Catechol-O-methyltransferase Val158Met polymorphism and altered COMT gene expression in the prefrontal cortex of suicide brains Lisheng Du a, ⁎, Zul Merali a , Michael O. Poulter b , Miklós Palkovits c , Gábor Faludi d , Hymie Anisman e a University of Ottawa Institute of Mental Health Research, Ottawa, Ontario, Canada b Molecular Brain Research Group, Robarts Research Institute, University of Western Ontario, London, Ontario, Canada c Neuromorphological and Neuroendocrine Research Laboratory, Hungarian Academy of Sciences and Semmelweis University, Budapest, Hungary d Department of Clinical and Theoretical Mental Health, Semmelweis University, Budapest, Hungary e Institute of Neuroscience, Carleton University, Ottawa, Ontario, Canada abstract article info Article history: Received 2 October 2013 Received in revised form 6 December 2013 Accepted 17 December 2013 Available online 2 January 2014 Keywords: COMT polymorphism Depression mRNA expression Prefrontal cortex Suicide Catechol-O-methyltransferase (COMT) plays a key role in the degradation of catecholamine neurotransmitters within the brain. A functional polymorphism COMT Val158Met has been associated with psychiatric disorders including suicidal behavior. In the present study we examined whether this polymorphism was related to COMT mRNA expression in frontal cortical regions, and whether the expression of COMT differed between depressed suicide victims and psychiatric healthy controls. The Val158Met polymorphism was determined by polymerase chain reaction and restriction fragment length polymorphism (PCR–RFLP) analysis. The levels of COMT mRNA expression in the frontopolar cortex (FPC; 29 suicides vs. 27 controls) and orbital frontal cortex (OFC; 19 suicides vs. 15 controls) were significantly increased among depressed individuals that died by suicide relative to those of controls, being up-regulated by approximately 60% and 65% in the FPC and OFC, respectively. Furthermore, among individuals with the Met allele (Met/Met and Met/Val genotypes) who died by suicide COMT mRNA expression was elevated relative to that of the nondepressed Met allele carriers. However, significant differences were not detected between suicides (n = 49) and controls (n = 72) with respect to the Val158Met genotypic distribution and allelic frequencies. These results are consistent with the perspective that altered COMT mRNA expression in frontal cortical brain regions might contribute to suicide and/ or depression, further supporting the role of dysregulation of catecholaminergic pathway genes in the pathophysiology of suicide behaviors. © 2014 Elsevier Inc. All rights reserved. 1. Introduction Suicide is a major public health concern, with approximately one million people committing suicide world-wide each year (WHO, 2012). Risk factors for suicidal behavior include, among other things, psychiatric and medical illness, impulsivity, aggression, alcohol and drug abuse, as well as family/social stress (Mann, 2003; Pandey, 2011). Of these, suicide is most prominent within the context of psychiatric illness, particularly depressive disorders. Dissociating to what extent any given neurobiolog- ical factor underlies depression versus suicide, however, is very difficult, although suicide is often preceded by negative life events that might interact with genetic factors (Roy, 2012). Neurobiological studies of depression/suicide have revealed several correlates of suicide/depression, including disturbances of serotonin (5-HT) receptors and of the serotonin transporter (5-HTT) (Jasinska et al., 2012; Snyder, 2011), growth factors, such as brain derived neurotrophic factors (BDNF) and fibroblast growth factor-2 (FGF-2) (Duman and Monteggia, 2006; Dwivedi et al., 2005; Jeon et al., 2012; Karege et al., 2005; Turner et al., 2012), as well as hormones, such as estrogen/progesterone (Baca-Garcia et al., (2010)), FSH (Kim et al., 2013), and corticotropin-releasing hormone (Austin et al., 2003). In addition, there has been evidence linking dysregulation of catechol- aminergic pathways with the development of depression/suicide (Pitchot et al., 2003; Ryding et al., 2008; Tsai et al., 2011). In this regard, particular attention has focused on the relationship between catechol- O-methyltransferase (COMT) and depression/suicide. Ordinarily, this enzyme catalyzes the transfer of a methyl group to catecholamines (e.g., dopamine and norepinephrine) and thus degradation of catecholamine Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183 Abbreviations: ANCOVA, analysis of covariance; ANOVA, analysis of variance; BDNF, brain derived neurotrophic