Acta Biologica Hungarica 61(4), pp. 401–410 (2010) DOI: 10.1556/ABiol.61.2010.4.4 0236-5383/$ 20.00 © 2010 Akadémiai Kiadó, Budapest EFFECTS OF ROTENONE AND OTHER MITOCHONDRIAL COMPLEX I INHIBITORS ON THE BRINE SHRIMP ARTEMIA ÁGNES VEHOVSZKY, 1 * HENRIETTE SZABÓ, 1 A. ÁCS, 2 J. Győri 1 and ANNA FARKAS 1 1 Balaton Limnological Research Institute, Hungarian Academy of Sciences, P. O. Box 35, H-8237 Tihany, Hungary 2 Department of Earth and Environmental Sciences, University of Pannonia, P. O. Box 158, H-8201 Veszprém, Hungary (Received: February 22, 2010; accepted: March 16, 2010) * Corresponding author; e-mail: [email protected] Brine shrimp (Artemia) nauplii was used to asses the toxicity of rotenone, MPTP (1-methyl-4-phenyl- 1,2,3,6-tetrahydropyridine), MPP + (1-methyl-4-phenylpyridinium) and the effect of L-DOPA co-treat- ment with rotenone. Rotenone had a dose dependent effect on mortality (LC 50 : 0.37 ± 0.04 µM mean ± SE, n = 24), while MPTP and MPP + proved to be toxic in millimolar range (LC 50 : 0.21 ± 0.09 mM and 0.20 ± 0.08 mM, respectively, n = 4). L-DOPA (50–200 µM) co-treatment increased the survival of the rotenone-treated animals (LC 50 : 0.51 ± 0.23 µM, 1.03 ± 0.66 µM, and 0.76 ± 0.52 µM, respectively). In the whole body tissue homogenates of Artemia, sublethal (up to 0.3 µM) concentrations of rotenone increased the glutathione S-transferase (GST) activity by up to 50 about percent (LC 50 : 53.3 ± 6.8 nM/min/mg pro- tein, against 34.7 ± 3.6 nM/min/mg protein, n = 4). Nauplii treated in 100 mM L-DOPA and rotenone together showed further increase of GST activity all across the range of rotenone concentrations. These results on Artemia nauplii show similarities with other animal models, when complex I inhibitors were tested. Biochemical measurements suggest a protective role of L-DOPA by increasing the GST activity as part of the intracellular defences during toxin-evoked oxidative stress. Keywords: Artemia – rotenone – MPP – MPTP + – complex I – glutathione S-transferase – L-DOPA INTRODUCTION Rotenone, a member of an isoflavonoid family is generally used as a pesticide [20], and also used for lake and pond management, as fishes show the lowest sensitivity to rotenone (LC 50 : 5–100 µg/l; reviewed by Ling) [20]. Moreover, in several pharmaco- logical and toxicological studies rotenone has also been used as a highly potent refer- ence complex I inhibitor [8, 10, 30]. Complex I inhibitors affect NADH-ubiquinone reductase, the first and largest enzyme of the five complexes of the mitochondrial respiratory electron transport chain, and also a major source of reactive oxygen species (free radicals) during cel- lular respiration. When complex I functions are impaired, the oxidizing effect of free radicals (generally regarded as oxidative stress) triggers a cascade of degenerative intracellular processes, potentially leading to cell death [27, 32]. In several human 04veho.indd 401 11/10/2010 1:40:07 PM

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Acta Biologica Hungarica 61(4), pp. 401–410 (2010)DOI: 10.1556/ABiol.61.2010.4.4
0236-5383/$ 20.00 © 2010 Aka dé mi ai Ki adó, Bu da pest
EffEcts of rotEnonE and othEr mitochondrial complEx i
inhibitors on thE brinE shrimp ArtemIAÁgnes VehoVszky,1* henriette szabó,1 a. Ács,2
J. Győri1 and anna Farkas1
1 balaton limnological research institute, hungarian academy of sciences, p. o. box 35, h-8237 tihany, hungary
2 department of Earth and Environmental sciences, University of pannonia, p. o. box 158, h-8201 Veszprém, hungary
(received: february 22, 2010; accepted: march 16, 2010)
* corresponding author; e-mail: [email protected]
brine shrimp (Artemia) nauplii was used to asses the toxicity of rotenone, mptp (1-methyl-4-phenyl- 1,2,3,6-tetrahydropyridine), mpp+ (1-methyl-4-phenylpyridinium) and the effect of l-dopa co-treat-ment with rotenone. rotenone had a dose dependent effect on mortality (lc50: 0.37 ± 0.04 µm mean ± sE, n = 24), while mptp and mpp+ proved to be toxic in millimolar range (lc50: 0.21 ± 0.09 mm and 0.20 ± 0.08 mm, respectively, n = 4). l-dopa (50–200 µm) co-treatment increased the survival of the rotenone-treated animals (lc50: 0.51 ± 0.23 µm, 1.03 ± 0.66 µm, and 0.76 ± 0.52 µm, respectively). in the whole body tissue homogenates of Artemia, sublethal (up to 0.3 µm) concentrations of rotenone increased the glutathione s-transferase (Gst) activity by up to 50 about percent (lc50: 53.3 ± 6.8 nm/min/mg pro-tein, against 34.7 ± 3.6 nm/min/mg protein, n = 4). nauplii treated in 100 mm l-dopa and rotenone together showed further increase of Gst activity all across the range of rotenone concentrations. these results on Artemia nauplii show similarities with other animal models, when complex i inhibitors were tested. biochemical measurements suggest a protective role of l-dopa by increasing the Gst activity as part of the intracellular defences during toxin-evoked oxidative stress.
