Nephrol Dial Transplant (2012) 27: 937–946 doi: 10.1093/ndt/gfr459 Advance Access publication 19 August 2011 Alanyl–glutamine dipeptide restores the cytoprotective stress proteome of mesothelial cells exposed to peritoneal dialysis fluids Klaus Kratochwill 1 , Michael Boehm 1 , Rebecca Herzog 1 , Anton Michael Lichtenauer 1 , Elisabeth Salzer 1 , Michael Lechner 2 , Lilian Kuster 1 , Konstantin Bergmeister 1 , Andreas Rizzi 2 , Bernd Mayer 3 and Christoph Aufricht 1 1 Department of Pediatrics and Adolescent Medicine, Medical University of Vienna, Vienna, Austria, 2 Institute of Analytical Chemistry, University of Vienna, Vienna, Austria and 3 Institute for Theoretical Chemistry, University of Vienna, Vienna, Austria Correspondence and offprint requests to: Christoph Aufricht; E-mail: [email protected] Abstract Background. Exposure of mesothelial cells to peritoneal dialysis fluids (PDF) results in cytoprotective cellular stress responses (CSR) that counteract PDF-induced damage. In this study, we tested the hypothesis that the CSR may be inadequate in relevant models of peritoneal dialysis (PD) due to insufficient levels of glutamine, resulting in in- creased vulnerability against PDF cytotoxicity. We partic- ularly investigated the role of alanyl–glutamine (Ala-Gln) dipeptide on the cytoprotective PDF stress proteome. Methods. Adequacy of CSR was investigated in two human in vitro models (immortalized cell line MeT-5A and mesothe- lial cells derived from peritoneal effluent of uraemic patients) following exposure to heat-sterilized glucose-based PDF (PD4-Dianeal, Baxter) diluted with medium and, in a compa- rative proteomics approach, at different levels of glutamine ranging from depletion (0 mM) via physiological (0.7 mM) to pharmacological levels (8 mM administered as Ala-Gln). Results. Despite severe cellular injury, expression of cytopro- tective proteins was dampened upon PDF exposure at phys- iological glutamine levels, indicating an inadequate CSR. Depletion of glutamine aggravated cell injury and further reduced the CSR, whereas addition of Ala-Gln at pharmaco- logical level restored an adequate CSR, decreasing cellular damage in both PDF exposure systems. Ala-Gln specifically stimulated chaperoning activity, and cytoprotective processes were markedly enhanced in the PDF stress proteome. Conclusions. Taken together, this study demonstrates an in- adequate CSR of mesothelial cells following PDF exposure associated with low and physiological levels of glutamine, indicating a new and potentially relevant pathomechanism. Supplementation of PDF with pharmacological doses of Ala-Gln restored the cytoprotective stress proteome, resulting in improved resistance of mesothelial cells to exposure to PDF. Future work will study the clinical relevance of CSR- mediated cytoprotection. Keywords: alanyl–glutamine; cytoprotection; heat shock proteins; peritoneal dialysis; stress proteome Introduction Peritoneal dialysis (PD) is a cost-effective and safe form of renal replacement therapy in end-stage renal disease. How- ever, peritoneal dialysis fluids (PDF) are cytotoxic and a third of PD patients will suffer from technical failure [1, 2]. Current research aims to increase the biocompatibility of PDF and to thereby reduce mesothelial cell damage during PD. Although new and improved formulats have recently been shown to be less toxic in several in vitro and in vivo experimental and clinical studies, the main working prin- ciple of PDF requires the use of a hypertonic solution for solute and water removal [3–5]. PDF will therefore never represent a physiologically inert or completely biocompat- ible fluid and repeated filling and drainage of the abdominal cavity with PDF will always infer some cytotoxic insults. Research from our and other laboratories have demon- strated a subtle balance between cytotoxic injury and coun- teracting cellular stress responses (CSR) in mesothelial cells exposed to PDF: acute exposure to cytotoxic proper- ties of PDF resulted in induction of cytoprotective cellular responses in mesothelial cells during recovery, counteract- ing toxic injury [6–10]. Heat shock proteins (HSPs), as main effectors of this stress response, protected cellular survival and increased resistance against repeated injuries [10–12]. The HSP are among the most conserved protein families in living organisms. Functioning as molecular chaperones, they mainly bind to other proteins, assisting in correct folding or stabilization. HSP can constitute up to 5% of the cells total protein content [13]. Recently, we studied mesothelial HSP expression in a more continuous PDF exposure system using diluted sol- utions, more closely reflecting intraperitoneal conditions during the PD dwell phase [14, 15]. In that setting, PDF exposure for up to 24 h unexpectedly resulted in down- regulation of HSPs in cultured human omental, mesothelial cells from healthy donors [16]. Ó The Author 2011. Published by Oxford University Press on behalf of ERA-EDTA. All rights reserved. For Permissions, please e-mail: [email protected] by guest on April 11, 2016 http://ndt.oxfordjournals.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Nephrol Dial Transplant (2012) 27: 937–946
doi: 10.1093/ndt/gfr459
Advance Access publication 19 August 2011
Alanyl–glutamine dipeptide restores the cytoprotective stress proteomeof mesothelial cells exposed to peritoneal dialysis fluids
Klaus Kratochwill1, Michael Boehm1, Rebecca Herzog1, Anton Michael Lichtenauer1, Elisabeth Salzer1,Michael Lechner2, Lilian Kuster1, Konstantin Bergmeister1, Andreas Rizzi2, Bernd Mayer3 andChristoph Aufricht1
1Department of Pediatrics and Adolescent Medicine, Medical University of Vienna, Vienna, Austria, 2Institute of Analytical Chemistry,University of Vienna, Vienna, Austria and 3Institute for Theoretical Chemistry, University of Vienna, Vienna, Austria
Correspondence and offprint requests to: Christoph Aufricht; E-mail: [email protected]
AbstractBackground. Exposure of mesothelial cells to peritonealdialysis fluids (PDF) results in cytoprotective cellular stressresponses (CSR) that counteract PDF-induced damage. Inthis study, we tested the hypothesis that the CSR may beinadequate in relevant models of peritoneal dialysis (PD)due to insufficient levels of glutamine, resulting in in-creased vulnerability against PDF cytotoxicity. We partic-ularly investigated the role of alanyl–glutamine (Ala-Gln)dipeptide on the cytoprotective PDF stress proteome.Methods. Adequacy of CSR was investigated in two humanin vitro models (immortalized cell line MeT-5A and mesothe-lial cells derived from peritoneal effluent of uraemic patients)following exposure to heat-sterilized glucose-based PDF(PD4-Dianeal, Baxter) diluted with medium and, in a compa-rative proteomics approach, at different levels of glutamineranging from depletion (0 mM) via physiological (0.7 mM)to pharmacological levels (8 mM administered as Ala-Gln).Results. Despite severe cellular injury, expression of cytopro-tective proteins was dampened upon PDF exposure at phys-iological glutamine levels, indicating an inadequate CSR.Depletion of glutamine aggravated cell injury and furtherreduced the CSR, whereas addition of Ala-Gln at pharmaco-logical level restored an adequate CSR, decreasing cellulardamage in both PDF exposure systems. Ala-Gln specificallystimulated chaperoning activity, and cytoprotective processeswere markedly enhanced in the PDF stress proteome.Conclusions. Taken together, this study demonstrates an in-adequate CSR of mesothelial cells following PDF exposureassociated with low and physiological levels of glutamine,indicating a new and potentially relevant pathomechanism.Supplementation of PDF with pharmacological doses ofAla-Gln restored the cytoprotective stress proteome, resultingin improved resistance of mesothelial cells to exposure toPDF. Future work will study the clinical relevance of CSR-mediated cytoprotection.