factors; COMT, catechol-O-methyltransferase; CSF, cerebrospi- nal fluid; DAT, dopamine transporter; DLPFC, dorsolateral prefrontal cortex; DSM-IV, Diagnostic and Statistical Manual of Mental Disorders, 4th Edition; FGF-2, fibroblast growth factor-2; FPC, frontopolar cortex; GABA, γ-aminobutyric acid; GAPDH, glyceralde- hyde 3-phosphate dehydrogenase; 5-HT, serotonin; 5-HTT, serotonin transporter; 5- HTTLPR, serotonin-transporter-linked polymorphic region; Met, methionine; OFC, orbital frontal cortex; PFC, prefrontal cortex; PMI, postmortem interval; RIN, RNA integrity num- ber; SNP, single-nucleotide polymorphism; Val, valine. ⁎ Corresponding author at: University of Ottawa Institute of Mental Health Research, 1145 Carling Ave., Ottawa, Ontario K1Z 7K4, Canada. Tel.: +1 613 722 6521x7052; fax: +1 613 792 3935. E-mail address: [email protected] (L. Du). 0278-5846/$ – see front matter © 2014 Elsevier Inc. All rights reserved. http://dx.doi.org/10.1016/j.pnpbp.2013.12.016 Contents lists available at ScienceDirect Progress in Neuro-Psychopharmacology & Biological Psychiatry journal homepage: www.elsevier.com/locate/pnp

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
Contents lists available at ScienceDirect
Progress in Neuro-Psychopharmacology & BiologicalPsychiatry
j ourna l homepage: www.e lsev ie r .com/ locate /pnp
Catechol-O-methyltransferase Val158Met polymorphism and alteredCOMT gene expression in the prefrontal cortex of suicide brains
Lisheng Du a,⁎, Zul Merali a, Michael O. Poulter b, Miklós Palkovits c, Gábor Faludi d, Hymie Anisman e
a University of Ottawa Institute of Mental Health Research, Ottawa, Ontario, Canadab Molecular Brain Research Group, Robarts Research Institute, University of Western Ontario, London, Ontario, Canadac Neuromorphological and Neuroendocrine Research Laboratory, Hungarian Academy of Sciences and Semmelweis University, Budapest, Hungaryd Department of Clinical and Theoretical Mental Health, Semmelweis University, Budapest, Hungarye Institute of Neuroscience, Carleton University, Ottawa, Ontario, Canada
Abbreviations: ANCOVA, analysis of covariance; ANObrain derived neurotrophic factors; COMT, catechol-O-metnal fluid; DAT, dopamine transporter; DLPFC, dorsolateDiagnostic and Statistical Manual of Mental Disorders,growth factor-2; FPC, frontopolar cortex; GABA, γ-aminobhyde 3-phosphate dehydrogenase; 5-HT, serotonin; 5-HTTLPR, serotonin-transporter-linked polymorphic regionfrontal cortex; PFC, prefrontal cortex; PMI, postmortem inber; SNP, single-nucleotide polymorphism; Val, valine.⁎ Corresponding author at: University of Ottawa Instit
1145 Carling Ave., Ottawa, Ontario K1Z 7K4, Canada. Tel+1 613 792 3935.
E-mail address: [email protected] (L. Du).
0278-5846/$ – see front matter © 2014 Elsevier Inc. All rihttp://dx.doi.org/10.1016/j.pnpbp.2013.12.016
a b s t r a c t
a r t i c l e i n f oArticle history:Received 2 October 2013Received in revised form 6 December 2013Accepted 17 December 2013Available online 2 January 2014
Keywords:COMT polymorphismDepressionmRNA expressionPrefrontal cortexSuicide
Catechol-O-methyltransferase (COMT) plays a key role in the degradation of catecholamine neurotransmitterswithin the brain. A functional polymorphism COMT Val158Met has been associated with psychiatric disordersincluding suicidal behavior. In the present study we examined whether this polymorphism was related toCOMT mRNA expression in frontal cortical regions, and whether the expression of COMT differed betweendepressed suicide victims and psychiatric healthy controls. The Val158Met polymorphism was determined bypolymerase chain reaction and restriction fragment length polymorphism (PCR–RFLP) analysis. The levels ofCOMT mRNA expression in the frontopolar cortex (FPC; 29 suicides vs. 27 controls) and orbital frontal cortex(OFC; 19 suicides vs. 15 controls) were significantly increased among depressed individuals that died by suiciderelative to those of controls, being up-regulated by approximately 60% and 65% in the FPC and OFC, respectively.Furthermore, among individuals with the Met allele (Met/Met and Met/Val genotypes) who died by suicideCOMT mRNA expression was elevated relative to that of the nondepressed Met allele carriers. However,significant differences were not detected between suicides (n = 49) and controls (n = 72) with respect tothe Val158Met genotypic distribution and allelic frequencies. These results are consistent with the perspectivethat altered COMT mRNA expression in frontal cortical brain regions might contribute to suicide and/or depression, further supporting the role of dysregulation of catecholaminergic pathway genes in thepathophysiology of suicide behaviors.
