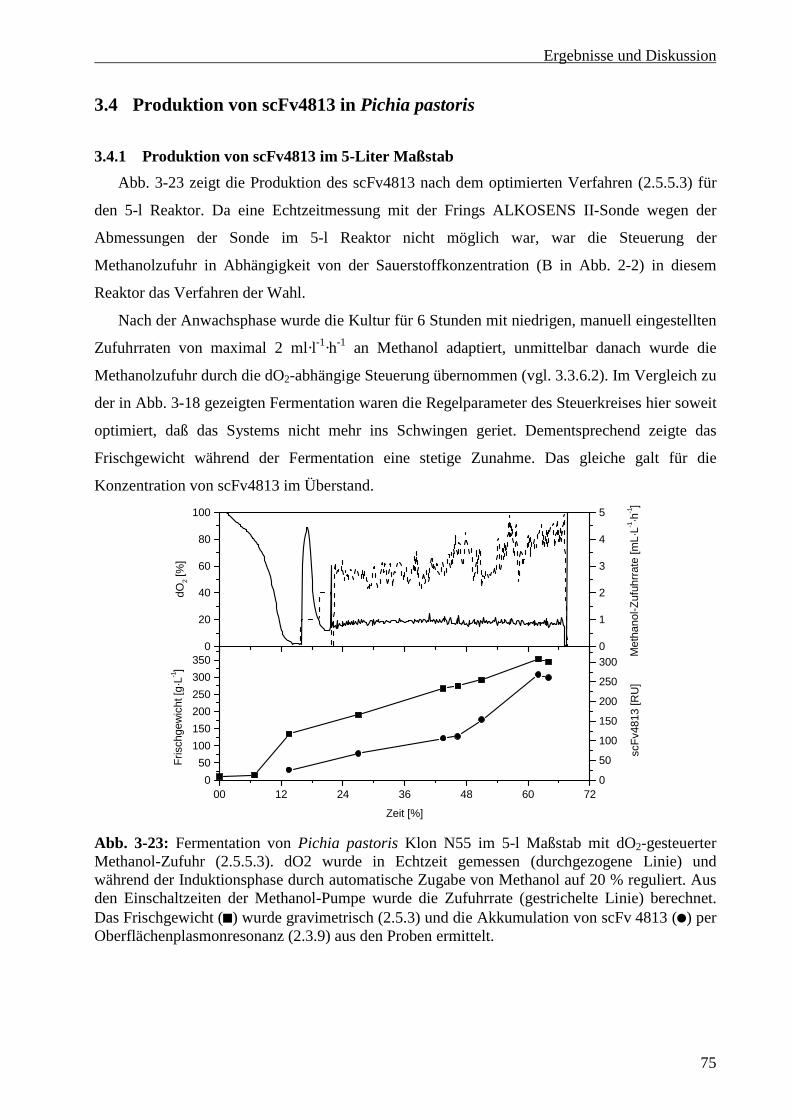
Ergebnisse und Diskussion
75
3.4 Produktion von scFv4813 in Pichia pastoris
3.4.1 Produktion von scFv4813 im 5-Liter Maßstab
Abb. 3-23 zeigt die Produktion des scFv4813 nach dem optimierten Verfahren (2.5.5.3) für
den 5-l Reaktor. Da eine Echtzeitmessung mit der Frings ALKOSENS II-Sonde wegen der
Abmessungen der Sonde im 5-l Reaktor nicht möglich war, war die Steuerung der
Methanolzufuhr in Abhängigkeit von der Sauerstoffkonzentration (B in Abb. 2-2) in diesem
Reaktor das Verfahren der Wahl.
Nach der Anwachsphase wurde die Kultur für 6 Stunden mit niedrigen, manuell eingestellten
Zufuhrraten von maximal 2 ml·l-1·h-1 an Methanol adaptiert, unmittelbar danach wurde die
Methanolzufuhr durch die dO2-abhängige Steuerung übernommen (vgl. 3.3.6.2). Im Vergleich zu
der in Abb. 3-18 gezeigten Fermentation waren die Regelparameter des Steuerkreises hier soweit
optimiert, daß das Systems nicht mehr ins Schwingen geriet. Dementsprechend zeigte das
Frischgewicht während der Fermentation eine stetige Zunahme. Das gleiche galt für die
Konzentration von scFv4813 im Überstand.
00 12 24 36 48 60 720
50
100
150
200
250
300
350
Fris
chge
wic
ht [g
·L-1]
Zeit [%]
0
20
40
60
80
100
dO2
[%]
0
50
100
150
200
250
300
scF
v481
3 [R
U]
0
1
2
3
4
5
Met
hano
l-Zuf
uhrr
ate
[mL·
L-1·h
-1]
Abb. 3-23: Fermentation von Pichia pastoris Klon N55 im 5-l Maßstab mit dO2-gesteuerterMethanol-Zufuhr (2.5.5.3). dO2 wurde in Echtzeit gemessen (durchgezogene Linie) undwährend der Induktionsphase durch automatische Zugabe von Methanol auf 20 % reguliert. Ausden Einschaltzeiten der Methanol-Pumpe wurde die Zufuhrrate (gestrichelte Linie) berechnet.Das Frischgewicht (!) wurde gravimetrisch (2.5.3) und die Akkumulation von scFv 4813 (") perOberflächenplasmonresonanz (2.3.9) aus den Proben ermittelt.
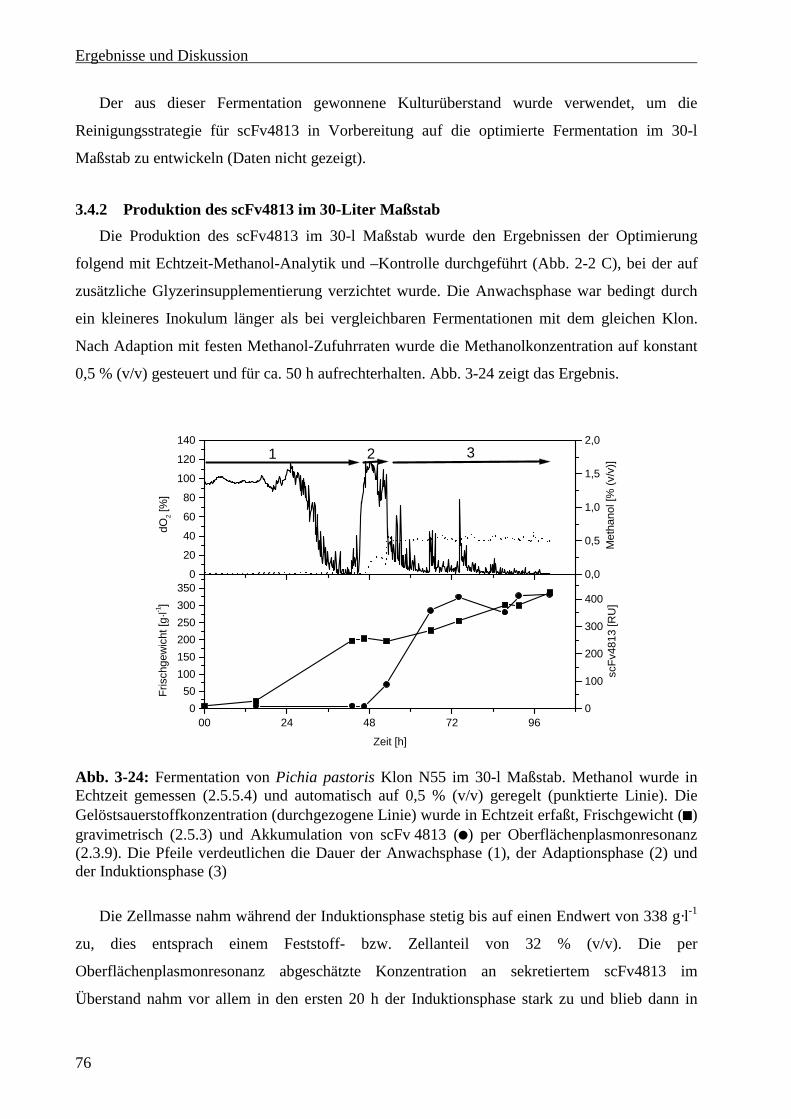
Ergebnisse und Diskussion
76
Der aus dieser Fermentation gewonnene Kulturüberstand wurde verwendet, um die
Reinigungsstrategie für scFv4813 in Vorbereitung auf die optimierte Fermentation im 30-l
Maßstab zu entwickeln (Daten nicht gezeigt).
3.4.2 Produktion des scFv4813 im 30-Liter Maßstab
Die Produktion des scFv4813 im 30-l Maßstab wurde den Ergebnissen der Optimierung
folgend mit Echtzeit-Methanol-Analytik und –Kontrolle durchgeführt (Abb. 2-2 C), bei der auf
zusätzliche Glyzerinsupplementierung verzichtet wurde. Die Anwachsphase war bedingt durch
ein kleineres Inokulum länger als bei vergleichbaren Fermentationen mit dem gleichen Klon.
Nach Adaption mit festen Methanol-Zufuhrraten wurde die Methanolkonzentration auf konstant
0,5 % (v/v) gesteuert und für ca. 50 h aufrechterhalten. Abb. 3-24 zeigt das Ergebnis.
00 24 48 72 960
50
100
150
200
250
300
350
Fris
chge
wic
ht [g
·l-1]
Zeit [h]
0
20
40
60
80
100
120
140321
dO2
[%]
0
100
200
300
400sc
Fv4
813
[RU
]
0,0
0,5
1,0
1,5
2,0
Met
hano
l [%
(v/
v)]
Abb. 3-24: Fermentation von Pichia pastoris Klon N55 im 30-l Maßstab. Methanol wurde inEchtzeit gemessen (2.5.5.4) und automatisch auf 0,5 % (v/v) geregelt (punktierte Linie). DieGelöstsauerstoffkonzentration (durchgezogene Linie) wurde in Echtzeit erfaßt, Frischgewicht (!)gravimetrisch (2.5.3) und Akkumulation von scFv 4813 (") per Oberflächenplasmonresonanz(2.3.9). Die Pfeile verdeutlichen die Dauer der Anwachsphase (1), der Adaptionsphase (2) undder Induktionsphase (3)
Die Zellmasse nahm während der Induktionsphase stetig bis auf einen Endwert von 338 g·l-1
zu, dies entsprach einem Feststoff- bzw. Zellanteil von 32 % (v/v). Die per
Oberflächenplasmonresonanz abgeschätzte Konzentration an sekretiertem scFv4813 im
Überstand nahm vor allem in den ersten 20 h der Induktionsphase stark zu und blieb dann in

Ergebnisse und Diskussion
77
etwa konstant bei einem Wert um 40 mg·l-1. Die Fermentation wurde nach 100,5 h gestoppt.
Abb. 3-25 zeigt ein Coomassie-gefärbtes PAA-Gel der Fermentationsüberstände. Die gesamte
Fermentationsbrühe wurde aufgearbeitet.
Abb. 3-25: Coomassie-gefärbtes PAA-Gel (2.3.5) von Fermentationsüberständen der in Abb.3-24 gezeigten Fermentation. Der Pfeil zeigt die Banden an, deren Größe mit der für scFv4813berechneten Größe übereinstimmt. Die Proben wurden zu den über den jeweiligen Spurengenannten Zeitpunkten gezogen. Pro Spur wurden 30 µl Überstand geladen.
3.5 Reinigung von scFv4813 im Gramm-Maßstab
3.5.1 Zellabtrennung
Vor der Zellabtrennung wurde die Fermentationsbrühe durch Zugabe von 10xPBS, festem
NaCl und NaOH auf 1 M NaCl, 1xPBS pH 7.5 eingestellt, um eine gute Bindung des scFv4813
in der nachfolgenden Ni-IDA Affinitätschromatographie zu erreichen. Wegen des erheblichen
Zellanteils von 32 % wurde die Zugabe von 10xPBS und NaCl auf 0,68 Liter Überstand pro
Liter zellhaltiger Fermentationsbrühe berechnet. Zur Abtrennung der Zellen wurde eine CARR
Powerfuge P6 verwendet. Die Zellabtrennung erfolgte mit einer Effizienz von > 99% in 11
Trennzyklen innerhalb von 80 Minuten. Die Zellmasse wurde verworfen und der Überstand
wurde bis zur Ni-IDA-Affinitätschromatographie (2.3.10.1) bei 4° C gelagert.
3.5.2 Metall-Affinitätschromatographie
Da die Kapazität der zur Verfügung stehenden Streamline-Säule zu gering war, wurden
insgesamt 23,4 l Fermentationsüberstand in 6 Chargen von etwa 4 l einer Metall-
Affinitätschromatographie unterzogen: Die Säule wurde im expandierten Modus (2.3.10.1) bei
einer Fließrate von 20 ml·min-1 äquilibriert (3 SV), mit Nickel beladen (2 SV) und gewaschen (3
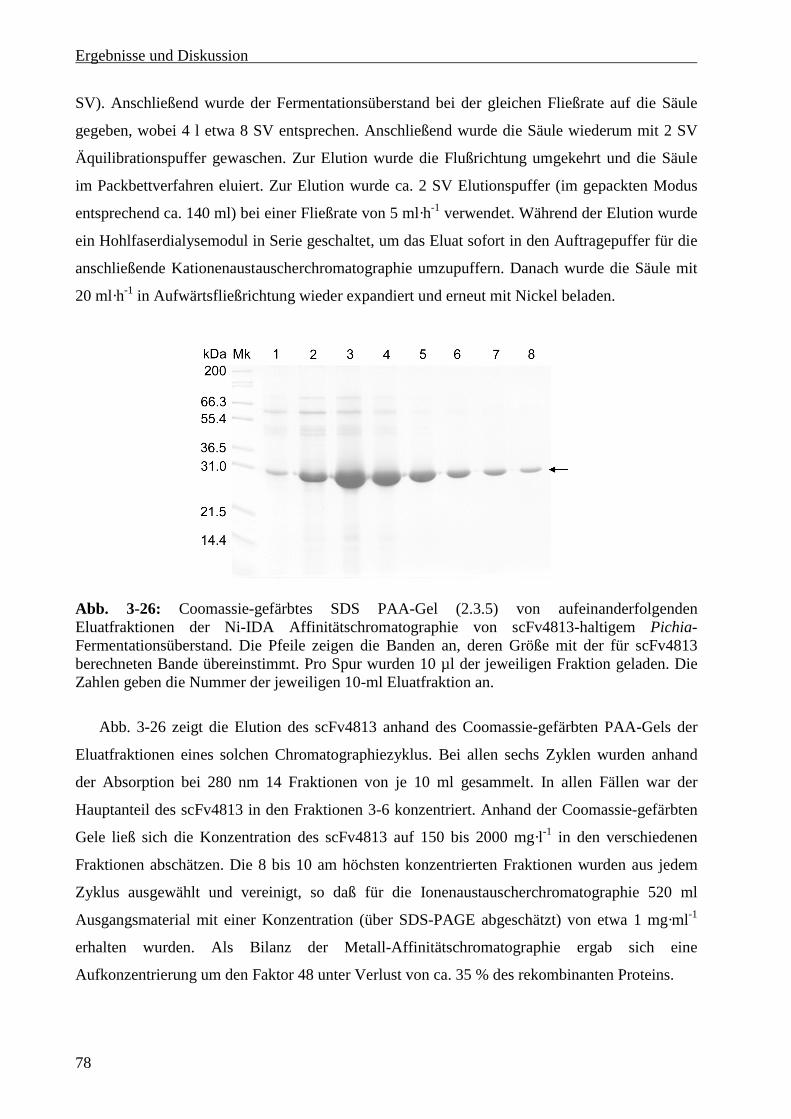
Ergebnisse und Diskussion
78
SV). Anschließend wurde der Fermentationsüberstand bei der gleichen Fließrate auf die Säule
gegeben, wobei 4 l etwa 8 SV entsprechen. Anschließend wurde die Säule wiederum mit 2 SV
Äquilibrationspuffer gewaschen. Zur Elution wurde die Flußrichtung umgekehrt und die Säule
im Packbettverfahren eluiert. Zur Elution wurde ca. 2 SV Elutionspuffer (im gepackten Modus
entsprechend ca. 140 ml) bei einer Fließrate von 5 ml·h-1 verwendet. Während der Elution wurde
ein Hohlfaserdialysemodul in Serie geschaltet, um das Eluat sofort in den Auftragepuffer für die
anschließende Kationenaustauscherchromatographie umzupuffern. Danach wurde die Säule mit
20 ml·h-1 in Aufwärtsfließrichtung wieder expandiert und erneut mit Nickel beladen.
Abb. 3-26: Coomassie-gefärbtes SDS PAA-Gel (2.3.5) von aufeinanderfolgendenEluatfraktionen der Ni-IDA Affinitätschromatographie von scFv4813-haltigem Pichia-Fermentationsüberstand. Die Pfeile zeigen die Banden an, deren Größe mit der für scFv4813berechneten Bande übereinstimmt. Pro Spur wurden 10 µl der jeweiligen Fraktion geladen. DieZahlen geben die Nummer der jeweiligen 10-ml Eluatfraktion an.
Abb. 3-26 zeigt die Elution des scFv4813 anhand des Coomassie-gefärbten PAA-Gels der
Eluatfraktionen eines solchen Chromatographiezyklus. Bei allen sechs Zyklen wurden anhand
der Absorption bei 280 nm 14 Fraktionen von je 10 ml gesammelt. In allen Fällen war der
Hauptanteil des scFv4813 in den Fraktionen 3-6 konzentriert. Anhand der Coomassie-gefärbten
Gele ließ sich die Konzentration des scFv4813 auf 150 bis 2000 mg·l-1 in den verschiedenen
Fraktionen abschätzen. Die 8 bis 10 am höchsten konzentrierten Fraktionen wurden aus jedem
Zyklus ausgewählt und vereinigt, so daß für die Ionenaustauscherchromatographie 520 ml
Ausgangsmaterial mit einer Konzentration (über SDS-PAGE abgeschätzt) von etwa 1 mg·ml-1
erhalten wurden. Als Bilanz der Metall-Affinitätschromatographie ergab sich eine
Aufkonzentrierung um den Faktor 48 unter Verlust von ca. 35 % des rekombinanten Proteins.

