Efficient myogenic reprogramming of adult white fat stem cells and bone marrow stem cells by freshly isolated skeletal muscle fibers HENRY COLLINS-HOOPER, GRAHAM LUKE, MARK CRANFIELD, WILLIAM R. OTTO, STEVE RAY, and KETAN PATEL READING AND LONDON, UK Stem cells that can be directed to differentiate into specific cell types offer the pros- pect of a renewable source of replacement cells to treat diseases. This study evalu- ates the reprogramming of 2 readily available stem cell populations into skeletal muscle. We show for the first time that freshly isolated muscle fibers reprogram bone marrow or white fat stem cells far more efficiently than muscle cell lines. In ad- dition, we show that the ability of muscle fibers to reprogram stem cells can be almost doubled through the use of chromatin remodeling reagents such as trichostatin A. This novel approach permits the generation of myogenic cells that could be used to treat a range of muscle-wasting diseases. (Translational Research 2011;158:334–343) Abbreviations: BMSC ¼ bone marrow stromal stem cell; DAPI ¼ 4’,6-diamidino-2-phenylindole; DMEM ¼ Dulbecco’s modified Eagle’s medium; DSHB ¼ Developmental Studies Hybridoma Bank; EDL ¼ extensor digitorum longus; FBS ¼ fetal bovine serum; GFP ¼ glial filament protein; MHC ¼ myosin heavy chain; PBS ¼ phosphate-buffered saline; PCR ¼ polymerase chain reaction; rBMSC ¼ rat bone marrow stromal stem cell; rWFSC ¼ rat white fat stem cell; IGF-1 ¼ insulin-like growth factor-1; SFCM ¼ single-fiber culture media; TSA ¼ trichostatin A; WFSC ¼ white fat stem cell M ultipotent stem cells have been identified from a variety of tissues and could be used as therapeutic reagents for the treatment of inherited diseases or restoring degenerated tissues. 1,2 For therapeutic value, stem cells should be readily available, ideally requiring minimal surgical intervention to obtain the desired tissue. Therefore, it is optimum if only a small tissue sample can be taken and that the resident stem cell population can be expanded in culture while maintaining their multipotency, normal karyotype, and ability to engraft at a variety of sites. Bone marrow stromal stem cells (BMSCs) meet some of these criteria and can be expanded for more than 70 population doublings. 3 However, their isolation is problematic, it is not only painful but also yields low numbers of suitable cells. Much attention has focused on white fat stem cells (WFSCs). These are not only multipotent but also present at readily available peripheral sites around the human body. 4 They also have a remarkable ability to divide while retaining their multipotency. These cells undergo more than 200 population doublings and retain their ability to differentiate into adipogenic, osteogenic, and myogenic lineages. 5 Many adult stem cell populations display multipo- tency but do not engraft at efficiencies likely to be of therapeutic value. One possible option to use adult stem cells to repair the tissue of choice is first to From School Biological Sciences, University of Reading, Reading, UK; Natural Bioscience, Animal and Microbial Building, Whiteknights, Reading, UK; Histopathology Laboratory, Cancer Research UK, London, UK. Supported by the BBSRC and Systems Biology Laboratory (SBL). Submitted for publication April 28, 2011; revision submitted July 26, 2011; accepted for publication July 29, 2011. Reprint requests: Ketan Patel, School Biological Sciences, Hopkins Building, University of Reading, Reading RG6 6UB, UK; e-mail: [email protected]. 1931-5244/$ - see front matter Ó 2011 Mosby, Inc. All rights reserved. doi:10.1016/j.trsl.2011.07.009 334

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Efficient myogenic reprogramming of adult white fatstem cells and bone marrow stem cells by freshlyisolated skeletal muscle fibers
HENRY COLLINS-HOOPER, GRAHAM LUKE, MARK CRANFIELD, WILLIAM R. OTTO, STEVE RAY,and KETAN PATEL
READING AND LONDON, UK
From School Biological Sciences, U
Natural Bioscience, Animal and M
Reading, UK; Histopathology La
London, UK.
Supported by the BBSRC and Syst
Submitted for publication April 28,
2011; accepted for publication July
Reprint requests: Ketan Patel, Sch
Building, University of Reading,
1931-5244/$ - see front matter
� 2011 Mosby, Inc. All rights rese
doi:10.1016/j.trsl.2011.07.009
334
Stem cells that can be directed to differentiate into specific cell types offer the pros-pect of a renewable source of replacement cells to treat diseases. This study evalu-ates the reprogramming of 2 readily available stem cell populations into skeletalmuscle. We show for the first time that freshly isolated muscle fibers reprogrambone marrow or white fat stem cells far more efficiently than muscle cell lines. In ad-dition, we show that the ability ofmuscle fibers to reprogram stemcells canbe almostdoubled through the use of chromatin remodeling reagents such as trichostatin A.This novel approach permits the generation of myogenic cells that could be usedto treat a range of muscle-wasting diseases. (Translational Research2011;158:334–343)
Abbreviations: BMSC ¼ bone marrow stromal stem cell; DAPI ¼ 4’,6-diamidino-2-phenylindole;DMEM ¼ Dulbecco’s modified Eagle’s medium; DSHB ¼ Developmental Studies HybridomaBank; EDL ¼ extensor digitorum longus; FBS ¼ fetal bovine serum; GFP ¼ glial filament protein;MHC¼myosin heavychain; PBS¼phosphate-buffered saline; PCR¼polymerasechain reaction;rBMSC ¼ rat bone marrow stromal stem cell; rWFSC ¼ rat white fat stem cell; IGF-1 ¼ insulin-likegrowth factor-1; SFCM¼ single-fiberculturemedia; TSA¼ trichostatinA;WFSC¼white fat stemcell
M ultipotent stem cells have been identifiedfrom a variety of tissues and could be used astherapeutic reagents for the treatment
of inherited diseases or restoring degenerated tissues.1,2
For therapeutic value, stem cells should be readilyavailable, ideally requiring minimal surgical intervention
niversity of Reading, Reading, UK;
icrobial Building, Whiteknights,
boratory, Cancer Research UK,
ems Biology Laboratory (SBL).
2011; revision submitted July 26,
29, 2011.
ool Biological Sciences, Hopkins
Reading RG6 6UB, UK; e-mail:
rved.
to obtain the desired tissue. Therefore, it is optimum ifonly a small tissue sample can be taken and that theresident stem cell population can be expanded in culturewhile maintaining their multipotency, normal karyotype,and ability to engraft at a variety of sites. Bone marrowstromal stem cells (BMSCs) meet some of these criteriaand can be expanded for more than 70 populationdoublings.3 However, their isolation is problematic, it isnot only painful but also yields low numbers of suitablecells. Much attention has focused on white fat stem cells(WFSCs). These are not only multipotent but also presentat readily available peripheral sites around the humanbody.4 They also have a remarkable ability to divide whileretaining their multipotency. These cells undergo morethan 200 population doublings and retain their ability todifferentiate into adipogenic, osteogenic, and myogeniclineages.5
Many adult stem cell populations display multipo-tency but do not engraft at efficiencies likely to be oftherapeutic value. One possible option to use adultstem cells to repair the tissue of choice is first to

AT A GLANCE COMMENTARY
Collins-Hooper H, et al.
