Genes, Brain and Behavior (2007) 6: 364–374 # 2006 The Authors Journal Compilation # 2006 Blackwell Publishing Ltd Effect of the catechol-O-methyltransferase val 158 met genotype on children’s early phases of facial stimuli processing M. Battaglia* ,†,‡,§ , A. Zanoni † , R. Giorda § , U. Pozzoli § , A. Citterio † , S. Beri § , A. Ogliari † , M. Nobile § , C. Marino ‡,§ and M. Molteni § † Department of Psychology, ‘Vita-Salute’ San Raffaele University at the Istituto Scientifico San Raffaele, Milan, Italy, ‡ Department of Psychiatry, Laval University, Que ´ bec, Canada, and § Department of Child Psychiatry, Istituto Scientifico Eugenio Medea, Bosisio Parini, Italy *Corresponding author: M. Battaglia, MD, UhSR-DNS, 20 via Stamira d’Ancona, 20127 Milan, Italy. E-mail: [email protected] The ability to process and identify human faces ma- tures early in life, is universal and is mediated by a distri- buted neural system. The temporal dynamics of this cognitive–emotional task can be studied by cerebral visual event-related potentials (ERPs) that are stable from midchildhood onwards. We hypothesized that part of individual variability in the parameters of the N170, a waveform that specifically marks the early, pre- categorical phases of human face processing, could be associated with genetic variation at the functional poly- morphism of the catechol-O-methyltransferase (val 158 met) gene, which influences information processing, cogni- tive control tasks and patterns of brain activation during passive processing of human facial stimuli. Forty-nine third and fourth graders underwent a task of implicit processing of other children’s facial expressions of emo- tions while ERPs were recorded. The N170 parameters (latency and amplitude) were insensitive to the type of expression, stimulus repetition, gender or school grade. Although limited by the absence of met- homozygotes among boys, data showed shorter N170 latency associ- ated with the presence of 1-2 met158 alleles, and family- based association tests (as implemented in the PBAT version 2.6 software package) confirmed the association. These data were independent of the serotonin trans- porter promoter polymorphism and the N400 waveform investigated in the same group of children in a previous study. Some electrophysiological features of face pro- cessing may be stable from midchildhood onwards. Different waveforms generated by face processing may have at least partially independent genetic architectures and yield different implications toward the understand- ing of individual differences in cognition and emotions. Keywords: Catechol- O-methyltransferase, children, development, ERP, face expressions of emotion, genetics Received 20 March 2006, revised 6 June 2006 and accepted for publication 26 June 2006 The act of processing a human face takes place following a specific temporal and topographic succession of events in the brain (Adolphs 2002): while imaging studies can identify precisely the brain areas involved in human face processing, electrophysiological studies are powerful in addressing the temporal dynamics of this complex cognitive–emotional task (Haxby et al. 2002). Cerebral visual event-related potentials (ERPs) techniques have identified some face-specific early potentials occurring within 200 milliseconds (range 120–200 milliseconds) from stimulus onset (Eimer 2000a; Eimer & Holmes 2002; Puce et al. 1999). These include the N170 waveform, a conspicuous negative peak that appears to be pre-eminently evoked by human faces but not, or to a much lesser extent, by other types of visual stimuli (Bentin et al. 1996; Itier & Taylor, 2004; Sagiv & Bentin 2001). Although the precise source of the N170 is not yet determined across ERP studies (Puce et al. 1998; Taylor et al. 1999) and through developmental stages (Taylor et al. 2004), it can be evoked in children from the age of 7 to 8 years (Taylor et al. 1999) and responds similarly to experimental manipulations in adults and children starting from midchildhood (Taylor et al. 2004). While in paradigms of facial stimuli presentation, the N170 amplitude and latency are typically unaffected by the type of expression (Eimer & Holmes 2002), stimulus’ repetition (Schweinberger et al. 2002) or a face’s degree of familiarity (Bentin & Deouell 2000; Eimer 2000a), inversion of facial stimuli increases significantly the N170 latency (Eimer 2000a; Rossion et al. 2000; Taylor et al. 2004). The bulk of these data led several, but not all, authors (see e.g. Eimer 2000a; Schweinberger & Burton 2003) to consider the N170 as a waveform reflecting the perceptual encoding analysis of the human face, and likely to mark the early, pre- categorical, rather than the subsequent recognition stage, processes. Consistent with this view, the N170 parameters also reflect the degree of cognitive control requested by the early perceptual encoding of the human face: experi- mental manipulations obtained by altering, and/or interfer- ing with, face stimuli encoding via demanding visual tasks 364 doi: 10.1111/j.1601-183X.2006.00265.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Genes, Brain and Behavior (2007) 6: 364–374 # 2006 The AuthorsJournal Compilation # 2006 Blackwell Publishing Ltd
Effect of the catechol-O-methyltransferase val158metgenotype on children’s early phases of facialstimuli processing
M. Battaglia*,†,‡,§, A. Zanoni†, R. Giorda§,
U. Pozzoli§, A. Citterio†, S. Beri§, A. Ogliari†,
M. Nobile§, C. Marino‡,§ and M. Molteni§
†Department of Psychology, ‘Vita-Salute’ San Raffaele University
at the Istituto Scientifico San Raffaele, Milan, Italy, ‡Department
of Psychiatry, Laval University, Quebec, Canada, and§Department of Child Psychiatry, Istituto Scientifico Eugenio
Medea, Bosisio Parini, Italy
*Corresponding author: M. Battaglia, MD, UhSR-DNS, 20 via
Stamira d’Ancona, 20127Milan, Italy. E-mail: [email protected]
The ability to process and identify human faces ma-
tures early in life, is universal and is mediated by a distri-
buted neural system. The temporal dynamics of this
cognitive–emotional task can be studied by cerebral
visual event-related potentials (ERPs) that are stable
from midchildhood onwards. We hypothesized that
part of individual variability in the parameters of the
N170, a waveform that specifically marks the early, pre-
categorical phases of human face processing, could be
associated with genetic variation at the functional poly-
morphism of the catechol-O-methyltransferase (val158met)
gene, which influences information processing, cogni-
tive control tasks and patterns of brain activation during
passive processing of human facial stimuli. Forty-nine
third and fourth graders underwent a task of implicit
processing of other children’s facial expressions of emo-
tions while ERPs were recorded. The N170 parameters
(latency and amplitude) were insensitive to the type of
expression, stimulus repetition, gender or school grade.
Although limited by the absence of met- homozygotes
among boys, data showed shorter N170 latency associ-
ated with the presence of 1-2 met158 alleles, and family-
based association tests (as implemented in the PBAT
version 2.6 software package) confirmed the association.
These data were independent of the serotonin trans-
porter promoter polymorphism and the N400 waveform
investigated in the same group of children in a previous
study. Some electrophysiological features of face pro-
cessing may be stable from midchildhood onwards.
Different waveforms generated by face processing may
have at least partially independent genetic architectures
and yield different implications toward the understand-
ing of individual differences in cognition and emotions.
Keywords: Catechol-O -methyltransferase, chi ldren,
development, ERP, face expressions of emotion, genetics
Received 20 March 2006, revised 6 June 2006 and accepted
for publication 26 June 2006
The act of processing a human face takes place following
a specific temporal and topographic succession of events in
the brain (Adolphs 2002): while imaging studies can identify
precisely the brain areas involved in human face processing,
electrophysiological studies are powerful in addressing the
temporal dynamics of this complex cognitive–emotional task
(Haxby et al. 2002).
