Fish & Shellfish Immunology (2000) 10, 47–59 Article No. fsim.1999·0229 Available online at http://www.idealibrary.com on Effects of sea lice (Lepeophtheirus salmonis Kröyer, 1837) infestation on macrophage functions in Atlantic salmon (Salmo salar L.) AHMED MUSTAFA 1 *, CHRIS MACWILLIAMS 1 ,NICOLE FERNANDEZ 1 , KELLY MATCHETT 1 ,GARY A. CONBOY 2 AND JOHN F. BURKA 1 1 Department of Anatomy and Physiology, and 2 Department of Pathology and Microbiology, Atlantic Veterinary College, University of Prince Edward Island, Charlottetown, PEI CIA 4P3, Canada (Received 1 December 1998, accepted after revision 7 June 1999) Experiments were conducted to determine the e#ects of sea lice, Lepeophthei- rus salmonis, on non-specific defence mechanisms in Atlantic salmon, Salmo salar, by experimentally infesting hatchery-reared 1 and 2 year old post- smolts, S 1 and S 2 , with laboratory grown infective copepodids at moderate to high infection intensities ranging from 15–285 lice per fish. The e#ects of sea lice-induced stress were investigated by measuring the blood levels of cortisol and glucose as indicators of primary and secondary stress responses, and by changes in macrophage respiratory burst activity and phagocytosis as indi- cators of tertiary stress responses as well as non-specific defence mechanisms. Fish were sampled prior to sea lice infestation at day 0 and at days 3, 7, 14 and 21 post-infestation. Sea lice were at copepodid stage at day 3, at chalimus stages at days 7 and 14, and at pre-adult stage at day 21. Blood levels of cortisol and glucose were found to be significantly increased at day 21 in fish-infested with the highest levels. Macrophage respiratory burst and phago- cytic activities were found to be significantly decreased only at day 21. These results indicate that sea lice do not suppress host defence mechanisms during the earlier stages of infestation. They do have e#ects on the development of chronic stress and on the host non-specific defence mechanisms soon after the lice reach the pre-adult stage. 2000 Academic Press Key words: sea lice, Atlantic salmon, stress, macrophage, respiratory burst, phagocytosis. I. Introduction The term sea lice refers to several species of marine ectoparasitic copepods of the genera Lepeophtheirus and Caligus that commonly infest salmonids. Of these, Lepeophtheirus salmonis is responsible for serious disease outbreaks and high economic losses to salmon farmers throughout the northern hemi- sphere. Pre-adult and adult sea lice browse on the surface of salmon, eating *Corresponding author 47 1050–4648/00/010047+13 $35.00/0 2000 Academic Press

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Fish & Shellfish Immunology (2000) 10, 47–59Article No. fsim.1999·0229Available online at http://www.idealibrary.com on
Effects of sea lice (Lepeophtheirus salmonis Kröyer, 1837)infestation on macrophage functions in Atlantic salmon
(Salmo salar L.)
AHMED MUSTAFA1*, CHRIS MACWILLIAMS
1, NICOLE FERNANDEZ1,
KELLY MATCHETT1, GARY A. CONBOY
2AND JOHN F. BURKA
1
1Department of Anatomy and Physiology, and 2Department of Pathology andMicrobiology, Atlantic Veterinary College, University of Prince Edward
Island, Charlottetown, PEI CIA 4P3, Canada
(Received 1 December 1998, accepted after revision 7 June 1999)
Experiments were conducted to determine the e#ects of sea lice, Lepeophthei-rus salmonis, on non-specific defence mechanisms in Atlantic salmon, Salmosalar, by experimentally infesting hatchery-reared 1 and 2 year old post-smolts, S1 and S2, with laboratory grown infective copepodids at moderate tohigh infection intensities ranging from 15–285 lice per fish. The e#ects of sealice-induced stress were investigated by measuring the blood levels of cortisoland glucose as indicators of primary and secondary stress responses, and bychanges in macrophage respiratory burst activity and phagocytosis as indi-cators of tertiary stress responses as well as non-specific defence mechanisms.Fish were sampled prior to sea lice infestation at day 0 and at days 3, 7, 14 and21 post-infestation. Sea lice were at copepodid stage at day 3, at chalimusstages at days 7 and 14, and at pre-adult stage at day 21. Blood levels ofcortisol and glucose were found to be significantly increased at day 21 infish-infested with the highest levels. Macrophage respiratory burst and phago-cytic activities were found to be significantly decreased only at day 21. Theseresults indicate that sea lice do not suppress host defence mechanisms duringthe earlier stages of infestation. They do have e#ects on the development ofchronic stress and on the host non-specific defence mechanisms soon after thelice reach the pre-adult stage. � 2000 Academic Press
Key words: sea lice, Atlantic salmon, stress, macrophage, respiratory burst,phagocytosis.