Background
Stem cells that can be directed to differentiate into
specific cell types offer the prospect of a renewable
source of replacement cells to treat diseases. This
study evaluates the reprogramming of 2 readily
available stem cell populations into skeletal muscle.
Translational Significance
We show that freshly isolated muscle fibers repro-
gram bone marrow or white fat stem cells far more
efficiently than muscle cell lines. Second, we show
that the ability of muscle fibers to reprogram stem
cells can be almost doubled through the use of
chromatin remodeling reagents.
Translational ResearchVolume 158, Number 6 Collins-Hooper et al 335
reprogram and then transplant the autologous cells intoan affected area. The reprogramming of adult stem cellscan be achieved by several approaches. Viral deliverymethods can be used to introduce master regulators ofspecific cell lineages. This approach has been shownto work efficiently for the production of skeletal musclecells.6 However, the use of viral vectors is not withoutsafety implications. Many nonviral mechanisms havebeen shown recently to reprogram stem cells includingthe use of conditioned medium, microvesicles as well asdirect introduction of RNA.7-9 Another regularly usedapproach to reprogram stem cells is through thecoculture technique. Such an approach has been usedpotentially to develop cell-based therapeutics to treatskeletal muscle wasting diseases.10
In this study, we aimed to improve the rate of myo-genic stem cell reprogramming. Recently, we developeda technique that permits the isolation of intact myofibersdecorated with functional myocytic satellite stem cellsfrom freshly dissected muscle. This technique hasbeen used to study features of muscle stem cell behaviorbecause it mimics closely the conditions found in anin vivo setting.11,12 We hypothesize that these fibersand their associated stem cells are likely to secretea spectrum of proteins that differ from those producedby long term muscle cell line cultures. Second, wesuggest that the factors produced by muscle fibers areconducive to promoting myogenic reprogramming ofadult stem cells.
MATERIALS AND METHODS
Animals. Green fluorescent protein positive (GFP1)and GFP negative (GFP2) BMSCs and WFSCs were
isolated from 8-week-old female Wistar GFPtransgenic rats or nontransgenic rats respectively. Allcells originating from rats are denoted by the prefix‘‘r’’ (rBMSCs and rWFSCs). RNA and conditionedmedium were isolated from 8-week-old female,nontransgenic cultured rat tissue. Briefly, the extensordigitorum longus (EDL) or tibialis anterior weremicrodissected and used immediately for RNAisolation, or the EDL was dispersed enzymaticallyinto single fibers and cultured for 5 days. Thereafter,the media were collected and the fibers were used asa source of RNA. Single myofibers for coculture wereisolated from the EDL of 4-month-old female wild-type or ubiquitous GFP C57Black6/J mice. Allanimals were killed according to local ethicscommittee approved methods.
WFSC and BMSC isolation and culture. The adipose tis-sue for rWFSC isolation was harvested from the visceralfat according to the method of Bunnell et al.13 Briefly,after washing with phosphate-buffered saline (PBS)containing 0.5% (vol/vol) penicillin/streptomycin (PSfrom stock containing 10,000 U/ml penicillin andstreptomycin 10,000 mg/ml) (Invitrogen Corp,Carlsbad, Calif) the tissue was finely minced in 0.1%(wt/vol) Type I collagenase (Sigma Chemical Co, StLouis, Mo) in PBS/2% PS. The minced tissue was thensubjected to trituration and incubated at 37�C, 5% CO2
for 30 min. Collagenase was then neutralized by theaddition of Alpha MEM (Fisher Scientific, Pittsburgh,Pa) containing, 20% (vol/vol) fetal bovine serum(FBS) (Fisher), 1% PS. The adipose tissue was thentriturated before being centrifuged twice at 1200 rpmfor 5 min, with vigorous shaking between stages todisrupt the pellet. The supernatant was aspirated andPBS/2% PS was added before being centrifuged againat 1200 rpm for 5 min. Aspiration of the supernatantwas followed by the subsequent resuspension of thepellet in Alpha MEM containing 20% FBS/1% PS Thecell suspension was then sieved through a cell strainerbefore plating out into T25 tissue culture flasks(NUNC; Thermo Scientific, Lafayette, Colo). The cellswere cultured in Dulbecco’s modified eagle’s medium(DMEM) containing Glutamax (GIBCO; Invitrogen)supplemented with 20% FBS/1% PS.rBMSCs were harvested from the tibiae and femurs of
rats according to the method of Soleimani and Nadri.14
The bones were dissected from the hind limbs andplaced into PBS/2% PS. The bone marrow was thenflushed with the Alpha MEM containing 10% FBS/1%PS using 21-gauge needle. The cell suspension wasthen filtered through a cell strainer before being platedinto T25 tissue culture flasks. The cells were culturedin DMEM containing Glutamax supplemented with10% FBS/1% PS. The initial plating out of cells was

Translational Research336 Collins-Hooper et al December 2011
defined as passage 0. Media changes for both cell typeswere carried out every 2–3 days.
Single muscle fiber isolation and culture. The isolationof single muscle fibers with associated satellite cellswas according to the method of Rosenblatt et al.15
Dissection of the EDL from either rats or mice wascarried out in the same manner. Care was taken not todamage the muscle fibers. Muscle connective tissuewas digested using 0.2% (wt/vol) Type 1 collagenasein DMEM containing Glutamax for 2 h to generatesingle myofibers. Single-fiber culture media (SFCM-DMEM containing Glutamax, 10% (vol/vol) horseserum, 0.5% (vol/vol) chick embryo extract, 1% PS)was used to neutralize the collagenase and forculturing the muscle fibers. The myofibers for RNAisolation were cultured for a period of 5 days, whereasothers were cocultured for 14 days at 37�C, 5% CO2.