Keywords: alanyl–glutamine; cytoprotection; heat shock proteins;peritoneal dialysis; stress proteome
Introduction
Peritoneal dialysis (PD) is a cost-effective and safe form ofrenal replacement therapy in end-stage renal disease. How-ever, peritoneal dialysis fluids (PDF) are cytotoxic and athird of PD patients will suffer from technical failure [1, 2].Current research aims to increase the biocompatibility ofPDF and to thereby reduce mesothelial cell damage duringPD. Although new and improved formulats have recentlybeen shown to be less toxic in several in vitro and in vivoexperimental and clinical studies, the main working prin-ciple of PDF requires the use of a hypertonic solution forsolute and water removal [3–5]. PDF will therefore neverrepresent a physiologically inert or completely biocompat-ible fluid and repeated filling and drainage of the abdominalcavity with PDF will always infer some cytotoxic insults.
Research from our and other laboratories have demon-strated a subtle balance between cytotoxic injury and coun-teracting cellular stress responses (CSR) in mesothelialcells exposed to PDF: acute exposure to cytotoxic proper-ties of PDF resulted in induction of cytoprotective cellularresponses in mesothelial cells during recovery, counteract-ing toxic injury [6–10]. Heat shock proteins (HSPs), asmain effectors of this stress response, protected cellularsurvival and increased resistance against repeated injuries[10–12]. The HSP are among the most conserved proteinfamilies in living organisms. Functioning as molecularchaperones, they mainly bind to other proteins, assistingin correct folding or stabilization. HSP can constitute up to5% of the cells total protein content [13].
Recently, we studied mesothelial HSP expression in amore continuous PDF exposure system using diluted sol-utions, more closely reflecting intraperitoneal conditionsduring the PD dwell phase [14, 15]. In that setting, PDFexposure for up to 24 h unexpectedly resulted in down-regulation of HSPs in cultured human omental, mesothelialcells from healthy donors [16].
� The Author 2011. Published by Oxford University Press on behalf of ERA-EDTA. All rights reserved.For Permissions, please e-mail: [email protected]
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

Comparable downregulation of HSP expression has beenfound in glutamine-starving cells under stressful conditions[17–19]. In these models, addition of glutamine (Gln) led toimproved cytoprotection and preservation of cell homeosta-sis, associated with activation of the CSR and increased HSPexpression [17–19]. Gln supplementation by the dipeptidealanyl–glutamine (Ala-Gln) is currently in wide clinical usein parenteral nutrition and was associated with improved clin-ical outcome in critically ill patients [17–19]. These reportsled us to the hypothesis that exposure to PDF with zero or lowGln levels may dampen cytoprotective cellular responses,resulting in an inadequate CSR that increases the vulnerabilityof mesothelial cells against PDF cytotoxicity and other insultsduring PD, which might be rescued by Gln supplementation.
Therefore, in this study, we investigated the inadequate CSRin a human in vitro model of extended PDF exposure. Pro-teome analysis represents a particularly powerful tool to definecellular responses upon PDF exposure, reflected by differentialprotein abundances. We hypothesized that inadequate Glnlevels disrupt the adequacy of the CSR in mesothelial cells.Using a proteomics approach, we compared exposure to PDFwith or without pharmacological levels of Gln, supplementedas Ala-Gln, to investigate the effects of this cytoprotectiveagent on the stress protein expression fingerprint.
Materials and methods
Standard chemicals were purchased from Sigma–Aldrich (St Louis, MO)if not specified otherwise.
Peritoneal effluent-derived mesothelial cell cultures
From June 2009 to January 2010, mesothelial cell cultures were grownfrom randomly collected peritoneal effluents in five male PD patients[aged 4.1 (range 0.1–11.7 years)] undergoing PD for 2–371 days withcommercial PD fluid. All patients were clinically stable and for >3 monthsfree of peritonitis at the time of effluent collection. Peritoneal cells wereconcentrated by centrifugation of dialysis fluid effluent and then culturedin Earle’s M199 medium supplemented to yield a final concentration of10% fetal calf serum (FCS), 50 U/mL penicillin and 50 lg/mL streptomy-cin. Non-adherent cells were removed after 24 h by two brief washes withmedium. When the primary mesothelial cultures reached confluence (after3–27 days), they were split two times and their morphologic features wereassessed microscopically. For each patient, a cobblestone-like epithelialculture was selected for the incubation experiments, showing stable mor-phology during two to three cell passages. Each experiment consisted ofthree independent observations in biological replicates on separate cultureplates in each of the five patients (n ¼ 15 per condition).
Immortalized human mesothelial cells
MeT-5A and ATCC-CRL-9444 (LGC Standards GmbH, Wesel, Ger-many) were cultured according to ATCC recommendations in M199 me-dium (M0650; Sigma–Aldrich) supplemented to yield 50 U/mL penicillin,50 lg/mL streptomycin as well as 5.958 g/L HEPES, 2.2 g/L NaHCO3, 0.1g/L Gln and 10% FCS. For continuous growth, the cultures were kept at37�C in 5% CO2 in 75 cm2 tissue culture flasks (TPP—Techno PlasticsProducts AG, Trasadigen, Switzerland). Cultures were passaged by tryp-sinization and passages 3–20 were used for experimental procedures. Dur-ing experimental procedures, the cultures grew on 12-well plates (TPP),the medium was changed every 2–3 days, and confluence was reachedbetween 5 and 7 days. Each experiment was repeated eight times (n ¼ 8).
Experimental PDF exposure setting
Mesothelial cells cultured in 12-well plates were incubated with one of theexperimental solutions mentioned below for up to 24 h. FCS was added toall final mixtures to yield a final concentration of 10%. All test fluids werefiltered with a 500 mL sterile filter unit (TPP). All cultures were kept at37�C. M199 medium was prepared from dry powder (M3769; Sigma–Aldrich), sodium bicarbonate and HEPES as prescribed by the manufacturer.
For PDF incubation, the commercially available 3.86% glucose basedPD4-Dianeal (Baxter AG, Vienna, Austria) was mixed either 1:1 or 2:1with medium. In a series of pilot experiments, it was determined thatdilution of medium components did not show effects on viability andHSP expression as long as the FCS level was maintained (data not shown).
To assess effects of Gln levels, Gln was either completely omitted(depletion) or the mixture contained a final concentration of 0.1 g/L Gln(¼0.7 mM) (physiological level) or was supplemented with the dipeptideAla-Gln to a final concentration of 1738 g/L Ala-Gln (¼8 mM Gln)(pharmacological level).
Normal media without PDF but with a final concentration of 0.1 g/LGln (¼0.7 mM) was set as the control. Control cultures underwent thesame ‘sham-procedures’ of fluid changes. This internal control was in-cluded as a condition in each experiment.
At the end of each protocol, the supernatant was saved for viabilityassessments, the cultures were washed three times with washing buffer(250 mM sucrose, 10 mM Tris/HCl (Merck KGaA, Darmstadt, Germany)and cells were subsequently shock frosted in liquid nitrogen and stored at�80�C for a maximum of 3 days.