I. Introduction
The term sea lice refers to several species of marine ectoparasitic copepods ofthe genera Lepeophtheirus and Caligus that commonly infest salmonids. Ofthese, Lepeophtheirus salmonis is responsible for serious disease outbreaksand high economic losses to salmon farmers throughout the northern hemi-sphere. Pre-adult and adult sea lice browse on the surface of salmon, eating
*Corresponding author
471050–4648/00/010047+13 $35.00/0 � 2000 Academic Press

48 A. MUSTAFA ET AL.
mucus, epidermal cells and blood, and eventually eroding the protectivesurface of the fish (Mackinnon, 1997). In severe cases, the head of the salmoncan be eroded su$ciently to expose the underlying tissue and even the skullroof (Egidius, 1985; Berland, 1993). These can lead to osmoregulatory problemsand predisposition to secondary bacterial infections and kill the fish (Woottenet al., 1982; MacKinnon, 1997).
The life cycle of Lepeophtheirus salmonis consists of 10 stages. These stagesinclude two free-swimming nauplii, one free-swimming infectious copepodid,four attached chalimus, two free-moving pre-adults, and one free-moving adult(Kabata, 1972; Pike, 1989; Johnson & Albright, 1991). After the fourthchalimus stage, the parasites are mobile and the cause of severe pathogenicitydue to their feeding activities on the fish (Grimnes & Jakobsen, 1996; Dawson,1998).
The absence of completely e#ective and safe methods for treating sea liceinfections emphasises the need to develop alternate methods. The develop-ment of such methods is limited by deficiencies in the understanding of manyaspects of the basic biology of Lepeophtheirus salmonis, especially its e#ectson host defence mechanisms. In all vertebrates, environmental stressors causeneuroendocrine and autonomic changes that modulate both non-specific andspecific defence mechanisms, which are often considered the cause of highersusceptibility of stressed individuals to disease (Ruis & Bayne, 1997). Inaquaculture, fish are exposed to stressors, such as transport, handling,marking, grading etc., on a regular basis which elicit the release of corticos-teroids from the interrenal cells and catecholamines from the chroma$n cellsby activating the hypothalamus-pituitary-interrenal axis (Schreck, 1996;Barton & Iwama, 1991). Though the immune systems of fish are sophisticatedand complex, it is believed that these stress hormones can modulatemacrophage functions (Ruis & Bayne, 1997). Correlation between increasedproduction of cortisol and glucose and decreased macrophage functions insalmonids have been well documented (Secombes, 1990; Schreck, 1996). Withrespect to sea lice, Johnson & Albright (1992) showed that coho salmonimplanted with cortisol, by intraperitoneal injection in an oil-based pellet,have a decreased inflammatory response and less epithelial hyperplasiawhen infested with L. salmonis, and increased susceptibility to the parasite.Mustafa & MacKinnon (1993) and Mustafa (1997) showed that Atlanticsalmon given cortisol implants acquired heavier infections with Caliguselongatus. While cortisol implantation experimentally stimulates some of thesecondary e#ects of stress, there have apparently been no reports on theassessment of stress on immune responses due to sea lice infestations anddevelopment in salmonids, neither have any reports compared S1 and S2
Atlantic salmon smolts. Thus, until now it has been impossible to predict whatstages of the sea lice would have e#ects on host defence mechanisms,especially in two di#erent age groups of Atlantic salmon smolts raised underaquaculture conditions.
In this study, the e#ects of sea lice infestations on host stress levels (i.e.plasma cortisol and glucose concentrations) and changes in the host non-specific defence mechanisms (i.e. respiratory burst and phagocytic activities ofmacrophages) have been examined along a sea lice development gradient

SEA LICE AND SALMON MACROPHAGE FUNCTIONS 49
in two di#erent age groups of Atlantic salmon smolts under laboratoryconditions.