Ergebnisse und Diskussion
79
Dieser relativ hohe Verlust an Ausgangsmaterial ist in erster Linie auf eine Überladung der
Säule zurückzuführen. Es hatte sich gezeigt, daß bei Verwendung von 70 ml Matrix nach
Auftrag von 2 Liter Pichia-Überstand zunehmende Mengen des scFv4813 im Durchfluß
nachweisbar waren. Da jedoch keine größere Expanded-bed Säule zur Verfügung stand und die
Prozessierung des in 6 Chargen von je 4 Litern aufgeteilten Überstandes ohnehin schon ca. 32
Stunden in Anspruch nahm, wurde dieser Verlust in Kauf genommen, um die Aufreinigung
möglichst zügig durchzuführen. Eventuelle Degradationsprodukte des Proteins wären unter
Umständen nur schwierig abzutrennen.
Der Haupteffekt der Affinitätschromatographie bestand in der Anreicherung des Produktes.
Die Reinheit des Eluats ist wie in Abb. 3-26 erkennbar, noch nicht allzu hoch. Zwischen 45 und
80 kDa sind noch mindestens 5 weitere Proteinbanden oberhalb der Bande des scFv4813 zu
erkennen, sowie ein kleines Protein bei ca. 15 kDa. Da ein Teil des produzierten Materials für
die Kristallisation und Strukturaufklärung verwendet werden sollte, wurden weitere
Reinigungsschritte durchgeführt.
3.5.3 Ionenaustauscherchromatographie
Aufgrund des anhand der Aminosäuresequenz ermittelten pI von 8,77 wurde als nächster
Reinigungschritt eine Kationenaustauscherchromatographie durchgeführt. Auch die für die
Ionenaustauscherchromatographie verfügbaren Säulen ließen die Prozessierung des vereinigten
Eluats aus 3.5.2 nur in drei Chargen zu. Es wurden jeweils 20-25 SV bei einer Fließrate von
10 ml·min-1 auf die Säule gegeben. Zur Elution wurde der Gradient in 100 % Elutionspuffer über
20 min gefahren. Die Säule wurde mit 100 % Elutionspuffer gewaschen (8-10 SV) und
anschließend mit ca. 10-15 SV Ladepuffer äquilibriert, bevor eine weitere Charge geladen
wurde.
Die Elution des scFv4813 erfolgte bei der gewählten Steilheit des Gradienten bei
Salzkonzentrationen zwischen 0,4 und 0,5 M NaCl. Das Eluat wurde in 10-ml Fraktionen
gesammelt. Abb. 3-27 zeigt je eine Eluatfraktionen der drei Chargen. Insgesamt wurden 450 ml
Eluat erhalten, die wiederum in 5 Chargen aufkonzentriert (2.3.10.3) und für die Gelfiltration
verwendet wurden.
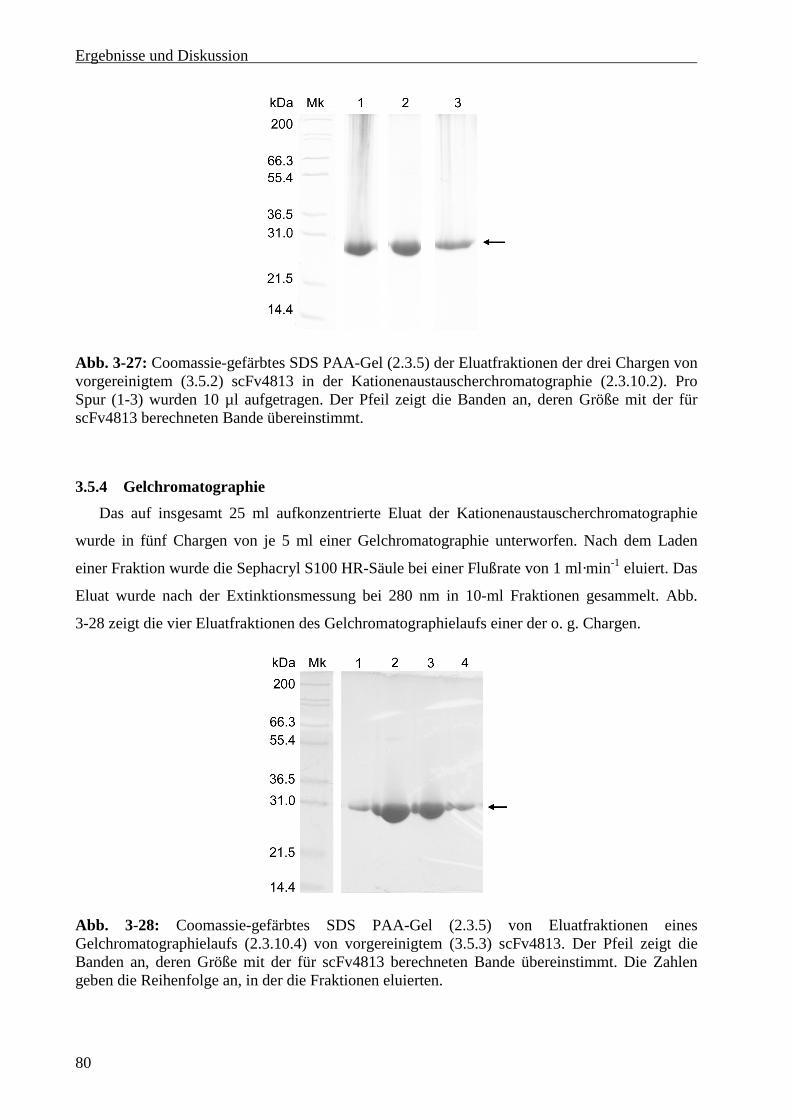
Ergebnisse und Diskussion
80
Abb. 3-27: Coomassie-gefärbtes SDS PAA-Gel (2.3.5) der Eluatfraktionen der drei Chargen vonvorgereinigtem (3.5.2) scFv4813 in der Kationenaustauscherchromatographie (2.3.10.2). ProSpur (1-3) wurden 10 µl aufgetragen. Der Pfeil zeigt die Banden an, deren Größe mit der fürscFv4813 berechneten Bande übereinstimmt.
3.5.4 Gelchromatographie
Das auf insgesamt 25 ml aufkonzentrierte Eluat der Kationenaustauscherchromatographie
wurde in fünf Chargen von je 5 ml einer Gelchromatographie unterworfen. Nach dem Laden
einer Fraktion wurde die Sephacryl S100 HR-Säule bei einer Flußrate von 1 ml·min-1 eluiert. Das
Eluat wurde nach der Extinktionsmessung bei 280 nm in 10-ml Fraktionen gesammelt. Abb.
3-28 zeigt die vier Eluatfraktionen des Gelchromatographielaufs einer der o. g. Chargen.
Abb. 3-28: Coomassie-gefärbtes SDS PAA-Gel (2.3.5) von Eluatfraktionen einesGelchromatographielaufs (2.3.10.4) von vorgereinigtem (3.5.3) scFv4813. Der Pfeil zeigt dieBanden an, deren Größe mit der für scFv4813 berechneten Bande übereinstimmt. Die Zahlengeben die Reihenfolge an, in der die Fraktionen eluierten.

Ergebnisse und Diskussion
81
Aus jedem Gelchromatographielauf wurden die beiden Hauptfraktionen gesammelt und
vereinigt, so daß zwei Eluatfraktionen ("A" und "B") erhalten wurden. Diese beiden Fraktionen
wurden einer genaueren Konzentrationsbestimmung unterworfen, um die Gesamtausbeute der
Fermentation nach der Reinigung des Proteins ermitteln zu können.
3.5.5 Konzentrationsbestimmung des scFv4813
Die Konzentrationsbestimmung des gereinigten scFv4813 (3.5.1-3.5.4) wurde mit mehreren
Methoden vorgenommen: Einerseits wurde das Protein mit Bradford-Reagenz gefärbt und die
Konzentration durch Korrelation mit einer aus RSA erstellten Kalibrationskurve bestimmt.
Zusätzlich wurde aus der Aminosäuresequenz der theoretische millimolare
Extinktionskoeffizient bei 280 nm ermittelt und die Konzentration des scFv4813 im Eluat der
Gelfiltration durch Anwendung des Lambert-Beer'schen Gesetzes bestimmt. Die Meßwerte sind
in Abb. 3-29 dargestellt.
0 10 20 30 40 500,00
0,05
0,10
0,15
0,20
0,25
0,30 A
Eluat A
1:100Eluat B1:100
E5
95
Konzentration RSA [µg·ml-1]
0,00 0,05 0,10 0,15 0,20 0,25 0,300,0
0,3
0,6
0,9
1,2
1,5
1,8 B
E28
0
1 / Verdünnung
Abb. 3-29: Konzentrationsbestimmung des gereinigten scFv4813. A: Messung der Extinktionbei 595 nm nach Anfärbung mit Bradford-Reagenz (2.3.9.3). Die Kalibrationskurve des RSA-Standards (!) und die mit 1:100 Verdünnungen der Eluatfraktionen A und B erhaltenenExtinktionen sind angegeben. B: Messung der Extinktion bei 280 nm (2.3.9.2). Die Extinktionender Eluate A (!) und B (#) sind über den Kehrwert der Verdünnung aufgetragen, um eine lineareKorrelation zu ermöglichen.
Bei der Konzentrationsbestimmung über den Bradford-Komplex lagen lediglich die 1:100
Verdünnungen im linearen Bereich der Kalibrationskurve mit RSA. Bei der Bestimmung der
Extinktion bei 280 nm zeigten die Verdünnungen über einen weiten Konzentrationsbereich eine
lineare Korrelation, so daß die Steigung der linearen Ausgleichsgeraden zur
Konzentrationsbestimmung verwendet wurde. Aus dem millimolaren Extinktionskoeffizienten

Ergebnisse und Diskussion
82
von 49170 ergibt sich eine Extinktion von 1,77 bei einer Konzentration von 1 mg·ml-1. Tabelle
3-3 zeigt die Ergebnisse der Konzentrationsbestimmungen.
Tabelle 3-3: Ergebnisse der Konzentrationsbestimmung der beiden Eluatfraktionen.
Bestimmungsmethode Eluat A [mg·ml-1] Eluat B [mg·ml-1]Bradford-Färbung 2,87 2,48E280 8,703 6,527
Die Ergebnisse der Konzentrationsbestimmungen wichen fast um den Faktor drei
voneinander ab, was aber nicht unüblich ist, da die Intensität der Bradford-Färbung vom
jeweiligen Protein abhängt, und als Standard RSA verwendet wurde, das ca. 1,5 mal so
empfindlich mit Bradford-Lösung reagiert, wie z. B. IgG (Angabe lt. Hersteller). Abschätzungen
der Konzentration über die Größe von Coomassie-gefärbten SDS-PAA-Gelen ergeben
Konzentrationen um 5 mg·ml-1 (Daten nicht gezeigt). Die Linearität der Bestimmung über dei
Extinktion be 280nm über einen Verdünnungsbereich von zwei Zehnerpotenzen (s. o.), die dabei
zugrunde liegende Verwendung eines proteinsequenzspezifischen Extinktionskoeffizienten und
die hohe Reinheit des Proteins, die in Abb. 3-28 erkennbar ist, sprechen dafür, daß in diesem Fall
die Methode der photometrischen Konzentrationsbestimmung das korrektere Ergebnis liefert.
Danach enthielt das gesamte Eluat "A" in 35 ml ca. 307 mg scFv4813, das gesamte Eluat "B" in
33 ml ca. 215 mg scFv4813.
Die Gesamtausbeute der 30-l Fermentation betrug demnach über 500 mg hochreinen
Proteins. Bei einer abgeschätzten Konzentration von ca. 40 mg·l-1 im Fermentationsüberstand
entspricht das einer Reinigungseffizienz von ca. 54 %. Für den größten Teil des Verlusts ist die
Überladung der Streamline-Säule bei der Metall-Affinitätschromatographie verantwortlich (s.o.),
weitere Verluste entstanden durch das großzügige "Abschneiden" der Elutionspeaks bei der
Auswahl der Fraktionen, die jeweils weiterverarbeitet wurden.
3.5.6 Stabilität und Lagerung von scFv4813
Die Lagerfähigkeit des gereinigten scFv4813 wurde überprüft, indem Aliquots der
Eluatfraktion A unter verschiedenen Bedingungen 3 Wochen lang gelagert und anschließend auf
ihr Erscheinungsbild im Coomassie-gefärbten PAA-Gel und ihre Reaktivität (BIACore-
Bestimmung über das his6-Motiv, Kap. 2.3.9.1) untersucht wurden.
Die unverdünnten Proben wurden bei RT, 4° C, -20° C und –70° C gelagert, wobei bei den
Aliquoten bei RT und bei 4° C jeweils ein Ansatz mit 0,1 % (w/v) NaN3 und ein Ansatz ohne
NaN3 untersucht wurde. Das Ergebnis ist in Abb. 3-30 gezeigt. Nach Lagerung bei

Ergebnisse und Diskussion
83
Raumtemperatur und ohne zusätzliche Konservierung mit Hilfe von Natriumazid war nach 3
Wochen eine deutliche Abnahme der mit Hilfe der Oberflächenplasmonresonanz gemessenen
Konzentration festzustellen. Zusätzlich waren bei den oberhalb von 0° C gelagerten Proben
leichte Abnahmen der Konzentration gegenüber den bei -20° C und bei -70° C gelagerten Proben
erkennbar. Die Ergebnisse waren in den 1:10 verdünnten Proben tendenziell gleich. Der erste
Befund deutet auf mikrobielle Zersetzung des Proteins hin, der zweite auf nicht-mikrobielle
Degradation. Vor der Lagerung der Proben ergab die Messung nach 2.3.9.1 ca. 11000 RU, ein
direkter Vergleich dieser Messung mit den in Abb. 3-30 gezeigten Werten ist aber nicht
aussagekräftig, da die Messungen mit Hilfe der Oberflächenplasmonresonanz vom Zustand des
verwendeten NTA-Chips abhängen, dessen Qualität über einen Zeitraum von drei Wochen nicht
konstant bleibt. Daher ließen sich nur Werte von zeitnah gemessenen Proben vergleichen. Im
Western-Blot der entsprechenden Proben waren keine Degradationsbanden zu erkennen,
obgleich die Bande des bei RT gelagerten Aliquots deutlich schwächer geworden war (Daten
nicht gezeigt).
RT RT (+Azid) 4° C 4° C (+Azid) -20° C -70° C0
2000
4000
6000
8000
10000
scF
v481
3 [R
U]
Abb. 3-30: Lagerstabilität des gereinigten scFv4813 (2.3.10). Aliquote des scFv4813 wurden beiverschiedenen Temperaturen für 3 Wochen z. T. unter Zusatz von Natriumazid gelagert. DieStabilität von 1:1 (helle Säulen) und 1:10 (dunkle Säulen) verdünntem, gereinigtem (3.5.2-3.5.4)scFv4813 (Eluat "A") anschließend durch Konzentrationsbestimmung mit Hilfe der Oberflächen-plasmonresonanz (2.3.9.1) bestimmt.
Die Bestimmung der Konzentration über das His6-Motiv ist nicht mit einer
Aktivitätsbestimmung des Proteins gleichzusetzen, sie läßt zuverlässige Rückschlüsse nur über
eine eventuelle Degradation in Form von proteolytischer Zersetzung zu, nicht aber Aussagen in
Bezug auf die Funktionalität des Proteins. Dennoch zeigen die Ergebnisse, daß der scFv4813 bei
-20° C und -70° C gut lagerfähig ist, mit Einschränkungen sogar bei 4° C.