© 2014 Elsevier Inc. All rights reserved.
1. Introduction
Suicide is a major public health concern, with approximately onemillion people committing suicide world-wide each year (WHO, 2012).Risk factors for suicidal behavior include, among other things, psychiatricand medical illness, impulsivity, aggression, alcohol and drug abuse, aswell as family/social stress (Mann, 2003; Pandey, 2011). Of these, suicide
VA, analysis of variance; BDNF,hyltransferase; CSF, cerebrospi-ral prefrontal cortex; DSM-IV,4th Edition; FGF-2, fibroblastutyric acid; GAPDH, glyceralde-HTT, serotonin transporter; 5-; Met, methionine; OFC, orbitalterval; RIN, RNA integrity num-
ute of Mental Health Research,.: +1 613 722 6521x7052; fax:
ghts reserved.
is most prominent within the context of psychiatric illness, particularlydepressive disorders. Dissociating to what extent any given neurobiolog-ical factor underlies depression versus suicide, however, is very difficult,although suicide is often preceded by negative life events that mightinteract with genetic factors (Roy, 2012).
Neurobiological studies of depression/suicide have revealed severalcorrelates of suicide/depression, including disturbances of serotonin(5-HT) receptors and of the serotonin transporter (5-HTT) (Jasinskaet al., 2012; Snyder, 2011), growth factors, such as brain derivedneurotrophic factors (BDNF) and fibroblast growth factor-2 (FGF-2)(Duman and Monteggia, 2006; Dwivedi et al., 2005; Jeon et al., 2012;Karege et al., 2005; Turner et al., 2012), as well as hormones, such asestrogen/progesterone (Baca-Garcia et al., (2010)), FSH (Kim et al.,2013), and corticotropin-releasing hormone (Austin et al., 2003). Inaddition, there has been evidence linking dysregulation of catechol-aminergic pathways with the development of depression/suicide(Pitchot et al., 2003; Ryding et al., 2008; Tsai et al., 2011). In this regard,particular attention has focused on the relationship between catechol-O-methyltransferase (COMT) and depression/suicide. Ordinarily, thisenzyme catalyzes the transfer of amethyl group to catecholamines (e.g.,dopamine and norepinephrine) and thus degradation of catecholamine

179L. Du et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
neurotransmitters, dopamine and norepinephrine. Although COMT isexpressed throughout the human brain (Hong et al., 1998), it plays amajor role in regulating dopamine levels in the prefrontal cortex (PFC)as other ways of terminating the dopamine signal, such as the presenceof the dopamine transporter (DAT), are limited in this region (Käenmäkiet al., 2010).
Given the fundamental role of COMT in modulating prefrontalcortical dopaminergic neurotransmission, the contribution of theCOMT gene in relation to behavior has attracted attention (Huotariet al., 2002; Matsumoto et al., 2003; Shehzad et al., 2012; Tunbridgeet al., 2004). COMT is encoded by a single gene localized on chromo-some 22q11.1–q11.2. A common single-nucleotide polymorphism(SNP) Val158Met in exon 4 of the COMT gene was studied intensivelyin relation to psychiatric disorders as well as in suicidal behavior andrelated traits, such as aggression (Crisafulli et al., 2010; Currier andMann, 2008). This SNP (rs4680) affects the functional ability of theenzyme to catabolize synaptic dopamine (Tunbridge, 2010; Witte andFlöel, 2012), such that in human postmortem PFC tissue the Val/Valgenotype is associated with about 38% greater enzyme activity thanthat evident in Met/Met homozyotes (Chen et al., 2004; Tunbridgeet al., 2004, 2006).