Cerebral visual event-related potentials (ERPs) techniques
have identified some face-specific early potentials occurring
within 200 milliseconds (range 120–200 milliseconds) from
stimulus onset (Eimer 2000a; Eimer & Holmes 2002; Puce
et al. 1999). These include the N170waveform, a conspicuous
negative peak that appears to be pre-eminently evoked by
human faces but not, or to a much lesser extent, by other
types of visual stimuli (Bentin et al. 1996; Itier & Taylor, 2004;
Sagiv & Bentin 2001). Although the precise source of the
N170 is not yet determined across ERP studies (Puce et al.
1998; Taylor et al. 1999) and through developmental stages
(Taylor et al. 2004), it can be evoked in children from the age
of 7 to 8 years (Taylor et al. 1999) and responds similarly to
experimental manipulations in adults and children starting
from midchildhood (Taylor et al. 2004). While in paradigms of
facial stimuli presentation, the N170 amplitude and latency are
typically unaffected by the type of expression (Eimer & Holmes
2002), stimulus’ repetition (Schweinberger et al. 2002) or a face’s
degree of familiarity (Bentin & Deouell 2000; Eimer 2000a),
inversion of facial stimuli increases significantly the N170 latency
(Eimer 2000a; Rossion et al. 2000; Taylor et al. 2004).
The bulk of these data led several, but not all, authors (see
e.g. Eimer 2000a; Schweinberger & Burton 2003) to consider
the N170 as a waveform reflecting the perceptual encoding
analysis of the human face, and likely to mark the early, pre-
categorical, rather than the subsequent recognition stage,
processes. Consistent with this view, the N170 parameters
also reflect the degree of cognitive control requested by the
early perceptual encoding of the human face: experi-
mental manipulations obtained by altering, and/or interfer-
ing with, face stimuli encoding via demanding visual tasks
364 doi: 10.1111/j.1601-183X.2006.00265.x

produced significantly delayed N170 latency in most
(Eimer 1998, 2000a; Holmes et al. 2003; Jemel et al. 1999),
but not all (Cauquil et al. 2000), studies of face perception
in adults.
At present, although metanalyses of twin studies found
substantial heritability for some ERP waveforms evoked by
cognitive tasks (van Beijsterveldt & van Baal 2002), little is
known about the genes that influence individual variation in
face-evoked ERPs in development, and so far no gene has
been associated with variation in the N170 parameters.
Inasmuch as experimental evidence shows that cognitive
control is a component of the individual variability of the N170
parameters (Eimer 1998, 2000a; Holmes et al. 2003; Jemel
et al. 1999), polymorphic genes that have been shown to
influence different processing tasks constitute reasonable
candidates for the molecular genetic investigation of the
N170.
We believed that the catechol-O-methyltransferase
(COMT) gene could be one such candidate because of its
involvement in several processing tasks, including trials that
implicate cognitive control (Goldberg & Weinberger 2004),
and tasks of passive processing of human facial stimuli
(Smolka et al. 2005). The COMT gene contains a G to A
missense variant (Lachman et al. 1996) that translates into a
substitution of methionine for valine at codon 158 (val158met),
with the enzyme containing met158 having one-third to
one-fourth of the activity of the val158 enzyme (Lotta et al.,
1995; Spielman & Weinshilboum 1981) in degrading dopa-
mine, epinephrine and norepinephrine. Consistently, there
are several reports of increased dopaminergic cortical signal-
ing, enhanced signal/noise ratio and optimized cortical activity
conferred by the met allele during different processing tasks
(Egan et al. 2001; Hariri & Weinberger 2003; Mattay et al.
1996, 2003; Smolka et al. 2005), including an ERP study of
P300 generated by an ‘oddball’ task in healthy controls and
in patients with schizophrenia (Gallinat et al. 2003). Most
recently, two brain imaging studies of healthy adults showed
an influence of COMT val158met genotype on task-related
brain activation. One study (Blasi et al. 2005) showed that the
met allele confers better cognitive performance and optimal
cingulate activation during visual cognitive control tasks,
while another study (Smolka et al. 2005) showed that the
met allele is associated with higher prefrontal, limbic and
cortical sensory (fusiform gyrus and inferior parietal lobule)
activation during the act of passive processing of emotional
stimuli.
We hypothesized that in a paradigm of face expression
presentation to third and fourth graders, neither the amplitude
nor the latency of the N170 would be sensitive to the type of
expression, but that variation of the N170 latency, as an index
of promptness to activate the early stages of face processing,
could be partially influenced by genetic variation at the
COMT val158met genotype, which has been found associated
with human variation in different aspects of information
processing.
Materials and methods
Subjects
This study is part of an ongoing project on the developmental
behavioral genetics of social interactions based on children
and their families randomly selected from the general pop-
ulation of the Lombard provinces of Milan and Lecco,
Northern Italy. The procedures were accepted by the ethical
committee of the participating institutions, and parents
signed a declaration of informed consent. The methods for
subjects’ selection and ERP recording have been described in
detail elsewhere (Battaglia et al. 2004, 2005). Briefly, 1 year
(time 0) before the ERP recording analyzed in this study, an
inception cohort of 149 schoolchildren was characterized by
different psychological variables and the ability to discriminate
other children’s facial expressions of emotions (Battaglia et al.
2004). After random selection and invitation to participate
sent to about 45% of subjects from the original sample, 49
white children of Italian ancestry with normal or corrected-to-
normal visual acuity agreed to take part in the ERP-genetic
study after parental written agreement. Post hoc compar-
isons showed no differences in several major demographic
and psychometric characteristics between 49 participating
and. 100 nonparticipating children (Battaglia et al. 2005).
Because a major focus of the project is set on the relation-
ships occurring between polymorphic genes, neurofunctional
responses to social–emotional stimuli and temperamental
traits, an extensive collection of behavioral data from teach-
ers, parents, direct observation of the children by psycholo-
gists and direct blind independent interviews to children and
their mothers through clinician-administered structured inter-
views was made at time 0 and on the day of the ERP recoding
(Battaglia et al. 2004, 2005).
DNA extraction and genotyping
The genomic DNA of all participating children and their
biological parents was extracted from mouthwash samples
collected in 4% sucrose by means of a reagent for isolation of
genomic DNA (DNAzol Genomic DNA Isolation Reagent,
Molecular Research Center Inc., Cincinnati, OH, USA).
We determined the subject’s COMT V158M genotypes
using the 50-exonuclease Taqman assay (Chenet al. 2004). Sub-
jects with known genotype, previously typed by sequencing,
were used as positive controls. The Taqman primers, probes
and reagents were purchased from Applied Biosystems (Monza,
Italy). The assays were performed and analyzed on a 9700HT
Sequence Detection System (Applied Biosystems).
Protocol
Because third- and fourth-grade schoolchildren spendmost of
their time among other children, not adults, we chose to use
standardized faces of other children of similar age (two
models, a boy and a girl, aged 8–9 years), instead of adults.
