Nephrol Dial Transplant (1996) 11: 1803 1809 Original Article Nephrology Dialysis Transplantation Effects of peritoneal effluents on mesothelial cells in culture: cell proliferation and extracellular matrix regulation F. Perfumo, P. Altieri, M. L. Degl'Innocenti, G. M. Ghiggeri, G. Caridi, A. Trivelli and R. Gusmano Nephrology Department, G. Gaslini Children's Hospital, Genoa, Italy Abstract Peritoneal fibrosis in patients on peritoneal dialysis is the result of interstitial collagen accumula- tion within the peritoneal membrane and in mural spaces. Hypothetically, collagen expression by target cells may be regulated by specific endoperitoneal fac- tors, though the existence of such factors has not yet been demonstrated. We evaluated the effects of cell-free peritoneal effluents obtained from six children undergoing peri- toneal dialysis on several mesothelial cell functions in vitro. Human peritoneal mesothelial cells (MC) were obtained from the omental tissue of six uraemic chil- dren who were undergoing surgery for insertion of a peritoneal catheter. Cells at confluence were utilized to determine cytotoxicity (LDH release), viability (trypan blue), proliferation ( 3 H-thymidine incorporation), col- lagen expression ( 3 H-proline incorporation, SDS-Page) and mRNA (dot-blot). A preliminary series of experiments, was undertaken to define which of the successive fluid collections during a dialytic procedure induces the greatest changes; this revealed maximal effects of the effluent from the long stasis period. Exposure to peritoneal effluents obtained from four patients with acute peritonitis induced marked changes in cell morphology, stimulated by ( 3 H)-thymi- dine incorporation into DNA by 300% and upregulated the expression and transcription of type III collagen (6-fold increment in COL3A1 mRNA). Qualitatively but not quantitatively comparable changes in cell pro- liferation (+100%) and collagen expression were induced by peritoneal effluents from patients without peritonitis. In an effort to reproduce the effect of peritoneal effluents in vitro, we exposed mesothelial cells to various cytokines putatively present in infected peritoneal effluents, namely IL-2, TGF)9 and TNFa; in no case did we find stimulation of cell proliferation. Finally TGF0 but not TNFa or IL2 upregulated collagen synthesis by these cells. Thesefindingsdemonstrate a direct influence of cell- free peritoneal effluents on mesothelial cell functions, including stimulation of interstitial collagen expression. Correspondence and offprint requests to: Francesco Perfumo MD, Department of Nephrology, G. Gaslini Children's Hospital, Largo G. Gaslini 5, 16148 Genoa, Italy. All these changes were more evident upon exposure to effluents collected during acute peritonitis, which sug- gests a link between recurrent peritoneal infection and collagen deposition, the most typical precursor of peritoneal fibrosis. Key words: peritoneal dialysis; peritonitis; fibrosis; collagen Introduction Peritoneal fibrosis is a major long-term complication of peritoneal dialysis and its occurrence seriously ham- pers the therapeutic potential of this technique [1,2]. From a pathological standpoint, peritoneal fibrosis occurring in patients on peritoneal dialysis appears as the extracellular deposition of interstitial collagens (type I and type III) with fibrous organization of the basement membrane (fibrous membrane) or derange- ment of the cellular organization within submembrane spaces (muralfibrosis)[3,4]. These two separate entities differ from classic thickening of the basement mem- brane due to collagen IV accumulation, a process which causes hyalinization of the superficial layer of the peritoneal membrane, probably as a result of the glycosylation of collagen. Although several pathogenetic aspects of peritoneal fibrosis are still open to discussion, it has long been recognized that recurrent episodes of peritoneal infec- tion often precede the fibrotic degeneration [5], thus suggesting the involvement of infectious agents per se and/or of infectious reactions. One reasonable possibil- ity is that nucleated blood cells (polymorphonuclears, macrophages) attracted into the peritoneal cavity during infection trigger a series of events culminating in the production of soluble factors (cytokines, growth factors) [6,7] which stimulate extracellular matrix (ECM) expression by mesothelial cells (MC) and/or fibroblasts [8]. Human peritoneal cells on their own are able to synthesize cytokines, such as interleukin 6, and this release is upregulated by an autocrine process [9]. Therefore cytokines derived from mesothelial cells might contribute directly to the cytokine network S 1996 European Renal Association-European Dialysis and Transplant Association by guest on July 6, 2011 ndt.oxfordjournals.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nephrol Dial Transplant (1996) 11: 1803 1809
Original Article
NephrologyDialysis
Transplantation
Effects of peritoneal effluents on mesothelial cells in culture:cell proliferation and extracellular matrix regulation
F. Perfumo, P. Altieri, M. L. Degl'Innocenti, G. M. Ghiggeri, G. Caridi, A. Trivelli and R. Gusmano
Nephrology Department, G. Gaslini Children's Hospital, Genoa, Italy
Abstract Peritoneal fibrosis in patients on peritonealdialysis is the result of interstitial collagen accumula-tion within the peritoneal membrane and in muralspaces. Hypothetically, collagen expression by targetcells may be regulated by specific endoperitoneal fac-tors, though the existence of such factors has not yetbeen demonstrated.