Keywords: Artemia – rotenone – mpp – mptp+ – complex i – glutathione s-transferase – l-dopa
introdUction
Rotenone, a member of an isoflavonoid family is generally used as a pesticide [20], and also used for lake and pond management, as fishes show the lowest sensitivity to rotenone (lc50: 5–100 µg/l; reviewed by Ling) [20]. Moreover, in several pharmaco-logical and toxicological studies rotenone has also been used as a highly potent refer-ence complex I inhibitor [8, 10, 30].
Complex I inhibitors affect NADH-ubiquinone reductase, the first and largest enzyme of the five complexes of the mitochondrial respiratory electron transport chain, and also a major source of reactive oxygen species (free radicals) during cel-lular respiration. When complex i functions are impaired, the oxidizing effect of free radicals (generally regarded as oxidative stress) triggers a cascade of degenerative intracellular processes, potentially leading to cell death [27, 32]. In several human
04veho.indd 401 11/10/2010 1:40:07 PM

402 Ágnes VehoVszky et al.
Acta Biologica Hungarica 61, 2010
neuromuscular and neurodegenerative disorders oxidative damage and the resulting mitochondrial failure is suggested to be involved (reviewed by fukui and moraes) [13]. For example, this provides a hypothesis for the selective death of dopaminergic neurons in substantia nigra of midbrain in Parkinson’s disease (PD) [22]. Experimentally, complex i inhibitors: rotenone, mptp (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) and mpp+ (1-methyl-4-phenylpyridinium) are often used to evoke the behavioral and cellular syndromes of parkinson’s disease both in verte-brates and invertebrates [3, 7, 18]. Therapeutic tools for both Parkinson’s patients and experimental rotenone models include preventing the disease-induced dopamine loss by adding the dopaminergic precursor levodopa (L-DOPA) [23, 28].
the glutathione s-transferase (Gst) family of enzymes is part of the intracellular defences when oxidative stress in involved by using the antioxidant glutathione (Gsh) molecule as its substrate to protect the cellular macromolecules from the oxidazing effect of free radicals (see rev. by Eaton and Bammler) [9]. One of the earliest detectable changes in the parkinsonian brain is the decreased glutathione level in substantia nigra [5], and in dopaminergic cell culture glutathione depletion results in a selective decrease in mitochondrial complex I activity [17]. GST enzyme, however, has a neuroprotective role, including the cellular symptoms of parkinson’s disease [2]. GST enzyme activity thus seems to be a good enzymatic marker to detect oxidative stress developed either endogenously or exogenously.
aquatic animals, living in water, get direct contact to harmful substances in their natural environment, as the toxic agents can be absorbed through the skin as well as swallowed during feeding. among aquatic invertebrates, specimens of the Artemia genus (both larvae and adults) represent a cost effective model organism for toxico-logical testing [11, 26, 33]. Biochemical analysis of the whole tissue preparations of Artemia samples, moreover, provides a straightforward assay of possible alterations in intracellular biomarkers (including oxidative stress enzymes) following the expo-sure to toxic substances [25, 34].
in our experiments we used 24-hours-old nauplius larvae of the brine shrimp Artemia to asses the toxic effect of complex i inhibitor neurotoxins: rotenone, mptp (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine), mpp+ (1-methyl-4-phenylpyridin-ium) and the effect of the dopaminergic precursor l-dopa. the rotenone-treated whole body tissue samples of Artemia were used for further testing the possible changes in the Gst enzymatic activity, involved in biotransformation of toxic sub-stances.
matErials and mEthods
Drugs, solutions
stock solution (10 mm) of rotenone was made in 1 ml dimethyl sulfoxide (dmso), and serial dilutions were made in distilled water containing 3% sodium chloride (our “standard brine”) up to the test (0–2 µM) concentration of rotenone. The final DMSO
04veho.indd 402 11/10/2010 1:40:07 PM

Complex I inhibitor effects in artemia 403
Acta Biologica Hungarica 61, 2010
concentration did not exceed 0.01 percent in any of the rotenone-containing test solu-tions. l-dopa, mptp and mpp+ test solutions were also freshly made in standard brine. prior to co-treatment experiments the rotenone (dmso) containing stock solu-tion was further diluted by salt water containing 50–200 mm l-dopa. all drugs were purchased from siGma.