Stimuli consisted of six black-and-white pictures standardized
Genes, Brain and Behavior (2007) 6: 364–374 365
COMT genotype and face processing in children
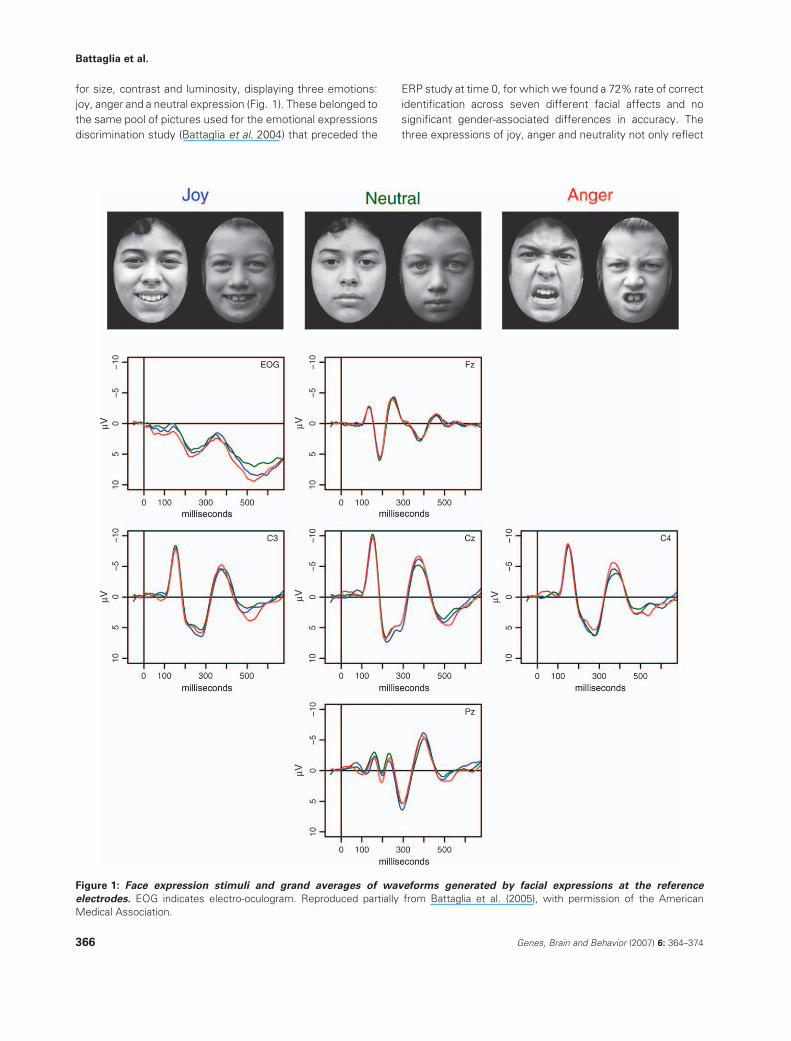
for size, contrast and luminosity, displaying three emotions:
joy, anger and a neutral expression (Fig. 1). These belonged to
the same pool of pictures used for the emotional expressions
discrimination study (Battaglia et al. 2004) that preceded the
ERP study at time 0, for which we found a 72% rate of correct
identification across seven different facial affects and no
significant gender-associated differences in accuracy. The
three expressions of joy, anger and neutrality not only reflect
Figure 1: Face expression stimuli and grand averages of waveforms generated by facial expressions at the reference
electrodes. EOG indicates electro-oculogram. Reproduced partially from Battaglia et al. (2005), with permission of the American
Medical Association.
366 Genes, Brain and Behavior (2007) 6: 364–374
Battaglia et al.

the conditions of acceptance, prosocialization, rejection hos-
tility and ambiguity, that characterize several possible social
interactions in childhood but also are processed by at least
partially independent brain networks (Adolphs 2002; Camras &
Allison 1985; Canli et al. 2002).
Children were told that this was a video game in which they
would get a gift ‘if they carefully followed the instructions and
performed well’.
Trials
On each trial, the children were first presented with a child’s
face (total time on screen, 1300 milliseconds), which they
were instructed to watch carefully until a blue circle appeared
superimposed around the center of the picture. As soon as
a blue circle appeared (700 milliseconds after the appearance
of the stimulus), they had to click a mouse. Thus, the ERPs
relevant to this study were all generated before the motor
task, which was merely set up to stimulate children’s
cognitive control and participation.
The monitor screen remained dark between trials for
periods that varied randomly from 1200 to 1600 milliseconds.
The stimuli were presented to all the children in a sequence
that alternated male and female pictures and avoided close
repetition of the same expression. To simulate a real video
game and sustain children’s participation, non-face screen
pictures with increasing scoring values (as reinforcements)
appeared about every six face pictures (stimuli). The possible
ERP signal evoked by such non-face stimuli was ignored and
therefore excluded from the baseline used to estimate the
waveforms. Each face stimulus was presented 20 times to
ensure sufficient ERP acquisition (total, 120 presentations
in a complete session). Every child was exposed to a pre-
experiment trial of six pictures not belonging to the same set
used for the experiment to make sure she or he understood
the procedure well. Each child received a gift of a value
equivalent to e30.
ERP acquisition and analysis
As in other studies of ERP and facial expressions of children
and adults (Batty & Taylor 2002; Eimer 2000a; Kestenbaum &
Nelson 1992; Pollak et al. 2001), electroencephalographic
activity was recorded at sites Fz, C3, Cz, C4 and Pz of the
10–20 systemwith the use of silver–silver chloride electrodes
referred to linked mastoids with an amplifier (Neuroscan
SynAmp, Neuroscan Labs, Sterling, VA, USA), with head
preamplified gain 150 and acquisition software (SCAN,
version 4.2, Neuroscan Labs). The ground electrode was
attached to the forehead. Electro-oculographic activity was
recorded from electrodes placed above and below the right
eye. Electrode impedance was maintained below 5 kV. The
electroencephalogram and electro-oculogram were amplified
(gain 500), analogically band-pass filtered (1–30 Hz), digitized
and acquired at a1000-Hz sampling rate. Electroencephalo-
gram and electro-oculogram epochs between –50 and
1300 milliseconds from the stimulus onset were obtained
by means of different trigger codes for each image, allowing
for later off-line artifact rejection, sorting and digital averaging
with the Neuroscan EDIT software (Neuroscan Labs). All
epochs from all electrodes were rejected if affected by
artifacts (greater than þ65 mV or less than –65 mV between
–50 and 700 milliseconds). The ERP averages were con-
structed from artifact-free epochs for each trigger code and
for each electrode. Amplitude was measured in relation to the
baseline mean voltage level preceding the onset of a wave-
form, and latency was defined as the time that occurred
between the appearance of the stimulus (set at time 0) and
the waveform’s peak.
Although there were 49 participants, results are shown for
number of participants that vary between 37 and 45 because
of removal of subjects who had artifacts such as eyeblinks in
recordings.
Genetic analyses
Between-individuals associations of phenotypes (N170 ampli-
tude and latency) with the COMT val158met genotype were
tested by analysis of variance (ANOVA).
Because between-individuals association tests are suscep-
tible to confounding effects due to population stratification,
those variables that yielded significant results in between-
individuals associations were submitted to within-family
analyses. The transmission disequilibrium test (TDT) is a test
of linkage in the presence of association that was originally
developed for dichotomous traits (Spielman et al. 1993). The
family-based association test (FBAT) was successively intro-
duced as an extension of the original TDT to deal with
quantitative phenotypes (Laird et al. 2000); the FBAT statistic
computes the distribution of the offspring’s genotype condi-
tioning on parental genotypes or family genotype configura-
tion, assuming that H0 is true (Laird et al. 2000).