We evaluated the effects of cell-free peritonealeffluents obtained from six children undergoing peri-toneal dialysis on several mesothelial cell functions invitro. Human peritoneal mesothelial cells (MC) wereobtained from the omental tissue of six uraemic chil-dren who were undergoing surgery for insertion of aperitoneal catheter. Cells at confluence were utilized todetermine cytotoxicity (LDH release), viability (trypanblue), proliferation (3H-thymidine incorporation), col-lagen expression (3H-proline incorporation,SDS-Page) and mRNA (dot-blot). A preliminaryseries of experiments, was undertaken to define whichof the successive fluid collections during a dialyticprocedure induces the greatest changes; this revealedmaximal effects of the effluent from the long stasisperiod. Exposure to peritoneal effluents obtained fromfour patients with acute peritonitis induced markedchanges in cell morphology, stimulated by (3H)-thymi-dine incorporation into DNA by 300% and upregulatedthe expression and transcription of type III collagen(6-fold increment in COL3A1 mRNA). Qualitativelybut not quantitatively comparable changes in cell pro-liferation (+100%) and collagen expression wereinduced by peritoneal effluents from patients withoutperitonitis. In an effort to reproduce the effect ofperitoneal effluents in vitro, we exposed mesothelialcells to various cytokines putatively present in infectedperitoneal effluents, namely IL-2, TGF)9 and TNFa; inno case did we find stimulation of cell proliferation.Finally TGF0 but not TNFa or IL2 upregulatedcollagen synthesis by these cells.
These findings demonstrate a direct influence of cell-free peritoneal effluents on mesothelial cell functions,including stimulation of interstitial collagen expression.
Correspondence and offprint requests to: Francesco Perfumo MD,Department of Nephrology, G. Gaslini Children's Hospital, LargoG. Gaslini 5, 16148 Genoa, Italy.
All these changes were more evident upon exposure toeffluents collected during acute peritonitis, which sug-gests a link between recurrent peritoneal infection andcollagen deposition, the most typical precursor ofperitoneal fibrosis.
Key words: peritoneal dialysis; peritonitis; fibrosis;collagen
Introduction
Peritoneal fibrosis is a major long-term complicationof peritoneal dialysis and its occurrence seriously ham-pers the therapeutic potential of this technique [1,2].From a pathological standpoint, peritoneal fibrosisoccurring in patients on peritoneal dialysis appears asthe extracellular deposition of interstitial collagens(type I and type III) with fibrous organization of thebasement membrane (fibrous membrane) or derange-ment of the cellular organization within submembranespaces (mural fibrosis) [3,4]. These two separate entitiesdiffer from classic thickening of the basement mem-brane due to collagen IV accumulation, a processwhich causes hyalinization of the superficial layer ofthe peritoneal membrane, probably as a result of theglycosylation of collagen.
Although several pathogenetic aspects of peritonealfibrosis are still open to discussion, it has long beenrecognized that recurrent episodes of peritoneal infec-tion often precede the fibrotic degeneration [5], thussuggesting the involvement of infectious agents per seand/or of infectious reactions. One reasonable possibil-ity is that nucleated blood cells (polymorphonuclears,macrophages) attracted into the peritoneal cavityduring infection trigger a series of events culminatingin the production of soluble factors (cytokines, growthfactors) [6,7] which stimulate extracellular matrix(ECM) expression by mesothelial cells (MC) and/orfibroblasts [8]. Human peritoneal cells on their ownare able to synthesize cytokines, such as interleukin 6,and this release is upregulated by an autocrine process[9]. Therefore cytokines derived from mesothelial cellsmight contribute directly to the cytokine network
S 1996 European Renal Association-European Dialysis and Transplant Association
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from

1804
which regulates the intraperitoneal inflammatoryresponse associated with bacterial peritonitis.
