Page 1 of 12 Journals of Gerontology: BIOLOGICAL SCIENCES Cite journal as: J Gerontol A Biol Sci Med Sci doi:10.1093/gerona/glu065 © The Author 2014. Published by Oxford University Press on behalf of The Gerontological Society of America. All rights reserved. For permissions, please e-mail: [email protected]. Endothelin-A-Mediated Vasoconstriction During Exercise With Advancing Age Zachary Barrett-O’Keefe, 1,2 Stephen J. Ives, 2,3 Joel D. Trinity, 2,3 Garrett Morgan, 2 Matthew J. Rossman, 1,2 Anthony J. Donato, 1,2,3,4 Sean Runnels, 5 David E. Morgan, 5 Benjamin S. Gmelch, 5 Amber D. Bledsoe, 5 Russell S. Richardson, 1,2,3,4 and D. Walter Wray 1,2,3,4 1 Department of Exercise and Sport Science, University of Utah, Salt Lake City. 2 Geriatric Research, Education, and Clinical Center, Salt Lake City Veterans Affairs Medical Center, Utah. 3 Department of Internal Medicine, University of Utah, Salt Lake City. 4 University of Utah Center on Aging, Salt Lake City. 5 Department of Anesthesiology, University of Utah, Salt Lake City. Address correspondence to D. Walter Wray, PhD, Department of Internal Medicine, Division of Geriatrics, University of Utah George E. Wahlen Department of Veterans Affairs Medical Center, Geriatric Research, Education, and Clinical Center, 500 Foothill Drive, Salt Lake City, UT 84148. Email: [email protected] The endothelin-1 vasoconstrictor pathway contributes to age-related elevations in resting peripheral vascular tone primar- ily through activation of the endothelin subtype A (ET A ) receptor. However, the regulatory influence of ET A -mediated vasoconstriction during exercise in the elderly is unknown. Thus, in 17 healthy volunteers (n = 8 young, 24 ± 2 years; n = 9 old, 70 ± 2 years), we examined leg blood flow, mean arterial pressure, leg arterial–venous oxygen (O 2 ) difference, and leg O 2 consumption (VO 2 ) at rest and during knee-extensor exercise before and after intra-arterial administration of the ET A antagonist BQ-123. During exercise, BQ-123 administration increased leg blood flow to a greater degree in the old (+29 ± 5 mL/min/W) compared with the young (+16 ± 3 mL/min/W). The increase in leg blood flow with BQ-123 was accompanied by an increase in leg VO 2 in both groups, suggesting a reduced efficiency following ET A receptor blockade. Together, these findings have identified an age-related increase in ET A -mediated vasoconstrictor activity that persists during exercise, suggesting an important role of this pathway in the regulation of exercising skeletal muscle blood flow and maintenance of arterial blood pressure in the elderly. Key Words: Endothelin-1—Exercise—Blood flow—Arterial blood pressure—Aging. Received February 20, 2014; Accepted March 31, 2014 Decision Editor: Rafael de Cabo, PhD H EALTHY aging is associated with an elevation in vas- cular tone (1,2), contributing to the attenuated resting skeletal muscle blood flow observed in the elderly (3–8). These age-related changes in vascular tone and the asso- ciated decline in skeletal muscle blood flow persist during exercise (4,8–10). Because adequate muscle perfusion is vital to meet the metabolic demand of the tissue and main- tain normal muscle function at rest and during exercise, a better understanding of the mechanisms that contribute to the regulation of vascular tone in the elderly is essential. The functional consequence of these adaptations are sub- stantial; indeed, impairment in perfusion of the exercising muscle may limit the capacity for physical activity, ulti- mately leading to immobility and an increased risk of car- diovascular disease in the elderly (11). The underlying mechanisms contributing to this age- related increase in vascular tone, and the apparent disparity in hemodynamic regulation between resting and exercis- ing states, has long been a topic of investigation. Although there are a host of pathways that collectively contribute to increased vascular tone in the elderly, changes in the endothelin-1 (ET-1) pathway may be particularly relevant to this age-related adaptation. Indeed, there is evidence from both animal and human studies for an increase in the activity of the ET-1 pathway with advancing age (12–14), which has been linked to the age-associated reduction in resting skeletal muscle blood flow (15,16) via the endothe- lin subtype A (ET A ) receptor pathway (16,17). Despite clear evidence implicating ET-1 as an important contributor to elevated resting vascular tone in the elderly, no studies to date have sought to examine the role of the ET A pathway in the regulation of exercising skeletal mus- cle blood flow with advancing age. Thus, this study evalu- ated the cardiovascular responses to knee-extensor exercise before and after inhibition of the ET A receptor via intra- arterial administration of BQ-123 in the exercising limb. We hypothesized that (a) ET A receptor blockade would enhance resting skeletal muscle blood flow in old, but not in young; (b) ET A receptor blockade during exercise would enhance skeletal muscle blood flow and reduce arterial blood pres- sure to a greater extent in old individuals, compared with their younger counterparts. by guest on April 28, 2016 http://biomedgerontology.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Page 1 of 12
Journals of Gerontology: BIOLOGICAL SCIENCESCite journal as: J Gerontol A Biol Sci Med Scidoi:10.1093/gerona/glu065
© The Author 2014. Published by Oxford University Press on behalf of The Gerontological Society of America. All rights reserved. For permissions, please e-mail: [email protected].
Endothelin-A-Mediated Vasoconstriction During Exercise With Advancing Age
Zachary Barrett-O’Keefe,1,2 Stephen J. Ives,2,3 Joel D. Trinity,2,3 Garrett Morgan,2 Matthew J. Rossman,1,2 Anthony J. Donato,1,2,3,4 Sean Runnels,5 David E. Morgan,5 Benjamin S. Gmelch,5 Amber D. Bledsoe,5
Russell S. Richardson,1,2,3,4 and D. Walter Wray1,2,3,4
1Department of Exercise and Sport Science, University of Utah, Salt Lake City.2Geriatric Research, Education, and Clinical Center, Salt Lake City Veterans Affairs Medical Center, Utah.
3Department of Internal Medicine, University of Utah, Salt Lake City.4University of Utah Center on Aging, Salt Lake City.
5Department of Anesthesiology, University of Utah, Salt Lake City.
Address correspondence to D. Walter Wray, PhD, Department of Internal Medicine, Division of Geriatrics, University of Utah George E. Wahlen Department of Veterans Affairs Medical Center, Geriatric Research, Education, and Clinical Center, 500 Foothill Drive, Salt Lake City, UT 84148.
Email: [email protected]
The endothelin-1 vasoconstrictor pathway contributes to age-related elevations in resting peripheral vascular tone primar-ily through activation of the endothelin subtype A (ET
A) receptor. However, the regulatory influence of ET
A-mediated
vasoconstriction during exercise in the elderly is unknown. Thus, in 17 healthy volunteers (n = 8 young, 24 ± 2 years; n = 9 old, 70 ± 2 years), we examined leg blood flow, mean arterial pressure, leg arterial–venous oxygen (O
2) difference,
and leg O2 consumption (VO
2) at rest and during knee-extensor exercise before and after intra-arterial administration of
the ETA antagonist BQ-123. During exercise, BQ-123 administration increased leg blood flow to a greater degree in the
old (+29 ± 5 mL/min/W) compared with the young (+16 ± 3 mL/min/W). The increase in leg blood flow with BQ-123 was accompanied by an increase in leg VO
2 in both groups, suggesting a reduced efficiency following ET
A receptor blockade.
Together, these findings have identified an age-related increase in ETA-mediated vasoconstrictor activity that persists
during exercise, suggesting an important role of this pathway in the regulation of exercising skeletal muscle blood flow and maintenance of arterial blood pressure in the elderly.