Cell characterization. A reverse-transcription poly-merase chain reaction (PCR) analysis was performedto determine cell surface marker expression. rWFSCswere characterized for the expression of CD44 andCD90, and to confirm negativity for CD56.16 rBMSCswere characterized for the expression of CD44 andCD45.17 Primer details:
rCD44 50 primer ACATGCTACAAGCACAACCTGGGCAGrCD44 30 primer GGAAGACTTCCACCTCTTCTTGCACCArCD90 50 primer AGCTAGGGGAGCCCAGACCCAGrCD90 30 primer CTCTTCAGCAGACCTTGAGGTTTCCmCD56 50 primer AGCAGGATGCCTCCATCCACCTCmCD56 30 primer TGGTGTTGCTGGCAGTGCAGATGrCD45 50 primer TGCACAAGGGCCCCGGGATGAGACrCD45 30 primer AGGTGCCTGTACGCCCAACGCCAG
Primers were optimized using a gradient PCR pro-gram, and the samples were amplified for 30 cycles.DNAwas resolved by electrophoresis in agarose gels.Antibody staining for caveolin 1 (gift from Dr D. Tan-
netta, Oxford University) and Pax7 (DevelopmentalStudies Hybridoma Bank [DSHB]) was used to charac-terize mouse satellite cells.18
RNA isolation and conditioned media preparation. TheSFCM from single EDL muscle fiber cultures was col-lected and filtered and used as the conditioned media.Fibers from the cultured and freshly isolated muscleswere processed for RNA isolation using Trizol Reagent(GIBCO) according to the manufacturer’s protocol, and
redissolved at 150 mg/mL RNA in PBS which was ap-plied to the mesenchymal stem cells for a period of 30min. The cells were then washed with DMEM contain-ing Glutamax and then maintained in either SFCM ordifferentiation media (DM) (DMEM containing Gluta-max, 2% horse serum/1% PS).Quantitative PCR was performed using an ABI7500
thermocycler (Applied BioSystems, Foster City, Calif).Individual PCRs were performed for a minimum of 3times using standard protocol.19 The following primerswere used:
rPax7 50 primer TCCGCCATCAACCATGCATCAGrPax7 30 primer CAGCCACAGGGTCCACACTGTAGCrMyoD 50 primer ACCTCTTTCCTTTCCTCACAGTCCCTArMyoD 30 primer CAGAAAGTGGGAACAGTGTCTCCACrMrf4 50 primer AGAAGTGGCAGAGGGCTCTCCTTTGTrMrf4 30 primer TCCACGATGGAAGAAAGGCGCTGrMyogenin 50 primer TCCTGTCCACCTTCAGGGCTTCrMyogenin 30 primer CAAGGGGGCTCTCTGGACTCC.
rMrf4 and rMyogenin products were distinguishedfrom mouse sequences by digesting the product ofPCR after the first cycle with AccI and NcoI, respec-tively. All amplifications were performed for a maxi-mum of 35 cycles after establishing optimalconditions using a temperature gradient program.
Culture of mesenchymal stem cells and muscle fibersin transwell dishes. Passage two to three rBMSCs orrWFSCs were trypsinized and plated out into 12-welldishes (Greiner Bio One, Monroe, NC) at 30,000 cellsper well and left overnight to adhere. In all, 2.5 mmol/L trichostatin A (TSA) was used when required(Sigma). The cells were then washed with PBS andmaintained in SFCM or DM. Ten mouse EDLmyofibers were then added to the required wells ortranswell dishes (Millipore, Bedford, Mass). The cellswere cultured for up to 14 days before being fixedwith 4% paraformaldehyde (PFA)/PBS for 15 min andstored in PBS at 4�C.
Immunocytochemistry. After fixation with 4% PFA,the cells were treated with permeabilization buffer (20mmol/L HEPES pH 7, 300 mmol/L sucrose, 50 mmol/L NaCl, 3 mmol/L MgCl2 and 0.5% Triton X-100) at4�C for 15min. Subsequent washes in PBS at room tem-perature were followed by blocking of nonspecific bind-ing using wash buffer (5% (vol/vol) fetal calf serum inPBS with 0.05% Triton X-100) for 30 min. The primary

Translational ResearchVolume 158, Number 6 Collins-Hooper et al 337
antibodies used were rat monoclonal antimyogenin(DSHB clone F5D [1:2]) and mouse monoclonal anti-myosin heavy chain (MHC) (DSHB clone A4.1025[1:1]). The primary antibodies were diluted with washbuffer and incubated with cells overnight at 4�C for20 h. Alexafluor 594-conjugated goat antimouseimmunoglobulin G (Molecular Probes [1:200])secondary antibody was used to visualize the primaryantibody. The samples were then mounted usingfluorescent mounting medium (DakoCytomation;DAKO Corp, Carpinteria, Calif) containing 2.5 mg/mL40,6-diamidino-2-phenylindole (DAPI) for nuclearvisualization. Images were taken at 203magnification using a Nikon Eclipse TE200 invertedmicroscope (Nikon Inc, Melville, NY).
Insulin-like growth factor-1 (IGF 1) and growth hormonemeasurements. Supernatants from cell culture experi-ments were frozen at –80�C until required for enzyme-linked immunosorbent assay (ELISA) measurementsof IGF-1 (product code MG100; R&D Systems,Minneapolis, Minn) and growth hormone (productcode EZRMGH-45K; Millipore) according to themanufacturers’ instructions.
Data quantification. An analysis was performed bymanual counting of the DAPI-stained myonuclei, totalnuclei, and the number of myotubes. Allquantification was based on the images that werecaptured after immunocytochemistry. A central regionof the culture dish was analyzed that containeda minimum of 300 nuclei. Myotubes were defined ashaving 3 or more myonuclei. All data were generatedfrom a minimum of 3 separate experiments, each ofwhich contained a minimum of 3 wells per condition.The data are shown as means 6 standard deviation.Significant differences were considered for P # 0.05.