Viability of cells
Viability of cells was investigated by lactate dehydrogenase (LDH) releaseand a LIVE/DEAD� viability/cytotoxicity kit (Invitrogen, MolecularProbes, Eugene, OR). For LDH analyses, 250 lL aliquots of supernatantswere removed as described and kept on �20�C until analysation within48 h. Measurements were performed in duplicates with Sigma TOX-7LDH Kit according to the manufacturer instructions. LDH efflux followingexposure to normal media without PDF but with 0.7 mM Gln was set as thecontrol, assigned 100% and data from other conditions with PDF with andwithout Gln were compared relative to this value.
For the LIVE/DEAD� viability/cytotoxicity kit, intracellular esteraseactivity (only live cells are stained) and plasma membrane integrity (onlydead cells are stained) were simultaneously stained with 250 lL of 4 lMcalcein AM and 2 lM ethidium homodimer (EthD-1) in PBS per well of astandard 12-well plate. The plates were then analysed using an LSM-780 laserscanning microscope (Carl Zeiss MicroImaging GmbH, Jena, Germany) withthe according settings for excitation (Ex) and emission (Em) wavelenghts ofcalcein AM (Ex/Em 494/517 nm) and EthD-1 (Ex/Em 528/617 nm).
Protein sample preparation
The cells were lysed by incubation with 1 mL lysis buffer [30 mM Tris, pH8.5, 7 M urea, 2 M thiourea, 4% 3-[(3-cholamidopropyl) dimethylammonio]-1-propanesulfonate (CHAPS), 1 mM ethylenediaminetetraacetic acid(EDTA), one tablet of Complete Protease Inhibitor (Roche, Basel; Switzer-land) per 100 mL and 10 lL/mL of each phosphatase inhibitor cocktail 1 and2 (Sigma–Aldrich)] per 33 107 cells for 10 min at 25�C. The resulting lysateswere centrifuged for 30 min (14 000 g, 4�C) and stored at�80�C until furtherprocessing. Total protein concentration was determined by the 2D-Quant kit(GE Healthcare, Uppsala, Sweden) according to the manufacturer’s manual.
Two-dimensional gel electrophoresis
Isoelectric focusing was conducted on a Protean� IEF Cell (Bio-Rad,Hercules, CA) with 200 lg of total protein per immobilized pH gradient(IPG) strip [ReadyStrip IPG Strips 24 cm pH 3–10 non-linear (Bio-Rad)].The rehydration mix containing the sample was brought to a final volumeof 450 lL and a final concentration of 5 M urea, 0.5%w/vol CHAPS,0.5%vol/vol Pharmalyte (GE Healthcare) and 12 lL/mL of DeStreak re-agent (GE Healthcare). The strips were actively rehydrated at 50 V for 12 hand afterward focused by increasing the voltage step by step up to 8000 V.The procedure was carried out under mineral oil at 20�C using a currentlimit of 30 lA/strip. Focused strips were stored at �80�C. For the seconddimension, the strips were consecutively incubated for 2 3 15 min in 10 mLequilibration buffer (6 M urea, 2%w/vol sodium dodecyl sulphate (SDS),25%w/vol glycerol, 3.3%vol/vol 50 mM Tris/HCl buffer pH 8.8, stained withbromphenolblue) first supplemented with 100 mg of dithiothreitol and thenwith 480 mg of 2-iodoacetamide. The following SDS–polyacrylamide gelelectrophoresis (PAGE) was accomplished using lab-cast gels (12%vol/vol
acrylamide, 0.375 M Tris/HCl pH 8.8) and a Dodeca Plucs cell (Bio-rad) at20�C with a current of 60 mA in the first phase of 100 Vh and then 200 mA inthe second phase until the tracking dye reached the end of the gel.
Visualization of proteins
Gels were stained using ruthenium II tris (bathophenanthroline disulfo-nate) (RuBPS) fluorescent dye (synthesized according to Rabilloud et al.
938 K. Kratochwill et al.
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

[20]) following an optimized protocol by Lamanda et al. [21]. In brief, gelswere fixed over night on a rotary shaker in an aqueous solution of 10%vol/vol
acetic acid and 30%vol/vol ethanol. After washing four times for 30 minwith 20%vol/vol ethanol, the gels were incubated for 6 h in RuBPS stainingsolution (50 lL/L stock solution in H2OUHQ) followed by two 10 minwashing steps in H2OUHQ and over night destaining in 10%vol/vol aceticacid and 40%vol/vol ethanol.
Image acquisition and data analysis
Gel images were aquired using a Typhoon Trio laser scanner (GE Health-care) at an excitation wavelength of 488 nm and an emission wavelengthof 610 nm. The images were analyzed using the Delta2D 3.6 software(Decodon GmbH, Greifswald, Germany) with group-wise image align-ment and spot detection on the resulting fused image. Protein identifica-tions of the PDF stress proteome from our recent work [22] accomplishedby mass spectrometry were reassigned from the original images to themaster image of the current study (see Supplemental Table S1). Normal-ized relative spot volumes (% volume) of these 60 marker proteins of thePDF stress proteome were quantified in the current extended exposuremodel with and without Ala–Gln. As the coefficient of variation (CV)was 9% for significantly altered spots (median overall CV 15%), proteinspots which showed a ratio in abundance increase >1.2 or a decrease <0.8compared control were considered as being differentially abundant. Cate-gorization of the proteins of interest in terms of biological processes wasderived from the PANTHER classification system (www.pantherdb.org).The level of activated biological processes was quantified by comparisonof the number of observed differentially abundant proteins to the expectednumber taking the total number of proteins assigned to each specific path-way as background (www.pantherdb.org). The expected number (set to1.0 for calculation of the ratio) represents the biological ‘normal’ state.Enrichment of proteins in a given process (yielding a ratio of >1.0) isinterpreted as activation, whereas depletion of proteins (yielding a ratio of<1.0) is interpreted as deactivation of this process. Significance values ofincreases or decreases in the number of proteins in enriched or depletedbiological processes upon treatment were calculated using Fisher’s exacttest or Chi-square test, where appropriate. P-values �0.05 were consideredas significant.
Western blotting
Equal amounts of protein lysates were separated by SDS–PAGE, on a Bio-Rad Criterion cell using Criterion precast 12.5% Tris–HCl gels of 1 mmthickness (Bio-Rad). Proteins were electroblotted onto polyvinylidene flu-oride (PVDF) membranes immediately after the run by tank blotting using aCriterion blotting cell (Bio-Rad) and the respective transfer buffer (200 mMglycine, 25 mM Tris–base, 0.1% SDS, 20% methanol). The membraneswere blocked with 5% dry milk in Tris-buffered saline containing 0.1%Tween-20 (TBST) and then incubated with the primary murine antibodyagainst Hsp72 (SPA-810, Stressgen/Assay Designs, Ann Arbor, MI),Hsp27 (SPA-801, Stressgen/Assay Designs) or beta-Tubulin (#691261;MP Biochemicals, Solon, OH) for 16 h. After incubation with secondary,peroxidase-coupled antibodies (Polyclonal Rabbit Anti-Mouse Ig/HRPP0260; Dako Cytomation, Carpinteria, CA) detection was accomplishedby using enhanced chemiluminescence solution (Western Lightning re-agent; Perkin Elmer, Boston, MA) and a ChemiDoc XRS chemilumines-cence detection system (Bio-Rad). Densitometric quantification of 1Dbands was accomplished using the Bio-Rad QuantityOne software, yieldingbackground-subtracted values of optical density that were in the linear rangefor the observed expression levels. Normal media without PDF but with 0.7mM Gln was set as the control, assigned 100% and data from other con-ditions with PDF with and without Gln were compared relative to this value.Values from different groups were compared using t-tests or ANOVAwhere appropriate using SPSS 17.0. In the case of ANOVA, Tukey’s testwas used as post-hoc test. P-values �0.05 were considered as significant.