Several studies demonstrated associations between the COMTVal158Met gene polymorphism and psychiatric disorders, includingmajor depression disorder and suicidal behavior as well as antidepres-sant treatment responses (Baune et al., 2007; Benedetti et al., 2009;Massat et al., 2004; Ono et al., 2004; Pivac et al., 2011), although contra-dictory results have also been reported (Hosák, 2007; Schosser et al.,2012). A meta-analysis of six related studies suggested an associationbetween COMT Val158Met polymorphism and suicidal behavior, andthis relationship was moderated by gender and the lethality of suicideattempt (Kia-Keating et al., 2007). In contrast, more recent meta-analyses that included new data questioned the association betweenCOMT Val158Met and suicidal behavior (Calati et al., 2011; Tovilla-Zarate et al., 2011). Although more than 16 case–control studies havebeen reported, the association between COMT Val158Met polymor-phism and suicidal behavior remains controversial, possibly owing todifferent end points (suicide vs. suicidal intent) that have been used inthese analyses.
Since suicidal behavior ranges from suicidal ideation to suicideattempts and completed suicide, it is possible that depressed individualsthat completed suicide, obviously the most severe form of suicidalbehavior, represent amore homogenous group than groups that includesuicidal ideation and suicide attempts (suicide attempts with non-fataloutcomes) (Kia-Keating et al., 2007; Supriyanto et al., 2011). Indeed, as-sociation studies suggested that the lethality of suicide attempt may beassociated with certain gene polymorphism, such as 5-HTTLPR (Gondaet al., 2011) and COMT Val158Met (Kia-Keating et al., 2007). However,only two studies concerning the association between COMT Val158Metpolymorphism and completed suicide have been reported (Ono et al.,2004; Pivac et al., 2011). Given the different outcomes observed in thepopulations studied, additional analyses of completed suicides areneeded in order to determine whether there is an association betweenCOMT Val158Met polymorphism and suicidal behavior, and important-ly, to determinewhether the Val158Met polymorphism is, in fact, relat-ed to altered COMT mRNA expression levels. In this regard, there havebeen few reports documenting the relationship between the Val158Met
Table 1Age and cause of death among male and female control and suicide participants in the associa
Subjects Age (year)
Control female 67.35 ± 17.23Control male 61.65 ± 14.60Suicide female 49.77 ± 16.89Suicide male 48.06 ± 15.28
ACF = acute cardiac failure; AMI = acute myocardial infarction; TC = traffic accident; poison
polymorphism and COMT mRNA expression levels. COMT mRNA ex-pression in human brainwas found to be related to a haplotype contain-ing Val158Met in schizophrenia, bipolar disorder and major depression(Bray et al., 2003; Dempster et al., 2006; Zhu et al., 2004), but, the ab-sence of a correlation between COMT genotypes and expression of itsmRNA was also reported (Chen et al., 2004; Tunbridge et al., 2004). In-terestingly, Matsumoto et al. (2003) found a disease-related laminardifference of COMT expression in the dorsolateral prefrontal cortex(DLPFC) of patients with schizophrenia; however, the relation betweenthe COMT Val158Met polymorphism and COMT expression levels with-in the depressed suicide brain is still uncertain.
The goal of the present study was to determine whether or notdifferences existed in the distribution of COMT Val158Met polymor-phism in depressed individuals that died by suicide relative to that evi-dent in nondepressed individuals that died of causes other than suicide.Aswell, in a subset of samples from the FPC andOFCwe assessed poten-tial differences of COMTmRNA expression in completed suicide relativeto control subjects, and determined the relation between COMTVal158Met polymorphism and COMT mRNA expression in both ofthese conditions.
2. Materials and methods
All procedures, including tissue harvesting, were approved by thelocal ethics committee (Semmelweis) and the Research Ethics Boardof the Royal Ottawa Health Care Group approved analyses of tissuesamples at University of Ottawa Institute of Mental Health Research.Analyses were performed in accordance with the Declaration ofHelsinki. Tissue harvesting occurred after written informed consentwas obtained from next of kin, which included the request to consultmedical charts and to conduct neurochemical and/or biochemicalanalyses.
2.1. Subjects
Brains from 49 depressed individuals that died by suicide (n = 35males and 14 females) and from 72 control participants (n = 46males and 26 females), who died from causes not directly involvingany diseases of the central nervous system, were obtained at autopsyat the Department of Forensic Medicine of the Semmelweis UniversityMedical School (Budapest), at the Department of Neuropathology,National Institute of Psychiatry and Neurology, Budapest, and at theDepartment of Pathology of Saint George Hospital, Székesfehérvár,Hungary. The microdissected brains were stored in the Human BrainTissue Bank, Budapest. Tissue samples from some of these brains werepreviously used in studies assessing serotonin receptors (Anismanet al., 2008), and epigenetic differences regarding the GABAA subunits(Poulter et al., 2008, 2010).