Key Words: Endothelin-1—Exercise—Blood flow—Arterial blood pressure—Aging.
Received February 20, 2014; Accepted March 31, 2014
Decision Editor: Rafael de Cabo, PhD
HEALTHy aging is associated with an elevation in vas-cular tone (1,2), contributing to the attenuated resting
skeletal muscle blood flow observed in the elderly (3–8). These age-related changes in vascular tone and the asso-ciated decline in skeletal muscle blood flow persist during exercise (4,8–10). Because adequate muscle perfusion is vital to meet the metabolic demand of the tissue and main-tain normal muscle function at rest and during exercise, a better understanding of the mechanisms that contribute to the regulation of vascular tone in the elderly is essential. The functional consequence of these adaptations are sub-stantial; indeed, impairment in perfusion of the exercising muscle may limit the capacity for physical activity, ulti-mately leading to immobility and an increased risk of car-diovascular disease in the elderly (11).
The underlying mechanisms contributing to this age-related increase in vascular tone, and the apparent disparity in hemodynamic regulation between resting and exercis-ing states, has long been a topic of investigation. Although there are a host of pathways that collectively contribute to increased vascular tone in the elderly, changes in the
endothelin-1 (ET-1) pathway may be particularly relevant to this age-related adaptation. Indeed, there is evidence from both animal and human studies for an increase in the activity of the ET-1 pathway with advancing age (12–14), which has been linked to the age-associated reduction in resting skeletal muscle blood flow (15,16) via the endothe-lin subtype A (ET
A) receptor pathway (16,17).
Despite clear evidence implicating ET-1 as an important contributor to elevated resting vascular tone in the elderly, no studies to date have sought to examine the role of the ET
A pathway in the regulation of exercising skeletal mus-
cle blood flow with advancing age. Thus, this study evalu-ated the cardiovascular responses to knee-extensor exercise before and after inhibition of the ET
A receptor via intra-
arterial administration of BQ-123 in the exercising limb. We hypothesized that (a) ET
A receptor blockade would enhance
resting skeletal muscle blood flow in old, but not in young; (b) ET
A receptor blockade during exercise would enhance
skeletal muscle blood flow and reduce arterial blood pres-sure to a greater extent in old individuals, compared with their younger counterparts.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 2 of 12 BARREtt-O’KEEFE Et AL.
Methods
ParticipantsEight young (24 ± 2 yrs) and nine old (70 ± 2 yrs) healthy
participants were enrolled in this study. A subset of the data from the young participants have been reported previously (18) and are presented for the purpose of comparison in this study. All participants were nonsmokers and were nor-mally active. Participants were not taking any prescription medication and were free of overt cardiovascular disease, as indicated by a health history. Protocol approval and written informed consent were obtained according to the University of Utah and the Salt Lake City Veterans Affairs Medical Center Institutional Review Board requirements. All data collection took place at the Veterans Affairs Salt Lake City Geriatric, Research, Education, and Clinical Center in the Utah Vascular Research Laboratory.
ProtocolsParticipants reported to the Utah Vascular Research
Laboratory at 0800 on the experimental day. After 30 min-utes of supine rest, two catheters (common femoral artery [CFA] and femoral vein) were placed using sterile technique, as described previously (19–21). After catheter placement, participants rested for about 30 minutes and then underwent the protocol as outlined in Figure 1. Participants reported to the lab in the fasted state and were given a small, standard-ized meal (1/2 cup of corn flakes and 1/2 cup of skim milk) 10 minutes prior to the start of the control (saline) and the ET
A receptor blockade (BQ-123) trials (Figure 1). All data
collection took place with participants in a semirecumbent position (60° reclined), and all studies were performed in a thermoneutral environment.
Due to the long-lasting effects of BQ-123, the drug administration portion of the protocol always occurred after the saline portion of the protocol. Thus, to determine whether the observed changes in leg blood flow and mean arterial pressure (MAP) were the consequence of an exer-cise ordering effect, rather than the effect of the drug per se, participants returned to the laboratory on a separate day to undergo an exercise time control study. For this visit, catheters were not placed, and no drugs were administered. Apart from these differences, the timeline for this time control study was identical to the drug infusion day, with the addition of leg blood flow measurements immediately before and after a light meal to examine possible postpran-dial hemodynamic effects.
Study DrugsThigh volumes were determined anthropometrically and
then used for the calculation of drug dosing. A selective ET
A receptor antagonist (BQ-123 Clinalfa, Calbiochem-
Novabiochem, Läufelfingen, Switzerland) was prepared in normal saline (0.9% NaCl) and administered intra-arte-rially (CFA) at 10 nmol/min/L of thigh volume (infusion rates 0.8–1.5 mL/min). This dose has been documented to induce an apparent plateau in vasodilation in both the forearm (22,23) and quadriceps (15), without affecting MAP. BQ-123 has been documented to have a high affin-ity for the ET
A receptor (24) and to effectively counteract
Figure 1. Experimental protocol. Arrows indicate points at which leg blood flow was recorded and arterial and venous blood samples were obtained.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Et-1 AND ExERCISE IN thE ELDERLy Page 3 of 12
the vasoconstrictor effect of ET-1 infusion in the human forearm (25). During the control trial, normal saline (0.9% NaCl) was administered intra-arterially into the CFA at the same infusion rates as BQ-123.
Exercise ModelThe knee-extensor paradigm implemented in this study
has been described previously (4,26–28). Briefly, partici-pants were seated in a semirecumbent position on an adjust-able chair with a cycle ergometer (model 828E; Monark Exercise AB, Vansbro, Sweden) positioned behind them. Resistance was created by applying friction to the flywheel, which was turned by the participant via a metal bar con-nected to the crank arm of the ergometer and a metal boot in which the participant’s foot was placed. Participants exer-cised for 3 minutes at 0 (ie, unloaded), 5, 10, and 15 W, as well as 20%, 40%, 60%, and 80% of their predetermined maximal work rate (WR
max) while maintaining 60 contrac-
tions per minute. A 3-minute recovery period was given following three successive exercise bouts (Figure 1). Both absolute and relative work rates were incorporated into the protocol to address the dichotomy that limb blood flow is regulated predominantly by absolute work performed (29), whereas exercise-induced changes in MAP are dependent upon relative exercise intensity (10,30).
Measurements
Ultrasound Doppler assessments.—Measurements of CFA blood velocity and vessel diameter were performed in the infused leg using a Logiq 7 ultrasound Doppler sys-tem (GE Medical Systems, Milwaukee, WI) operating in duplex mode. The Logic 7 was equipped with a linear array transducer operating at an imaging frequency of 14 MHz. The CFA was insonated 2–3 cm proximal to the bifurca-tion of the CFA into the superficial and deep branches. The blood velocity profile was obtained using the same transducer with a Doppler frequency of 5 MHz, operated in the high-pulsed repetition frequency mode (2–25 kHz). Care was taken to avoid aliasing the blood velocity spectra by using scale adjustments, especially during exercise. All blood velocity measurements were obtained with the probe appropriately positioned to maintain an insonation angle of 60° or less (31). The sample volume was maximized according to vessel size and was centered within the vessel on the basis of real-time ultrasound visualization. At all sample points, arterial diameter (cm) and angle-corrected, time-averaged, and intensity-weighted mean blood veloc-ity (V
mean) values were calculated using commercially avail-
able software (Logic 7). Using measured arterial diameter and V
mean, leg blood flow was calculated according to the
equation: leg blood flow (mL/min) = (Vmean
× π (vessel diameter/2)2 × 60).
