RESEARCH PAPER ENOD40 affects elongation growth in tobacco Bright Yellow-2 cells by alteration of ethylene biosynthesis kinetics Tom Ruttink 1, *, Kees Boot 2 , Jan Kijne 2 , Ton Bisseling 1 and Henk Franssen 1,† 1 Laboratory of Molecular Biology, Department of Plant Sciences, Wageningen University, Dreijenlaan 3, 6703 HA, Wageningen, The Netherlands 2 Institute of Biology, Leiden University, Wassenaarseweg 64, 2333 AL, Leiden, The Netherlands Received 31 March 2006; Accepted 21 June 2006 Abstract Plant developmental processes are controlled by co- ordinated action of phytohormones and plant genes encoding components of developmental response pathways. ENOD40 was identified as a candidate for such a plant factor with a regulatory role during nod- ulation. Although its mode of action is poorly under- stood, several lines of evidence suggest interaction with phytohormone response pathways. This hypothe- sis was investigated by analysing cytokinin-, auxin-, and ethylene-induced responses on cell growth and cell division in transgenic 35S:NtENOD40 Bright Yellow- 2 (BY-2) tobacco cell suspensions. It was found that cell division frequency is controlled by the balance between cytokinin and auxin in wild-type cells and that this regulation is not affected in 35S:NtENOD40 lines. Elongation growth, on the other hand, is reduced upon overexpression of NtENOD40. Analysis of ethy- lene homeostasis shows that ethylene accumulation is accelerated in 35S:NtENOD40 lines. ENOD40 action can be counteracted by an ethylene perception blocker, indicating that ethylene is a negative regulator of elongation growth in 35S:NtENOD40 cells, and that the NtENOD40-induced response is mediated by alter- ation of ethylene biosynthesis kinetics. Key words: BY-2 cells, elongation growth, ENOD40, ethylene. Introduction ENOD40 homologues have been identified in plant spe- cies across the plant kingdom, including monocots like rice (Kouchi et al., 1999), rye grass, barley (Larsen et al., 2003), Zea mays (Compaan et al., 2003), and sorghum, and dicots such as tomato (Vleghels et al., 2003), tobacco (Matvienko et al., 1996), citrus, and numerous leguminous species. The highest expression levels of ENOD40 have been found during legume nodule formation, and therefore its func- tion has been studied in most detail during this process. Misregulation of ENOD40 in Medicago truncatula by co- suppression reduces the number of nodules and nodule development is arrested, indicating that ENOD40 has a regulatory role in nodule organogenesis (Crespi et al., 1994; Charon et al., 1999). Ectopic expression of ENOD40, on the other hand, induces cortical cell divisions in Medicago roots and accelerates nodule development (Charon et al., 1997, 1999). However, ENOD40 expression alone is not sufficient for nodule primordium formation (Minami et al., 1996; Mathesius et al., 2000), and inter- action with other plant factors is probably required for the initiation of nodule development. Several observations (Hirsch et al., 1989; Peters and Crist-Estes, 1989; Lee and LaRue, 1992; Cooper and Long, 1994; Heidstra et al., 1997) show the involvement of phytohormones, in par- ticular auxin, cytokinin, and ethylene, and of ENOD40 (Charon et al., 1997) suggesting that, during nodule devel- opment, cross-talk between ENOD40 and phytohormone signalling exists. The expression of ENOD40 homologues in developmental processes in non-leguminous plant species, for example, during lateral root formation, flower development, * Present address: Department of Plant Systems Biology, VIB/Gent University, Technologiepark 927, 9052, Gent, Belgium. y To whom correspondence should be addressed. E-mail: [email protected] Journal of Experimental Botany, Vol. 57, No. 12, pp. 3271–3282, 2006 doi:10.1093/jxb/erl089 Advance Access publication 6 September, 2006 ª The Author [2006]. Published by Oxford University Press [on behalf of the Society for Experimental Biology]. All rights reserved. For Permissions, please e-mail: [email protected] by guest on June 1, 2013 http://jxb.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

RESEARCH PAPER
ENOD40 affects elongation growth in tobaccoBright Yellow-2 cells by alteration of ethylenebiosynthesis kinetics
Tom Ruttink1 Kees Boot2 Jan Kijne2 Ton Bisseling1 and Henk Franssen1dagger
1 Laboratory of Molecular Biology Department of Plant Sciences Wageningen University Dreijenlaan 36703 HA Wageningen The Netherlands2 Institute of Biology Leiden University Wassenaarseweg 64 2333 AL Leiden The Netherlands
Received 31 March 2006 Accepted 21 June 2006
Abstract
Plant developmental processes are controlled by co-
ordinated action of phytohormones and plant genes
encoding components of developmental response
pathways ENOD40 was identified as a candidate for
such a plant factor with a regulatory role during nod-
ulation Although its mode of action is poorly under-
stood several lines of evidence suggest interaction
with phytohormone response pathways This hypothe-
sis was investigated by analysing cytokinin- auxin-
and ethylene-induced responses on cell growth and
cell division in transgenic 35SNtENOD40 Bright Yellow-
2 (BY-2) tobacco cell suspensions It was found that
cell division frequency is controlled by the balance
between cytokinin and auxin in wild-type cells and
that this regulation is not affected in 35SNtENOD40
lines Elongation growth on the other hand is reduced
upon overexpression of NtENOD40 Analysis of ethy-
lene homeostasis shows that ethylene accumulation
is accelerated in 35SNtENOD40 lines ENOD40
action can be counteracted by an ethylene perception
blocker indicating that ethylene is a negative regulator
of elongation growth in 35SNtENOD40 cells and that
the NtENOD40-induced response is mediated by alter-
ation of ethylene biosynthesis kinetics
Key words BY-2 cells elongation growth ENOD40 ethylene
Introduction
ENOD40 homologues have been identified in plant spe-cies across the plant kingdom including monocots like rice(Kouchi et al 1999) rye grass barley (Larsen et al 2003)Zea mays (Compaan et al 2003) and sorghum and dicotssuch as tomato (Vleghels et al 2003) tobacco (Matvienkoet al 1996) citrus and numerous leguminous speciesThe highest expression levels of ENOD40 have been foundduring legume nodule formation and therefore its func-tion has been studied in most detail during this processMisregulation of ENOD40 in Medicago truncatula by co-suppression reduces the number of nodules and noduledevelopment is arrested indicating that ENOD40 has aregulatory role in nodule organogenesis (Crespi et al1994 Charon et al 1999) Ectopic expression of ENOD40on the other hand induces cortical cell divisions inMedicago roots and accelerates nodule development(Charon et al 1997 1999) However ENOD40 expressionalone is not sufficient for nodule primordium formation(Minami et al 1996 Mathesius et al 2000) and inter-action with other plant factors is probably required for theinitiation of nodule development Several observations(Hirsch et al 1989 Peters and Crist-Estes 1989 Leeand LaRue 1992 Cooper and Long 1994 Heidstra et al1997) show the involvement of phytohormones in par-ticular auxin cytokinin and ethylene and of ENOD40(Charon et al 1997) suggesting that during nodule devel-opment cross-talk between ENOD40 and phytohormonesignalling exists The expression of ENOD40 homologues indevelopmental processes in non-leguminous plant species forexample during lateral root formation flower development
Present address Department of Plant Systems Biology VIBGent University Technologiepark 927 9052 Gent Belgiumy To whom correspondence should be addressed E-mail HenkFranssenwurnl
Journal of Experimental Botany Vol 57 No 12 pp 3271ndash3282 2006
doi101093jxberl089 Advance Access publication 6 September 2006
ordf The Author [2006] Published by Oxford University Press [on behalf of the Society for Experimental Biology] All rights reservedFor Permissions please e-mail journalspermissionsoxfordjournalsorg
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
and vascular tissue development (Kouchi et al 1999Varkonyi-Gasic and White 2002 Vleghels et al 2003)indicates that the function of ENOD40 is not confinedto nodule development in leguminous species and suggeststhat ENOD40 has a general role in plant development Thisnotion is supported by the observation that ectopic expres-sion of ENOD40 affected formation of somatic embryosof alfalfa under in vitro culture conditions (Crespi et al1994) Also overexpression of ENOD40 led to reducedapical dominance in tobacco (van de Sande et al 1996)Both observations indicate that phytohormone signalling isaffected by ENOD40 Up to now the function of ENOD40and its mode of action have been poorly understoodAlthough observations in both legumes and non-legumesare pointing to a cross-talk between ENOD40 activity andphytohormone signalling pathways direct evidence forsuch an interaction is lacking Establishing whether thefunction of ENOD40 involves interaction with phyto-hormone signalling pathways could be an important steptowards unravelling the role of ENOD40 during organo-genesis Therefore a search was made for a system thatwould make it possible to test whether cross-talk betweenENOD40 and phytohormone signalling occurs The to-bacco Bright Yellow-2 (BY-2) cell suspension was chosenas a model system as it is convenient for studying phyto-hormone responses on a cellular level In BY-2 cellselongation growth and cell division are regulated by thebalance between cytokinin and auxin in the culture medium(Hasezawa and Syono 1983) Thus cell elongation growthand cell division frequency can be used as morphologicalmarkers to study whether overexpression of ENOD40affects the response of BY-2 cells to phytohormones Itwas found that overexpression of ENOD40 negatively af-fects cell elongation growth whereas cytokinin- or auxin-dependent control of cell division frequency is not affectedin 35SNtENOD40 transgenic cell lines It was shownfurther that the altered ethylene biosynthesis kinetics ob-served in ENOD40-overexpressing cells is a primary causeof the reduction in cell elongation growth
Materials and methods
Construction of binary vector p35SNtENOD40
Nicotiana tabacum contains two ENOD40 homologues that are96 identical at the nucleotide level (Matvienko et al 1996) Thecauliflower mosaic virus 35S promoter from pMON999 (Monsanto)was transferred to pCambia 1390 (Cambia Australia) yieldingp35STnos A 470 bp PCR fragment corresponding to the Nt-ENOD40-1 cDNA sequence was then cloned in p35STnos
Liquid BY-2 cultures and BY-2 transformation
Nicotiana tabacum BY-2 cell suspensions were subcultured weeklyby 403 dilution in fresh medium (Nagata et al 1981) BY-2transformation was performed using a modification of the procedurereported by Gu and Verma (1997) Five millilitres of a 3-d-old BY-2cell suspension was co-cultivated for 2 d at 25 8C in the dark with
60 ll of log-phase Agrobacterium tumefaciens strain C58C1harbouring the binary vector Cells were washed three times beforeplating on culture medium supplemented with 08 Daishin agar200 lg l1 ticarcilineclavuline and 40 lg l1 hygromycin BTransgenic calli that appeared after 3ndash4 weeks were cultured on freshselection plates for 1 more week and were subsequently transferredto liquid selection medium Six independent 35SNtENOD40 BY-2cell lines were generated and named lines Nt1 to Nt6 Each transgenicline was derived from a different callus which means that they can-not be siblings Transgenic lines were continuously maintained inselection medium
Protoplast isolation
Protoplasts were obtained from 6-d-old suspension cultures using1 (wv) cellulase-YC and 01 (wv) pectolyase Y23 in 04 MD-mannitol pH 55 (Nagata et al 1981) Cells were incubated in theenzyme solution for 3 h at room temperature filtered through 63 lmnylon mesh washed twice with 02 M KCl purified over a one-step18 (wv) sucrose gradient and subsequently washed three timeswith protoplast culture medium (PCM) containing 43 g l1 MS salts(without vitamins) supplemented with 1 mg l1 thiamine-HCl100 mg l1 myo-inositol 10 g l1 sucrose 255 mg l1 KH2PO4and 04 M D-mannitol at pH 57 Elongation growth-inducingPCM contained 01 mg l1 1-naphthalene-acetic acid (NAA) and10 mg l1 benzyl-adenine (BA) Protoplasts were cultured in 3 mlliquid medium at a density of approximately 105 ml1 in smallsealed Petri dishes at 25 8C in the dark (Kuss and Cyr 1992)
Protoplast assay growth parameter measurements
Growth parameter measurements were performed on random photo-graphs of protoplast-derived cells after 4 d of culture Viable cellswere selected for measurements using FDA (fluorescein-diacetate)staining (Fig 1) Fluorescent images were captured using a cooledCCD camera mounted on a Leica DMR microscope with a 320objective The digital fluorescent images facilitated computer-basedmorphometric measurements using the NIH-IMAGE program(httprsbinfonihgovnih-image) in which objects can be contouredby applying the invertthreshold option The parameters measuredwere number of cells per file cell width and cell file length For eachsample the average cell division frequency was calculated as (totalnumber of cellsnumber of cell files)ndash1 The cell file length was firstexpressed in width units by calculating the lengthwidth ratio forindividual cells and then averaged over 100ndash150 cells per sampleElongation growth was calculated as increase in cell file length duringthe culture time as cell file lengthendndashcell file lengthbegin where cellfile lengthbegin=1 for a spherical protoplast Values presented in thegraphs represent average values and standard deviations for eachlinecondition calculated over a number of independent repetitionsas indicated in the textIn order to determine elongation growth capacity of the six
