Chemico-Biological Interactions 130 – 132 (2001) 659 – 671 Engineering steroid hormone specificity into aldo-keto reductases Trevor M. Penning *, Haiching Ma 1 , Joseph M. Jez 2 Department of Pharmacology, Uni6ersity of Pennsyl6ania School of Medicine, 3620 Hamilton Walk, Philadelphia, PA 19104 -6084, USA Abstract Steroid hormone transforming aldo-keto reductases (AKRs) include virtually all mam- malian 3a-hydroxysteroid dehydrogenases (3a-HSDs), 20a-HSDs, as well as the 5b-reduc- tases. To elucidate the molecular determinants of steroid hormone recognition we used rat liver 3a-HSD (AKR1C9) as a starting structure to engineer either 5b-reductase or 20a-HSD activity. 5b-Reductase activity was introduced by a single point mutation in which the conserved catalytic His (H117) was mutated to Glu117. The H117E mutant had a k cat comparable to that for homogeneous rat and human liver 5b-reductases. pH versus k cat profiles show that this mutation increases the acidity of the catalytic general acid Tyr55. It is proposed that the increased TyrOH 2 + character facilitates enolization of the D 4 -3-ketos- teroid and subsequent hydride transfer to C5. Since 5b-reductase precedes 3a-HSD in steroid hormone metabolism it is likely that this metabolic pathway arose by gene duplication and point mutation. 3a-HSD is positional and stereospecific for 3-ketosteroids and inactivates androgens. The enzyme was converted to a robust 20a-HSD, which is positional and stereospecific for 20-ketosteroids and inactivates progesterone, by the generation of loop- chimeras. The shift in log 10 (k cat /K m ) from androgens to progestins was of the order of 10 11 . This represents a rare example of how steroid hormone specificity can be changed at the enzyme level. Protein engineering with predicted outcomes demonstrates that the molecular determinants of steroid hormone recognition in AKRs will be ultimately rationalized. © 2001 Elsevier Science Ireland Ltd. All rights reserved. Keywords: 3a-Hydroxysteroid dehydrogenase; 5b-Reductase; 20a-Hydroxysteroid dehydrogenase; Protein engineering www.elsevier.com/locate/chembiont * Corresponding author. Tel.: +1-215-8989445; fax: +1-215-5732236. E-mail address: [email protected] (T.M. Penning). 1 Current address: Institute of Medicine in Engineering, 1150 Vagelos Laboratory, University of Pennsylvania, PA 19014, USA. 2 Current address: The Salk Institute for Biological Studies, Structural Biology Laboratory, 10010 North Torrey Pines Road, La Jolla, CA 92037, USA. 0009-2797/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved. PII:S0009-2797(00)00257-X

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Chemico-Biological Interactions 130–132 (2001) 659–671
Engineering steroid hormone specificity intoaldo-keto reductases
Trevor M. Penning *, Haiching Ma 1, Joseph M. Jez 2
Department of Pharmacology, Uni6ersity of Pennsyl6ania School of Medicine, 3620 Hamilton Walk,Philadelphia, PA 19104-6084, USA
Abstract
Steroid hormone transforming aldo-keto reductases (AKRs) include virtually all mam-malian 3a-hydroxysteroid dehydrogenases (3a-HSDs), 20a-HSDs, as well as the 5b-reduc-tases. To elucidate the molecular determinants of steroid hormone recognition we used ratliver 3a-HSD (AKR1C9) as a starting structure to engineer either 5b-reductase or 20a-HSDactivity. 5b-Reductase activity was introduced by a single point mutation in which theconserved catalytic His (H117) was mutated to Glu117. The H117E mutant had a kcat
comparable to that for homogeneous rat and human liver 5b-reductases. pH versus kcat
profiles show that this mutation increases the acidity of the catalytic general acid Tyr55. Itis proposed that the increased TyrOH2
+ character facilitates enolization of the D4-3-ketos-teroid and subsequent hydride transfer to C5. Since 5b-reductase precedes 3a-HSD in steroidhormone metabolism it is likely that this metabolic pathway arose by gene duplication andpoint mutation. 3a-HSD is positional and stereospecific for 3-ketosteroids and inactivatesandrogens. The enzyme was converted to a robust 20a-HSD, which is positional andstereospecific for 20-ketosteroids and inactivates progesterone, by the generation of loop-chimeras. The shift in log10(kcat/Km) from androgens to progestins was of the order of 1011.This represents a rare example of how steroid hormone specificity can be changed at theenzyme level. Protein engineering with predicted outcomes demonstrates that the moleculardeterminants of steroid hormone recognition in AKRs will be ultimately rationalized.© 2001 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: 3a-Hydroxysteroid dehydrogenase; 5b-Reductase; 20a-Hydroxysteroid dehydrogenase;Protein engineering
www.elsevier.com/locate/chembiont
* Corresponding author. Tel.: +1-215-8989445; fax: +1-215-5732236.E-mail address: [email protected] (T.M. Penning).1 Current address: Institute of Medicine in Engineering, 1150 Vagelos Laboratory, University of
Pennsylvania, PA 19014, USA.2 Current address: The Salk Institute for Biological Studies, Structural Biology Laboratory, 10010
North Torrey Pines Road, La Jolla, CA 92037, USA.
0009-2797/01/$ - see front matter © 2001 Elsevier Science Ireland Ltd. All rights reserved.