MAP, vascular conductance, and heart rate assess-ment.—Arterial blood pressure measurements were col-lected continuously from the indwelling catheter placed in the CFA with the pressure transducer placed at the level of the catheter (Transpac IV, Abbott Laboratories). MAP (mmHg) was calculated as diastolic arterial pressure + (arterial pulse pressure × 0.33). On the time control study day, MAP was determined noninvasively using finger pho-toplethysmography (Finometer, Finapres Medical Systems BV, Amsterdam, The Netherlands). Leg vascular conduct-ance was calculated as leg blood flow/MAP. Heart rate was monitored from a standard three-lead electrocardio-gram recorded in duplicate on the data acquisition device (Biopac, Goleta, CA) and the Logic 7.
Blood chemistry.—A lipid panel was obtained for all par-ticipants by standard techniques. In the last 30 seconds of each exercise intensity, femoral arterial and venous blood samples (3–4 mL) were collected. One milliliter of arterial and venous blood were presented anaerobically to a GEM 4000 blood–gas analyzer and co-oximeter (Instrumentation Laboratories, Bedford, MA) to obtain arterial and venous total hemoglobin (tHb) oxyhemoglobin saturation (SO
2),
partial pressure of oxygen (PO2), hematocrit (hct), lac-
tate, and pH. Arterial and venous blood oxygen content (CaO
2 and CvO
2; mL/dL) were calculated as the following:
Blood oxygen content (mL/dL) = 1.39 (tHb) × (SO2/100) +
0.003 × PO2. Leg oxygen consumption (VO
2; mL/min) was
calculated as the following: VO2 (mL/min) = (C
AO
2 − C
VO
2)
× leg blood flow. The remaining blood was centrifuged, and plasma stored at −80°C until analysis.
Plasma ET-1 concentrations were evaluated via quanti-tative enzyme immunoassay (R&D Systems, Minneapolis, MN), with a sensitivity of less than 1 pg/mL. Intra-assay and interassay coefficient of variation were 4.5% and 5.5%, respectively. Using arterial (C
A) and venous (C
V) plasma
ET-1 or lactate concentrations, with corrections for leg blood flow, “net ET-1 release” and “net lactate release” were calculated (18,32) according to the following equation: Net ET-1 (ng/min) or lactate release (mmol/min) = (C
V − C
A) ×
(leg blood flow × ((101 − (hct/100))).
Data Analysis and StatisticsBQ-123-induced changes in leg blood flow, MAP, and
leg vascular conductance were calculated as the difference between the BQ-123 trial value and the saline trial value. Statistics were performed with the use of commercially available software (SigmaStat 3.10; Systat Software, Point Richmond, CA). Sample size calculations were performed using the estimated effect size and standard deviation for BQ-induced changes in leg blood flow and leg vascular conductance. For these outcome measures, it was estimated that six participants per group were needed for a desired power of 0.8 (α = 0.05). A 2 × 9 repeated measure analysis
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 4 of 12 BARREtt-O’KEEFE Et AL.
of variance was used to identify significant changes in measured variables within and between drug groups and across exercise intensities. When a significant main effect was found (α = 0.05), the Holm-Sidak method was used for alpha adjustment and post hoc analysis. To further explore the relationship between absolute work rate and the hemodynamic effects of ET-1 receptor blockade at higher exercise intensities, we also evaluated responses at 80% of maximal work rate in those participants whose maximal work rate was greater than 15 W (n = 8, young; n = 5, old). Linear regression analysis was performed on individual data, with the slope and y-intercept determined to evaluate the increase in leg blood flow, MAP, and leg vascular con-ductance in response to BQ-123 infusion across work rates. A paired t test was used to compare the effect of BQ-123-induced changes on slope and y-intercept values. All group data are expressed as mean ± SE.
Results
Participant CharacteristicsParticipant characteristics are presented in Table 1. All
participants were normotensive, with lab values within normal range.
Resting ResponsesPrior to BQ-123 administration, resting leg blood flow
was significantly lower in the old compared with the young (Figure 2). In both young and old groups, the continuous administration of the ET
A receptor antagonist BQ-123
(10 nmol/min/L of thigh volume; infusion rate: 0.8–1.5 mL/min) did not significantly change heart rate (Δ −2 ± 3 bpm, young; Δ −4 ± 2 bpm, old) or MAP (Δ −1 ± 3 mmHg, young; Δ −2 ± 2 mmHg, old) after 45 minutes of infusion. However, BQ-123 did provoke a significant increase in leg blood flow (Figure 2) and leg vascular conductance in the old
(Δ + 0.9 ± 0.3 mL/min/mmHg) but not in the young (Figure 2; Δ 0.0 ± 0.4 mL/min/mmHg). This observed BQ-123-mediated increase leg blood flow effectively restored leg blood flow in the old to that of the young (Figure 2).
Exercise ResponsesA subset of the exercise data investigating the contribution
of the ETA receptor in the regulation of vascular tone in the
young during knee-extensor exercise of absolute work rates has been previously reported (18). This study extends these previously published findings through inclusion of absolute and relative exercise intensities, as well as quantifying the association between work rate and BQ-123-Induced changes in leg blood flow, MAP, and leg vascular conductance.
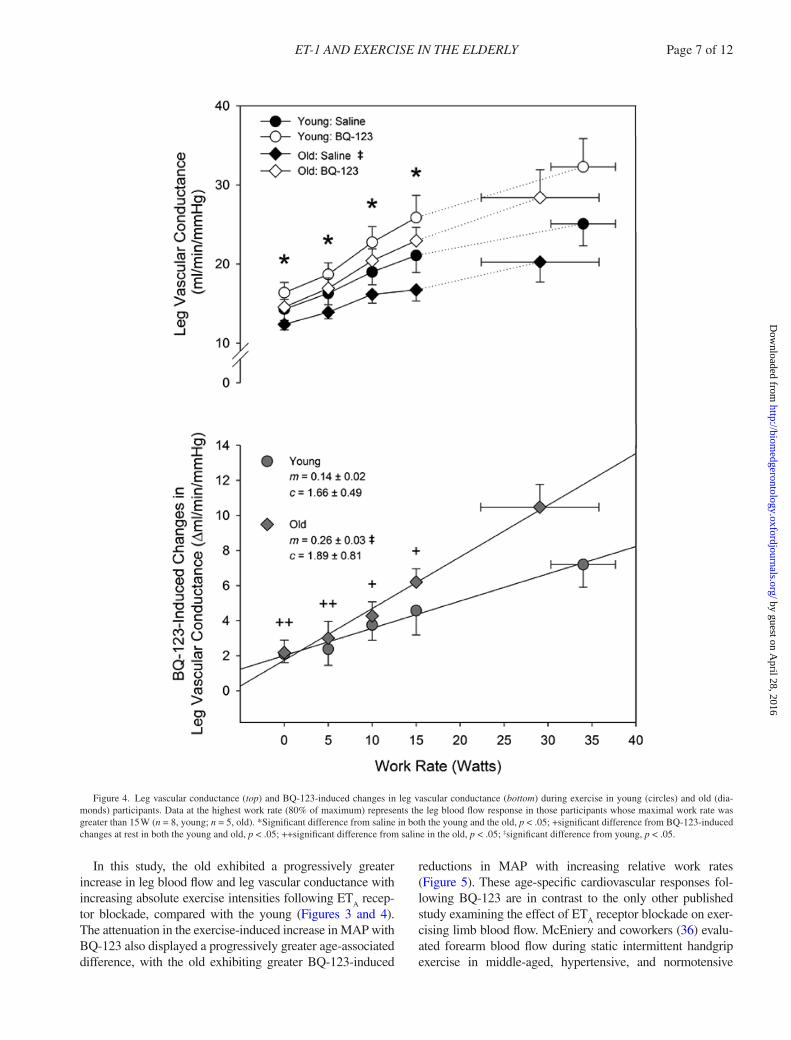
A significant difference in delta leg blood flow was observed between young and old (p < .001). The slope of the relationship between BQ-123-induced increases in leg blood flow and work rate was significantly greater in the old compared with the young (+29 ± 3 and +16 ± 3 mL/min/W, respectively, p = .037; Figure 3, bottom). A significant dif-ference in delta leg vascular conductance was also observed between young and old (p < .001). The slope of the rela-tionship between BQ-123-induced increases in leg vascular conductance and work rate was significantly greater in the old compared with the young (+0.14 ± 0.02 vs +0.26 ± 0.03 mL/min/mmHg/W, respectively, p = .012; Figure 4, bot-tom). Delta MAP was similar between groups (p = .605), with the slope of the relationship between the BQ-123-induced reductions in MAP and work rate significantly greater in the old compared with the young (−0.16 ± 0.02 vs −0.08 ± 0.02 mmHg/W, respectively, p = .032; Figure 5, bottom). In both the young and the old, the vasodilation pro-voked by BQ-123 during exercise contributed to an increase in leg VO
2 (Tables 2 and 3). No significant changes in net
lactate release or pH were observed during BQ-123 infu-sion compared with the saline trial (Tables 2 and 3).