individual transgenic lines protoplasts from the wild-type and theNt1ndashNt6 cell lines were cultured for 4 d in PCM in the presence of10 mg l1 BA and 01 mg l1 NAA At least five independent ex-periments were performed Pairwise comparisons between trans-genic lines and the wild type were performed using a two-tailedStudentrsquos t test Significance values were adjusted for multiple tests
RNA gel blot analysis
Total RNA was isolated using the TRIzol method (GibcoBRL) A16 lg aliquot of total RNA was subjected to electrophoresis ona 1 agarose gel in 001 M NaH2PO4 (pH 70) using the glyoxalDMSO method RNA was subsequently transferred to a genescreenmembrane in 203 SSC RNA gel blots were hybridized with radio-labelled PCR fragments of the respective transcripts in formamidehybridization buffer overnight at 42 8C Autoradiograms were obtainedusing a Molecular Dynamics phosphorimager (Sunnyvale CA USA)
3272 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Reverse transcriptase-mediated PCR
Total RNA was isolated using the TRIzol method (GibcoBRL) AfterDNase I (Promega) treatment to remove chromosomal DNA cDNAis synthesized from 25 lg of total RNA in a volume of 20 ll [10 mMTRIS-HCl pH 88 50 mM KCl 5 mM MgCl2 1 mM dNTPs 1 lgoligo-dT(12)V anchor primer 20 U RNA guard (Pharmacia) and200 U MuMLV reverse transcriptase (RT Stratagene)] The sampleswere incubated for 1 h at 37 8C and subsequently at 95 8C for 5 minto inactivate the enzyme The samples were then diluted to 100 lland 1 or 2 ll of the cDNA were used for PCR analysis [10 mMTRIS-HCl pH 83 50 mM KCl 25 mM MgCl2 100 lM dNTPs50 ng primer and 1 U Taq polymerase (Boehringer MannheimUSA) in a total volume of 50 ll]Primer sets were designed for RT-mediated PCR-based transcript
quantification for each of the genes analysed Specificities of theprimer sets were verified by sequencing the RT-PCR products Thenumber of PCR cycles was adapted to the linear range of the PCRamplification reaction for each gene corresponding to the relativeexpression levels All samples were normalized on ubiquitin levelsThe following primers were used for RT-PCR UBI-f 59-ATGCA-GAT(CT)TTTGTGAAGAC-39 UBI-r 59-ACCACCACG(GA)A-GACGGAG-39 ACS-f 59-GATTTAATACAAGAATGGG-39 ACS-r59-GAACAATGAAAAGAACAAC-39 ACO-f 59-GGGCTTCTTTGAGTTGGTG-39 ACO-r 59-CTCCGCTGCCTCTTTCTC-39 Ampli-fied DNA fragments were run on a 1 agarose gel alkaline blottedto Hybond-N+ membrane (Amersham Pharmacia) and hybridized toradiolabelled PCR fragments of the corresponding cloned cDNAsAutoradiograms were obtained by using a Molecular DynamicsphosphorimagerRACE-PCR on NtENOD40 transcripts was as follows cDNA was
synthesized from RNA isolated from the transgenic lines using theRACE-T anchor primer 59-CATCTAGAGGATCGAATTC-T(16)-39The PCR cycles were 94 8C for 5 min 30 cycles of 94 8C for 20 s50 8C for 20 s 72 8C for 30 s and a final extension at 72 8C for 5 minusing in the first run primers RACE-A 59-CATCTAGAGGATC-GAATTC-39 and reverse primer 59-CGGGATCCTAGTTGGAGT-GAATTAAGGA-39 and in the second run RACE-A primer andreverse primer 59-AAGCTTTTGGAGTCTTTCTTGGCCTTT-39After the second PCR the total RACE-PCR product mixture waspurified using a PCR purification kit (Boehringer) and was cloned inpGEM-T (Promega)
Dosendashresponse curves
Protoplasts from three lines (WT Nt1 and Nt2) were cultured for 4 din PCM with various concentrations of cytokinin (00 mg l1 BA01 mg l1 BA 05 mg l1 BA 10 mg l1 BA and 20 mg l1 BA)and a fixed concentration of auxin (01 mg l1 NAA) or with vari-ous concentrations of auxin (00 mg l1 NAA 005 mg l1 NAA01 mg l1 NAA 05 mg l1 NAA and 10 mg l1 NAA) and afixed concentration of cytokinin (10 mg l1 BA) The auxincytokinindosendashresponse curve (DRC) experiment was repeated six three andfive times for the wild-type Nt1 and Nt2 lines respectivelySignificant line (PL) dosage (PD) and the interaction between lineand dosage (PL3D) effects were obtained by two-way analysis ofvariance (ANOVA) using the SAS glm procedure (windows version91 SAS Inc North Carolina USA) The model applied wasyijk=l+Li+Dj+L3Dij+eijk where yijk is either the elongation growthor the cell division frequency from line i (i=1 2 3) dosage j ( j=1 5) and observation k (k=1 6) l represents the overall meanL is the main effect for lines D is the concentration or dosage ef-fect for BA or NAA L3D is the interaction effect and e is the stocha-stic error Differences of the least square means (LSMeans) for theline3dosage effects along with associated t-tests and P-values werecalculated For the DRC experiment of the wild type single factoranalysis was performed to estimate the significance of the dosage
effect using the model yjk=l+Dj+ejk Duncanrsquos multiple range testwas applied to compare the effects of the different concentrationsIn a similar set-up the ethylene perception blocker AgNO3 was ap-plied at a concentration range from 108 M to 105 M with 10-foldincrements to protoplasts of wild-type Nt1 and Nt2 lines culturedin PCM supplemented with 01 mg l1 NAA and 10 mg l1 BA Togive the appropriate final concentrations of AgNO3 30 ll of a serialdilution of AgNO3 in water was transferred to the culture mediumcontaining the protoplasts just before sealing the Petri dishes at thestart of the culture period Three independent sets of experimentswere performed Regression analysis of each line was performedusing SPSS 90 (SPSS Chicago IL USA) The slopes (means 6standard error) of the linear functions obtained for the lines Nt1 Nt2and wild type were 54500624000 (P=0041 R2=028) 90500613000 (P lt0001 R2=080) and 2790628300 (P=0923R2=0004) respectively
Ethylene measurements
For each line protoplasts were divided over six Petri dishes at thestart of the experiment and were cultured in parallel Each Petri dishwas sampled every 24 h for 7 d In order not to severely alteraccumulating ethylene levels gas samples of 1 ml from a totalof 30 ml headspace volume were taken with a syringe through arubber gasket in the lid of the Petri dish without opening thesealed Petri dishes Ethylene concentration was determined directlyby standard GC-analysis on a gas chromatograph equipped withan alumina column and a flame ionization detector (Gilissen andHoekstra 1984) Ethylene accumulation at each time-point wasdetermined as the average ethylene concentration in the headspaceof these six cultures
Distribution of materials
Upon request all novel materials described in this publication willbe made available in a timely manner for non-commercial re-search purposes
Results
Cytokininndashauxin DRCs of BY-2 cells
To create a reference to determine the effect of NtENOD40overexpression on the phytohormone response of BY-2cells cytokinin and auxin DRCs of wild-type BY-2 cellswere made by measuring elongation growth and celldivision as a function of increasing cytokinin or auxinconcentration It has been shown previously that the mostaccurate data concerning cytokinin- and auxin-regulatedelongation growth and cell division in BY-2 cells areobtained with a bioassay starting from BY-2 protoplastswhich subsequently divide and elongate (Hasezawa andSyono 1983) The reason for using protoplasts in theseassays is that the composition of the BY-2 cell suspensionis heterogeneous with respect to cell file length and numberof cells per file By preparing protoplasts from the cell sus-pension a population of single cells with a similar diameteris obtained Analysing growth parameters of these culturedprotoplasts has an advantage over using the cell suspensiondirectly as it allows the effects on elongation growth andcell division to be separated Firstly under the conditionsof the present study the number of cell files during culture
ENOD40 and ethylene biosynthesis kinetics 3273
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
remains similar to the number of protoplasts at the start ofthe experiment So cells remain attached to each other afterdivision Hence by starting from protoplasts the increasein the number of cells per cell file directly reflects thenumber of cell divisions that took place during the incu-bation time and this parameter is from here onwards calledthe lsquocell division frequencyrsquo This parameter is expressed asthe average number of cells per cell filendash1 ie culturedprotoplasts remain single cells when no cell division takesplace whereas finding two cells per file means that oneround of cell division has occurred during the incubationtime Secondly the width of cells remains similar duringculture to the diameter of protoplasts at the start of theincubation (Fig 1A B) This means that no radial ex-pansion growth occurs and the length of the cell files at theend of the culture period is a measure of elongation growthThis parameter is hereafter called the lsquocell file lengthrsquo andis expressed in width units Elongation growth is calculatedas the increase in cell file length during the culture timeDuring the incubation time of 4 d used in these bioas-says wild-type cells can become on average about four tofive times as long as a protoplast So the increase in celllength is about three to four times the initial size of aprotoplast
DRCs were made using protoplasts prepared from thewild-type cell line (see Materials and methods) For thecytokinin DRC elongation growth and cell division fre-quency were determined as a function of cytokinin (BA)concentration at a fixed concentration of auxin (01 mg l1
NAA) For the auxin DRC the same parameters weredetermined as a function of auxin (NAA) concentration at afixed concentration of cytokinin (10 mg l1 BA) (seeMaterials and methods) Although data for the wild-type line were obtained in parallel with that of the trans-genic lines (see below) the results for the wild-type line arediscussed first to illustrate the action of cytokinin and auxinin wild-type BY-2 cells The DRCs were made six timesin independent experiments for the wild-type line The re-sults from independent experiments were similar and theaverage value (6standard deviation) was calculated foreach parameter (Fig 2A B) The cytokinin DRC for elong-ation growth (Fig 2A) shows that elongation growth ismaximal when cells are grown in the absence of exogenouscytokinin Application of increasing concentrations of
Fig 1 Representative photographs of protoplasts and protoplast-derivedcells of wild-type and 35SNtENOD40 transgenic lines (A) Wild-typeprotoplasts directly after protoplast isolation (B) Wild-type cells after 4 dof culture in elongation growth-inducing medium (C) 35SNtENOD40(Nt1) cells after 4 d of culture showing a reduction in elongation growth(D) 35SNtENOD40 (Nt1) cells cultured for 4 d in the presence of 10 lMAgNO3 showing a recovery of elongation growth The pictures weretaken with a fluorescence microscope after FDA staining of cells This fac-ilitated selection of viable protoplasts for measurements and aided objectrecognition with the NIH-image software for quantification of growthparameters Scale bars=100 lm
3274 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
cytokinin gradually reduces cell file length of wild-typecells suggesting that addition of cytokinin has a mildnegative effect on elongation growth (one-way ANOVAPD=00985) Application of 2 mg l1 BA can provoke areduction of cell file length of 078 units (corresponding to a21 reduction of elongation growth) compared with thecell file length reached when cells are grown in the absenceof exogenous cytokinin Duncanrsquos multiple range testindicated that this difference is significant at the 005 levelThus exogenous cytokinin is not essential for elongationgrowth when auxin is present and exogenous applicationof cytokinin has a mild negative effect on elongationgrowth The auxin DRC for cell file length (Fig 2B) showsthat cell file length is not affected (one-way ANOVAPD=08991) by the concentration of auxin as in the absenceof auxin it is similar to that at the various concentrationsof exogenous auxin This shows that elongation growthneither requires auxin nor does auxin markedly affect itwhen cytokinin is applied to the medium These data showthat the presence of either cytokinin or auxin is sufficientto sustain the growth rates achieved under the cultureconditions of the present study and that only addition ofcytokinin has a mild negative effect on elongation growth
The cytokinin DRC for cell division frequency (Fig 2A)shows that the cell division frequency is reduced from078 to 012 at increasing concentrations of cytokinin andthat the cell division frequency is highest in the absenceof exogenous cytokinin The auxin DRC for cell divisionfrequency (Fig 2B) shows that the cell division frequencyincreases from 000 to 064 at increasing concentrationsof auxin One-way ANOVA analysis indicated that thecell division responses provoked by cytokinin and auxinare highly significant (PD lt00001) In the absence ofauxin all cell files still consist of a single cell after 4 d ofculture which means that cells have not divided during
the culture period These results show that exogenouslyapplied auxin is essential for cell division in BY-2 cellsTaken together these observations show that exogenouslyapplied cytokinin has an inhibitory effect on cell divi-sion and that exogenous auxin has a stimulating effect oncell division whereas elongation growth is only mildlyreduced by exogenous application of cytokinin in wild-typeBY-2 cells
Generation of stably transformed BY-2 cell linescarrying 35SNtENOD40
