Research article Biosynthesis of flavan 3-ols by leucoanthocyanidin 4-reductases and anthocyanidin reductases in leaves of grape (Vitis vinifera L.), apple (Malus x domestica Borkh.) and other crops Judith Pfeiffer a , Christiane Kühnel a , Jeannette Brandt a , Daniela Duy a , P.A. Nimal Punyasiri b , Gert Forkmann a , Thilo C. Fischer a, * a Chair of Floriculture Crops and Horticultural Plant Breeding, Technical University München, Am Hochanger 4, 85350 Freising, Germany b Biochemistry Division, Tea Research Institute of Sri Lanka, Talawakelle, Sri Lanka Received 12 October 2005 Available online 13 June 2006 Abstract Catechin and epicatechin biosyntheses were studied of grape (Vitis vinifera L.), apple (Malus x domestica Borkh.) and other crop leaves, since these monomers and the derived proanthocyanidins are important disease resistance factors. Grape and apple leucoanthocyanidin 4-reductase (LAR; EC 1.17.1.3) enzymes were characterized on basis of plant and recombinant enzymes. In case of grape, two LAR cDNAs were cloned by assembling available EST sequences. Grape and apple leaf anthocyanidin reductase (ANR; EC 1.3.1.77) cDNAs were also obtained and the respective plant and recombinant enzymes were characterized. Despite general low substrate specificity, within the respective flavonoid biosynth- eses of grape and apple leaves, both enzyme types deliver differently hydroxylated catechins and epicatechins, due to substrate availability in vivo. Furthermore, for LAR enzymes conversion of 3-deoxyleucocyanidin was shown. Beside relevance for plant protection, this restricts the number of possible reaction mechanisms of LAR. ANR enzyme activity was demonstrated for a number of other crop plants and its correlation with (–)-epicatechin and obvious competition with UDP-glycosyl:flavonoid-3-O-glycosyltransferases was considered. © 2006 Elsevier SAS. All rights reserved. Keywords: Apple; Catechin biosynthesis; Epicatechin biosynthesis; Flavonoids; Grape; Plant resistance; Proanthocyanidins 1. Introduction The elucidation of flavanol ((+)-catechins, (–)-epicatechins and proanthocyanidins) biosynthesis in plants has made pro- gress in two important ways. Firstly, a gene of a leucoantho- cyanidin 4-reductase (LAR; (2R,3S)-catechin:NADP + 4- oxidoreductase; EC 1.17.1.3) has recently been cloned from a legume [41]. Furthermore, with the anthocyanidin reductase (ANR; flavan-3-ol:NAD(P) + oxidoreductase; EC 1.3.1.77) reaction, a previously unknown enzymatic reaction was identi- fied with a recombinant enzyme from an Arabidopsis gene (banyuls) involved in proanthocyanidin biosynthesis [45]. Twenty years ago, LAR was described as an enzymatic reaction from conifers (Pseudotsuga menziesii, Cryptomeria japonica) [23,38], Ginkgo biloba [37], barley (Hordeum vul- gare) [27], and later from legumes (Onobrychis viciifolia) [40,41]. It converts the flavan 3,4-diol products of the dihydro- flavonol 4-reductase (DFR) reaction to the corresponding cate- chins (Fig. 1). The legume LAR shows some substrate specifi- cities with respect to B-ring hydroxylation [41]. The first lar gene, recently cloned from the legume Desmodium uncinatum [41], was found to belong to the superfamily of isoflavone reductases. www.elsevier.com/locate/plaphy Plant Physiology and Biochemistry 44 (2006) 323–334 Abbreviations: ANR, anthocyanidin reductase; DFR, dihydroflavonol 4- reductase; DHK, dihydrokaempferol; DHQ, dihydroquercetin; DMACA, dimethylaminocinnamicaldehyde; EGME, ethylenglycolmonomethylether; ERI, eriodictyol; EtOAc, ethylacetate; EtOH, ethanol; HOAc, acetic acid; HPLC, high performance liquid chromatography; LAR, leucoanthocyanidin 4-reductase; MeOH, methanol; NADPH, nicotinamide adenine dinucleotide phosphate; RT-PCR, reverse transcription/polymerase chain reaction; TLC, thin layer chromatography; UFGT, UDP- glycosyl:flavonoid-3-O-glycosyltransferase. * Corresponding author. Tel.: +49 8161 71 3419; fax: +49 8161 71 3886. E-mail address: [email protected] (T.C. Fischer). 0981-9428/$ - see front matter © 2006 Elsevier SAS. All rights reserved. doi:10.1016/j.plaphy.2006.06.001

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

www.elsevier.com/locate/plaphy
Plant Physiology and Biochemistry 44 (2006) 323–334
Research article
Biosynthesis of flavan 3-ols by leucoanthocyanidin 4-reductases
AbreductdimethERI, eHPLC4-reduphosphthin laglycos
* CoE-m
0981-9doi:10
and anthocyanidin reductases in leaves of grape (Vitis vinifera L.),
apple (Malus x domestica Borkh.) and other cropsJudith Pfeiffera, Christiane Kühnela, Jeannette Brandta, Daniela Duya, P.A. Nimal Punyasirib,Gert Forkmanna, Thilo C. Fischera,*
aChair of Floriculture Crops and Horticultural Plant Breeding, Technical University München, Am Hochanger 4, 85350 Freising, GermanybBiochemistry Division, Tea Research Institute of Sri Lanka, Talawakelle, Sri Lanka
Received 12 October 2005Available online 13 June 2006
Abstract
Catechin and epicatechin biosyntheses were studied of grape (Vitis vinifera L.), apple (Malus x domestica Borkh.) and other crop leaves, sincethese monomers and the derived proanthocyanidins are important disease resistance factors. Grape and apple leucoanthocyanidin 4-reductase(LAR; EC 1.17.1.3) enzymes were characterized on basis of plant and recombinant enzymes. In case of grape, two LAR cDNAs were clonedby assembling available EST sequences. Grape and apple leaf anthocyanidin reductase (ANR; EC 1.3.1.77) cDNAs were also obtained and therespective plant and recombinant enzymes were characterized. Despite general low substrate specificity, within the respective flavonoid biosynth-eses of grape and apple leaves, both enzyme types deliver differently hydroxylated catechins and epicatechins, due to substrate availability invivo. Furthermore, for LAR enzymes conversion of 3-deoxyleucocyanidin was shown. Beside relevance for plant protection, this restricts thenumber of possible reaction mechanisms of LAR. ANR enzyme activity was demonstrated for a number of other crop plants and its correlationwith (–)-epicatechin and obvious competition with UDP-glycosyl:flavonoid-3-O-glycosyltransferases was considered.© 2006 Elsevier SAS. All rights reserved.
Keywords: Apple; Catechin biosynthesis; Epicatechin biosynthesis; Flavonoids; Grape; Plant resistance; Proanthocyanidins
1. Introduction
The elucidation of flavanol ((+)-catechins, (–)-epicatechinsand proanthocyanidins) biosynthesis in plants has made pro-gress in two important ways. Firstly, a gene of a leucoantho-cyanidin 4-reductase (LAR; (2R,3S)-catechin:NADP+ 4-
breviations: ANR, anthocyanidin reductase; DFR, dihydroflavonol 4-ase; DHK, dihydrokaempferol; DHQ, dihydroquercetin; DMACA,ylaminocinnamicaldehyde; EGME, ethylenglycolmonomethylether;riodictyol; EtOAc, ethylacetate; EtOH, ethanol; HOAc, acetic acid;, high performance liquid chromatography; LAR, leucoanthocyanidinctase; MeOH, methanol; NADPH, nicotinamide adenine dinucleotideate; RT-PCR, reverse transcription/polymerase chain reaction; TLC,yer chromatography; UFGT, UDP-yl:flavonoid-3-O-glycosyltransferase.rresponding author. Tel.: +49 8161 71 3419; fax: +49 8161 71 3886.ail address: [email protected] (T.C. Fischer).
428/$ - see front matter © 2006 Elsevier SAS. All rights reserved..1016/j.plaphy.2006.06.001
oxidoreductase; EC 1.17.1.3) has recently been cloned from alegume [41]. Furthermore, with the anthocyanidin reductase(ANR; flavan-3-ol:NAD(P)+ oxidoreductase; EC 1.3.1.77)reaction, a previously unknown enzymatic reaction was identi-fied with a recombinant enzyme from an Arabidopsis gene(banyuls) involved in proanthocyanidin biosynthesis [45].