II. Materials and Methods
FISH AND THEIR MAINTENANCE
The experimental design consists of two studies. The first study used 3002 year old Atlantic salmon smolts (S2), with a mean weight of 680 g and a meanlength of 37·2 cm; whilst the second study used 300 1 year old Atlantic salmonsmolts (S1), with a mean weight of 180 g and a mean length of 26·2 cm. In bothcases, Atlantic salmon smolts (Saint John river stocks) were obtained from aPrince Edward Island hatchery and randomly assigned to two di#erent groups(controls and tests) in four di#erent tanks (i.e. two control groups and two testgroups for each study). Fish were maintained in 1500 1 tanks and wereacclimated gradually to artificial seawater (Instant Ocean�, Aquarium Sys-tems, Mentor, OH, U.S.A.) over a 1 week period and were then maintained in30�2 ppt at 10�1� C for a further 2 weeks prior to sea lice infestation.Dissolved oxygen levels were monitored and maintained at �8 ppm. Thephoto-period was maintained at 14 h light: 10 h dark. Fish were fed daily tosatiation with the appropriate pelleted salmon diet to suit the fish size (CoreyFeed Mills, Frederiction, N. B., Canada) and cared for according to theguidelines of the Canadian Council on Animal Care.
SEA LICE CULTURE
Infective copepodids were grown from egg-strings removed from ovigeroussea lice collected from Atlantic salmon in aquaculture sites in the lower Bayof Fundy. Egg-strings were placed in three, 20 l white plastic buckets withsieved static sea water (27 ppt) collected from the same locality where the licewere collected. The buckets were then placed in an environmental chamber ata constant temperature of 10�2� C with gentle aeration supplied fromaquarium pumps and a 12 h light: 12 h dark cycle maintained by a timer. Theeggs hatched on day 1 and the maximum number of active copepodids wereobtained on day 12.
LABORATORY INFESTATION
The fish in the designated test tanks were infested with sea lice by addinginfective copepodids (30 000 in the first study and 20 000 in the second study) ineach of the two test tanks. During infestation the water level in all tanks waslowered to one third of the normal and maintained with decreased water flowbut constant aeration. The water outlets were screened with 70 � Nytex mesh(Valox, Fredericton, N.B., Canada) and the room was kept dark. The controltanks were treated similarly but no copepodids were added. After 21 h ofexposure, water circulation and light cycles were returned to normal.
SAMPLING
On day 0, i.e. prior to sea lice infestation, and days 3, 7, 14, and 21post-infestation, 10 fish from each group were randomly sampled. Each fish

50 A. MUSTAFA ET AL.
was collected individually with a white 70 � Nytex mesh-net and placedimmediately into a white plastic bucket containing a lethal dose of anaes-thetic (MS-222; �200 mg l�1). Each fish was measured for length and weightand bled to measure plasma levels of cortisol and glucose. Head kidneys fromeach fish were then removed aseptically for macrophage assays. Fish werethen individually bagged and sea lice were counted later from each fish as wellas from each net and bucket corresponding to that particular fish.
PLASMA CORTISOL AND GLUCOSE ASSAYS
Plasma levels of cortisol and glucose of the representative fish from eachexperimental group were analysed using validated and characterised radio-immunoassays (Coat-A-Count� RIA and Glucose Oxidase Method) by theAtlantic Veterinary College Diagnostic Services Unit.
ISOLATION OF MACROPHAGE CELLS
The head kidney samples that had been removed from fish were placed inLeibovitz-15 medium (L-15) containing 2% foetal calf serum (FCS) on ice andbrought back to the laboratory. The samples were then macerated through acoarse mesh in order to tease apart the larger fibres. The cells were centri-fuged at 1000�g, the supernatants discarded, and the cells then resuspendedin fresh L-15 containing 0·1% FCS. This process was repeated once againand adjusted to obtain a cell concentration of 106 cells ml�1. Trypan blueexclusion test was used to test for viability.
RESPIRATORY BURST ACTIVITY
The respiratory burst activity of phagocytic macrophage cells was measuredby the reduction of nitro-blue tetrazolium (NBT) by intracellular superoxideradicals produced by leucocytes stimulated with phorbol myristate acetate(PMA). Macrophage respiratory burst and phagocytic activity were measuredfollowing the methods described by Secombes (1990) and Brown et al. (1996).Briefly, aliquots of each sample (100 �l) were dropped, in duplicate, onto glassslides. The glass slides were then incubated in a moist chamber for 90 min at15� C. The slides were then rinsed in a stream of phosphate bu#ered saline(PBS), and incubated again for 15 min at 15� C with NBT (dissolved in L-15 at1 mg l�1, and PMA added at 1 �g ml�1). During this reaction NBT is reducedby O�
2 into an insoluble blue formazan. After incubation, the slides wereexamined at 100� magnification to determine the proportion of activatedcells. At least 100 cells per sample were examined and the proportion ofactivated cells was recorded.