This possibility requires further experimental sup-port. The first task is to determine whether peritonealeffluents from infected patients contain soluble factorsable to stimulate ECM; the second is to characterizethe target cell of this effect. In the present study weevaluated the effects of peritoneal effluents obtainedfrom children undergoing CPD on mesothelial cellproliferation and ECM expression focusing on differ-ences between stable conditions and conditions ofperitonitis.
Subjects and methods
Patients
Peritoneal effluents were obtained from six children (meanage 10 years, range 6-18) who had been undergoing chronicperitoneal dialysis (CPD) for at least 1 year. The dialysisschedule was automated peritoneal dialysis using commer-cially available dialysis fluid (Baxter Viaflex LCB 4800 L).Two children had had negative effluent cultures for at least6 months and were in a stable clinical and metabolic balance.The other four patients had been affected by peritonitis forat least 2 days. The pathogens isolated were Staphylococcusaureus in two children and Streptococcus epidermidis andE. coli in the remaining two. In one of these four patientseffluent was collected before the start of treatment, while inthe other intraperitoneal treatment with vancomycin orcephalosporins had already begun. This study was approvedby the Ethics Committee of G. Gaslini Institute forBiomedical Research. Informed consent was obtained fromomental donors.
Peritoneal effluent collection
In a preliminary approach we determined the effects on MCfunctions of 10 successive effluent collections dwell obtainedfrom two patients during a dialytic session. In a second seriesof experiments, only effluents obtained at 9 p.m. at the startof dialysis were tested (12-h dwell).
Cell isolation and culture
Human peritoneal mesothelial cells were obtained from theomental tissue of six uraemic children who were undergoingsurgery to insert a peritoneal catheter. The omental tissuewas cut into pieces with a razor-blade, and after an initialwash in sterile polysaline buffer and digestion with 1% trypsinat 37°C for 20 min, cells were maintained in Medium 199(Seromed, Berlin, Germany) plus 15% FCS, 5 ng/ml insulin,5 |ig/ml transferrin, 5% sodium selenite, under 5% CO2 at37°C [10]. The following parameters were utilized to assessthe purity of our mesothelial preparations: (1) uniformcobblestone appearance; (2) lack of staining for factor VIII;(3) positivity for staining with anti-cytokeratin 8 antibodies[10]. All data presented here derive from experiments per-formed with cells utilized at confluence at the second passage.Effluents were mixed for 10 min on a rotor at 37;C and afterfiltration (see below) were stored at — 20°C. Each experimentwas performed in triplicate, with parallel incubation ofpathological and non-infected effluents in each setting.
F. Perfumo et al.
Preparation of free effluents
After collection, peritoneal effluents were mixed for 10 minat 37°C on a rotor and immediately centrifuged at lOOOOgfor 30 min at + 5°C; supernatants were filtered through0.2 urn cellulose acetate filters (Corning, Cambridge,Massachussetts) to eliminate cells and cell debris. Cell-freeeffluents were then diluted 1:5 with serum-free medium andwere incubated with MC for 12, 24 and 48 h at 37CC in5% CO2.
Effects ofcytokines
In an effort to tentatively reproduce the effect of infectedperitoneal effluents on MC in vitro, we incubated severalcytokines (TGFjS, TNFa, IL2) with MC in standardizedconditions. The range of concentrations for each cytokinewas chosen on the basis of their effects on other cell types.
Cell viability
Cell viability was assayed with trypan blue. After digestionwith 0.05% trypsin—0.02% EDTA, cells were incubated with0.05% trypan blue for 2 h at 37°C in 5% CO2, and afterthorough washing were counted in a haemacytometer.
LDH assay
The acute cytolytic effect of cell-free peritoneal effluents onmesothelial cells was evaluated as LDH release, the enzymebeing determined by a colorimetric method which utilizesthe oxidation of NADH as a measure of LDH activity inthe presence of pyruvate [11] (Kit Roche, Milan, Italy).Maximal cytolysis was induced by treating cells with 1%Triton x-100. LDH activity was determined in mU/ml, wheremU is the enzyme required to transform 0.0167 nMNADH/min.