Twenty years ago, LAR was described as an enzymaticreaction from conifers (Pseudotsuga menziesii, Cryptomeriajaponica) [23,38], Ginkgo biloba [37], barley (Hordeum vul-gare) [27], and later from legumes (Onobrychis viciifolia)[40,41]. It converts the flavan 3,4-diol products of the dihydro-flavonol 4-reductase (DFR) reaction to the corresponding cate-chins (Fig. 1). The legume LAR shows some substrate specifi-cities with respect to B-ring hydroxylation [41]. The first largene, recently cloned from the legume Desmodium uncinatum[41], was found to belong to the superfamily of isoflavonereductases.

Fig. 1. Enzymatic steps in (+)-catechin (a) and (–)-epicatechin biosynthesis (b) performed by LAR and ANR, respectively.
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334324
The LAR enzymatic reaction was recently found as a sub-sequent step in DFR reactions with enzyme preparations fromsome major crop plants, such as tea (Camellia sinensis) [31],rose (Rosa x hybrida) [26], and strawberry (Fragaria x ana-nassa) [unpublished results].
The banyuls gene of Arabidopsis, whose loss of function—mutation leads to a proanthocyanidin deficient seed testa, wasfirst supposed to represent an lar gene [11], it is like isoflavonereductases distantly related to the DFR. However, upon hetero-logous expression of the respective Arabidopsis and Medicagotruncatula cDNAs, the yet unknown reaction was identified forthe enzymes, leading from anthocyanidins to the respectiveepicatechins by NADPH-dependent reduction [44,45]. Thisanthocyanidin reductase (ANR) seems to be the enzymeresponsible for (–)-epicatechin biosynthesis in many plants,among them tea as an outstanding example [31].
Flavan 3-ols (catechins and epicatechins) are precursors ofthe polymeric proanthocyanidins [12, and citations therein];there are several features that make the monomers and theproanthocyanidins relevant as plant secondary metabolites:the protein precipitation by them influences the brewery pro-cess as well as quality of pasture. In fruits they cause astrin-gency [12, and citations therein]. Furthermore, catechins andproanthocyanidins are also important antioxidants and alsohave other beneficial effects on human health [2,24]. In leavescatechins, epicatechins and the polymeric proanthocyanidinsare especially important as plant resistance factors [8,14,34].For the analysis of the abovementioned functions of the flava-nols, in particular in delivering plant resistance factors, theplant and the respective recombinant LAR and ANR enzymeswere obtained from grape and apple leaves and studied withrespect to their enzyme characteristics. The respective LARand ANR enzymes from grape and apple possess similar char-acteristics and substrate specificities, but considering the quitedifferent flavonoid biosyntheses in both plants, they performpartly different biosynthetic reactions. Additionally, a screen-ing for ANR enzyme activity and its correlation with(–)-epicatechin content was performed in a number of crops,to examine prevalence of this biosynthetic pathway.
2. Results
Enzyme activities of both, LAR and ANR, were found to bepresent in crude enzyme preparations of young grape and appleleaves accumulating catechins and epicatechins. Sequences ofcloned genes and available database sequence informationwere used to amplify the lar and anr cDNAs from grape andapple leaves by RT-PCR. Heterologous expression in yeastwas performed to obtain the recombinant enzymes. Theenzymes prepared from leaves and the recombinant enzymeswere used for characterization of the respective reactions.
2.1. Biosynthesis of grape catechins and epicatechins
2.1.1. Leucoanthocyanidin 4-reductaseAn LAR enzyme assay with enzyme extract from young
grape (Vitis vinifera) leaves of cultivar Regent confirmedLAR enzyme activity in leaves. Such leaves were then usedfor cDNA preparation and subsequent cloning of lar cDNAs.The cloning of lar genes from grape was based on EST data-base information of various Vitis species identified by BLASTN searches. One lar cDNA sequence (v-lar1) could be comple-tely reconstructed by overlapping EST sequences from Vitisshuttleworthii, Vitis riparia and Vitis vinifera and was subse-quently RT-PCR-amplified from grape leaf cDNA. For a sec-ond lar cDNA (v-lar2) only the sequence of the 3′ region couldbe assembled from three Vitis vinifera EST sequences, the 5′region was obtained by a RACE protocol, subsequently thewhole coding region was RT-PCR-amplified. Grape leaf larsequences showed 57% (v-lar1) and 54% (v-lar2) identity atthe nucleotide level and 55% (v-lar1) and 52% (v-lar2) identityat the amino acid level to the lar of Desmodium. To each otherthe grape leaf lar sequences showed 65% (nucleotide) and 58%(amino acid) identity. The recombinant enzymes produced byheterologous yeast expression were both active with14C-labelled leucocyanidin as a substrate. Both recombinantenzymes have flat temperature optima between 30 °C and37 °C and pH optima of 6.5 ± 0.5. With respect to substratespecificity, v-LAR1 and v-LAR2 of grape are both acceptingthe three leucoanthocyanidins leucopelargonidin, leucocyani-
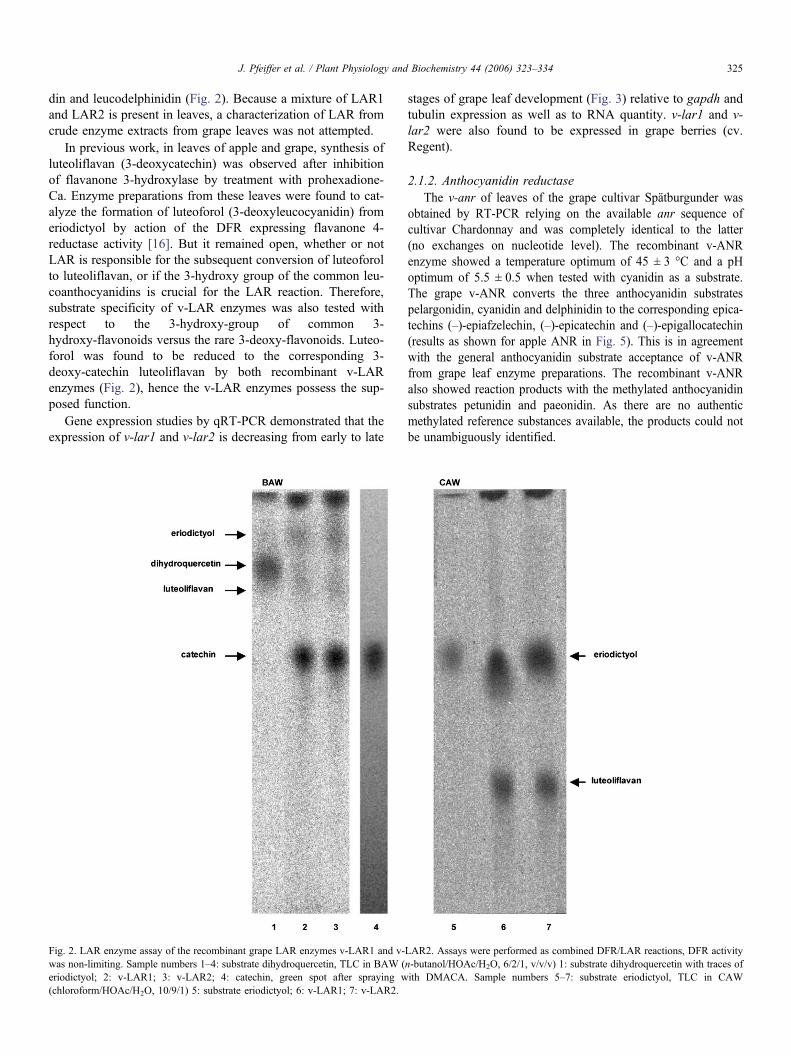
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334 325
din and leucodelphinidin (Fig. 2). Because a mixture of LAR1and LAR2 is present in leaves, a characterization of LAR fromcrude enzyme extracts from grape leaves was not attempted.
In previous work, in leaves of apple and grape, synthesis ofluteoliflavan (3-deoxycatechin) was observed after inhibitionof flavanone 3-hydroxylase by treatment with prohexadione-Ca. Enzyme preparations from these leaves were found to cat-alyze the formation of luteoforol (3-deoxyleucocyanidin) fromeriodictyol by action of the DFR expressing flavanone 4-reductase activity [16]. But it remained open, whether or notLAR is responsible for the subsequent conversion of luteoforolto luteoliflavan, or if the 3-hydroxy group of the common leu-coanthocyanidins is crucial for the LAR reaction. Therefore,substrate specificity of v-LAR enzymes was also tested withrespect to the 3-hydroxy-group of common 3-hydroxy-flavonoids versus the rare 3-deoxy-flavonoids. Luteo-forol was found to be reduced to the corresponding 3-deoxy-catechin luteoliflavan by both recombinant v-LARenzymes (Fig. 2), hence the v-LAR enzymes possess the sup-posed function.