Exercise time Control StudyFifteen of the original 17 participants (n = 8 young; n = 7
old) who participated in the drug infusion protocol returned to the lab for an exercise time control protocol. There was no significant difference in leg blood flow or MAP between the two exercise bouts (Figure 6). Using this time con-trol study and the one exercise bout (saline trial) from the catheter-based study day, we identified a within-participant coefficient of variation for leg blood flow of 3%–4% across work rates in both young and old groups.
DiscussionThis study has identified a functional role of the ET
A
vasoconstrictor pathway in the regulation of vascular tone with advancing age. At rest, ET
A blockade reversed the
age-associated reduction in leg blood flow, confirming a
Table 1. Participant Characteristics
young Old
Age, y 24 ± 2 70 ± 2*Height, cm 172 ± 2 172 ± 3Weight, kg 71 ± 6 75 ± 3Body mass index, kg/m2 24 ± 5 25 ± 1Systolic blood pressure, mmHg 107 ± 5 116 ± 3Diastolic blood pressure, mmHg 73 ± 3 78 ± 2Quadriceps muscle mass, kg 2.0 ± 0.1 2.1 ± 0.2Maximum knee-extensor, W 42 ± 4 29 ± 4‡Glucose, mg/dL 68 ± 6 67 ± 2Total cholesterol, mg/dL 153 ± 10 173 ± 8Triglycerides, mg/dL 69 ± 12 96 ± 14HDL, mg/dL 60 ± 9 55 ± 8LDL, mg/dL 78 ± 9 83 ± 10
Notes: HDL = high-density lipoprotein; LDL = low-density lipoprotein. Data are expressed as means ± SE.
*Significantly different than young, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Et-1 AND ExERCISE IN thE ELDERLy Page 5 of 12
significant contribution of the ETA receptor to the elevation
in resting vascular tone in the elderly. During exercise, ETA
receptor inhibition augmented leg blood flow to a greater degree in the old compared with young, demonstrating an age-associated increase in ET
A-mediated restraint of exer-
cising skeletal muscle blood flow. ETA blockade also attenu-
ated the exercise-induced increase in arterial blood pressure to a greater degree in the old compared with their younger counterparts. These hemodynamic changes were accompa-nied by increases in leg VO
2 in both groups, suggesting a
decrease in intramuscular efficiency in the presence of ETA
blockade. Taken together, these findings have identified an age-related increase in resting ET
A vasoconstrictor activity
that persists during exercise, suggesting an important role of this pathway in the regulation of skeletal muscle blood flow and arterial blood pressure during physical activity with advancing age.
EtA Receptor–Mediated Restraint of Skeletal Muscle
Blood Flow at RestA number of studies from our group (7,29) and others
(1,3,33,34) have collectively demonstrated that healthy aging is associated with a reduction in resting limb blood flow. Although there are many structural, autonomic, and neuroendocrine factors responsible for the decline in blood flow, findings from this study implicate the ET-1 pathway as a significant contributor to this age-related change. As presented in Figure 2, we observed an approximately 30% reduction in resting leg blood flow in the old compared with their younger counterparts. Administration of BQ-123 did not affect leg blood flow in the young group, but in the elderly, it resulted in a progressive increase in leg blood flow that ultimately restored resting leg blood flow to a level that was similar to the young (Figure 2). Interestingly, prior to BQ-123 administration, net ET-1 release was
Figure 2. Resting leg blood flow expressed as percent change (top) and absolute units (bottom) during continuous infusion of BQ-123 in the young (circles) and old (diamonds). ‡Significant difference from young, p < .05; ##significant difference from minute 0 in the old, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 6 of 12 BARREtt-O’KEEFE Et AL.
similar between groups (Tables 2 and 3). However, fol-lowing ET
A receptor blockade, circulating ET-1 increased
almost twofold in the old but remained unchanged in the young (Tables 2 and 3). This suggests that present find-ings build upon recent studies reporting a significantly greater contribution of the ET
A receptor to vascular tone in
the old compared with the young at rest (15,16), but with the important addition of arterial and venous ET-1 levels, data that provide further insight concerning the relation-ship between circulating ET-1 and hemodynamic measure-ments. Together, these former and present findings present an aging vascular phenotype characterized by an increase in the endogenous contribution of ET-1 to vascular tone, accompanied by an overall increase in bound ET-1 and/or enhanced ET-1 bioavailability in old individuals.
EtA Receptor–Mediated Restraint of Skeletal Muscle
Blood Flow during ExerciseFrom rest to exercise, perfusion of the exercising skel-
etal muscle increases dramatically in order to match the metabolic demand of the active muscle, achieved through the vasoconstriction of less metabolically active tissue and the modulation of vasodilator and vasoconstrictor influ-ences within the exercising muscle vasculature (35). With advancing age, there is an apparent shift in favor of greater vasoconstriction during exercise, resulting in attenuation of exercise hyperemia (4,8–10). Based on these previous findings and on the known role of ET
A inhibition to modu-
late vascular tone in the elderly at rest (Figure 2) (15,16), the ET-1 pathway seems a likely contributor to blood flow regulation during exercise in this cohort.
Figure 3. Leg blood flow (top) and BQ-123-induced changes in leg blood flow (bottom) during exercise in young (circles) and old (diamonds) participants. Data at the highest work rate (80% of maximum) represents the leg blood flow response in those participants whose maximal work rate was greater than 15 W (n = 8, young; n = 5, old). *Significant difference from saline in both the young and the old, p < .05; **significant difference from saline in the old, p < .05; +significant difference from BQ-123-induced changes at rest in both the young and old, p < .05; ++significant difference from saline in the old, p < .05; ‡significant difference from young, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
Et-1 AND ExERCISE IN thE ELDERLy Page 7 of 12
In this study, the old exhibited a progressively greater increase in leg blood flow and leg vascular conductance with increasing absolute exercise intensities following ET
A recep-
tor blockade, compared with the young (Figures 3 and 4). The attenuation in the exercise-induced increase in MAP with BQ-123 also displayed a progressively greater age-associated difference, with the old exhibiting greater BQ-123-induced
reductions in MAP with increasing relative work rates (Figure 5). These age-specific cardiovascular responses fol-lowing BQ-123 are in contrast to the only other published study examining the effect of ET
A receptor blockade on exer-
cising limb blood flow. McEniery and coworkers (36) evalu-ated forearm blood flow during static intermittent handgrip exercise in middle-aged, hypertensive, and normotensive
Figure 4. Leg vascular conductance (top) and BQ-123-induced changes in leg vascular conductance (bottom) during exercise in young (circles) and old (dia-monds) participants. Data at the highest work rate (80% of maximum) represents the leg blood flow response in those participants whose maximal work rate was greater than 15 W (n = 8, young; n = 5, old). *Significant difference from saline in both the young and the old, p < .05; +significant difference from BQ-123-induced changes at rest in both the young and old, p < .05; ++significant difference from saline in the old, p < .05; ‡significant difference from young, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 8 of 12 BARREtt-O’KEEFE Et AL.
individuals before and after intra-arterial (brachial) BQ-123 administration and reported no difference between the blocked and control trials in the normotensive group, suggesting the ET
A receptor did not contribute significantly to the modulation
of vascular tone during handgrip exercise. However, although this previous study included a wide age range (30–50 years), it was not designed to examine age-related changes in limb blood flow in the response to BQ-123. Thus, the present find-ings substantially extend this earlier work, clearly identifying the ET
A pathway as an important component in age-associated
changes in the cardiovascular response to dynamic exercise.