Table 1 provides the age and cause of death of the control partici-pants and those that died by suicide for which genotypingwas conduct-ed. Of these participants, only a subset was used for the determinationsof COMTmRNA expression in the FPC and OFC. The sample comprised 3independent sets of tissue obtained over a 4 year period. Table 2 showsthe sex, age, postmortem interval and cause of death for each samplethat went into the qPCR analysis. Overall, control participants usedfor genotyping were older than those that died by suicide [F (1,
tion study.
Cause of death
n = 26: 17 ACF, 3 AMI, 2 stroke, 4 bronchopneumonian = 46: 31 ACF, 10 AMI, 2 TC, 1 stroke, 1 electric shock, 1 CO intoxicationn = 14: 6 hanging, 8 poisoningn = 35: 24 hanging, 3 jumping, 8 poisoning
ing (includes prescription drug overdose).
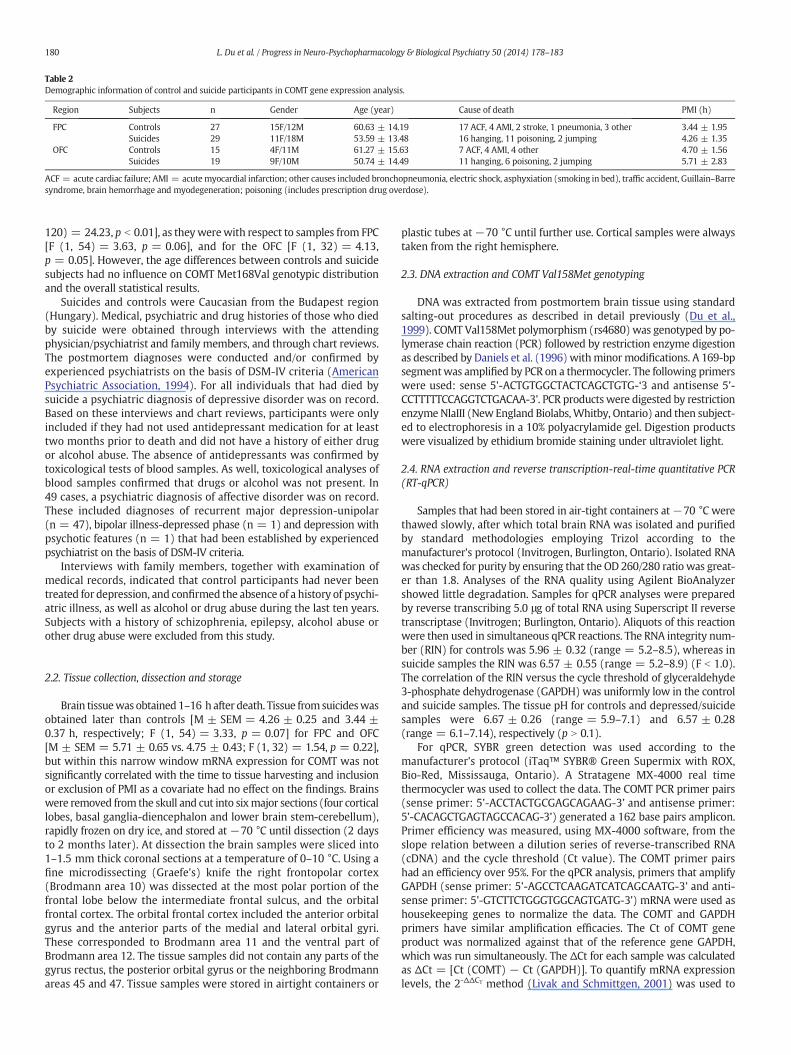
Table 2Demographic information of control and suicide participants in COMT gene expression analysis.
Region Subjects n Gender Age (year) Cause of death PMI (h)
FPC Controls 27 15F/12M 60.63 ± 14.19 17 ACF, 4 AMI, 2 stroke, 1 pneumonia, 3 other 3.44 ± 1.95Suicides 29 11F/18M 53.59 ± 13.48 16 hanging, 11 poisoning, 2 jumping 4.26 ± 1.35
OFC Controls 15 4F/11M 61.27 ± 15.63 7 ACF, 4 AMI, 4 other 4.70 ± 1.56Suicides 19 9F/10M 50.74 ± 14.49 11 hanging, 6 poisoning, 2 jumping 5.71 ± 2.83
ACF = acute cardiac failure; AMI = acutemyocardial infarction; other causes included bronchopneumonia, electric shock, asphyxiation (smoking in bed), traffic accident, Guillain–Barresyndrome, brain hemorrhage and myodegeneration; poisoning (includes prescription drug overdose).