Gene expression studies by qRT-PCR demonstrated that theexpression of v-lar1 and v-lar2 is decreasing from early to late
Fig. 2. LAR enzyme assay of the recombinant grape LAR enzymes v-LAR1 and v-was non-limiting. Sample numbers 1–4: substrate dihydroquercetin, TLC in BAW (eriodictyol; 2: v-LAR1; 3: v-LAR2; 4: catechin, green spot after spraying w(chloroform/HOAc/H2O, 10/9/1) 5: substrate eriodictyol; 6: v-LAR1; 7: v-LAR2.
stages of grape leaf development (Fig. 3) relative to gapdh andtubulin expression as well as to RNA quantity. v-lar1 and v-lar2 were also found to be expressed in grape berries (cv.Regent).
2.1.2. Anthocyanidin reductaseThe v-anr of leaves of the grape cultivar Spätburgunder was
obtained by RT-PCR relying on the available anr sequence ofcultivar Chardonnay and was completely identical to the latter(no exchanges on nucleotide level). The recombinant v-ANRenzyme showed a temperature optimum of 45 ± 3 °C and a pHoptimum of 5.5 ± 0.5 when tested with cyanidin as a substrate.The grape v-ANR converts the three anthocyanidin substratespelargonidin, cyanidin and delphinidin to the corresponding epica-techins (–)-epiafzelechin, (–)-epicatechin and (–)-epigallocatechin(results as shown for apple ANR in Fig. 5). This is in agreementwith the general anthocyanidin substrate acceptance of v-ANRfrom grape leaf enzyme preparations. The recombinant v-ANRalso showed reaction products with the methylated anthocyanidinsubstrates petunidin and paeonidin. As there are no authenticmethylated reference substances available, the products could notbe unambiguously identified.
LAR2. Assays were performed as combined DFR/LAR reactions, DFR activityn-butanol/HOAc/H2O, 6/2/1, v/v/v) 1: substrate dihydroquercetin with traces ofith DMACA. Sample numbers 5–7: substrate eriodictyol, TLC in CAW
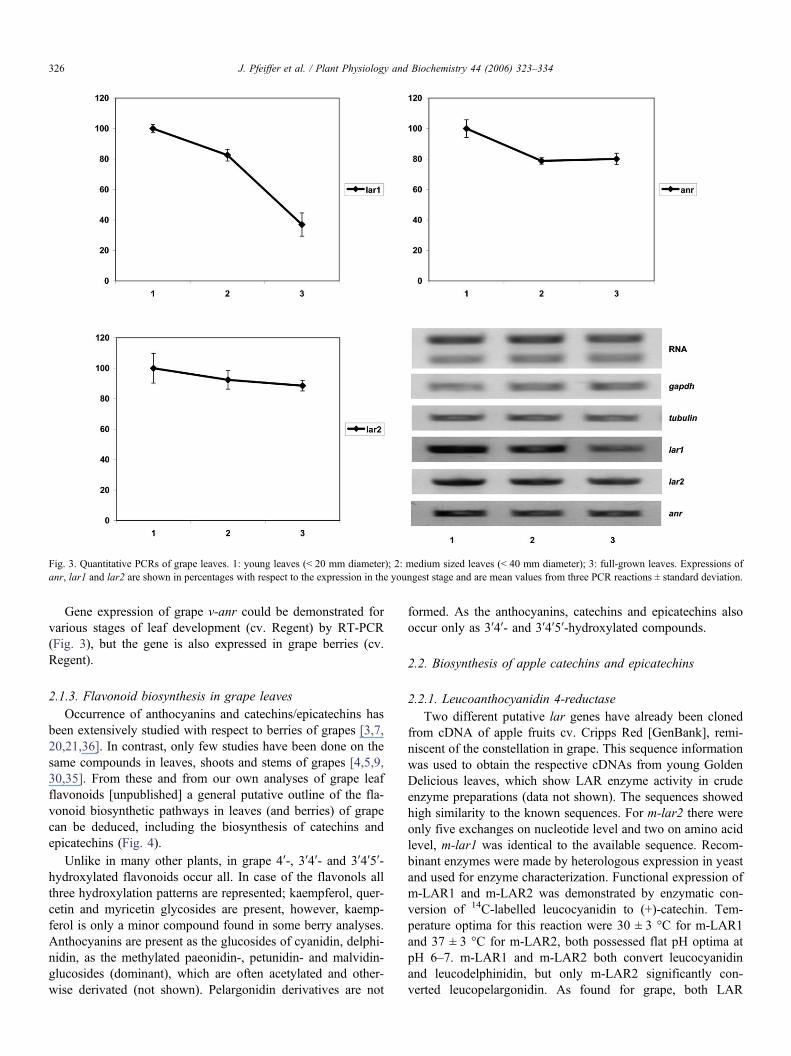
Fig. 3. Quantitative PCRs of grape leaves. 1: young leaves (< 20 mm diameter); 2: medium sized leaves (< 40 mm diameter); 3: full-grown leaves. Expressions ofanr, lar1 and lar2 are shown in percentages with respect to the expression in the youngest stage and are mean values from three PCR reactions ± standard deviation.
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334326
Gene expression of grape v-anr could be demonstrated forvarious stages of leaf development (cv. Regent) by RT-PCR(Fig. 3), but the gene is also expressed in grape berries (cv.Regent).
2.1.3. Flavonoid biosynthesis in grape leavesOccurrence of anthocyanins and catechins/epicatechins has
been extensively studied with respect to berries of grapes [3,7,20,21,36]. In contrast, only few studies have been done on thesame compounds in leaves, shoots and stems of grapes [4,5,9,30,35]. From these and from our own analyses of grape leafflavonoids [unpublished] a general putative outline of the fla-vonoid biosynthetic pathways in leaves (and berries) of grapecan be deduced, including the biosynthesis of catechins andepicatechins (Fig. 4).
Unlike in many other plants, in grape 4′-, 3′4′- and 3′4′5′-hydroxylated flavonoids occur all. In case of the flavonols allthree hydroxylation patterns are represented; kaempferol, quer-cetin and myricetin glycosides are present, however, kaemp-ferol is only a minor compound found in some berry analyses.Anthocyanins are present as the glucosides of cyanidin, delphi-nidin, as the methylated paeonidin-, petunidin- and malvidin-glucosides (dominant), which are often acetylated and other-wise derivated (not shown). Pelargonidin derivatives are not
formed. As the anthocyanins, catechins and epicatechins alsooccur only as 3′4′- and 3′4′5′-hydroxylated compounds.
2.2. Biosynthesis of apple catechins and epicatechins
2.2.1. Leucoanthocyanidin 4-reductaseTwo different putative lar genes have already been cloned
from cDNA of apple fruits cv. Cripps Red [GenBank], remi-niscent of the constellation in grape. This sequence informationwas used to obtain the respective cDNAs from young GoldenDelicious leaves, which show LAR enzyme activity in crudeenzyme preparations (data not shown). The sequences showedhigh similarity to the known sequences. For m-lar2 there wereonly five exchanges on nucleotide level and two on amino acidlevel, m-lar1 was identical to the available sequence. Recom-binant enzymes were made by heterologous expression in yeastand used for enzyme characterization. Functional expression ofm-LAR1 and m-LAR2 was demonstrated by enzymatic con-version of 14C-labelled leucocyanidin to (+)-catechin. Tem-perature optima for this reaction were 30 ± 3 °C for m-LAR1and 37 ± 3 °C for m-LAR2, both possessed flat pH optima atpH 6–7. m-LAR1 and m-LAR2 both convert leucocyanidinand leucodelphinidin, but only m-LAR2 significantly con-verted leucopelargonidin. As found for grape, both LAR

Fig. 4. General outline of the flavonoid biosynthetic pathways in grape leaves deduced from flavonoid occurrences and contents, for comparison biosynthetic steps inberries are also shown.
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334 327
enzymes also converted the 3-deoxy-leucoanthocyanidin luteo-forol to the 3-deoxy-catechin luteoliflavan (results as shown forgrape LAR enzymes in Fig. 2). This reaction occurs in appleleaves after artificial induction of luteoforol formation [32].