Results
As shown in Figure 1, PDF exposure of mesothelial cellsresulted in low cytoprotective responses, as reflected byHSP expression, despite marked cellular injury, as reflectedby LDH release. Exposure of human cultured mesothelialcells (MeT-5A) to heat-sterilized glucose based PDF (mixed
Fig. 1. Influence of glutamine levels on cell injury and stress response inhuman mesothelial cells upon PDF exposure. In panel A, LDH release isshown as marker of cellular injury following exposure to medium or to 1:1dilution of medium with PDF. Western blot-based densitometric expres-sion profiles for Hsp72 are demonstrated in Panel B and for Hsp27 in PanelC. Compared to physiological levels (0.7 mM, light gray bar), depletion ofGln (0 mM, dark bar) results in exacerbated LDH release (Panel A) anddepressed levels of Hsp72 (Panel B) and Hsp27 (Panel C), whereas addi-tion of Ala-Gln at pharmacological levels (8 mM, hatched bars) results inreduced LDH release and increased levels of Hsp72 and Hsp27. Glnstarvation exacerbates cellular injury and HSP depression, whereas phar-macological supplementation of Ala-Gln improved adequacy of the CSR.Asterisks (*) indicate P < 0.05 versus control. All data are related to theinternal control (set as 100%), included as condition in each experiment,i.e. exposure to normal medium without PDF at physiological levels ofGln (0.7 mM). The data are representative for eight experiments withMeT-5A cells (n ¼ 8).
Cytoprotective effects of alanyl–glutamine 939
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

1:1 with medium) at physiological Gln levels (0.7 mM)resulted in LDH release of 387 � 93% relative to control(P < 0.05 versus control). Despite this severe cellular in-jury, counteracting Hsp72 expression was decreased to92.6 � 6.9% (P < 0.05 versus control) and Hsp27 expres-sion was decreased to 91.4 � 6.9% (P ¼ not significant(n.s.) versus control).
To investigate the influence of varying Gln levels on cellsurvival and the CSR, mesothelial cells were additionallyexposed in vitro to 1:1 PDF:medium mixture under com-plete Gln depletion or with addition of Ala-Gln dipeptide(final Gln concentration 8 mM). As shown in Figure 1,depletion of Gln resulted in increased mesothelial cell vul-nerability and further decreased HSP expression, whereasaddition of Ala-Gln at pharmacological levels resulted innormalized cellular survival and increased HSP expression:Following Gln depletion, LDH release increased to 601 �55% (P < 0.05 versus control), associated with depressedHsp72 and Hsp27 levels of 89.6 � 10.7% and 75.9 �15.8%, respectively (both: P < 0.05 versus control). Incontrast, pharmacological Gln supplementation (8 mMGln administered as Ala-Gln) decreased LDH release to94 � 74% (n.s. versus control), associated with increasedHsp72 and Hsp27 levels of 111.7 � 13.0% and 180 �43.9%, respectively (both: P < 0.05 versus control).
Using two-dimensional gel electrophoresis (2DGE), theproteome of mesothelial cells (MeT-5A) was assessed. Thereason for using this mesothelial cell line for proteomic-type experiments is the excellent reproducibility and com-parability to previously published 2DGE results. Sixtymarker proteins recently identified as PDF stress proteome(see Supplemental Table S1) were analysed for changes inabundance following exposure to 1:1 PDF:medium mixtureat ‘physiological’ Gln levels. As given in Table 1, only 10of the 60 marker proteins were changed in abundance.Gene ontology information from the PANTHER databasewas exploited to categorize the proteins according to thebiological processes they are involved in. In Figure 2A, thelevel of activated biological processes was quantified bycomparison of the number of observed differentially abun-dant proteins to the number expected for this size of dataset‘Protein folding’ was the only enriched (activated) process.All other processes were either maintained at control levelsor even depleted (<50% of the expected proteins werefound, yielding a ratio of <0.5). Figure 2B shows that only1 of 10 marker proteins assigned to the biological process‘stress response’ showed differential abundance versuscontrol.
Upon addition of Ala-Gln (8 mM) to PDF, the number ofdifferentially expressed proteins in the PDF stress pro-teome increased from 10 to 21 of the 60 marker proteins(17–35%; Chi-square test P-value ¼ 0.022) (see Table 1).As shown in Figure 3A, addition of Ala-Gln did not resultin a general, non-specific enrichment of biological pro-cesses, but in specifically enhanced chaperoning activity.The processes stress response, ‘protein complex assembly’and protein folding were markedly enriched (activated),indicated by the increased number of differentially regu-lated proteins assigned to these processes (see also Table 1).‘Carbohydrate metabolism’ was maintained at controllevels, ‘cell structure’ appeared activated, ‘immunity and
defense’, ‘cell structure and motility’ and ‘protein metabo-lism and modification’ returned to the biological normalstate. Only ‘signal transduction’, ‘cell cycle’ and ‘proteinmodification’ remained depleted, likely reflecting the gen-eral effects of PDF stress. Under these conditions, 7 of the10 marker proteins assigned to the biological process stressresponse showed differential abundance (Figure 3B).
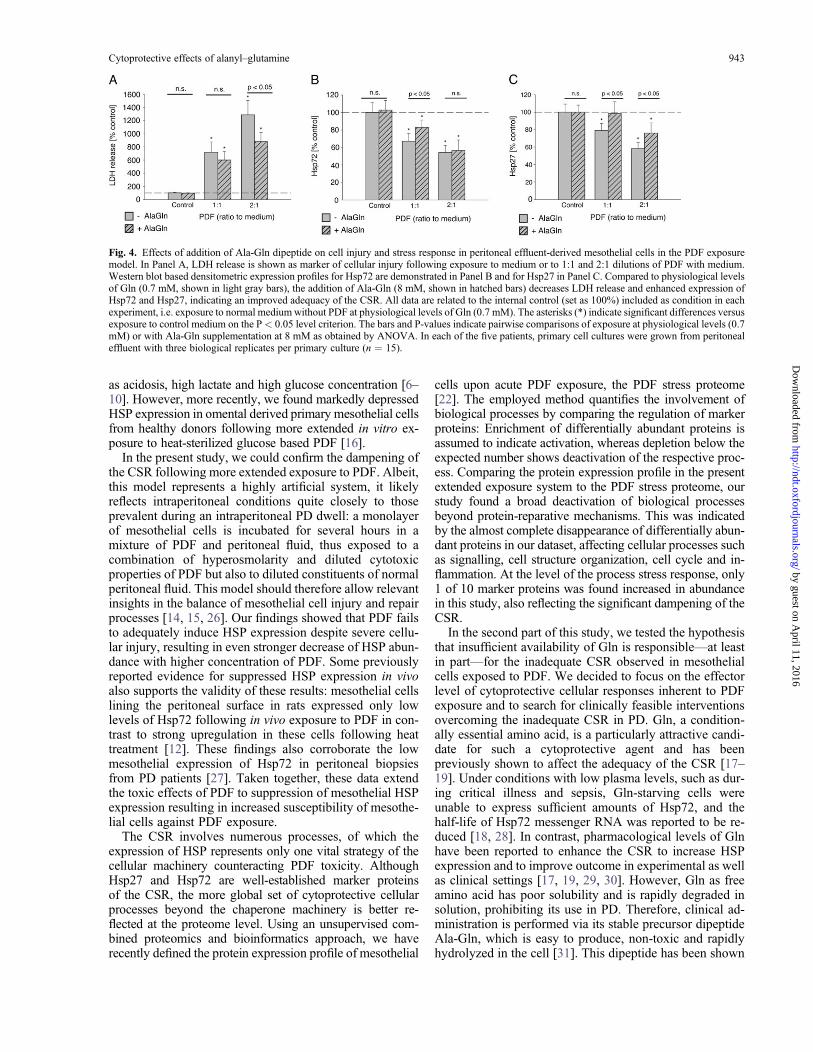
Finally, we validated these cytoprotective effects of Glnsupplementation during in vitro PDF exposure in the bio-logically relevant ex vivo system of human peritoneal mes-othelial cells derived from the dialyzed peritoneal cavity ofPD patients. As shown in Figure 4, at ‘physiological Glnlevels’, LDH release relative to control increased to 726 �155% upon exposure to 1:1 PDF:medium mixture and to1286 � 236% upon exposure to 2:1 PDF:medium mixture(for both changes: P < 0.05 versus control). Despite thissevere cellular injury, counteracting Hsp72 expression wasdecreased to 67.1 � 9.2% and 54.1 � 8.4%, (both: P <0.05 versus control) and Hsp27 expression was decreasedto 78.9 � 8.0% and 58.3 � 7.1%, respectively (both: P <0.05 versus control).