To determine whether overexpression of NtENOD40 af-fects responses to phytohormones in BY-2 cells a set ofsix independent 35SNtENOD40 BY-2 cell lines calledlines Nt1 to Nt6 was generated by Agrobacterium-mediatedtransformation (see Materials and methods) Expression ofthe transgenes was determined by RNA gel blot analysis(Fig 3A) In the wild-type line NtENOD40 mRNA couldnot be detected indicating a very low expression level of theendogenous NtENOD40 gene In the six 35SNtENOD40lines NtENOD40 transcripts were expressed at variouslevels In all lines except line Nt6 this level was muchhigher than in the wild-type line with the highest expres-sion in lines Nt1 and Nt2 and the lowest expression in Nt5and Nt6 Further hybridization with the NtENOD40 proberesulted in two bands on the RNA gel blot To characterizethe nature of these two RNAs further 39-RACE-PCR wasperformed on NtENOD40 transcripts of the transgenic linesAnalysis of nucleotide sequences of 11 cloned RACE-PCR fragments revealed that all sequences are fully iden-tical to the transgene sequence and that read-through occursat the NOS terminator that flanks the NtENOD40 cDNAsequence in the construct resulting in transcripts of twodifferent lengths (data not shown)
Fig 2 Dosendashresponse curves for cytokinin and auxin in wild-type BY-2 cells (A) Wild-type elongation growth and cell division frequency atincreasing concentrations of BA each in the presence of 01 mg l1 NAA (B) Wild-type elongation growth and cell division frequency at increasingconcentrations of NAA each in the presence of 10 mg l1 BA Data represent average (6standard deviation) of six independent repetitions
ENOD40 and ethylene biosynthesis kinetics 3275
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

and vascular tissue development (Kouchi et al 1999Varkonyi-Gasic and White 2002 Vleghels et al 2003)indicates that the function of ENOD40 is not confinedto nodule development in leguminous species and suggeststhat ENOD40 has a general role in plant development Thisnotion is supported by the observation that ectopic expres-sion of ENOD40 affected formation of somatic embryosof alfalfa under in vitro culture conditions (Crespi et al1994) Also overexpression of ENOD40 led to reducedapical dominance in tobacco (van de Sande et al 1996)Both observations indicate that phytohormone signalling isaffected by ENOD40 Up to now the function of ENOD40and its mode of action have been poorly understoodAlthough observations in both legumes and non-legumesare pointing to a cross-talk between ENOD40 activity andphytohormone signalling pathways direct evidence forsuch an interaction is lacking Establishing whether thefunction of ENOD40 involves interaction with phyto-hormone signalling pathways could be an important steptowards unravelling the role of ENOD40 during organo-genesis Therefore a search was made for a system thatwould make it possible to test whether cross-talk betweenENOD40 and phytohormone signalling occurs The to-bacco Bright Yellow-2 (BY-2) cell suspension was chosenas a model system as it is convenient for studying phyto-hormone responses on a cellular level In BY-2 cellselongation growth and cell division are regulated by thebalance between cytokinin and auxin in the culture medium(Hasezawa and Syono 1983) Thus cell elongation growthand cell division frequency can be used as morphologicalmarkers to study whether overexpression of ENOD40affects the response of BY-2 cells to phytohormones Itwas found that overexpression of ENOD40 negatively af-fects cell elongation growth whereas cytokinin- or auxin-dependent control of cell division frequency is not affectedin 35SNtENOD40 transgenic cell lines It was shownfurther that the altered ethylene biosynthesis kinetics ob-served in ENOD40-overexpressing cells is a primary causeof the reduction in cell elongation growth
Materials and methods
Construction of binary vector p35SNtENOD40
Nicotiana tabacum contains two ENOD40 homologues that are96 identical at the nucleotide level (Matvienko et al 1996) Thecauliflower mosaic virus 35S promoter from pMON999 (Monsanto)was transferred to pCambia 1390 (Cambia Australia) yieldingp35STnos A 470 bp PCR fragment corresponding to the Nt-ENOD40-1 cDNA sequence was then cloned in p35STnos
Liquid BY-2 cultures and BY-2 transformation
Nicotiana tabacum BY-2 cell suspensions were subcultured weeklyby 403 dilution in fresh medium (Nagata et al 1981) BY-2transformation was performed using a modification of the procedurereported by Gu and Verma (1997) Five millilitres of a 3-d-old BY-2cell suspension was co-cultivated for 2 d at 25 8C in the dark with
60 ll of log-phase Agrobacterium tumefaciens strain C58C1harbouring the binary vector Cells were washed three times beforeplating on culture medium supplemented with 08 Daishin agar200 lg l1 ticarcilineclavuline and 40 lg l1 hygromycin BTransgenic calli that appeared after 3ndash4 weeks were cultured on freshselection plates for 1 more week and were subsequently transferredto liquid selection medium Six independent 35SNtENOD40 BY-2cell lines were generated and named lines Nt1 to Nt6 Each transgenicline was derived from a different callus which means that they can-not be siblings Transgenic lines were continuously maintained inselection medium
Protoplast isolation
Protoplasts were obtained from 6-d-old suspension cultures using1 (wv) cellulase-YC and 01 (wv) pectolyase Y23 in 04 MD-mannitol pH 55 (Nagata et al 1981) Cells were incubated in theenzyme solution for 3 h at room temperature filtered through 63 lmnylon mesh washed twice with 02 M KCl purified over a one-step18 (wv) sucrose gradient and subsequently washed three timeswith protoplast culture medium (PCM) containing 43 g l1 MS salts(without vitamins) supplemented with 1 mg l1 thiamine-HCl100 mg l1 myo-inositol 10 g l1 sucrose 255 mg l1 KH2PO4and 04 M D-mannitol at pH 57 Elongation growth-inducingPCM contained 01 mg l1 1-naphthalene-acetic acid (NAA) and10 mg l1 benzyl-adenine (BA) Protoplasts were cultured in 3 mlliquid medium at a density of approximately 105 ml1 in smallsealed Petri dishes at 25 8C in the dark (Kuss and Cyr 1992)
Protoplast assay growth parameter measurements
Growth parameter measurements were performed on random photo-graphs of protoplast-derived cells after 4 d of culture Viable cellswere selected for measurements using FDA (fluorescein-diacetate)staining (Fig 1) Fluorescent images were captured using a cooledCCD camera mounted on a Leica DMR microscope with a 320objective The digital fluorescent images facilitated computer-basedmorphometric measurements using the NIH-IMAGE program(httprsbinfonihgovnih-image) in which objects can be contouredby applying the invertthreshold option The parameters measuredwere number of cells per file cell width and cell file length For eachsample the average cell division frequency was calculated as (totalnumber of cellsnumber of cell files)ndash1 The cell file length was firstexpressed in width units by calculating the lengthwidth ratio forindividual cells and then averaged over 100ndash150 cells per sampleElongation growth was calculated as increase in cell file length duringthe culture time as cell file lengthendndashcell file lengthbegin where cellfile lengthbegin=1 for a spherical protoplast Values presented in thegraphs represent average values and standard deviations for eachlinecondition calculated over a number of independent repetitionsas indicated in the textIn order to determine elongation growth capacity of the six
individual transgenic lines protoplasts from the wild-type and theNt1ndashNt6 cell lines were cultured for 4 d in PCM in the presence of10 mg l1 BA and 01 mg l1 NAA At least five independent ex-periments were performed Pairwise comparisons between trans-genic lines and the wild type were performed using a two-tailedStudentrsquos t test Significance values were adjusted for multiple tests
RNA gel blot analysis
Total RNA was isolated using the TRIzol method (GibcoBRL) A16 lg aliquot of total RNA was subjected to electrophoresis ona 1 agarose gel in 001 M NaH2PO4 (pH 70) using the glyoxalDMSO method RNA was subsequently transferred to a genescreenmembrane in 203 SSC RNA gel blots were hybridized with radio-labelled PCR fragments of the respective transcripts in formamidehybridization buffer overnight at 42 8C Autoradiograms were obtainedusing a Molecular Dynamics phosphorimager (Sunnyvale CA USA)
3272 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Reverse transcriptase-mediated PCR
Total RNA was isolated using the TRIzol method (GibcoBRL) AfterDNase I (Promega) treatment to remove chromosomal DNA cDNAis synthesized from 25 lg of total RNA in a volume of 20 ll [10 mMTRIS-HCl pH 88 50 mM KCl 5 mM MgCl2 1 mM dNTPs 1 lgoligo-dT(12)V anchor primer 20 U RNA guard (Pharmacia) and200 U MuMLV reverse transcriptase (RT Stratagene)] The sampleswere incubated for 1 h at 37 8C and subsequently at 95 8C for 5 minto inactivate the enzyme The samples were then diluted to 100 lland 1 or 2 ll of the cDNA were used for PCR analysis [10 mMTRIS-HCl pH 83 50 mM KCl 25 mM MgCl2 100 lM dNTPs50 ng primer and 1 U Taq polymerase (Boehringer MannheimUSA) in a total volume of 50 ll]Primer sets were designed for RT-mediated PCR-based transcript
quantification for each of the genes analysed Specificities of theprimer sets were verified by sequencing the RT-PCR products Thenumber of PCR cycles was adapted to the linear range of the PCRamplification reaction for each gene corresponding to the relativeexpression levels All samples were normalized on ubiquitin levelsThe following primers were used for RT-PCR UBI-f 59-ATGCA-GAT(CT)TTTGTGAAGAC-39 UBI-r 59-ACCACCACG(GA)A-GACGGAG-39 ACS-f 59-GATTTAATACAAGAATGGG-39 ACS-r59-GAACAATGAAAAGAACAAC-39 ACO-f 59-GGGCTTCTTTGAGTTGGTG-39 ACO-r 59-CTCCGCTGCCTCTTTCTC-39 Ampli-fied DNA fragments were run on a 1 agarose gel alkaline blottedto Hybond-N+ membrane (Amersham Pharmacia) and hybridized toradiolabelled PCR fragments of the corresponding cloned cDNAsAutoradiograms were obtained by using a Molecular DynamicsphosphorimagerRACE-PCR on NtENOD40 transcripts was as follows cDNA was
synthesized from RNA isolated from the transgenic lines using theRACE-T anchor primer 59-CATCTAGAGGATCGAATTC-T(16)-39The PCR cycles were 94 8C for 5 min 30 cycles of 94 8C for 20 s50 8C for 20 s 72 8C for 30 s and a final extension at 72 8C for 5 minusing in the first run primers RACE-A 59-CATCTAGAGGATC-GAATTC-39 and reverse primer 59-CGGGATCCTAGTTGGAGT-GAATTAAGGA-39 and in the second run RACE-A primer andreverse primer 59-AAGCTTTTGGAGTCTTTCTTGGCCTTT-39After the second PCR the total RACE-PCR product mixture waspurified using a PCR purification kit (Boehringer) and was cloned inpGEM-T (Promega)
Dosendashresponse curves
Protoplasts from three lines (WT Nt1 and Nt2) were cultured for 4 din PCM with various concentrations of cytokinin (00 mg l1 BA01 mg l1 BA 05 mg l1 BA 10 mg l1 BA and 20 mg l1 BA)and a fixed concentration of auxin (01 mg l1 NAA) or with vari-ous concentrations of auxin (00 mg l1 NAA 005 mg l1 NAA01 mg l1 NAA 05 mg l1 NAA and 10 mg l1 NAA) and afixed concentration of cytokinin (10 mg l1 BA) The auxincytokinindosendashresponse curve (DRC) experiment was repeated six three andfive times for the wild-type Nt1 and Nt2 lines respectivelySignificant line (PL) dosage (PD) and the interaction between lineand dosage (PL3D) effects were obtained by two-way analysis ofvariance (ANOVA) using the SAS glm procedure (windows version91 SAS Inc North Carolina USA) The model applied wasyijk=l+Li+Dj+L3Dij+eijk where yijk is either the elongation growthor the cell division frequency from line i (i=1 2 3) dosage j ( j=1 5) and observation k (k=1 6) l represents the overall meanL is the main effect for lines D is the concentration or dosage ef-fect for BA or NAA L3D is the interaction effect and e is the stocha-stic error Differences of the least square means (LSMeans) for theline3dosage effects along with associated t-tests and P-values werecalculated For the DRC experiment of the wild type single factoranalysis was performed to estimate the significance of the dosage
effect using the model yjk=l+Dj+ejk Duncanrsquos multiple range testwas applied to compare the effects of the different concentrationsIn a similar set-up the ethylene perception blocker AgNO3 was ap-plied at a concentration range from 108 M to 105 M with 10-foldincrements to protoplasts of wild-type Nt1 and Nt2 lines culturedin PCM supplemented with 01 mg l1 NAA and 10 mg l1 BA Togive the appropriate final concentrations of AgNO3 30 ll of a serialdilution of AgNO3 in water was transferred to the culture mediumcontaining the protoplasts just before sealing the Petri dishes at thestart of the culture period Three independent sets of experimentswere performed Regression analysis of each line was performedusing SPSS 90 (SPSS Chicago IL USA) The slopes (means 6standard error) of the linear functions obtained for the lines Nt1 Nt2and wild type were 54500624000 (P=0041 R2=028) 90500613000 (P lt0001 R2=080) and 2790628300 (P=0923R2=0004) respectively
Ethylene measurements
For each line protoplasts were divided over six Petri dishes at thestart of the experiment and were cultured in parallel Each Petri dishwas sampled every 24 h for 7 d In order not to severely alteraccumulating ethylene levels gas samples of 1 ml from a totalof 30 ml headspace volume were taken with a syringe through arubber gasket in the lid of the Petri dish without opening thesealed Petri dishes Ethylene concentration was determined directlyby standard GC-analysis on a gas chromatograph equipped withan alumina column and a flame ionization detector (Gilissen andHoekstra 1984) Ethylene accumulation at each time-point wasdetermined as the average ethylene concentration in the headspaceof these six cultures
Distribution of materials
Upon request all novel materials described in this publication willbe made available in a timely manner for non-commercial re-search purposes
Results
Cytokininndashauxin DRCs of BY-2 cells