2.2.2. Anthocyanidin reductase
The m-anr of leaves of the apple cultivar Golden Deliciouswas obtained by RT-PCR relying on the available putative anrsequence young fruits of cultivar Cripps Red and was highlysimilar to the latter (two exchanges on nucleotide level, one onamino acid level). The recombinant m-ANR has a pH optimumof 5.0 ± 0.5 and a temperature optimum of 45 ± 3 °C with thesubstrate cyanidin. It has low substrate specificity and convertsthe three anthocyanidins pelargonidin, cyanidin and delphini-din to the respective epicatechins (Fig. 5). The same enzymecharacteristics as for the recombinant m-ANR enzyme werealso found for the ANR in enzyme preparations from appleleaves. The methylated anthocyanidin substrate petunidin isalso reduced by the recombinant m-ANR, but the methylatedanthocyanidins paeonidin and malvidin are not converted.
During normal apple (Golden Delicious) leaf developmentfrom the unrolled leaf to first full-grown leaf only a slight
decrease of specific ANR enzyme activity (referring to protein)was found.
2.2.3. Flavonoid biosynthesis in apple leavesFlavonoid biosynthesis in apple leaves has previously been
studied by flavonoid analyses from adult leaves [42]. Fromthese results the outline of flavonoid biosynthesis in appleleaves can be deduced (Fig. 6), which is also valid for applefruits.
Generally, in both, apple leaves and fruits, almost exclu-sively 3′4′-dihydroxylated anthocyanins (cyanidin-glycosides)as well as the 3′4′-dihydroxylated (+)-catechin and (–)-epicatechin are formed. Leaves of apple cultivars possessinganthocyanins (the red-leaved cultivars Niedzwetzkyana,Weirouge) and those without anthocyanins (the green-leavedcultivars Klarapfel, Golden Delicious, Red Delicious, Elstar,Holsteiner Cox, Pinova, Pilot) were studied to test for correla-tion between the presence of cyanidin-glycosides and (–)-epicatechin, both derived from the common precursor cyanidin,and the corresponding enzyme activities of ANR and UFGT.ANR enzyme activity and (–)-epicatechin are present inboth types of leaves (Table 1). Anthocyanin biosynthesis inthe respective tissues occurs only in cultivars with red leaves
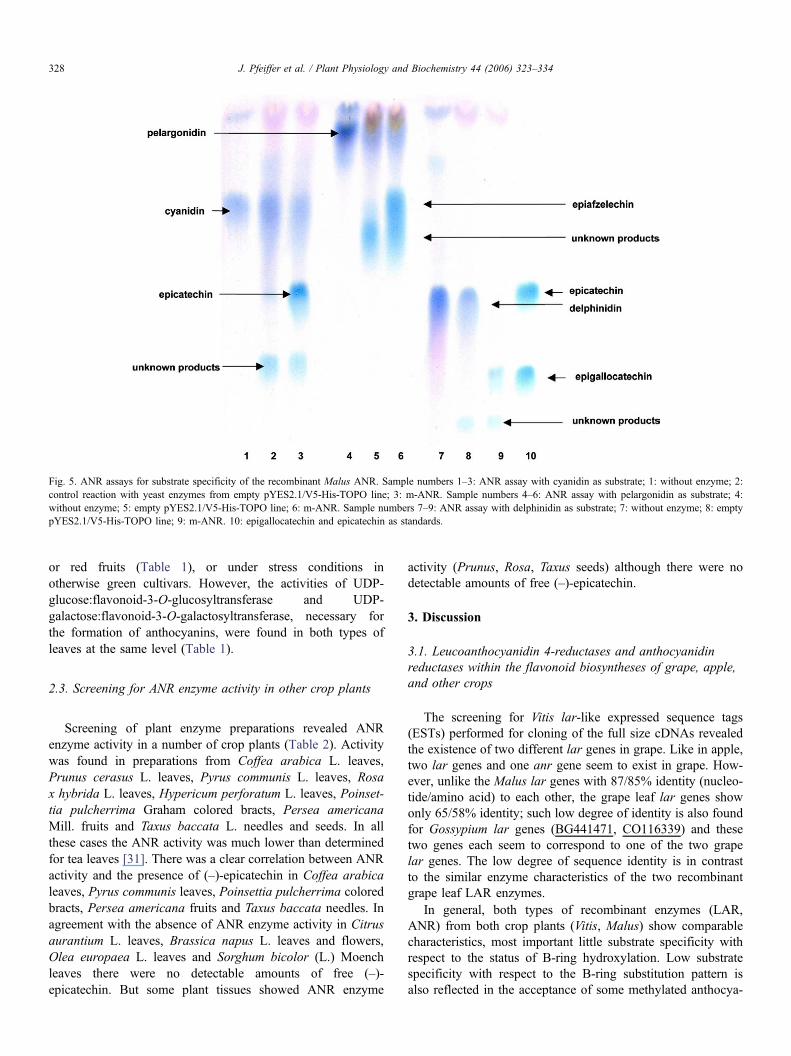
Fig. 5. ANR assays for substrate specificity of the recombinant Malus ANR. Sample numbers 1–3: ANR assay with cyanidin as substrate; 1: without enzyme; 2:control reaction with yeast enzymes from empty pYES2.1/V5-His-TOPO line; 3: m-ANR. Sample numbers 4–6: ANR assay with pelargonidin as substrate; 4:without enzyme; 5: empty pYES2.1/V5-His-TOPO line; 6: m-ANR. Sample numbers 7–9: ANR assay with delphinidin as substrate; 7: without enzyme; 8: emptypYES2.1/V5-His-TOPO line; 9: m-ANR. 10: epigallocatechin and epicatechin as standards.
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334328
or red fruits (Table 1), or under stress conditions inotherwise green cultivars. However, the activities of UDP-glucose:flavonoid-3-O-glucosyltransferase and UDP-galactose:flavonoid-3-O-galactosyltransferase, necessary forthe formation of anthocyanins, were found in both types ofleaves at the same level (Table 1).
2.3. Screening for ANR enzyme activity in other crop plants
Screening of plant enzyme preparations revealed ANRenzyme activity in a number of crop plants (Table 2). Activitywas found in preparations from Coffea arabica L. leaves,Prunus cerasus L. leaves, Pyrus communis L. leaves, Rosax hybrida L. leaves, Hypericum perforatum L. leaves, Poinset-tia pulcherrima Graham colored bracts, Persea americanaMill. fruits and Taxus baccata L. needles and seeds. In allthese cases the ANR activity was much lower than determinedfor tea leaves [31]. There was a clear correlation between ANRactivity and the presence of (–)-epicatechin in Coffea arabicaleaves, Pyrus communis leaves, Poinsettia pulcherrima coloredbracts, Persea americana fruits and Taxus baccata needles. Inagreement with the absence of ANR enzyme activity in Citrusaurantium L. leaves, Brassica napus L. leaves and flowers,Olea europaea L. leaves and Sorghum bicolor (L.) Moenchleaves there were no detectable amounts of free (–)-epicatechin. But some plant tissues showed ANR enzyme
activity (Prunus, Rosa, Taxus seeds) although there were nodetectable amounts of free (–)-epicatechin.
3. Discussion
3.1. Leucoanthocyanidin 4-reductases and anthocyanidinreductases within the flavonoid biosyntheses of grape, apple,and other crops
The screening for Vitis lar-like expressed sequence tags(ESTs) performed for cloning of the full size cDNAs revealedthe existence of two different lar genes in grape. Like in apple,two lar genes and one anr gene seem to exist in grape. How-ever, unlike the Malus lar genes with 87/85% identity (nucleo-tide/amino acid) to each other, the grape leaf lar genes showonly 65/58% identity; such low degree of identity is also foundfor Gossypium lar genes (BG441471, CO116339) and thesetwo genes each seem to correspond to one of the two grapelar genes. The low degree of sequence identity is in contrastto the similar enzyme characteristics of the two recombinantgrape leaf LAR enzymes.
In general, both types of recombinant enzymes (LAR,ANR) from both crop plants (Vitis, Malus) show comparablecharacteristics, most important little substrate specificity withrespect to the status of B-ring hydroxylation. Low substratespecificity with respect to the B-ring substitution pattern isalso reflected in the acceptance of some methylated anthocya-

Fig. 6. General outline of the flavonoid biosynthetic pathways in leaves andfruits of apple.