The addition of ‘pharmacological levels’ of Ala-Gln toPDF decreased cellular damage and enhanced HSP expres-sion in peritoneal effluent-derived primary mesothelialcells. As shown in Figure 4, LDH release was attenuatedto 601 � 130% during exposure to 1:1 PDF:medium mix-ture (P < 0.05 versus control, n.s. versus PDF exposure at‘physiological’ Gln levels), and to 879 � 144% duringexposure to 2:1 PDF:medium mixture (P <0.05 versus con-trol, P < 0.05 versus PDF exposure at physiological Glnlevels). This reduced cellular damage was associated withHsp72 expression levels of 83.0 � 8.3% following exposureto 1:1 PDF:medium mixture (P <0.05 versus control, P <0.05 versus PDF exposure at physiological Gln levels) and of56.7 � 11.8 following exposure to 2:1 PDF:medium mixture(P < 0.05 versus control, n.s. versus PDF exposure at phys-iological Gln levels). Hsp27 expression levels improved to98.4 � 13.6 following exposure to 1:1 PDF:medium mixture(n.s. versus control, P < 0.05 versus PDF exposure at phys-iological Gln levels) and to 76.1 � 11.6 following exposure to2:1 PDF:medium mixture (P < 0.05 versus control, P < 0.05versus PDF exposure at physiological Gln levels). Individualdata for each of the five patients is given in SupplementalFigures 1 and 2.
Figure 5 compares the effects of Ala-Gln supplementa-tion during PDF exposure between the mesothelial cellline (MeT-5a) and the primary mesothelial cells grownfrom PD effluates: as shown in the western blot analysis,exposure to PDF mixed 1:1 with medium at physiologicallevels of Gln resulted in reduced HSP expression in bothcell types that was restored by supplementation of PDFwith pharmacological levels of Ala-Gln. Comparing thedata shown in Figure 1 to those shown in Figure 4, Hsp72expression levels increased by 20.6 � 9.5% in MeT-5A ver-sus 23.7 � 2.8% in primary cells and Hsp27 expression levelsincreased by 97.0 � 31.6% in MeT-5A versus 24.6 � 3.0in primary cells upon pharmacological Gln supplementation(8 mM Gln administered as Ala-Gln). As shown in the live/dead analysis, exposure to PDF mixed 1:1 with medium atphysiological levels of Gln resulted in reduced cellular via-bility in both cell types that was improved by supplementation
940 K. Kratochwill et al.
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

Table 1. Differentially abundant marker proteins of the PDF stress proteome of mesothelial cells after extended exposure to PDF in dependence of Ala-Gln supplementation as compared to exposure to controlmedium at physiological glutamine levels
Protein nameSwissProtentry name
Ala-Glna
Cellstructure
Cellstructureandmotility
Carbohydratemetabolism
Stressresponse
Immunityanddefense
Proteinfolding
Proteinmodification
Proteincomplexassembly
Proteinmetabolismandmodification
Cellcycle
Signaltransduction� 1
Protein phosphatasemethylesterase 1
PPME1_HUMAN x x
Trifunctional enzyme subunitalpha,mitochondrial
ECHA_HUMAN x x
Lamin-B1 LMNB1_HUMAN x x14-3-3 protein epsilon 1433E_HUMAN x xEndoplasmin, HSP 90 kDa beta ENPL_HUMAN x x x xHeat shock 70 kDa protein 4 HSP74_HUMAN x xSerpin H1 SERPH_HUMAN xTropomyosin alpha-1 chain TPM1_HUMAN x xIsocitrate dehydrogenase[NADP] cytoplasmic
IDHC_HUMAN x
Serine–threonine kinasereceptor-associated protein
STRAP_HUMAN x
Stress-70 protein, mitochondrial GRP75_HUMAN x x x x xHeat shock 70 kDa protein 1 HSP71_HUMAN x x x x xHSP beta-1 HSPB1_HUMAN x x x xNeutral alpha-glucosidase AB GANAB_HUMAN x x xCytochrome c-type heme lyase CCHL_HUMAN x xProcollagen-lysine,2-oxoglutarate5-dioxygenase 3
PLOD3_HUMAN x
Tropomyosin alpha-3 chain TPM3_HUMAN x xMoesin MOES_HUMAN x xRadixin RADI_HUMAN x xEzrin EZRI_HUMAN x xGelsolin GELS_HUMAN x xAnnexin A2 ANXA2_HUMAN xAlpha-enolase ENOA_HUMAN xFructose-bisphosphate aldolase A ALDOA_HUMAN xProtein C14orf166 CN166_HUMAN xDifferentially abundant proteins(� Ala-Gln)
2 2 2 1 1 2 1 0 5 1 2
Differentially abundant proteins(1 Ala-Gln)
6 7 4 4 4 5 2 2 9 1 1
Differentially abundant proteins(expected)b
2.7 4.5 2.3 0.8 5.2 0.7 4.6 0.3 12 4 13.4
aExtended exposure to PDF at physiological (0.7 mM) glutamine levels (- Ala-Gln) or with addition of Ala-Gln (8 mM) (1Ala-Gln).bAssigned biological processes (marked by x) and the expected number of differentially abundant proteins for that size of dataset was obtained from the PANTHER database.
Cytoprotective
effectsof
alanyl–glutamine
941
by guest on April 11, 2016 http://ndt.oxfordjournals.org/ Downloaded from

of PDF with pharmacological levels of Ala-Gln. Comparingthe LDH data shown in Figure 1 to those shown in Figure 4,PDF-induced LDH release decreased by 75.6 � 12.5% in theMeT-5a cell line versus 16.4 � 18.1% in primary cells uponpharmacological Gln supplementation. Addition of Ala-Glnresulted in significantly improved adequacy of the CSR andcytoprotection against PDF exposure in both cell types.