RESULTS
The field of cellular reprogramming has been con-fused by experimental designs that have failed to re-solve the difference between reprogramming andheterogenic cell fusion. This issue is important becauseoften it has led to the overestimation of the true degreeof reprogramming. To determine the degree of reprog-ramming and any other type of event that would yieldmyogenic cells, we carried out 2 complementary exper-iments. In the first, wild-type rWFSCs/GFP2 (orrBMSCs/GFP2) were cultured with GFP1 mouse skel-etal muscle fibers. rWFSCs and rBMSCs (and musclestem cells on fibers) were isolated according to standardprotocols, and analyses showed that they expressed thecorrect cell surface markers: rWFSCs were CD441/CD901/CD562, rBMSCs were CD441,CD451, andmuscle stem cells were Caveolin11/Pax71. The cellswere cultured in single-fiber culture medium up to 14
days and then processed for immunocytochemistry todetect the marker of terminally differentiated muscleMHCs. The numbers of cells that expressed MHC andwere either GFP1 or GFP2 were counted. Our analysesshowed that the culture of rWFSCs (or rBMSCs) withGFP1 mouse muscle fibers resulted in 60% of MHC1
cells being GFP2. These cells could have arisen throughreprogramming. The remaining 40% were MHC1/GFP1 (Fig 1, A). This latter population could representmouse satellite cells that had undergone differentiationor satellite cells that had fused with rat nonmuscle cells.We performed the complementary experiment in whichwild-type mouse muscle fibers were cultured with eitherrWFSCs/GFP1 or rBMSCs/GFP1 for 14 days prior toMHC profiling. In these experiments, we found that60% of MHC1 cells were also GFP1 for either WFSCsor BMSCs (Fig 1, A). These could have arisen from ei-ther the reprogramming of rWFSCs or rBMSCs or fu-sion between rWFSCs or rBMSCs with satellite cellsfrom mouse muscle fibers. However, given that the firstexperiment showed that reprogramming occurred in60% of the cells, we conclude that most of the MHC-positive cells that have formed when either rWFSCsor rBMSCs that are cultured with muscle fibers developthrough reprogramming. We confirmed these results bytaking advantage of the fact that DAPI staining of rat nu-clei shows smooth uniform distribution, whereas it ap-pears speckled in mouse cells.20,21 In experimentswhere GFP1 mouse muscle fibers were cultured withwild-type rWFSCs, all nuclei in cells (total of 18 myo-tubes) that expressed MHC and were GFP- displayedsmooth nuclei (Fig 1, B and C). Furthermore, onlyspeckled nuclei were found MHC1/GFP1 myotubes(n 5 18) (Fig 1, B and D). These results also allow usto conclude that fusion between mouse satellite cellsand rWFSCs or rBMSCs is a rare occurrence.We next determined the reprogramming rate of
rWFSCs and rBMSCs (both GFP2) when coculturedwith the mouse muscle cell line GFP1/C2C12. The cellswere grown in conditions identical to those with musclefibers, but these experiments gave rise to only 1–2 toweakly expressing MHC1/GFP2 cells per 1000 cells.Therefore, the muscle fibers are more potent at promot-ing the reprogramming of rWFSCs and rBMSCs thanC2C12 cells.Primary cultures of rBMSCs and rWFSCs can be
maintained for many passages without losing the pluri-potency. However, single muscle fibers have to be iso-lated freshly. For all future experiments, we usedGFP1/rWFSCs or GFP1/rBMSCs with wild-typemouse muscle fibers to minimize the use of transgenicanimals. In the first set of experiments, we aimed tocompare the ability of mouse muscle fibers to repro-gram rWFSCs with rBMSCs. The expression of MHC
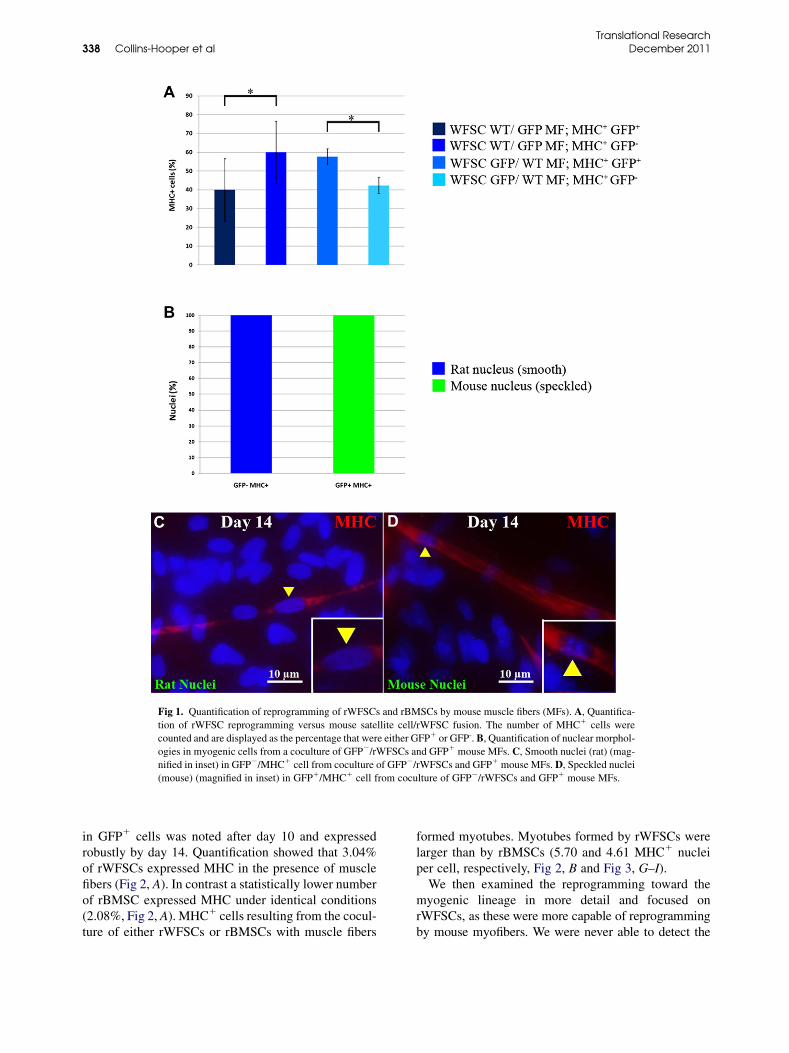
Fig 1. Quantification of reprogramming of rWFSCs and rBMSCs by mouse muscle fibers (MFs). A, Quantifica-
tion of rWFSC reprogramming versus mouse satellite cell/rWFSC fusion. The number of MHC1 cells were
counted and are displayed as the percentage that were either GFP1 or GFP-. B, Quantification of nuclear morphol-
ogies in myogenic cells from a coculture of GFP2/rWFSCs and GFP1 mouse MFs. C, Smooth nuclei (rat) (mag-
nified in inset) in GFP2/MHC1 cell from coculture of GFP2/rWFSCs and GFP1 mouse MFs. D, Speckled nuclei
(mouse) (magnified in inset) in GFP1/MHC1 cell from coculture of GFP2/rWFSCs and GFP1 mouse MFs.