Table 2Correlation of the presence of ANR enzyme activity and the occurrence of free(–)-epicatechin in a selected range of crop plants. (-: no enzyme activity; +: lowenzyme activity; ++: moderate enzyme activity)
Crop plant tissue ANR enzymeactivity
Free (–)-epicatechin(ng mg–1 dry weight)
Coffea arabica leaves + + (not quantified)Prunus cerasus leaves ++ 0Pyrus communis leaves ++ 30Rosa x hybrida leaves + TracesHypericum perforatum leaves ++ UnidentifiedPoinsettia pulcherrima bracts ++ 4Persea americana fruits + 20Taxus baccata needles ++ 260Taxus baccata seeds + 0Citrus aurantium leaves – 0Brassica napus leaves – 0Brassica napus flowers – 0Olea europaea leaves – 0Sorghum bicolor leaves – 0
Table 1Anthocyanin, (–)-epicatechin and (+)-catechin contents and presence of UFGT and ANR activities in leaves of red and green-leaved cultivars. Flavonoid contents aregiven in mg g–1 dried adult apple leaves. +: enzyme activity present
Cultivar Niedzwetz-kyana Weirouge Klarapfel GoldenDelicious
RedDelicious
Elstar HolsteinerCox
Pinova Pilot
(+)-Catechin 0 0.1 0 0.1 0.1 0.1 0.1 0.2 0.1(–)-Epicatechin 0.3 0.2 0.1 0.2 0.3 0.3 0.3 0.4 0.2ANR activity + + + + + + + + +Cyanidin O-glycosides 1.9 2.9 0 0 0 0 0 0 0UFGT activity + + + + + + + + +
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334 329
nidins as substrates in case of the ANR enzymes. The broadsubstrate acceptance is in contrast to the presence of specificand differently hydroxylated flavan 3-ols in grape and apple.Obviously, for explanation of the respective functions of theLAR and ANR enzymes, the whole flavonoid metabolismshave to be taken in consideration, with differing substrateavailabilities and different enzymatic steps, which are in com-petition to flavan 3-ol formation.
In grape 4′-, 3′4′- and 3′4′5′-hydroxylated flavonols (-glycosides) are formed (Fig. 4), with kaempferol (mono-hydroxylated) being only a minor compound in berries. Antho-cyanins (and their various derivates), catechins and epicate-chins occur only as 3′4′- and 3′4′5′-hydroxylated flavonoids.With respect to the general acceptance of 4′-, 3′4′- and 3′4′5′-hydroxylated substrates by grape ANR and LAR1/LAR2, butexclusive formation of 3′4′- and 3′4′5′-hydroxylated flavan 3-ols ((+)-catechin, (+)-gallocatechin and (–)-epicatechin, (–)-epigallocatechin) in vivo, substrate availability for ANR andLAR1/LAR2 has to be decisive for flavan 3-ol biosynthesisin grape leaves. As the grape DFR shows no substrate specifi-city [16], the absence of 4′-hydroxylated flavan 3-ols andanthocyanins could be due to strong flavonoid 3′- and 3′5′-hydroxylase activities.
Flavonoid biosynthesis in apple fruit has been the subject ofmany studies, mainly at the level of flavonoid composition andcontents and including also (+)-catechins, (–)-epicatechins andproanthocyanidins [1,17,18,42]. In addition, some studies atthe level of gene expression and enzyme activity were per-formed [13,22 and citations therein, 25]. In contrast, flavonoidbiosynthesis in apple leaves is less investigated but is importantwith regard to flavanols as pathogen resistance factors. In appleleaves (and fruits), like for the other classes of flavonoid endproducts (flavonol glycosides, anthocyanins), only the 3′4′dihydroxylated flavan 3-ols (+)-catechin and (–)-epicatechinare formed (Fig. 6). With flavonoid 3′-hydroxylase acting onnaringenin and dihydrokaempferol, the derived 3′4′ dihydroxy-lated leucocyanidin is the common precursor for (+)-catechin,cyanidin (-glycosides) and (–)-epicatechin. In green-leaved cul-tivars, (–)-epicatechin is regularly found, indicating that cyani-din is formed and reduced to (–)-epicatechin, but cyanidin isnot glycosylated to the respective anthocyanins. However, fla-vonoid UDP-glycosyl-3-O-glycosyltransferase (UFGT) activitywas unexpectedly demonstrated from leaf extracts of red- aswell as from green-leaved apple cultivars. The presence of(–)-epicatechin demonstrates the principal availability of cyani-din as a substrate for ANR and UFGT also in green leaves.

Fig. 7. Possible reaction mechanisms of the leucoanthocyanidin 4-reductasereaction as supported by the conversion of the 3-deoxyleucoanthocyanidinluteoforol. R=H demonstrates the intermediates of 3-deoxyleucoanthocyanidinconversion, R=OH the intermediates of normal leucoanthocyanidin conversion.
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334330
With UFGT being present in red and green leaves, the relativeenzymatic activity of ANR to UFGT, or a temporal or spatialseparation (compartmentalization), will be crucial for directingthese final steps towards anthocyanin and/or (–)-epicatechin.The transport of anthocyans or proanthocyanidins to thevacuole may also be decisive. Seemingly, in young leaves ofrose, which are red due to presence of cyanidin-glycosides, thereverse constellation as in green apple leaves is realized.Despite the presence of ANR enzyme activity, almost no (–)-epicatechin is formed there, but only anthocyanin instead [26].This question has to be subjected to further investigations.
With regard to practical relevance, for apple catechins, epi-catechins and proanthocyanidins are important secondary meta-bolites active against apple scab [43], a major fungal disease ofapple leaves and fruits. The conversion of the 3-deoxy-leucoanthocyanidin luteoforol to luteoliflavan by grapeand apple LAR enzymes is also of practical relevance. Theinduction of the antimicrobial luteoforol [19] in leaves ofapple and other crops [16,32] is accompanied by partial resis-tance to the fireblight agent Erwinia amylovora, but isobviously counteracted by LAR activity which leads to theinactive luteoliflavan. Thus, like DFR, which can act as a fla-vanone 4-reductase (FNR) in luteoforol formation, LAR showslow substrate specificity with respect to the 3-hydroxy functionin its substrate and can perform other reactions.
Biosynthesis of secondary metabolites in different plantsmay occur via different pathways or, in case of comparablepathways, simply via different enzymes. For flavonoid bio-synthesis the reaction from flavanones to flavones is an exam-ple, where the oxidation may either be performed by NADPH-dependent cytochrome P450 monooxygenases in some plants(flavone synthase II [28], most plant species), but also by a 2-oxoglutarate-dependent dioxygenase (flavone synthase I inApiaceae [29]). For this reason, the correlation of (–)-epicatechin occurrence and presence of ANR enzyme activitywas studied in a small series of crop plants. Correlation ofANR enzyme activity with presence of (–)-epicatechin wasfound, but in two crops ANR activity was demonstrated with-out presence of detectable amounts of (–)-epicatechin. How-ever, (–)-epiafzelechin or (–)-epigallocatechin, which were notanalyzed for, may be the natural ANR products in these cases,or, alternatively, (–)-epicatechin may be completely convertedto proanthocyanidins or other derivatives.
The biological relevance of the quite different characteris-tics of LAR, ANR and UFGT concerning pH optima and tem-perature optima has to be considered. LAR and UFGTenzymes possess optima at 6–7 and 7–8, respectively, whereasANR enzymes have optima approximately at pH 5, with Ara-bidopsis ANR with pH 8.0 being an obvious exception [44].However, ANR is still active at pH 7.0, so that parallel activ-ities of the three enzymes in the cytosol seem possible. Theextremely high temperature optimum of ANRs at 45 °C (withconsiderable activity at even higher temperatures) may possessa biological function. With anthocyanin formation by UFGT asthe competing process, for high temperatures a reduction ofanthocyanin formation could reduce light absorption. Conver-
sely, at low temperatures (stress, autumn colors) anthocyaninformation could be the favored process.
3.2. Substrate specificity and possible reaction mechanismsof leucoanthocyanidin 4-reductases
v-LAR and m-LAR enzymes are able to convert the3-deoxy-leucoanthocyanidin luteoforol (1) to the 3-deoxy-catechin luteoliflavan (6) (Fig. 7). This demonstrates asubstrate recognition not necessarily including the 3-hydroxyfunction and supports a LAR reaction mechanism involving the4-hydroxy function, but without direct participation of the 3-hydroxy function.
Assuming that reduction does not occur directly on leu-coanthocyanidins (2), but on a double bond or on a carbocationinstead, it will depend on the LAR reaction sequence whether a

J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334 331
carbocation (3) is first formed by protonation/water eliminationat C-4, or if water elimination from C-4 leads first to the flav-3-en/flav-3-en 3-ol (4) or, alternatively, to the p-quinonemethide (5) (or o-quinone methide, not shown). Both, theflav-3-en (3-ol) (4) and the quinone methides could also resultin the carbocation (3) after protonation at O-3 or O-7, O-5.Subsequently, the intermediates (3, 4 or 5) would be reducedby NADPH. In case of the flav-3-en (3-ol) (4), the reductionwould have to take place stereospecifically by the LARenzyme to lead to the flavan 3β-ol (natural (+)-catechin) (7)[39]; for the carbocation (3) and the p-quinone methide (5)the configuration at C-3 would be preserved during the reac-tion.