[3H]-thymidine incorporation
Proliferation of mesangial cells was evaluated as incorpora-tion of [3H]-thymidine (Amersham, Little Chalfont; UK)into growing cells. Thymidine incorporation was determinedby incubating 104/ml adherent cells with 0.5 uCi/ml [3H]-thymidine for 24 and 48 h at 37°C in 5% CO2. After thoroughwashing in RPMI, cells were counted in a Packard /?-scintillation counter after transfer to filter-paper sheetsunder vacuum.
Collagen characterization
Collagen synthesis by mesothelial cells was evaluated as theincorporation of [3H]-proline into partial pepsin digests.Briefly, after incubation with 50 ng/ml sodium ascorbate for18 h, 80 ug/ml of /?-aminopropionitrile and 50 uCi/ml of[3H]-proline (Amersham) were added to the cells, which werethen incubated for another 48 h. Cells and extracellularmatrix were scraped off and digested for 24 h with 250 ug/mlpepsin in 0.5 M acetic acid and then absorbed on glassmicrofibre filter membranes (Whatman, Maidstone, UK)under vacuum. Radioactivity was counted in a scintillationmedium; results are expressed as c.p.m. per 105 cells. Thecollagen composition of [3H] radiolabelled extracellular
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from
Collagen expression by mesothelial cells
2m0 H-thymidine incorporation
1805
i.wo ^
I10000 -
3000 -
Patient 1
Patient 2
Effluent number
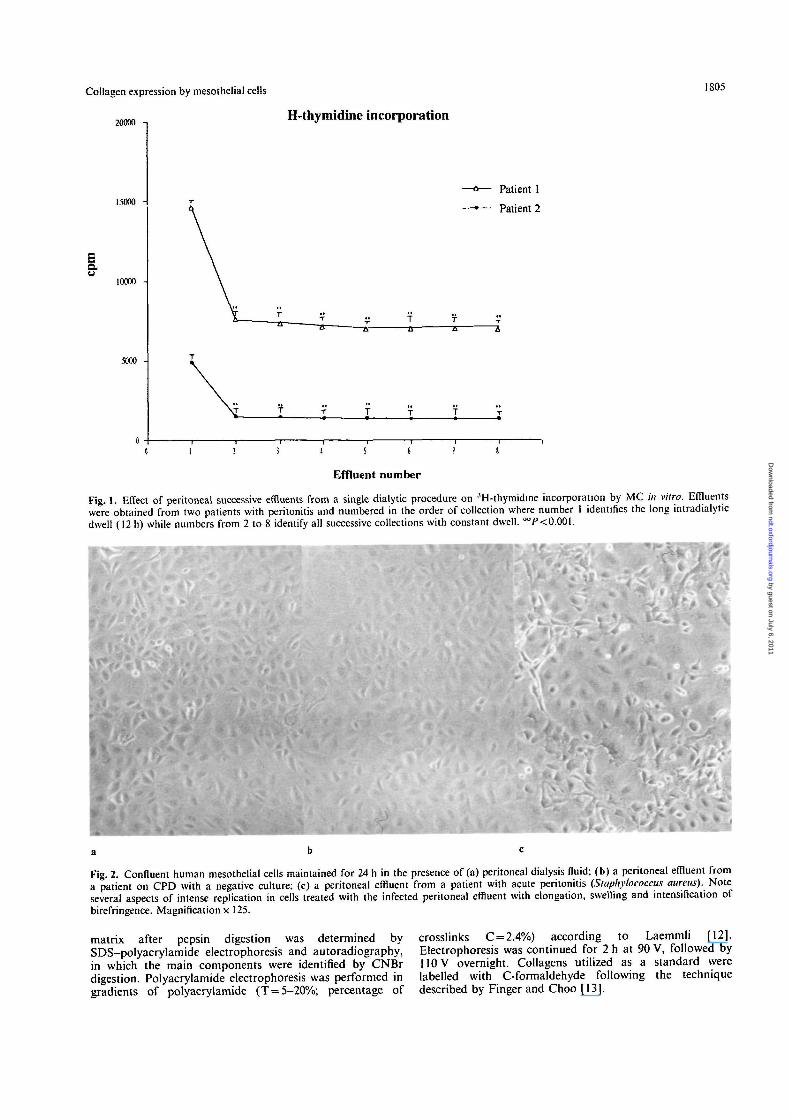
Fig. 1. Effect of peritoneal successive effluents from a single dialytic procedure on 3H-thymidine incorporation by MC in vitro. Effluentswere obtained from two patients with peritonitis and numbered in the order of collection where number 1 identifies the long intradialyticdwell (12 h) while numbers from 2 to 8 identify all successive collections with constant dwell. °°P< 0.001.