Brine shrimp (artemia) bioassay
cyst-based mortality tests were carried out on newly hatched (24 hours) Artemia nauplii in 24-well polystyrene culture plates. cysts were obtained from a pet shop (novotemia, product no 730901 00 Jbl, Germany) and hydrated at 4 °c for over-night in 100 ml standard brine. next day, the container was put in a thermostat (26 ± 1 °c, continuous aeration) for 24 hours. the freshly hatched Artemia nauplii (about 60–80 individuals) were separated from their stock cultures using a 100 µl Gilson pipette and added into the wells of the culture plate, containing 0.5 ml of con-trol solution (standard brine) or test solution (rotenone, l-dopa, mptp or mpp+
diluted in standard brine). the bioassay plates were then kept on standard conditions (26 °c, continuous aeration) for another 24 hours, and the number of dead animals was counted in each well under dissecting microscope. the mortality data of treated samples were corrected by the control data (mortality in standard brine) (abbott’s formula [16]) and the median lethal concentration (LC50) was determined by finney’s probit analysis [12]. Four replicate test series were used in each individual experi-ment.
enzymatic assay of glutathione S-transferase (GSt) activity
tissue homogenates of the whole body samples of Artemia larvae were prepared in ice cold phosphate buffer (0.1 m, ph 7.4) by grinding and the supernatant collected after centrifugation (5 min at 2500× g at 4 °c). Gst activity was measured spectro-photometrically using 10 mm 1-chloro-2,4-dinitrobenzene (cdnb) as a substrate and 10 mm reduced glutathion (Gsh) in potassium phosphate buffer (0.1 m, ph 6.5), by the method of Habig et al. [14]. The absorbance of the samples was measured at a wavelength of 340 nm every 20 s for 5 min using a Wallace 1420 Victor³™ (perkin Elmer co.) microplate reader. the enzymatic activity was calculated from the slope of the absorbance curve and expressed as nmoles of formed product per minute per milligram protein. the protein concentrations of samples were determined by the Bradford method [6] adapted to microplates, using bovine serum albumine as protein standard (sigma®, Germany) at a wavelength of 595 nm.
04veho.indd 403 11/10/2010 1:40:07 PM
404 Ágnes VehoVszky et al.
Acta Biologica Hungarica 61, 2010
Statistical analysis
Data are presented as mean ± SE, and significance was calculated by using Student’s t-test.
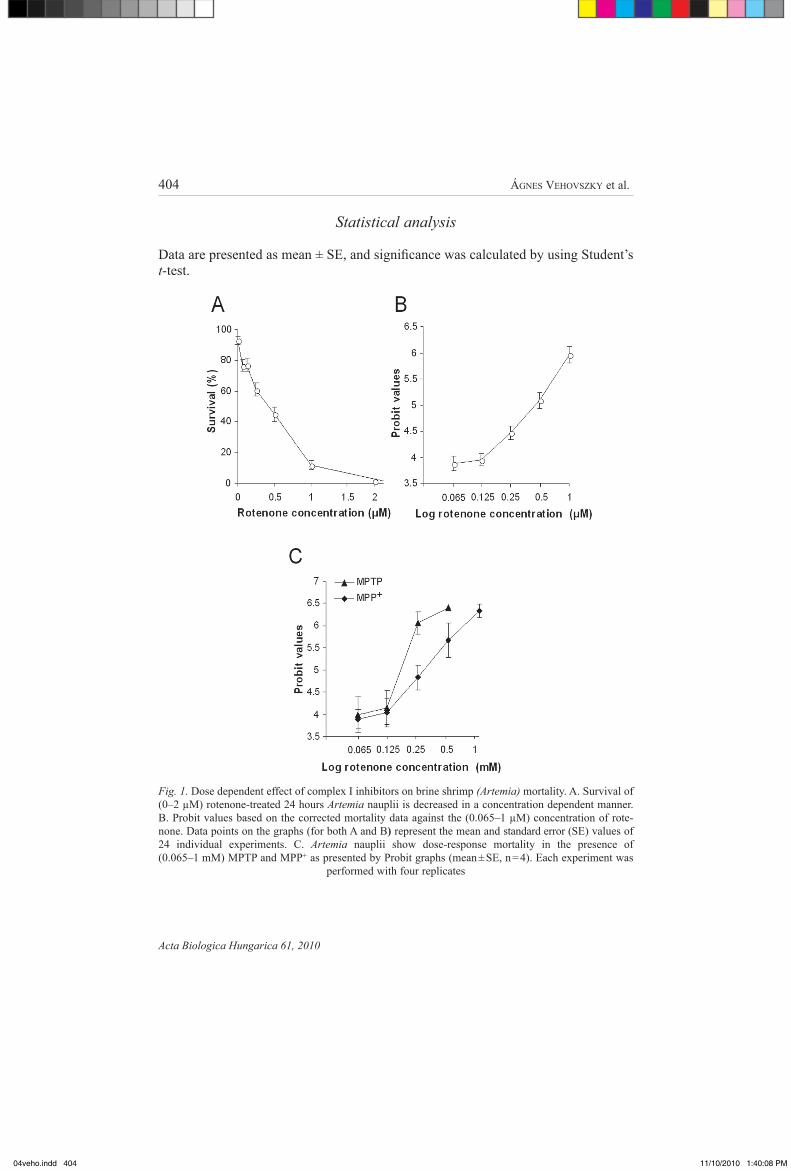
Fig. 1. dose dependent effect of complex i inhibitors on brine shrimp (Artemia) mortality. a. survival of (0–2 µm) rotenone-treated 24 hours Artemia nauplii is decreased in a concentration dependent manner. b. probit values based on the corrected mortality data against the (0.065–1 µm) concentration of rote-none. data points on the graphs (for both a and b) represent the mean and standard error (sE) values of 24 individual experiments. c. Artemia nauplii show dose-response mortality in the presence of (0.065–1 mm) mptp and mpp+ as presented by probit graphs (mean ± sE, n = 4). Each experiment was
performed with four replicates
04veho.indd 404 11/10/2010 1:40:08 PM
Complex I inhibitor effects in artemia 405
Acta Biologica Hungarica 61, 2010
rEsUlts
artemia acute toxicity tests
in standard brine, about 90 percent of the animals were hatched then transferred to the test solutions (rotenone, mptp, mpp+, l-dopa, or standard brine) for the next 24 hours (see: materials and methods).