Improved Exercising Muscle Blood Flow and Intramuscular Efficiency
The increase in leg blood flow following ETA recep-
tor blockade was accompanied by a comparable decline
in intramuscular efficiency during exercise in both groups, as evident by greater leg VO
2 for a similar amount of work
(Tables 2 and 3). These data indicate that in both young and old, the maintenance of vascular tone not only is vital in the support of MAP but also might play a role in limiting the perfusion of the exercising muscle during submaximal work in order to prevent an exaggerated O
2 delivery in excess of
metabolic demand. This increase in leg VO2 induced by ET
A
receptor blockade occurred in the absence of an alteration in net lactate release (Tables 2 and 3), further suggesting that the increase in leg blood flow following BQ-123 did not appear to confer beneficial effect on muscle metabolism. Thus, it appears that the ET
A pathway may provide an impor-
tant governing influence on exercising limb blood flow in the elderly in order to optimize the matching of blood flow with metabolic demand in the exercising skeletal muscle.
Figure 5. Mean arterial pressure (top) and BQ-123-induced changes in mean arterial pressure (bottom) during exercise of relative intensity work rates in young (circles) and old (diamonds). *Significant difference from saline in both the young and the old, p < .05; +significant difference from BQ-123-induced changes at rest in both the young and old, p < .05; ‡significant difference from young, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
Et-1 AND ExERCISE IN thE ELDERLy Page 9 of 12
PerspectivesThis study has identified a functional role of the ET
A
vasoconstrictor pathway in the regulation of vascular tone during exercise in older, healthy adults. To our knowledge, this is the first study to perform ET
A receptor inhibition dur-
ing exercise in the elderly and to report a greater augmenta-tion in leg blood flow in the old compared with young in response to ET
A blockade. These findings demonstrating
an age-associated increase in ETA-mediated restraint of
exercising skeletal muscle blood flow add significantly to our understanding of how this pathway contributes to the sequelae of vascular dysfunction in the elderly. The high prevalence of exercise intolerance in the elderly (37), cou-pled with the recognition of inactivity as a risk factor in the development of cardiovascular disease (38), makes findings from this study highly relevant to the growing body of litera-ture linking excessive vascular tone and impaired capacity for physical activity in older, healthy adults.
Although there are several factors that may modulate ET
A-mediated vasoconstriction during exercise in the
elderly, one noteworthy candidate is the vasodilator nitric oxide (NO). Indeed, it is now well established that NO may limit the production and action of ET-1 (39,40), a reciprocal relationship that may be particularly relevant to age-related changes in the regulation of exercising skeletal muscle blood flow. Although there is some ambiguity in the litera-ture regarding the importance of NO release on exercising muscle blood flow (41–44), recent work from our group identified a marked reduction in forearm blood flow during handgrip exercise following intra-arterial (brachial) admin-istration of the nitric oxide synthase inhibitor L-NMMA (21). This finding in young, healthy humans suggests that the NO pathway contributes significantly to hyperemia of the exercising skeletal muscle and thus may play an important role in opposition to vasoconstrictor pathways such as ET-1. Thus, considering the age-related decline in
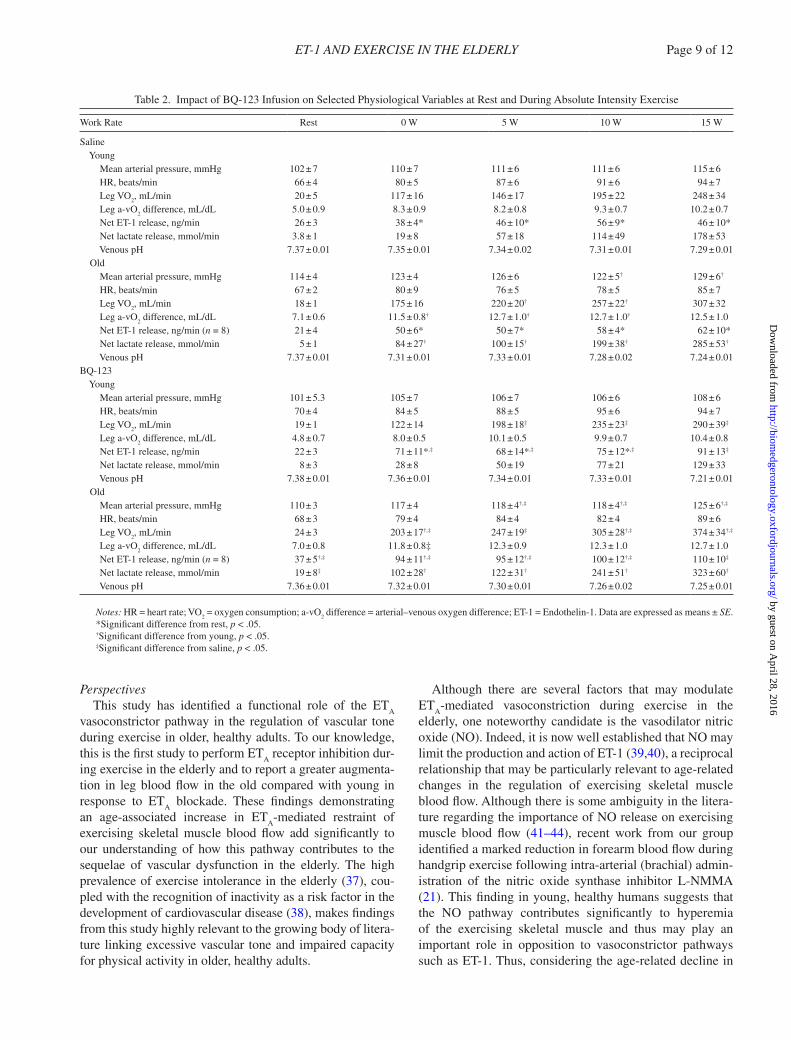
Table 2. Impact of BQ-123 Infusion on Selected Physiological Variables at Rest and During Absolute Intensity Exercise
Work Rate Rest 0 W 5 W 10 W 15 W
Saline young Mean arterial pressure, mmHg 102 ± 7 110 ± 7 111 ± 6 111 ± 6 115 ± 6 HR, beats/min 66 ± 4 80 ± 5 87 ± 6 91 ± 6 94 ± 7 Leg VO
2, mL/min 20 ± 5 117 ± 16 146 ± 17 195 ± 22 248 ± 34
Leg a-vO2 difference, mL/dL 5.0 ± 0.9 8.3 ± 0.9 8.2 ± 0.8 9.3 ± 0.7 10.2 ± 0.7
Net ET-1 release, ng/min 26 ± 3 38 ± 4* 46 ± 10* 56 ± 9* 46 ± 10* Net lactate release, mmol/min 3.8 ± 1 19 ± 8 57 ± 18 114 ± 49 178 ± 53 Venous pH 7.37 ± 0.01 7.35 ± 0.01 7.34 ± 0.02 7.31 ± 0.01 7.29 ± 0.01 Old Mean arterial pressure, mmHg 114 ± 4 123 ± 4 126 ± 6 122 ± 5† 129 ± 6†
HR, beats/min 67 ± 2 80 ± 9 76 ± 5 78 ± 5 85 ± 7 Leg VO
2, mL/min 18 ± 1 175 ± 16 220 ± 20† 257 ± 22† 307 ± 32
Leg a-vO2 difference, mL/dL 7.1 ± 0.6 11.5 ± 0.8† 12.7 ± 1.0† 12.7 ± 1.0† 12.5 ± 1.0
Net ET-1 release, ng/min (n = 8) 21 ± 4 50 ± 6* 50 ± 7* 58 ± 4* 62 ± 10* Net lactate release, mmol/min 5 ± 1 84 ± 27† 100 ± 15† 199 ± 38† 285 ± 53†