Abschlußdiskussion und Ausblick
84
4 Abschlußdiskussion und Ausblick
4.1 Produktion des scFv4813 durch Fermentation von Nicotiana tabacum cvBY-2
Erste Versuche zur Entwicklung einer Fermentationsstrategie für die Suspensionskultur
Nicotiana tabacum BY-2 wurden anhand der schon vor Beginn dieser Arbeit am Institut für
Biologie I der RWTH Aachen etablierten Zellinie biscFv2429KDEL (2.1.7) durchgeführt
(Schumann, 1996). Diese Zellinie exprimiert das Gen für ein bispezifisches "single-chain"-
Antikörperfragment unter der Kontrolle des CaMV 35SS Promotors. Obwohl später für die
Produktion des scFv4813 in Nicotiana tabacum cv BY-2 mit der Zellinie R1-25 der (ocs)3mas
Promotor verwendet wurde (Ni et al., 1995), konnten die mit diesem System gewonnenen Daten
weitgehend auf die Produktion des scFv4813 übertragen werden, da es sich bei beiden
rekombinanten Proteinen um von murinen IgG abgeleitete Antikörperfragmente handelte, beide
Promotoren konstitutive Promotoren sind und das rekombinante Protein in beiden Fällen durch
die C-terminale KDEL-Sequenz im ER retardiert wurde. Beide Proteine waren im ER der
Pflanzenzellen stabil und als funktionale scFv im ELISA nachweisbar.
Die Fermentation wurde mit der Zellinie R1-25, die den scFv4813 exprimierte, erfolgreich
bis in den 30-l Maßstab durchgeführt. Der Prozeß war verfahrenstechnisch relativ einfach: Die
Inokulation erfolgte mit 15 bis 20 % (v/v) Vorkultur mit einem PCV von 50 %. Die Temperatur
wurde konstant auf 26° C geregelt und die Gelöstsauerstoffkonzentration wurde durch getaktete
Zugabe von Luft oberhalb von 10-20 % gehalten. Die Durchmischung durch Schrägblattrührer
bei relativ langsamen, konstanten Drehzahlen von 130 (5-l Arbeitsvolumen) bzw. 80 (30-l
Arbeitsvolumen) war effektiv. Zellschädigungen wurden nicht beobachtet. Der pH-Wert wurde
nicht geregelt. Mit dieser Strategie ergab sich eine Fermentationsdauer von 160 bis 200 h. Die
für die BY-2 Zellinie typische Synchronizität der Zellteilungen wurde dabei beobachtet. Aus den
im Überstand leicht analysierbaren Nährstoffkonzentrationen (Phosphat, Saccharose, Glukose,
Fruktose) ergaben sich typische Fermentationskinetiken.
Die intrazellulär im ER akkumulierten Antikörperfragmente wurden nach dem Aufbrechen
der in Extraktionspuffer resuspendierten Zellen durch Ultraschall in ELISA und Western Blot
analysiert sowie per Oberflächenplasmonresonanz analysiert. Dabei ergaben sich für den
biscFv2429 in Verbindung mit dem 35SS Promotor Expressionslevel bis maximal 2,9 mg·l-1
(bezogen auf das Fermentationsvolumen), mit dem (ocs)3mas Promotor lagen die maximal für

Abschlußdiskussion und Ausblick
85
den scFv4813 per Western Blot abgeschätzten Expressionslevel bei maximal 11 mg·l-1. Diese
Werte entsprechen einer Expressionsrate von etwa 0,2 bis 1 % des gesamtlöslichen Proteins.
Diese Werte liegen im Rahmen der von anderen Forschern für die Nicotiana tabacum cv BY-2
Zellinie erhaltenen Ergebnisse (Hein et al., 1991; Firek et al., 1993; Magnuson et al., 1996;
Francisco et al., 1997; Lee et al., 1997; des Molles et al., 1999; Sehnke und Ferl, 1999; Sharp
und Doran, 1999; Fischer et al., 1999a; Doran, 2000). Am Institut für Biologie I der RWTH
Aachen durchgeführte Versuche zur Expression von Proteinen in Nicotiana tabacum cv BY-2
und deren Sekretion ins Medium waren zwar prinzipiell erfolgreich (Drossard, 1996; Fischer et
al., 1999a; Fischer et al., 1999e; Fischer et al., 1999f), aber die bezogen auf das
Fermentationsvolumen erreichten Ausbeuten waren im Einklang mit den Ergebnissen anderer
Gruppen (Doran, 2000) zu gering, um diesen Weg, der eine deutlich einfachere Reinigung der
Proteine ermöglicht hätte, zu rechtfertigen. Der einzige Bericht, der die Expression eines
rekombinanten Proteins in sekretierter Form in pflanzlichen Zellkulturen in größeren Mengen
(Ausbeute vor Reinigung 85 mg·l-1 bezogen auf das Fermentationsvolumen) beschreibt,
behandelt die Expression von humanem α-1-Antitrypsin (hAAT) in einer Kultur von Orhiza
japonica (Terashima et al., 1999).
Versuche, die Expression in Pflanzensuspensionskulturen durch Zugabe von Elicitoren,
Aminosäuren, Licht oder Pflanzenhormone zu steigern, haben zwar in einzelnen Fällen Erfolg
gezeigt (Sehnke und Ferl, 1999; Fischer et al., 1999a), wie sich im Falle der Zellinie R1-25 im
Rahmen dieser Arbeit zeigte (3.1.4), sind diese Effekte aber nicht ohne weiteres auf bestimmte
Kombinationen von Promotor und zu exprimierendem Protein übertragbar. In diesem Bereich
besteht noch weiterer Handlungsbedarf.
Die Suche nach stärkeren, eventuell auch induzierbaren Promotoren wie dem (ocs)3mas-
Promotor (Ni et al., 1995) oder den von Shinmyo et al. beschriebenen "heat-shock"-Promotoren
(Shinmyo et al., 1998) scheint gerade für die pflanzliche Suspensionskultur von besonderer
Bedeutung, da sich in vielen Fällen gezeigt hat, daß die Expressionsraten (in % des
gesamtlöslichen Proteins) in Suspensionskulturen deutlich unter denen liegen, die sich mit dem
gleichen Promotor in intakten Pflanzen der gleichen Art erreichen lassen (Doran, 2000).
Die Entwicklung und Optimierung der Zellinien biscFv2429KDEL und R1-25 wurde im
Rahmen dieser Arbeit nicht in Betracht gezogen. Zwar zeigten beiden Zellinien auch bei
langfristiger Subkultivierung unter nicht-selektiven Bedingungen keine Variabilität in den
erreichten Expressionsmengen, es ist aber denkbar, daß die Zellinien, die stets in Schüttelkolben
subkultiviert wurden, auf die Bedingungen im Fermenter nicht angepaßt waren. Eine ständige
Kultivierung im Fermenter, etwa durch kontinuierliche bzw. semikontinuierliche Fermentation

Abschlußdiskussion und Ausblick
86
("draw-fill") oder durch Subkultivierung innerhalb eines Netzwerks von Rührkesseln könnte
langfristige Adaptions- und Selektionseffekte zur Ausprägung bringen. Diese Effekte könnten
sich in besserem Wachstum im Fermenter niederschlagen, es besteht aber auch die Gefahr, daß
Selektionsdruck hin zu z. B. schneller wachsenden Zellen mit einer Verringerung der
Expressionslevel einhergeht. Für derart langfristige und aufwendige Untersuchungen standen im
Rahmen der vorliegenden Arbeit allerdings weder ausreichend Zeit noch Fermentationskapazität
zur Verfügung. Entsprechende Langfristversuche stellen aber eine weitere, interessante
Perspektive zur Etablierung pflanzlicher Suspensionskulturen als Alternative neben mikrobiellen
und tierischen Expressionssystemen dar.
4.2 Produktion des scFv4813 durch Fermentation von Pichia pastoris
4.2.1 Fermentationsstrategie
Die Entwicklung einer Fermentationsstrategie für Pichia pastoris erfolgte teilweise nicht
anhand des späteren Expressorklons für den scFv4813 (Klon N55), sondern mit dem Klon
PP599, der ein Fusionsprodukt zweier Domänen des humanen Glykoproteins CEA exprimierte
(You et al., 1998; Hellwig et al., 1999). In beiden Fällen wurde die Expression des
rekombinanten Proteins über den AOX1 Promotor gesteuert und beide Proteine wurden über das
prepro-Sekretionssignal des α-Paarungsfaktors von S. cerevisiae ins Medium sekretiert. Die mit
dem Klon PP599 entwickelte Fermentationsstrategie ließ sich in wesentlichen Teilen auf die
Fermentation von des Klons N55 anwenden. Die vom Hersteller des Expressionssystems
vorgeschlagene "Standard"-Fermentationsstrategie wurde dabei in folgenden Punkten
abgeändert.
4.2.1.1 Medium
Das verwendete Medium "B" (2.5.5.1) enthielt um 50 % reduzierte Konzentrationen aller
Makro-Nährstoffe, ohne daß eine Einschränkung der Wachstumsgeschwindigkeit, der erreichten
Biomassekonzentrationen oder der Expressionsraten zu beobachten gewesen wäre. Daraus
ergeben sich erhebliche Vorteile: Erstens sind die Komponenten des Fermentationsmediums "A"
(2.5.5.1) bei pH 6 nicht vollständig gelöst und führen dadurch zu Fehlern in der
Frischgewichtsbestimmung zu Beginn der Fermentation und zu möglicher Beschädigung der
Fermenteroberflächen durch abrasive Effekte. Zweitens fielen beim Einstellen des pH-Werts im
Überstand auf leicht basische Werte für die Aufarbeitung stets erhebliche Mengen an nicht
metabolisierten Salzen aus. Diese Effekte wurden mit dem Fermentationsmedium "B" nicht

Abschlußdiskussion und Ausblick
87
beobachtet. Abgesehen von den positiven Auswirkungen auf die Fermentation und die
Aufbereitung stellt die Reduktion der Salzmenge einen ökonomischen Vorteil dar, der zwar im
Pilotmaßstab kaum ins Gewicht fällt, im industriellen Maßstab aber durchaus Bedeutung hat. Die
Ausgangs-Glyzerinkonzentration wurde auf 5 % (v/v) erhöht, wodurch sich keine negativen
Einflüsse auf die Wachstumsgeschwindigkeit ergaben (siehe auch 4.2.1.2).
4.2.1.2 Glyzerin-Zufuhrphase
Im "Standard"-Fermentationsprotokoll ist nach dem Verbrauch des vorgelegten Glyzerins
eine Glyzerin-Zufuhrphase vorgesehen. Damit wird beabsichtigt, die zur Induktion erwünschten
hohen Zelldichten zügig zu erreichen, ohne die Wachstumsrate durch eventuelle
Substratinhibierung infolge zu hoher initiale Glyzerinkonzentrationen zu beeinträchtigen. Dabei
soll die Glyzerin-Zufuhrrate so niedrig sein, daß Glyzerin limitierend ist, da unter diesen
Umständen möglicherweise der AOX1-Promotor dereprimiert wird und die Adaption an
Methanol schneller erfolgt (Stratton et al., 1998). Weder die Derepression des AOX1-Promotors
(etwa durch im Western-Blot oder ELISA nachweisbare basale Expression von rekombinanten
Proteinen während einer Phase limitierender Glyzerinzufuhr) noch die schnellere Anpassung an
Methanol konnten in entsprechenden Versuchen festgestellt werden. Diese Ergebnisse
entsprechen den Modellen, nach denen die Regulation des AOX1-Promotors sowohl Repression
durch leichter verfügbare Kohlenstoffquellen als auch Induktion durch Methanol involviert (Ellis
et al., 1985; Tschopp et al., 1987; Waterham et al., 1997). In diesem Punkt unterscheidet sich die
Regulation des AOX1 von Pichia pastoris entscheidend von der Regulation des MOX-Promotor
der methylotrophen Hefe Hansenula polymorpha, der tatsächlich die Expression unter
limitierender Glyzerinzufuhr ohne Methanol erlaubt (Gellissen et al., 1995; Hollenberg und
Gellissen, 1997). Die Adaption an Methanol war, gemessen an der Zeit, die im Rührkessel vom
Beginn der Methanol-Zufuhr bis zum Erreichen der maximalen Methanol-Verbrauchsrate
notwendig war, unabhängig davon, ob der Methanol-Zufuhrphase eine Phase mit limitierender
Glyzerinzufuhr vorgeschaltet war. Die zu Beginn der Induktion erwünschten Zelldichten von ca.
180 g·l-1 wurden bei den im Rahmen dieser Arbeit durchgeführten Fermentationen dadurch
erreicht, daß die initiale Glyzerinkonzentration von 4 % (v/v) auf 5 % erhöht wurde. Diese
Ausgangs-Glyzerinkonzentration wurde aus ähnlichen Gründen auch von anderen Gruppen
verwendet (Digan et al., 1989; Clare und Romanos, 1995; Loewen et al., 1997). Dazu kommt,
daß die verwendeten Mut+-Stämme auch auf Methanol als einziger Kohlenstoffquelle während
der Induktion noch Wachstumsraten bis 0,02 h-1 aufwiesen und somit die Biomasse auch
während der Induktion signifikant zunahm.

Abschlußdiskussion und Ausblick
88
4.2.1.3 Methanol-Zufuhrphase
Die maximale Methanol-Zufuhrrate lag mit 4-5 ml·l-1·h-1 deutlich unter den von anderen
Autoren beschriebenen Werten, die mit bis zu 15 ml·l-1·h-1 angegeben werden (Nilsen et al.,
1997). Die Obergrenze von 4-5 ml·l-1·h-1 hatte sich bereits in den Fermentationen mit manuell
eingestellter, fester Zufuhrrate ergeben, bei denen höhere Zufuhrraten zur Akkumulation von
Methanol und letztlich zum Absterben der Zellen führten (Daten nicht gezeigt), und bestätigte
sich bei den Fermentationen, in denen die Methanolzufuhr über den Gelöstsauerstoff oder on-
line Methanolanalyse gesteuert wurde.
Für die Differenz gibt es mehrere mögliche Gründe: Eine mögliche Erklärung wären
unterschiedliche Sauerstoffübergangsraten in den verschiedenen Rührreaktoren. Aufgrund der
hohen Zelldichten wurden die Fermentationen während der Induktionsphase – trotz des relativ
langsamen Wachstums auf Methanol – unter Sauerstofflimitierung oder nahe daran geführt. Da
die Verwertung von Methanol stöchiometrisch an die Sauerstoffverfügbarkeit gekoppelt ist,
würde eine niedrigere Sauerstoffübergangsrate unmittelbar zu niedrigerer Methanol-
Verwertungsrate und Wachstumsrate führen. Gegen diese Möglichkeit sprechen mehrere Punkte.
Zum einen waren die Eckdaten der Prozessführung (Durchmischung mit 3 Rushton-
Scheibenblattrührern bei hohen Umdrehungszahlen und Belüftungsraten zwischen 1,5 und 2,3
vvm) mit den Daten anderer Autoren vergleichbar. Zum anderen wurden Fermentationen
durchgeführt, in denen die Sauerstoffübergangsrate durch zusätzliche Zufuhr von reinem
Sauerstoff zum Begasungsstrom erhöht wurde (z. B. Abb. 3-19, 46 h - 64 h). Damit konnten
Gelöstsauerstoffkonzentration von ca. 15-20 % erreicht werden, also Werte die oberhalb des
dO2krit liegen (normalerweise schwankte der dO2 während der Fermentation zwischen 1 % und
5 %, die Sauerstoffübergangsrate war also fast maximal). Obwohl diese Versuche mehrfach
wiederholt wurden, waren keine damit verbundenen Steigerungen der Wachstumsrate oder des
Methanolverbrauchs erkennbar, ein Zeichen dafür, daß die Sauerstoffübergangsrate auch ohne
Zusatz von reinem Sauerstoff zum Begasungsstrom knapp oberhalb der maximalen
Sauerstoffaufnahmerate lag. Damit bleiben als Erklärung entweder unterschiedliche Mengen an
mit der Abluft ausgetragenen Methanols infolge anderer Reaktorgeometrien oder
Bestimmungsfehler bezüglich der Methanol-Verbrauchsraten.
Die wesentlichste Verbesserung der Fermentationsstrategie stellte die Implementierung der
on-line Methanolsteuerung dar. Die Vorteile liegen in der gleichmäßigen Methanolkonzentration
von 0,5 % (Abb. 3-19) im Vergleich zu schwankenden Methanolkonzentrationen bei manueller
Steuerung (Abb. 3-16) bzw. nahe bei 0 liegenden Methanolkonzentrationen bei dO2-abhängiger
Steuerung (Abb. 3-23). Eine gleichmäßige Methanolkonzentration schafft optimale