Translational Research338 Collins-Hooper et al December 2011
in GFP1 cells was noted after day 10 and expressedrobustly by day 14. Quantification showed that 3.04%of rWFSCs expressed MHC in the presence of musclefibers (Fig 2, A). In contrast a statistically lower numberof rBMSC expressed MHC under identical conditions(2.08%, Fig 2, A). MHC1 cells resulting from the cocul-ture of either rWFSCs or rBMSCs with muscle fibers
formed myotubes. Myotubes formed by rWFSCs werelarger than by rBMSCs (5.70 and 4.61 MHC1 nucleiper cell, respectively, Fig 2, B and Fig 3, G–I).We then examined the reprogramming toward the
myogenic lineage in more detail and focused onrWFSCs, as these were more capable of reprogrammingby mouse myofibers. We were never able to detect the

Translational ResearchVolume 158, Number 6 Collins-Hooper et al 339
expression of Pax7, which is a myogenic stem cellmarker11,22 with either an immunocytochemical ora PCR approach in rat cells. Rat MyoD expression wasdetected by PCR from 2 days onward with expressiongradually decreasing over time (Fig 2, G). In contrast,the expression of 2 other members of the Mrf familyshowed a more dynamic expression. We detected proteinexpression ofMyogenin, which is amarker ofmuscle dif-ferentiation in rat cells, after 3 days of coculture, becom-ing strong by 5 days and maintained until day 14 albeit atlower levels (Figs 3, A, B, D, and E). Molecular profilingof rat myogenin mirrored the abundance of the protein(Fig 2, G). rBMSC also expressed Myogenin at days 5and 14 (Figs 3, C and F). The expression profile of ratMrf-4was similar to that of ratmyogenin, with a peak ex-pression at 3 days followed by a steep decline in transcriptabundance (Fig 2, G).Next, we determined whether reprogramming could
be mediated indirectly. We found that rWFSCs exposedto a continuous presence of medium conditioned bymouse muscle fibers (achieved using transwells) alsounderwent myogenic conversion, with 2.94% of all nu-clei showing robust myogenin distribution (Fig 2, C andFig 3, J). Significantly myogenin-positive cells from thetranswell experiments did not go on to express MHCand did not form myotubes (Fig 2,D). Mouse muscle fi-ber conditioned medium or RNA frommuscle fibers didinduce myogenin expression in rWFSCs (Fig 2, C).We next investigated whether the chromatin remodel-
ing of target cell DNA could promote myogenic reprog-ramming. rWFSCs were treated with TSA for 24 priorto the exposure to muscle fibers. The cells were washedwith culture medium to remove TSA before the intro-duction of muscle fibers. In a second variation, somecultures were exposed to a differentiation medium thatwas substituted for SFCM 7 days into the experiment.We found that the number of nuclei in MHC-expressing cells increased from 3.04% to 5.69% whenWFSCs had been pretreated with TSA (Fig 2, E andFig 3, L). rWFSCs cultured with muscle fibers in a sin-gle-fiber culture medium for 7 days before switching todifferentiation medium had no significant outcome inthe reprogramming of rWFSCs (Fig 2, E). However,the rWFSCs that had been pretreated with TSA andthen exposed to differentiation medium gave rise toMHC1 cells, which accounted for 7.63% of all nuclei.(Fig 2, E and Fig 3, K). Finally, our analyses showedthat treatment of rWFSC with TSA led to the formationof larger myotubes, irrespective of the culture medium(Fig 2, F).Last, we examined whether the paracrine-mediated
reprogramming of rWFSCs by mouse muscle fiberswas through the production of IGF-1, which is a factorthat has been shown to direct the differentiation
program of multipotent neural stem cells.23,24 ELISA-based measurements showed that muscle fibers pro-duced little IGF-1 (maximum level of 32.66 pg/mL)during the reprogramming window (Fig 2, H). In con-trast, rWFSCs produced over 10 times as much IGF-1(maximum level of 352 pg/mL) (Fig 2,H). A synergisticeffect was observed between rWFSCs and muscle fibersthat led to a maximum IGF-1 level of 660 pg/mL (Fig 2,H). However, the pretreatment of rWFSCs with TSAprior to coculture led to little IGF-1 production (maxi-mum level of 37.37 pg/mL) even though these cells re-programmed more efficiently than without exposure toTSA (Fig 2, H). We conclude that IGF-1 is not playinga significant role in the reprogramming of rWFSCs bymouse muscle fibers. In addition, we could not measureany significant production of growth hormone byELISA in any of the preceding culture permutations(data not shown).
DISCUSSION
Our study aimed at exploiting the development ofa technique that permits the isolation of intact viablemuscle fibers, which only contain the resident stemcell population called satellite cells and the syncytiallyorganized myofiber. We showed previously that suchisolated fibers can be grown in coculture with a varietyof cells and that they secrete numerous growth factorsfor extended periods of time.12 Furthermore, the main-tenance of muscle stem cells on their original substra-tum permits these cells to display characteristicsfound in an in vivo setting. Many of these normal traitsare lost by muscle cell lines. To our knowledge, this isthe first study that has used single muscle fibers to repro-gram adult stem cells.We compared the myogenic reprogramming potential