rotenone (0.1–2 µm) killed the 24 hours old Artemia nauplii in a concentration dependent manner (fig. 1a, b). summarizing all data of mortality tests in rotenone the median lethal concentration (lc50) value was found to be 0.37 ± 0.04 µm (mean ± sE, n = 24), equivalent to 0.146 mg/l rotenone in water.
the other two complex i inhibitors tested (mptp and mpp+) also proved to be toxic to Artemia nauplii in a dose-dependent manner (fig. 1c), but to reach the lc50 value a much higher concentration was required for both mptp and mpp+ (0.21 ± 0.09 mm, and 0.20 ± 0.08 mm, respectively, n = 4) compared to rotenone solution (0.37 ± 0.04 µm).
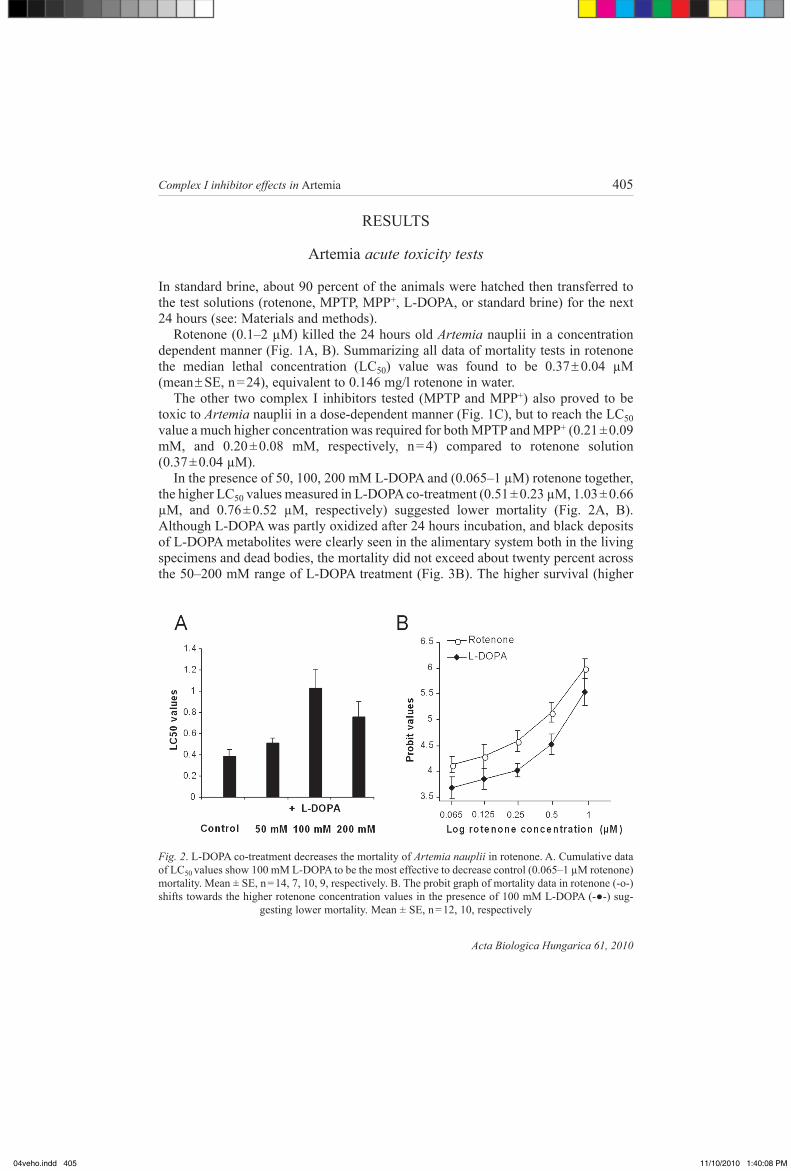
in the presence of 50, 100, 200 mm l-dopa and (0.065–1 µm) rotenone together, the higher lc50 values measured in l-dopa co-treatment (0.51 ± 0.23 µm, 1.03 ± 0.66 µm, and 0.76 ± 0.52 µm, respectively) suggested lower mortality (fig. 2a, b). although l-dopa was partly oxidized after 24 hours incubation, and black deposits of l-dopa metabolites were clearly seen in the alimentary system both in the living specimens and dead bodies, the mortality did not exceed about twenty percent across the 50–200 mm range of l-dopa treatment (fig. 3b). the higher survival (higher
Fig. 2. l-dopa co-treatment decreases the mortality of Artemia nauplii in rotenone. a. cumulative data of lc50 values show 100 mm l-dopa to be the most effective to decrease control (0.065–1 µm rotenone) mortality. mean ± sE, n = 14, 7, 10, 9, respectively. b. the probit graph of mortality data in rotenone (-o-) shifts towards the higher rotenone concentration values in the presence of 100 mM L-DOPA (-●-) sug-
gesting lower mortality. mean ± sE, n = 12, 10, respectively
04veho.indd 405 11/10/2010 1:40:08 PM
406 Ágnes VehoVszky et al.