PII: S0009 -2797 (00 )00257 -X

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671660
1. Introduction
The aldo-keto reductase (AKR) superfamily contains steroid hormone transform-ing enzymes that play pivotal roles in the inactivation and mechanism of action ofsex steroids [1]. These enzymes cluster to the AKR1 family and include virtually all3a- and 20a-hydroxysteroid dehydrogenases (HSDs) as well as the 5b-reductases[2,3].
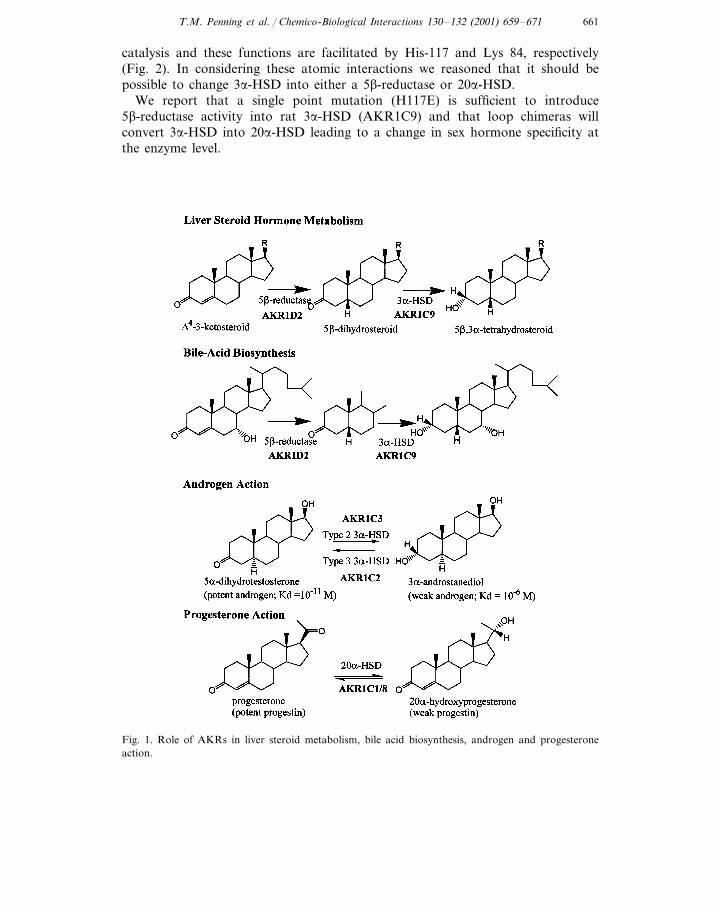
Apart from estrogens, all steroid hormones contain the D4-3-ketosteroid func-tionality and are converted to the 5a/5b-dihydrosteroids by the action of 5a/5b-re-ductases. The resultant dihydrosteroids are metabolized further by 3a-HSD to yieldthe 5a/3a- and 5b/3a-tetrahydrosteroids. In the liver, these reactions result in thereductive elimination of steroid hormones and protect against circulating steroidhormone excess, (Fig. 1). The enzymes responsible for this sequence in rodent liverare AKR1D2 and AKR1C9, while the corresponding enzymes in human liver areAKR1D1 and AKR1C4 (type 1 3a-HSD) [2,3].
In steroid target tissues these reactions are significant since the metabolism oftestosterone by 5a-reductase leads to the production of the potent androgen,5a-dihydrotestosterone (Kd=10−11 M for the androgen receptor), which is thenconverted by 3a-HSD isoforms to the inactive androgen, 3a-androstanediol (Kd=10−6 M for the androgen receptor). Here 3a-HSD isoforms interconvert a potentandrogen with its cognate inactive metabolite and regulate the occupancy andtrans-activation of the androgen receptor [1]. In human prostate, type 2 3a-HSD(AKR1C3) eliminates 5a-DHT from the gland while type 3 3a-HSD (AKR1C2)catalyzes the reverse reaction and forms the potent androgen 5a-DHT, Fig. 1 [4,5].The ability of these human isoforms to switch androgen action ‘off and on’,provides a pre-receptor regulation of steroid hormone action.
Rat ovarian 20a-HSD (AKR1C8) also catalyzes the inactivation of a potent sexhormone. It converts the female sex hormone, progesterone to yield the inactiveprogestin, 20a-hydroxyprogesterone. Induction of AKR1C8 contributes to luteoly-sis and the termination of pregnancy in rodents [6]. In humans the dominant20a-HSD is AKR1C1. Tissue distribution of AKR1C1 also suggests that it mayregulate the occupancy and trans-activation of the progesterone receptor since it ishighly expressed in the mammary gland [5]. Cluster analysis using pair-wisecomparisons shows that there is high sequence identity (\70%) between theseimportant sex hormone metabolizing AKRs suggesting that structural informationon one will act as a template for the rest [3].