In this study, we adopted the FBAT methodology to cal-
culate the FBAT statistics as implemented in the phenotype-
based extension of the family-based association test (PBAT),
which allows for the use of covariates; the PBAT software
package version 2.6 is available at http://www.biostat.
harvard.edu/~clange/default.htm (Lange et al. 2004; Steen &
Lange 2005).
The N170 waveform is likely to be a multifactorial condi-
tion, and the transmission pattern of inheritance of N170
parameters is likely to be complex. No information is avail-
able to choose a priori the proper model of transmission of
N170 parameters; therefore, we performed the analyses
with all the four possible models (i.e. additive, dominant, re-
cessive and codominant) of transmission. As a consequence,
the nominal alpha level of the FBAT statistics was adjusted
for the number of FBAT statistics. The offset was set in order
to maximize the power of the FBAT statistic (Lange et al.
2004).
Genes, Brain and Behavior (2007) 6: 364–374 367
COMT genotype and face processing in children

Results
ERP analyses
Analyses of general effects
Analyses of the waveforms generated by the facial expres-
sions (Fig. 1, Table 1) showed an enhanced early negativity
occurring after stimulus presentation at a mean of 136 milli-
seconds for the Fz, 153 milliseconds for the Cz, 152 milli-
seconds for the C4, 154 milliseconds for C3, and
159 milliseconds for the Pz electrode. These latency values
are compatible with those reported by studies of N170
elicitation by facial stimuli in midchildhood (Taylor et al.
2004) and adulthood (Eimer & Holmes 2002), and thus these
waveforms were putatively identified as N170. By visual
inspection (Fig. 1), the N170 had maximum amplitudes and
clearest waveforms at the central (Cz) and centrolateral (C3,
C4) electrodes, similarl to other studies of N170 elicitation by
face perception (Eimer 2000b), while on the posterior
electrode (Pz), the N170 assumed the bifid shape typically
recorded by posterior electrodes in children of the 8–11 age
span, the so-called N170a and N170b (Taylor et al. 2004)
waveforms. According to ANOVA, the mean amplitudes of the
three expressions (joy, neutral and anger) were significantly
different across the five electrodes (ANOVA-repeated meas-
ures: F4,96 ¼ 47.14, P ¼ 0.00001) and significantly larger at
Cz, C3 and C4 than the amplitudes at Fz and Pz (Post hoc
Newman–Keuls P range ¼ 0.00010–0.00014); moreover,
the mean amplitude at Cz was larger than mean amplitude
at C3 and C4 (Post hoc Newman–Keuls: Cz vs. C3 P ¼ 0.03;
Cz vs. C4 P ¼ 0.06).
Similarly, ANOVA of the mean N170 latencies of the three
expressions (joy, neutral and anger) showed significant
differences across the five electrodes (ANOVA-repeated
measures: F4,96 ¼ 19.34, P ¼ 0.00001), with latencies at
central electrodes not significantly different from each other
(Post hoc Newman–Keuls P ¼ nonsignificant, for all Cz, C3
and C4 cross-comparisons) but significantly different from
both Fz and Pz latencies (Post hoc Newman–Keuls compar-
isons of Cz, C3 and C4 vs. Fz and Pz p range ¼ 0.00010–
0.00033).
In the light of these data and of the fact that Cz was the
electrode with the highest number of valid observations
(n ¼ 45), we focused the rest of the analyses on N170
parameters (latency and amplitude) as obtained at Cz.
Consistently, a visual inspection analysis of the topography
of the N170 waveform in our children showed a clear
prevalence of negativity at a central location compatible
with the choice of the Cz, and with the topographic
distribution found by Taylor et al. (2004, fig. 8) in children
aged 8–9 years in the context of an implicit emotional task.
To ascertain the degree of variability of measures across
trials (habituation effects), we performed ANOVA with ‘time’
(on two levels: first block, encompassing the first 50% of
repetitions; and second block, with the remaining repeti-
tions) as a repeated-measures factor, separately for N170 Table
1:N170amplitudesandlatenciesevokedbyfacialexpressionsatthereferenceelectrodesin
thesample
Joy
Neutral
Anger
Meanofthreeexpressions
Amplitude�
SD
(mV)
Latency
�SD
(milliseconds)
Amplitude�
SD
(mV)
Latency
�SD
(milliseconds)
Amplitude�
SD
(mV)
Latency�
SD
(milliseco
nds)
Amplitude�
SD
(mV)
Latency
�SD
(milliseconds)
Fz
�4.58�
2.25
(n¼
37)
135.92�
8.05
(n¼
37)
�4.35�
2.46
(n¼
38)
133.77�
11.96
(n¼
38)
�4.23�
2.37
(n¼
38)
135.92�
22.59
(n¼
38)
�4.33�
1.65
(n¼
37)
135.60�
8.56
(n¼
37)
Cz
�10.92�
4.21
(n¼
45)
154.16�
8.64
(n¼
45)
�11.81�
5.16
(n¼
45)
152.76�
10.53
(n¼
45)
�11.13�
4.10
(n¼
45)
151.90�
9.04
(n¼
45)
�11.29�
3.46
(n¼
45)
152.94�
7.85
(n¼
45)
Pz
�6.48�
4.83
(n¼
38)
161.65�
28.13
(n¼
38)
�8.07�
4.02
(n¼
38)
158.33�
32.84
(n¼
38)
�7.29�
4.83
(n¼
38)
155.62�
31.84
(n¼
38)
�7.28�
3.07
(n¼
38)
158.88�
28.98
(n¼
38)
C3
�9.80�
4.02
(n¼
40)
152.16�
8.32
(n¼
40)
�10.28�
4.40
(n¼
41)
153.43�
12.65
(n¼
40)
�9.67�
4.04
(n¼
41)
154.89�
10.50
(n¼
41)
�9.89�
3.16
(n¼
40)
153.55�
7.40
(n¼
39)
C4
�9.88�
4.37
(n¼
39)
153.62�
10.23
(n¼
39)
�10.34�
4.13
(n¼
40)
152.41�
9.91
(n¼
40)
�10.62�
3.96
(n¼
40)
149.61�
8.98
(n¼
40)
�10.23�
3.10
(n¼
39)
151.93�
7.65
(n¼
39)
368 Genes, Brain and Behavior (2007) 6: 364–374
Battaglia et al.

amplitude and latency for each expression recorded at Cz. We
found no evidence of significant habituation effects on either
amplitude or latency (with the possible exception of a trend
toward significantly decreased amplitude in the second vs.
first block of stimulations with the neutral expression ANOVA-
repeated measures: F1,41 ¼ 3.85, P ¼ 0.060), confirming
previous findings that the face-evoked N170 waveform is
unaffected by stimulus repetition.
To ascertain the possible effect of the type of expression
on the N170 parameters at Cz, we performed two separate
ANOVA-repeated measures tests on latency and amplitude,
with the three different expressions (joy, neutral and anger)
as a repeated measures factor. Results (latency: F2,88 ¼ 1.43,
P ¼ 0.24; amplitude: F2,88 ¼ 0.78, P ¼ 0.46) confirmed
that the N170 waveform is insensitive to the type of face
expression. Consequently, we focused all the following
analyses on the N170 latency and amplitude values calcu-
lated as a mean of the three expressions. The distributions
of these two parameters did not deviate significantly
from normality (N170 amplitude mean of three expressions
skewness ¼ �0.007, kurtosis ¼ �0.640, Lilliefors test
P ¼ 0.20; N170 latency mean of three expressions: skew-
ness ¼ 0.650, kurtosis ¼ 1.330; Lilliefors test P ¼ 0.20).