Acta Biologica Hungarica 61, 2010
lc50 values) after co-treatment of l-dopa and rotenone suggests a protective role of l-dopa against rotenone toxicity. this protective effect of l-dopa was also dem-onstrated by the parallel shift of the dose-response curves calculated by probit analy-sis of the mortality data (fig. 2b).
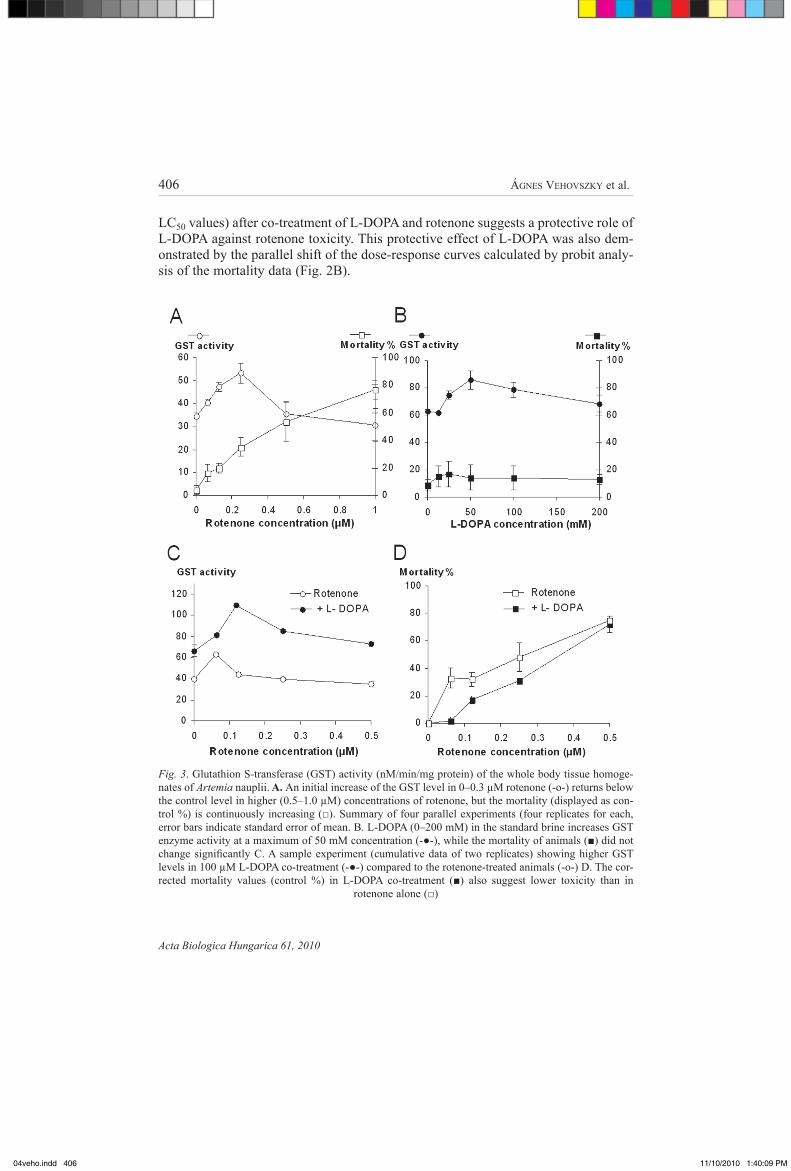
Fig. 3. Glutathion s-transferase (Gst) activity (nm/min/mg protein) of the whole body tissue homoge-nates of Artemia nauplii. A. an initial increase of the Gst level in 0–0.3 µm rotenone (-o-) returns below the control level in higher (0.5–1.0 µm) concentrations of rotenone, but the mortality (displayed as con-trol %) is continuously increasing (□). Summary of four parallel experiments (four replicates for each, error bars indicate standard error of mean. b. l-dopa (0–200 mm) in the standard brine increases Gst enzyme activity at a maximum of 50 mM concentration (-●-), while the mortality of animals (■) did not change significantly C. A sample experiment (cumulative data of two replicates) showing higher GST levels in 100 µM L-DOPA co-treatment (-●-) compared to the rotenone-treated animals (-o-) D. The cor-rected mortality values (control %) in L-DOPA co-treatment (■) also suggest lower toxicity than in
rotenone alone (□)
04veho.indd 406 11/10/2010 1:40:09 PM

Complex I inhibitor effects in artemia 407
Acta Biologica Hungarica 61, 2010
Glutathione S-transferase activity in the brine shrimp nauplii
for biochemical analysis the 24 hours Artemia nauplii were exposed to sublethal (0–1.0 µm) concentrations of rotenone (four wells of the microplate for each concen-tration) and after 24 hours incubation the animals were collected from each well and further processed for Gst measurement. in parallel series of experiments the mortal-ity of nauplii was determined in rotenone (0–1.0 µm) as described for the toxicity tests (see above). control wells contained standard brine.