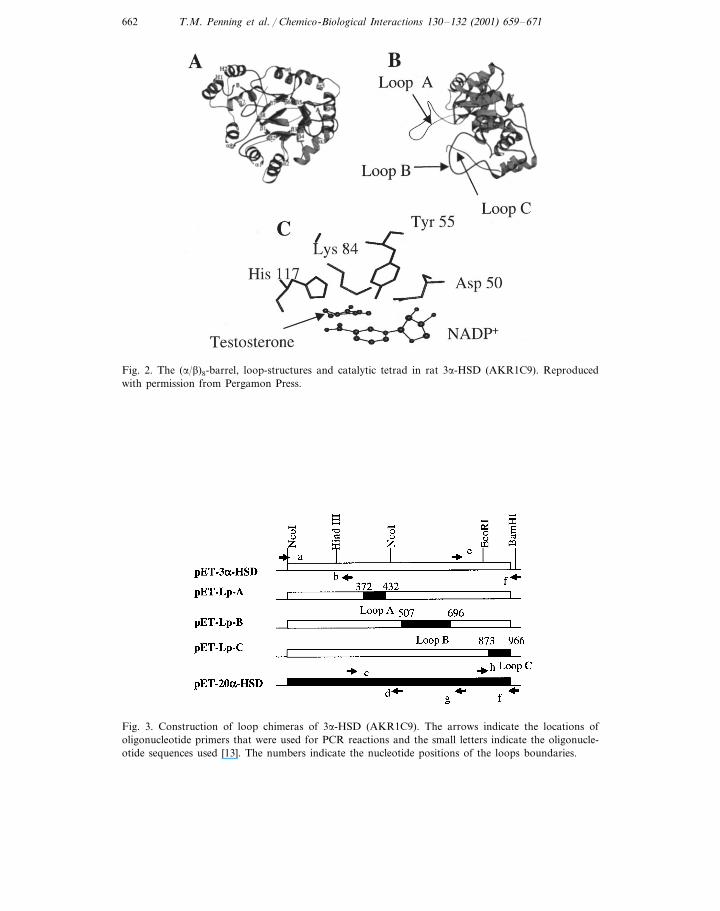
The crystal structure for the rat liver 3a-HSD (AKR1C9) NADP+–testosteroneternary complex has been determined, where testosterone is a competitive inhibitor[7]. As in other AKRs, the protein has an (a/b)8-barrel motif, the cofactor andtestosterone are bound at the back of the barrel and lie perpendicular to oneanother. The steroid binding site consists of ten residues from five loops includingthree large flexible loops (Loops, A, B and C) at the back of the barrel. The C4position of the nicotinamide ring and the C3-ketone of the steroid lie in closeproximity to a catalytic tetrad: Tyr 55, Lys 84, Asp 50 and His 117. Previoussite-directed mutagenesis identified Tyr 55 as the general acid/base involved in
T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 661
catalysis and these functions are facilitated by His-117 and Lys 84, respectively(Fig. 2). In considering these atomic interactions we reasoned that it should bepossible to change 3a-HSD into either a 5b-reductase or 20a-HSD.
We report that a single point mutation (H117E) is sufficient to introduce5b-reductase activity into rat 3a-HSD (AKR1C9) and that loop chimeras willconvert 3a-HSD into 20a-HSD leading to a change in sex hormone specificity atthe enzyme level.
Fig. 1. Role of AKRs in liver steroid metabolism, bile acid biosynthesis, androgen and progesteroneaction.
T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671662
Fig. 2. The (a/b)8-barrel, loop-structures and catalytic tetrad in rat 3a-HSD (AKR1C9). Reproducedwith permission from Pergamon Press.
Fig. 3. Construction of loop chimeras of 3a-HSD (AKR1C9). The arrows indicate the locations ofoligonucleotide primers that were used for PCR reactions and the small letters indicate the oligonucle-otide sequences used [13]. The numbers indicate the nucleotide positions of the loops boundaries.

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 663
2. Methods and methods
Materials and methods for these studies have been published elsewhere and thereader is referred to these sources [8–12]. The construction of the loop chimeras isshown in Fig. 3.
3. Results and discussion
3.1. Engineering 5b-reductase acti6ity into rat li6er 3a-HSD
Alignment of the catalytic tetrad residues in members of the AKR superfamilyindicate that these are invariant except in the rat and human 5b-reductases. In theseinstances His117 is now a glutamic acid. This suggested that 5b-reductase activitymay result from a single amino acid substitution in the catalytic tetrad. To test thishypothesis wild-type 3a-HSD was converted to the H117E mutant. The previouslydescribed H117A was also examined.
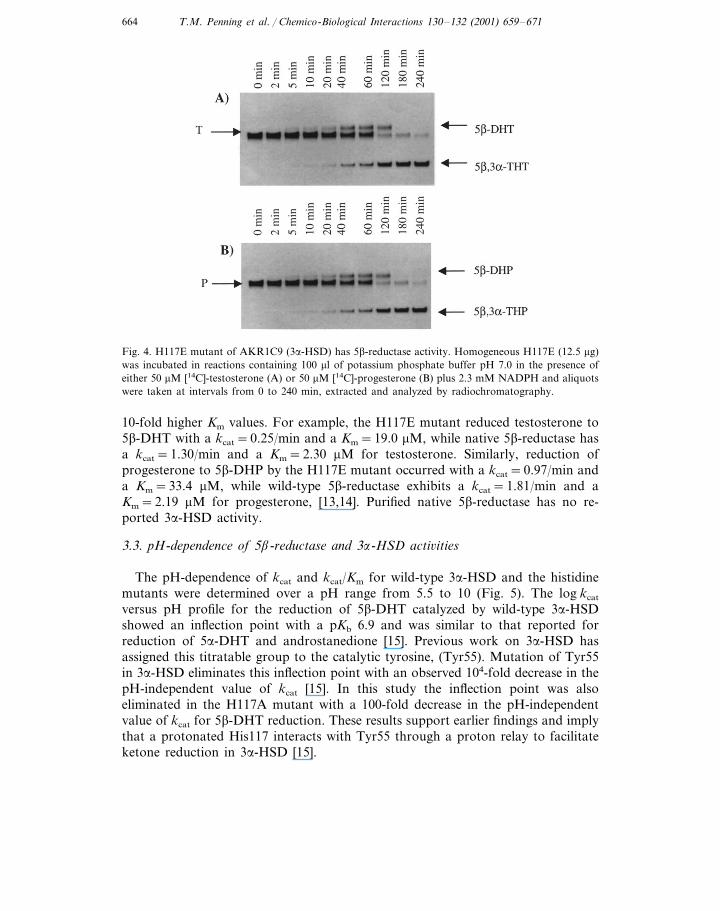
[14C]-Testosterone (T) was used as a substrate to assay the purified wild-type3a-HSD and the H117E and H117A mutants for 5b-reductase activity. As expected,wild-type 3a-HSD exhibited no 5b-reductase activity. The H117E mutant generateda product that co-migrated on TLC analysis with 5b-androstan-17b-ol-3-one (5b-DHT). However, this mutant also reduced 5b-DHT to 5b-androstan-3a,17b-diol[5b,3a-tetrahydrotestosterone; (5b,3a-THT)] during the assay, Fig. 4. Thus, theHis117E mutant introduced a stereospecific 5b-reductase activity, but also retained3a-HSD activity. Surprisingly, the H117A mutant also converted testosterone to5b,3a-THT but no 5b-DHT could be detected. This indicated that while 3a-HSDand 5b-reductase activities were also present, the 3a-HSD activity dominated in thismutant [10].