Abschlußdiskussion und Ausblick
89
Wachstumsbedingungen und führt zu verbesserten Expressionsergebnissen (Stratton et al.,
1998). Daneben ermöglicht die on-line Methanolkontrolle eine vereinfachte und sicherere
Fermentationsführung, da die Gefahr einer Über- oder Unterdosierung mit den entsprechenden
Folgen nicht besteht, weil die Zufuhrrate sich automatisch dem Methanolverbrauch anpaßt. Im
Rahmen dieser Arbeit wurde die von der Fa. Heinrich Frings GmbH & Co KG (Bonn) für die
Ethanolmessung in Essigsäurefermentation entwickelte und in dieser Branche im industriellen
Maßstab etablierte Technologie erstmalig im Zusammenhang mit der Methanol-Zufuhrsteuerung
in Fermentation von Pichia pastoris eingesetzt. Zum Einsatz kamen dabei die Sonde
ALKOSENS II sowie ein Prototyp mit veränderter Geometrie, beide in Verbindung mit der
ACETOMAT Methanolsteuerung. Die direkte Messung im Medium durch eine
Pervaporationsmembran (Abb. 2-3) erlaubt die Veränderung der Belüftungsrate während der
Fermentation, was bei im Abluftstrom messenden Sonden, wie der von Katakura et al.
verwendeten, nicht möglich ist (Katakura et al., 1998). Das gemessene Signal stellt einen
Summenparameter dar, der alle membrangängigen, oxidierbaren Substanzen widerspiegelt, die
von der Spülluft zum Halbleitersensor transportiert werden. In dem verwendeten Mineralmedium
waren keine störenden Substanzen enthalten. Da Pichia pastoris nicht oder nur in minimalem
Maßstab zur Gärung befähigt ist, bestand auch nicht die Gefahr, die Messung durch biogenes
Ethanol zu verfälschen (Higgins und Cregg, 1998). Die verwendete Membran schloß darüber
hinaus die Pervaporation eventueller organischer Säuren aufgrund ihrer Selektivität aus (Dr.
Frank Emde, Frings, pers. Mitteilung). Die in Abb. 3-22 und Abb. 3-24 gezeigte Fermentations-
strategie ist – soweit diese publiziert sind – anderen Pichia-Fermentationsstrategien in puncto
Betriebssicherheit, Dokumentierbarkeit und Reproduzierbarkeit überlegen.
Die durch die Implementierung der Methanol-Analytik verfügbare Technologie wurde
genutzt, um die gemischte Zufuhr von Glyzerin und Methanol während der Induktion zu
untersuchen. Diese Strategie ist in einigen Arbeiten als vorteilhaft im Hinblick auf die
volumetrische Produktivität beschrieben worden (Sreekrishna et al., 1989; Loewen et al., 1997;
Sreekrishna et al., 1997; Katakura et al., 1998). In diesen Untersuchungen wurden Methanol-
und Glyzerinzufuhr nicht voneinander entkoppelt, d. h. die beiden Substrate wurden in
verschiedenen Verhältnissen gemischt und die Mischung wurde mit manuell gesteuerten Raten
zugeführt. Im Gegensatz dazu wurde die Zufuhr der beiden Kohlenstoffquellen in dieser Arbeit
erstmals völlig entkoppelt, indem der über die ALKOSENS II-Sonde gesteuerten konstanten
Methanolkonzentration eine manuell gesteuerte Glyzerinzufuhr überlagert wurde. Dadurch
wurde die Möglichkeit einer präzisen Untersuchung des Einflusses einer zusätzlichen
Glyzerinverfügbarkeit während der Induktionsphase geschaffen.

Abschlußdiskussion und Ausblick
90
Es konnte gezeigt werden, daß selbst geringste spezifische Glyzerin-Zufuhrraten, die weit
unter der maximalen Glyzerin-Aufnahmerate lagen, sich negativ auf die Induktion des AOX1-
Promotors auswirkten. Dieser Effekt wurde an der drastisch verringerten Konzentration des
scFv4813 im Überstand deutlich, aber auch daran, daß die Wachstumsrate bei sehr niedrigen
Glyzerin-Zufuhrraten noch niedriger war, als die Wachstumsrate mit einer konstanten
Konzentration von 0,5 % Methanol als einziger Kohlenstoffquelle (vgl. Tabelle 3-2). Der
letztgenannte Effekt kann nur auf eine Inhibierung der Expression der Alkoholoxidase 1
zurückzuführen sein.
Diese Folgerung kann verallgemeinert werden. Dennoch muß der beobachtete deutliche
Effekt auf die spezifische (und in diesem Fall auch volumetrische) Produktivität nicht bei allen
Klonen und in allen Bioreaktoren in dieser Ausprägung auftreten. Im Falle von MutS- Stämmen
oder bei aus anderen Gründen, wie z. B. Sauerstofflimitierung, nicht optimal laufenden
Prozessen kann der Effekt der zusätzlichen Glyzerinversorgung sich auf die volumetrische
Produktivität unter Umständen positiv auswirken, nämlich dann, wenn der Einfluß anderer
limitierender Faktoren von der Zugänglichkeit einer leichter verwertbaren Kohlenstoffquelle wie
Glyzerin überlagert wird.
Die Frage, ob die Expression über den AOX1-Promotor durch Glyzerin inhibiert wird oder
nicht, ist in den letzten Jahren in verschiedenen Veröffentlichungen gegensätzlich beantwortet
worden. Während einige Arbeiten ganz klar von einer Repression des AOX1 Promotors durch
Glyzerin sprechen (Buckholz und Gleeson, 1991; Hollenberg und Gellissen, 1997; Lal et al.,
1997; Sreekrishna et al., 1997), gehen andere vom genauen Gegenteil aus (Loewen et al., 1997;
Katakura et al., 1998), darauf beruht schließlich das Konzept des "mixed feed".
Interessanterweise gibt es auch Ergebnisse, die sogar bei gemischter Zufuhr von Glucose und
Methanol unter bestimmten Umständen vorteilhafte Expressionsergebnisse zeigen (pers.
Mitteilung, R. Takors., Forschungszentrum Jülich, IBT-2)
Unabhängig von solchen möglicherweise spezifischen Effekten ist zu erwarten, daß sich die
Fermentation von Pichia mit Hilfe der on-line Methanol-Analytik zumindest im professionellen
Bereich durchsetzen wird.
4.2.1.4 Vergleich mit anderen in Pichia pastoris exprimierten scFvs
Die bei der Produktion des scFv4813 im Fermenter erreichten Expressionslevel von ca.
40 mg·l-1 (3.4.2) liegen im Vergleich mit anderen, in Pichia exprimierten single-chain
Antikörpern etwa im Mittelfeld. Die von Freyre et al. mit einem sekretierten scFv erreichten
1,2 g·l-1 (Freyre et al., 2000) stellen unter den bisher publizierten (Fischer et al., 1999b; Eldin et

Abschlußdiskussion und Ausblick
91
al., 1997; Gram et al., 1998) Arbeiten zur Produktion solcher Fragmente in Pichia den höchsten
Wert dar (Vgl. Tabelle 1-7). Daneben kann von einer Reihe nicht publizierter Ergebnisse (pers.
Mitteilung, Dr. P. van der Logt, Unilever, Bedford, GB bzw. J. Drossard, RWTH Aachen)
abgeleitet werden, daß bei einer Reihe von Versuchen, sekretierte scFvs in Pichia zu
produzieren, Expressionslevel im Bereich von 1-5 mg·l-1 erreicht wurden. Die Verwendung des
Pichia-Expressionssystems stellt also keine Garantie für die erfolgreiche Überexpression eines
Proteins im größeren Maßstab dar. Selbst innerhalb strukturell relativ ähnlicher Proteine
entscheiden neben der erfolgreichen Selektion eines "guten" Expressorklons bisher nicht näher
bekannte, proteinspezifische Faktoren über den Erfolg oder Mißerfolg der Produktion in Pichia.
4.3 Reinigung und Lagerung des in Pichia produzierten scFv4813
Bei der Reinigung des scFv4813 aus dem Fermentationsüberstand von Pichia pastoris erwies
sich die Ni-IDA Affinitätschromatographie in Verbindung mit der "expanded bed"-Technologie
für den ersten Reinigungsschritt als sehr geeignet. Ausschlaggebend dafür war zum ersten die
Möglichkeit, den nach Entfernung von ca. 99,9 % der Zellen noch relativ trüben Überstand über
die Säule zu geben. Eine Filtration zur vollständigen Entfernung der Zellen, die erheblichen
zeitlichen und materiellen Aufwand bedeutet hätte (s. u.) war nicht notwendig. Zum zweiten
erlaubte der durch den Charakter der Affinitätschromatographie gegebene hohe
Aufkonzentrierungsfaktor die weiteren Schritte mit einem um den Faktor 50 reduzierten
Volumen. Eine dem Reaktorvolumen angepaßte, größere Chromatographiesäule hätte eine
insgesamt höhere Ausbeute ermöglicht (3.5.2) und die Prozessierungszeit verkürzt.
Der ungewöhnlich hohe theoretische isoelektrische Punkt des Proteins von 8,77 erlaubte die
weitere Reinigung über eine Kationenaustauscherchromatographie bei pH 5,0, wobei die in der
Ni-IDA Affinitätschromatographie im Eluat enthaltenen kontaminierenden 80, 60 und 45-50 kDa
Proteine entfernt wurden (vgl. Abb. 3-27 und Abb. 3-28). Der abschließende Gelfiltrationsschritt
diente der Entfernung eventueller Dimere des scFv. Die Reinigungseffizienz läßt sich aufgrund
der nicht exakt bestimmten Konzentration des scFv4813 im Fermentationsüberstand nur relativ
grob abschätzen. Der Wert von 54 % (3.5.5) ist angesichts der zu klein dimensionierten Säule in
der Affinitätschromatographie und der hohen Reinheit des Produkts nach einer 3-Schritt-
Prozedur, die vor der Ausbeutemaximierung Vorrang hatte, akzeptabel. Das Ergebnis ließe sich
aber durch Verwendung größerer Säulen noch deutlich verbessern.
Die Stabilität des gereinigten scFv4813 ist als sehr gut einzustufen, da selbst ohne Zusatz von
Proteaseinhibitoren oder Natriumazid sogar bei 4° C innerhalb von 3 Wochen keine

Abschlußdiskussion und Ausblick
92
nennenswerten proteolytischen Effekte oder Aktivitätsverluste (bezogen auf die Funktionalität in
der Ni-NTA Oberflächenplasmonresonanzanalyse) festzustellen waren.
Die Reinheit und Qualität des in dieser Weise gewonnen scFv4813 war ausreichend zur
Kristallisierung des Proteins (pers. Mitteilung, Dr. Kurt Hoffmann, RWTH Aachen, Inst. f.
Biologie I). Diese Arbeit stellt daher neben anderen (O'Donohue et al., 1996; Kim et al., 1997;
Linnevers et al., 1997; Zhan et al., 1999) ein weiteres Beispiel für die Eignung des Pichia-
Expressionssystems zur Produktion von rekombinanten Proteinen zur Strukturanalyse dar.
4.4 Vergleich der Expressionssysteme unter Einbeziehung ökonomischerGesichtspunkte
Die Expression des diagnostisch relevanten scFv4813 durch Fermentation ist, wie gezeigt,
sowohl in Nicotiana tabacum BY-2 als auch in Pichia pastoris möglich. Im vorliegenden Fall
wurde das Pichia Expressionssystem gewählt, weil das Protein in diesem System in relativ
hohen Konzentrationen ins Medium sekretiert wurde. Im BY-2 System wäre durch Verwendung
entsprechender Sekretionssignale anstelle des KDEL-Retentionssignals auch eine sekretierte
Expression möglich gewesen, aber aufgrund der Erfahrung, daß die intrazelluläre Expression
höhere Expressionsraten hervorbringt (Firek et al., 1993; Fischer et al., 1999a; Doran, 2000),
war dieser Weg gewählt worden. Natürlich wäre die Reinigung des scFv 4813 auch aus
Pflanzenzellextrakten möglich gewesen, die guten Ergebnisse in Pichia machten diesen deutlich
aufwendigeren Prozeß aber unnötig.
Tabelle 4-1 zeigt eine Gegenüberstellung der beiden Expressionssysteme anhand der
Eckdaten einer 30-l Fermentation. Dabei wird deutlich, daß die bessere Eignung des Pichia-
Systems nicht uneingeschränkt gültig ist
Tabelle 4-1: Gegenüberstellung der beiden Expressionssysteme
Pichia pastoris Nicotiana tabacum cv BY-2
Medium preiswert teuer*
Fermentationsdauer 4 Tage 7-8 Tage
Technischer Aufwand hoch niedrig
Personeller Aufwand hoch niedrig
Expressionslevel** max. 30 mg·l-1 max. 11mg·l-1
Volumetrische Produktivität 7,5 mg·l-1·d-1 1,6 mg·l-1·d-1
Lokalisierung sekretiert intrazellulär
Reinigung einfach schwierig*Verwendung von Fertigmedium **bezogen auf Fermentationsvolumen

Abschlußdiskussion und Ausblick
93
Das für die Pflanzenzellkultur verwendete Fertigmedium in Pulverform schlägt sich im 30-l
Maßstab deutlich im Preis nieder, bei einer Fermentation im Großmaßstab würde die Herstellung
dieses Mediums aus den Einzelkomponenten aber effektiver und der Preis würde dem des
Pichia-Mediums näher kommen. Trotz der kürzeren Fermentationsdauer erfordert die
Fermentation von Pichia pastoris aufgrund der aufwendigen Prozessführung (pH-Regulation,
Schaumkontrolle, Zufuhrverfahren, Methanolmessung) einen erheblich höheren personellen und
materiellen Aufwand als die verfahrenstechnisch denkbar einfach gestalteten Fermentationen
von Nicotiana tabacum cv BY-2. Aus diesem Grund liegen die für eine Pflanzenzellkultur zu
kalkulierenden Arbeitskosten und Geräteabschreibungen deutlich niedriger als die Kosten einer
Pichia-Fermentation.
Die Produktion des scFv4813 in Pichia zeigte einen um den Faktor 3 höheren
Expressionslevel sowie eine um den Faktor 4,7 höhere volumetrische Produktivität als die
Produktion in Pflanzensuspensionskultur. Diese beiden Werte sind aber stark von
proteinspezifischen Faktoren abhängig, so daß sie nicht generalisiert werden können. Die
Entwicklung stärkerer Promotoren für die Pflanzensuspensionskultur läßt für dieses System noch
Perspektiven nach oben zu, während die Machbarkeit noch stärkerer Promotoren für das
Hefesystem unwahrscheinlich erscheint.
Die erfolgreiche Optimierung der Taxol-Produktion in Suspensionskulturen von Taxus spec.
durch die Fa. Phyton (pers. Mitteilung, Dr. Rainer Fischer) zeigt, daß Pflanzensuspensions-
kulturen durch Medien- und Stammoptimierung durchaus um den Faktor 100 in ihrer
Produktivität gesteigert werden können. Obwohl sich dieses Ergebnis auf die Produktion eines
Sekundärmetaboliten bezieht, zeigt es auch für die Produktion rekombinanter Proteine in
Pflanzensuspensionskulturen Perspektiven auf.
Für Proteine, deren Produktion in mikrobiellen Wirtsorganismen nicht möglich ist, weil diese
z. B. essentielle Faltungsschritte oder posttranslationale Modifikationen nicht korrekt ausführen
können, bietet sich die Pflanzensuspensionskultur als Alternative zu tierischen Zellkulturen an.
Im wirtschaftlichen Vergleich dieser beiden Systeme schneidet die Pflanzensuspensionskultur
deutlich besser ab, weil zum einen die Kultivierungskosten niedriger sind, zum anderen z. B. für
die Produktion von Therapeutika der Nachweis des Nichtvorhandenseins von Humanpathogenen
und Onkogenen einfacher und kostengünstiger ausfällt (Fischer et al., 1999a; Doran, 2000), .
Auch gegenüber der Expression in intakten Pflanzen weist die Pflanzensuspensionskultur
bestimmte Vorteile auf (Fischer et al., 1999a; Fischer et al., 1999d). Intakte Pflanzen sind
insbesondere wegen der Möglichkeit, im Feldanbau sehr große Mengen an Biomasse zu äußerst

Abschlußdiskussion und Ausblick
94
niedrigen Kosten zu erzeugen, attraktiv. Die Nachteile dieses Systems liegen aber in der
kostenintensiven Reinigung rekombinanter Produkte aus dem Pflanzenmaterial. Für den Fall des
rekombinanten Avidins aus Mais (Hood et al., 1999) machen die Aufarbeitungskosten 94 % der
Gesamtkosten aus (Doran, 2000). Bei Pflanzensuspensionskulturen sind die Aufarbeitungskosten
grundsätzlich niedriger, weil das Ausgangsmaterial homogener ist und viele bei intakten
Pflanzen vorhandene störende Begleitstoffe (Wachse, Öle, Chlorophyll, Alkaloide) nicht oder
nur in geringerem Umfang vorhanden sind. Dazu kommen regulatorische Aspekte, die bei der
Produktion therapeutisch wirksamer Proteine eine Rolle spielen: Lückenlose Dokumentation der
Wachstumsphasen ist im Feldanbau ebensowenig denkbar wie exakt reproduzierbare
Bedingungen während mehrwöchiger Wachstumsphasen. Demgegenüber sind die Bedingungen
bei der Pflanzensuspensionskultur sehr leicht reproduzierbar und dokumentierbar, was der
Produktion nach GMP entgegenkommt. Dazu kommt die Unabhängigkeit der
Suspensionskulturen von klimatischen Einflüssen und Vegetationszeiten sowie die Möglichkeit,
Biomasse vergleichsweise schnell zu produzieren. Pflanzliche Suspensionskulturen bieten also
insbesondere für die Produktion komplexer Proteine, die in kleinen bis mittleren Mengen für
therapeutische Zwecke benötigt werden, besondere Vorteile.
Die relativ niedrigen Expressionslevel in Pflanzensuspensionskulturen machen trotz der
genannten Vorteile die Weiterentwicklung dieses Expressionssystems erforderlich. Auch in
diesem Fall wird allerdings die Frage, welches Expressionssystem für ein bestimmtes Protein das
Mittel der Wahl ist, von proteinspezifischen Eigenschaften abhängen, und möglicherweise nur
empirisch zu klären sein. Diese Entscheidung fiel im vorliegenden Fall auf der Basis der mit dem
Pichia-System erzielten Expressionslevel, der benötigten Menge des rekombinanten Proteins
und der deutlich leichteren Reinigung des rekombinanten Produkts aus dem Pichia-
Fermentationsüberstandes für das mikrobielle System.