To create a reference to determine the effect of NtENOD40overexpression on the phytohormone response of BY-2cells cytokinin and auxin DRCs of wild-type BY-2 cellswere made by measuring elongation growth and celldivision as a function of increasing cytokinin or auxinconcentration It has been shown previously that the mostaccurate data concerning cytokinin- and auxin-regulatedelongation growth and cell division in BY-2 cells areobtained with a bioassay starting from BY-2 protoplastswhich subsequently divide and elongate (Hasezawa andSyono 1983) The reason for using protoplasts in theseassays is that the composition of the BY-2 cell suspensionis heterogeneous with respect to cell file length and numberof cells per file By preparing protoplasts from the cell sus-pension a population of single cells with a similar diameteris obtained Analysing growth parameters of these culturedprotoplasts has an advantage over using the cell suspensiondirectly as it allows the effects on elongation growth andcell division to be separated Firstly under the conditionsof the present study the number of cell files during culture
ENOD40 and ethylene biosynthesis kinetics 3273
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
remains similar to the number of protoplasts at the start ofthe experiment So cells remain attached to each other afterdivision Hence by starting from protoplasts the increasein the number of cells per cell file directly reflects thenumber of cell divisions that took place during the incu-bation time and this parameter is from here onwards calledthe lsquocell division frequencyrsquo This parameter is expressed asthe average number of cells per cell filendash1 ie culturedprotoplasts remain single cells when no cell division takesplace whereas finding two cells per file means that oneround of cell division has occurred during the incubationtime Secondly the width of cells remains similar duringculture to the diameter of protoplasts at the start of theincubation (Fig 1A B) This means that no radial ex-pansion growth occurs and the length of the cell files at theend of the culture period is a measure of elongation growthThis parameter is hereafter called the lsquocell file lengthrsquo andis expressed in width units Elongation growth is calculatedas the increase in cell file length during the culture timeDuring the incubation time of 4 d used in these bioas-says wild-type cells can become on average about four tofive times as long as a protoplast So the increase in celllength is about three to four times the initial size of aprotoplast
DRCs were made using protoplasts prepared from thewild-type cell line (see Materials and methods) For thecytokinin DRC elongation growth and cell division fre-quency were determined as a function of cytokinin (BA)concentration at a fixed concentration of auxin (01 mg l1
NAA) For the auxin DRC the same parameters weredetermined as a function of auxin (NAA) concentration at afixed concentration of cytokinin (10 mg l1 BA) (seeMaterials and methods) Although data for the wild-type line were obtained in parallel with that of the trans-genic lines (see below) the results for the wild-type line arediscussed first to illustrate the action of cytokinin and auxinin wild-type BY-2 cells The DRCs were made six timesin independent experiments for the wild-type line The re-sults from independent experiments were similar and theaverage value (6standard deviation) was calculated foreach parameter (Fig 2A B) The cytokinin DRC for elong-ation growth (Fig 2A) shows that elongation growth ismaximal when cells are grown in the absence of exogenouscytokinin Application of increasing concentrations of
Fig 1 Representative photographs of protoplasts and protoplast-derivedcells of wild-type and 35SNtENOD40 transgenic lines (A) Wild-typeprotoplasts directly after protoplast isolation (B) Wild-type cells after 4 dof culture in elongation growth-inducing medium (C) 35SNtENOD40(Nt1) cells after 4 d of culture showing a reduction in elongation growth(D) 35SNtENOD40 (Nt1) cells cultured for 4 d in the presence of 10 lMAgNO3 showing a recovery of elongation growth The pictures weretaken with a fluorescence microscope after FDA staining of cells This fac-ilitated selection of viable protoplasts for measurements and aided objectrecognition with the NIH-image software for quantification of growthparameters Scale bars=100 lm
3274 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
cytokinin gradually reduces cell file length of wild-typecells suggesting that addition of cytokinin has a mildnegative effect on elongation growth (one-way ANOVAPD=00985) Application of 2 mg l1 BA can provoke areduction of cell file length of 078 units (corresponding to a21 reduction of elongation growth) compared with thecell file length reached when cells are grown in the absenceof exogenous cytokinin Duncanrsquos multiple range testindicated that this difference is significant at the 005 levelThus exogenous cytokinin is not essential for elongationgrowth when auxin is present and exogenous applicationof cytokinin has a mild negative effect on elongationgrowth The auxin DRC for cell file length (Fig 2B) showsthat cell file length is not affected (one-way ANOVAPD=08991) by the concentration of auxin as in the absenceof auxin it is similar to that at the various concentrationsof exogenous auxin This shows that elongation growthneither requires auxin nor does auxin markedly affect itwhen cytokinin is applied to the medium These data showthat the presence of either cytokinin or auxin is sufficientto sustain the growth rates achieved under the cultureconditions of the present study and that only addition ofcytokinin has a mild negative effect on elongation growth
The cytokinin DRC for cell division frequency (Fig 2A)shows that the cell division frequency is reduced from078 to 012 at increasing concentrations of cytokinin andthat the cell division frequency is highest in the absenceof exogenous cytokinin The auxin DRC for cell divisionfrequency (Fig 2B) shows that the cell division frequencyincreases from 000 to 064 at increasing concentrationsof auxin One-way ANOVA analysis indicated that thecell division responses provoked by cytokinin and auxinare highly significant (PD lt00001) In the absence ofauxin all cell files still consist of a single cell after 4 d ofculture which means that cells have not divided during
the culture period These results show that exogenouslyapplied auxin is essential for cell division in BY-2 cellsTaken together these observations show that exogenouslyapplied cytokinin has an inhibitory effect on cell divi-sion and that exogenous auxin has a stimulating effect oncell division whereas elongation growth is only mildlyreduced by exogenous application of cytokinin in wild-typeBY-2 cells
Generation of stably transformed BY-2 cell linescarrying 35SNtENOD40
To determine whether overexpression of NtENOD40 af-fects responses to phytohormones in BY-2 cells a set ofsix independent 35SNtENOD40 BY-2 cell lines calledlines Nt1 to Nt6 was generated by Agrobacterium-mediatedtransformation (see Materials and methods) Expression ofthe transgenes was determined by RNA gel blot analysis(Fig 3A) In the wild-type line NtENOD40 mRNA couldnot be detected indicating a very low expression level of theendogenous NtENOD40 gene In the six 35SNtENOD40lines NtENOD40 transcripts were expressed at variouslevels In all lines except line Nt6 this level was muchhigher than in the wild-type line with the highest expres-sion in lines Nt1 and Nt2 and the lowest expression in Nt5and Nt6 Further hybridization with the NtENOD40 proberesulted in two bands on the RNA gel blot To characterizethe nature of these two RNAs further 39-RACE-PCR wasperformed on NtENOD40 transcripts of the transgenic linesAnalysis of nucleotide sequences of 11 cloned RACE-PCR fragments revealed that all sequences are fully iden-tical to the transgene sequence and that read-through occursat the NOS terminator that flanks the NtENOD40 cDNAsequence in the construct resulting in transcripts of twodifferent lengths (data not shown)
Fig 2 Dosendashresponse curves for cytokinin and auxin in wild-type BY-2 cells (A) Wild-type elongation growth and cell division frequency atincreasing concentrations of BA each in the presence of 01 mg l1 NAA (B) Wild-type elongation growth and cell division frequency at increasingconcentrations of NAA each in the presence of 10 mg l1 BA Data represent average (6standard deviation) of six independent repetitions
ENOD40 and ethylene biosynthesis kinetics 3275
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

Reverse transcriptase-mediated PCR
Total RNA was isolated using the TRIzol method (GibcoBRL) AfterDNase I (Promega) treatment to remove chromosomal DNA cDNAis synthesized from 25 lg of total RNA in a volume of 20 ll [10 mMTRIS-HCl pH 88 50 mM KCl 5 mM MgCl2 1 mM dNTPs 1 lgoligo-dT(12)V anchor primer 20 U RNA guard (Pharmacia) and200 U MuMLV reverse transcriptase (RT Stratagene)] The sampleswere incubated for 1 h at 37 8C and subsequently at 95 8C for 5 minto inactivate the enzyme The samples were then diluted to 100 lland 1 or 2 ll of the cDNA were used for PCR analysis [10 mMTRIS-HCl pH 83 50 mM KCl 25 mM MgCl2 100 lM dNTPs50 ng primer and 1 U Taq polymerase (Boehringer MannheimUSA) in a total volume of 50 ll]Primer sets were designed for RT-mediated PCR-based transcript
quantification for each of the genes analysed Specificities of theprimer sets were verified by sequencing the RT-PCR products Thenumber of PCR cycles was adapted to the linear range of the PCRamplification reaction for each gene corresponding to the relativeexpression levels All samples were normalized on ubiquitin levelsThe following primers were used for RT-PCR UBI-f 59-ATGCA-GAT(CT)TTTGTGAAGAC-39 UBI-r 59-ACCACCACG(GA)A-GACGGAG-39 ACS-f 59-GATTTAATACAAGAATGGG-39 ACS-r59-GAACAATGAAAAGAACAAC-39 ACO-f 59-GGGCTTCTTTGAGTTGGTG-39 ACO-r 59-CTCCGCTGCCTCTTTCTC-39 Ampli-fied DNA fragments were run on a 1 agarose gel alkaline blottedto Hybond-N+ membrane (Amersham Pharmacia) and hybridized toradiolabelled PCR fragments of the corresponding cloned cDNAsAutoradiograms were obtained by using a Molecular DynamicsphosphorimagerRACE-PCR on NtENOD40 transcripts was as follows cDNA was
synthesized from RNA isolated from the transgenic lines using theRACE-T anchor primer 59-CATCTAGAGGATCGAATTC-T(16)-39The PCR cycles were 94 8C for 5 min 30 cycles of 94 8C for 20 s50 8C for 20 s 72 8C for 30 s and a final extension at 72 8C for 5 minusing in the first run primers RACE-A 59-CATCTAGAGGATC-GAATTC-39 and reverse primer 59-CGGGATCCTAGTTGGAGT-GAATTAAGGA-39 and in the second run RACE-A primer andreverse primer 59-AAGCTTTTGGAGTCTTTCTTGGCCTTT-39After the second PCR the total RACE-PCR product mixture waspurified using a PCR purification kit (Boehringer) and was cloned inpGEM-T (Promega)
Dosendashresponse curves
Protoplasts from three lines (WT Nt1 and Nt2) were cultured for 4 din PCM with various concentrations of cytokinin (00 mg l1 BA01 mg l1 BA 05 mg l1 BA 10 mg l1 BA and 20 mg l1 BA)and a fixed concentration of auxin (01 mg l1 NAA) or with vari-ous concentrations of auxin (00 mg l1 NAA 005 mg l1 NAA01 mg l1 NAA 05 mg l1 NAA and 10 mg l1 NAA) and afixed concentration of cytokinin (10 mg l1 BA) The auxincytokinindosendashresponse curve (DRC) experiment was repeated six three andfive times for the wild-type Nt1 and Nt2 lines respectivelySignificant line (PL) dosage (PD) and the interaction between lineand dosage (PL3D) effects were obtained by two-way analysis ofvariance (ANOVA) using the SAS glm procedure (windows version91 SAS Inc North Carolina USA) The model applied wasyijk=l+Li+Dj+L3Dij+eijk where yijk is either the elongation growthor the cell division frequency from line i (i=1 2 3) dosage j ( j=1 5) and observation k (k=1 6) l represents the overall meanL is the main effect for lines D is the concentration or dosage ef-fect for BA or NAA L3D is the interaction effect and e is the stocha-stic error Differences of the least square means (LSMeans) for theline3dosage effects along with associated t-tests and P-values werecalculated For the DRC experiment of the wild type single factoranalysis was performed to estimate the significance of the dosage
effect using the model yjk=l+Dj+ejk Duncanrsquos multiple range testwas applied to compare the effects of the different concentrationsIn a similar set-up the ethylene perception blocker AgNO3 was ap-plied at a concentration range from 108 M to 105 M with 10-foldincrements to protoplasts of wild-type Nt1 and Nt2 lines culturedin PCM supplemented with 01 mg l1 NAA and 10 mg l1 BA Togive the appropriate final concentrations of AgNO3 30 ll of a serialdilution of AgNO3 in water was transferred to the culture mediumcontaining the protoplasts just before sealing the Petri dishes at thestart of the culture period Three independent sets of experimentswere performed Regression analysis of each line was performedusing SPSS 90 (SPSS Chicago IL USA) The slopes (means 6standard error) of the linear functions obtained for the lines Nt1 Nt2and wild type were 54500624000 (P=0041 R2=028) 90500613000 (P lt0001 R2=080) and 2790628300 (P=0923R2=0004) respectively
Ethylene measurements
For each line protoplasts were divided over six Petri dishes at thestart of the experiment and were cultured in parallel Each Petri dishwas sampled every 24 h for 7 d In order not to severely alteraccumulating ethylene levels gas samples of 1 ml from a totalof 30 ml headspace volume were taken with a syringe through arubber gasket in the lid of the Petri dish without opening thesealed Petri dishes Ethylene concentration was determined directlyby standard GC-analysis on a gas chromatograph equipped withan alumina column and a flame ionization detector (Gilissen andHoekstra 1984) Ethylene accumulation at each time-point wasdetermined as the average ethylene concentration in the headspaceof these six cultures