3.2. Kinetic characterization of the H117E and H117A mutants
Time courses were conducted using [14C]-T and [14C]-progesterone (P) as sub-strates to monitor the progress of the 5b-reductase and 3a-HSD activities in thehistidine mutants. The H117E mutant reduced testosterone to 5b-DHT (Fig. 4).However, as the concentration of 5b-DHT increased, the 3a-HSD activity of themutant reduced 5b-DHT to 5b,3a-THT. A similar transition was seen for theconversion of P to 5b-DHP and 5b,3a-THP. Based on the initial rates of D4-3-ke-tosteroid disappearance and appearance of 5b-dihydrosteroid products (Fig. 4), thespecific activities of the H117E mutant were 4.1 mM testosterone reduced/min permg protein and 9.8 mM progesterone reduced/min per mg protein.
The kinetic constants for 5b-reductase catalyzed by the H117E mutant weredetermined spectrophotometrically over the time course in which radiochromatog-raphy validated that only the 5b-dihydrosteroids were produced (0–2 min). Re-markably, the H117E mutant had similar kcat values to those reported forhomogeneous wild-type 5b-reductase purified from rat liver and demonstrated only
T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671664
Fig. 4. H117E mutant of AKR1C9 (3a-HSD) has 5b-reductase activity. Homogeneous H117E (12.5 mg)was incubated in reactions containing 100 ml of potassium phosphate buffer pH 7.0 in the presence ofeither 50 mM [14C]-testosterone (A) or 50 mM [14C]-progesterone (B) plus 2.3 mM NADPH and aliquotswere taken at intervals from 0 to 240 min, extracted and analyzed by radiochromatography.
10-fold higher Km values. For example, the H117E mutant reduced testosterone to5b-DHT with a kcat=0.25/min and a Km=19.0 mM, while native 5b-reductase hasa kcat=1.30/min and a Km=2.30 mM for testosterone. Similarly, reduction ofprogesterone to 5b-DHP by the H117E mutant occurred with a kcat=0.97/min anda Km=33.4 mM, while wild-type 5b-reductase exhibits a kcat=1.81/min and aKm=2.19 mM for progesterone, [13,14]. Purified native 5b-reductase has no re-ported 3a-HSD activity.
3.3. pH-dependence of 5b-reductase and 3a-HSD acti6ities
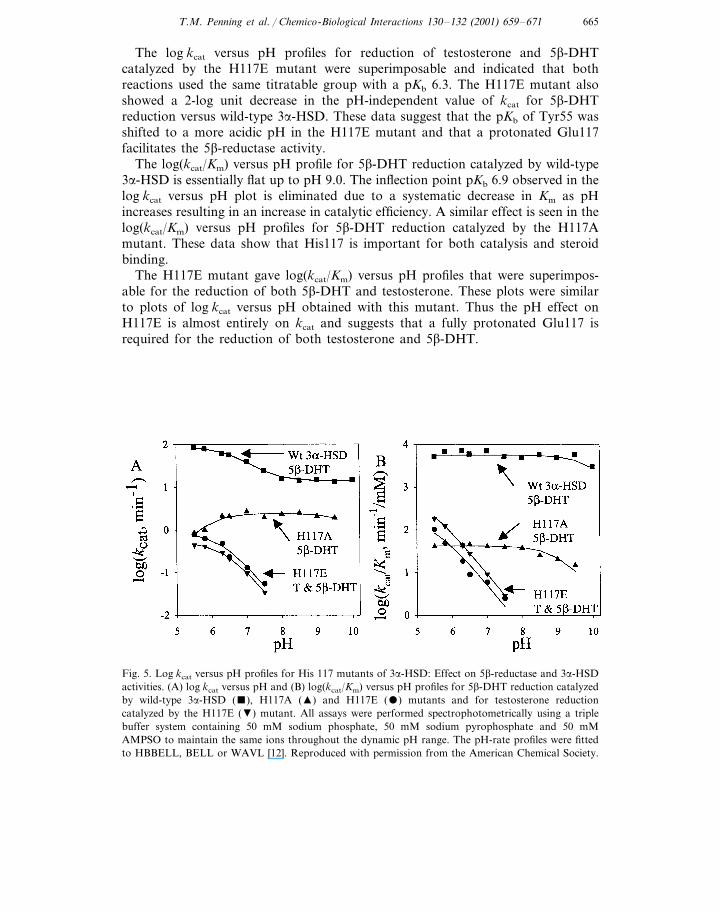
The pH-dependence of kcat and kcat/Km for wild-type 3a-HSD and the histidinemutants were determined over a pH range from 5.5 to 10 (Fig. 5). The log kcat
versus pH profile for the reduction of 5b-DHT catalyzed by wild-type 3a-HSDshowed an inflection point with a pKb 6.9 and was similar to that reported forreduction of 5a-DHT and androstanedione [15]. Previous work on 3a-HSD hasassigned this titratable group to the catalytic tyrosine, (Tyr55). Mutation of Tyr55in 3a-HSD eliminates this inflection point with an observed 104-fold decrease in thepH-independent value of kcat [15]. In this study the inflection point was alsoeliminated in the H117A mutant with a 100-fold decrease in the pH-independentvalue of kcat for 5b-DHT reduction. These results support earlier findings and implythat a protonated His117 interacts with Tyr55 through a proton relay to facilitateketone reduction in 3a-HSD [15].