Discussion
CSR is a highly conserved adaptive response to diverseenvironmental and physiological stressors, classically suchas temperature, chemical toxicants, energy depletion andmechanical stress [23]. The CSR results in the immediateinduction of effector proteins such as HSPs to prevent theimproper folding of newly translated proteins, and, in
response to stress, to protect critical stabilization of proteins[23]. HSPs and other molecular chaperones, the major sub-group of effector proteins involved in cytoprotection, rep-resent a large family of soluble proteins that can constituteup to 5% of total cellular proteins [24].
Due to their position as barrier between cytotoxic PDFand the peritoneal wall, mesothelial cells are exposed torepeated metabolic, osmotic and biomechanical insults dur-ing PD that can result in severe functional and/or structuralinjuries [1, 2, 25]. In earlier studies, we have demonstratedthat the cytotoxic properties of PDF not only result in dis-ruption of cell homeostasis but also induce the CSR: theexpression of Hsp27 and Hsp72, the best described HSPs inhumans, were upregulated in mesothelial cells followingrecovery from acute short-term exposure to stressors such
Fig. 2. Effects of PDF exposure at physiological glutamine levels on thestress proteome of mesothelial cells. Panel A represents the extent ofenrichment of biological processes with differentially abundant proteinsin mesothelial cells exposed to a 1:1 dilution of medium with PDF. Anal-ysis of the mesothelial stress proteome reveals a reduced activation ofbiological processes. In Panel B, the bars represent the expression profileof 10 proteins and their isoforms assigned to the process ‘stress response’following exposure to 1:1 diluted PDF compared to exposure to controlmedium at physiological levels of Gln (0.7 mM). Only 1 of 10 markerproteins involved in this biological process was enhanced in abundance asindicated by the hash mark (#). The dashed line represents the controllevel. The data are representative for eight experiments using MeT-5Acells (n ¼ 8).
Fig. 3. Effects of the addition of Ala-Gln at pharmacological doses on thestress proteome of mesothelial cells in the PDF exposure model. Panel Ashows the extent of enrichment of biological processes with differentiallyabundant proteins in mesothelial cells exposed to a 1:1 dilution of PDFwith medium. The addition of Ala-Gln specifically improved chaperoningactivity, as reflected by selected enrichment of those biological processesthat are involved in cytoprotection. In Panel B, the bars represent theexpression profile of 10 proteins and their isoforms assigned to stressresponse following exposure to 1:1 diluted PDF with supraphysiologicallevels of Ala-Gln (hatched bar) compared to exposure to control mediumat physiological levels of Gln (0.7 mM). Seven of 10 marker proteinsinvolved in this biological process were found as enhanced in abundanceas indicated by the hash mark (#). The dashed line represents the controllevel. The data are representative for eight experiments using MeT-5Acells (n ¼ 8).
942 K. Kratochwill et al.
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from
as acidosis, high lactate and high glucose concentration [6–10]. However, more recently, we found markedly depressedHSP expression in omental derived primary mesothelial cellsfrom healthy donors following more extended in vitro ex-posure to heat-sterilized glucose based PDF [16].
In the present study, we could confirm the dampening ofthe CSR following more extended exposure to PDF. Albeit,this model represents a highly artificial system, it likelyreflects intraperitoneal conditions quite closely to thoseprevalent during an intraperitoneal PD dwell: a monolayerof mesothelial cells is incubated for several hours in amixture of PDF and peritoneal fluid, thus exposed to acombination of hyperosmolarity and diluted cytotoxicproperties of PDF but also to diluted constituents of normalperitoneal fluid. This model should therefore allow relevantinsights in the balance of mesothelial cell injury and repairprocesses [14, 15, 26]. Our findings showed that PDF failsto adequately induce HSP expression despite severe cellu-lar injury, resulting in even stronger decrease of HSP abun-dance with higher concentration of PDF. Some previouslyreported evidence for suppressed HSP expression in vivoalso supports the validity of these results: mesothelial cellslining the peritoneal surface in rats expressed only lowlevels of Hsp72 following in vivo exposure to PDF in con-trast to strong upregulation in these cells following heattreatment [12]. These findings also corroborate the lowmesothelial expression of Hsp72 in peritoneal biopsiesfrom PD patients [27]. Taken together, these data extendthe toxic effects of PDF to suppression of mesothelial HSPexpression resulting in increased susceptibility of mesothe-lial cells against PDF exposure.
The CSR involves numerous processes, of which theexpression of HSP represents only one vital strategy of thecellular machinery counteracting PDF toxicity. AlthoughHsp27 and Hsp72 are well-established marker proteinsof the CSR, the more global set of cytoprotective cellularprocesses beyond the chaperone machinery is better re-flected at the proteome level. Using an unsupervised com-bined proteomics and bioinformatics approach, we haverecently defined the protein expression profile of mesothelial
cells upon acute PDF exposure, the PDF stress proteome[22]. The employed method quantifies the involvement ofbiological processes by comparing the regulation of markerproteins: Enrichment of differentially abundant proteins isassumed to indicate activation, whereas depletion below theexpected number shows deactivation of the respective proc-ess. Comparing the protein expression profile in the presentextended exposure system to the PDF stress proteome, ourstudy found a broad deactivation of biological processesbeyond protein-reparative mechanisms. This was indicatedby the almost complete disappearance of differentially abun-dant proteins in our dataset, affecting cellular processes suchas signalling, cell structure organization, cell cycle and in-flammation. At the level of the process stress response, only1 of 10 marker proteins was found increased in abundancein this study, also reflecting the significant dampening of theCSR.
In the second part of this study, we tested the hypothesisthat insufficient availability of Gln is responsible—at leastin part—for the inadequate CSR observed in mesothelialcells exposed to PDF. We decided to focus on the effectorlevel of cytoprotective cellular responses inherent to PDFexposure and to search for clinically feasible interventionsovercoming the inadequate CSR in PD. Gln, a condition-ally essential amino acid, is a particularly attractive candi-date for such a cytoprotective agent and has beenpreviously shown to affect the adequacy of the CSR [17–19]. Under conditions with low plasma levels, such as dur-ing critical illness and sepsis, Gln-starving cells wereunable to express sufficient amounts of Hsp72, and thehalf-life of Hsp72 messenger RNA was reported to be re-duced [18, 28]. In contrast, pharmacological levels of Glnhave been reported to enhance the CSR to increase HSPexpression and to improve outcome in experimental as wellas clinical settings [17, 19, 29, 30]. However, Gln as freeamino acid has poor solubility and is rapidly degraded insolution, prohibiting its use in PD. Therefore, clinical ad-ministration is performed via its stable precursor dipeptideAla-Gln, which is easy to produce, non-toxic and rapidlyhydrolyzed in the cell [31]. This dipeptide has been shown
Fig. 4. Effects of addition of Ala-Gln dipeptide on cell injury and stress response in peritoneal effluent-derived mesothelial cells in the PDF exposuremodel. In Panel A, LDH release is shown as marker of cellular injury following exposure to medium or to 1:1 and 2:1 dilutions of PDF with medium.Western blot based densitometric expression profiles for Hsp72 are demonstrated in Panel B and for Hsp27 in Panel C. Compared to physiological levelsof Gln (0.7 mM, shown in light gray bars), the addition of Ala-Gln (8 mM, shown in hatched bars) decreases LDH release and enhanced expression ofHsp72 and Hsp27, indicating an improved adequacy of the CSR. All data are related to the internal control (set as 100%) included as condition in eachexperiment, i.e. exposure to normal medium without PDF at physiological levels of Gln (0.7 mM). The asterisks (*) indicate significant differences versusexposure to control medium on the P < 0.05 level criterion. The bars and P-values indicate pairwise comparisons of exposure at physiological levels (0.7mM) or with Ala-Gln supplementation at 8 mM as obtained by ANOVA. In each of the five patients, primary cell cultures were grown from peritonealeffluent with three biological replicates per primary culture (n ¼ 15).