Zusammenfassung
95
5 Zusammenfassung
Ziel dieser Arbeit war es, im Rahmen des EU Projektes "Processing technology for recovery
of recombinant antibody produced in crop plants" (FAIR-PL-CT96-1039) etwa 500 mg des
rekombinanten scFv4813 zur Verfügung zu stellen. Das Projekt beschäftigte sich mit der
Entwicklung von affinitätschromatographischen Reinigungsstrategien für in pflanzlichen
Systemen produzierte Antikörperfragmente. Als immobilisierte Liganden sollten synthetische
Peptide dienen, die aus der Aminosäuresequenz des Antigens (im Falle des scFv4813
follikelstimulierendes Hormon, FSH) oder aus synthetischen Peptidbibliotheken gewonnen
werden sollten. Der Antikörper sollte entweder direkt in Pflanzensuspensionskulturen oder in
einem mikrobiellen Wirt produziert werden.
Die pflanzliche Zellinie Nicotiana tabacum cv BY-2 war am Institut für Biologie I
einschließlich der notwendigen Techniken zur Transformation und Selektion schon seit
mehreren Jahren etabliert. Als mikrobieller Wirt wurde zu Beginn des Projekts noch relativ neue
Pichia pastoris Expressionssystem gewählt, mit dem bei einem der Kooperationspartner gute
Erfahrungen gemacht worden waren.
Schon während die notwendigen Klonierungsarbeiten im Vorfeld der Expression des
scFv4813 in Tabak und Pflanzenzellen (die nicht zum Umfang dieser Arbeit gehören) anliefen,
wurde anhand einer bereits bestehenden transgenen Zellinie von Nicotiana tabacum cv BY-2
(biscFv2429) sowie eines transgenen Pichia-Stammes (PP599) Fermentationsstrategien für die
beiden unterschiedlichen Expressionssysteme entwickelt. Im Fall der Pflanzensuspensions-
kulturen umfaßte dies die Untersuchung der Fermentationsbedingungen, der Wachstumskinetik,
Substrataufnahmekinetik und Versuche zur Stimulation der Expression. Die Ergebnisse
mündeten in der erfolgreichen Maßstabsvergrößerung in den 30-l Maßstab und die Produktion
des scFv4813 diesem System. Parallel dazu wurden Fermentationsstrategien für Pichia pastoris
entwickelt, wobei sich die Arbeit hier nach der Untersuchung grundlegender Parameter auf die
Entwicklung optimaler Zufuhrstrategien für Methanol während der im "fed-batch" Modus
erfolgenden Induktionsphase konzentrierte. Auch das Pichia-System wurde bis in den 30-l
Maßstab vergrößert.
Initiale 30-l Fermentationen zur Produktion des scFv4813 in beiden Systemen (Zellinie
R1-25 bzw. Pichia-Klon N55) führte zu der Entscheidung, dem mikrobiellen System den
Vorrang zu geben, da hier der scFv4813 in sekretierter Form vorlag und verhältnismäßig leicht

Zusammenfassung
96
zu reinigen war. Zudem waren die Expressionslevel deutlich höher als in der
Pflanzensuspensionskultur.
Nach der Entwicklung einer Reinigungsstrategie (nicht Bestandteil dieser Arbeit) wurden
annähernd 500 mg des scFv4813 aus einer 30-l Fermentation von Pichia pastoris mit Hilfe eines
3-Schritt-Verfahrens gereinigt. Die Konzentration und Stabilität des Produkts wurden bestimmt.
Die Reinheit des Produktes war ausreichend zu dessen Kristallisation.

Literatur
97
6 Literatur
Adams, G. P., Schier, R., McCall, A. M., Crawford, R. S., Wolf, E. J., Weiner, L. M., Marks, J. D. (1998).
Prolonged in vivo tumour retention of a human diabody targeting the extracellular domain of human
HER2/neu. Br J Cancer, 77, 1405-1412.
Altmann, F., Staudacher, E., Wilson, I. B., Marz, L. (1999). Insect cells as hosts for the expression of
recombinant glycoproteins. Glycoconj J, 16, 109-123.
Arakawa, T., Yu, J., Langridge, W. H. (1999). Food plant-delivered cholera toxin B subunit for vaccination
and immunotolerization. Adv Exp Med Biol, 464, 161-178.
Arndt, M. A., Krauss, J., Kipriyanov, S. M., Pfreundschuh, M., Little, M. (1999). A bispecific diabody that
mediates natural killer cell cytotoxicity against xenotransplantated human Hodgkin's tumors. Blood, 94,
2562-2568.
Artsaenko, O., Peisker, M., Nieden, U., Fiedler, U., Weiler, E. W., Küntz, K., Conrad, U. (1995).
Expression of a single-chain Fv antibody against abscisic acid creates a wilty phenotype in transgenic
tobacco. Plant J, 8, 745-750.
Atwell, J. L., Breheney, K. A., Lawrence, L. J., McCoy, A. J., Kortt, A. A., Hudson, P. J. (1999). scFv
multimers of the anti-neuraminidase antibody NC10: length of the linker between VH and VL domains
dictates precisely the transition between diabodies and triabodies. Protein Eng, 12, 597-604.
Ausubel, F. M., Brent, R., Kingstone, R. E. (1994). Current Protocols in Molecular Biology New York:
Wiley Interscience
Baneyx, F. (1999). Recombinant protein expression in Escherichia coli. Curr Opin Biotechnol, 10, 411-
421.
Benvenuto, E., Ordas, R. J., Tavazza, R., Ancora, G., Biocca, S., Cattaneo, A., Galeffi, P. (1991).
'Phytoantibodies': a general vector for the expression of immunoglobulin domains in transgenic plants.
Plant Mol Biol, 17, 865-874.
Bird, R. E., Hardman, K. D., Jacobson, J. W., Johnson, S., Kaufman, B. M., Lee, S. M., Lee, T., Pope, S.
H., Riordan, G. S., Whitlow, M. (1988). Single-chain antigen-binding proteins. Science, 242, 423-426.
Boleti, E., Deonarain, M. P., Spooner, R. A., Smith, A. J., Epenetos, A. A., George, A. J. (1995).
Construction, expression and characterisation of a single chain anti- tumour antibody (scFv)-IL-2 fusion
protein. Ann Oncol, 6, 945-947.
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of
protein utilizing the principle of protein-dye binding. Anal Biochem, 72, 248-254.

Literatur
98
Brankamp, R. G., Sreekrishna, K., Smith, P. L., Blankenship, D. T., Cardin, A. D. (1995). Expression of a
synthetic gene encoding the anticoagulant- antimetastatic protein ghilanten by the methylotropic yeast
Pichia pastoris. Protein Expr Purif, 6, 813-820.
Buckholz, R. G., Gleeson, M. A. G. (1991). Yeast systems for the commercial production of heterologous
proteins. BIO/TECHNOLOGY, 9, 1067-1072.
Burnette, W. N. (1981). Electrophoretic transfer of proteins from sodium dodecyl sulfate-polyacrylamide
gels to unmodified nitrocellulose and radiographic detection with antibody and radioionidated protein.
Anal Biochem, 112, 195-203.
Byun, S. Y., Pedersen, H. (1994). Two-phase airlift fermentor operation with elicitation for the enhanced
production of benzophenanthridine alkaloids in cell suspensions of Eschcholtzia californica. Biotechnol
Bioeng, 44, 14-20.
Clare, J., Scorer, C., Buckholz, R., Romanos, M. (1998). Expression of EGF and HIV Envelope
Glycoprotein. In: Pichia Protocols, D. R. Higgins, J. M. Cregg, eds. Humana Press, Totowa, NJ, 209-225.
Clare, J. J., Rayment, F. B., Ballantine, S. P., Sreekrishna, K., Romanos, M. A. (1991a). High-level
expression of tetanus toxin fragment C in Pichia pastoris strains containing multiple tandem integrations
of the gene. BIO/TECHNOLOGY, 9, 455-460.
Clare, J. J., Romanos, M. A. (1995). Expression of cloned genes in the yeasts Saccharomyces cerevisiae
and Pichia pastoris. Methods Mol Cell Biol, 5, 319-329.
Clare, J. J., Romanos, M. A., Rayment, F. B., Rowedder, J. E., Smith, M. A., Payne, M. M., Sreekrishna,
K., Henwood, C. A. (1991b). Production of mouse epidermal growth factor in yeast: high-level secretion
using Pichia pastoris strains containing multiple gene copies. Gene, 105, 205-212.
Clark, M. F., Adams, A. N. (1977). Characteristics of the microplate method of Enzyme-linked
Immunosorbent Assay for the detection of plant viruses. J Gen Virol., 34, 475-483.
Colcher, D., Bird, R., Roselli, M., Hardman, K. D., Johnson, S., Pope, S., Dodd, S. W., Pantoliano, M. W.,
Milenic, D. E., Schlom, J. (1990). In vivo tumor targeting of a recombinant single-chain antigen-binding
protein. J Natl Cancer Inst, 82, 1191-1197.
Couderc, R., Baratti, J. (1980). Oxidation of methanol by the yeast Pichia pastoris: purification and
properties of alcohol oxidase. Agric Biol Chem, 44, 2279-2289.
Cramer, C. L., Boothe, J. G., Oishi, K. K. (1999). Transgenic plants for therapeutic proteins: linking
upstream and downstream strategies. Curr Top Microbiol Immunol, 240, 95-118.
Cregg, J. M., Madden, K. R., Barringer, K. J., Thill, G. P., Stillman, C. A. (1989). Functional
characterization of the two alcohol oxidase genes from the yeast Pichia pastoris. Mol Cell Biol, 9, 1316-
1323.

Literatur
99
Cregg, J. M., Tschopp, J. F., Stillman, C., Siegel, R., Akong, W., Craig, W. S., Buckholz, R. G., Madden,
K. R., Kellaris, P. A., Davis, G. R., Smiley, B. L. et al. (1987). High-level expression and efficient
assembly of hepatitis B surface antigen in the methylotrophic yeast, Pichia pastoris. BIO/TECHNOLOGY,
5, 479-485.
De Neve, M., De Buck, S., De Wilde, C., Van Houdt, H., Strobbe, I., Jacobs, A., Van, M., Depicker, A.
(1999). Gene silencing results in instability of antibody production in transgenic plants. Mol Gen Genet,
260, 582-592.
De Wilde, C., De Rycke, R., Beeckman, T., De Neve, M., Van Montagu, M., Engler, G., Depicker, A.
(1998). Accumulation pattern of IgG antibodies and Fab fragments in transgenic Arabidopsis thaliana
plants. Plant Cell Physiol, 39, 639-646.
Denbow, D. M., Grabau, E. A., Lacy, G. H., Kornegay, E. T., Russell, D. R., Umbeck, P. F. (1998).
Soybeans transformed with a fungal phytase gene improve phosphorus availability for broilers. Poult Sci,
77, 878-881.
Denecke, J., de Rycke, R., Bottermann, J. (1992). Plant and mammalian sorting signals for protein
retention in the endoplasmic reticulum contain a conserved epitope. EMBO J, 11, 2345-2355.
Denecke, J., Goldman, M. H. S., Demolder, J., Seurinck, J., Botterman, J. (1991). The tobacco luminal
binding protein is encoded by a multigen family. Plant Cell, 3, 1025-1035.
des Molles, D. V., Gomord, V., Bastin, M., Faye, L., Courtois, D. (1999). Experession of a carrot invertase
gene in tobacco suspension cells cultivated in batch and continuous culture conditions. J Biosci Bioeng,
87.
Digan, M. E., Lair, S. V., Brierley, R. A., Siegel, R. S., Williams, M. E., Ellis, S. B., Kellaris, P. A., Provow,
S. A., Craig, W. S., Velicelebi, G., Harpold, M. M. et al. (1989). Continuous production of a novel
lysozyme via secretion from the yeast, Pichia pastoris. BIO/TECHNOLOGY, 7, 160-164.
Dombrowski, J. E., Raikhel, N. V. (1996). Protein targeting to the plant vacuole - a historical perspective.
Braz J Med Biol Res, 29, 413-430.
Doran, P. (1993). Design of reactors for plant cells and organs. In: Bioprocess design and control, A.
Fiechter, ed. Springer, Berlin, Heidelberg, New York, 115-168.
Doran, P. M. (2000). Foreign protein production in plant tissue cultures. Curr Opin Biotechnol, 11, 199-
204.
Dörnenburg, H., Knorr, D. (1995). Strategies for the improvement of secondary metabolite production in
plant cell cultures. Enz Microb Technol, 17, 674-684.
Drapeau, D., Blanch, H. W., Wilke, C. R. (1986). Growth kinetics of Dioscorea deltoidea and
Catharanthus roseus in batch culture. Biotechnol Bioeng, 28, 1555-1563.

Literatur
100
Drossard, J. (1996). Expression und Reinigung von rekombinanten Antikörpern aus Pflanzenzellkulturen.
Diplomarbeit; RWTH Aachen, Institut für Biologie I, .
Drossard, J., Nähring, J. M., Hellwig, S., Fischer, R. (1997). Production of engineered antibodies in
tobacco plants and cell suspension cultures. In: Antibody engineering, New technology, Application and
Commercialization IBC library services, 69-93.
Ducos, J. P., Pareilleux, A. (1986). Effect of aeration rate and influence of pCO2 in large-scale cultures of
Catharanthus roseus cells. Appl Microbiol Biotechnol, 25, 101-105.
Düring, K., Hippe, S., Kreuzaler, F., Schell, J. (1990). Synthesis and self-assembly of a functional
monoclonal antibody in transgenic Nicotiana tabacum. Plant Mol Biol, 15, 281-293.
Eckart, M. R., Bussineau, C. M. (1996). Quality and authenticity of heterologous proteins synthesized in
yeast. Curr Opin Biotechnol, 7, 525-530.
Eldin, P., Pauza, M. E., Hieda, Y., Lin, G., Murtaugh, M. P., Pentel, P. R., Pennell, C. A. (1997). High-
level secretion of two antibody single chain Fv fragments by Pichia pastoris. J Immunol Methods, 201, 67-
75.
Ellis, S. B., Brust, P. F., Koutz, P. J., F., W. A., Harpold, M. M., R., G. T. (1985). Isolation of alcohol
oxidase and two other methanol regulatable genes from the yeast Pichia pastoris. Mol Cell Biol, 5, 1111-
1121.
Engvall, E., Perlmann, P. (1971). Enzyme-linked immunosorbent assay (ELISA): Quantitative assay of
Immunglobulin G. Immunochemistry, 8, 871-879.
Eyk, H. G. v., Terhorst, C., Vijlder, M. M. d. (1967). Fragmentation of human IgG globulin with papain,
trypsin and pepsin. Clin Chim Acta, 16, 429-431.
Fecker, L. F., Kaufmann, A., Commandeur, U., Commandeur, J., Koenig, R., Burgermeister, W. (1996).
Expression of single-chain antibody fragments (scFv) specific for beet necrotic yellow vein virus coat
protein or 25 kDa protein in Escherichia coli and Nicotiana benthamiana. Plant Mol Biol, 32, 979-986.
Fiedler, U., Conrad, U. (1995). High-level production and long-term storage of engineered antibodies in
transgenic tobacco seeds. Biotechnology (N Y), 13, 1090-1093.
Fiedler, U., Phillips, J., Artsaenko, O., Conrad, U. (1997). Optimization of scFv antibody production in
transgenic plants. Immunotechnology, 3, 205-216.
Fierobe, H. P., Mirgorodskaya, E., Frandsen, T. P., Roepstorff, P., Svensson, B. (1997). Overexpression
and characterization of Aspergillus awamori wild-type and mutant glucoamylase secreted by the
methylotrophic yeast Pichia pastoris: comparison with wild-type recombinant glucoamylase produced
using Saccharomyces cerevisiae and Aspergillus niger as hosts. Protein Expr Purif, 9, 159-170.