Moreover, two separate two-way ANOVA with age (8–
10 years) and sex as independent variables conducted on
Cz N170 latency and amplitude (mean of three expressions)
showed no significant influence of either of these demo-
graphic variables (latency: age F2,39 ¼ 0.52, P ¼ 0.60, sex
F1,39 ¼ 3.13, P ¼ 0.10, sex by age interaction P ¼ 0.62;
amplitude: sex F1,39 ¼ 0.56, P ¼ 0.46, age F2,39 ¼ 0.43, P
¼ 0.65, sex by age interaction P ¼ 0.73). However, as
commonly reported in the literature (e.g. Taylor et al. 2004),
a Student’s t-test showed that girls had slightly shorter N170
latency than boys (150.19 � 6.33 vs. 155.36 � 8.37, t ¼�2.30, P ¼ 0.03).
Genetic analyses
Genotyping
Forty-nine index children and their biological parents were
genotyped for the COMT val158met polymorphism. Both
parents were available for 42 children; only the mother was
available for two children; neither parent was available for five
children. The allele frequencies in children were 55 (56%) for
the val allele and 43 (44%) for the met allele, and the
genotypic frequencies were 14 (29%) val/val, 27 (55%) val/
met and 8 (16%)met/met, consistent with those expected in
European populations (Palmatier et al. 1999) and in Hardy–
Weinberg equilibrium (P ¼ 0.60). There were no differences
in the genotypes’ distribution of children in the study owing to
age, education (based on school grade 3 or 4) or socioeco-
nomic status (divided into lower, middle and upper and
calculated on the basis of the Hollingshead scale, 1975).
There was, however, a significant difference in the genotype-
by-sex distribution (w2 ¼ 17.360, df ¼ 2, P ¼ 0.001), owing
to a relative excess of boys with the val/met, and absence of
boys with the met/met genotype (Table 2). To better clarify
the nature of genotype distribution among boys, and to
conduct further analyses with the FBAT methodology, we
analyzed allelic and genotypic frequencies in parents. Hardy–
Weinberg equilibrium was also observed among parents
(P ¼ 0.37); frequencies for the val allele in parents were
52% among fathers and 64% among mothers (total fre-
quency among parents: 58%) and for the met allele were
48% among fathers and 36% among mothers (total fre-
quency among parents: 42%). The distribution of COMT
Table 2: Demographic and N170 electrophysiological features across genotypes in children
Catechol-O-methyltransferase val158met genotype
val/val (n ¼ 14) val/met (n ¼ 27) met/met (n ¼ 8)
Age, years (mean � SD) 8.79 � 0.80 8.78 � 0.70 9.12 � 0.64
Sex, girls (%)* 5 (64.28) 6 (22.22) 8 (100)
Education
Grade 3 8 17 3
Grade 4 6 10 5
Socioeconomic status (%)
Lower 2 (14.29) 3 (11.11) 1 (12.5)
Middle 7 (50) 13 (48.15) 4 (50)
Upper 5 (35.71) 11 (40.74) 3 (37.5)
N170 amplitude (mV) mean � SD
of three expressions
�11.44 � 2.51 (n ¼ 11) �11.37 � 3.79 (n ¼ 26) �10.79 � 3.81 (n ¼ 8)
N170 latency (ms)† mean � SD
of three expressions
154.98 � 8.09 (n ¼ 11) 153.96 � 7.43 (n ¼ 26) 146.83 � 6.60 (n ¼ 8)
All differences nonsignificant except *w2 ¼ 17.35, df ¼ 2, P ¼ 0.001, and †ANOVA F2,42 ¼ 3.33, P ¼ 0.044.
Genes, Brain and Behavior (2007) 6: 364–374 369
COMT genotype and face processing in children

val158met alleles in parents (Table 3) was entirely consis-
tent with that of children, indicating that the skewed
distribution of alleles among boys was simply due to
chance, as it can occur in samples of small size like this
one. Call rate was>99% and error rate<1% in the population
tested.
Between-subjects association tests
Two separate ANOVA conducted on N170 latency and ampli-
tude at Cz (mean of three expressions), with the COMT
val158met polymorphism as the independent variable on three
levels (0, 1 and 2 met alleles), showed no influence on
amplitude (F2,42 ¼ 1.00, P ¼ 0.91), but showed a significant
effect on latency, in the direction of a reduced time of onset of
the N170 waveform predicted by the number of met alleles
(F2,42 ¼ 3.33, P ¼ 0.044; regression beta ¼ �.32, adjusted
R2 ¼ 0.08; Newman–Keuls post hoc: met/met vs. val/val
P ¼ 0.03, met/met vs. val/met P ¼ 0.025, val/met vs. val/
val P ¼ 0.74), consistent with our hypothesis (Table 2,
Fig. 2). There was one outlier (a boy) in the val/val group with
a latency of 177.40 milliseconds (Fig. 2): when ANOVA on Cz
N170 latency was repeated after removal of this subject,
differences remained significant (F2,41 ¼ 3.58, P ¼ 0.037,
Newman–Keuls post hoc: met/met vs. val/val P ¼ 0.039,
met/met vs. val/met P ¼ 0.036, val/met vs. val/val P ¼ 0.66).
When ANOVA was repeated following the dichotomic
approach of Meyer-Lindenberg et al. (2005), i.e. contrasting
met homozygotes vs. val carriers (which was possible in
female subjects only, due to the absence of met homozy-
gotes among boys), the differences in N170 latency by
genotype were confirmed (met homozygotes 146.83 � 6.60
milliseconds vs. val carriers 152.26 � 5.41 milliseconds;
F1,23 ¼ 4.230, P ¼ 0.054) again.
Within-family association test
Following the between-subjects associations tests, the N170
latency was further tested for family-based association with
the COMT val158met polymorphism. Univariate analyses
were run with the N170 latency as a quantitative phenotype
and sex as a covariate (Lange et al. 2004), given the difference
in genotype-by-sex distribution among children in the study.
Among the four alternative models of transmission (domi-
nant, recessive, codominant and additive), only the codomi-
nant model yielded a significant result for the COMT
val158met polymorphism (FBAT statistic P ¼ 0.0034) with
25 informative families, and a nominal alpha level adjusted
to 0.013.
Relationships of the N170 to other salient
waveforms, and of the COMT genotype
to behavioral variables
The distribution of the COMT val158met genotypes was
independent of the distribution of the serotonin transporter
(5HTT) promoter polymorphism (5-HTTLPR, w2 ¼ 1.453,
df ¼ 4, P ¼ 0.834), which we (Battaglia et al. 2005) had
previously investigated in this group of children in relation to
social shyness and another waveform, namely the N400,
associated with face processing.
The correlations of the N170 and N400 amplitudes and
latencies across the three expressions were modest to
moderate and varied between .07 and .29 for latency, and
between .19 and .35 for amplitude (all correlations nonsignifi-
cant for this sample size). Likewise, by ANOVAs, we found no
significant association of the 5-HTTLPR genotype with N170
latency or amplitude (P ¼ 0.18 and P ¼ 0.17, respectively) or
COMT val158met genotype with N400 latency or amplitude
(P ¼ 0.25 and P ¼ 0.91, respectively).