PHAGOCYTIC ACTIVITY
The phagocytic activity of macrophage cells were evaluated using a micro-scopic counting technique as described by Mathews et al. (1990) and Brownet al. (1996). This assay determined the proportion of phagocytic cells thatwere able to take up opsonised formalin-killed bacteria, Yersinia ruckeri.

SEA LICE AND SALMON MACROPHAGE FUNCTIONS 51
Briefly, a Y. ruckeri culture was grown in the laboratory and added to slidescontaining attached macrophage cells that had been isolated from fish headkidneys and incubated for 90 min at 15� C to yield a final particle to cell ratio100:1. These slides were then incubated for another 60 min at 15� C. Followingincubation, the slides were washed with PBS, air dried, fixed in methanol andstained with Di#-Quick� (Leukostat Stain kit) for microscopic examination at100� magnification. At least 100 cells were examined to determine phagocyticcapacity (the percentage of macrophage cells containing five or more bacteria)(Enane et al., 1993).
CHEMICALS
MS-222, L-15, heparin, FCS, penicillin/streptomycin, trypan blue, NBT,PMA were purchased from Sigma Chemical Co., St. Louis, MO, U.S.A.Di#-Quick-Leukostat Stain Kit was purchased from Fisher Scientific Ltd.,Oshawa, Ont., Canada.
DATA ANALYSIS
The means and standard errors of the means were calculated for each assay.Analyses were carried out using Student’s t-test, ANOVA and post-ANOVAmultiple comparison test after checking the data for normal distribution.Di#erences were considered significant when P<0·05. All values shown in thisinvestigation are mean�standard error of means.
III. Results
Only copepodids were found on fish at day 3 post-infestation. On days 7 and14 all lice found were at chalimus stages. On day 21, all lice were pre-adults.Most of the copepodid and chalimus larvae were found attached to the gillsand fins and a few on the operculum and body surface. Pre-adults on the otherhand were mostly found on the body surface. During the first study, thenumber of lice per fish ranged from 15–285, with the mean intensity of 106.During the second study, the lice number ranged from 23–74, with the meanintensity of 52. In both studies, the prevalence was 100%.
During the first study where S2 smolts were used, plasma cortisol concen-trations increased following sea lice infestation and remained significantlyelevated (P<0·05) in the infested groups throughout the experiment, except onday 14. The highest level of plasma cortisol was recorded on day 21 (Fig. 1).During the second study where S1 smolts were used, plasma cortisol concen-tration increased significantly (P<0·05) in the infested groups on day 7post-infestation and remained elevated for the rest of the experimental periodwith the highest level on day 21 (Fig. 2).
Plasma glucose concentrations also increased with time post-infestation insea lice infested groups, both in the case of S2 and S1 smolts, with control

52 A. MUSTAFA ET AL.
250
00
Day of sampling
Pla
sma
cort
isol
con
cen
trat
ion
(n
mol
l–1)
143 7
200
150
100
50
21
ControlInfested
*
*
*
Fig. 1. Plasma cortisol concentrations in control and sea lice infested Atlantic salmonsmolts (S2). *Significantly di#erent from controls.
100
00
Day of sampling
Pla
sma
cort
isol
con
cen
trat
ion
(n
mol
l–1)
143 7
75
50
25
21
ControlInfested
*
*
*
Fig. 2. Plasma cortisol concentrations in control and sea lice infested Atlantic salmonsmolts (S1). *Significantly di#erent from controls.
groups remaining relatively constant (Figs 3 and 4, respectively). However,glucose concentrations in both infested and control groups appeared to fallwithin the ‘ normal ’ reference range values (3·65–7·40 mmol l�1) for Atlanticsalmon in seawater (Standards developed by Diagnostic Services at theAtlantic Veterinary College).
In both studies, respiratory burst activity remained unchanged until day 14between infested and control groups but a significant suppression (P<0·05)was observed in the infested groups on day 21 (Figs 5 and 6). Phagocyticcapacity (>5 intracellular bacteria) showed a similar pattern to respiratoryburst activity with a significant suppression (P<0·05) on day 21 in the infestedgroups compared to that of controls (Figs 7 and 8).

SEA LICE AND SALMON MACROPHAGE FUNCTIONS 53
5
00
Day of sampling
Pla
sma
glu
cose
con
cen
trat
ion
(m
mol
l–1)
143 7
4
3
2
1
21
ControlInfested
*
*
*
Fig. 3. Plasma glucose concentrations in control and sea lice infested Atlantic salmonsmolts (S2). *Significantly di#erent from controls.