180 L. Du et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
120) = 24.23, p b 0.01], as theywerewith respect to samples from FPC[F (1, 54) = 3.63, p = 0.06], and for the OFC [F (1, 32) = 4.13,p = 0.05]. However, the age differences between controls and suicidesubjects had no influence on COMT Met168Val genotypic distributionand the overall statistical results.
Suicides and controls were Caucasian from the Budapest region(Hungary). Medical, psychiatric and drug histories of those who diedby suicide were obtained through interviews with the attendingphysician/psychiatrist and family members, and through chart reviews.The postmortem diagnoses were conducted and/or confirmed byexperienced psychiatrists on the basis of DSM-IV criteria (AmericanPsychiatric Association, 1994). For all individuals that had died bysuicide a psychiatric diagnosis of depressive disorder was on record.Based on these interviews and chart reviews, participants were onlyincluded if they had not used antidepressant medication for at leasttwo months prior to death and did not have a history of either drugor alcohol abuse. The absence of antidepressants was confirmed bytoxicological tests of blood samples. As well, toxicological analyses ofblood samples confirmed that drugs or alcohol was not present. In49 cases, a psychiatric diagnosis of affective disorder was on record.These included diagnoses of recurrent major depression-unipolar(n = 47), bipolar illness-depressed phase (n = 1) and depression withpsychotic features (n = 1) that had been established by experiencedpsychiatrist on the basis of DSM-IV criteria.
Interviews with family members, together with examination ofmedical records, indicated that control participants had never beentreated for depression, and confirmed the absence of a history of psychi-atric illness, as well as alcohol or drug abuse during the last ten years.Subjects with a history of schizophrenia, epilepsy, alcohol abuse orother drug abuse were excluded from this study.
2.2. Tissue collection, dissection and storage
Brain tissuewas obtained1–16 hafter death. Tissue fromsuicideswasobtained later than controls [M ± SEM = 4.26 ± 0.25 and 3.44 ±0.37 h, respectively; F (1, 54) = 3.33, p = 0.07] for FPC and OFC[M ± SEM = 5.71 ± 0.65 vs. 4.75 ± 0.43; F (1, 32) = 1.54, p = 0.22],but within this narrow window mRNA expression for COMT was notsignificantly correlated with the time to tissue harvesting and inclusionor exclusion of PMI as a covariate had no effect on the findings. Brainswere removed from the skull and cut into sixmajor sections (four corticallobes, basal ganglia-diencephalon and lower brain stem-cerebellum),rapidly frozen on dry ice, and stored at −70 °C until dissection (2 daysto 2 months later). At dissection the brain samples were sliced into1–1.5 mm thick coronal sections at a temperature of 0–10 °C. Using afine microdissecting (Graefe's) knife the right frontopolar cortex(Brodmann area 10) was dissected at the most polar portion of thefrontal lobe below the intermediate frontal sulcus, and the orbitalfrontal cortex. The orbital frontal cortex included the anterior orbitalgyrus and the anterior parts of the medial and lateral orbital gyri.These corresponded to Brodmann area 11 and the ventral part ofBrodmann area 12. The tissue samples did not contain any parts of thegyrus rectus, the posterior orbital gyrus or the neighboring Brodmannareas 45 and 47. Tissue samples were stored in airtight containers or
plastic tubes at−70 °C until further use. Cortical samples were alwaystaken from the right hemisphere.
2.3. DNA extraction and COMT Val158Met genotyping
DNA was extracted from postmortem brain tissue using standardsalting-out procedures as described in detail previously (Du et al.,1999). COMT Val158Met polymorphism (rs4680) was genotyped by po-lymerase chain reaction (PCR) followed by restriction enzyme digestionas described by Daniels et al. (1996)withminormodifications. A 169-bpsegmentwas amplified by PCR on a thermocycler. The following primerswere used: sense 5’-ACTGTGGCTACTCAGCTGTG-‘3 and antisense 5’-CCTTTTTCCAGGTCTGACAA-3’. PCR products were digested by restrictionenzymeNlaIII (New England Biolabs,Whitby, Ontario) and then subject-ed to electrophoresis in a 10% polyacrylamide gel. Digestion productswere visualized by ethidium bromide staining under ultraviolet light.