T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 665
The log kcat versus pH profiles for reduction of testosterone and 5b-DHTcatalyzed by the H117E mutant were superimposable and indicated that bothreactions used the same titratable group with a pKb 6.3. The H117E mutant alsoshowed a 2-log unit decrease in the pH-independent value of kcat for 5b-DHTreduction versus wild-type 3a-HSD. These data suggest that the pKb of Tyr55 wasshifted to a more acidic pH in the H117E mutant and that a protonated Glu117facilitates the 5b-reductase activity.
The log(kcat/Km) versus pH profile for 5b-DHT reduction catalyzed by wild-type3a-HSD is essentially flat up to pH 9.0. The inflection point pKb 6.9 observed in thelog kcat versus pH plot is eliminated due to a systematic decrease in Km as pHincreases resulting in an increase in catalytic efficiency. A similar effect is seen in thelog(kcat/Km) versus pH profiles for 5b-DHT reduction catalyzed by the H117Amutant. These data show that His117 is important for both catalysis and steroidbinding.
The H117E mutant gave log(kcat/Km) versus pH profiles that were superimpos-able for the reduction of both 5b-DHT and testosterone. These plots were similarto plots of log kcat versus pH obtained with this mutant. Thus the pH effect onH117E is almost entirely on kcat and suggests that a fully protonated Glu117 isrequired for the reduction of both testosterone and 5b-DHT.
Fig. 5. Log kcat versus pH profiles for His 117 mutants of 3a-HSD: Effect on 5b-reductase and 3a-HSDactivities. (A) log kcat versus pH and (B) log(kcat/Km) versus pH profiles for 5b-DHT reduction catalyzedby wild-type 3a-HSD (), H117A (�) and H117E () mutants and for testosterone reductioncatalyzed by the H117E (�) mutant. All assays were performed spectrophotometrically using a triplebuffer system containing 50 mM sodium phosphate, 50 mM sodium pyrophosphate and 50 mMAMPSO to maintain the same ions throughout the dynamic pH range. The pH-rate profiles were fittedto HBBELL, BELL or WAVL [12]. Reproduced with permission from the American Chemical Society.

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671666
Fig. 6. Proposed mechanism of 5b-reduction: facilitation of acid-catalyzed enolization by H117E mutant.Reproduced with permission from Pergamon Press.
3.4. Tyr55 as the general acid for the 5b-reductase and 3a-HSD acti6ities
To test the role of Tyr55 as the general acid for both 3a-HSD and 5b-reductaseactivities, the homogeneous Y55F/H117E double mutant was assayed for bothenzymatic activities. As expected, wild-type 3a-HSD did not reduce [14C]-testos-terone and the H117E mutant functioned as both a 5b-reductase and 3a-HSD. TheY55F mutant was severely impaired for reduction of [14C]-5a-DHT. However, theY55F/H117E mutant did not catalyze the reduction of either 3-ketosteroids orD4-3-ketosteroids (data not shown). These results are consistent with Tyr55 actingas the general acid for both 3a-HSD and 5b-reductase activities.
On this basis a mechanism for steroid double bond reduction catalyzed by theH117E mutant and the 5b-reductases of the AKR superfamily is proposed (Fig. 6).Following the binding of NADPH and testosterone, Tyr55 promotes acid-catalyzedenolization of the D4-3-ketosteroid. Similar to the role of His117 in 3-ketosteroidreduction [15], a protonated Glu117 hydrogen bonds with Tyr55 to lower the pKb
of the tyrosine making it a stronger acid to facilitate the enolization step. Theresultant 3,4-enol produces cationic character at C5 and facilitates hydride transferfrom NADPH to the b-face of the steroid C5. Finally, the enol can tautomerize togenerate the 5b-dihydrosteroid product.
Since 5b-reductase precedes 3a-HSD in steroid hormone metabolism and bothproteins are members of the AKR superfamily, the introduction of 5b-reductaseinto 3a-HSD by a single point mutation suggests that this major pathway of steroidmetabolism may have arisen by gene duplication and point mutation of an AKR.
3.5. Engineering 3a-HSD into 20a-HSD using crystal structures as templates
The availability of a crystal structure for the 3a-HSD–NADP+–testosteronecomplex, and the observation that HSDs in the AKR superfamily share an excessof 67% sequence identity, suggested that it should be possible to convert 3a-HSD(AKR1C9) into 20a-HSD (AKR1C8) and change sex hormone specificity fromandrogens to progestins.