Fig. 2. Confluent human mesothelial cells maintained for 24 h in the presence of (a) peritoneal dialysis fluid; (b) a peritoneal effluent froma patient on CPD with a negative culture; (c) a peritoneal effluent from a patient with acute peritonitis (Staphylococcus aureus). Noteseveral aspects of intense replication in cells treated with the infected peritoneal effluent with elongation, swelling and intensification ofbirefringence. Magnification x 125.
matrix after pepsin digestion was determined bySDS-polyacrylamide electrophoresis and autoradiography.in which the main components were identified by CNBrdigestion. Polyacrylamide electrophoresis was performed ingradients of polyacrylamide (T = 5-20%; percentage of
crosslinks C = 2.4%) according to Laemmli [12].Electrophoresis was continued for 2 h at 90 V, followed by110 V overnight. Collagens utilized as a standard werelabelled with C-formaldehyde following the techniquedescribed by Finger and Choo [13].
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from

1806
COL3AlmRNA
The expression of al(III) mRNAs by mesothelial cells wasdetermined by dot-blot hybridization. Total mRNA (5 ug)was extracted in 6 M guanidine-isothiocyanate and trans-ferred to HYBOND-C membranes (Amersham).
Analysis of COL3A1 mRNA was performed by hybridiza-tion with pDACl clone containing the cDNA correspondingto the amino-terminal region of pre-pro al(III) collagen(kind gift from Dr F. Ramirez). A probe for detection ofglyceraldehyde 3 phosphate dehydrogenase (GAPDH)mRNA was prepared by RT-PCR from human RNA usingcommercial primers (Clonotech, Palo Alto, California) andwas utilized to normalize COL3A1 mRNA.
Statistical analysis
Data were analysed using the t test for unpaired sequences.Results are given as mean + SD.
Results
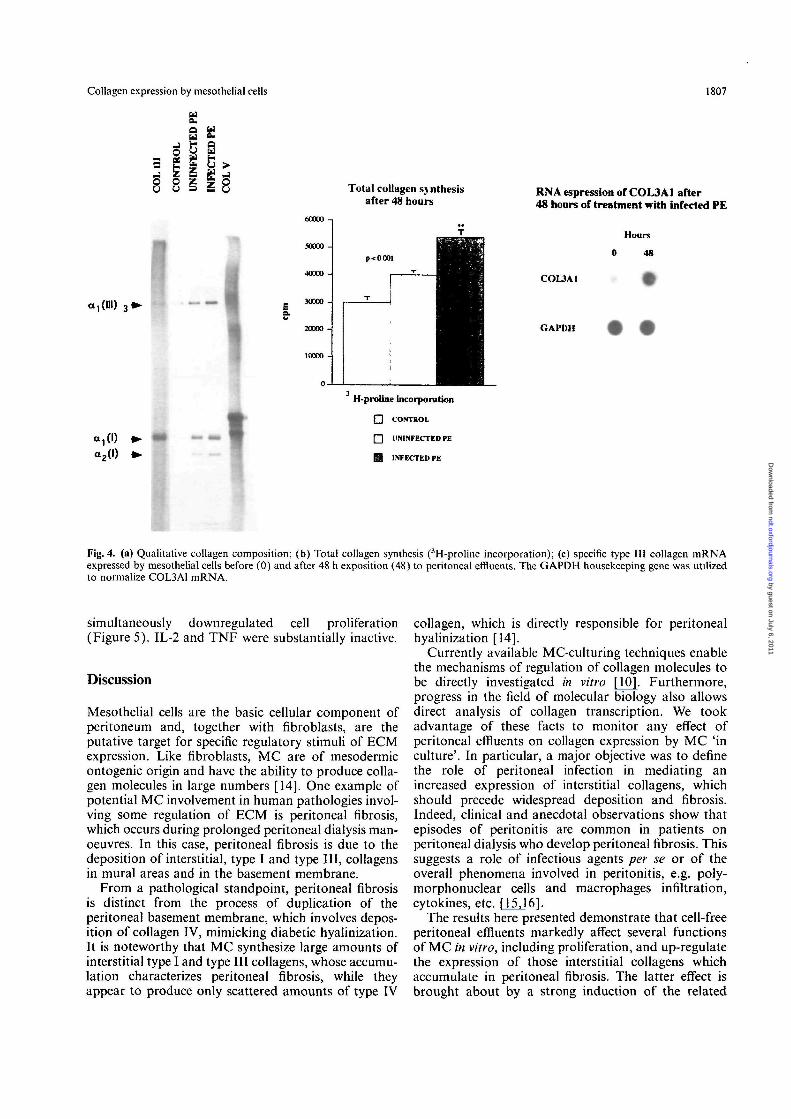
Our first approach was to evaluate the effect on MCfunctions of 10 successive effluent collections within asingle dialytic session. The most pronounced effect wasinduced by the effluent (Figure 1) obtained after a 12-hdwell period. All subsequent experiments were thenperformed utilizing the long-dwelling effluent. Asshown in Figure 2, MC exposed to infected effluentsfor 24 h acquired typical features of active prolifera-tion, such as elongation, swelling, and intensificationof birefringence. The elongated morphology acquiredby cells exposed to infected effluents mimics the mor-phology of MC 'in culture' in an initial growth phase.Compared with dialysis solution, cell-free peritonealeffluents from patients on CPD with peritonitis induceda three-to fourfold increment in [3H]-thymidine