6
00
Day of sampling
Pla
sma
glu
cose
con
cen
trat
ion
(m
mol
l–1)
143 7
4
3
2
1
21
ControlInfested
*
*5
Fig. 4. Plasma glucose concentrations in control and sea lice infested Atlantic salmonsmolts (S1). *Significantly di#erent from controls.
IV. Discussion
The prevalence of sea lice infestation in these studies was 100%. Betweenday 0 and day 14, most sea lice were at copepodid and chalimus stages. Duringthese stages, most lice were recorded from gills and fins, and during mobilepre-adult stages, most lice were recorded from body surfaces. These findingsare similar to those of Johnson & Albright (1991, 1992), Grimnes & Jakobsen(1996), Dawson et al. (1997) and Dawson (1998). On the body surface, licewere commonly found on the head, external operculum, areas betweendorsal and adipose fins, and peri-anal regions. In most fish, the head and the

54 A. MUSTAFA ET AL.
100
00
Day of sampling
Cel
ls p
osit
ive
for
resp
irat
ory
burs
t (%
)
143 7
75
50
25
21
ControlInfested
*
Fig. 5. Respiratory burst activity of macrophage cells isolated from head kidneys ofAtlantic salmon smolts (S2). *Significantly di#erent from controls.
100
00
Day of sampling
Cel
ls p
osit
ive
for
resp
irat
ory
burs
t (%
)
143 7
50
25
21
ControlInfested
*
Fig. 6. Respiratory burst activity of macrophage cells isolated from head kidneys ofAtlantic salmon smolts (S1). *Significantly di#erent from controls.
external opercular regions were preferred. The preference of these regions bythe mobile stages is likely feeding related, because these areas have thinepidermis with no or fewer scales (Wootten et al., 1982; Jónsdóttir et al., 1992).
Details on the changes in blood parameters of S2 smolts in response to sealice have been submitted elsewhere (Bowers et al., 1999). Briefly, plasmacortisol concentration, an indicator of primary stress, increased significantlyin the sea lice-infested fish, in both S2 and S1, with the highest levels on day 21when most lice were at pre-adult stages. Plasma glucose concentration, anindicator of secondary stress, also increased similarly with the highest levelon day 21. On the same day (i.e. day 21) macrophage respiratory burst and
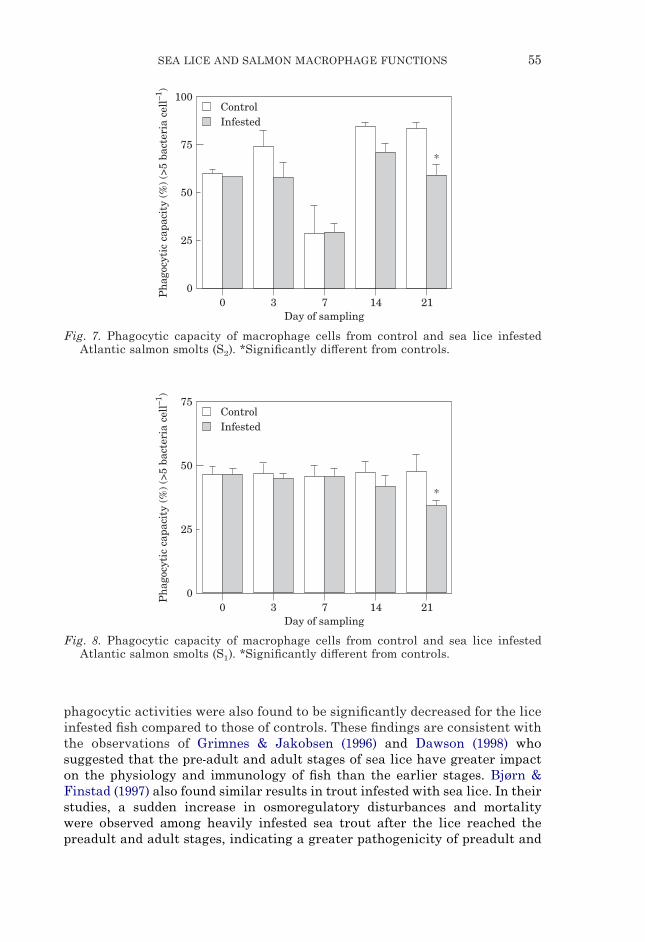
SEA LICE AND SALMON MACROPHAGE FUNCTIONS 55
100
00
Day of sampling
Ph
agoc
ytic
cap
acit
y (%
) (>
5 ba
cter
ia c
ell–1
)
143 7
50
25
21
ControlInfested
*75
Fig. 7. Phagocytic capacity of macrophage cells from control and sea lice infestedAtlantic salmon smolts (S2). *Significantly di#erent from controls.