To further clarify possible relationships of the COMT
val158met genotypes to other variables that we had collected
in our children, we ran separate ANOVAs with the number of
COMTmet alleles as the independent variable on three levels
(0, 1 and 2 met alleles) and found no significant effect
pertaining to a comprehensive index of observed degree of
shyness (Battaglia et al. 2005), the number of spontaneous
Figure 2: Scatterplot of subjects in the experiment ordered
by catechol-O-methyltransferase (COMT) val158met geno-
type. The graph shows the latency of the N170waveform evoked
by face expressions (mean of three expressions) at the Cz
electrode for each subject.
Table 3: Results of catechol-O-methyltransferase val158met
parental genotyping
Subjects Typing n
Total (n ¼ 86) val 32
val/met 36
met 18
Fathers (n ¼ 42) val 13
val/met 18
met 11
Mothers (n ¼ 44) val 19
val/met 18
met 7
370 Genes, Brain and Behavior (2007) 6: 364–374
Battaglia et al.

comments made on the ERP experiment day, all the Children
Behavior CheckList (CBCL 4–18, Achenbach, 1991) scales [(1)
withdrawn, (2) somatic complaints, (3) anxious/depressed, (4)
social problems, (5) thought problems, (6) attention problems,
(7) delinquent behavior, (8) aggressive behavior, and the
broadband internalizing and externalizing scores encompass-
ing syndromes 1, 2, 3, and 7, 8, respectively), or the number
of misclassifications made in a face expression discrimination
task completed by children 1 year before the ERP study.
Discussion
Our results indicate that variation at the COMT val158met
genotype influences the time of onset of the N170 waveform,
an electrophysiological event mapping the early phases of the
cognitive–emotional act of human face processing in the
brain. Specifically, the met allele is associated with a signifi-
cantly reduced latency of the N170, the earliest detectable
electrophysiological waveform in our experiment, and awave-
form that has been repeatedly shown to be specifically
evoked by human face in children and adults.
The finding of better promptness to face processing
associated with the met allele is consistent with the notion
that the enzyme containing the met allele has one-third to
one-fourth of the activity of the enzyme containing the val
allele (Chen et al. 2004; Lotta et al. 1995; Spielman &
Weinshilboum 1981) in degrading dopamine. The enzyme
containing the met allele is thus likely to confer increased
cortical dopamine signaling (Weinberger et al. 2001), which in
turn can explain better cortical signal-to-noise ratio (Gallinat
et al. 2003; Mattay et al. 1996), more effective interaction
between prefrontal activity and midbrain dopaminergic syn-
thesis (Meyer-Lindenberg et al. 2005), better performances in
tests of executive cognition (Goldberg & Weinberger 2004)
and shorter reaction time in visual cognitive control tasks
(Blasi et al. 2005) reported in adults with one or two copies of
the met allele. Likewise, children homozygous for the met
allele perform better in cognitive tasks that specifically
engage the dorsolateral prefrontal cortex (Diamond et al.
2004). Due to chance, the distribution of genotypes is
significantly different between boys and girls in this sample;
however, there is robust enough evidence that gender affects
only modestly the function of this gene, and only after
puberty: functional analyses of COMT genetic variation on
dorsolateral prefrontal cortex specimens show only small
effects of sex on COMT enzyme activity, with adult women
having slightly but significantly lower activity than adult men
(Chen et al. 2004), likely due to the regulatory activity of
estrogen on COMT promoter activity (Chen et al. 2004;
Weinshilboum et al. 1999). Because all our subjects were
prepubertal, an influence of sexual hormones on COMT
enzymatic activity should be excluded, and the uneven
distribution of genotypes by gender should therefore be
considered only a relative limitation to the generalizability of
these findings in midchildhood.
Turning our attention to the electrophysiological data, they
confirm earlier findings (Taylor et al. 1999) that even in
midchildhood the face-evoked N170 waveform is a reliable
marker of the early phases of face processing. The lack of
influence of the type of emotional expression and the
absence of habituation effects on N170 amplitude or latency
in our data are consistent with experimental evidence
that this waveform is not affected by emotional valence
(Eimer & Holmes 2002; McCarthy et al. 1999) or repetition
(Schweinberger et al. 2002) of stimuli in adults and extends
the same notion to children.
In a previous report based on the same group of children,
we have shown that a pattern of decreased centroparietal
cortical activation (as indexed by the N400 waveform) in
response to peers’ expressions of anger is associated with
social shyness and the –S allele of the 5-HTTLPR (Battaglia
et al. 2005), a finding independent of the present association
of N170 latency with COMT val158met genotype.
In our effort to investigate the causal architecture of the
complex cognitive–emotional act of human face processing,
we are thus focusing on different electrophysiological
phases, each of which may be underpinned by relatively
separate biological mechanisms and independent genetic
causes. The use of N170 latency as a measure of early face
structural encoding influenced by the COMT val158met geno-
type, however, is exposed to some limitations and caveats,
which need to be taken into account.
First, several previous ERP studies have typically examined
an earlier and more occipital response, such as the P1 (see
Luck et al. 2000, for review), and we cannot rule out defini-
tively the possibility that our data may reflect in part some
more general aspects of perceptual processing, perhaps even
arising at P1; however, the N170 is clearly the earliest
waveform evoked by our paradigm (Fig. 1) at the available
electrodes. Moreover, the evidences of rapid emotional face
processing, mostly at posterior sensors, found within the
100–200-millsecond time range (e.g. Pizzagalli et al. 1999,
2002; Vuillemier 2002) were based on different paradigms,
which makes direct comparisons between studies difficult:
the strongest evidence that early ERP activity can be influ-
enced by the type of emotional facial stimulus rests on data
(Pizzagalli et al. 1999, 2002) recorded to individually assessed
liked, neutral and disliked faces in adults, while we used
a paradigm of implicit processing of other children’s faces in
prepubertal subjects. A conservative interpretation of the par-
tial inconsistencies regarding the effects of affective com-
ponents of facial stimuli on early ERP waveforms is offered
by Eimer & Holmes (2002), who suggest that structural
encoding of faces and emotional expression analysis may
be parallel processes indexed by different waveforms.
Second, the relationship between the source of the N170
and the brain areas for which there is clearer evidence for
expression of the COMT gene deserves a specific caveat.
While the precise source for the N170 is still debated,
a reanalysis of combined samples (Taylor et al. 2004) found
Genes, Brain and Behavior (2007) 6: 364–374 371
COMT genotype and face processing in children

two separate N170 sources for children aged 10–11: one
occipital and the other lateral temporal. This may sound
problematic when one considers that most studies of COMT
expression and COMT val158met genotype influence on brain
activation are focused on prefrontal and frontal cortices
(Goldberg & Weinberger 2004). However, the functional
magnetic resonance data by Smolka et al. (2005) show that
a task of passive processing of human faces elicits a pattern
of brain activation that is significantly affected by the COMT
val158met genotype at the levels of the fusiform gyrus,
inferior parietal lobule, amygdala and thalamus, in addition
to the ventrolateral prefrontal cortex (where it is most pro-
nounced) and middlefrontal gyrus. Thus, inasmuch as the role
of the COMT val158met genotype in influencing cerebral
activation in response to passive viewing of human faces
extends to a wider network of brain areas and functions
beyond the prefrontal cortex, the relationship between N170
latency and the met158 allele can be viable.
Third, although our study group was under many social,
behavioral and genetic vantage points, an unbiased sample
of a cohort of schoolchildren characterized by their ability to
discriminate facial expressions of emotions (Battaglia et al.