incorp-oration into DNA by MC (from 2100 + 190 to7400 + 320 c.p.m. per 104 cells) (Figure 3). However,the effluents from uninfected patients also induced asignificant albeit smaller increment in [3H] thymidineincorporation into MC (4050 + 250 c.p.m.), thus sug-gesting that substances which regulate cell proliferationare also present in the peritoneum of patients under-going CPD in sterile conditions. In spite of the afore-mentioned marked effects on cell proliferation,peritoneal effluents did not affect cell viability, nor didthey exert cytotoxic phenomena, as judged by trypanblue incorporation and LDH release from cells after48-h incubation, in any of the different experimentalconditions (Figure 3). Figure 4 shows the effect ofperitoneal effluents on collagen synthesis, as evaluatedby [3H]-proline incorporation, in all and in singlecollagenic components. The results indicate that overallcollagen synthesis doubles in the presence of infectedperitoneal effluents (51800 + 2800 versus30 300 ±1500 c.p.m. for 10s cells), while a smallerincrement takes place in the presence of uninfectedeffluents (40800 + 500 c.p.m.). The greatest changeswere observed on type III collagen expression, whichincreased by a factor of 8-10 with infected effluents
F. Perfumo el al.
H-thymidine incorporation
Cell viability
l«h _ r T
uS ao I 2--
Q control
D uninfected PE
0 infected PE
incubation 48 hours
LDH release
1 5
IX
a
incubation 48 hours
Fig. 3. Thymidine incorporation (a) cell viability (b) and LDHrelease (c) by mesothelial cells upon incubation for 24 and for 48 hwith the first peritoneal effluents obtained from four patients withperitonitis and two with a normal effluent culture. Six different cellcultures were grown as described in Subjects and Methods andincubated with different effluents each experiment incorporatinginfected, non-infected effluents and peritoneal solutions. Results arethe mean±SD. */'<0.006 **P<0.0l versus control (peritonealdialysis fluid).
compared with dialysis fluid alone, and by a factor oftwo compared with uninfected effluents (Figure 4B).In order to characterize the mechanisms of regulationmore accurately, we determined the mRNA relative totype III collagen by utilizing a molecular probe specificfor the amino-terminal region of pre-pro al (III) colla-gen. A six-fold increment in COL3A1 mRNA expres-sion by MC incubated for 48 h with infected peritonealeffluents was found, thus indicating a direct effect onthe relevant promoter (Figure 4C).
A further series of experiments was undertaken tocharacterize the putative substance involved in theaforementioned cellular effects.