of rBMSCs with rWFSCs and found that they both gaverise to cells that not only expressed markers of termi-nally differentiated skeletal muscle but also displayedcellular characteristics of normal muscle; both ex-pressed MHC and fused to form myotubes.We found that rWFSCs were reprogrammed more
readily by muscle fibers into the myogenic lineage com-pared with rBMSCs (3.04% compared with 2.08%). Thereprogramming was also rapid, as we detected the ex-pression of species-specific expression of MyoD in thefirst 48 h of coculture. However, it should be notedthat expression ofMyoD alone does not indicate a com-mitment to the myogenic differentiation because acti-vated satellite cells can express this member of theMrf family but subsequently revert back to a stem cellrather than form myotubes.11 Many reprogrammedstem cells went on to fuse into myotubes, often compris-ing up to 6 nuclei. This number is considerably higher
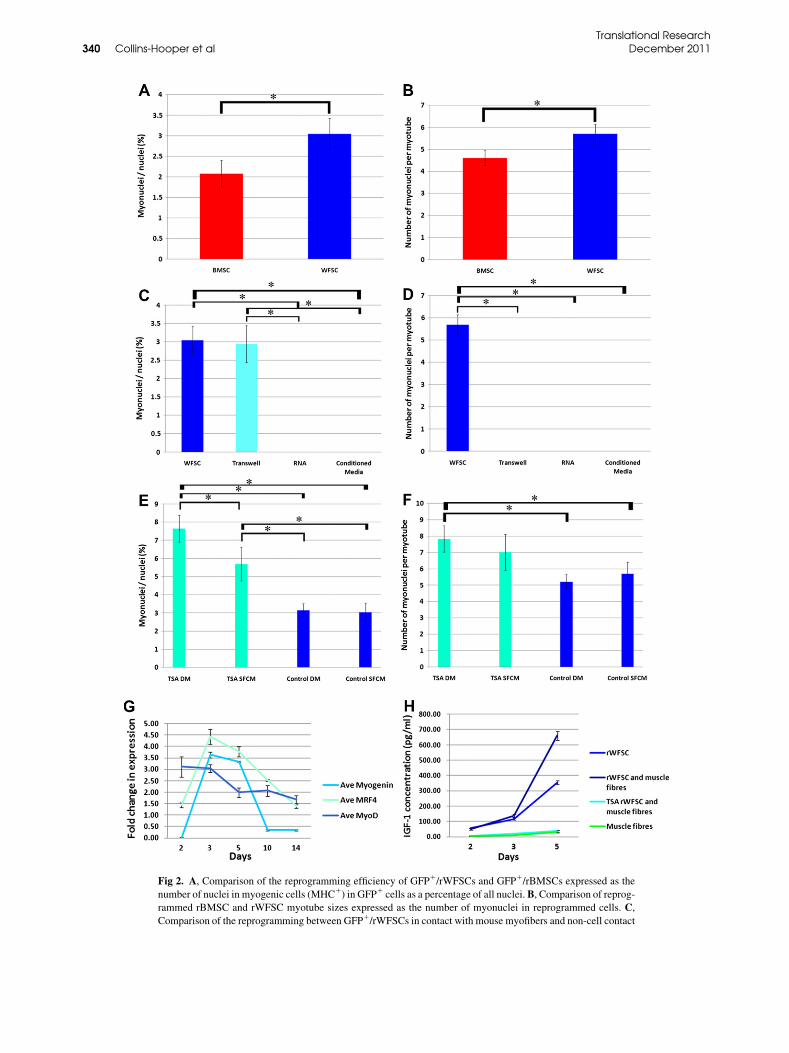
Fig 2. A, Comparison of the reprogramming efficiency of GFP1/rWFSCs and GFP1/rBMSCs expressed as the
number of nuclei in myogenic cells (MHC1) in GFP1 cells as a percentage of all nuclei.B, Comparison of reprog-
rammed rBMSC and rWFSC myotube sizes expressed as the number of myonuclei in reprogrammed cells. C,
Comparison of the reprogramming between GFP1/rWFSCs in contact with mouse myofibers and non-cell contact
Translational Research340 Collins-Hooper et al December 2011

Translational ResearchVolume 158, Number 6 Collins-Hooper et al 341
than previously reported by Di Rocco et al,25 who re-ported only 0.2% conversion of fat into muscle by pri-mary mouse myoblasts. Furthermore, we found thatfusion between mouse satellite cells and the rWFSCsor rBMSCs was a rare occurrence. This could be causedby species specificity or that satellite cells have a prefer-ence of staying in contact with the muscle fiber. Interest-ingly, using our coculture conditions, we found that thesingle fibers were much more potent at reprogrammingstem cells than the skeletal muscle stem cell lineC2C12. This outcome could have arisen for several rea-sons. First, 2 cell populations are present in the singlefibers, the myofiber and the satellite cells, both of whichhave unique gene activity. Second, these cells are iso-lated directly from whole animals and so they do notsuffer the disadvantages of cells that have been main-tained in vitro for extended periods. The most com-monly used skeletal muscle cell line C2C12 is welldocumented to contain numerous genetic alterations inits DNA.26
Several cell-free methods have been developed forreprogramming stem cells. The advantage of these isthat it allows the development of cell populations solelyof stem cell origin. We found that rWFSCs could be re-programmed by continual exposure to factors secretedinto media by single fibers. The number of cells con-verted into muscle in this cell-free system was thesame as in coculture experiments. This again was sur-prising because others have found that the use of trans-well systems decreased myogenic conversion of MSCby an order of magnitude.25 However, these cells failedto fuse into myotubes. Currently, we do not know whythemyogenic cells failed to fuse but point to the possibil-ity that the conversion takes place in 2 phases: 1 that in-volves small readily diffusing molecules which canconvert stem cells into mononuclear muscle cells andthe second phase thatmay require direct contact betweenmuscle cells and the converted cell. Alternatively, thetranswell membrane (pore size, 0.4 mm) may preventthe passage of larger biologically active entities. An ex-ample of which includes microvesicles, which can be1 mm in size27 and have been shown to be potent media-tors of cellular reprogramming.8 However, we showed
(transwell dishes, RNA, and conditioned media) conditions
GFP1 cells as a percentage of all nuclei. D, Myotube sizes
contact mediated reprogrammed cells and non-cell contact (
ditions.E, Quantification of the effect of TSA on the reprogra
nuclei in myogenic cells (MHCs1) within GFP1 cells, as a pe
rWFSCs for 24 h prior to coculture with the mouse myofiber
dium for the entire 14 days of coculture. DM indicates the use
coculture. F, Quantification of the effect of TSA on myotub
programmed cells. G, Quantitative PCR profile of rat myog
and mouse muscle fibers. H, Secretion of IGF-1 by coculture
tistical significance is at P , 0.05.
that conditioned medium from our muscle fibers couldnot reprogram WFSCs. This may not be totally surpris-ing if themicrovesicle concentration in themediumweretoo low to bring about a biologic response. In the future,wewill examinewhether a continual supply of microve-sicles is more efficacious at promoting myogenic re-programming of WFSCs.Several previous studies highlighted the importance