Cytoprotective effects of alanyl–glutamine 943
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

to be advantageous compared to other Gln-releasing dipep-tides regarding its uptake and intracellular Gln availability[32, 33].
In our study, Gln depletion also aggravated HSP depres-sion in mesothelial cells and resulted in increased vulner-ability against PDF exposure. In contrast, addition of Glnat a pharmacological level via the dipeptide Ala-Glnenhanced mesothelial HSP expression and improved cellu-lar survival, confirming our previous findings in the acutePDF exposure system [34]. Interestingly, supraphysiolog-ical doses of Gln conferred stronger cytoprotection in our
extended PDF exposure model than simple supplementa-tion of physiological levels. These findings are in agree-ment with the hypothesis of increased needs of Gln understressful conditions, likely resulting in relative Gln defi-ciency at the cellular levels [18, 28]. Comparable rates oftransperitoneal transport have been reported for Gln andcreatinine due to similar molecular weights and chargestates [35]. Peritoneal levels of Gln can therefore be ex-pected to be lower than the corresponding plasma levelslikely resulting in actual Gln starvation of peritoneal cellsin the clinical setting of PD.
Fig. 5. Comparison of cytoprotective effects of Ala-Gln at pharmacological doses between the MeT-5A mesothelial cell line and peritoneal effluent-derived primary mesothelial cell cultures in the PDF exposure model. As shown in the western blot analysis in Panel A, exposure to PDF mixed 1:1 withmedium at physiological levels of Gln (0.7 mM) resulted in reduced HSP expression in both cell types that was restored by supplementation of PDF withpharmacological levels of Ala-Gln (8 mM). As shown in the live/dead analysis in Panel B, exposure to PDF mixed 1:1 with medium at physiologicallevels of Gln (0.7 mM) resulted in reduced cellular viability in both cell types that was improved by supplementation of PDF with pharmacological levelsof Ala-Gln (8 mM). The data for western blot analysis are representative for eight experiments with MeT-5A cells (n¼ 8) and experiments in triplicates inindependent cultures from five donors (n ¼ 15) in primary cells. The data for live/dead analysis are representative for four experiments (n ¼ 4) for eachcell type.
944 K. Kratochwill et al.
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

Addition of Ala-Gln ameliorated the dampening of cy-toprotective responses during PDF exposure, analysed byproteomic analysis of involved biological processes, ex-plaining at least in part the increased survival of mesothe-lial cells. Although the addition of Ala-Gln dipeptide alsoresulted in a higher enrichment in unspecific biologicalprocesses, the majority of proteins with restored expressionprofiles were involved in chaperoning processes, in partic-ular the stress response, ‘protein complex assembly’ andprotein folding. In the biological process stress response, asignificantly higher number of marker proteins demonstrateddifferential expression following PDF exposure at pharma-cological Gln levels, reflecting a release of the dampening ofthis cellular response. Therefore, our findings clearly supporta specific role of Ala–Gln supplementation on the cellularprotein repair machinery in experimental PD.
Despite the obvious therapeutic potential of CSR-mediatedcytoprotection, there are only few reports on successfultranslation of this approach into clinically relevant models[23, 30]. PD should represent a unique opportunity for theimplementation of such a therapeutic concept, as PD isstrictly based on predictable, perfectly timed and dosed‘repetitive insults’ to the peritoneal membrane. In thisstudy, cytoprotection was tested in two different mesothe-lial cell types, the MeT-5A cell line, in which most of ourprevious CSR research had been performed, and a biolog-ically more relevant primary cell system derived from thedialysed peritoneal cavity of PD patients. Both mesothelialcell types demonstrated clear evidence for a suppressedCSR during injurious exposure to cytotoxic hyperosmolarPDF, the primary cells showed a higher vulnerability and amore dampened HSP expression. In both cell types, addi-tion of Ala-Gln to PDF ex vivo restored HSP expression,which is associated with improved cellular integrity. Ourresults therefore extend previous reports on cytoprotectionin the acute setting of PDF exposure, where addition ofcytoprotective additives had resulted in further enhance-ment of an upregulated CSR [34, 36]. As a novel finding,our study suggests that restoring the adequacy of a re-pressed CSR might reduce peritoneal damage by increasingresistance to PD related injury. Previous clinical researchfurther increases the attractivity of this approach as paren-teral Ala-Gln supplementation was associated with in-creased cellular expression of HSPs and better survival incritically ill patients [17, 28, 29].
Certainly, the design of this study does not allow theexclusion of other potentially cytoprotective effects ofAla-Gln: restoration of Gln levels has been also reportedto preserve cell homeostasis, to alter immune regulationand to increase antioxidant capacity via the glutathionepathway [37]. In particular, the latter mechanism mightbe relevant in PD, as evidenced by addition of the gluta-thione precursor procysteine to PDF [38]. Gln has also beenreported to lead to the activation of intracellular signallingpathways including differential regulation of genes relatedto metabolism, apoptosis and signal transduction [39] How-ever, the precise functional mechanism of Gln-mediatedcytoprotection remains to be validated in deepened mo-lecular biological experiments, such as effects of knock-out and knockdown of key regulators of the CSR inmesothelial cells upon PDF exposure.
Taken together, the results of this study confirm the in-adequate CSR of mesothelial cells following PDF exposureas a new and potential relevant pathomechanism in PD.Depressed HSP expression caused by—at least relative—Gln starvation increased the susceptibility of mesothelialcells during in vitro PD. Supplementation of PDF withpharmacological doses of Ala-Gln increased HSP expres-sion and restored the cytoprotective stress proteome, re-sulting in improved resistance of mesothelial cells to invitro exposure to PDF. Future work is needed to study theclinical relevance of CSR-mediated cytoprotection and tofurther elucidate the underlying regulatory cytoprotectivecellular mechanisms of Ala-Gln.
Supplementary data
Supplementary data are available online at http://ndt.oxfordjournals.org
Acknowledgements. This research work was in part supported by the FWF(Austrian Science Fund) Project P18130-B13 (to C.A.).
Figure 5 was performed with the technical advice and help from theImaging Unit (Head: Marion Groeger) of the Anna Spiegel Center ofTranslational Research.
We are grateful for the technical support by Astrid Scherr in preparingthe manuscript and figures.
Conflict of interest statement. C.A. is co-founder of Zytoprotec GmbH,aspin-off of the Medical University Vienna that holds the patentCarbohydrate-based PDF comprising glutamine residue (InternationalPublicationNumber: WO 2008/106702 A1).