Among cytokines, only TGF/? was effective in upreg-ulating the synthesis of total collagen by MC, but this
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from
Collagen expression by mesothelial cells 1807
Total collagen synthesisafter 48 hours
RNA espression of COL3A1 after48 hoars of treatment with infected PE
Ia ,(111) 3 ^
Hours
0 48
COL3A1
GAPDH
a , (I) • •
0t2(D • >
II
H-proline incorporation
• CONTROL
• UNINFECTED PE
ffl INFECTED PE
Fig. 4. (a) Qualitative collagen composition; (b) Total collagen synthesis (3H-proline incorporation); (c) specific type III collagen mRNAexpressed by mesothelial cells before (0) and after 48 h exposition (48) to peritoneal effluents. The GAPDH housekeeping gene was utilizedto normalize COL3A1 mRNA.
simultaneously downregulated cell proliferation(Figure 5). IL-2 and TNF were substantially inactive.
Discussion
Mesothelial cells are the basic cellular component ofperitoneum and, together with fibroblasts, are theputative target for specific regulatory stimuli of ECMexpression. Like fibroblasts, MC are of mesodermicontogenic origin and have the ability to produce colla-gen molecules in large numbers [14]. One example ofpotential MC involvement in human pathologies invol-ving some regulation of ECM is peritoneal fibrosis,which occurs during prolonged peritoneal dialysis man-oeuvres. In this case, peritoneal fibrosis is due to thedeposition of interstitial, type I and type III, collagensin mural areas and in the basement membrane.
From a pathological standpoint, peritoneal fibrosisis distinct from the process of duplication of theperitoneal basement membrane, which involves depos-ition of collagen IV, mimicking diabetic hyalinization.It is noteworthy that MC synthesize large amounts ofinterstitial type I and type III collagens, whose accumu-lation characterizes peritoneal fibrosis, while theyappear to produce only scattered amounts of type IV
collagen, which is directly responsible for peritonealhyalinization [14].
Currently available MC-culturing techniques enablethe mechanisms of regulation of collagen molecules tobe directly investigated in vitro [10]. Furthermore,progress in the field of molecular biology also allowsdirect analysis of collagen transcription. We tookadvantage of these facts to monitor any effect ofperitoneal effluents on collagen expression by MC 'inculture'. In particular, a major objective was to definethe role of peritoneal infection in mediating anincreased expression of interstitial collagens, whichshould precede widespread deposition and fibrosis.Indeed, clinical and anecdotal observations show thatepisodes of peritonitis are common in patients onperitoneal dialysis who develop peritoneal fibrosis. Thissuggests a role of infectious agents per se or of theoverall phenomena involved in peritonitis, e.g. poly-morphonuclear cells and macrophages infiltration,cytokines, etc. [15,16].
The results here presented demonstrate that cell-freeperitoneal effluents markedly affect several functionsof MC in vitro, including proliferation, and up-regulatethe expression of those interstitial collagens whichaccumulate in peritoneal fibrosis. The latter effect isbrought about by a strong induction of the related
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from

1808 F. Perfumo et al.
H-proline60000-j
50000-
40000-
30000-
20000-
10000-
0
incubation 48 hours
' H-thymidine
• control
• TNF-a
• TGF-6
E2 IL-26000-1
4000
3001)
2001H•
lOOd-i
a _ _
T 1
ii
1—
ji
_. i
Y/////•
incubation 48 hours
Fig. 5. Effect of TGF/?, TNFa and IL2 on proliferation(3H-thymidine incorporation) by mesothelial cells in culture.
mRNAs, which means a direct regulation of transcrip-tion of collagen. Both cellular phenomena, namely theinduction of proliferation and the regulation of colla-gen expression, can also be detected in sterile condi-tions but are maximal during endoperitoneal infectiousepisodes. A second point of interest is the markeddifference in cellular effects determined by differenteffluents deriving from a single dialytic procedure; sucheffects are maximal for the longest exposure times.This finding suggests that short interdialytic stasisand/or increased volume may be more appropriate.
Although the identity of the endoperitoneal sub-stance with fibrogenic activity is still to be clarified, afew hypotheses can be put forward. One of theseenvisions a role of cytokines and growth factors, whichregulate both cell proliferation and ECM expressionin many cell systems [17]. The list includes bFGF,TGF/?, TNF, interleukins, and eicosanoids, which maybe produced by polymorphonuclear cells and macro-phages in peritoneal cavity [7,15,16]. Our data, how-ever, indicate that among the most common cytokines,no candidate can explain the concomitant changes incell proliferation and collagen synthesis. Only TGF/?is able to upregulate the synthesis of collagen by MC;
at the same time, however, it inhibits cell proliferation.The most plausible explanation is the occurrence of amultifactorial regulation of cell functions [10]. Thepresence of osmotic solutes, such as glucose and aminoacids, in the medium is known to affect these autocrinephenomena as well as MC functions and other factors[18,19]. In conclusion, the data here presented demon-strate that in patients on peritoneal dialysis, bioactivesubstances are produced and released within the peri-toneal cavity and that this phenomenon is greatlyenhanced during infectious episodes. These unidentifiedsubstances stimulate cell proliferation and also upregul-ate the synthesis of collagen by MC in culture. It maytherefore be claimed that they play a role in periton-eal fibrosis.