of inducing the expression of MyoD to initiate reprog-ramming to a myogenic lineage. Classic experimentsfrom the Weintraub laboratory showed that the forcedexpression of MyoD alone could convert certain celltypes to forming muscle.27,28 However, this was notpossible for all cell types including hepatocytes,possibly because of modification of the MyoDchromatin organization.29 Histone acetylation of chro-matin supports transcription, whereas deacetylation in-hibits transcription.30 We have used the histonedeacetylase inhibitor TSA and found that pretreatmentof rWFSCs enhanced their reprogramming potentialby almost 100%. The enhanced-level TSA inducedmyogenic reprogramming of WFSC was similar to theoutcome of applying the reagent to other cell types in-cluding hepatocytes.31
We increased the level of myogenic reprogrammingby manipulating the culture conditions. Myogenic dif-ferentiation can be induced by growing committed mus-cle cells in low serum conditions by a mechanism drivenby selective interactions of transcription factors on theserum response element at the MyoD locus.32 Our studypoints to an interesting property of TSA and the reprog-ramming process. We found no increase in the numberof myogenic cells after the introduction of differentia-tion medium to rWFSCs that had been cocultured inthe presence of myofibers compared with cultures ex-posed to normal serum-rich conditions. These resultsimply that a similar number of myogenic-conversionevents took place in both conditions. However, wefound that the number of myogenic cells increased inrWFSCs that had been treated with TSA and that thisnumber could be increased by growing the cells in dif-ferentiation medium. Therefore, TSA induces more re-programming events and that some of these cells are
expressed as the number of myogenin1 nuclei in
expressed as the number of myonuclei in MHC1 in
transwell dishes, RNA, and conditioned media) con-
mming of GFP1/rWFSCs expressed as the number of
rcentage of all nuclei. TSA indicates the treatment of
s. SFCM indicates the use of single-fiber culture me-
of differentiationmedium for the last 7 of 14 days of
e sizes expressed as the number of myonuclei in re-
enic gene expression in coculture of GFP1/rWFSCs
s of GFP1/rWFSCs and mouse muscle fibers. *Sta-
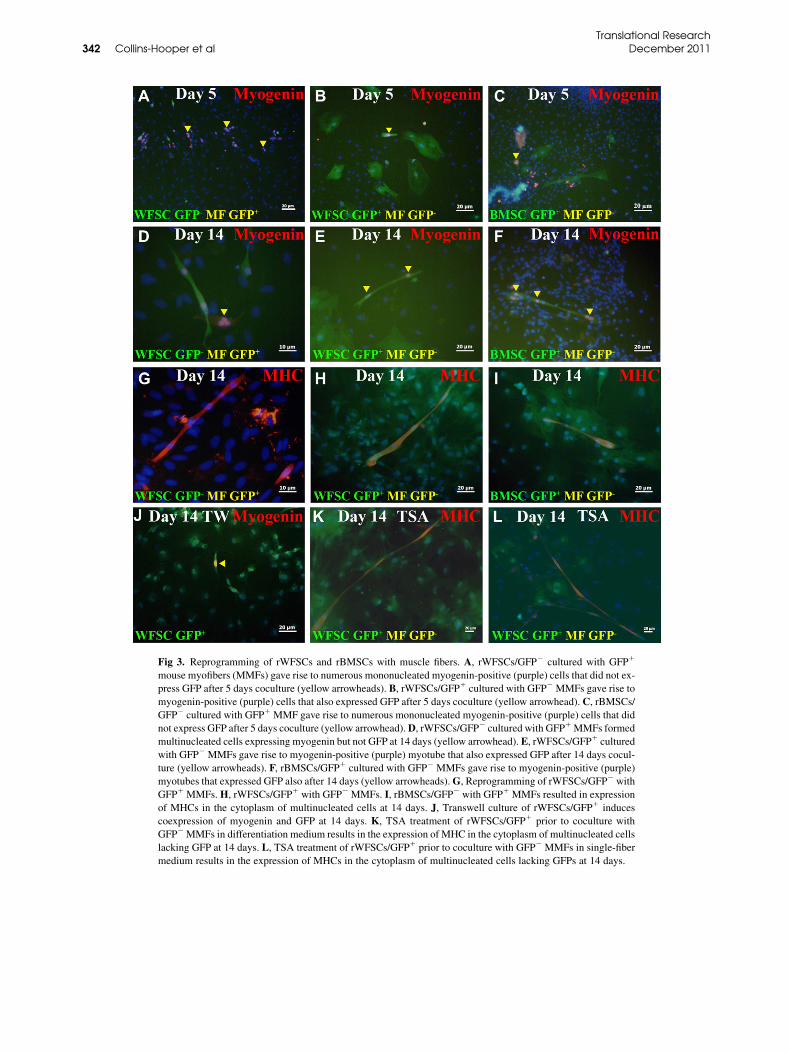
Fig 3. Reprogramming of rWFSCs and rBMSCs with muscle fibers. A, rWFSCs/GFP2 cultured with GFP1
mouse myofibers (MMFs) gave rise to numerous mononucleated myogenin-positive (purple) cells that did not ex-
press GFP after 5 days coculture (yellow arrowheads). B, rWFSCs/GFP1 cultured with GFP2 MMFs gave rise to
myogenin-positive (purple) cells that also expressed GFP after 5 days coculture (yellow arrowhead). C, rBMSCs/
GFP2 cultured with GFP1 MMF gave rise to numerous mononucleated myogenin-positive (purple) cells that did
not express GFP after 5 days coculture (yellow arrowhead).D, rWFSCs/GFP2 cultured with GFP1MMFs formed
multinucleated cells expressing myogenin but not GFP at 14 days (yellow arrowhead). E, rWFSCs/GFP1 cultured
with GFP2 MMFs gave rise to myogenin-positive (purple) myotube that also expressed GFP after 14 days cocul-
ture (yellow arrowheads). F, rBMSCs/GFP1 cultured with GFP2 MMFs gave rise to myogenin-positive (purple)
myotubes that expressed GFP also after 14 days (yellow arrowheads).G, Reprogramming of rWFSCs/GFP2 with
GFP1 MMFs.H, rWFSCs/GFP1 with GFP2 MMFs. I, rBMSCs/GFP2 with GFP1 MMFs resulted in expression
of MHCs in the cytoplasm of multinucleated cells at 14 days. J, Transwell culture of rWFSCs/GFP1 induces
coexpression of myogenin and GFP at 14 days. K, TSA treatment of rWFSCs/GFP1 prior to coculture with
GFP2MMFs in differentiation medium results in the expression of MHC in the cytoplasm of multinucleated cells
lacking GFP at 14 days. L, TSA treatment of rWFSCs/GFP1 prior to coculture with GFP2 MMFs in single-fiber
medium results in the expression of MHCs in the cytoplasm of multinucleated cells lacking GFPs at 14 days.
Translational Research342 Collins-Hooper et al December 2011

Translational ResearchVolume 158, Number 6 Collins-Hooper et al 343
accelerated into differentiation by reducing serumlevels. Previous work has shown that TSA can promotethe fusion of muscle cells.33 However, our data suggestthat this is not the main effect of TSA.We found that thenumber of myogenic nuclei increased by 90% in thepresence of TSA compared with control conditions,whereas the size of the myotubes (number of nucleiper myotube) increased by approximately 27%.Work underway in our laboratory will determine
whether readily available human WFSCs can be reprog-rammed to the skeletal myogenic lineage, and wewill as-say their efficacy in promoting repair to damagedmuscle.
We are grateful to Dr Mike Fischer of SBL (Didcot, UK) for his
continued support for academic-based research focusing on improving
human health.
REFERENCES
1. Verfaillie CM. Adult stem cells: assessing the case for pluripo-
tency. Trends Cell Biol 2002;11:502–8.
2. Jiang Y, Jahagirdar BN, Reinhardt RL, et al. Pluripotency of mes-
enchymal stem cells derived from adult marrow. Nature 2002;418:
41–9.
3. Reyes M, Lund T, Lenvik T, et al. Purification and ex vivo expan-
sion of postnatal human marrow mesodermal progenitor cells.
Blood 2001;98:2615–25.
4. Zuk PA, ZhuM, Ashjian P, et al. Human adipose tissue is a source
of multipotent stem cells. Mol Biol Cell 2002;13:4279–95.
5. Rodriguez AM, Elabd C, Delteil F, et al. Adipocyte differentiation
of multipotent cells established from human adipose tissue. Bio-
chem Biophys Res Commun 2004;315:255–63.