75
00
Day of sampling
Ph
agoc
ytic
cap
acit
y (%
) (>
5 ba
cter
ia c
ell–1
)
143 7
50
25
21
ControlInfested
*
Fig. 8. Phagocytic capacity of macrophage cells from control and sea lice infestedAtlantic salmon smolts (S1). *Significantly di#erent from controls.
phagocytic activities were also found to be significantly decreased for the liceinfested fish compared to those of controls. These findings are consistent withthe observations of Grimnes & Jakobsen (1996) and Dawson (1998) whosuggested that the pre-adult and adult stages of sea lice have greater impacton the physiology and immunology of fish than the earlier stages. Bjørn &Finstad (1997) also found similar results in trout infested with sea lice. In theirstudies, a sudden increase in osmoregulatory disturbances and mortalitywere observed among heavily infested sea trout after the lice reached thepreadult and adult stages, indicating a greater pathogenicity of preadult and

56 A. MUSTAFA ET AL.
adult lice than the chalimus stages. When salmon lice reach pre-adultstage, they change from an immobile to mobile phase. Transformation ofchalimus to preadult induces significant change in louse activities anddistribution on the host and likely explains the sudden increase in thepathogenicity.
The cortisol elevation on days 3 and 7 in the case of S2 smolts and on days7 and 14 in the case of S1 smolts, even though significant compared to those ofcontrols, were at levels comparable to levels found in acutely stressed browntrout (Pickering & Pottinger, 1989) and other teleost fish (Barton & Iwama,1991). Plasma glucose concentrations, which similarly increased over controlspost infestation, were also within the normal range. The sedentary nature ofchalimus larvae may explain these results. During the chalimus stages, licestay attached to the gill lamellae and to scales of the fish, and since theyremain attached, the area of skin they feed on is localised and the impactlimited (Pike, 1989; Bjørn & Finstad, 1997).
Since fish are poikilothermic in nature, specific immune responses are oftendelayed and smaller than in other vertebrates (Blazer, 1991). The first, andoften most important, responses of fish to infectious agents are thereforenonspecific. These include various soluble and cellular factors, enzymes,other proteins and cells for phagocytosis, particularly macrophages. Duringphagocytosis, fish macrophages respond with an oxidative burst when theyencounter an appropriate stimulus. They convert molecular oxygen to anumber of highly reactive oxygen intermediates and damaging organic mol-ecules (Bayne & Levy, 1991). Macrophage functions, as mentioned earlier, canbe a#ected by numerous environmental and physiological factors, especiallystress. In their studies with rainbow trout, Narnaware et al., (1994) showed adepression of phagocytic activities within 3 h of an acute stress but could notcorrelate the depression of phagocytosis with plasma cortisol concentrations.They concluded that the depression due to acute stress was caused bycatecholamines, not corticosteroids which actually have e#ects only in thelonger term. This is consistent with the studies by Barton & Iwama (1991).Persistence of stressors generally causes biological tolerance limits to beexceeded, resulting in a maladaptation with adverse physiological and behav-ioral consequences. Prolonged increase in cortisol concentration can induce ageneralised immune suppression (Maule et al., 1989; Schreck, 1996; Ruis &Bayne, 1997). Thus, the decrease in macrophage respiratory burst and phago-cytic activities in the present studies could be a consequence of increasedcortisol concentrations over a long period. It is more likely that fish wereacutely stressed, perhaps due to a discomfort caused by chalimus larvae(Bjørn & Finstad, 1997). During the third week stress became chronic as thesealice increased size and matured into mobile stages, which eventuallysuppressed the immune system, reducing the respiratory burst and phagocyticactivities of macrophage cells.