However, the intermediates (3, 4, 5) could also represent theactual electrophilic substrates of the chain elongation inproanthocyanidin synthesis, as it was already discussed indetail [46]. For this, the reaction would either have to occurby LAR itself as an alternative reaction to reduction of an inter-mediate, or an intermediate would have to be released and reactseparately from LAR chemically or enzymatically to formproanthocyanidins. As it was stated [46], it is possible that cor-responding intermediates are formed by the ANR reaction too.These are also potential substrates for chain elongation inproanthocyanidin biosynthesis.
3.3. Perspective
Elucidation of the polymerization reaction formingproanthocyanidins is of course a central part of future studies.Currently, availability of lar and anr gene sequences from cropplants and reliable enzyme tests allow studies on gene expres-sions and corresponding enzyme activities, their correlationwith biosynthetic products and their biological effects. Influ-ences on biosynthesis by environmental parameters may alsohave an impact on crop cultivation practices with respect tocrop pathogen resistances and promotion of ingredients bene-ficial for human health.
4. Methods
4.1. Plant material
Plant material from the various apple cultivars was kindlyprovided by Iris Szankowski (Vegetable and Fruit Science,University of Hannover) and Henryk Flachowsky (FederalCentre for Breeding Research on Cultivated Plants (BAZ),Dresden/Pillnitz). Plant material from the grape cultivarsRegent and Spätburgunder was collected from plants cultivatedin Freising (Fruit Science, Technical University Munich). Allplant material was frozen in liquid nitrogen immediately afterharvest and transported or stored at –80 °C until use.
4.2. Flavonoid analysis
(–)-Epicatechins and (+)-catechins were analyzed by HPLCas described in [33]. Cyanidin-glycosides were identified by
their UV/VIS spectra and by using authentic reference sub-stances.
4.3. Extraction of enzymes
The enzymes of the divergent plant tissues were isolatedapplying a method suitable for lignocellulosic tissues [10].For enzyme preparations from grape young leaves of cv.Regent were used, various stages of developing cv. GoldenDelicious leaves were used for enzyme preparations fromapple. Protein concentrations were determined according to [6].
4.4. Cloning of grape v-lar1 and v-lar2
Leaves from Vitis vinifera cv. Regent were used to cloneboth lar cDNAs. Total RNA from grape leaves was extractedwith Concert (TM) Plant RNA Reagent (Invitrogen) accordingto the manufacturer’s protocol. cDNA was synthesized withMMLV Reverse Transcriptase (Promega) using 1 μg of totalRNA. EST sequences from Vitis similar to lar from Desmo-dium (AJ550154) or apple (AY830131, AY830132) were iden-tified by BLAST N searches. From the alignment of the result-ing ESTs two distinctly different lar-like sequences could beassembled, a complete CDS of one lar (v-lar1) and the 3′region of a second lar (v-lar2).
v-lar1: Primers 5′-GTAGAGAAGAAGCATCATCAG-3′for the 5′ and 5′-GGAGGACTAATCGCATGCATG-3′ forthe 3′ non-coding region of v-lar1 were designed from analignment of the following EST Sequences from GenBank:Vitis shuttleworthii CV092592, CV094089, CV097143,CV100198, CV100739, Vitis riparia CN725607, CN725609and Vitis vinifera CF207251, CF514437.
v-lar2: Alignment of three EST Sequences from Vitis vini-fera (CB979225, CF207282, CF514225) yielded only the 3′region of v-lar2. Using a forward primer 5′-GCAAYTCMATYGCHKCTTGGCC-3′, degenerated to fit toboth, v-lar1 and v-lar2, and two nested reverse primers 5′-CACCAATATTACTAGATTCCAATAAG-3′ (3ViLAR4) and5′-GAWTRGGGAGCTGGCCACATC-3′ (3ViLAR3) from the3′ non-coding region specific to v-lar2, the 3′ region of v-lar2was cloned and sequenced. From this sequence new primerswere designed to perform a 5′-RACE. A shortened RLM-RACE protocol (Invitrogen GeneRacer), using only one preci-pitation, was used for the RACE-cDNA. The 5′ region of v-lar2 was cloned by nested PCRs, using the GeneRacer 5′primer and nested 5′ primer and as reverse primers 3ViLAR4and 5′-GAGCCATCACCGTAGATATGGAAACG-3′, respec-tively. The amplification of the full-size v-lar2 was performedwith the primers 5′-CTCAGAAAATTAACACCGAT-3′ fromthe 5′ and 3ViLAR3 from the 3′ non-coding region. Thesequences were deposited as DQ129685 (v-lar1) andDQ129686 (v-lar2).
4.5. Heterologous expression in yeast
Proof-reading RT-PCR amplification (High Fidelity®,Roche) of the v-anr from Vitis vinifera cv. Spätburgunder leaf

J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334332
cDNA relied on the available GenBank sequence of a putativeanr from cv. Chardonnay (BN000166) with the derived pri-mers 5′-GCGAATTTGGAAGTGAGG-3′ from the 5′ and5′-GAGGGATAAGATAGTGCTCC-3′ from the 3′ non-coding region (Spätburgunder v-anr sequence deposited asDQ129684). The v-lar1 and v-lar2 cDNAs cloned from Vitisvinifera cv. Regent leaf cDNA were directly PCR-amplified forthe cloning in the yeast expression vector (pYes2.1 TOPO®,Invitrogen) using proofreading Taq-Polymerase. Malusx domestica anr (AY830130), lar1 (AY830131) and lar2(AY830132) were RT-PCR amplified from cDNA of cv.Golden Delicious leaves [15] with primers derived from therespective 5′ and 3′ non-coding regions (5MANR 5′-GACAGAGGAAGAGGAAACCATG-3′, 3MANR 5′-GCCAGACAATAGAGAGATAGC-3′, 5MLAR1 5′-TCTGTAGCAGGAARGAAGAGC-3′, 3MLAR1 5′-CRGTCYTGATTCKGATAGATTG-3′, 5MLAR2 5′-CTTTTCAAACTTGAGGGCA-3′, 3MLAR2 5′-CACCTCACATAATTTACCAGAC-3′). The sequences weredeposited as DQ139835, DQ139836 and DQ139837. Hetero-logous expressions of the cDNAs in yeast were performed aspreviously described [15].
4.6. Gene expression in plant tissues
Expression of the respective mRNAs of lar1, lar2 and anrin leaves and fruits of grape were determined by end-pointquantitative RT-PCR amplification using the primers that hadbeen used for the cDNA cloning. Young (< 20 mm diameter),medium (< 40 mm diameter), and full-grown leaves of grapecv. Regent were used. For each specific PCR reliable proce-dures, PCR conditions, and a cycle number within thelinear/half-logarithmic range of amplification were determinedwith several series of cDNA dilutions. Amplification of tubulinand gapdh as house-keeping genes with the primers 5′-TTGTTGAGCCATACAATGC-3′ and 5′-AGTACCAATGCAAGAAAGC-3′ (tubulin), and 5′-GATGTTGARCTHGTYGCWGTBAAYGAYCC-3′ and 5′-GCAAGRCARTTRGTRGTRCARCTRGC-3′ (gapdh) wereused as controls for equal RNA amounts and even reverse tran-scription efficacy in the qPCR tests series. For each gene, theexpression was determined threefold in separate PCRs with amixture of three separate cDNAs to avoid errors induced by thevariability of cDNA synthesis as well as variation between dif-ferent PCR reactions. Expression data were quantified by gra-phically integrating amounts of ethidiumbromide stained PCRproduct from UV image files (Quantity One Basic, Biorad), allvalues were within the dynamic range of the procedure.
4.7. Substrate synthesis
The 14C-labelled flavonoid substrates ERI, DHK, DHQ andDHM were synthesized from 14C-malonyl-CoA andcoumaroyl-CoA according to [15,31].