Venous pH 7.37 ± 0.01 7.31 ± 0.01 7.33 ± 0.01 7.28 ± 0.02 7.24 ± 0.01BQ-123 young Mean arterial pressure, mmHg 101 ± 5.3 105 ± 7 106 ± 7 106 ± 6 108 ± 6 HR, beats/min 70 ± 4 84 ± 5 88 ± 5 95 ± 6 94 ± 7 Leg VO
2, mL/min 19 ± 1 122 ± 14 198 ± 18‡ 235 ± 23‡ 290 ± 39‡
Leg a-vO2 difference, mL/dL 4.8 ± 0.7 8.0 ± 0.5 10.1 ± 0.5 9.9 ± 0.7 10.4 ± 0.8
Net ET-1 release, ng/min 22 ± 3 71 ± 11*,‡ 68 ± 14*,‡ 75 ± 12*,‡ 91 ± 13‡
Net lactate release, mmol/min 8 ± 3 28 ± 8 50 ± 19 77 ± 21 129 ± 33 Venous pH 7.38 ± 0.01 7.36 ± 0.01 7.34 ± 0.01 7.33 ± 0.01 7.21 ± 0.01 Old Mean arterial pressure, mmHg 110 ± 3 117 ± 4 118 ± 4†,‡ 118 ± 4†,‡ 125 ± 6†,‡
HR, beats/min 68 ± 3 79 ± 4 84 ± 4 82 ± 4 89 ± 6 Leg VO
2, mL/min 24 ± 3 203 ± 17†,‡ 247 ± 19‡ 305 ± 28†,‡ 374 ± 34†,‡
Leg a-vO2 difference, mL/dL 7.0 ± 0.8 11.8 ± 0.8‡ 12.3 ± 0.9 12.3 ± 1.0 12.7 ± 1.0
Net ET-1 release, ng/min (n = 8) 37 ± 5†,‡ 94 ± 11†,‡ 95 ± 12†,‡ 100 ± 12†,‡ 110 ± 10‡
Net lactate release, mmol/min 19 ± 8‡ 102 ± 28† 122 ± 31† 241 ± 51† 323 ± 60†
Venous pH 7.36 ± 0.01 7.32 ± 0.01 7.30 ± 0.01 7.26 ± 0.02 7.25 ± 0.01
Notes: HR = heart rate; VO2 = oxygen consumption; a-vO
2 difference = arterial–venous oxygen difference; ET-1 = Endothelin-1. Data are expressed as means ± SE.
*Significant difference from rest, p < .05.†Significant difference from young, p < .05.‡Significant difference from saline, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 10 of 12 BARREtt-O’KEEFE Et AL.
NO-mediated vasodilation present at rest (45) and during exercise (46), it is tempting to speculate that a reduction in NO bioavailability in the elderly may be partially responsi-ble for the exaggerated increase in leg blood flow following ET
A receptor inhibition (Figure 3). However, further studies
utilizing both NO and ETA antagonism are required to fur-
ther explore this interesting possibility.
Experimental ConsiderationsDue to the slow acting kinetics of BQ-123 binding and
clearance, the study was ordered so the control (saline) trial always preceded the BQ-123 trial. However, a time control trial on a separate day demonstrated comparable measure-ments of leg blood flow and MAP during repeated bouts of knee-extensor exercise (Figure 6), providing confidence in
the comparison of the control and the BQ-123 trials. We cannot exclude the possibility that some of the observed changes in peripheral hemodynamics are due to ET-1 sub-type B (ET
B) receptor activation, as BQ-123 is highly selec-
tive for ETA receptors. A recent study utilizing combined
ETA and ET
B receptor inhibition in the arm vasculature have
failed to identify a significant role for ETB receptors in the
regulation of vascular tone in humans (16). However, with the recognition that significant limb specificity exists in the regulation of vascular tone (47), additional studies utilizing both ET
A and ET
B inhibitions are required to fully charac-
terize the role of ET-1 during lower limb exercise in the elderly. Finally, although all participants were identified as normally active, we acknowledge that differences in aerobic fitness between groups may have influenced the observed exercising leg blood flow responses.
Table 3. Impact of BQ-123 Infusion on Selected Physiological Variables at Rest and During Relative Intensity Exercise
Work Rate Rest 20% 40% 60% 80%
Saline young Leg blood flow, mL/min 382 ± 38 1949 ± 131 2454 ± 176 2882 ± 245 3220 ± 275 Leg vascular conductance, mL/min/mmHg 4.0 ± 0.5 17.4 ± 2.0 22.1 ± 2 24.5 ± 2.9 25.1 ± 2.8 HR, beats/min 66 ± 4 88 ± 5 94 ± 5 101 ± 8 112 ± 6 Leg VO
2, mL/min 20 ± 5 174 ± 17 264 ± 41 346 ± 43 373 ± 39
Leg a-vO2 difference, mL/dL 5.0 ± 0.9 8.7 ± 0.7 10.2 ± 0.8 11.6 ± 0.5 11.2 ± 0.4
Net ET-1 release, ng/min 26 ± 3 58 ± 13* 66 ± 17* 68 ± 20* 63 ± 15* Net lactate release, mmol/min 4 ± 1 74 ± 25 195 ± 45 354 ± 54 698 ± 138 Venous pH 7.37 ± 0.01 7.31 ± 0.01 7.28 ± 0.01 7.25 ± 0.01 7.20 ± 0.02 Old Leg blood flow, mL/min 257 ± 13† 1847 ± 123 2253 ± 208 2683 ± 241 2963 ± 385 Leg vascular conductance, mL/min/mmHg 2.3 ± 0.1† 15.0 ± 1.2 17.8 ± 1.6 19.5 ± 1.7 20.2 ± 2.5 HR, beats/min 67 ± 3 77 ± 6† 80 ± 6† 89 ± 10† 98 ± 15 Leg VO
2, mL/min 18 ± 2 230.7 ± 22 287 ± 36 341 ± 48 399 ± 59
Leg a-vO2 difference, mL/dL 7.1 ± 0.6 12.7 ± 1.0 12.7 ± 1.1 12.7 ± 1.1 12.5 ± 1.2
Net ET-1 release, ng/min (n = 8) 21 ± 4 51 ± 4* 58 ± 9* 69 ± 4* 77 ± 13* Net lactate release, mmol/min 5 ± 1 173 ± 47 216 ± 62 437 ± 76 629 ± 136 Venous pH 7.37 ± 0.01 7.31 ± 0.02 7.22 ± 0.01 7.21 ± 0.01 7.24 ± 0.23BQ-123 young Leg blood flow, mL/min 382 ± 31 2262 ± 139‡ 2863 ± 258‡ 3416 ± 314‡ 3874 ± 355‡
Leg vascular conductance, mL/min/mmHg 3.9 ± 0.4 20.1 ± 1.7 27.8 ± 2.9‡ 30.9 ± 3.9‡ 32.3 ± 3.6‡