Literatur
101
Firek, S., Draper, J., Owen, M. R., Gandecha, A., Cockburn, B., Whitelam, G. C. (1993). Secretion of a
functional single-chain Fv protein in transgenic tobacco plants and cell suspension cultures. Plant Mol
Biol, 23, 861-870.
Fischer, R. (1990). Herstellung, Charakterisierung und Klonierung der Immunglobulingene von TMV-
neutralisierenden Antikörpern (Tübingen: Eberhard-Karls-Universität), .
Fischer, R., Drossard, J., Commandeur, U., Schillberg, S., Emans, N. (1999c). Towards molecular
farming in the future: moving from diagnostic protein and antibody production in microbes to plants.
Biotechnol Appl Biochem, 30, 101-108.
Fischer, R., Drossard, J., Emans, N., Commandeur, U., Hellwig, S. (1999b). Towards molecular farming
in the future: Pichia pastoris-based production of single-chain antibody fragments. Biotechnol Appl
Biochem, 30, 117-120.
Fischer, R., Emans, N., Schuster, F., Hellwig, S., Drossard, J. (1999a). Towards molecular farming in the
future: using plant-cell-suspension cultures as bioreactors. Biotechnol Appl Biochem, 30, 109-112.
Fischer, R., Liao, Y. C., Drossard, J. (1999f). Affinity-purification of a TMV-specific recombinant full-size
antibody from a transgenic tobacco suspension culture. J Immunol Methods, 226, 1-10.
Fischer, R., Liao, Y. C., Hoffmann, K., Schillberg, S., Emans, N. (1999d). Molecular farming of
recombinant antibodies in plants. Biol Chem, 380, 825-839.
Fischer, R., Schumann, D., Zimmermann, S., Drossard, J., Sack, M., Schillberg, S. (1999e). Expression
and characterization of bispecific single-chain Fv fragments produced in transgenic plants. Eur J
Biochem, 262, 810-816.
FitzGerald, K., Holliger, P., Winter, G. (1997). Improved tumour targeting by disulphide stabilized
diabodies expressed in Pichia pastoris. Protein Eng, 10, 1221-1225.
Fontes, E. B. P., Shank, B. B., Wrobel, R. L., Moose, S. P., O'Brian, G. R., Wurtzel, E. T., Boston, R. S.
(1991). Characterization of an immunoglobulin binding protein homolog in the maize floury-2 endosperm
mutant. Plant Cell, 3, 483-496.
Francisco, J. A., Gawlak, S. L., Miller, M., Bathe, J., Russell, D., Chace, D., Mixan, B., Zhao, L., Fell, H.
P., Siegall, C. B. (1997). Expression and characterization of bryodin 1 and a bryodin 1-based single-chain
immunotoxin from tobacco cell culture. Bioconjug Chem, 8, 708-713.
Freyre, F. M., Vazquez, J. E., Ayala, M., Canaan-Haden, L., Bell, H., Rodriguez, I., Gonzalez, A., Cintado,
A., Gavilondo, J. V. (2000). Very high expression of an anti-carcinoembryonic antigen single chain Fv
antibody fragment in the yeast Pichia pastoris. J Biotechnol, 76, 157-63.
Gellissen, G., Hollenberg, C. P. (1997). Application of yeasts in gene expression studies: a comparison of
Saccharomyces cerevisiae, Hansenula polymorpha and Kluyveromyces lactis -- a review. Gene, 190, 87-
97.

Literatur
102
Gellissen, G., Hollenberg, C. P., Janowicz, Z. A. (1995). Gene expression in methylotrophic yeasts. In:
Gene expression in recombinant microorganisms, A. Smith, ed., New York, 195-239.
Govardhan, C. P. (1999). Crosslinking of enzymes for improved stability and performance. Curr Opin
Biotechnol, 10, 331-335.
Gram, H., Schmitz, R., Ridder, R. (1998). Secretion of scFv antibody fragments. In: Pichia Protocols, D.
R. Higgins, J. M. Cregg, eds. Humana Press, Totowa, NJ, 179-192.
Graus, Y. F., de Baets, M. H., Parren, P. W. H. I., Berrih-Aknin, S., Wokke, J., van Breda Vriesman, P. J.,
Burton, D. R. (1997). Human anti-Nicotinic acetylcholine receptor recombinant Fab fragments isolated
from thymus-derived phage display libraries from Myasthenia Gravis patients reflect predominant
specificities in serum and block the action of pathogenic serum antibodies. The Journal of Immunology,
158, 1919-1929.
Han, Y., Lei, X. G. (1999). Role of glycosylation in the functional expression of an Aspergillus niger
phytase (phyA) in Pichia pastoris. Arch Biochem Biophys, 364, 83-90.
Harper, K., Kerschbaumer, R. J., Ziegler, A., Macintosh, S. M., Cowan, G. H., Himmler, G., Mayo, M. A.,
Torrance, L. (1997). A scFv-alkaline phosphatase fusion protein which detects potato leafroll luteovirus in
plant extracts by ELISA. J Virol Methods, 63, 237-242.
Hasslacher, M., Schall, M., Hayn, M., Bona, R., Rumbold, K., Luckl, J., Griengl, H., Kohlwein, S. D.,
Schwab, H. (1997). High-level intracellular expression of hydroxynitrile lyase from the tropical rubber tree
Hevea brasiliensis in microbial hosts. Protein Expr Purif, 11, 61-71.
Heimo, H., Palmu, K., Suominen, I. (1997). Expression in Pichia pastoris and purification of Aspergillus
awamori glucoamylase catalytic domain. Protein Expr Purif, 10, 70-79.
Hein, M. B., Tang, Y., McLeod, D. A., Janda, K. D., Hiatt, A. (1991). Evaluation of immunoglobulins from
plant cells. Biotechnol Prog, 7, 455-461.
Helfrich, W., Kroesen, B. J., Roovers, R. C., Westers, L., Molema, G., Hoogenboom, H. R., de Leij, L.
(1998). Construction and characterization of a bispecific diabody for retargeting T cells to human
carcinomas. Int J Cancer, 76, 232-239.
Hellwig, S., Robin, F., Drossard, J., Raven, N. P., Vaquero-Martin, C., Shively, J. E., Fischer, R. (1999).
Production of carcinoembryonic antigen (CEA) N-A3 domain in Pichia pastoris by fermentation.
Biotechnol Appl Biochem, 30, 267-275.
Henry, A., Masters, C. L., Beyreuther, K., Cappai, R. (1997). Expression of human amyloid precursor
protein ectodomains in Pichia pastoris: analysis of culture conditions, purification, and characterization.
Protein Expr Purif, 10, 283-291.
Herbers, K., Sonnewald, U. (1999). Production of new/modified proteins in transgenic plants. Curr Opin
Biotechnol, 10, 163-168.

Literatur
103
Hiatt, A., Ma, J. K.-C. (1992). Monoclonal antibody engineering in plants. FEBS Letters, 307, 71-75.
Hiatt, A., Robert, C., Bowdish, K. (1989). Production of antibodies in transgenic plants. Nature, 342, 76-
78.
Higgins, D. R., Cregg, J. M. (1998). Introduction to Pichia pastoris. In: Pichia Protocols, D. R. Higgins, J.
M. Cregg, eds. Humana Press, Totowa, NJ, 1-15.
Hochman, J., Inbar, D., Givol, D. (1973). An active antibody fragment (Fv) composed of the variable
portions of heavy and light chains. Biochemistry, 12, 1130-1135.
Hollenberg, C. P., Gellissen, G. (1997). Production of recombinant proteins by methylotrophic yeasts.
Curr Opin Biotechnol, 8, 554-560.
Holliger, P., Brissinck, J., Williams, R. L., Thielemans, K., Winter, G. (1996). Specific killing of lymphoma
cells by cytotoxic T-cells mediated by a bispecific diabody. Protein Eng, 9, 299-305.
Holmquist, M., Tessier, D. C., Cygler, M. (1997). High-level production of recombinant Geotrichum
candidum lipases in yeast Pichia pastoris. Protein Expr Purif, 11, 35-40.
Hood, E. E., Jilka, J. M. (1999). Plant-based production of xenogenic proteins. Curr Opin Biotechnol, 10,
382-386.
Hood, E. E., Kusnadi, A., Nikolov, Z., Howard, J. A. (1999). Molecular farming of industrial proteins from
transgenic maize. Adv Exp Med Biol, 464, 127-147.
Hooker, B. S., Lee, J. M., An, G. (1990). Cultivation of plant cells in a stirred tank reactor. Biotechnol
Bioeng, 35, 296-304.
Hoppmann, V. (2000). Targeting von rekombinanten Antikörpern in verschieden Kompartimente der
Pflanzenzelle. Diplomarbeit; RWTH Aachen, Institut für Biologie I, .
Hu, S., Shively, L., Raubitschek, A., Sherman, M., Williams, L. E., Wong, J. Y., Shively, J. E., Wu, A. M.
(1996). Minibody: A novel engineered anti-carcinoembryonic antigen antibody fragment (single-chain Fv-
CH3) which exhibits rapid, high-level targeting of xenografts. Cancer Res, 56, 3055-3061.
Huston, J. S., Levinson, D., Mudgett-Hunter, M., Tai, M. S., Novotny, J., Margolies, M. N., Ridge, R. J.,
Bruccoleri, R. E., Haber, E., Crea, R., et al. (1988). Protein engineering of antibody binding sites:
recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc
Natl Acad Sci U S A, 85, 5879-5883.
Ikegaya, K., Hirose, M., Ohmura, T., Nokihara, K. (1997). Complete determination of disulfide forms of
purified recombinant human serum albumin, secreted by the yeast Pichia pastoris. Anal Chem, 69, 1986-
1991.
Inbar, D., Hochman, J., Givol, D. (1972). Localization of antibody-combining sites within the variable
portions of heavy and light chains. Proc Natl Acad Sci U S A, 69, 2659-2662.

Literatur
104
Jensen, L. G., Olsen, O., Kops, O., Wolf, N., Thomsen, K. K., von Wettstein, D. (1996). Transgenic barley
expressing a protein-engineered, thermostable (1,3- 1,4)-beta-glucanase during germination. Proc Natl
Acad Sci U S A, 93, 3487-3491.
Jolicoeur, M., Chavarie, C., Carreau, P. J., Archambault, J. (1990). Development of a helical-ribbon
impeller bioreactor for high-density plant cell suspension culture. Biotechnol Bioeng, 39, 511-517.
Jones, P. T., Dear, P. H., Foote, J., Neuberger, M. S., Winter, G. (1986). Replacing the complementarity-
determining regions in a human antibody with those from a mouse. Nature, 321, 522-525.
Jonsson, U., Fagerstam, L., Ivarsson, B., Johnsson, B., Karlsson, R., Lundh, K., Lofas, S., Persson, B.,
Roos, H., Ronnberg, I., et al. (1991). Real-time biospecific interaction analysis using surface plasmon
resonance and a sensor chip technology. Biotechniques, 11, 620-627.
Juge, N., Andersen, J. S., Tull, D., Roepstorff, P., Svensson, B. (1996). Overexpression, purification, and
characterization of recombinant barley alpha-amylases 1 and 2 secreted by the methylotrophic yeast
Pichia pastoris. Protein Expr Purif, 8, 204-214.
Kabat, E. A., Wu, T. T., Reid-Miller, M., Perry, H. M., Gottesman, K. S. (1990). Sequences of proteins of
immunological interest, 5 Edition: U. S. Department of Health
Kapusta, J., Modelska, A., Figlerowicz, M., Pniewski, T., Letellier, M., Lisowa, O., Yusibov, V., Koprowski,
H., Plucienniczak, A., Legocki, A. B. (1999). A plant-derived edible vaccine against hepatitis B virus.
Faseb J, 13, 1796-1799.
Katakura, Y., Zhang, W., Zhuang, G., Omasa, T., Kishimoto, M., Goto, Y., Suga, K. (1998). Effect of
methanol concentration on the production of human ß2-glycoprotein Domain V by a recombinant Pichia
pastoris: a simple system for the control of methanol concentration using a semiconductor gas sensor. J
Ferm Bioeng, 86, 482-487.
Kay, R., Chan, A., Daly, M., McPherson, J. (1987). Duplication of CaMV 35S promoter sequences creates
a strong enhancer for plant genes. Science, 236, 1299-1302.
Ketchum, R. E. B., Gibson, D. M., Croteau, R. B., Shuler, L. (1999). The kinetics of taxoid accumulation in
cell suspension cultures of Taxus following elicitation with methyl jasmonate. Biotechnol Bioeng, 62, 91-
105.
Kim, D. I., Cho, G. H., Pedersen, H., Chin, C. K. (1991). A hybrid bioreactor for high density cultivation of
plant cell suspensions. Appl Microbiol Biotechnol, 34, 726-729.
Kim, T. R., Goto, Y., Hirota, N., Kuwata, K., Denton, H., Wu, S. Y., Sawyer, L., Batt, C. A. (1997). High-
level expression of bovine beta-lactoglobulin in Pichia pastoris and characterization of its physical
properties. Protein Eng, 10, 1339-1345.

Literatur
105
Kipriyanov, S. M., Moldenhauer, G., Schuhmacher, J., Cochlovius, B., Von der Lieth, C. W., Matys, E. R.,
Little, M. (1999). Bispecific tandem diabody for tumor therapy with improved antigen binding and
pharmacokinetics. J Mol Biol, 293, 41-56.
Kipriyanov, S. M., Moldenhauer, G., Strauss, G., Little, M. (1998). Bispecific CD3 x CD19 diabody for T
cell-mediated lysis of malignant human B cells. Int J Cancer, 77, 763-772.
Köhler, G., Milstein, C. (1975). Continuous cultures of fused cells secreting antibody of predefined
specificity. Nature, 256, 495-497.
Kost, T. A., Condreay, J. P. (1999). Recombinant baculoviruses as expression vectors for insect and
mammalian cells. Curr Opin Biotechnol, 10, 428-433.
Kusnadi, A. R., Evangelista, R. L., Hood, E. E., Howard, J. A., Nikolov, Z. L. (1998). Processing of
transgenic corn seed and its effect on the recovery of recombinant beta-glucuronidase. Biotechnol
Bioeng, 60, 44-52.
Kusnadi, A. R., Hood, E. E., Witcher, D. R., Howard, J. A., Nikolov, Z. L. (1998). Production and
purification of two recombinant proteins from transgenic corn. Biotechnol Prog, 14, 149-155.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage
T4. Nature, 227, 680-685.
Lal, S. K., Tulasiram, P., Jameel, S. (1997). Expression and characterization of the hepatitis E virus
ORF3 protein in the methylotrophic yeast, Pichia pastoris. Gene, 190, 63-67.
Laroche, Y., Storme, V., De Meutter, J., Messens, J., Lauwereys, M. (1994). High-level secretion and
very efficient isotopic labeling of tick anticoagulant peptide (TAP) expressed in the methylotrophic yeast,
Pichia pastoris. Biotechnology (N Y), 12, 1119-1124.
Leckie, F., Scragg, A. H., Cliffe, K. R. (1991). Effect of impeller design and speed on the large-scale
cultivation of suspension cultures of Catharanthus roseus. Enz Microb Technol, 13, 801-810.
Lee, J. S., Choi, S. J., Kang, H. S., Oh, W. G., Cho, K. H., Kwon, T. H., Kim, D. H., Jang, Y. S., Yang, M.
S. (1997). Establishment of a transgenic tobacco cell suspension culture system for producing murine
granulocyte-macrophage colony stimulating factor. Mol Cells, 7, 783-787.
Lerouge, P., Cabanes-Macheteau, M., Rayon, C., Fischette-Laine, A. C., Gomord, V., Faye, L. (1998). N-
glycoprotein biosynthesis in plants: recent developments and future trends. Plant Mol Biol, 38, 31-48.
Li, J., Hegeman, C. E., Hanlon, R. W., Lacy, G. H., Denbow, M. D., Grabau, E. A. (1997). Secretion of
active recombinant phytase from soybean cell-suspension cultures. Plant Physiol, 114, 1103-1111.
Lin Cereghino, G. P., Cregg, J. M. (1999). Applications of yeast in biotechnology: protein production and
genetic analysis. Curr Opin Biotechnol, 10, 422-427.