Distribution of materials
Upon request all novel materials described in this publication willbe made available in a timely manner for non-commercial re-search purposes
Results
Cytokininndashauxin DRCs of BY-2 cells
To create a reference to determine the effect of NtENOD40overexpression on the phytohormone response of BY-2cells cytokinin and auxin DRCs of wild-type BY-2 cellswere made by measuring elongation growth and celldivision as a function of increasing cytokinin or auxinconcentration It has been shown previously that the mostaccurate data concerning cytokinin- and auxin-regulatedelongation growth and cell division in BY-2 cells areobtained with a bioassay starting from BY-2 protoplastswhich subsequently divide and elongate (Hasezawa andSyono 1983) The reason for using protoplasts in theseassays is that the composition of the BY-2 cell suspensionis heterogeneous with respect to cell file length and numberof cells per file By preparing protoplasts from the cell sus-pension a population of single cells with a similar diameteris obtained Analysing growth parameters of these culturedprotoplasts has an advantage over using the cell suspensiondirectly as it allows the effects on elongation growth andcell division to be separated Firstly under the conditionsof the present study the number of cell files during culture
ENOD40 and ethylene biosynthesis kinetics 3273
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
remains similar to the number of protoplasts at the start ofthe experiment So cells remain attached to each other afterdivision Hence by starting from protoplasts the increasein the number of cells per cell file directly reflects thenumber of cell divisions that took place during the incu-bation time and this parameter is from here onwards calledthe lsquocell division frequencyrsquo This parameter is expressed asthe average number of cells per cell filendash1 ie culturedprotoplasts remain single cells when no cell division takesplace whereas finding two cells per file means that oneround of cell division has occurred during the incubationtime Secondly the width of cells remains similar duringculture to the diameter of protoplasts at the start of theincubation (Fig 1A B) This means that no radial ex-pansion growth occurs and the length of the cell files at theend of the culture period is a measure of elongation growthThis parameter is hereafter called the lsquocell file lengthrsquo andis expressed in width units Elongation growth is calculatedas the increase in cell file length during the culture timeDuring the incubation time of 4 d used in these bioas-says wild-type cells can become on average about four tofive times as long as a protoplast So the increase in celllength is about three to four times the initial size of aprotoplast
DRCs were made using protoplasts prepared from thewild-type cell line (see Materials and methods) For thecytokinin DRC elongation growth and cell division fre-quency were determined as a function of cytokinin (BA)concentration at a fixed concentration of auxin (01 mg l1
NAA) For the auxin DRC the same parameters weredetermined as a function of auxin (NAA) concentration at afixed concentration of cytokinin (10 mg l1 BA) (seeMaterials and methods) Although data for the wild-type line were obtained in parallel with that of the trans-genic lines (see below) the results for the wild-type line arediscussed first to illustrate the action of cytokinin and auxinin wild-type BY-2 cells The DRCs were made six timesin independent experiments for the wild-type line The re-sults from independent experiments were similar and theaverage value (6standard deviation) was calculated foreach parameter (Fig 2A B) The cytokinin DRC for elong-ation growth (Fig 2A) shows that elongation growth ismaximal when cells are grown in the absence of exogenouscytokinin Application of increasing concentrations of
Fig 1 Representative photographs of protoplasts and protoplast-derivedcells of wild-type and 35SNtENOD40 transgenic lines (A) Wild-typeprotoplasts directly after protoplast isolation (B) Wild-type cells after 4 dof culture in elongation growth-inducing medium (C) 35SNtENOD40(Nt1) cells after 4 d of culture showing a reduction in elongation growth(D) 35SNtENOD40 (Nt1) cells cultured for 4 d in the presence of 10 lMAgNO3 showing a recovery of elongation growth The pictures weretaken with a fluorescence microscope after FDA staining of cells This fac-ilitated selection of viable protoplasts for measurements and aided objectrecognition with the NIH-image software for quantification of growthparameters Scale bars=100 lm
3274 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
cytokinin gradually reduces cell file length of wild-typecells suggesting that addition of cytokinin has a mildnegative effect on elongation growth (one-way ANOVAPD=00985) Application of 2 mg l1 BA can provoke areduction of cell file length of 078 units (corresponding to a21 reduction of elongation growth) compared with thecell file length reached when cells are grown in the absenceof exogenous cytokinin Duncanrsquos multiple range testindicated that this difference is significant at the 005 levelThus exogenous cytokinin is not essential for elongationgrowth when auxin is present and exogenous applicationof cytokinin has a mild negative effect on elongationgrowth The auxin DRC for cell file length (Fig 2B) showsthat cell file length is not affected (one-way ANOVAPD=08991) by the concentration of auxin as in the absenceof auxin it is similar to that at the various concentrationsof exogenous auxin This shows that elongation growthneither requires auxin nor does auxin markedly affect itwhen cytokinin is applied to the medium These data showthat the presence of either cytokinin or auxin is sufficientto sustain the growth rates achieved under the cultureconditions of the present study and that only addition ofcytokinin has a mild negative effect on elongation growth
The cytokinin DRC for cell division frequency (Fig 2A)shows that the cell division frequency is reduced from078 to 012 at increasing concentrations of cytokinin andthat the cell division frequency is highest in the absenceof exogenous cytokinin The auxin DRC for cell divisionfrequency (Fig 2B) shows that the cell division frequencyincreases from 000 to 064 at increasing concentrationsof auxin One-way ANOVA analysis indicated that thecell division responses provoked by cytokinin and auxinare highly significant (PD lt00001) In the absence ofauxin all cell files still consist of a single cell after 4 d ofculture which means that cells have not divided during
the culture period These results show that exogenouslyapplied auxin is essential for cell division in BY-2 cellsTaken together these observations show that exogenouslyapplied cytokinin has an inhibitory effect on cell divi-sion and that exogenous auxin has a stimulating effect oncell division whereas elongation growth is only mildlyreduced by exogenous application of cytokinin in wild-typeBY-2 cells
Generation of stably transformed BY-2 cell linescarrying 35SNtENOD40
To determine whether overexpression of NtENOD40 af-fects responses to phytohormones in BY-2 cells a set ofsix independent 35SNtENOD40 BY-2 cell lines calledlines Nt1 to Nt6 was generated by Agrobacterium-mediatedtransformation (see Materials and methods) Expression ofthe transgenes was determined by RNA gel blot analysis(Fig 3A) In the wild-type line NtENOD40 mRNA couldnot be detected indicating a very low expression level of theendogenous NtENOD40 gene In the six 35SNtENOD40lines NtENOD40 transcripts were expressed at variouslevels In all lines except line Nt6 this level was muchhigher than in the wild-type line with the highest expres-sion in lines Nt1 and Nt2 and the lowest expression in Nt5and Nt6 Further hybridization with the NtENOD40 proberesulted in two bands on the RNA gel blot To characterizethe nature of these two RNAs further 39-RACE-PCR wasperformed on NtENOD40 transcripts of the transgenic linesAnalysis of nucleotide sequences of 11 cloned RACE-PCR fragments revealed that all sequences are fully iden-tical to the transgene sequence and that read-through occursat the NOS terminator that flanks the NtENOD40 cDNAsequence in the construct resulting in transcripts of twodifferent lengths (data not shown)
Fig 2 Dosendashresponse curves for cytokinin and auxin in wild-type BY-2 cells (A) Wild-type elongation growth and cell division frequency atincreasing concentrations of BA each in the presence of 01 mg l1 NAA (B) Wild-type elongation growth and cell division frequency at increasingconcentrations of NAA each in the presence of 10 mg l1 BA Data represent average (6standard deviation) of six independent repetitions
ENOD40 and ethylene biosynthesis kinetics 3275
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

remains similar to the number of protoplasts at the start ofthe experiment So cells remain attached to each other afterdivision Hence by starting from protoplasts the increasein the number of cells per cell file directly reflects thenumber of cell divisions that took place during the incu-bation time and this parameter is from here onwards calledthe lsquocell division frequencyrsquo This parameter is expressed asthe average number of cells per cell filendash1 ie culturedprotoplasts remain single cells when no cell division takesplace whereas finding two cells per file means that oneround of cell division has occurred during the incubationtime Secondly the width of cells remains similar duringculture to the diameter of protoplasts at the start of theincubation (Fig 1A B) This means that no radial ex-pansion growth occurs and the length of the cell files at theend of the culture period is a measure of elongation growthThis parameter is hereafter called the lsquocell file lengthrsquo andis expressed in width units Elongation growth is calculatedas the increase in cell file length during the culture timeDuring the incubation time of 4 d used in these bioas-says wild-type cells can become on average about four tofive times as long as a protoplast So the increase in celllength is about three to four times the initial size of aprotoplast
DRCs were made using protoplasts prepared from thewild-type cell line (see Materials and methods) For thecytokinin DRC elongation growth and cell division fre-quency were determined as a function of cytokinin (BA)concentration at a fixed concentration of auxin (01 mg l1
NAA) For the auxin DRC the same parameters weredetermined as a function of auxin (NAA) concentration at afixed concentration of cytokinin (10 mg l1 BA) (seeMaterials and methods) Although data for the wild-type line were obtained in parallel with that of the trans-genic lines (see below) the results for the wild-type line arediscussed first to illustrate the action of cytokinin and auxinin wild-type BY-2 cells The DRCs were made six timesin independent experiments for the wild-type line The re-sults from independent experiments were similar and theaverage value (6standard deviation) was calculated foreach parameter (Fig 2A B) The cytokinin DRC for elong-ation growth (Fig 2A) shows that elongation growth ismaximal when cells are grown in the absence of exogenouscytokinin Application of increasing concentrations of
Fig 1 Representative photographs of protoplasts and protoplast-derivedcells of wild-type and 35SNtENOD40 transgenic lines (A) Wild-typeprotoplasts directly after protoplast isolation (B) Wild-type cells after 4 dof culture in elongation growth-inducing medium (C) 35SNtENOD40(Nt1) cells after 4 d of culture showing a reduction in elongation growth(D) 35SNtENOD40 (Nt1) cells cultured for 4 d in the presence of 10 lMAgNO3 showing a recovery of elongation growth The pictures weretaken with a fluorescence microscope after FDA staining of cells This fac-ilitated selection of viable protoplasts for measurements and aided objectrecognition with the NIH-image software for quantification of growthparameters Scale bars=100 lm
3274 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
cytokinin gradually reduces cell file length of wild-typecells suggesting that addition of cytokinin has a mildnegative effect on elongation growth (one-way ANOVAPD=00985) Application of 2 mg l1 BA can provoke areduction of cell file length of 078 units (corresponding to a21 reduction of elongation growth) compared with thecell file length reached when cells are grown in the absenceof exogenous cytokinin Duncanrsquos multiple range testindicated that this difference is significant at the 005 levelThus exogenous cytokinin is not essential for elongationgrowth when auxin is present and exogenous applicationof cytokinin has a mild negative effect on elongationgrowth The auxin DRC for cell file length (Fig 2B) showsthat cell file length is not affected (one-way ANOVAPD=08991) by the concentration of auxin as in the absenceof auxin it is similar to that at the various concentrationsof exogenous auxin This shows that elongation growthneither requires auxin nor does auxin markedly affect itwhen cytokinin is applied to the medium These data showthat the presence of either cytokinin or auxin is sufficientto sustain the growth rates achieved under the cultureconditions of the present study and that only addition ofcytokinin has a mild negative effect on elongation growth