HR, beats/min 70 ± 4 93 ± 5 97 ± 5 106 ± 9 116 ± 7 Leg VO
2, mL/min 19 ± 1 277 ± 24‡ 315 ± 51‡ 348 ± 51‡ 381 ± 48‡
Leg a-vO2 difference, mL/dL 4.8 ± 0.7 10.1 ± 0.6 10.2 ± 0.8 11.1 ± 0.4 11.0 ± 0.5
Net ET-1 release, ng/min 22 ± 3 72 ± 13*,‡ 88 ± 24*,‡ 92 ± 20*,‡ 92 ± 10*,‡
Net lactate release, mmol/min 8 ± 3 74 ± 20 136 ± 24 406 ± 86 707 ± 153 Venous pH 7.38 ± 0.01 7.32 ± 0.01 7.32 ± 0.01 7.27 ± 0.01 7.25 ± 0.02 Old Leg blood flow, mL/min 375 ± 30‡ 2247 ± 141‡ 2593.29 ± 176‡ 3002.95 ± 221‡ 3815 ± 478‡
Leg vascular conductance, mL/min/mmHg 3.4 ± 0.4‡ 18.7 ± 1.4‡ 21.3 ± 1.6†,‡ 23.7 ± 1.8†,‡ 28.4 ± 3.6‡
HR, beats/min 68 ± 3 86 ± 7 86 ± 5† 89 ± 5† 97 ± 9†
Leg VO2, mL/min 24 ± 3 269 ± 28‡ 319 ± 32‡ 386 ± 52‡ 478 ± 70‡
Leg a-vO2 difference, mL/dL 7.0 ± 0.9 12 ± 1.0 12.4 ± 1.0 12.7 ± 1.2 13.0 ± 1.3
Net ET-1 release, ng/min (n = 8) 37 ± 5†,‡ 99 ± 10*,†,‡ 95 ± 8*,‡ 105 ± 11*,‡ 115 ± 14*,‡
Net lactate release, mmol/min 9 ± 20†,‡ 155 ± 47 295 ± 54 502 ± 82 562 ± 95 Venous pH 7.36 ± 0.01 7.29 ± 0.01 7.28 ± 0.02 7.24 ± 0.02 7.23 ± 0.01
Notes: HR = heart rate; VO2 = oxygen consumption; a-vO
2 difference = arterial–venous oxygen difference; ET-1 = Endothelin-1. Data are expressed as means ± SE.
*Significant difference from rest, p < .05.†Significant difference from young, p < .05.‡Significant difference from saline, p < .05.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Et-1 AND ExERCISE IN thE ELDERLy Page 11 of 12
Funding
This work was supported by the National Institute of Health (PO1 HL-091830, R01 HL118313-01), the United States Veterans Administration (RR&D E6910R), and the American Heart Association (0835209N).
Conflict of Interest
None.
References 1. Dinenno FA, Tanaka H, Stauffer BL, Seals DR. Reductions in basal
limb blood flow and vascular conductance with human ageing: role for augmented alpha-adrenergic vasoconstriction. J Physiol. 2001;536(Pt 3):977–983.
2. Sundlöf G, Wallin BG. Human muscle nerve sympathetic activity at rest. Relationship to blood pressure and age. J Physiol. 1978;274:621–637.
3. Dinenno FA, Jones PP, Seals DR, Tanaka H. Limb blood flow and vascular conductance are reduced with age in healthy humans: rela-tion to elevations in sympathetic nerve activity and declines in oxygen demand. Circulation. 1999;100:164–170.
4. Lawrenson L, Poole JG, Kim J, Brown C, Patel P, Richardson RS. Vascular and metabolic response to isolated small muscle mass exercise:
effect of age. Am J Physiol heart Circ Physiol. 2003;285:H1023–H1031. doi:10.1152/ajpheart.00135.2003
5. Lawrenson L, Hoff J, Richardson RS. Aging attenuates vascular and metabolic plasticity but does not limit improvement in muscle VO(2) max. Am J Physiol heart Circ Physiol. 2004;286:H1565–H1572.
6. Parker BA, Smithmyer SL, Pelberg JA, Mishkin AD, Proctor DN. Sex-specific influence of aging on exercising leg blood flow. J Appl Physiol. 2008;104:655–664. doi:10.1152/japplphysiol.01150.2007
7. Wray DW, Nishiyama SK, Harris RA, Richardson RS. Angiotensin II in the elderly: impact of angiotensin II type 1 receptor sensitivity on peripheral hemodynamics. hypertension. 2008;51:1611–1616. doi:HyPERTENSIONAHA.108.111294 [pii]
8. Poole JG, Lawrenson L, Kim J, Brown C, Richardson RS. Vascular and metabolic response to cycle exercise in sedentary humans: effect of age. Am J Physiol heart Circ Physiol. 2003;284:H1251–H1259. doi:10.1152/ajpheart.00790.2002
9. Beere PA, Russell SD, Morey MC, Kitzman DW, Higginbotham MB. Aerobic exercise training can reverse age-related periph-eral circulatory changes in healthy older men. Circulation. 1999;100:1085–1094.
Pre
-foo
dP
ost-
food
0 W
5 W
10 W
15 W
20%
40%
60%
80%
Pre
-foo
dP
ost-
food
0 W
5 W
10 W
15 W
20%
40%
60%
80%
Leg
Blo
od F
low
(m
l/min
)
0
500
1000
1500
2000
2500
3000
3500
4000
4500
Mea
n A
rter
ial P
ress
ure
(mm
Hg)
0
80
100
120
140
160YoungOld
Figure 6. Leg blood flow and mean arterial pressure during multiple time points during the time control study in the young (black circles) and old (white circles) are displayed. No significant differences were observed between measurements at rest or at equivalent exercise intensities.
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from

Page 12 of 12 BARREtt-O’KEEFE Et AL.
10. Proctor DN, Shen PH, Dietz NM, et al. Reduced leg blood flow dur-ing dynamic exercise in older endurance-trained men. J Appl Physiol. 1998;85:68–75.
11. Buchner DM. Physical activity and prevention of cardiovascular disease in older adults. Clin Geriatr Med. 2009;25:661–675, viii. 10.1016/j.cger.2009.08.002
12. Ishihata A, Katano y, Morinobu S, Endoh M. Influence of aging on the contractile response to endothelin of rat thoracic aorta. Eur J Pharmacol. 1991;200:199–201.