4.8. Leucoanthocyanidin 4-reductase (LAR) enzyme assays
Because of the high instability of leucoanthocyanidins, allLAR assays were performed as combined DFR/LAR reactions.The LAR assays for substrate specificity contained2R,3R-[4a,6,8-14C] dihydrokaempferol, or 2R,3R-[4a,6,8-14C]dihydroquercetin, or 2R,3R-[4a,6,8-14C] dihydromyricetin(0.0375 nmol, specific activity 1111 bq nmol–1), 10 μl ofNADPH (200 nmol), 110 μl 0.1 M Tris–HCl buffer (pH 7.5),recombinant DFR enzyme from Malus x domestica [15], andthe respective recombinant LAR to be tested (300 μg total pro-tein each) in a final volume of 200 μl and was incubated for30 min at 25 °C. Correspondingly, for the LAR tests with the3-deoxy-leucoanthocyanidin luteoforol, the labeled luteoforolwas produced by incubation of 2S-[4a,6,8-14C] eriodictyolwith Malus x domestica DFR acting as flavanone 4-reductase,the incubation was in 0.1 M KPi (pH 6.0) at 30 °C for 30 min,and another 30 min after addition of the respective LAR. TheLAR assays for determination of the LAR temperature opti-mum were performed by preincubation of 2R,3R-[4a,6,8-14C]dihydroquercetin with the recombinant DFR enzyme fromMalus x domestica [15] for 30 min at 25 °C (generating labeledleucocyanidin in vitro) followed by the addition of recombi-nant LAR and continued incubation for further 30 min at var-ious temperatures. The LAR assays for determination of the pHoptimum were performed in the same way, but the buffer forthe DFR preincubation was 20 mM KPi (pH 7.5), for subse-quent LAR incubation 110 μl of 0.1 mM KPi buffer of variouspH values were added, resulting in the respective pH values forthe test. Control reactions containing only the DFR enzymewere also carried out for all assays to demonstrate sufficientavailability of LAR substrate. All assays were extracted twicewith 200 μl EtOAc, analyzed by TLC and the products wereidentified and quantified as described in [31].
4.9. Anthocyanidin reductase (ANR) assay and (–)-epicatechinanalysis
The ANR enzyme assays contained 50 μl of 20 mMNADPH, 50 μl cyanidin chloride (Roth) in EGME (1 mgml–1) and 100 μg enzyme in a total of 500 μl 0.1 M KPi(pH 5.5) and were incubated for 60 min at 45 °C. Reactionproducts were extracted twice with 200 μl ethylacetate(EtOAc), vacuum-dried and redissolved in 60 μl methanol forHPLC analysis. For determination of ANR substrate specificitythe enzymatic reactions with the anthocyanidins pelargonidin,cyanidin, delphinidin, and the methylated anthocyanidins paeo-nidin, petunidin and malvidin were performed and the reactionproducts were EtOAc extracted and subjected to TLC on cellu-lose plates in BAW (n-butanol/HOAc/H2O, 6/2/1, v/v/v).Authentic (–)-epicatechin and (–)-epigallocatechin were avail-able as reference substances and were co-analyzed by chroma-tography, all epicatechin products were subsequently stainedand identified with the specific reagent DMACA (0.1% inEtOH/6 N HCl, 1/1, v/v). The ANR products (–)-epiafzelechin, (–)-epicatechin and (–)-epigallocatechin detectedas DMACA positive blue-green spots after TLC had previously

[2]
[3]
[4]
[5]
[6]
[7]
[8]
[9]
[10
[11
[12
[13
[14
[15
[16
[17
[18
[19
[20
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334 333
been additionally confirmed for the tea ANR by HPLC/MS/MS[31]. To screen for the presence of ANR enzyme activity inselected crop plants 100 μg total protein each was used forthe enzyme tests, which were incubated for 60 min at 37 °C;analysis was by HPLC and by TLC/DMACA. For demonstra-tion of the presence of ANR activities in enzyme preparationsfrom the different apple cultivars, analysis of product formationwas also performed by TLC/DMACA. For direct (–)-epicatechin analysis from plant tissues, the dried plant tissueswere homogenized, extracted with methanol supported byultrasonic sound treatment, and the supernatant organic phaseapplied for HPLC-analysis as described for the ANR enzymeassays.
4.10. UDP-glucose:flavonoid-3-O-glucosyltransferaseand UDP-galactose:flavonoid-3-O-galactosyltransferaseassays (UFGT)
For the UFGT enzyme tests 5 μl 15 mM UDP-glucose orUDP-galactose, 10 μl 6 mM cyanidin chloride in EGME and50 μg enzyme in a final volume of 200 μl 50 mM KPi (pH 7.0)were incubated for 30 min at 30 °C. The reaction was stoppedwith 100 μl CHCl3/MeOH (0.5% HCl) (2/1, v/v) by vortexingand centrifuged for 2 min. 100 μl of the upper phase wereapplied to a cellulose TLC-plate (Merck). After additionalapplication of authentic cyanidin 3-glucoside or cyanidin 3-galactoside as references, the TLC was run in GT(H2O/HOAc/HCl, 85/15/3, v/v/v). Anthocyanin reaction pro-ducts were detected visually.
Acknowledgements
The “Bayerisches Staatsministerium für Landesentwicklungund Umweltfragen” is acknowledged for the financial supportof the project “Induzierte Pathogenabwehr”, the Department ofResearch Cooperation (SAREC) of the Swedish InternationalDevelopment Cooperation, Sida, for financial support throughBiochemical Pest Control Project for P.A.N. Punyasiri to carryout work at the Technical University of Munich, Germany. Dr.Thorsten Strissel and Professor Dr. Dieter Treutter (FruitScience, Technical University Munich) are acknowledged forhelp with HPLC analyses performed in their lab, Professor Dr.Iris Szankowski (Vegetable and Fruit Science, University ofHannover) and Dr. Henryk Flachowsky (Federal Centre forBreeding Research on Cultivated Plants (BAZ), Dresden/Pill-nitz) for providing us with frozen apple leaf material of variouscultivars. Bettina Deiml is thanked for manifold support. Dr.Christian Seitz (Floriculture, TU Munich) is acknowledgedfor providing a flavonoid-3′5′-hydroxylase expression clonefor substrate synthesis. We are grateful to Dr. Henriette A.Remmer (University of Michigan) and Dr. Uta von Rad(BIOP, Neuherberg) for proofreading the manuscript.
References
[1] R.M. Alonso-Salces, K. Ndjoko, E.F. Queiroz, J.R. Ioset, K. Hostett-mann, L.A. Berrueta, B. Gallo, F. Vicente, On-line characterisation of
apple polyphenols by liquid chromatography coupled with mass spectro-metry and ultraviolet absorbance detection, J. Chromatogr. A. 1046(2004) 89–100.G.R. Beecher, Proanthocyanidins, Biological activities associated withhuman health, Pharmaceutical Biology 42 (2004) 2–20.P.K. Boss, C. Davies, S.P. Robinson, Analysis of the expression ofanthocyanin pathway genes in developing Vitis vinifera L. cv shirazgrape berries and the implications for pathway regulation, Plant Physiol.111 (1996) 1059–1066.M. Boukharta, M. Girardin, M. Metche, Procyanidines galloylées du sar-ment de vigne (Vitis vinifera) : séparation et identification par chromato-graphie liquide haute performance et chromatographie en phase gazeuse,J. Chromatogr. A. 455 (1988) 406–409.M. Bourzeix, D. Weyland, N. Heredia, Study of catechins and procyani-dins in grape clusters, wine, and other vine products, Bull. OIV 59(1986) 1171–1254.M.M. Bradford, A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding, Anal. Biochem. 72 (1976) 248–254.S. Carando, P.-L. Teissedre, L. Pascual-Martinez, J.-C. Cabanis, Levelsof flavan-3-ols in French wines, J. Agric. Food Chem. 47 (1999) 4161–4166.G.H. Dai, C. Andary, L. Mondolot-Cosson, D. Boubals, Histochemicalstudies on the interaction between three species of grapevine, Vitis vini-fera, V. rupestris and V. rotundifolia and the downy mildew fungus, Plas-mopara viticola, Physiol. Mol. Plant Pathol. 46 (1995) 3.G. Darné, Y. Glories, Anthocyanins in leaves of some Vitis vinifera L.cultivars between veraison and leaf fall, Vitis 27 (1988) 71–78.
] V. Dellus, W. Heller, Heinrich Sandermann (Jr.), A. Scalbert, Dihydro-flavonol 4-reductase activity in lignocellulosic tissues, Phytochemistry 45(1997) 1415–1418.
] M. Devic, J. Guilleminot, I. Debeaujon, N. Bechtold, E. Bensaude, M.Koornneef, G. Pelletier, M. Delseny, The BANYULS gene encodes aDFR-like protein and is a marker of early seed coat development, PlantJ. 19 (1999) 387–398.
] R.A. Dixon, D.-Y. Xie, S.B. Sharma, Proanthocyanidins—a final frontierin flavonoid research?, New Phytol. 165 (2005) 9–28.