Atlantic salmon in nature usually take a minimum of 2 years to become asmolt. In aquaculture, this period can be reduced to 1 year as a result ofenvironmental manipulation and optimum feeding with high energy diets.There has been speculation that commercially raised S1 smolts would performless satisfactorily than naturally occurring S2 smolts when released in sea

SEA LICE AND SALMON MACROPHAGE FUNCTIONS 57
water, due to their di#erences in physiology, behavior and genetics (Saunders,1991). Such speculation contradicts opinions of other researchers who believethat such di#erences are not important when fish are put in sea cages, fed welland looked after during their marine phase. In the present studies, thereappeared to be no di#erence between the two age groups in terms of bloodparameters (i.e. plasma cortisol and glucose concentrations) or in macrophagefunctions (i.e. respiratory burst and phagocytic activities). The only di#erencein plasma cortisol concentrations on day 21, i.e. the lower cortisol concen-tration in S1 smolts, could be attributed, at least in part, to a lower infectionintensity of sea lice.
In conclusion, these studies indicate that sea lice induce a stress responseand immune suppression in their fish host, having greater e#ect during thelater stages of their life cycle when they are mobile and able to cause the mostdamage. Other studies have found similar results and demonstrate that laterlife stages are more detrimental physically and have a high physiological andimmunological impact on the fish (Pike, 1989; MacKinnon, 1997; Grimnes &Jakobsen, 1996; Bjørn & Finstad, 1997). Further study will allow this infor-mation to be integrated into attempts to develop alternate methods to controlsea lice.
We thank Joanne Bowers, Pablo Gonzalez, Cheryl Wartman, Margaret Horne and DrDavid Sims for their help in collecting, counting and sampling the fish and sea lice. Wealso thank Dr Laura Brown and Dr Simon Jones for useful discussion and adviceregarding macrophage assays. We are grateful to Dr Stewart Johnson and Dr Neil Rossfor their advice on the experimental design and for constructive comments on earlierdrafts of this manuscript. This research was funded by a grant from the NSERC/NRCPartnership Program. The Salmon Health Consortium also provided useful advice andfinancial support.
References
Barton, B. A. & Iwama, G. K. (1991). Physiological changes in fish from stress inaquaculture with emphasis on the response and e#ects of corticosteroids.Annual Reiew of Fish Diseases 1, 3–26.
Bayne, C. J. & Levy, S. (1991). The respiratory burst of rainbow trout, Oncorhynchusmykiss (Walbaum), phagocytes is modulated by sympathetic neurotransmittersand the ‘neuro’ peptide ACTH. Journal of Fish Biolology 38, 609–619.
Berland, B. (1993). Salmon lice on wild salmon (Salmo salar) in western Norway. InPathogens of wild and farmed fish: sea lice. (Boxshall, G. A. & Defaye, D., eds),Ellis Horwood, West Sussex, UK. pp 179–187.
Bjørn, P. A. & Finstad, B. (1997). The physiological e#ects of salmon lice infection onsea trout post smolts. Nordic Journal of Freshwater Research 73, 60–72.
Blazer, V. S. (1991). Piscine macrophage functions and nutritional influences: a review.Journal of Aquatic Animal Health 3, 77–86.
Bowers, J. M., Mustafa, A., Speare, D. J., Conboy, G. A., Brimacombe, M., Sims, D. &Burka, J. F. (1999). The e#ects of a single experimental challenge of sea lice,Leophtheirus salmonis, on the stress response of Atlantic salmon, Salmo salar.Journal of Fish Diseases (In press).
Brown, L. L., Iwama, G. K. & Evelyn, T. P. T. (1996). The e#ects of early exposure ofCoho salmon (Oncorhynchus kisutch) eggs to the p57 protein of Renibacteriumsalmoninarum on the development of immunity to the pathogen. Fish & ShellfishImmunology 6, 149–165.

58 A. MUSTAFA ET AL.
Dawson, L. H. J., Pike, A. W., Houlihan, D. F. & McVicar, A. H. (1997). Comparison ofthe susceptibility of sea trout (Salmo trutta L.) And Atlantic salmon (Salmo salarL.) To sea lice (Lepeophtheirus salmonis (Kröyer, 1837)) infections. ICES Journalof Marine Sciences 54, 1129–1139.
Dawson, L. H. J. (1998). The physiological e#ects of salmon lice (Lepeophtheirussalmonis) infections on returning post-smolt sea trout (Salmo trutta L.) Inwestern Ireland, 1996. ICES Journal of Marine Sciences 55, 193–200.
Egidius, E. (1985). Salmon lice, Lepeophtheirus salmonis. ICES identification leaflets fordisease and parasites of fish and shellfish. Leaflet No. 26. pp 1–4.
Enane, N. A., Frenkel, K., O’Connor, J. M., Squibb, K. S. & Zeliko#, J. T. (1993).Biological markers of macrophage activation: application for fish phagocytes.Immunology 80, 68–72.