in control conditions the Gst activity in the whole body tissue of the Artemia nauplii was measured between 30–70 nm/min/mg protein (51.0 ± 2.6 nm/min/mg protein, mean ± sE, n = 20, individual experiments). rotenone treatment had a dual effect on the Gst activity: low concentrations of rotenone (below 0.3 µm) increased the Gst activity up to about the 50 percent (from 34.7 ± 3.6 nm/min/mg protein to 53.3 ± 6.8 nm/min/mg protein; mean ± sE, n = 4), while with higher concentrations (0.5 – 1.0 µm) of rotenone the Gst activity declined (down to 30.9 ± 7.2 nm/min/mg protein) (fig. 3a). the mortality of the animals, however, continuously increased in rotenone in a dose-dependent manner (fig. 3a), in agreement with our previous results obtained from rotenone toxicity tests (fig. 1a, b).
l-dopa (up to 200 mm concentrations) applied in the standard brine also increased the Gst activity with a maximum around 50 percent of the control (62.9 ± 1.1 nm/min/mg protein, to 85.6 ± 6.9 nm/min/mg protein, mean ± sE, n = 3), but the mortality of the nauplii did not change significantly in this range of L-DOPA concentration (fig. 3b). in parallel experiments we compared the Gst activity levels of rotenone-treated animals with those which additionally received 100 mm l-dopa. l-dopa elicited a higher level of Gst activity (up to 109.8 nm/min/mg protein againts to 63.0 nm/min/mg protein) over the entire range of rotenone concentrations (fig. 3c), and at the same time, lower mortality was observed (fig. 3c), suggesting the protective effect of l-dopa treatment.
discUssion
our results showed a relatively high sensitivity of 1-day-old Artemia nauplii to roten-one (lc50: 0.37 µm or 0.146 mg/l water), well inside the range of the other crusta-ceans reported (lc50: 0.05–0.5 mg/l, reviewed by Ling [20]). A recent publication [36] reporting similar toxicological testing of related “rotenoids” (natural extracts from a mangrove plant) on brine shrimps gave similar sensitivity (lc50 = 1.6 and 4.0 µg/ml), although we do not know how pure the toxins in these extracts are.
rotenone (along with mptp and mpp+) is also used in experimental medicine to selectively destroy dopaminergic neurons in the cns, thus mimicking the behav-ioural and cellular symptoms of the Parkinson’s disease [3, 18, 35]. Although Artemia larvae do not allow direct study of the individual dopaminergic neurons, our current results suggest some similarities with those animal models, as Artemia nauplii were killed by all the dopaminergic neurotoxins, rotenone, mptp and mpp+ in a dose-
04veho.indd 407 11/10/2010 1:40:09 PM

408 Ágnes VehoVszky et al.
Acta Biologica Hungarica 61, 2010
dependent manner. in vertebrates mptp is converted by monoamine oxidase b (mao b) enzyme into mpp+, which is taken up by the dopamine transporter system and accumulated intracellularly in the dopamine-containing neurons [31]. Invertebrates may have some similar mptp conversion and uptake mechanisms as both mptp and mpp+ are toxic to different invertebrate species [18]. Moreover, rotenone is a much more potent neurotoxin, than mptp or mpp+, both in vertebrates and invertebrates [19, 24]. We also found that for the same mortality three magnitudes higher concen-trations were required for mptp and mpp+ (lc50 was 0.21 ± 0.09 mm, and 0.2 ± 0.08 mm, respectively) compared to rotenone (lc50: 0.37 ± 0.04 µm).
rotenone decreases the level of the tyrosine hydroxylase enzyme which produces the precursor (DOPA) for the further dopamine synthesis [15, 21], thus L-DOPA treatment is often used to enhance dopamine level in the cns both in therapy and experimental models [1, 23]. In agreement with these data our Artemia tests also showed higher lc50 value of rotenone in the presence of 100 mm l-dopa (1.03 ± 0.66 µm against 0.37 ± 0.04 µm) suggesting its neuroprotective effect against rotenone toxicity.
biochemical results demonstrated that lower doses (up to 0.3 µm) of rotenone increase glutathione s-transferase activity in Artemia (fig. 3a, c). similar results were also reported in Artemia preparations or in Penaeus prawn tissues when toxic effects and oxidative stress were involved [4, 37]. Higher concentrations of rotenone, however, decreased the enzyme activity in the whole body tissue homogenates. this suggest that above a particular concentration of rotenone the oxidative damage evoked by the neurotoxin impairs the intracellular compensatory mechanisms, including Gst system, therefore the activity of the enzymes responsible for biotrans-formation would decrease. similar results are reported on cyanotoxin-treated prawn tissue preparations, when elevation of the Gst activity is measured in lower toxin concentrations, while higher doses seem to decrease the enzyme activity [29].