2.4. RNA extraction and reverse transcription-real-time quantitative PCR(RT-qPCR)
Samples that had been stored in air-tight containers at−70 °C werethawed slowly, after which total brain RNA was isolated and purifiedby standard methodologies employing Trizol according to themanufacturer's protocol (Invitrogen, Burlington, Ontario). Isolated RNAwas checked for purity by ensuring that the OD 260/280 ratio was great-er than 1.8. Analyses of the RNA quality using Agilent BioAnalyzershowed little degradation. Samples for qPCR analyses were preparedby reverse transcribing 5.0 μg of total RNA using Superscript II reversetranscriptase (Invitrogen; Burlington, Ontario). Aliquots of this reactionwere then used in simultaneous qPCR reactions. The RNA integrity num-ber (RIN) for controls was 5.96 ± 0.32 (range = 5.2–8.5), whereas insuicide samples the RIN was 6.57 ± 0.55 (range = 5.2–8.9) (F b 1.0).The correlation of the RIN versus the cycle threshold of glyceraldehyde3-phosphate dehydrogenase (GAPDH) was uniformly low in the controland suicide samples. The tissue pH for controls and depressed/suicidesamples were 6.67 ± 0.26 (range = 5.9–7.1) and 6.57 ± 0.28(range = 6.1–7.14), respectively (p N 0.1).
For qPCR, SYBR green detection was used according to themanufacturer's protocol (iTaq™ SYBR® Green Supermix with ROX,Bio-Red, Mississauga, Ontario). A Stratagene MX-4000 real timethermocycler was used to collect the data. The COMT PCR primer pairs(sense primer: 5’-ACCTACTGCGAGCAGAAG-3’ and antisense primer:5’-CACAGCTGAGTAGCCACAG-3’) generated a 162 base pairs amplicon.Primer efficiency was measured, using MX-4000 software, from theslope relation between a dilution series of reverse-transcribed RNA(cDNA) and the cycle threshold (Ct value). The COMT primer pairshad an efficiency over 95%. For the qPCR analysis, primers that amplifyGAPDH (sense primer: 5’-AGCCTCAAGATCATCAGCAATG-3’ and anti-sense primer: 5’-GTCTTCTGGGTGGCAGTGATG-3’) mRNA were used ashousekeeping genes to normalize the data. The COMT and GAPDHprimers have similar amplification efficacies. The Ct of COMT geneproduct was normalized against that of the reference gene GAPDH,which was run simultaneously. The ΔCt for each sample was calculatedas ΔCt = [Ct (COMT) − Ct (GAPDH)]. To quantify mRNA expressionlevels, the 2-ΔΔCT method (Livak and Schmittgen, 2001) was used to

181L. Du et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
convert ΔCt values to mRNA fold changes relative to the female controlgroup (calibrator).
2.5. Statistical analyses
All statistical analyses were performed using the Statistical Packagefor Social Sciences (SPSS) for windows version 11.5 (SPSS, Chicago,IL). Allele and genotype frequencieswere compared between depressedsuicides and control subjects by a chi-square analysis. Hardy–Weinbergequilibrium for the distribution of genotypes was tested with the chi-square test for quality of fit. The level of significancewas set at p = 0.05.
Effects of Val158Met variation on gene expression were examinedusing ANCOVA with genotype and diagnosis as independent variablesand age and postmortem interval as covariates. The mRNA expressionof COMT was analyzed by a 2 (Suicide vs. Control) × 2 (Male vs.Female) ANOVA. Using Grubb's test, one identified outlier was deleted.Deleting this one outlier did not change overall statistical results.
3. Results
3.1. Association study
Genotypic distributions of those that died by suicide and controlsubjects were in Hardy–Weinberg equilibrium (χ2 = 2.44, p = 0.12,and χ2 = 2.36, p = 0.13, respectively). No significant differenceswere observed between suicides and controls in genotypic distribution(χ2 = 0.06, p = 0.97) and allelic frequencies (χ2 = 0.01, p = 0.92).Likewise, the genotypic distribution did not differ when the analysisincluded gender (Table 3).