Grimnes, A. & Jakobsen, P. J. (1996). The physiological e#ects of salmon lice infectionon post-smolt of Atlantic salmon. Journal of Fish Biology 48, 1179–1194.
Jónsdóttir, H., Bron, J. E., Wootten, R., & Turnbull, J. F. (1992). The histopathologyassociated with the preadult stages of Lepeophtheirus salmonis on the Atlanticsalmon, Salmo salar L. Journal of Fish Diseases 13, 303–310.
Johnson, S. C. & Albright, L. J. (1991). Development, growth, and survival ofLepeophtheirus salmnonis (Copepoda: Caligidae) under laboratory conditions.Journal of the Marine Biology Association, U.K. 71, 425–436.
Johnson, S. C. & Albright, L. J. (1992). E#ects of cortisol implants on the susceptibilityand the histopathology of the responses of naive coho salmon Oncorhynchuskisutch to experimental infection with Lepeophtheirus salmonis (Copepoda:Caligidae). Diseases of Aquatic Organisms 14, 195–205.
Kabata, Z. (1972). Developmental stages of Caligus clemensi (Copepoda: Caligidae).Journal of Fisheries Research Board of Canada 29, 1571–1593.
MacKinnon, B. M. (1997). Sea lice: a review. World Aquaculture 28, 5–10.Mathews, E. S., Warinner, J. E. & Weeks, B. A. (1990). Assays of immune function in
fish macrophage. In: Techniques in Fish Immunology. (Stolen, J. C., Fletcher,T. C., Anderson, D. P., Robertson, B. S., vanMuiswinkle, W. B. eds), SOSPublications: NJ pp. 155–163.
Maule, A. G., Tripp, R. A., Kaattari, S. L. & Schreck, C. B. (1989). Stress alters immunefunction and disease resistance in chinook salmon (Oncorhynchus tshawtscha).Journal of Endocrinology 120, 135–142.
Mustafa, A. & MacKinnon, B. M. (1993). Sea lice and salmonids: preliminary obser-vations on modulation of the e#ects of host stress using iodine. Bulletin of theAquaculture Association of Canada 93, 99–101.
Mustafa, A. (1997). Host factors important in determining infection intensity with thesea louse, Caligus elongatus Nordmann, 1932, in Atlantic salmon, Salmo salar L.and Arctic charr, Salvelinus alpinus (L.). Ph.D. Thesis, University of NewBrunswick, Fredericton, N. B., Canada, 175 pp.
Narnaware, Y. K., Baker, B. I. & Tomlinson, M. G. (1994). The e#ect of various stresses,corticosteroids and adrenergic agents on phagocytosis in the rainbow trout,Oncorhynchus mykiss. Fish Physiology and Biochemistry 13, 31–40.
Pickering, A. D. & Pottinger, T. G. (1989). Stress responses and disease resistance insalmonid fish: e#ects of chronic elevation of plasma cortisol. Fish Physiologyand Biochemistry 7, 253–258.
Pike, A. W. (1989). Sea lice-major pathogens of farmed Atlantic salmon. ParasitologyToday 5, 291–297.
Ruis, M. A. W. & Bayne, C. J. (1997). E#ects of acute stress on blood clotting and yeastkilling by phagocytes of rainbow trout. Journal of Aquatic Animal Health 9,190–195.3.
Saunders, R. L. (1991). Stresses encounter during production of Atlantic salmon smolts.In Stress in salmonid aquaculture workshop proceedings. University of NewBrunswick, NB Department of Fisheries and Aquaculture and NB Research andProductivity Council. pp. 12–17.

SEA LICE AND SALMON MACROPHAGE FUNCTIONS 59
Schreck, C. B. (1996). Immunomodulation: endogenous factors. In: Fish Physiology –Vol 15. Iwama, G. K., Nakanishi, T., eds), Organisms pathogen and environment.Academic Press, London, pp. 311–337.
Secombes, C. J. (1990). Isolation of salmonid macrophages and analysis of their killingactivity. In: Techniques in Fish Immunology (Stolen J. C., Fletcher, T. C.,Anderson, D. P., Robertson, B. S., vanMuiswinkle, W. B. eds), SOS Publications.NJ. pp. 137–154.
Wootten, R., Smith, J. W. & Needham, E. A. (1982). Aspects of the biology of theparasitic copepods Lepeophtheirus salmonis and Caligus elongatus on farmedsalmonids, and their treatment. Proceedings of the Royal Society, Edinburgh. 81,185–197.
Related Documents