our biochemical results on the Gst levels of the Artemia tissue samples suggest an additional, intracellular effect of the dopaminergic precursor l-dopa. We demon-strated, that l-dopa in a particular (50–200 mm) dose range increased the Gst activity in the Artemia nauplii, while l-dopa co-treament with rotenone resulted an even more increased Gst activity in the tissue homogenates, and lower mortality of the animals (fig. 3c, d). the higher level of Gst activity, thus, may partly explain the protective effect of l-dopa co-treatment, as Gst emzymes are part of the intra-cellular defense, which contributing to eliminate the effect of free radicals after com-plex i inhibition.
We conclude that Artemia shows similarities with other animal models when com-plex i inhibitors are applied to selectively destroy the dopaminergic system in the cns. all the dopaminergic neurotoxins (rotenone, mptp and mpp+) resulted con-centration-dependent mortality, while l-dopa co-treatment increased the survival of the rotenone-treated animals. the biochemical results, moreover, demonstrated that l-dopa triggers an increase of Gst activity, as the suggested intracellular mecha-nism againts the rotenone-induced toxicity.
04veho.indd 408 11/10/2010 1:40:09 PM

Complex I inhibitor effects in artemia 409
Acta Biologica Hungarica 61, 2010
acknoWlEdGEmEnts
this work was supported by the hungarian otka t63451 grant for a. V. and mEh Öktm 12002 grant for a. f. We are grateful to christopher Elliott for reading the draft of the manuscipt.
rEfErEncEs
1. alam, m., schmidt, W. J. (2004) l-dopa reverses the hypokinetic behaviour and rigidity in rote-none-treated rats. Behav. Brain res. 153, 439–446.
2. baez, s., segura-aguilar, J., Widersten, m., Johansson a., mannervick, b. (1997) Glutathione trans-ferases catalyse the detoxication of oxidized metabolites (o-quinones) of catecholamines and may serve as an antioxidant system preventing degenerative cellular processes. Biochem. J. 324, 25–28.
3. beal, m. f. (2001) Experimental models of parkinson’s disease. Nature reviews Neuroscience 2, 325–332.
4. Beattie, K. A., Ressler, J., Wiegand, C., Krause, E., Codd, G. A., Steinberg, C. E. W., Pflugmacher, S. (2003) comparative effects and metabolism of two microcystins and nodularin in the brine shrimp Artemia salina. Aquatic toxicology 62, 219–226.
5. bharath, s., hsu, m., kaur, d., rajagopalan, s., andersen, J. k. (2002) Glutathione, iron and parkinson’s disease. Biochem. Pharmacol. 64, 1037–1048.
6. Bradford, M. (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254.
7. coulom, h., birman, s. (2004) chronic exposure to rotenone models sporadic parkinson’s disease in Drosophila melanogaster. J. Neurosci. 24, 10993–10998.
8. delaney, J. m., Wilkins, r. m. (1995) toxicity of microcystin-lr, isolated from microcystis aerugi-nosa, against various insect species. toxicon 33, 771–778.
9. Eaton, d. l., bammler, t. k. (1999) concise review of the glutathione s-transferases and their sig-nificance to toxicology. toxicol. Sci. 49, 156–164.
10. Esposti, m. d. (1998) inhibitors of nadh-ubiquinone reductase: an overview. Biochemical et Biophysica Acta-Bioenergetics 1364, 222–235.
11. favilla, m., macchia l., Gallo, a., altomare, c. (2006) toxicity assessment of metabolites of fungal biocontrol agents using two different (Artemia salina and Daphnia magna) invertebrate bioassays. Food Chem. toxicol. 44, 1922–1931.
12. finney, d. J. (1971) Probit Analysis. cambridge University press, cambridge, Uk.13. fukui, h., moraes, c. t. (2008) the mitochondrial impairment, oxidative stress and neurodegenera-
tion connection: reality or just an attractive hypothesis? trend in Neurosciences 31, 251–256.14. Habig, W. H., Pabst, M. J., Jakoby, W. B. (1974) Glutathione S-transferases. The first enzymatic step
in mercapturic acid formation. J. Biol. Chem. 249, 7130–7139.15. hirata, Y., nagatsu, t. (2005) rotenone and cccp inhibit tyrosine hydroxylation in rat striatal tissue
slices. toxicology 216, 9–14.16. hoekstra, J. a. (1987) acute bioassays with control mortality water air and soil pollution. Water, Air,
and Soil Pollution 35, 311–317.17. Jha, n., kumar, m. J., boonplueang, r., andersen, J. k. (2002) Glutathione decreases in dopaminer-
gic pc 12 cells interfere with the ubiquitin protein degradation pathway: relevance for parkinson’s disease? J. Neurochem. 80, 555–561.
18. kitamura, Y., shimohama, s., akaike, a., taniguchi, t. (2000) the parkinsonian models: invertebrates to mammals. Jap. J. Pharmacol. 84, 237–243.