2004, 2005), there were distributional differences in the
COMT val158met genotype, which we addressed in the ana-
lyses but call for replication in a sample without genotype-
by-sex distribution differences, thus leaving this first study
as a pilot and tentative investigation.
Altogether, while our data need replication in a more
balanced sample and information from more electrodes, they
do suggest that the complex emotional behavioral task of
human face processing can be partitioned into different
phases, each characterized by at least partially independent
genetic architectures. While earlier phases of face processing
are associated with the earlier ’exogenous‘ potentials, the
later potentials may reflect more the nature of interaction
between the subject and the event (Battaglia et al. 2005) and
may covary with other psychophysiological indexes of affec-
tive processing (Palomba et al. 2000); thus earlier and later
phases of face processing yield different implications for
psychology and psychopathology.
From a developmental behavioral genetic perspective, the
study of the molecular genetic bases of face processing can
open new avenues to understanding individual differences in
human interpersonal abilities. Changes in patterns of brain
activation in response to facial stimuli together with modifi-
cations of genes’ expression along development may consti-
tute new instruments for the implementation of more
scientifically based treatments of mental disorders, including
psychotherapies (Etkin et al. 2005).
References
Achenbach, T.M. (1991) Manual for the Child Behavior Checklist
4-18 and Profile. Department of Psychiatry, University of
Vermont, Burlington.
Adolphs, R. (2002) Neural systems for recognizing emotion. Curr
Opin Neurobiol 12, 169–177.
Battaglia, M., Ogliari, A., Zanoni, A., Villa, F., Citterio, A., Binagli, F.,
Fossati, A. & Maffei, C. (2004) Children’s discrimination of
expressions of emotions: relationship with indices of social
anxiety and shyness. J Am Acad Child Adolesc Psychiatry 43,
358–365.
Battaglia, M., Ogliari, A., Zanoni, A., Citterio, A., Bozzoli, U.,
Giorda, R., Maffei, C. & Marino, C. (2005) Influence of the
serotonin transporter promoter gene and shyness on children’s
cerebral responses to facial expressions. Arch Gen Psychiatry
62, 85–94.
Batty, M. & Taylor, M.J. (2002) Visual categorization during
childhood: an ERP study. Psychophysiology 39, 482–490.
Bentin, S. & Deouell, L.Y. (2000) Structural encoding and identi-
fication in face processing: ERP evidence for separate mech-
anisms. Cogn Neuropsychol 17, 35–54.
Bentin, S., Allison, T., Puce, A., Perez, E. & McCarthy, G. (1996)
Electrophysiological studies of face perception in humans.
J Cogn Neurosci 8, 551–565.
Blasi, G., Mattay, V.S., Bertolino, A., Elvevag, B., Callicott, J.H.,
Das, S., Kolachana, B.S., Egan, M.F., Goldberg, T.E. &
Weinberger, D.R. (2005) Effect of catechol-O-methyltransferase
val158met genotype on attentional control. J Neurosci 25,
5038–5045.
Camras, L.A. & Allison, K. (1985) Children’s understanding of
emotional facial expressions and verbal labels. J Nonverbal
Behav 9, 84–94.
Canli, T., Sivers, H., Whitfield, S.L., Gotlib, I.H. & Gabrieli, J.D.
(2002) Amygdala response to happy faces as a function of
extraversion. Science 296, 2191.
Cauquil, A.S., Edmonds, G.E. & Taylor, M.J. (2000) Is the face--
sensitive N170 the only ERP not affected by selective atten-
tion? Neuroreport 11, 2167–2171.
Chen, J., Lipska, B.K., Halim, N., Ma, Q.D., Matsumoto, M.,
Melhem, S., Kolachana, B.S., Hyde, T.M., Herman, M.M.,
Apud, J., Egan, M.F., Kleinman, J.E. & Weinberger, D.R.
(2004) Functional analysis of genetic variation in catechol-O--
methyltransferase (COMT): effects on mRNA, protein, and
enzyme activity in postmortem human brain. Am J Hum Genet
75, 807–821.
Diamond, A., Briand, L., Fossella, J. & Gehlbach, L. (2004) Genetic
and neurochemical modulation of prefrontal cognitive functions
in children. Am J Psychiatry 161, 125–132.
Egan, M.F., Goldberg, T.E., Kolachana, B.S., Callicott, J.H.,
Mazzanti, C.M., Straub, R.E., Goldman, D. & Weinberger,
D.R. (2001) Effect of COMT Val108/158 Met genotype on
frontal lobe function and risk for schizophrenia. Proc Natl Acad
Sci USA 98, 6917–6922.
Eimer, M. (1998) Does the face-specific N170 component reflect
the activity of a specialized eye processor? Neuroreport 9,
2945–2948.
Eimer, M. (2000a) Effects of face inversion on the structural
encoding and recognition of faces: evidence from event-related
brain potentials. Brain Res Cogn Brain Res 10, 145–158.
Eimer, M. (2000b) Event-related brain potentials distinguish
processing stages involved in face perception and recognition.
Clin Neurophysiol 111, 694–705.
Eimer, M. & Holmes, A. (2002) An ERP study on the time course
of emotional face processing. Neuroreport 13, 427–431.
Etkin, A., Pittenger, C., Polan, H.J. & Kandel, E.R. (2005) Toward
a neurobiology of psychotherapy: basic science and clinical
applications. J Neuropsychiatry Clin Neurosci 17, 145–158.
372 Genes, Brain and Behavior (2007) 6: 364–374
Battaglia et al.

Gallinat, J., Bajbouj, M., Sander, T., Schlattmann, P., Xu, K., Ferro,
E.F., Goldman, D. & Winterer, G. (2003) Association of the
G1947A COMT (Val(108/158)Met) gene polymorphism with
prefrontal P300 during information processing. Biol Psychiatry
54, 40–48.
Goldberg, T.E. & Weinberger, D.R. (2004) Genes and the parsing
of cognitive processes. Trends Cogn Sci 8, 325–335.
Hariri, A.R. &Weinberger, D.R. (2003) Imaging genomics. Br Med
Bull 65, 259–270.
Haxby, J.V., Hoffman, E.A. & Gobbini, M.I. (2002) Human neural
systems for face recognition and social communication. Biol
Psychiatry 51, 59–67.
Hollingshead, A.B. (1975) Four-Factor Index of Social Status.
Department of Sociology, Yale University, New Haven.
Holmes, A., Vuilleumier, P. & Eimer, M. (2003) The processing
of emotional facial expression is gated by spatial attention:
evidence from event-related brain potentials. Brain Res Cogn
Brain Res 16, 174–184.
Itier, R.J. & Taylor, M.J. (2004) N170 or N1? Spatiotemporal
differences between object and face processing using ERPs.
Cereb Cortex 14, 132–142.
Jemel, B., George, N., Chaby, L., Fiori, N. & Renault, B. (1999)
Differential processing of part-to-whole and part-to-part face
priming: an ERP study. Neuroreport 10, 1069–1075.
Kestenbaum, R. & Nelson, C.A. (1992) Neural and behavioral
correlates of emotion recognition in children and adults. J Exp
Child Psychol 54, 1–18.
Lachman, H.M., Papolos, D.F., Saito, T., Yu, Y.M., Szumlanski,
C.L. & Weinshilboum, R.M. (1996) Human catechol-O-methyl-
transferase pharmacogenetics: description of a functional poly-
morphism and its potential application to neuropsychiatric
disorders. Pharmacogenetics 6, 243–250.