Literatur
106
Linder, S., Schliwa, M., Kube-Granderath, E. (1996). Direct PCR screening of Pichia pastoris clones.
Biotechniques, 20, 980-982.
Linnevers, C. J., McGrath, M. E., Armstrong, R., Mistry, F. R., Barnes, M. G., Klaus, J. L., Palmer, J. T.,
Katz, B. A., Bromme, D. (1997). Expression of human cathepsin K in Pichia pastoris and preliminary
crystallographic studies of an inhibitor complex. Protein Sci, 6, 919-921.
Loewen, M. C., Liu, X., Davies, P. L., Daugulis, A. J. (1997). Biosynthetic production of type II fish
antifreeze protein: fermentation by Pichia pastoris. Appl Microbiol Biotechnol, 48, 480-486.
Lu, Q., Bauer, J. C., Greener, A. (1997). Using Schizosaccharomyces pombe as a host for expression
and purification of eukaryotic proteins. Gene, 200, 135-144.
Luo, D., Geng, M., Schultes, B., Ma, J., Xu, D. Z., Hamza, N., Qi, W., Noujaim, A. A., Madiyalakan, R.
(1998). Expression of a fusion protein of scFv-biotin mimetic peptide for immunoassay. J Biotechnol, 65,
225-228.
Luo, D., Mah, N., Krantz, M., Wilde, K., Wishart, D., Zhang, Y., Jacobs, F., Martin, L. (1995). Vl-linker-Vh
orientation-dependent expression of single chain Fv- containing an engineered disulfide-stabilized bond in
the framework regions. J Biochem (Tokyo), 118, 825-831.
Ma, J. K., Hein, M. B. (1995b). Plant antibodies for immunotherapy. Plant Physiol, 109, 341-346.
Ma, J. K., Hiatt, A., Hein, M., Vine, N. D., Wang, F., Stabila, P., van Dolleweerd, C., Mostov, K., Lehner,
T. (1995a). Generation and assembly of secretory antibodies in plants. Science, 268, 716-719.
Ma, J. K., Hikmat, B. Y., Wycoff, K., Vine, N. D., Chargelegue, D., Yu, L., Hein, M. B., Lehner, T. (1998).
Characterization of a recombinant plant monoclonal secretory antibody and preventive immunotherapy in
humans. Nat Med, 4, 601-606.
Ma, J. K., Lehner, T., Stabila, P., Fux, C. I., Hiatt, A. (1994). Assembly of monoclonal antibodies with
IgG1 and IgA heavy chain domains in transgenic tobacco plants. Eur J Immunol, 24, 131-138.
Magnuson, N. S., Linzmaier, P. M., Gao, J.-W., Reeves, R., An, G., Lee, J. M. (1996). Enhanced recovery
of a secreted mammalian protein from suspension culture of genetically modified tobacco cells. Protein
Expr Purif, 7, 220-228.
Mallender, W. D., Voss, E. W. (1994). Construction, expression and activity of a bivalent bispecific single-
chain antibody. J Biol Chem, 269, 199-206.
Malmqvist, M. (1999). BIACORE: an affinity biosensor system for characterization of biomolecular
interactions. Biochem Soc Trans, 27, 335-340.
Malmqvist, M., Karlsson, R. (1997). Biomolecular interaction analysis: affinity biosensor technologies for
functional analysis of proteins. Curr Opin Chem Biol, 1, 378-383.

Literatur
107
Markaryan, A., Morozova, I., Lee, B. S., Kaplan, A. (1999). Atypical processing of amyloid precursor
fusion protein by proteolytic activity in Pichia pastoris. Biochem Biophys Res Commun, 262, 263-268.
Mason, H. S., Arntzen, C. J. (1995). Transgenic plants as vaccine production systems. Trends
Biotechnol, 13, 388-392.
Mason, H. S., Ball, J. M., Shi, J. J., Jiang, X., Estes, M. K., Arntzen, C. J. (1996). Expression of Norwalk
virus capsid protein in transgenic tobacco and potato and its oral immunogenicity in mice. Proc Natl Acad
Sci U S A, 93, 5335-5340.
Mason, H. S., Haq, T. A., Clements, J. D., Arntzen, C. J. (1998). Edible vaccine protects mice against
Escherichia coli heat-labile enterotoxin (LT): potatoes expressing a synthetic LT-B gene. Vaccine, 16,
1336-1343.
Mason, H. S., Lam, D. M., Arntzen, C. J. (1992). Expression of hepatitis B surface antigen in transgenic
plants. Proc Natl Acad Sci U S A, 89, 11745-9.
Miele, R. G., Nilsen, S. L., Brito, T., Bretthauer, R. K., Castellino, F. J. (1997). Glycosylation properties of
the Pichia pastoris-expressed recombinant kringle 2 domain of tissue-type plasminogen activator.
Biotechnol Appl Biochem, 25, 151-157.
Modelska, A., Dietzschold, B., Sleysh, N., Fu, Z. F., Steplewski, K., Hooper, D. C., Koprowski, H.,
Yusibov, V. (1998). Immunization against rabies with plant-derived antigen. Proc Natl Acad Sci U S A, 95,
2481-2485.
Muller, S., Sandal, T., Kamp-Hansen, P., Dalboge, H. (1998). Comparison of expression systems in the
yeasts Saccharomyces cerevisiae, Hansenula polymorpha, Klyveromyces lactis, Schizosaccharomyces
pombe and Yarrowia lipolytica. Cloning of two novel promoters from Yarrowia lipolytica. Yeast, 14, 1267-
1283.
Munshi, C. B., Fryxell, K. B., Lee, H. C., Branton, W. D. (1997). Large-scale production of human CD38 in
yeast by fermentation. Methods Enzymol, 280, 318-330.
Murphy, K. P., Jr., Gagne, P., Pazmany, C., Moody, M. D. (1998). Expression of human interleukin-17 in
Pichia pastoris: purification and characterization. Protein Expr Purif, 12, 208-214.
Nagata, T., Nemoto, Y., Seiichiro, H. (1992). Tobacco BY-2 cell line as the "HeLa" cell in the cell biology
of higher plants. Int Rev Cytol, 132, 1-30.
Ni, M., Cui, D., Einstein, J., Narasimhulu, S., Vergara, C. E., Gelvin, S. B. (1995). Strength and tissue
specificity of chimeric promoters derived from the octopine and mannopine synthase genes. Plant J, 7,
661-676.
Nilsen, S. L., DeFord, M. E., Prorok, M., Chibber, B. A., Bretthauer, R. K., Castellino, F. J. (1997). High-
level secretion in Pichia pastoris and biochemical characterization of the recombinant kringle 2 domain of
tissue-type plasminogen activator. Biotechnol Appl Biochem, 25, 63-74.

Literatur
108
O'Donohue, M. J., Boissy, G., Huet, J. C., Nespoulous, C., Brunie, S., Pernollet, J. C. (1996).
Overexpression in Pichia pastoris and crystallization of an elicitor protein secreted by the
phytopathogenic fungus, Phytophthora cryptogea. Protein Expr Purif, 8, 254-261.
Ohsawa, I., Hirose, Y., Ishiguro, M., Imai, Y., Ishiura, S., Kohsaka, S. (1995). Expression, purification, and
neurotrophic activity of amyloid precursor protein-secreted forms produced by yeast. Biochem Biophys
Res Commun, 213, 52-58.
Paifer, E., Margolles, E., Cremata, J., Montesino, R., Herrera, L., Delgado, J. M. (1994). Efficient
expression and secretion of recombinant alpha amylase in Pichia pastoris using two different signal
sequences. Yeast, 10, 1415-1419.
Parham, P. (1986). Chapter 14: Immunochemistry. In: Handbook of Experimental Immunology Blackwell
Scientific Publication, 1-23.
Park, C. S., Chang, C. C., Kim, J. Y., Ogrydziak, D. M., Ryu, D. D. (1997). Expression, secretion, and
processing of rice alpha-amylase in the yeast Yarrowia lipolytica. J Biol Chem, 272, 6876-6881.
Parmenter, D. L., Boothe, J. G., van Rooijen, G. J., Yeung, E. C., Moloney, M. M. (1995). Production of
biologically active hirudin in plant seeds using oleosin partitioning. Plant Mol Biol, 29, 1167-1180.
Pen, J., Molendijk, L., Quax, W. J., Sijmons, P. C., van Ooyen, A. J., van den Elzen, P. J., Rietveld, K.,
Hoekema, A. (1992). Production of active Bacillus licheniformis alpha-amylase in tobacco and its
application in starch liquefaction. Biotechnology (N Y), 10, 292-296.
Perisic, O., Webb, P. A., Holliger, P., Winter, G., Williams, R. L. (1994). Crystal structure of a diabody, a
bivalent antibody fragment. Structure, 2, 1217-1226.
Pflanz, S., Tacken, I., Grötzinger, J., Jacques, Y., Dahmen, H., Heinrich, P. C., Müller-Newen, G. (1999).
A fusion protein of interleukin-11 and soluble interleukin-11 receptor acts as a superagonist on cells
expressing gp130. FEBS Lett, 450, 117-122.
Piontek, M., Hagedorn, J., Hollenberg, C. P., Gellissen, G., Strasser, A. W. (1998). Two novel gene
expression systems based on the yeasts Schwanniomyces occidentalis and Pichia stipitis. Appl Microbiol
Biotechnol, 50, 331-338.
Porter, R. R. (1959). The hydrolysis of rabbit gamma-globulin and antibodies with crystalline papain.
Biochem J, 73, 119-126.
Raven, N. (1999). Vergleichende Expression rekombinanter Einzelketten-Antikörper (scFv) in
E. coli, P. pastoris und Pflanzenzellen. Diplomarbeit; RWTH Aachen, Institut für Biologie I, .
Raymond, C. K., Bukowski, T., Holderman, S. D., Ching, A. F., Vanaja, E., Stamm, M. R. (1998).
Development of the methylotrophic yeast Pichia methanolica for the expression of the 65 kilodalton
isoform of human glutamate decarboxylase. Yeast, 14, 11-23.

Literatur
109
Reiter, Y., Wright, A. F., Tonge, D. W., Pastan, I. (1996). Recombinant single-chain and disulfide-
stabilized Fv-immunotoxins that cause complete regression of a human colon cancer xenograft in nude
mice. Int J Cancer, 67, 113-123.
Richter, L., Mason, H. S., Arntzen, C. J. (1996). Transgenic plants created for oral immunization against
diarrheal diseases. J Travel Med, 3, 52-56.
Rittershaus, E., Ulrich, J., Weiss, A., Westphal, K. (1989). Großtechnische Fermentation von pflanzlichen
Zellkulturen: Betriebserfahrungen bei der Kultivierung von Pflanzenzellen in einer Fermentationskaskade
bis zu 75000 Liter. BioEngineering, 5, 28-34.
Robin, F. (1998). Expression chez la levure Pichia pastoris du domaine NA3 de l´antigène
carcinoembryonnaire humain et d´un fragment de l´anticorps correspondant T84.66 constitué
uniquement des domaines variables. Diplomarbeit; RWTH-Aachen, Institut für Biologie I, .
Rodriguez, M., Rubiera, R., Penichet, M., Montesinos, R., Cremata, J., Falcon, V., Sanchez, G., Bringas,
R., Cordoves, C., Valdes, M. (1994). High level expression of the B. microplus Bm86 antigen in the yeast
Pichia pastoris forming highly immunogenic particles for cattle. J Biotechnol, 33, 135-146.
Romanos, M. A., Clare, J. J., Beesley, K. M., Rayment, F. B., Ballantine, S. P., Makoff, A. J., Dougan, G.,
Fairweather, N. F., Charles, I. G. (1991). Recombinant Bordetella pertussis pertactin (P69) from the yeast
Pichia pastoris: high-level production and immunological properties. Vaccine, 9, 901-906.
Sack, M. (1998). Untersuchungen zum Anwendungspotential rekombinanter Antikörper aus Nicotiana
tabacum. Diplomarbeit; RWTH Aachen, Institut für Biologie I, .
Saiki, R. K., Scharf, S., Faloona, F., Mullis, K. B., Horn, G. T., Ehrlich, H. A., Arnheim, N. (1985).
Enzymatic amplification of b-globin genomic sequences and restriction site analysis for diagnosis of sickle
cell anemia. Science, 230, 1350-1354.
Sakai, Y., Akiyama, M., Kondoh, H., Shibano, Y., Kato, N. (1996). High-level secretion of fungal
glucoamylase using the Candida boidinii gene expression system. Biochim Biophys Acta, 1308, 81-87.
Sambrook, J., Fritsch, E. F., Maniatis, T. (1989). Molecular cloning - A laboratory manual, 2nd Edition:
Cold Spring Harbour Laboratory, Cold Spring Harbour, NY
Sato, K., Nakayama, M., Shigeta, J. (1996). Culturing conditions affecting the production of anthocyanin
in suspended cell cultures of strawberry. Plant Science, 113, 91-98.
Schiffer, M., Girling, R. L., Ely, K. R., Edmundson, A. B. (1973). Structure of a lambda-type Bence-Jones
protein at 3.5-A resolution. Biochemistry, 12, 4620-4631.
Schillberg, S., Zimmermann, S., Voss, A., Fischer, R. (1999). Apoplastic and cytosolic expression of full-
size antibodies and antibody fragments in Nicotiana tabacum. Transgenic Res, 8, 255-263.
Schumann, D. (1996). Herstellung und Expression bispezifischer "Single chain" Antikörper. Diplomarbeit;
RWTH Aachen, Institut für Biologie I, .

Literatur
110
Scorer, C. A., Buckholz, R. G., Clare, J. J., Romanos, M. A. (1993). The intracellular production and
secretion of HIV-1 envelope protein in the methylotrophic yeast Pichia pastoris. Gene, 136, 111-119.
Scragg, A. H. (1991). Plant and animal cell bioreactors. In: Bioreactors in biotechnology: a practical
approach, A. H. Scragg, ed. Ellis Horwood, London, 271-301.
Scragg, A. H. (1992). Large-scale plant cell culture: methods, applications and products. Curr Opin
Biotechnol, 3, 105-109.
Scragg, A. H. (1997). The production of aromas by plant cell cultures. In: Advances in biochemical
engineering/Biotechnology, T. Scheper, ed. Springer, Berlin, Heidelberg, 239-263.
Sehnke, P. C., Ferl, R. J. (1999). Processing of preproricin in transgenic tobacco. Protein Expr Purif, 15,
188-195.
Sharp, J. M., Doran, P. M. (1999). Effect of bacitracin on growth and monoclonal antibody production by
tobacco hairy roots and cell suspensions. Biotechnol Bioprocess Eng, 4, 253-258.
Shinmyo, A., Shoji, T., Bando, E., Nagaya, S., Nakai, Y., Kato, K., Sekine, M., Yoshida, K. (1998).
Metabolic engineering of cultured tobacco cells. Biotechnol Bioeng, 58, 329-332.
Siegel, R. S., Brierley, R. A. (1989). Methylotrophic yeast Pichia pastoris produced in high-cell-density
fermentations with high cell yields as vehicle for recombinant protein production. Biotechnol Bioeng, 34,
403-404.
Singh, G., Curtis, W. R. (1995). Reactor design for plant cell suspension cultures. In: Biotechnological
applications of plant cultures, P. D. Shargool, T. T. Ngo, eds. CRC Press, Boca Raton, FL, USA, 151-
183.
Smart, N. J., Fowler, M. W. (1981). Effect of aeration on large-scale cultures of plant cells. Biotechnol
Lett, 3, 171-176.
Spiegel, H., Schillberg, S., Sack, M., Holzem, A., Nähring, J. M., Monecke, M., Liao, Y.-C., Fischer, R.
(1999). Accumulation of antibody fusion proteins in the cytoplasm and ER of plant cells. Plant Sci, 149,
63-71.
Spieler, H., Alferman, A. W., Reinhard, E. (1985). Biotransformation of ß-methyldigitoxin by cell cultures
of Digitalis lanata in airlift and stirred tank reactors. Appl Microbiol Biotechnol, 23, 1-4.
Sreekrishna, K., Brankamp, R. G., Kropp, K. E., Blankenship, D. T., Tsay, J. T., Smith, P. L., Wierschke,
J. D., Subramaniam, A., Birkenberger, L. A. (1997). Strategies for optimal synthesis and secretion of
heterologous proteins in the methylotrophic yeast Pichia pastoris. Gene, 190, 55-62.
Sreekrishna, K., Nelles, L., Potenz, R., Cruze, J., Mazzaferro, P., Fish, W., Fuke, M., Holden, K., Phelps,
D., Wood, P., Parker, K. (1989). High-level expression, purification and characterization of recombinant

Literatur
111
human tumor necrosis factor synthesized in the methylotrophic yeast Pichia pastoris. Biochemistry, 28,
4117-4125.
Staub, J. M., Maliga, P. (1992). Long regions of homologous DNA are incorporated into the tobacco
plastid genome by transformation. Plant Cell, 4, 39-45.
Stenberg, E., Persson, B., Roos, H., Urbaniczky, C. (1991). Quantitative determination of surface
concentration of protein with surface plasmon resonance by using radiolabelled proteins. J Colloid
Interface Sci, 143, 513-526.
Stoger, E., Vaquero, C., Torres, E., Sack, M., Nicholson, L., Drossard, J., Williams, S., Keen, D., Perrin,
Y., Christou, P., Fischer, R. (2000). Cereal crops as viable production and storage systems for
pharmaceutical scFv antibodies. Plant Mol Biol, 42, 583-590.
Stratton, J., Chiruvolu, V., Meagher, M. (1998). High cell-density fermentation. In: Pichia Protocols, D. R.
Higgins, J. M. Cregg, eds. Humana Press, Totowa, NJ, 107-121.
Sun, J., Bird, C. H., Buzza, M. S., McKee, K. E., Whisstock, J. C., Bird, P. I. (1999). Expression and
purification of recombinant human granzyme B from Pichia pastoris. Biochem Biophys Res Commun,
261, 251-255.
Swinkels, B. W., van Ooyen, A. J., Bonekamp, F. J. (1993). The yeast Kluyveromyces lactis as an
efficient host for heterologous gene expression. Antonie Van Leeuwenhoek, 64, 187-201.
Tai, M. S., Mudgett-Hunter, M., Levinson, D., Wu, G. M., Haber, E., Oppermann, H., Huston, J. S. (1990).
A bifunctional fusion protein containing Fc-binding fragment B of staphylococcal protein A amino terminal
to antidigoxin single-chain Fv. Biochemistry, 29, 8024-8030.
Tanaka, H., Nishijima, F., Suwa, M., Iwamoto, T. (1983). Rotating Drum Fermentor for plant cell
suspension cultures. Biotechnol Bioeng, 25, 2359-2370.
Taticek, R. A., Moo-Young, M., Legge, R. L. (1991). The scale-up of plant cell culture: Engineering
considerations. Plant Cell, Tissue and Organ Culture, 24, 139-158.
Terashima, M., Murai, Y., Kawamura, M., Nakanishi, S., Stoltz, T., Chen, L., Drohan, W., Rodriguez, R.
L., Katoh, S. (1999). Production of functional human alpha 1-antitrypsin by plant cell culture. Appl
Microbiol Biotechnol, 52, 516-523.
Torres, E., Vaquero, C., Nicholson, L., Sack, M., Stoger, E., Drossard, J., Christou, P., Fischer, R., Perrin,
Y. (1999). Rice cell culture as an alternative production system for functional diagnostic and therapeutic
antibodies. Transgenic Res, 8, 441-449.
Towbin, H., Staehlin, T., Gordon, T. (1979). Electrophoretic transfer of proteins from polyacrylamide gels
to nitrocellulose sheets. Procedure and some applications. Proc Natl Acad Sci USA, 76, 4350-4354.
Tschopp, J. F., Brust, P. F., Cregg, J. M., Stillman, C. A., Gingeras, T. R. (1987). Expression of the lacZ
gene from two methanol-regulated promotors in Pichia pastoris. Nucl Acids Res, 15, 3859-3877.