19. kitamura, Y., inden, m., sanada, h., takata, k., taniguchi, t., shimohama, s., orii, h., mochii, m., agata, k., Watanabe, k. (2003) inhibitory effects of antiparkinsonian drugs and caspase inhibitors in a parkinsonian flatworm model. J. Pharmacol. Sci. 92, 137–142.
20. ling, n. (2002) Rotenone – a review of its use for fisheries management. department of conservation science publishing and service Unit, Wellington.
04veho.indd 409 11/10/2010 1:40:10 PM

410 Ágnes VehoVszky et al.
Acta Biologica Hungarica 61, 2010
21. milusheva, E., baranyi, m., kittel, a., sperlagh, b., Vizi, E. s. (2005) increased sensitivity of striatal dopamine release to h(2)o(2) upon chronic rotenone treatment. Free radic. Biol. med. 39, 133–142.
22. moore, d. J., West, a. b., dawson, V. l., dawson, t. m. (2005) molecular pathophysiology of parkinson’s disease. Ann. rev. Neurosci. 28, 57–87.
23. nagatsu, t., sawada, m. (2009) l-dopa therapy for parkinson’s disease: past, present, and future. Parkinsonism and related Disorders 15, s3–s8
24. nieto, m., Gil-beab, f. J., dalfó, E., cuadrado, m., cabodevilla, f., sánchez, b., catena, s., sesma, t., ribé, E., ferrer, i. ramírez m. J., Gómez-isla, t. (2006) increased sensitivity to mptp in human alpha-synuclein a30p transgenic mice. Neurobiol. of Aging 7, 848–856.
25. nunes, b. s., carvalho, f. d., Guilhermino, l. m. (2006) Effects of widely used pharmaceuticals and a detergent on oxidative stress biomarkers of the crustacean Artemia parthenogenetica. Chemosphere 62, 581–594.
26. nunes, b. s., carvalho, f. d., Guilhermino, l. m., Van stappen, G. (2006) Use of the genus Artemia in ecotoxicity testing. environ. Poll. 144, 453–462.
27. obata, t. (2006) nicric oxide and mpp+-induced hydroxyl radical generation. J. Neural transmission 113, 1131–1144.
28. Olanow, C. W. (2004) The scientific basis for the current treatment of Parkinson’s disease. Annu. rev. med. 55, 41–60.
29. Pflugmacher, S., Wiegand, C., Werner, S., Schroder, H., Kankaanpaa, H. (1995) Activity and substrate specificity of cytosolic and microsomal glutathione S-transferase in Australian black tiger prawns (Penaeus monodon) after exposure to cyanobacterial toxins. env. toxicol. 20, 301–307.
30. da silva, E. m., soares, a. m. V. m., moreno, a. J. m. (1998) the use of the mitochondrial trans-membrane electric potential as an effective biosensor in ecotoxicological research. Chemosphere 36, 2375–2390.
31. storch, a., ludolph, a. c., schwarz, J. (2004) dopamine transporter: involvement in selective dop-aminergic neurotoxicity and degeneration. J. Neural transmission 111, 1267–1286.
32. testa, c. m., sherer, t. b., Greenamyre, J. t. (2005) rotenone induces oxidative stress and dopamin-ergic neuron damage in organotypic substantia nigra cultures. mol. Brain res. 134, 109–118.
33. Vanhaecke, p., persoone, G., claus, c., sorgeloos, p. (1981) proposal for a short-term toxicity test with Artemia nauplii. ecotoxicology and environmental Safety 5, 382–387.
34. Varó, i., navarro, J. c., amat, f., Guilhermino, l. (2002) characterisation of cholinesterases and evaluation of the inhibitory potential of chlorpyrifos and dichlorvos to Artemia salina and Artemia parthenogenetica. Chemosphere 48, 563–569.
35. Vehovszky, Á., szabó, h., hiripi, l., Elliott, c. J. h., hernádi, l. (2007) behavioural and neural deficits induced by rotenone in the pond snail Lymnaea stagnalis. a possible model for parkinson’s disease in an invertebrate. eur. J. Neurosci. 25, 2123–2130.
36. Wangensteen, h., alamgir, m., rajia, s., meza, t. J., samuelsen, a. b., malterud, k. E. (2007) cytotoxicity and brine shrimp lethality of rotenoids and extracts from Sarcolobus globosus. Natural Product Communications 2, 841–844.
37. Wang, W.-n., Wang, Y., Wang, a.-l., Wangensteen, h., alamgir, m., rajia, s., meza, t. J., samuelsen, a. b., malterud, k. E. (2006) Effect of supplemental l-ascorbyl-2-polyphosphate (app) in enriched live food on the immune response of Penaeus vannamei exposed to ammonia-n. Aquaculture 256, 552–557.
04veho.indd 410 11/10/2010 1:40:10 PM
Related Documents











![The occurrence of the brine shrimp, Artemia franciscana ... · America [25], other Artemia species include: Artemia persimilis of South America [26], Artemia salina in the Mediterranean](https://static.cupdf.com/doc/110x72/5c4b6a7093f3c3117d72c1b0/the-occurrence-of-the-brine-shrimp-artemia-franciscana-america-25-other.jpg)