] Y.-H. Dong, L. Beuning, K. Davies, D. Mitra, B. Morris, A. Kootstra,Expression of pigmentation genes and photo-regulation of anthocyaninbiosynthesis in developing Royal Gala apple flower, Austr. J. Plant Phy-siol. 25 (1998) 245–252.
] W. Feucht, D. Treutter, Principles and Practices of Plant Ecology, in: S.Inderjit, K.M.M. Dakshini, C.L. Foy (Eds.), CRC Press, Bota Raton,Florida, 1999, pp. 307–338.
] T.C. Fischer, H. Halbwirth, B. Meisel, K. Stich, G. Forkmann, Molecularcloning, substrate specificity of the functionally expressed dihydroflavo-nol 4-reductases from Malus domestica and Pyrus communis cultivarsand the consequences for flavonoid metabolism, Arch. Biochem. Bio-phys. 412 (2003) 223–230.
] C. Gosch, I. Puhl, H. Halbwirth, K. Schlangen, S. Roemmelt, C.Andreotti, G. Costa, T.C. Fischer, D. Treutter, K. Stich, G. Forkmann,Effect of prohexadione-Ca on various fruit crops: flavonoid compositionand substrate specificity of their dihydroflavonol 4-reductases, Eur. J.Hort. Sci. 68 (2003) 144–151.
] S. Guyot, N. Marnet, D. Laraba, P. Sanoner, J.-F. Drilleau, Reversed-phase HPLC following thiolysis for quantitative estimation and charac-terization of the four main classes of phenolic compounds in differenttissue zones of a French cider apple variety (Malus domestica Var. Ker-merrien), J. Agric. Food Chem. 46 (1998) 1698–1705.
] S. Guyot, N. Marnet, J.-F. Drilleau, Thiolysis-HPLC characterization ofapple procyanidins covering a large range of polymerization states, J.Agric. Food Chem. 49 (2001) 14–20.
] H. Halbwirth, T.C. Fischer, S. Roemmelt, F. Spinelli, K. Schlangen, S.Peterek, E. Sabatini, C. Messina, J.-B. Speakman, C. Andreotti, W.Rademacher, C. Bazzi, G. Costa, D. Treutter, G. Forkmann, K. Stich,Induction of antimicrobial 3-deoxyflavonoids in pome fruit trees controlsfire blight, Z. Naturforsch. 58c (2003) 765–770.
] J.B. Harborne, The Flavonoids, Chapman & Hall, London, 1994.

[21
[22
[23
[24
[25
[26
[27
[28
[29
[30
[31
[32
[33
[34
[35
[36
[37
[38
[39
[40
[41
[42
[43
[44
[45
[46
J. Pfeiffer et al. / Plant Physiology and Biochemistry 44 (2006) 323–334334
] K. Herrmann, Inhaltsstoffe von Obst und Gemüse, Verlag Eugen Ulmer,Hohenheim, Stuttgart, 2001.
] C. Honda, N. Kotoda, M. Wada, S. Kondo, S. Kobayashi, J. Soejima, Z.Zhang, T. Tsuda, T. Moriguchi, Anthocyanin biosynthetic genes arecoordinately expressed during red coloration in apple skin, Plant Physiol.Biochem. 40 (2002) 955–962.
] N. Ishikura, H. Murakami, Y. Fujii, Conversion of (+)-dihydroquercetinto 3,4-cis-leucocyanidin by a reductase extracted from cell suspensioncultures of Cryptomeria japonica, Plant Cell Physiol. 29 (1988) 795–799.
] W.-S. Jeong, A.-N.T. Kong, Biological Properties of monomeric andpolymeric catechins, Green Tea Catechins and Procyanidins, Pharmaceu-tical Biology 42 (2004) 84–93.
] S.-H. Kim, J.-R. Lee, S.-T. Hong, Y.-K. Yoo, G. An, S.-R. Kim, Mole-cular cloning and analysis of anthocyanin biosynthesis genes preferen-tially expressed in apple skin, Plant Sci. 165 (2003) 403–413.
] J. Knott, Biochemische Untersuchungen zur Flavonoidbiosynthese beiRose und Kartoffel, Technical University of Munich, 2004.
] K.N. Kristiansen, Conversion of (+)-dihydroquercetin to (+)-2,3-trans-3,4-cis-leucocyanidin and (+)-catechin with an enzyme extractfrom maturing grains of barley, Carlsberg Res. Comm. 51 (1986) 51–60.
] S. Martens, G. Forkmann, Cloning and expression of flavone synthase IIfrom Gerbera hybrids, Plant J. 20 (1999) 611–618.
] S. Martens, G. Forkmann, U. Matern, R. Lukačin, Cloning of parsleyflavone synthase I, Phytochemistry 58 (2001) 43–46.
] H.-J. Park, H.-C. Cha, Flavonoids from leaves and exocarps of the grapeKyoho, Korean J. Biol. Sci. 7 (2003) 327–330.
] P.A.N. Punyasiri, I.S.B. Abeysinghe, V. Kumar, D. Treutter, D. Duy, C.Gosch, S. Martens, G. Forkmann, T.C. Fischer, Flavonoid biosynthesisin the tea plant Camellia sinensis: properties of enzymes of the promi-nent epicatechin and catechin pathways, Arch. Biochem. Biophys. 431(2004) 22–30.
] S. Roemmelt, T.C. Fischer, H. Halbwirth, S. Peterek, K. Schlangen, J.B.Speakman, D. Treutter, G. Forkmann, K. Stich, Effect of dioxygenaseinhibitors on the resistance-related flavonoid metabolism of apple andpears: chemical, biochemical and molecular biological aspects, Eur. J.Hort. Sci. 68 (2003) 129–136.
] S. Roemmelt, N. Zimmermann, W. Rademacher, D. Treutter, Formationof novel flavonoids in apple (Malus×domestica) treated with the 2-
oxoglutarate-dependent dioxygenase inhibitor prohexadione-Ca, Phyto-chemistry 64 (2003) 709–716.
] A. Scalbert, Antimicrobial properties of tannins, Phytochemistry 30(1991) 3875–3883.
] J.-M. Souquet, B. Labarbe, C.L. Guernevé, V. Cheynier, M. Moutounet,Phenolic composition of grape stems, J. Agric. Food Chem. 48 (2000)1076–1080.
] J.-M. Souquet, V. Cheynier, F. Brossaud, M. Moutounet, Polymericproanthocyanidins from grape skins, Phytochemistry 43 (1996) 509–512.
] H.A. Stafford, H.H. Lester, Flavan-3-ol biosynthesis. The conversion of(+)-dihydromyricetin to its flavan-3,4-Diol (leucodelphinidin) and to (+)-gallocatechin by reductases extracted from tissue cultures ofGinkgo biloba and Pseudotsuga menziesii, Plant Physiol. 78 (1985)791–794.
] H.A. Stafford, H.H. Lester, Flavan-3-ol biosynthesis. The conversion of(+)-dihydroquercetin and flavan-3,4-cis-Diol (leucocyanidin) to (+)-catechin by reductases extracted from cell suspension cultures of Douglasfir, Plant Physiol. 76 (1984) 184–186.
] H.A. Stafford, Enzymic regulation of procyanidin biosynthesis; lack of aflav-3-en-3-ol intermediate, Phytochemistry 22 (1983) 2643–2646.
] G.J. Tanner, K.N. Kristiansen, Synthesis of 3,4-cis-[3H]leucocyanidinand enzymatic reduction to catechin, Anal. Biochem. 209 (1993) 274–277.
] G.J. Tanner, K.T. Francki, S. Abrahams, J.M. Watson, P.J. Larkin, A.R.Ashton, Proanthocyanidin biosynthesis in plants, J. Biol. Chem. 278(2003) 31647–31656.
] D. Treutter, Biosynthesis of phenolic compounds and its regulation inapple, Plant Growth Regul. 34 (2001) 71–89.
] D. Treutter, W. Feucht, The pattern of flavan-3-ols in relation to scabresistance of apple cultivars, J. Hortic. Sci. 65 (1990) 511–518.
] D.-Y. Xie, S.B. Sharma, R.A. Dixon, Anthocyanidin reductases fromMedicago truncatula and Arabidopsis thaliana, Arch. Biochem. Biophys.422 (2004) 91–102.
] D.-Y. Xie, S.B. Sharma, N.L. Paiva, D. Ferreira, R.A. Dixon, Role ofanthocyanidin reductase, encoded by BANYULS in plant flavonoid bio-synthesis, Science 299 (2003) 396–399.
] D.-Y. Xie, R.A. Dixon, Proanthocyanidin biosynthesis—still more ques-tions than answers?, Phytochemistry 66 (2005) 2127–2144.
Related Documents