Literatur
112
Tschopp, J. F., Sverlow, G., Kosson, R., Craig, W., Grinna, L. (1987). High-level secretion of glycosylated
invertase in the methylotrophic yeast, Pichia pastoris. BIO/TECHNOLOGY, 5, 1305-1308.
Van Nostrand, W. E., Schmaier, A. H., Neiditch, B. R., Siegel, R. S., Raschke, W. C., Sisodia, S. S.,
Wagner, S. L. (1994). Expression, purification, and characterization of the Kunitz-type proteinase inhibitor
domain of the amyloid beta-protein precursor-like protein-2. Biochim Biophys Acta, 1209, 165-170.
Vaquero, C., Sack, M., Chandler, J., Drossard, J., Schuster, F., Monecke, M., Schillberg, S., Fischer, R.
(1999). Transient expression of a tumor-specific single-chain fragment and a chimeric antibody in tobacco
leaves. Proc Natl Acad Sci U S A, 96, 11128-11133.
Verch, T., Yusibov, V., Koprowski, H. (1998). Expression and assembly of a full-length monoclonal
antibody in plants using a plant virus vector. J Immunol Methods, 220, 69-75.
Verhoeyen, M., Milstein, C., Winter, G. (1988). Reshaping human antibodies: grafting an antilysozyme
activity. Science, 239, 1534-1536.
Verwoerd, T. C., van Paridon, P. A., van Ooyen, A. J., van Lent, J. W., Hoekema, A., Pen, J. (1995).
Stable accumulation of Aspergillus niger phytase in transgenic tobacco leaves. Plant Physiol, 109, 1199-
1205.
Vitale, A., Ceriotti, A., Denecke, J. (1993). The role of the endoplasmic reticulum in protein synthesis,
modification and intracellular transport. J Exp Bot, 44, 1417-1444.
Waterham, H. R., Digan, M. E., Koutz, P. J., Lair, S. V., Cregg, J. M. (1997). Isolation of the Pichia
pastoris glyceraldehyde-3-phosphate dehydrogenase gene and regulation and use of its promoter. Gene,
186, 37-44.
Weiss, S., Famulok, M., Edenhofer, F., Wang, Y. H., Jones, I. M., Groschup, M., Winnacker, E. L. (1995).
Overexpression of active Syrian golden hamster prion protein PrPc as a glutathione S-transferase fusion
in heterologous systems. J Virol, 69, 4776-4783.
Werten, M. W., van den Bosch, T. J., Wind, R. D., Mooibroek, H., de Wolf, F. A. (1999). High-yield
secretion of recombinant gelatins by Pichia pastoris. Yeast, 15, 1087-1096.
White, C. E., Hunter, M. J., Meininger, D. P., White, L. R., Komives, E. A. (1995). Large-scale expression,
purification and characterization of small fragments of thrombomodulin: the roles of the sixth domain and
of methionine 388. Protein Eng, 8, 1177-1187.
Winter, G., Milstein, C. (1991). Man-made antibodies. Nature, 349, 293-299.
Wong, J. Y., Williams, L. E., Yamauchi, D. M., Odom-Maryon, T., Esteban, J. M., Neumaier, M., Wu, A.
M., Johnson, D. K., Primus, F. J., Shively, J. E., et al. (1995). Initial experience evaluating 90yttrium-
radiolabeled anti- carcinoembryonic antigen chimeric T84.66 in a phase I radioimmunotherapy trial.
Cancer Res, 55, 5929s-5934s.

Literatur
113
Wurm, F., Bernard, A. (1999). Large-scale transient expression in mammalian cells for recombinant
protein production. Curr Opin Biotechnol, 10, 156-159.
You, Y. H., Hefta, L. J., Yazaki, P. J., Wu, A. M., Shively, J. E. (1998). Expression, purification, and
characterization of a two domain carcinoembryonic antigen minigene (N-A3) in Pichia pastoris. The
essential role of the N-domain. Anticancer Res, 18, 3193-3201.
Yusibov, V., Modelska, A., Steplewski, K., Agadjanyan, M., Weiner, D., Hooper, D. C., Koprowski, H.
(1997). Antigens produced in plants by infection with chimeric plant viruses immunize against rabies virus
and HIV-1. Proc Natl Acad Sci U S A, 94, 5784-5788.
Zeitlin, L., Olmsted, S. S., Moench, T. R., Co, M. S., Martinell, B. J., Paradkar, V. M., Russell, D. R.,
Queen, C., Cone, R. A., Whaley, K. J. (1998). A humanized monoclonal antibody produced in transgenic
plants for immunoprotection of the vagina against genital herpes. Nat Biotechnol, 16, 1361-1364.
Zhan, H., Liu, B., Reid, S. W., Aoki, K. H., Li, C., Syed, R. S., Karkaria, C., Koe, G., Sitney, K., Hayenga,
K., Mistry, F. et al. (1999). Engineering a soluble extracellular erythropoietin receptor (EPObp) in Pichia
pastoris to eliminate microheterogeneity, and its complex with erythropoietin. Protein Eng, 12, 505-13.
Zhu, A., Monahan, C., Zhang, Z., Hurst, R., Leng, L., Goldstein, J. (1995). High-level expression and
purification of coffee bean alpha-galactosidase produced in the yeast Pichia pastoris. Arch Biochem
Biophys, 324, 65-70.
Ziegler, M. T., Thomas, S. R., Danna, K. J. (2000). Accumulation of a thermostable endo-1,4-beta-D-
glucanase in the apoplast of Arabidopsis thaliana leaves. Mol Breed, 6, 37-46.

Anhang
114
Anhang A. Abkürzungen
2,4 D 2,4-Di-Chlor-phenoxyessigsäure
AOX1 Alkoholoxidase 1
AP Alkalische Phosphatase
APS Ammoniumpersulfat
ATCC "American Type Culture Collection"
BMGY Komplexes, gepuffertes Pichia-Medium mit Glyzerin
BMMY Komplexes, gepuffertes Pichia-Induktionsmedium mit Methanol
CaMV "Cauliflower Mosaic Virus", Blumenkohlmosaikvirus
CEA "Carcinoembryonic antigen", Darmkrebs-assoziiertes Oberflächenantigen
CH Konstante Region der schweren Kette
CL Konstante Region der leichten Kette
DMSO Dimethylsulfoxid
DTT Dithiothreitol
EDTA Ethylendiamintetraacetat
ELISA Enzyme-linked immunosorbent assay
ER Endoplasmatisches Retikulum
et al. und andere
Fab "Fragment antigen binding", Antigen-bindendes Fragment
FAD Flavin-Adenin Dinukleotid
Fc "Fragment crystalisable", kristallisierbares Fragment
FSH Follikelstimulierendes Hormon (Peptidhormon)
FW Frischgewicht
GAP Glyzerinaldehydphosphat-Dehydrogenase
GMP "Good Manufacturing Practice"
GUS β-Glucuronidase
hAAT humanes α-1-Antitypsin
hCG "human chorionic gonadotrophin", Peptidhormon
HEPES N-2-Hydroxyethylpiperazin-N'-2-ethansulfonsäure
ID Innendurchmesser
IDA "Imino-diacetic acid"
mAk monoklonaler Antikörper

Anhang
115
mGM-CSF muriner Granulocyten Makrophagen-Koloniestimulierender Faktor
MSMO "Murashige & Skoog with minimal organics", Pflanzenzellkulturmedium
Mut+ "Methanol-utilization" Wildtyp
MutS "Methanol utilization slow"
NRRL "Northern Regional Research Laboratories", Peoria, IL
NTA "Nitrilo-triacetic acid"
OD Optische Dichte
PAA Polyacrylamid
PMSF Polymethylsulfon
PNPP p-Nitrophenylphosphat
rAk rekombinanter Antikörper
RIP "ribosome-inactivating protein"
RSA Rinderserumalbumin, hier: Fraktion V
scFv "single chain Fragment variable", Einzelkettenantikörper
SDS "Sodium-dodecyl-sulfate ", Natrium-dodecyl-sulfate
SDS-PAA SDS-Polyacrylamid
SDS-PAGE SDS-Polyacrylamid-Gelelektrophorese
SV Säulenvolumen
TBE Tris/Borsäure/EDTA Puffer (Elektrophorese von DNA)
TEMED N,N,N',N'-Tetramethylethylenediamin
TMV Tabakmosaikvirus
TRIS Tris-(hydroxylmethyl)-aminomethan
Tween 20 poly-oxyethylen(20) sorbitan-monolaurate
vvm "volume per volume per minute", Belüftungsrate
v/v "volume per volume", Volumenprozent
VH Variable Region der schweren Kette
VL Variable Region der leichten Kette
w/v "weight per volume", Gewicht pro Volumen
YPD Komplexes, ungepuffertes Pichia-Medium mit Glukose
YPG Komplexes, ungepuffertes Pichia-Medium mit Glyzerin
YPM Komplexes, ungepuffertes Pichia-Induktionsmedium mit Methanol

Anhang
116
Anhang B. Veröffentlichungen, die direkt oder indirekt aus der Promotionsarbeitresultieren
• Drossard, J., Nähring, J. M., Hellwig, S., Fischer, R. (1997). Production of engineeredantibodies in tobacco plants and cell suspension cultures. In: Antibody engineering, Newtechnology, Application and Commercialization: IBC library services, 69-93.
• Fischer, R., Drossard, J., Emans, N., Commandeur, U., Hellwig, S. (1999). Towardsmolecular farming in the future: Pichia pastoris-based production of single-chain antibodyfragments. Biotechnol Appl Biochem: 30, 117-120.
• Fischer, R., Drossard, J., Hellwig, S., Emans, N., Schillberg, S. (1998). Transgenic plants asbioreactors for the expression of recombinant antibodies. In: 10th International Congress ofImmunology (New Delhi, India: Monduzzi Editore, S. p. A., Bologna, Italy), 307-313.
• Fischer, R., Emans, N., Schuster, F., Hellwig, S., Drossard, J. (1999). Towards molecularfarming in the future: using plant-cell-suspension cultures as bioreactors. Biotechnol ApplBiochem: 30, 109-112.
• Hellwig, S., Robin, F., Drossard, J., Raven, N. P., Vaquero-Martin, C., Shively, J. E.,Fischer, R. (1999). Production of carcinoembryonic antigen (CEA) N-A3 domain in Pichiapastoris by fermentation. Biotechnol Appl Biochem: 30, 267-275.
• Hellwig, S., Emde, F., Henke, M., Raven, N. P. G., van der Logt, P., Fischer, R.: Analysis ofsingle-chain antibody production in Pichia pastoris using on-line methanol control in fed-batch and mixed-feed fermentations. Eingereicht (Biotechnol Bioeng)

Anhang
117
Anhang C. Poster und Vorträge
• Hellwig, S., Drossard, J., Schillberg, S., Fischer, R.: Production of recombinant antibodies byfermentation of transgenic plant suspension cell lines. 8th IBC Conference on "Recombinantantibody engineering", San Diego, CA, U. S. A. Dezember 1997
• Hellwig, S., Drossard, J., Fischer, S.: Production of antibody fragments in suspensioncultures of Nicotiana tabacum: From shake flask to stirred tank reactor. Vortrag, DECHEMAJahrestagung der Biotechnologen, Wiesbaden, 1998
• Hellwig, S., Drossard, J., Sack, M., Chandler, J., Vaquero, C., Fischer, R.: Production ofrecombinant antibodies by fermentation of plant suspension cell lines. Poster, XthInternational virology conference, Jerusalem, Israel, 1998
• Hellwig, S., Emde, F., Raven, N. P. G., Fischer, R.: Production of scFv antibody fragments inthe methylotrophic yeast Pichia pastoris using online methanol control in fed batchfermentations. Poster, DECHEMA 1999

Anhang
118
Danksagung
Prof. Fritz Kreuzaler danke ich für die Möglichkeit, diese Arbeit am Institut für Biologie Ianfertigen zu können und für seine wunderbaren Vorlesungen.
Prof. Winfried Hartmeier danke ich für die Übernahme des Korreferates.
Ich danke Dr. Rainer Fischer für die Aufnahme in seine Arbeitsgruppe, seinen unermüdlichenund selbstlosen Einsatz für den Erfolg der Arbeitsgruppe und das in mich gesetzte Vertrauen.
Ich danke neben allen anderen Mitarbeitern der "Antibody group" vor allem (in alphabeticalorder) Olga Artsaenko (Unilever, intakte Pflanzen), Jürgen Drossard (Tabak,Reinigungsstrategien), Mario Henke (versteht auch was von Fermentern), Torsten Natelberg(Zerstörungsexperte), Nicole Raven (N55, PCR), Frederic Robin (PP599, Kultur, retraite auxflambeaux), und Flora Schuster (Vorkulturen) für die hervorragende Zusammenarbeit.
In meiner Zeit bei Unilever in Bedford und während des gesamten "Unilever" EU-Projekteswaren Prof. Philip Porter, Prof. Paul Davis, Dr. Glen Hughes und Dr. Paul van der Logt vonUnilever sowie Prof. Rob Meloen und Jerry Slootstra vom ID-DLO auf wissenschaftlicher undmenschlicher Basis hervorragende Kollegen.
Mit Dr. Frank Emde (Frings GmbH & Co KG, Bonn) etablierte sich eine ausgesprochenfruchtbare Zusammenarbeit auf dem Gebiet der Online-Methanol-Analytik
Holger, Jörg, Monne und Achim haben mich in völliger Ignoranz der Unwahrscheinlichkeit, daßich es schaffen könnte, ermutigt, ernsthaft für einen Marathon zu trainieren. Mein Erfolg gehörteuch.
Anke Krüger hat mich trotz häufig unmöglicher Arbeitszeiten ("Wann bist du eigentlich zuletztnach Hause gekommen?") stets unterstützt und schließlich sogar geheiratet. Das ist tollkühn undverdient Dank und Anerkennung.

Anhang
119
Lebenslauf
Name: Stephan Hellwig
Geburtsdatum/-ort: 1. April 1971 in Birkesdorf (jetzt Düren)
Anschrift: Abteistraße 8
52066 Aachen
Staatsangehörigkeit: deutsch
Familienstand: verheiratet
Schulbildung: 8/80 – 8/89 Franziskusgymnasium Vossenack
Abschluß: Abitur (2,3)
Zivildienst: 10/89 –10/90 Altenheim St. Nikolaus, Düren
Studium: 10/90 –7/96 RWTH Aachen
Abschluß: Diplom-Biologe
Prüfungsfächer: Biotechnologie (Prof. Hartmeier)
Mikrobiologie (Prof. Wolf)
Zoologie (Prof. Scriba)
Organische Chemie (Prof. Höcker)
Titel der Diplomarbeit: Versuche zur Optimierung der kontinuierlichen Milchsäurefermentation
mit Lactobacillus casei in der Blenke-Kaskade (RWTH Aachen, Lehrstuhl für Biotechnologie)
Berufstätigkeit: seit 10/96 Wiss. Angestellter,
RWTH Aachen, Institut für Biologie I
Arbeitsgruppe Dr. Fischer
Related Documents