The crystal structure predicts that atomic interactions that bind the cofactor arehighly conserved within the superfamily so that the stereochemistry of hydride

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 667
transfer to the substrate carbonyl is maintained. To alter sex hormone specificity itis the position of the steroid that must change. In 3a-HSD, it is the C3 ketone ofandrogens that is reduced, whereas in 20a-HSD it is the C20 ketone of progestinsthat is reduced. This implies that in 20a-HSD the steroid must bind backwardsrelative to the position of the steroid in 3a-HSD, i.e. the D-ring must bind in theA-ring position. To achieve this 180° rotation we examined how testosterone wasbound in 3a-HSD. This steroid pocket is comprised of three large loops (an orderedloop A and the mobile loops B and C), Fig. 2. Additional residues are recruitedfrom two smaller loops that connect two b-strands and two a-helices (b1–a1 andb2–a2) in the (a/b)8 barrel motif. These five loops contribute ten residues that formpoints of contact with testosterone. Alignment of these residues with those presentin 20a-HSD indicates that six out of ten residues are different (Table 1). Conservedresidues were either part of the catalytic tetrad Tyr 55 and His 117 or their nearestneighbors Leu 54 and Phe 118. To predict which of the six residues conferspecificity, the steroid binding pockets of 3a-HSD and 20a-HSD were compared atthe atomic level. A 20a-HSD–NADP+–testosterone complex was built using thecoordinates for the 3a-HSD–NADP+–testosterone complex. The 3a-HSD ternarycomplex depicts a perfect pocket for testosterone but substitution of 20a-HSDresidues resulted in significant clashes between the mutated residues and testos-terone (Fig. 7). Progesterone was then substituted in the 20a-HSD–NADP+–testosterone complex to determine whether progesterone could fit the alteredpocket. The resultant model predicted that altered residues could accommodate thebinding of progesterone in the backwards orientation (D-ring at the A-ring posi-tion) and place the C17 side chain near the active site residues, which are essentialchanges if 20a-HSD activity is to be introduced.
Fig. 7. Homology model of the steroid-binding site of 20a-HSD. Reproduced with permission fromPergamon Press.

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671668
Table 1Alignment of steroid binding residues in AKR1C9 (3a-HSD) and AKR1C8 (20a-HSD)
Loop A Loop B Loop CLoopa b1–a1 b2–a2
54 55 117 118 129 226 227 306 31024
YbLbT Hb FbRat liver F T Wb N Y3a-HSD
MFCYLFbHbRat ovarian YbYb Lb
20a-HSD
a The secondary structure motifs are defined by the ternary complex of 3a-HSD.b Conserved in other mammalian 3a-HSDs.
3.6. Specific acti6ities of point mutants in the substrate binding pocket
Based on the sequence alignment and modeling studies, individual and multipleamino acids in the pocket were mutated in an attempt to convert 3a-HSD to20a-HSD. All mutants were expressed in E. coli and immunoblot analysis was usedto ensure that all mutants were similarly expressed (data not shown). Rather thanpurifying each of these mutants to homogeneity, the E. coli sonicates were assayedspectrophotometrically for 3a- and 20a-HSD. The single mutants, T24Y, F129Land the double mutant, N306F/Y310M, all retained activity (47–91 mM/min permg) that was similar to a sonicate of wt 3a-HSD (52 mM/min per mg). By contrast,the double mutant T226Y/W227C retained low 3a-HSD activity (2 mM/min permg). None of the mutants had 20a-HSD activity. Even when all the predictedmutations necessary to convert 3a-HSD into 20a-HSD were introduced, theresultant mutant T24Y/F129L/T226Y/W227C/N306F/Y310M had no 20a-HSDactivity. Based on the sensitivity of our assay an inactive mutant had less than 0.01mM/min per mg activity for 3a or 20a-HSD substrates.
3.7. Initial characterization of chimeric proteins
Chimeric proteins were then constructed in which the steroid binding loops of3a-HSD were replaced with loops of 20a-HSD. In designing these chimeras the twosmall loops b1–a1 and b2–a2 were not replaced. Loop b1–a1 contributes only oneresidue to the binding pocket and with the exception of rat 3a-HSD is a Tyr inother 3a, 17b and 20a-HSDs of the superfamily. Loop b2–a2 was not swappedsince it contains the catalytic residue Tyr 55. Other chimeric HSDs were overex-pressed and purified to homogeneity from E. coli for kinetic characterization. TheLp-B chimera and double loop chimeras Lp-AB and Lp-BC were not characterizedsince their activity was too low. It is estimated that these chimeras oxidized lessthan 3 mM androsterone/min per mg as compared to 1.6 mM androsteroneoxidized/min per mg by homogenous recombinant 3a-HSD. These chimeras alsodisplayed no 20a-HSD activity where the limit of detection was 0.01 mM 20a-OHPoxidized/min per mg.