Acknowledgements The authors are indebted to Mrs Silvana Varrialeand Alessandra Trivelli for their secretarial support in a hot Liguriansummer. This paper was presented at the meeting of EDTA inVienna and appeared as an abstract in Nephrology DialysisTransplantation. The study was supported by a grant from the ItalianMinistry of Health to G. Gaslini Institute.
References
1. Dobbie JW. Pathogenesis of peritoneal fibrosing syndromes(sclerosing peritonitis) in peritoneal dialysis. Peril Dial Int 1992;12: 14-27
2. Gotch FA. Adequacy of peritoneal dialysis. Am J Kidney Dis1993; 21: 96-98
3. Dobbie JW, Anderson JD, Hind C. Long-term effects of periton-eal dialysis on peritoneal morphology. Peril Dial hit 1994; 14[Suppl. 3]: 916-920
4. Dobbie JW. Monitoring peritoneal histopathology in peritonealdialysis: the role of a biopsy registry. Dial Transplant 1989;18: 319-335
5. Oules R, Challah S, Brunner FP. Case control study to determinethe cause of sclerosing peritoneal disease. Nephrol DialTransplant 1988; 3: 66-69
6. Goldman M, Vandenabeele P, Mulart J et al. Intraperitonealsecretion of interleukin-6 during continuous ambulatory periton-eal dialysis. Nephron 1990; 56: 277-280
7. Mackenzie RK, Coles GA, Williams JD. Eicosanoid synthesisin human peritoneal macrophages stimulated with S. epidermidis.Kidney Int 1990; 37: 1316-1324
8. Kovacs EJ. Fibrogenic cytokines: the role of immune mediatorsin the development of scar tissue. Immunol Today 1991; 12: 17-23
9. Topley N. Jorres A, Luttman W et al. Human peritonealmesothelial cells synthesize interleukin-6: induction by IL-I/? andTNFa. Kidney Int 1993; 43: 226-233
10. Stylianou E, Jenner LA, Davies M et al. Isolation, culture andcharacterization of human peritoneal mesothelial cells. KidneyInt 1990; 37: 1563-1570
11. Recommendations of the German Society for clinical chemistry.Z Klin Chem Biochem 1972; 10: 281-291
12. Laemmli UK. Cleavage of structural proteins during the assem-bly of the head bacteriophage T4. Nature 1970; 227: 680-685
13. Finger JM, Choo KH. Double-label reductive methylation oftissue proteins for precision two-dimensional polyacrylamide-gelelectrophoresis analysis. Biochem J 1981; 193: 371-374
14. Harvey W, Amlot PL. Collagen production by human mesothel-ial cells in vitro. J Pathol 1983; 139: 337-347
15. Fieren M, Van Den Bend G, Bonta IL. Endotoxin-stimulatedperitoneal macrophages obtained from continuous ambulatory
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from

Collagen expression by mesothelial cells
peritoneal dialysis patients show increased capacity to releaseinterleukin-I/? in vitro during peritonitis. Eur J Clin Invest 1990;20: 453-457
16. Andreoli SP, Langefeld CD, Stadler S ex al. Risks of peritonealmembrane failure in children undergoing long-term peritonealdialysis. Pediatr Nephrol 1993; 7: 543-547
1809
17. Breborowicz A, Rodela M, Oreopulos DG. Toxicity of osmoticsolutes on human mesothelial cells in vitro. Kidney Int 1992; 41:1280-1285
18. Brunkhorst R, Mahiout A. Piruvate neutralizes peritoneal dialys-ate cytotoxicity: maintained integrity and proliferation of cul-tured human mesothelial cells. Kidney Int 1995: 48: 177-181
Received for publication: 27.10.95Accepted in revised form: 17.5.96
by guest on July 6, 2011ndt.oxfordjournals.org
Dow
nloaded from
Related Documents