The cytokinin DRC for cell division frequency (Fig 2A)shows that the cell division frequency is reduced from078 to 012 at increasing concentrations of cytokinin andthat the cell division frequency is highest in the absenceof exogenous cytokinin The auxin DRC for cell divisionfrequency (Fig 2B) shows that the cell division frequencyincreases from 000 to 064 at increasing concentrationsof auxin One-way ANOVA analysis indicated that thecell division responses provoked by cytokinin and auxinare highly significant (PD lt00001) In the absence ofauxin all cell files still consist of a single cell after 4 d ofculture which means that cells have not divided during
the culture period These results show that exogenouslyapplied auxin is essential for cell division in BY-2 cellsTaken together these observations show that exogenouslyapplied cytokinin has an inhibitory effect on cell divi-sion and that exogenous auxin has a stimulating effect oncell division whereas elongation growth is only mildlyreduced by exogenous application of cytokinin in wild-typeBY-2 cells
Generation of stably transformed BY-2 cell linescarrying 35SNtENOD40
To determine whether overexpression of NtENOD40 af-fects responses to phytohormones in BY-2 cells a set ofsix independent 35SNtENOD40 BY-2 cell lines calledlines Nt1 to Nt6 was generated by Agrobacterium-mediatedtransformation (see Materials and methods) Expression ofthe transgenes was determined by RNA gel blot analysis(Fig 3A) In the wild-type line NtENOD40 mRNA couldnot be detected indicating a very low expression level of theendogenous NtENOD40 gene In the six 35SNtENOD40lines NtENOD40 transcripts were expressed at variouslevels In all lines except line Nt6 this level was muchhigher than in the wild-type line with the highest expres-sion in lines Nt1 and Nt2 and the lowest expression in Nt5and Nt6 Further hybridization with the NtENOD40 proberesulted in two bands on the RNA gel blot To characterizethe nature of these two RNAs further 39-RACE-PCR wasperformed on NtENOD40 transcripts of the transgenic linesAnalysis of nucleotide sequences of 11 cloned RACE-PCR fragments revealed that all sequences are fully iden-tical to the transgene sequence and that read-through occursat the NOS terminator that flanks the NtENOD40 cDNAsequence in the construct resulting in transcripts of twodifferent lengths (data not shown)
Fig 2 Dosendashresponse curves for cytokinin and auxin in wild-type BY-2 cells (A) Wild-type elongation growth and cell division frequency atincreasing concentrations of BA each in the presence of 01 mg l1 NAA (B) Wild-type elongation growth and cell division frequency at increasingconcentrations of NAA each in the presence of 10 mg l1 BA Data represent average (6standard deviation) of six independent repetitions
ENOD40 and ethylene biosynthesis kinetics 3275
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

cytokinin gradually reduces cell file length of wild-typecells suggesting that addition of cytokinin has a mildnegative effect on elongation growth (one-way ANOVAPD=00985) Application of 2 mg l1 BA can provoke areduction of cell file length of 078 units (corresponding to a21 reduction of elongation growth) compared with thecell file length reached when cells are grown in the absenceof exogenous cytokinin Duncanrsquos multiple range testindicated that this difference is significant at the 005 levelThus exogenous cytokinin is not essential for elongationgrowth when auxin is present and exogenous applicationof cytokinin has a mild negative effect on elongationgrowth The auxin DRC for cell file length (Fig 2B) showsthat cell file length is not affected (one-way ANOVAPD=08991) by the concentration of auxin as in the absenceof auxin it is similar to that at the various concentrationsof exogenous auxin This shows that elongation growthneither requires auxin nor does auxin markedly affect itwhen cytokinin is applied to the medium These data showthat the presence of either cytokinin or auxin is sufficientto sustain the growth rates achieved under the cultureconditions of the present study and that only addition ofcytokinin has a mild negative effect on elongation growth
The cytokinin DRC for cell division frequency (Fig 2A)shows that the cell division frequency is reduced from078 to 012 at increasing concentrations of cytokinin andthat the cell division frequency is highest in the absenceof exogenous cytokinin The auxin DRC for cell divisionfrequency (Fig 2B) shows that the cell division frequencyincreases from 000 to 064 at increasing concentrationsof auxin One-way ANOVA analysis indicated that thecell division responses provoked by cytokinin and auxinare highly significant (PD lt00001) In the absence ofauxin all cell files still consist of a single cell after 4 d ofculture which means that cells have not divided during
the culture period These results show that exogenouslyapplied auxin is essential for cell division in BY-2 cellsTaken together these observations show that exogenouslyapplied cytokinin has an inhibitory effect on cell divi-sion and that exogenous auxin has a stimulating effect oncell division whereas elongation growth is only mildlyreduced by exogenous application of cytokinin in wild-typeBY-2 cells
Generation of stably transformed BY-2 cell linescarrying 35SNtENOD40
To determine whether overexpression of NtENOD40 af-fects responses to phytohormones in BY-2 cells a set ofsix independent 35SNtENOD40 BY-2 cell lines calledlines Nt1 to Nt6 was generated by Agrobacterium-mediatedtransformation (see Materials and methods) Expression ofthe transgenes was determined by RNA gel blot analysis(Fig 3A) In the wild-type line NtENOD40 mRNA couldnot be detected indicating a very low expression level of theendogenous NtENOD40 gene In the six 35SNtENOD40lines NtENOD40 transcripts were expressed at variouslevels In all lines except line Nt6 this level was muchhigher than in the wild-type line with the highest expres-sion in lines Nt1 and Nt2 and the lowest expression in Nt5and Nt6 Further hybridization with the NtENOD40 proberesulted in two bands on the RNA gel blot To characterizethe nature of these two RNAs further 39-RACE-PCR wasperformed on NtENOD40 transcripts of the transgenic linesAnalysis of nucleotide sequences of 11 cloned RACE-PCR fragments revealed that all sequences are fully iden-tical to the transgene sequence and that read-through occursat the NOS terminator that flanks the NtENOD40 cDNAsequence in the construct resulting in transcripts of twodifferent lengths (data not shown)
Fig 2 Dosendashresponse curves for cytokinin and auxin in wild-type BY-2 cells (A) Wild-type elongation growth and cell division frequency atincreasing concentrations of BA each in the presence of 01 mg l1 NAA (B) Wild-type elongation growth and cell division frequency at increasingconcentrations of NAA each in the presence of 10 mg l1 BA Data represent average (6standard deviation) of six independent repetitions
ENOD40 and ethylene biosynthesis kinetics 3275
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

Cytokininndashauxin DRCs of 35SNtENOD40 transgenicBY-2 cells
To select lines in which the effect of NtENOD40 over-expression on hormone responses can be studied first theeffect of ENOD40 overexpression was determined on
elongation growth and division frequency of cells grownin the presence of 10 mg l1 BA and 01 mg l1 NAAThese conditions represent the intersection of the two setsof conditions used in the DRCs (Fig 2) To this endprotoplasts were obtained from the wild-type and Nt1ndashNt6cell lines and subsequently cultured for 4 d in PCM (seeMaterials and methods) Representative photographs takendirectly after protoplast isolation as well as after 4 d ofculture of wild-type cells and cells of a transgenic line witha strong phenotype (line Nt1) are presented in Fig 1AndashCThese pictures show the typical elongated appearanceof wild-type cells after 4 d of culture whereas cells aremarkedly smaller in line Nt1 Wild-type and transgeniccells are of similar size during propagation of the cellsuspension culture and so the size of isolated protoplastsis equal Cell file length and cell division frequency weredetermined for each line in at least five independentexperiments (see Materials and methods) The resultsfrom independent experiments were similar and the aver-age value (6standard deviation) was calculated for eachparameter (Fig 3B C) It was found that four lines Nt1Nt2 Nt3 and Nt4 have a strongly reduced cell file length ascompared with the wild type (Fig 3B) Elongation growthin these lines ranged from 166 units (Nt1) to 084 units(Nt3) corresponding to 56 and 29 respectively of theelongation growth of the wild-type line and is statisticallysignificant (two-tailed Studentrsquos t-test P lt0001) for thesefour lines The elongation growth of lines Nt5 and Nt6 issimilar to the wild type (Fig 3B) The cell divisionfrequency in the wild-type line is 0156009 This meansthat about 15 of the cells have divided once The celldivision frequency in the transgenic lines is similar tothat of the wild type (Fig 3C) This shows that di-vision frequency is not affected in the transgenic linesTaken together these data show that the lines with thehighest expression level (Nt1 and Nt2) display the strongestphenotype and that in the lines with the lowest level ofNtENOD40 expression no phenotypical change is ob-served This indicates that the observed suppression ofelongation growth most likely is correlated to overexpres-sion of NtENOD40
To analyse the effect of NtENOD40 overexpression oncell division frequency and elongation growth in moredetail and to test for a possible effect of ENOD40 on auxinor cytokinin responses DRCs were made for the transgeniclines Nt1 and Nt2 the two lines that have the highest levelof NtENOD40 expression and a strong phenotype andare compared with those of the wild-type line (Fig 4AndashD)As in the wild-type line the cell division frequency inboth transgenic lines shows a strong dose-dependentresponse to both BA and NAA The division frequencydrops from 042 to 015 for Nt1 cells and from 070 to 018for Nt2 cells grown in the presence of 01 mg l1 NAA atincreasing concentrations of BA For the wild type thedivision frequency decreases from 078 to 012 under these
Fig 3 Transgene expression levels elongation growth and divisionfrequency of cultured cells of wild-type (Wt) and 35SNtENOD40 BY-2cell lines Growth parameters of protoplast-derived cells were determinedafter 4 d of culture in medium supplemented with 01 mg l1 NAA and10 mg l1 BA Data represent average (6standard deviation) of 11 (wildtype) eight (Nt1) nine (Nt2) and five (Nt3ndashNt6) independent repetitionsAn asterisk marks transgenic cell lines with a significant reduction ofelongation growth compared with the wild type (P lt0001) (A) The levelof transgene expression was determined at the start of protoplast cultureby RNA gel blot analysis Hybridization with the NtENOD40 probe andHPTII probes shows expression of the transgene transcripts Hybridiza-tion with the ubiquitin (UBI) probe was performed in order to compareloading of the separate samples RNA gel blot analysis was performed inthree independent experiments each time with similar results Onerepresentative set of data is presented (B) Elongation growth in wild-typeand 35SNtENOD40 cell lines (C) Division frequency in wild-type and35SNtENOD40 cell lines
3276 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
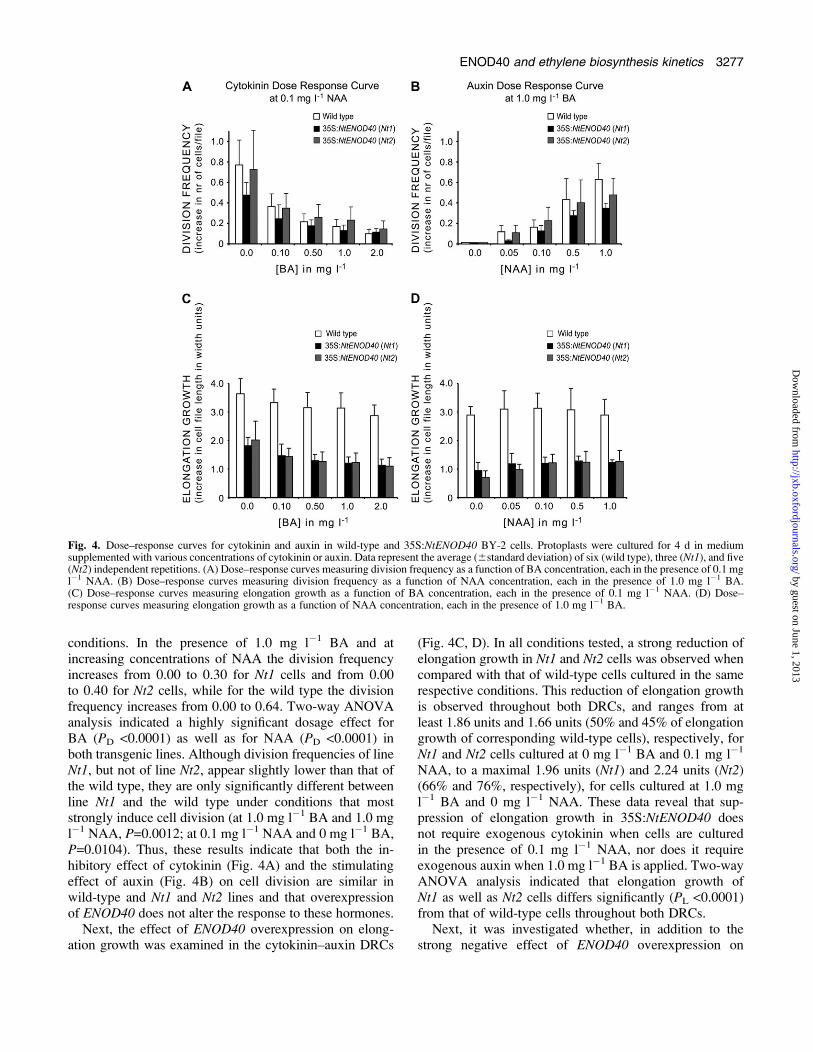
conditions In the presence of 10 mg l1 BA and atincreasing concentrations of NAA the division frequencyincreases from 000 to 030 for Nt1 cells and from 000to 040 for Nt2 cells while for the wild type the divisionfrequency increases from 000 to 064 Two-way ANOVAanalysis indicated a highly significant dosage effect forBA (PD lt00001) as well as for NAA (PD lt00001) inboth transgenic lines Although division frequencies of lineNt1 but not of line Nt2 appear slightly lower than that ofthe wild type they are only significantly different betweenline Nt1 and the wild type under conditions that moststrongly induce cell division (at 10 mg l1 BA and 10 mgl1 NAA P=00012 at 01 mg l1 NAA and 0 mg l1 BAP=00104) Thus these results indicate that both the in-hibitory effect of cytokinin (Fig 4A) and the stimulatingeffect of auxin (Fig 4B) on cell division are similar inwild-type and Nt1 and Nt2 lines and that overexpressionof ENOD40 does not alter the response to these hormones
Next the effect of ENOD40 overexpression on elong-ation growth was examined in the cytokininndashauxin DRCs
(Fig 4C D) In all conditions tested a strong reduction ofelongation growth in Nt1 and Nt2 cells was observed whencompared with that of wild-type cells cultured in the samerespective conditions This reduction of elongation growthis observed throughout both DRCs and ranges from atleast 186 units and 166 units (50 and 45 of elongationgrowth of corresponding wild-type cells) respectively forNt1 and Nt2 cells cultured at 0 mg l1 BA and 01 mg l1
NAA to a maximal 196 units (Nt1) and 224 units (Nt2)(66 and 76 respectively) for cells cultured at 10 mgl1 BA and 0 mg l1 NAA These data reveal that sup-pression of elongation growth in 35SNtENOD40 doesnot require exogenous cytokinin when cells are culturedin the presence of 01 mg l1 NAA nor does it requireexogenous auxin when 10 mg l1 BA is applied Two-wayANOVA analysis indicated that elongation growth ofNt1 as well as Nt2 cells differs significantly (PL lt00001)from that of wild-type cells throughout both DRCs
Next it was investigated whether in addition to thestrong negative effect of ENOD40 overexpression on
Fig 4 Dosendashresponse curves for cytokinin and auxin in wild-type and 35SNtENOD40 BY-2 cells Protoplasts were cultured for 4 d in mediumsupplemented with various concentrations of cytokinin or auxin Data represent the average (6standard deviation) of six (wild type) three (Nt1) and five(Nt2) independent repetitions (A) Dosendashresponse curves measuring division frequency as a function of BA concentration each in the presence of 01 mgl1 NAA (B) Dosendashresponse curves measuring division frequency as a function of NAA concentration each in the presence of 10 mg l1 BA(C) Dosendashresponse curves measuring elongation growth as a function of BA concentration each in the presence of 01 mg l1 NAA (D) Dosendashresponse curves measuring elongation growth as a function of NAA concentration each in the presence of 10 mg l1 BA