13. Goettsch W, Lattmann T, Amann K, et al. Increased expression of endothelin-1 and inducible nitric oxide synthase isoform II in aging arteries in vivo: implications for atherosclerosis. Biochem Biophys Res Commun. 2001;280:908–913. doi:10.1006/bbrc.2000.4180
14. Donato AJ, Gano LB, Eskurza I, et al. Vascular endothelial dysfunc-tion with aging: endothelin-1 and endothelial nitric oxide synthase. Am J Physiol heart Circ Physiol. 2009;297:H425–H432. doi:10.1152/ajpheart.00689.2008
15. Thijssen DH, Rongen GA, van Dijk A, Smits P, Hopman MT. Enhanced endothelin-1-mediated leg vascular tone in healthy older subjects. J Appl Physiol. 2007;103:852–857. doi:10.1152/japplphysiol.00357.2007
16. Van Guilder GP, Westby CM, Greiner JJ, Stauffer BL, DeSouza CA. Endothelin-1 vasoconstrictor tone increases with age in healthy men but can be reduced by regular aerobic exercise. hypertension. 2007;50:403–409. doi: 10.1161/hypertensionaha.107.088294
17. Krum H, Viskoper RJ, Lacourciere y, Budde M, Charlon V. The effect of an endothelin-receptor antagonist, bosentan, on blood pres-sure in patients with essential hypertension. Bosentan Hypertension Investigators. N Engl J Med. 1998;338:784–790. doi:10.1056/NEJM199803193381202
18. Barrett-O’Keefe Z, Ives SJ, Trinity JD, et al. Taming the “sleeping giant”: the role of endothelin-1 in the regulation of skeletal mus-cle blood flow and arterial blood pressure during exercise. Am J Physiol heart Circ Physiol. 2013;304:H162–H169. doi:10.1152/ajpheart.00603.2012
19. Amann M, Runnels S, Morgan DE, et al. On the contribution of group III and IV muscle afferents to the circulatory response to rhyth-mic exercise in humans. J Physiol. 2011;589(Pt 15):3855–3866. doi:10.1113/jphysiol.2011.209353
20. Barden J, Lawrenson L, Poole JG, et al. Limitations to vasodila-tory capacity and.VO2 max in trained human skeletal muscle. Am J Physiol heart Circ Physiol. 2007;292:H2491–H2497. doi:10.1152/ajpheart.01396.2006
21. Wray DW, Witman MA, Ives SJ, et al. Progressive handgrip exercise: evidence of nitric oxide-dependent vasodilation and blood flow regu-lation in humans. Am J Physiol heart Circ Physiol. 2011;300:H1101–H1107. doi:10.1152/ajpheart.01115.2010
22. Goddard J, Webb DJ. Endothelin antagonists and hypertension: a question of dose? hypertension. 2002;40:e1–2; author reply e1–2.
23. Helmy A, Newby DE, Jalan R, Hayes PC, Webb DJ. Enhanced vasodilatation to endothelin antagonism in patients with compensated cirrhosis and the role of nitric oxide. Gut. 2003;52:410–415.
24. Ihara M, Noguchi K, Saeki T, et al. Biological profiles of highly potent novel endothelin antagonists selective for the ETA receptor. Life Sci. 1992;50:247–255.
25. Haynes WG, Webb DJ. Contribution of endogenous generation of endothelin-1 to basal vascular tone. Lancet. 1994;344:852–854.
26. Andersen P, Adams RP, Sjøgaard G, Thorboe A, Saltin B. Dynamic knee extension as model for study of isolated exercising muscle in humans. J Appl Physiol. 1985;59:1647–1653.
27. Richardson RS, Saltin B. Human muscle blood flow and metabolism studied in the isolated quadriceps muscles. Med Sci Sports Exerc. 1998;30:28–33.
28. Wray DW, Uberoi A, Lawrenson L, Richardson RS. Heterogeneous limb vascular responsiveness to shear stimuli during dynamic exer-cise in humans. J Appl Physiol. 2005;99:81–86. doi:01285.2004 [pii] 10.1152/japplphysiol.01285.2004
29. Donato AJ, Uberoi A, Wray DW, Nishiyama S, Lawrenson L, Richardson RS. Differential effects of aging on limb blood flow in humans. Am J Physiol heart Circ Physiol. 2006;290:H272–H278. doi:00405.2005 [pii]10.1152/ajpheart.00405.2005
30. Rowell LB. human Cardiovascular Control. New york: Oxford University Press; 1993:xv, 500.
31. Logason K, Bärlin T, Jonsson ML, Boström A, Hårdemark HG, Karacagil S. The importance of Doppler angle of insonation on differentiation between 50-69% and 70-99% carotid artery ste-nosis. Eur J Vasc Endovasc Surg. 2001;21:311–313. doi:10.1053/ejvs.2001.1331
32. Jern C, Seeman-Lodding H, Biber B, Winsö O, Jern S. An experimen-tal multiple-organ model for the study of regional net release/uptake rates of tissue-type plasminogen activator in the intact pig. thromb haemost. 1997;78:1150–1156.
33. Smith EG, Voyles WF, Kirby BS, Markwald RR, Dinenno FA. Ageing and leg postjunctional alpha-adrenergic vasoconstrictor responsive-ness in healthy men. J Physiol. 2007;582(Pt 1):63–71. doi:10.1113/jphysiol.2007.130591
34. Newcomer SC, Leuenberger UA, Hogeman CS, Handly BD, Proctor DN. Different vasodilator responses of human arms and legs. J Physiol. 2004;556(Pt 3):1001–1011.
35. Laughlin MH, Korthuis RJ, Duncker DJ, Bache RJ. Control of blood flow to cardiac and skeletal muscle during exercise. In: LB Rowell & JT Shepherd (Eds), handbook of Physiology. Exercise: Regulation and Integration of Multiple Systems. New york: Oxford University Press; 1996:705–769.
36. McEniery CM, Wilkinson IB, Jenkins DG, Webb DJ. Endogenous endothelin-1 limits exercise-induced vasodilation in hypertensive humans. hypertension. 2002;40:202–206.
37. Landin RJ, Linnemeier TJ, Rothbaum DA, Chappelear J, Noble RJ. Exercise testing and training of the elderly patient. Cardiovasc Clin. 1985;15:201–218.
38. Fletcher G. Physical inactivity as a risk factor for cardiovascular dis-ease. Am J Med. 1999;107(2A):10S–11S.
39. Goligorsky MS, Tsukahara H, Magazine H, Andersen TT, Malik AB, Bahou WF. Termination of endothelin signaling: role of nitric oxide. J Cell Physiol. 1994;158:485–494. doi:10.1002/jcp.1041580313
40. Bender SB, Klabunde RE. Altered role of smooth muscle endothelin receptors in coronary endothelin-1 and alpha1-adrenoceptor-mediated vasoconstriction in Type 2 diabetes. Am J Physiol heart Circ Physiol. 2007;293:H2281–H2288. doi:10.1152/ajpheart.00566.2007
41. Rådegran G, Saltin B. Nitric oxide in the regulation of vasomo-tor tone in human skeletal muscle. Am J Physiol. 1999;276(6 Pt 2):H1951–H1960.
42. McAllister RM, Newcomer SC, Pope ER, Turk JR, Laughlin MH. Effects of chronic nitric oxide synthase inhibition on responses to acute exercise in swine. J Appl Physiol. 2008;104:186–197. doi:10.1152/japplphysiol.00731.2007
43. Tschakovsky ME, Joyner MJ. Nitric oxide and muscle blood flow in exercise. Appl Physiol Nutr Metab. 2008;33:151–161. doi:10.1139/H07-148
44. Dyke CK, Proctor DN, Dietz NM, Joyner MJ. Role of nitric oxide in exercise hyperaemia during prolonged rhythmic handgripping in humans. J Physiol. 1995;488(Pt 1):259–265.
45. Taddei S, Virdis A, Mattei P, et al. Aging and endothelial function in normotensive subjects and patients with essential hypertension. Circulation. 1995;91:1981–1987.
46. Schrage WG, Eisenach JH, Joyner MJ. Ageing reduces nitric-oxide- and prostaglandin-mediated vasodilatation in exercis-ing humans. J Physiol. 2007;579(Pt 1):227–236. doi:10.1113/jphysiol.2006.124313
47. Richardson RS, Secher NH, Tschakovsky ME, Proctor DN, Wray DW. Metabolic and vascular limb differences affected by exer-cise, gender, age, and disease. Med Sci Sports Exerc. 2006;38: 1792–1796. doi:10.1249/01.mss.0000229568.17284.ab 00005768-200610000-00014 [pii]
by guest on April 28, 2016
http://biomedgerontology.oxfordjournals.org/
Dow
nloaded from
Related Documents