Laird, N.M., Horvath, S. & Xu, X. (2000) Implementing a unified
approach to family-based tests of association. Genet Epidemiol
19 (Suppl 1), S36–S42.
Lange, C., DeMeo, D., Silverman, E., Weiss, S. & Laird, N.M.
(2004) PBAT: tools for family-based association studies. Am J
Hum Genet 74, 367–369.
Lotta, T., Vidgren, J., Tilgmann, C., Ulmanen, I., Melen, K.,
Julkunen, I. & Taskinen, J. (1995) Kinetics of human soluble
and membrane-bound catechol O-methyltransferase: a revised
mechanism and description of the thermolabile variant of the
enzyme. Biochemistry 34, 4202–4210.
Luck, S.J., Woodman, G.F. & Vogel, E.K. (2000) Event-
related potential studies of attention. Trends Cogn Sci 4,
432–440.
Mattay, V.S., Berman, K.F., Ostrem, J.L., Esposito, G., Van Horn,
J.D., Bigelow, L.B. & Weinberger, D.R. (1996) Dextroamphet-
amine enhances ‘‘neural network-specific’’ physiological sig-
nals: a positron-emission tomography rCBF study. J Neurosci
16, 4816–4822.
Mattay, V.S., Goldberg, T.E., Fera, F., Hariri, A.R., Tessitore, A.,
Egan, M.F., Kolachana, B., Callicott, J.H. & Weinberger, D.R.
(2003) CatecholO-methyltransferase val158-met genotype and
individual variation in the brain response to amphetamine. Proc
Natl Acad Sci USA 100, 6186–6191.
McCarthy, G., Puce, A., Belger, A. & Allison, T. (1999) Electro-
physiological studies of human face perception. II: response
properties of face-specific potentials generated in occipito-tem-
poral cortex. Cereb Cortex 9, 431–444.
Meyer-Lindenberg, A., Kohn, P.D., Kolachana, B., Kippenhan,
S., McInerney-Leo, A., Nussbaum, R., Weinberger, D.R. &
Berman, K.F. (2005) Midbrain dopamine and prefrontal function
in humans: interaction and modulation by COMT genotype.
Nat Neurosci 8, 594–596.
Palmatier, M.A., Kang, A.M. & Kidd, K.K. (1999) Global vari-
ation in the frequencies of functionally different catechol-O-
methyltransferase alleles. Biol Psychiatry 46, 557–567.
Palomba, D., Sarlo, M., Angrilli, A., Mini, A. & Stegagno, L. (2000)
Cardiac responses associated with affective processing of
unpleasant film stimuli. Int J Psychophysiol 36, 45–57.
Pizzagalli, D., Regard, M. & Lehmann, D. (1999) Rapid emotional
face processing in the human right and left brain hemispheres:
an ERP study. Neuroreport 10, 2691–2698.
Pizzagalli, D., Lehmann, D., Hendrick, A.M., Regard, M. &
Pascual-Marqui, R.D. (2002) Affective judgments of faces
modulate early activity (approximately 160 ms) within the
fusiform gyri. Neuroimage 16, 663–677.
Pollak, S.D., Klorman, R., Thatcher, J.E. & Cicchetti, D. (2001) P3b
reflects maltreated children’s reactions to facial displays of
emotion. Psychophysiology 38, 267–274.
Puce, A., Allison, T., Bentin, S. & McCarthy, G. (1998) Temporal
cortex activation in humans viewing eye and mouth move-
ments. J Neurosci 8, 2188–2199.
Puce, A., Truett, A. & McCarthy, G. (1999) Electrophysiological
studies of human face perception. III: Effect of top-down
processing on face-specific potentials. Cereb Cortex 9, 445–458.
Rossion, B., Gauthier, I., Tarr, M.J., Despland, P., Bruyer, R.,
Linotte, S. & Crommelinck,M. (2000) TheN170 occipito-temporal
component is delayed and enhanced to inverted faces but not
to inverted objects: an electrophysiological account of face--
specific processes in the human brain. Neuroreport 11, 69–74.
Sagiv, N. & Bentin, S. (2001) Structural encoding of human and
schematic faces: holistic and part-based processes. J Cogn
Neurosci 13, 937–951.
Schweinberger, S.R. & Burton, A.M. (2003) Covert recognition
and the neural system for face processing. Cortex 39, 9–30.
Schweinberger, S.R., Pickering, E.C., Burton, A.M. & Kaufmann,
J.M. (2002) Human brain potential correlates of repetition priming
in face and name recognition. Neuropsychologia 40, 2057–2073.
Smolka, M.N., Schumann, G., Wrase, J., Grusser, S.M., Flor, H.,
Mann, K., Braus, D.F., Goldman, D., Buchel, C. & Heinz, A.
(2005) Catechol-O-methyltransferase val158met genotype af-
fects processing of emotional stimuli in the amygdala and
prefrontal cortex. J Neurosci 25, 836–842.
Spielman, R.S. & Weinshilboum, R.M. (1981) Genetics of red cell
COMT activity: analysis of thermal stability and family data. Am
J Med Genet 10, 279–290.
Spielman, R.S., McGinnis, R.E. & Ewens, W.J. (1993) Trans-
mission test for linkage disequilibrium: the insulin gene region
and insulin-dependent diabetes mellitus (IDDM). Am J Hum
Genet 52, 506–516.
Steen, K.V. & Lange, C. (2005) PBAT: a comprehensive software
package for genome-wide association analysis of complex
family-based studies. Hum Genomics 2, 1–3.
Taylor, M.J., McCarthy, G., Saliba, E. & Degiovanni, E. (1999) ERP
evidence of developmental changes in processing of faces.
Clin Neurophysiol 110, 910–915.
Taylor, M.J., Batty, M. & Itier, R.J. (2004) The faces of develop-
ment: a review of early face processing over childhood. J Cogn
Neurosci 16, 1426–1442.
van Beijsterveldt, C.E. & Van Baal, G.C. (2002) Twin and
family studies of the human electroencephalogram: a review
and a meta-analysis. Biol Psychol 61, 111–129.
Vuillemier, P. (2002) Facial expression and selective attention.
Curr Opin Psychiatry 15, 291–300.
Genes, Brain and Behavior (2007) 6: 364–374 373
COMT genotype and face processing in children

Weinberger, D.R., Egan, M.F., Bertolino, A., Callicott, J.H.,
Mattay, V.S., Lipska, B.K., Berman, K.F. & Goldberg, T.E.
(2001) Prefrontal neurons and the genetics of schizophrenia.
Biol Psychiatry 50, 825–844.
Weinshilboum, R.M., Otterness, D.M. & Szumlanski, C.L. (1999)
Methylation pharmacogenetics: catechol O-methyltransferase,
thiopurine methyltransferase, and histamine N-methyltransfer-
ase. Annu Rev Pharmacol Toxicol 39, 19–52.
Acknowledgments
Supported by the Italian Ministry of University and Research (Co-
Fin grant 11/2001-113555_004 to M. B. ), the Anna Villa & Felice
Rusconi Foundation and the LiberaMente Foundation. The au-
thors of this paper do not recognize any form of a conflict of
interest. We thank Linda Camras, L. Brenna, N. Stillitano, and all
children, parents and teachers who took part in this study.
Battaglia et al.
374 Genes, Brain and Behavior (2007) 6: 364–374
Related Documents