T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 669
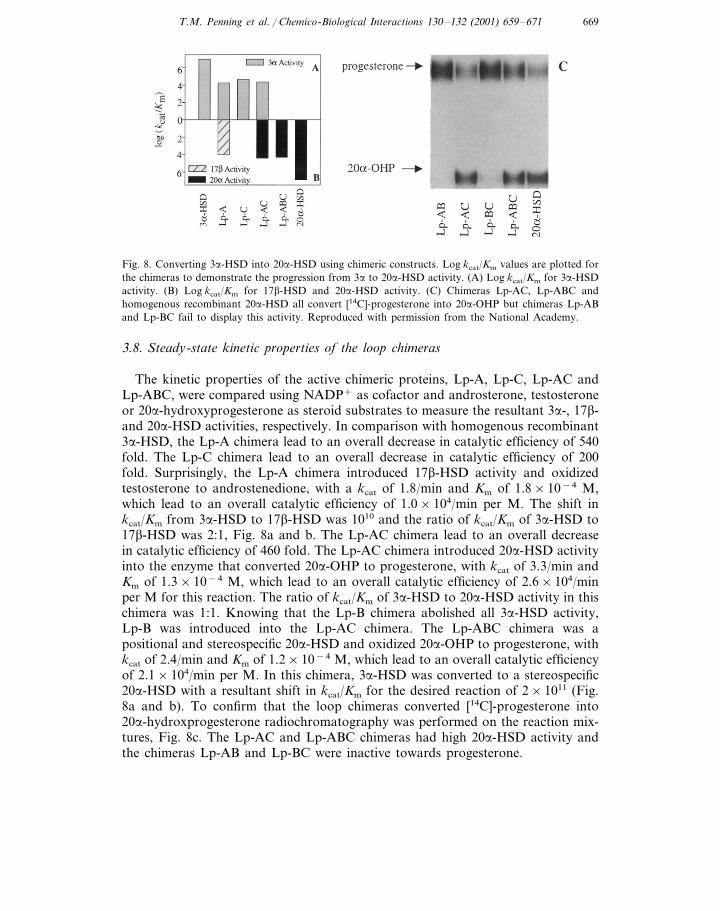
Fig. 8. Converting 3a-HSD into 20a-HSD using chimeric constructs. Log kcat/Km values are plotted forthe chimeras to demonstrate the progression from 3a to 20a-HSD activity. (A) Log kcat/Km for 3a-HSDactivity. (B) Log kcat/Km for 17b-HSD and 20a-HSD activity. (C) Chimeras Lp-AC, Lp-ABC andhomogenous recombinant 20a-HSD all convert [14C]-progesterone into 20a-OHP but chimeras Lp-ABand Lp-BC fail to display this activity. Reproduced with permission from the National Academy.
3.8. Steady-state kinetic properties of the loop chimeras
The kinetic properties of the active chimeric proteins, Lp-A, Lp-C, Lp-AC andLp-ABC, were compared using NADP+ as cofactor and androsterone, testosteroneor 20a-hydroxyprogesterone as steroid substrates to measure the resultant 3a-, 17b-and 20a-HSD activities, respectively. In comparison with homogenous recombinant3a-HSD, the Lp-A chimera lead to an overall decrease in catalytic efficiency of 540fold. The Lp-C chimera lead to an overall decrease in catalytic efficiency of 200fold. Surprisingly, the Lp-A chimera introduced 17b-HSD activity and oxidizedtestosterone to androstenedione, with a kcat of 1.8/min and Km of 1.8×10−4 M,which lead to an overall catalytic efficiency of 1.0×104/min per M. The shift inkcat/Km from 3a-HSD to 17b-HSD was 1010 and the ratio of kcat/Km of 3a-HSD to17b-HSD was 2:1, Fig. 8a and b. The Lp-AC chimera lead to an overall decreasein catalytic efficiency of 460 fold. The Lp-AC chimera introduced 20a-HSD activityinto the enzyme that converted 20a-OHP to progesterone, with kcat of 3.3/min andKm of 1.3×10−4 M, which lead to an overall catalytic efficiency of 2.6×104/minper M for this reaction. The ratio of kcat/Km of 3a-HSD to 20a-HSD activity in thischimera was 1:1. Knowing that the Lp-B chimera abolished all 3a-HSD activity,Lp-B was introduced into the Lp-AC chimera. The Lp-ABC chimera was apositional and stereospecific 20a-HSD and oxidized 20a-OHP to progesterone, withkcat of 2.4/min and Km of 1.2×10−4 M, which lead to an overall catalytic efficiencyof 2.1×104/min per M. In this chimera, 3a-HSD was converted to a stereospecific20a-HSD with a resultant shift in kcat/Km for the desired reaction of 2×1011 (Fig.8a and b). To confirm that the loop chimeras converted [14C]-progesterone into20a-hydroxprogesterone radiochromatography was performed on the reaction mix-tures, Fig. 8c. The Lp-AC and Lp-ABC chimeras had high 20a-HSD activity andthe chimeras Lp-AB and Lp-BC were inactive towards progesterone.

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671670
3.9. Lessons from the loop chimeras
The ability of the Lp-A chimera to act as both a 3a- and 17b-HSD wasunexpected. For the enzyme to display 17b-HSD activity, the pocket must havebeen altered so that it binds testosterone not only in the backwards orientation(D ring in the A ring position) but also upside down (b-face in the a-faceorientation, angular methyl groups inverted) to preserve the stereochemistryof hydride transfer. Thus the replacement of loop A of 3a-HSD with thecorresponding loop in 20a-HSD produces a flexible binding pocket. Cry-stallographic studies on 3a-HSD and its complexes indicated that loop A wasordered and its position was indepen-dent of the binding of cofactor or steroid[7]. Our data indicate that loop A residues may be more plastic than originallyproposed.
The Lp-B chimera eliminated all 3a-HSD activity but the Lp-C chimera main-tained relatively high 3a-HSD activity. These results were also unexpected sinceloop C undergoes the most movement upon the binding of testosterone, andloop C has been implicated in substrate and inhibitor specificity in other AKRs.