ENOD40 and ethylene biosynthesis kinetics 3277
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
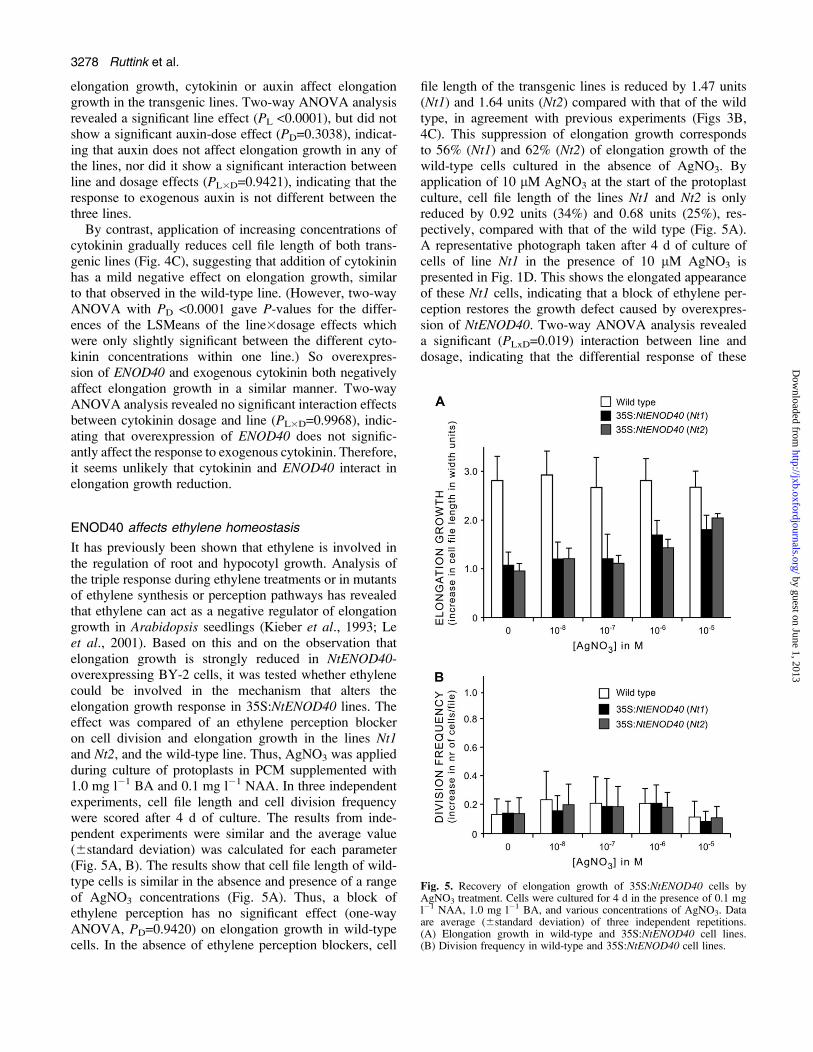
elongation growth cytokinin or auxin affect elongationgrowth in the transgenic lines Two-way ANOVA analysisrevealed a significant line effect (PL lt00001) but did notshow a significant auxin-dose effect (PD=03038) indicat-ing that auxin does not affect elongation growth in any ofthe lines nor did it show a significant interaction betweenline and dosage effects (PL3D=09421) indicating that theresponse to exogenous auxin is not different between thethree lines
By contrast application of increasing concentrations ofcytokinin gradually reduces cell file length of both trans-genic lines (Fig 4C) suggesting that addition of cytokininhas a mild negative effect on elongation growth similarto that observed in the wild-type line (However two-wayANOVA with PD lt00001 gave P-values for the differ-ences of the LSMeans of the line3dosage effects whichwere only slightly significant between the different cyto-kinin concentrations within one line) So overexpres-sion of ENOD40 and exogenous cytokinin both negativelyaffect elongation growth in a similar manner Two-wayANOVA analysis revealed no significant interaction effectsbetween cytokinin dosage and line (PL3D=09968) indic-ating that overexpression of ENOD40 does not signific-antly affect the response to exogenous cytokinin Thereforeit seems unlikely that cytokinin and ENOD40 interact inelongation growth reduction
ENOD40 affects ethylene homeostasis
It has previously been shown that ethylene is involved inthe regulation of root and hypocotyl growth Analysis ofthe triple response during ethylene treatments or in mutantsof ethylene synthesis or perception pathways has revealedthat ethylene can act as a negative regulator of elongationgrowth in Arabidopsis seedlings (Kieber et al 1993 Leet al 2001) Based on this and on the observation thatelongation growth is strongly reduced in NtENOD40-overexpressing BY-2 cells it was tested whether ethylenecould be involved in the mechanism that alters theelongation growth response in 35SNtENOD40 lines Theeffect was compared of an ethylene perception blockeron cell division and elongation growth in the lines Nt1and Nt2 and the wild-type line Thus AgNO3 was appliedduring culture of protoplasts in PCM supplemented with10 mg l1 BA and 01 mg l1 NAA In three independentexperiments cell file length and cell division frequencywere scored after 4 d of culture The results from inde-pendent experiments were similar and the average value(6standard deviation) was calculated for each parameter(Fig 5A B) The results show that cell file length of wild-type cells is similar in the absence and presence of a rangeof AgNO3 concentrations (Fig 5A) Thus a block ofethylene perception has no significant effect (one-wayANOVA PD=09420) on elongation growth in wild-typecells In the absence of ethylene perception blockers cell
file length of the transgenic lines is reduced by 147 units(Nt1) and 164 units (Nt2) compared with that of the wildtype in agreement with previous experiments (Figs 3B4C) This suppression of elongation growth correspondsto 56 (Nt1) and 62 (Nt2) of elongation growth of thewild-type cells cultured in the absence of AgNO3 Byapplication of 10 lM AgNO3 at the start of the protoplastculture cell file length of the lines Nt1 and Nt2 is onlyreduced by 092 units (34) and 068 units (25) res-pectively compared with that of the wild type (Fig 5A)A representative photograph taken after 4 d of culture ofcells of line Nt1 in the presence of 10 lM AgNO3 ispresented in Fig 1D This shows the elongated appearanceof these Nt1 cells indicating that a block of ethylene per-ception restores the growth defect caused by overexpres-sion of NtENOD40 Two-way ANOVA analysis revealeda significant (PLxD=0019) interaction between line anddosage indicating that the differential response of these
Fig 5 Recovery of elongation growth of 35SNtENOD40 cells byAgNO3 treatment Cells were cultured for 4 d in the presence of 01 mgl1 NAA 10 mg l1 BA and various concentrations of AgNO3 Dataare average (6standard deviation) of three independent repetitions(A) Elongation growth in wild-type and 35SNtENOD40 cell lines(B) Division frequency in wild-type and 35SNtENOD40 cell lines
3278 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
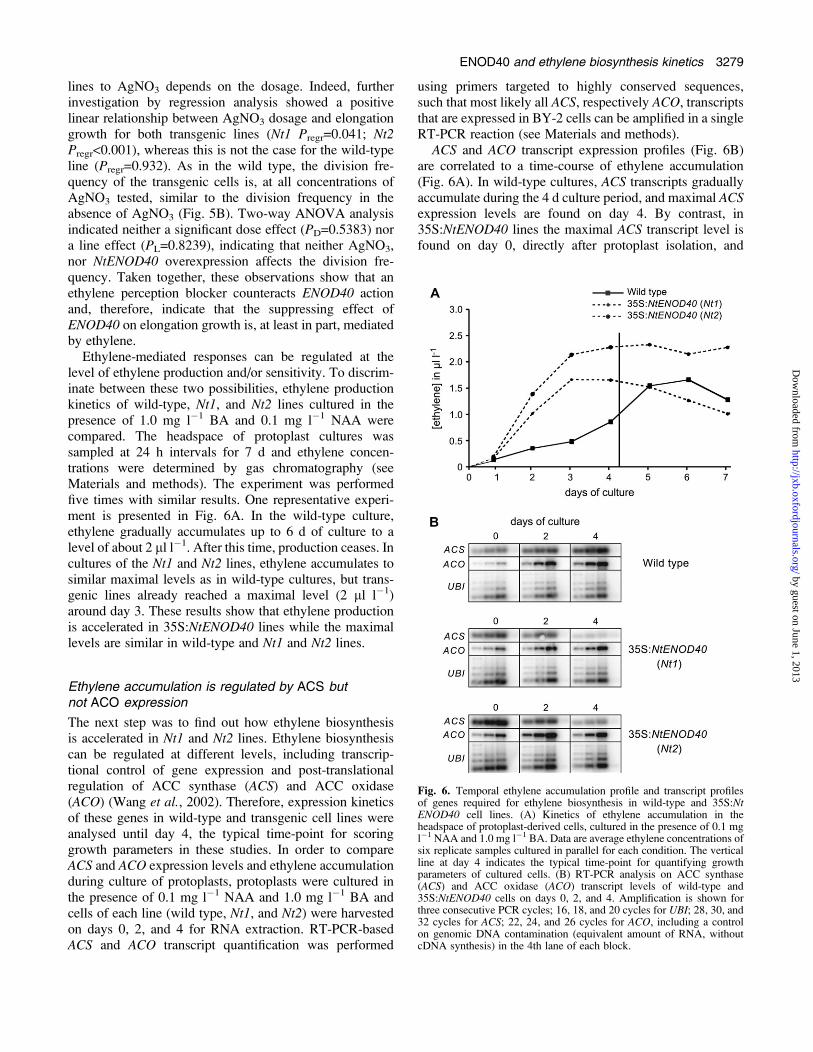
lines to AgNO3 depends on the dosage Indeed furtherinvestigation by regression analysis showed a positivelinear relationship between AgNO3 dosage and elongationgrowth for both transgenic lines (Nt1 Pregr=0041 Nt2Pregrlt0001) whereas this is not the case for the wild-typeline (Pregr=0932) As in the wild type the division fre-quency of the transgenic cells is at all concentrations ofAgNO3 tested similar to the division frequency in theabsence of AgNO3 (Fig 5B) Two-way ANOVA analysisindicated neither a significant dose effect (PD=05383) nora line effect (PL=08239) indicating that neither AgNO3nor NtENOD40 overexpression affects the division fre-quency Taken together these observations show that anethylene perception blocker counteracts ENOD40 actionand therefore indicate that the suppressing effect ofENOD40 on elongation growth is at least in part mediatedby ethylene
Ethylene-mediated responses can be regulated at thelevel of ethylene production andor sensitivity To discrim-inate between these two possibilities ethylene productionkinetics of wild-type Nt1 and Nt2 lines cultured in thepresence of 10 mg l1 BA and 01 mg l1 NAA werecompared The headspace of protoplast cultures wassampled at 24 h intervals for 7 d and ethylene concen-trations were determined by gas chromatography (seeMaterials and methods) The experiment was performedfive times with similar results One representative experi-ment is presented in Fig 6A In the wild-type cultureethylene gradually accumulates up to 6 d of culture to alevel of about 2 ll l1 After this time production ceases Incultures of the Nt1 and Nt2 lines ethylene accumulates tosimilar maximal levels as in wild-type cultures but trans-genic lines already reached a maximal level (2 ll l1)around day 3 These results show that ethylene productionis accelerated in 35SNtENOD40 lines while the maximallevels are similar in wild-type and Nt1 and Nt2 lines
Ethylene accumulation is regulated by ACS butnot ACO expression
The next step was to find out how ethylene biosynthesisis accelerated in Nt1 and Nt2 lines Ethylene biosynthesiscan be regulated at different levels including transcrip-tional control of gene expression and post-translationalregulation of ACC synthase (ACS) and ACC oxidase(ACO) (Wang et al 2002) Therefore expression kineticsof these genes in wild-type and transgenic cell lines wereanalysed until day 4 the typical time-point for scoringgrowth parameters in these studies In order to compareACS and ACO expression levels and ethylene accumulationduring culture of protoplasts protoplasts were cultured inthe presence of 01 mg l1 NAA and 10 mg l1 BA andcells of each line (wild type Nt1 and Nt2) were harvestedon days 0 2 and 4 for RNA extraction RT-PCR-basedACS and ACO transcript quantification was performed
using primers targeted to highly conserved sequencessuch that most likely all ACS respectively ACO transcriptsthat are expressed in BY-2 cells can be amplified in a singleRT-PCR reaction (see Materials and methods)
ACS and ACO transcript expression profiles (Fig 6B)are correlated to a time-course of ethylene accumulation(Fig 6A) In wild-type cultures ACS transcripts graduallyaccumulate during the 4 d culture period and maximal ACSexpression levels are found on day 4 By contrast in35SNtENOD40 lines the maximal ACS transcript level isfound on day 0 directly after protoplast isolation and
Fig 6 Temporal ethylene accumulation profile and transcript profilesof genes required for ethylene biosynthesis in wild-type and 35SNtENOD40 cell lines (A) Kinetics of ethylene accumulation in theheadspace of protoplast-derived cells cultured in the presence of 01 mgl1 NAA and 10 mg l1 BA Data are average ethylene concentrations ofsix replicate samples cultured in parallel for each condition The verticalline at day 4 indicates the typical time-point for quantifying growthparameters of cultured cells (B) RT-PCR analysis on ACC synthase(ACS) and ACC oxidase (ACO) transcript levels of wild-type and35SNtENOD40 cells on days 0 2 and 4 Amplification is shown forthree consecutive PCR cycles 16 18 and 20 cycles for UBI 28 30 and32 cycles for ACS 22 24 and 26 cycles for ACO including a controlon genomic DNA contamination (equivalent amount of RNA withoutcDNA synthesis) in the 4th lane of each block
ENOD40 and ethylene biosynthesis kinetics 3279
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

gradually decreases during the culture period (Fig 6B)These results show that in 35SNtENOD40 lines ACStranscripts accumulate at an earlier time-point and thisis consistent with the accelerated ethylene production Inwild-type cultures as well as in 35SNtENOD40 culturesACS expression profiles correlate with the timing ofethylene production In wild-type cultures ACO transcriptsare present directly after protoplast culture has started andtheir level only slightly increases during the 4 d cultureperiod The ACO transcript accumulation profiles in both35SNtENOD40 lines are similar to that in the wild type(Fig 6B) The temporal regulation of ACO transcript ac-cumulation does not correlate with the timing of ethyleneproduction in the different lines Since a tight correlationbetween ethylene biosynthesis and ACS but not ACOtranscript accumulation is found regulation of ethylenebiosynthesis can be largely attributed to transcriptionalregulation of ACS
Discussion
In this report the interaction of ENOD40 with hormonesignalling pathways was studied by analysing hormone-induced responses in tobacco BY-2 cell lines For thispurpose the BY-2 protoplast assay described by Hasezawaand Syono (1983) was adapted and the dependence of cellelongation growth and cell division on cytokinin auxinand ethylene in wild-type and transgenic 35SNtENOD40BY-2 cells was investigated
It was found in agreement with other studies in BY-2cells (Hasezawa and Syono 1983 Hasezawa et al 1988Nagata et al 1992) that cell division frequency is con-trolled by the cytokininauxin ratio Exogenous cytokininhas an inhibitory effect whereas auxin stimulates celldivision (Fig 2) The response to different cytokinin orauxin concentrations is comparable in wild-type and35SNtENOD40 lines (Fig 4) suggesting that ENOD40overexpression in BY-2 cells does not affect cytokinin-or auxin-dependent control on cell division In additionevidence is shown that under the conditions of the presentstudy alterations of ethylene production or perceptiondo not affect cell division frequency Similar cell divisionfrequencies are observed in wild-type and transgenic lines(Figs 3ndash5) despite the differences in their ethylene pro-duction levels during early days of protoplast culture (Fig6) Also no effect of ethylene perception blockers oncell division frequency was found (Fig 5) Therefore itis concluded that ethylene is not involved in the control ofcell division under the conditions of the present study
Elongation growth is not affected by exogenous auxin(Fig 2) nor is this process sensitive to application ofethylene perception blockers in wild-type BY-2 cells (Fig5) Thus elongation growth does not seem to be limitedby either of these two hormones in wild-type cells By
contrast cytokinin has a mild negative effect as addition ofcytokinin reduces elongation growth of wild-type cells ina dose-dependent manner (Fig 2)
Upon overexpression of NtENOD40 elongation growthis strongly reduced (Figs 3 4) Analysis of ethylenehomeostasis showed that ENOD40-provoked phenotypicresponses are mediated by ethylene Ethylene biosynthesisis accelerated in 35SNtENOD40 lines under conditionswhere ENOD40 reduces elongation growth Constitutiveexpression of ENOD40 does not lead to constitutiveethylene production but rather accelerates a transientaccumulation of ethylene during cell growth (Fig 6) Inaddition it was observed that the ENOD40-induced re-sponse is counteracted by an ethylene perception blocker(Fig 5) but this recovery appears not to be complete Thiscould imply that the complete block of ethylene percep-tion requires a higher AgNO3 concentration that might betoxic to BY-2 cells or that ethylene is not the only factorthat is responsible for reduced elongation growth in35SNtENOD40 cells As cytokinin also negatively af-fects elongation growth an effect of NtENOD40 overexpre-ssion on the endogenous cytokinin concentration cannotbe excluded as a factor by which overexpression of Nt-ENOD40 affects elongation growth However an increasein the endogenous cytokinin concentration is expected tosuppress elongation growth (Fig 4A) and cell division(Fig 4C) simultaneously Since cell division frequenciesare not significantly different between wild-type and trans-genic lines it appears unlikely that endogenous cytokininconcentrations are markedly affected by overexpressionof NtENOD40 Hence if besides ethylene a second factoris responsible for reduced elongation growth the nature ofthis factor remains unknown