References
1. Davies SJ, Phillips L, Griffiths AM et al. What really happens to peopleon long-term peritoneal dialysis? Kidney Int 1998; 54: 2207–2217
2. Devuyst O, Topley N, Williams JD. Morphological and functionalchanges in the dialysed peritoneal cavity: impact of more biocompat-ible solutions. Nephrol Dial Transplant 2002; 17 (Suppl 3): 12–15
3. Aufricht C, Ardito T, Thulin G et al. Heat-shock protein 25 inductionand redistribution during actin reorganization after renal ischemia. AmJ Physiol 1998; 274: F215–F222
4. Aufricht C, Lu E, Thulin G et al. ATP releases HSP-72 from proteinaggregates after renal ischemia. Am J Physiol 1998; 274: F268–F274
5. Arbeiter K, Greenbaum L, Balzar E et al. Reproducible erythroid aplasiacaused by mycophenolate mofetil. Pediatr Nephrol 2000; 14: 195–197
6. Arbeiter K, Bidmon B, Endemann M et al. Peritoneal dialysate fluidcomposition determines heat shock protein expression patterns inhuman mesothelial cells. Kidney Int 2001; 60: 1930–1937
7. Arbeiter K, Bidmon B, Endemann M et al. Induction of mesothelialHSP-72 upon in vivo exposure to peritoneal dialysis fluid. Perit DialInt 2003; 23: 499–501
8. Aufricht C, Endemann M, Bidmon B et al. Peritoneal dialysis fluidsinduce the stress response in human mesothelial cells. Perit Dial Int2001; 21: 85–88
9. Ruffingshofer D, Endemann M, Arbeiter K et al. Induction of heatshock protein 72 in mesothelial cells exposed to peritoneal dialysateeffluent. Perit Dial Int 2003; 23: 74–77
10. Kuo HT, Chen HW, Hsiao HH et al. Heat shock response protectshuman peritoneal mesothelial cells from dialysate-induced oxidativestress and mitochondrial injury. Nephrol Dial Transplant 2009; 24:1799–1809
11. Bidmon B, Endemann M, Arbeiter K et al. Overexpression of HSP-72confers cytoprotection in experimental peritoneal dialysis. Kidney Int2004; 66: 2300–2307
Cytoprotective effects of alanyl–glutamine 945
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from

12. Endemann M, Bergmeister H, Bidmon B et al. Evidence for HSP-mediated cytoskeletal stabilization in mesothelial cells during acuteexperimental peritoneal dialysis. Am J Physiol Renal Physiol 2007;292: F47–F56
13. Morimoto RI. Cells in stress: transcriptional activation of heat shockgenes. Science 1993; 259: 1409–1410
14. Topley N. What is the ideal technique for testing the biocompatibilityof peritoneal dialysis solutions? Perit Dial Int 1995; 15: 205–209
15. Holmes CJ. Pre-clinical biocompatibility testing of peritoneal dialysissolutions. Perit Dial Int 2000; 20 (Suppl 5): S5–S9
16. Bender TO, Bohm M, Kratochwill K et al. Peritoneal dialysis fluidscan alter HSP expression in human peritoneal mesothelial cells. Neph-rol Dial Transplant 2011; 26: 1046–1052
17. Wischmeyer PE. Glutamine and heat shock protein expression.Nutrition 2002; 18: 225–228
18. Roth E. Nonnutritive effects of glutamine. J Nutr 2008; 138:S2025–S20–31
19. Weitzel LR, Wischmeyer PE. Glutamine in critical illness: the timehas come, the time is now. Crit Care Clin 2010; 26: 515–525, ix-x
20. Rabilloud T, Strub JM, Luche S et al. A comparison between syproruby and ruthenium II tris (bathophenanthroline disulfonate) as fluo-rescent stains for protein detection in gels. Proteomics 2001; 1:699–704
21. Lamanda A, Zahn A, Roder D et al. Improved ruthenium II tris (bath-ophenantroline disulfonate) staining and destaining protocol for abetter signal-to-background ratio and improved baseline resolution.Proteomics 2004; 4: 599–608
22. Kratochwill K, Lechner M, Siehs C et al. Stress responses and con-ditioning effects in mesothelial cells exposed to peritoneal dialysisfluid. J Proteome Res 2009; 8: 1731–1747
23. Aufricht C. Heat-shock protein 70: molecular supertool? PediatrNephrol 2005; 20: 707–713
24. Morimoto RI. The Biology of Heat Shock Proteins and MolecularChaperones. Cold Spring Harbor: Laboratory Press, 1994,1–594
25. Jorres A, Topley N, Gahl GM. Biocompatibility of peritoneal dialysisfluids. Int J Artif Organs 1992; 15: 79–83
26. Topley N. Peritoneal dialysis solution biocompatibility testing: a real-istic alternative? Perit Dial Int 2005; 25: 348–351
27. Shioshita K, Miyazaki M, Ozono Y et al. Expression of heat shockproteins 47 and 70 in the peritoneum of patients on continuous am-bulatory peritoneal dialysis. Kidney Int 2000; 57: 619–631
28. Eliasen MM, Brabec M, Gerner C et al. Reduced stress tolerance ofglutamine-deprived human monocytic cells is associated with selec-tive down-regulation of Hsp70 by decreased mRNA stability. J MolMed 2006; 84: 147–158
29. Morrison AL, Dinges M, Singleton KD et al. Glutamine’s protectionagainst cellular injury is dependent on heat shock factor-1. Am JPhysiol Cell Physiol 2006; 290: C1625–C1632
30. Westerheide SD, Morimoto RI. Heat shock response modulators astherapeutic tools for diseases of protein conformation. J Biol Chem2005; 280: 33097–33100
31. Furst P, Albers S, Stehle P. Glutamine-containing dipeptides in paren-teral nutrition. JPEN J Parenter Enteral Nutr 1990; 14: S118–S1–24
32. Hubl W, Druml W, Roth E et al. Importance of liver and kidney forthe utilization of glutamine-containing dipeptides in man. Metabolism1994; 43: 1104–1107
33. Ollenschlager G, Simmel A, Roth E. Availability of glutamine frompeptides and acetylglutamine for human tumor-cell cultures. Metab-olism 1989; 38: 40–42
34. Bender TO, Bohm M, Kratochwill K et al. Hsp-mediated cytoprotec-tion of mesothelial cells in experimental acute peritoneal dialysis.Perit Dial Int 2010; 30: 294–299
35. Nakao T, Ogura M, Takahashi H et al. Charge-affected transperito-neal movement of amino acids in CAPD. Perit Dial Int 1996; 16(Suppl 1): S88–S90
36. Boehm M, Bergmeister H, Kratochwill K et al. Cellular stress-response modulators in the acute rat model of peritoneal dialysis.Pediatr Nephrol 2010; 25: 169–172
37. Preiser JC, Wernerman J. Glutamine, a life-saving nutrient, but why?Crit Care Med 2003; 31: 2555–2556
38. Breborowicz A, Witowski J, Polubinska A et al. L-2-oxothiazolidine-4-carboxylic acid reduces in vitro cytotoxicity of glucose degradationproducts. Nephrol Dial Transplant 2004; 19: 3005–3011
39. Curi R, Newsholme P, Procopio J et al. Glutamine, gene expression,and cell function. Front Biosci 2007; 12: 344–357
Received for publication: 22.11.10; Accepted in revised form: 4.7.11
946 K. Kratochwill et al.
by guest on April 11, 2016
http://ndt.oxfordjournals.org/D
ownloaded from
Related Documents