6. Goudenege S, Pisani DF, Wdziekonski B, et al. Enhancement of
myogenic and muscle repair capacities of human adipose-
derived stem cells with forced expression of MyoD. Mol Ther
2009;17:1064–72.
7. Hao HN, Zhao J, Thomas RL, Parker GC, Lyman WD. Fetal hu-
man hematopoietic stem cells can differentiate sequentially into
neural stem cells and then astrocytes in vitro. J Hematother
Stem Cell Res 2003;12:23–32.
8. Camussi G, Deregibus MC, Tetta C. Paracrine/endocrine mecha-
nism of stem cells on kidney repair: role of microvesicle-mediated
transfer of genetic information. Curr Opin Nephrol Hypertens
2010;19:7–12.
9. Warren L,Manos PD, Ahfeldt T, et al. Highly efficient reprogram-
ming to pluripotency and directed differentiation of human cells
with synthetic modified mRNA. Cell Stem Cell 2010;7:618–30.
10. Salvatori G, Lattanzi L, Coletta M, et al. Myogenic conversion of
mammalian fibroblasts induced by differentiating muscle cells.
J Cell Sci 1995;108:2733–9.
11. Zammit PS, Golding JP, Nagata Y, et al. Muscle satellite cells
adopt divergent fates: a mechanism for self-renewal? J Cell Biol
2004;166:347–57.
12. Otto A, Schmidt C, Luke G, et al. Canonical Wnt signalling in-
duces satellite-cell proliferation during adult skeletal muscle re-
generation. J Cell Sci 2008;121:2939–50.
13. Bunnell BA, Flaat M, Gagliardi C, Patel B, Ripoll C. Adipose-de-
rived stem cells: isolation, expansion and differentiation. Methods
2008;45:115–20.
14. SoleimaniM, Nadri S. A protocol for isolation and culture of mes-
enchymal stem cells from mouse bone marrow. Nat Protoc 2009;
4:102–6.
15. Rosenblatt JD, Lunt AI, Parry DJ, Partridge TA. Culturing satel-
lite cells from living single muscle fiber explants. In Vitro Cell
Dev Biol Anim 1995;31:773–9.
16. Alipour R, Sadeghi F, Hashemi-Beni B, et al. Phenotypic charac-
terizations and comparison of adult dental stem cells with
adipose-derived stem cells. Int J Prev Med 2010;1:164–71.
17. Zhang XM, Du F, Yang D, et al. Transplanted bone marrow stem
cells relocate to infarct penumbra and co-express endogenous pro-
liferative and immature neuronal markers in a mouse model of is-
chemic cerebral stroke. BMC Neurosci 2010;11:138.
18. Gnocchi VF, White RB, Ono Y, Ellis JA, Zammit PS. Further
characterisation of the molecular signature of quiescent and acti-
vated mouse muscle satellite cells. PLoS One 2009;4:e5205.
19. Amthor H, Otto A, Vulin A, et al. Muscle hypertrophy driven by
myostatin blockade does not require stem/precursor-cell activity.
Proc Natl Acad Sci U S A 2009;106:7479–84.
20. Davis MA, SmithMW, Chang SH, Trump BF. Characterization of
a renal epithelial cell model of apoptosis using okadaic acid and
the NRK-52E cell line. Toxicol Pathol 1994;22:595–605.
21. Moser FG, Dorman BP, Ruddle FH. Mouse-human heterokaryon
analysis with a 33258 Hoechst-Giemsa technique. J Cell Biol
1975;66:676–80.
22. Seale P, Sabourin LA, Girgis-Gabardo, et al. Pax7 is required for
the specification of myogenic satellite cells. Cell 2000;102:
777–86.
23. Aberg MA, Aberg ND, Hedbacker H, Oscarsson J, Eriksson PS.
Peripheral infusion of IGF-I selectively induces neurogenesis in
the adult rat hippocampus. J Neurosci 2000;20:2896–903.
24. Dentremont KD, Ye P, D’Ercole AJ, O’Kusky JR. Increased
insulin-like growth factor-I (IGF-I) expression during early post-
natal development differentially increases neuron number and
growth in medullary nuclei of the mouse. Brain Res Dev Brain
Res 1999;114:135–41.
25. Di Rocco G, Iachininoto MG, Tritarelli A, et al. Myogenic po-
tential of adipose-tissue-derived cells. J Cell Sci 2006;119:
2945–52.
26. Pajcini KV, Corbel SY, Sage J, Pomerantz JH, Blau HM. Transient
inactivation of Rb and ARF yields regenerative cells from postmi-
totic mammalian muscle. Cell Stem Cell 2010;7:198–213.
27. Davis RL, Cheng PF, Lassar AB, Weintraub H. The MyoD DNA
binding domain contains a recognition code for muscle-specific
gene activation. Cell 1990;60:733–46.
28. Tapscott SJ, Davis RL, Thayer MJ, et al. MyoD1: a nuclear phos-
phoprotein requiring a Myc homology region to convert fibro-
blasts to myoblasts. Science 1988;242:405–11.
29. Choi J, Costa ML, Mermelstein CS, et al. MyoD converts primary
dermal fibroblasts, chondroblasts, smooth muscle, and retinal pig-
mented epithelial cells into striated mononucleated myoblasts and
multinucleated myotubes. Proc Natl Acad Sci U S A 1990;87:
7988–92.
30. Mal A, HarterML.MyoD is functionally linked to the silencing of
a muscle-specific regulatory gene prior to skeletal myogenesis.
Proc Natl Acad Sci U S A 2003;100:1735–9.
31. Pomerantz JH, Mukherjee S, Palermo AT, Blau HM. Reprogram-
ming to amuscle fate by fusion recapitulates differentiation. J Cell
Sci 2009;122:1045–53.
32. L’Honore A, Lamb NJ, Vandromme M, et al. MyoD distal regula-
tory region contains an SRF binding CArG element required for
MyoD expression in skeletal myoblasts and during muscle regen-
eration. Mol Biol Cell 2003;14:2151–62.
33. Iezzi S, Di Padova M, Serra C, et al. Deacetylase inhibitors
increase muscle cell size by promoting myoblast recruitment
and fusion through induction of follistatin. Dev Cell 2004;6:
673–84.
Related Documents



![Skeletal Muscle-derived Hematopoietic Stem Cells: Muscular ... · injury or exercise, they initiate proliferation and give rise to daughter myogenic precursor cells [1-3]. After multiple](https://static.cupdf.com/doc/110x72/5e77ef32e6c8d136ca3ba872/skeletal-muscle-derived-hematopoietic-stem-cells-muscular-injury-or-exercise.jpg)