Nevertheless the observations made in the present studythat ENOD40-provoked responses are mediated by ethyleneand that these responses can be counteracted by additionof an ethylene perception blocker show that acceleration ofethylene accumulation is a primary cause rather than a con-sequence of the reduced cell elongation in the transgeniclines Therefore it seems likely that altered regulation ofethylene biosynthesis is part of the mechanism that affectselongation growth in NtENOD40-overexpressing BY-2cells As accelerated accumulation of ACS transcript levelsis correlated with accelerated ethylene accumulation inENOD40-overexpressing BY-2 cells (Fig 6) ACS couldbe a primary inducer of the ethylene accumulation Thiswould be consistent with ACS as the rate-limiting step ofethylene biosynthesis (Yang and Hoffman 1984) anda well-known primary target of ethylene biosynthesis re-gulation (Yang and Hoffman 1984 Theologis 1992)However since ethylene accumulation is measured in-directly by analysing the gas phase above the cell cultureit cannot be excluded that the cells have been exposed toa higher ethylene concentration before ACS expression isup-regulated Since ethylene can induce ACS expression in
3280 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

the latter case ACS may act as a secondary inducer in apositive feedback mechanism
Although cytokinin and ENOD40 both negatively affectelongation growth providing the possibility that theirsignalling pathways share common downstream compo-nents no indication was found that interaction betweenthese factors indeed occurs This is based on the observa-tions that ENOD40-provoked growth suppression doesnot require exogenous cytokinin nor is cytokinin dose-dependent growth suppression significantly different in thetransgenic and wild-type lines (Fig 4C)
A major question now is whether the observations madein this cellular model system can be extrapolated to thewhole-plant level Ectopic expression of GmENOD40 inArabidopsis did not lead to severe changes in overall plantarchitecture but led to a decrease in cell size of epidermalinternode and leaf mesophyll cells (Guzzo et al 2005)In addition a subpopulation of protoplasts isolated fromArabidopsis cell suspension culture displayed reducedexpansion growth after either transient expression ofGmENOD40 or direct administration of GmENOD40 pe-ptides (Guzzo et al 2005) Thus these data suggest that thephenotypic effect of ectopic expression of ENOD40 maydepend on the cell type andor environmental or develop-mental context Upon inoculation of leguminous plantswith rhizobia ENOD40 is highly induced in cortical cellsseveral hours prior to the first cell division leading to noduleprimordium formation suggesting that ENOD40is involved in the control over cortical cell division InMedicago plants ectopically expressing MtENOD40 over-expression of MtENOD40 leads to proliferation of cellsin the upper region of the root (Charon et al 1997) How-ever upon inoculation of plants with the symbiontSinorhizobium meliloti cell proliferation is induced in theregion close to the root tip (Charon et al 1999) showingthat overexpression of ENOD40 does not lead to celldivision per se and that ENOD40 expression alone is notsufficient for nodule primordium formation Likewise inBY-2 cells overexpression of ENOD40 did not lead toan increase in cell division frequency Thus the pheno-typic response to ectopic expression of ENOD40 may bedependent on the cellular context andor on local actionof other plant factors such as phytohormones
Since ENOD40 is highly induced and acts as a positiveregulator during nodulation the observation that ENOD40overexpression leads to an acceleration of ethylene bio-synthesis in BY-2 cells appears counter-intuitive to thefunction of ethylene as a negative regulator of nodulationduring legumendashRhizobium interaction It has been demon-strated that ethylene acts at a multitude of steps in thenodulation pathway including nodule formation (Petersand Crist-Estes 1989 Lee and LaRue 1992) infectionthread formation (Penmetsa and Cook 1997) root hair de-formation early gene expression and calcium spiking inresponse to Nod factor (Oldroyd et al 2001) and regulation
of the maintenance of nodule meristems in Sesbania rost-rata (Fernandez-Lopez et al 1998) In addition ethylenehas a possible role in defining the position at which noduleprimordia are initiated (Heidstra et al 1997) Togetherthese observations have led to the hypothesis (Oldroydet al 2001) that ethylene could function as a dynamicregulator in the nodulation process by acting at an earlypoint in Nod factor signalling at or upstream of calciumspiking in response to Nod factor resulting in multipleethylene-regulated developmental effects downstream in theNod factor-dependent pathway Alternatively ethylenecould inhibit several components of the nodulation pathwaydirectly and independently (Oldroyd et al 2001) Althoughthe underlying mechanism remains unclear the presentstudies in BY-2 cells suggest appropriate timing of ethylenebiosynthesis as a critical factor for cellular responses andindicate that ENOD40 may participate in the control overthis timing In the absence of studies describing the effectof either endogenous or ectopic expression of ENOD40 onethylene biosynthesis the existence of these interactions inwhole-plant systems or indeed during nodule developmentstill needs to be confirmed Due to the diversity of possibleresponses induced by alterations of ethylene levels in whole-plant systems the phenotypic outcome of such interactionsis difficult to predict and may be dependent on the cellularcontext Nevertheless it is conceivable that local expressionof ENOD40 during early steps of the legumendashRhizobiuminteraction (Kouchi and Hata 1993 Yang et al 1993Compaan et al 2001) would provide the plant with a meansto attenuate in a dynamic manner ethylene production inkeeping with the proposed role of ethylene as a dynamicregulator of cellular responses (Oldroyd et al 2001) duringnodulation Therefore effects of ENOD40 on the ethylenebiosynthesis pathway could be considered as an importantcomponent of the complex regulatory pathway controllingnodule development
Acknowledgements
We thank Danny Geelen for kindly supplying the BY-2 cellsuspension We gratefully acknowledge Gerard van der Krogt forhis valuable contribution to the BY-2 transformations VeroniqueStorme for support with data analysis and the excellent technicalassistance of Ciska Braam and Maelle Lorvellec We are thankful toMark Hink and Jan-Willem Borst (Wageningen University Micro-scopy Center) for use of the microscope facilities This work wassupported by the Netherlands Organization for Scientific Research(NWO 80549004)
References
Charon C Johansson C Kondorosi E Kondorosi A Crespi M1997 ENOD40 induces dedifferentiation and division of rootcortical cells in legumes Proceedings of the National Academy ofSciences USA 94 8901ndash8906
Charon C Sousa C Crespi M Kondorosi A 1999 Alteration ofENOD40 expression modifies Medicago truncatula root nodule
ENOD40 and ethylene biosynthesis kinetics 3281
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from

development induced by Sinorhizobium meliloti The Plant Cell11 1953ndash1965
Compaan B Ruttink T Albrecht C Meeley R Bisseling TFranssen H 2003 Identification and characterisation of a Zeamays line carrying a transposon-tagged ENOD40 Biochimica etBiophysica Acta 1629 84ndash91
Compaan B Yang WC Bisseling T Franssen H 2001 ENOD40expression in the pericycle precedes cortical cell division inRhizobiumndashlegume interaction and the highly conserved internalregion of the gene does not encode a peptide Plant and Soil 2301ndash8
Cooper JB Long SR 1994 Morphogenetic rescue of Rhizobiummeliloti nodulation mutants by trans-zeatin secretion The PlantCell 6 215ndash225
Crespi MD Jurkevitch E Poiret M drsquoAubenton-Carafa YPetrovics G Kondorosi E Kondorosi A 1994 ENOD40a gene expressed during nodule organogenesis codes for a non-translatable RNA involved in plant growth EMBO Journal 135099ndash5112
Fernandez-Lopez M Goormachtig S Gao M DrsquoHaeze WVan Montagu M Holsters M 1998 Ethylene-mediated pheno-typic plasticity in root nodule development on Sesbaniarostrata Proceedings of the National Academy of SciencesUSA 95 12724ndash12728
Gilissen LJW Hoekstra FA 1984 Pollination-induced corollawilting in Petunia hybrida rapid transfer through the style ofa wilting-inducing substance Plant Physiology 75 496ndash498
Gu X Verma DPS 1997 Dynamics of phragmoplastin in livingcells during cell plate formation and uncoupling of cell elongationfrom the plane of cell division The Plant Cell 9 157ndash169
Guzzo F Portaluppi P Grisi R Barone S Zampieri SFranssen H Levi M 2005 Reduction of cell size inducedby enod40 inArabidopsis thaliana Journal of Experimental Botany56 507ndash513
Hasezawa S Syono K 1983 Hormonal control of elongation oftobacco cells derived from protoplasts Plant and Cell Physiology24 127ndash132
Hasezawa S Hogetsu T Syono K 1988 Rearrangement of corticalmicrotubules in elongating cells derived from tobacco protoplastsa time-course observation by immunofluorescence microscopyPlant Physiology 133 46ndash51
Heidstra R Yang WC Yalcin Y Peck S Emons AM vanKammen A Bisseling T 1997 Ethylene provides positionalinformation on cortical cell division but is not involved in Nodfactor-induced root hair tip growth in Rhizobiumndashlegume in-teraction Development 124 1781ndash1787
Hirsch AM Bhuvaneswari TV Torrey JG Bisseling T1989 Early nodulin genes are induced in alfalfa root outgrowthselicited by auxin transport inhibitors Proceedings of theNational Academy of Sciences USA 86 1244ndash1249
Kieber JJ Rothenberg M Roman G Feldmann KA Ecker JR1993 CTR1 a negative regulator of the ethylene response pathwayin Arabidopsis encodes a member of the raf family of proteinkinases Cell 72 427ndash441
Kouchi H Hata S 1993 Isolation and characterization of novelnodulin cDNAs representing genes expressed at early stages ofsoybean nodule development Molecular and General Genetics238 106ndash119
Kouchi H Takane K So RB Ladha JK Reddy PM 1999 RiceENOD40 isolation and expression analysis in rice and transgenicsoybean root nodules The Plant Journal 18 121ndash129
Kuss WCL Cyr RJ 1992 Tobacco protoplasts differentiate intoelongate cells without new microtubule depolymerization Proto-plasma 168 64ndash72
Larsen K 2003 Molecular cloning and characterisation of a cDNAencoding a ryegrass (Lolium perenne) ENOD40 homologueJournal of Plant Physiology 160 675ndash687
Le J Vandenbussche F Van Der Straeten D Verbelen J2001 In the early response of Arabidopsis roots to ethylene cellelongation is up- and down-regulated and uncoupled from dif-ferentiation Plant Physiology 125 519ndash522
Lee KH LaRue TA 1992 Exogenous ethylene inhibits nodula-tion of Pisum sativum L cv Sparkle Plant Physiology 1001759ndash1763
Mathesius U Charon C Rolfe BG Kondorosi A Crespi M2000 Temporal and spatial order of events during the induc-tion of cortical cell divisions in white clover by Rhizobiumleguminosarum bv trifolii inoculation or localized cyto-kinin addition Molecular PlantndashMicrobe Interaction 13617ndash628
Matvienko M van de Sande K Pawlowski K van Kammen AFranssen H Bisseling T 1996 Nicotiana tabacum SR1 containstwo ENOD40 homologs In Stacey G Mullin B Gresshoff PMeds Biology of plantndashmicrobe interactions St Paul USA Int SocMPMI 387ndash391
Minami E Kouchi H Cohn JR Ogawa T Stacey G 1996Expression of the early nodulin ENOD40 in soybean roots inresponse to various lipo-chitin signal molecules The Plant Journal10 23ndash32
Nagata T Okada K Tabeke I Matsui C 1981 Delivery of to-bacco mosaic virus RNA into plant protoplasts mediated byreverse-phase evaporation vesicles (liposomes) Molecular andGeneral Genetics 184 161ndash165
Nagata T Nemoto Y Hasezawa S 1992 Tobacco BY-2 cell lineas the He-la cell in the cell biology of higher plants Inter-national Reviews in Cytology 132 1ndash30
Oldroyd GED Engstrom EM Long SR 2001 Ethylene inhibitsthe Nod factor signal transduction pathway of Medicago trunca-tula The Plant Cell 13 1835ndash1849
Penmetsa RV Cook RV 1997 A legume ethylene-insensitivemutant hyperinfected by its rhizobial symbiont Science 275527ndash530
Peters NK Crist-Estes DK 1989 Nodule formation is stimulatedby the ethylene inhibitor aminoethoxyvinylglycine Plant Physi-ology 91 690ndash693
Theologis A 1992 One rotten apple spoils the whole bushel therole of ethylene in fruit ripening Cell 70 181ndash184
van de Sande K Pawlowski K Czaja I et al 1996 Mod-ification of phytohormone response by a peptide encodedby ENOD40 of legumes and a nonlegume Science 273370ndash373
Varkonyi-Gasic E White DWR 2002 The white cloverENOD40 gene family expression patterns of two types of genesindicate a role in vascular function Plant Physiology 1291107ndash1118
Vleghels I Hontelez J Ribeiro A Fransz P Bisseling TFranssen H 2003 Expression of ENOD40 during tomato plantdevelopment Planta 218 42ndash49
Wang KL Li H Ecker JR 2002 Ethylene biosynthesis andsignaling networks The Plant Cell 14 131ndash151
Yang SF Hoffman NE 1984 Ethylene biosynthesis and itsregulation in higher plants Annual Review of Plant Physiology35 155ndash189
Yang WC Katinakis P Hendriks P Smolders A de Vries FSpee J van Kammen A Bisseling T Franssen H 1993Characterization of GmENOD40 a gene showing novel pat-terns of cell-specific expression during soybean nodule develop-ment The Plant Journal 3 573ndash585
3282 Ruttink et al
by guest on June 1 2013httpjxboxfordjournalsorg
Dow
nloaded from
Related Documents

![New Therban Compound Development - SGF · 2018. 9. 17. · Therban LT2007 Change in Elongation [%] 14d 21d 42d. 14 ARLANXEO RubberCon 2016-06-08 Formulation 1: Ageing in Water/Ethylene](https://static.cupdf.com/doc/110x72/611c37f48070c147f6655f19/new-therban-compound-development-2018-9-17-therban-lt2007-change-in-elongation.jpg)