Double loop chimeras Lp-AB and Lp-BC had such low 3a- and 20a-HSDactivity that complete kinetic characterization was not possible. This was alsosurprising since Lp-BC contains both loops that undergo ligand-induced move-ment. Surprisingly, the Lp-AC chimera gave an enzyme that used both andros-terone and 20a-OHP as substrates. This double loop chimera thus had a newsubstrate-binding pocket that permits the backwards binding of C21 steroids (Dring in the A ring position), but it did not allow the binding of steroids upsidedown (b-face in the a-face orientation). Thus the introduction of loop C of20a-HSD made the pocket of the Lp-A chimera more discriminating since testos-terone was no longer oxidized.
The Lp-ABC chimera that contained all three loops of 20a-HSD had no3a-HSD activity but converted 3a-HSD into a positional and stereospecific 20a-HSD. This chimera displayed a kcat comparable to the human 20a-HSD(AKR1C1), only its Km was ten-fold higher [13]. The catalytic efficiency of thischimera was still 404-fold less than homogenous recombinant rat ovarian 20a-HSD, but a change in kcat/Km of 1011 was achieved for the desired reaction.There are few cases in protein engineering where such a large change in kcat/Km
has been achieved in generating a target activity. Protein engineering in AKRswith predicted outcomes will ultimately demonstrate that the molecular determi-nants of steroid hormone recognition are understood in this superfamily.
Acknowledgements
Supported in part by a grant from the National Institutes of Health(DK47015 to T.M.P.).

T.M. Penning et al. / Chemico-Biological Interactions 130–132 (2001) 659–671 671
References
[1] T.M. Penning, Molecular endocrinology of hydroxysteroid dehydrogenases, Endocr. Rev. 18 (1997)281–305.
[2] J.M. Jez, M.J. Bennett, B.P. Schlegel, M. Lewis, T.M. Penning, Comparitive anatomy of thealdo-keto reductase superfamily, Biochem. J. 326 (1997) 625–636.
[3] J.M. Jez, T.G. Flynn, T.M. Penning, A new nomenclature for the aldo-keto reductase superfamily,Biochem. Pharmacol. 54 (1997) 639–647.
[4] H.K. Lin, J.M. Jez, B.P. Schlegel, D.M. Peehl, J.A. Pachter, T.M. Penning, Expression andcharacterization of recombinant type 2 3a-hydroxysteroid dehydrogenase (HSD) from humanprostate: demonstration of bifunctional 3a/17b-HSD activity and cellular distribution, Mol. En-docrinol. 11 (1997) 1971–1984.
[5] T.M. Penning, M.E. Burczynski, J.M. Jez, C-F. Hung, H-K. Lin, H. Ma, M. Moore, N. Palackal,K. Ratnam, Human 3a-hydroxysteroid dehydrogenase isoforms (AKR1C1-AKR1C4) of the aldo-keto reductase superfamily: functional plasticity and tissue distribution reveals roles in the inactiva-tion and formation of male and female sex hormones, Biochem. J. 351 (2000) 67–77.
[6] J.F. Strauss, Jr, III, R.L. Stambaugh, Induction of 20a-hydroxysteroid dehydrogenase in ratcorpora lutea of pregnancy by prostaglandin F-2a, Prostaglandins 5 (1974) 73–85.
[7] M.J. Bennett, R.H. Albert, J.M. Jez, H. Ma, T.M. Penning, M. Lewis, Steroid recognition andregulation of hormone action: crystal structure of testosterone and NADP+ bound to 3a-hydroxys-teroid/dihydrodiol dehydrogenase, Structure 5 (1997) 799–812.
[8] J.E. Pawlowski, T.M. Penning, Overexpression and mutagenesis of the cDNA for rat liver3a-hydroxysteroid/dihydrodiol dehydrogenase. Role of cysteines and tyrosines in catalysis, J. Biol.Chem. 269 (1994) 13502–13510.
[9] H. Ma, T.M. Penning, Characterization of homogeneous recombinant rat ovarian 20a-hydroxys-teroid dehydrogenase: fluorescent properties and inhibition profile, Biochem. J. 341 (1999) 853–859.
[10] J.M. Jez, T.M. Penning, Engineering steroid 5b-reductase activity into rat liver 3a-hydroxysteroiddehydrogenase, Biochemistry 37 (1998) 9695–9703.
[11] H. Ma, T.M. Penning, Conversion of mammalian 3a-hydroxysteroid dehydrogenase to 20a-hydrox-ysteroid dehydrogenase using loop chimeras: changing specificity from androgens to progestins,Proc. Natl. Acad. Sci. U.S.A. 96 (1999) 11161–11166.
[12] W.W. Cleland, Statistical analysis of enzyme kinetic data, Methods Enzymol. 63 (1979) 103–117.[13] A. Okuda, K. Okuda, Purification and characterization of D4-3-ketosteroid 5b-reductase, J. Biol.
Chem. 259 (1984) 7519–7524.[14] K. Kondo, M. Kai, Y. Setoguchi, G. Eggertsen, P. Sjoborn, T. Setoguchi, K. Okuda, I. Bjorkhem,
Cloning and expression of cDNA of human D4-3-oxosteroid-5b-reductase and substrate specificityof the expressed enzyme, Eur. J. Biochem. 219 (1994) 357–363.
[15] B.P. Schlegel, J.M. Jez, T.M. Penning, Mutagenesis of 3a-hydroxysteroid dehydrogenase reveals a‘push–pull’ mechanism for proton transfer in aldo-keto reductases, Biochemistry 37 (1998) 3583–3584.
.
Related Documents