3.2. COMT expression in the frontopolar cortex and orbital frontal cortex
The age of participants at the time of death and postmortem interval(PMI) were not significantly correlated with COMT mRNA expression(r = −0.10, p = 0.46 and r = 0.13, p = 0.33, respectively) in theFPC. Analyses of COMT mRNA expression within the FPC indicatedthat expression levels were significantly higher in suicides than in con-trols [F (1, 54) = 15.29, p = 0.001]. Therewas no expression differencebetween genders [F (1, 52) = 0.02, p = 0.88], and no Group × Genderinteraction was detected [F (1, 52) = 0.09, p = 0.76]. Altered COMTmRNA expression was also observed in the OFC [F (1, 32) = 13.17,p = 0.001]. The levels of COMT mRNA expression in this region weresignificantly increased in individuals that died by suicide relative tothose that died of other causes, being up-regulated by approximately60% and 65% in the FPC and OFC, respectively.
3.3. Relationship between Val158Met genotype and COMT expression
Since prior studies on the relation of suicide and COMT gene focusedon gene polymorphisms rather than on gene function, we also explored
Table 3Genotype and allele distributions of COMT 158Val/Met polymorphism in depressed/suicide indiv
COMT Val108/158Met
Genotypes
Val/Val (%) Val/Met (%)
Controls 22 (30.5) 30 (41.7)Suicides 16 (32.7) 19 (38.8)
χ2 = 0.11; df = 2; p = 0.95
Male controls 15 (31.9) 18 (38.3)Male suicides 13 (37.1) 16 (45.7)
χ2 = 1.74; df = 2; p = 0.42
Female controls 7 (29.2) 11 (45.8)Female suicides 3 (21.4) 3 (21.4)
χ2 = 4.13; df = 2; p = 0.13
the relation between COMT Val158Met genotypes and their mRNAexpression in the FPC and OFC. No association was found betweengenotype and mRNA expression [F(2,53) = 0.55, p = 0.58], and genderhad no influence in relation to COMT expression [F(1,54) = 0.24,p = 0.63] in the analyses of either the FPC or the OFC in the pooledsuicides and controls [mRNA expression: F(2,31) = 0.77, p = 0.47and gender effect: F(1,32) = 3.3, p = 0.08]. Importantly, however, aGroup (suicides or controls) × Genotype interaction onCOMTexpressionwas apparent within the FPC [F(2,50) = 5.46, p = 0.007; Table 4].Individuals with the Met allele (Met/Met and Met/Val genotypes) whodied by suicide displayed higher COMT mRNA expression than did theMet allele carriers who were not depressed (Fig. 1). A similar differencewas not apparent among those carrying the Val/Val genotype. In contrastto the FPC, there was no Group × Genotype interaction on gene expres-sion in the OFC [F(2,28) = 2.1, p = 0.14]. Correlational analyses alsorevealed that genotype was correlated with COMT gene expression inthe FPC of control samples (Spearman's rho = 0.63, p = .0001), butnot in the FPC of suicide samples (Spearman's rho = −0.25, p = 0.19).In the OFC, significant correlations were not detected in either controlsor suicides (p N 0.30; data not shown).
4. Discussion
As reported in recent meta-analyses relating COMT polymorphismand suicidal intentions (Calati et al., 2011; Tovilla-Zarate et al., 2011),in the present study, significant associations were not observedbetween suicide completers and either the genotype distribution orallelic frequencies of the COMT Val158Met polymorphism. To ourknowledge, only two prior studies investigated the association betweenCOMT Val158Met and suicide completion, as opposed to suicideattempts (Ono et al., 2004; Pivac et al., 2011). It had been suggestedbased on a Japanese sample that the Val/Val genotype served as aprotective factor against suicide in males, as the Val/Val genotypeoccurred at a higher frequency in male controls than in male suicides.In contrast, Pivac et al. (2011) showed that in a Caucasian populationof Slovenian origin there was an overrepresentation of the Met/Met inmale control subjects compared with males that died by suicide,suggesting that Met/Met genotype might be a protective factor againstsuicide. The discrepancies between these studies and with that of thepresent investigation in which an association was not observedbetween the genotype and the occurrence of suicide might be due toany number of factors. Significantly, the suicide cases in the presentstudy were determined on the basis of psychological autopsy andincluded only individuals who had been depressed, whereas the twoearlier studies also involved individuals that had died by suicide, butwithout a psychological autopsy having been performed for diagnosticpurposes. Thus, it is uncertain whether these studies, in fact, involvedsuicides related to depression or suicides associated with other condi-tions. Furthermore, ethnic differences in genotype and allele frequencydistributions exist between Caucasian and Japanese male control
iduals and nondepressed individuals.
Alleles
Met/Met (%) Val (%) Met (%)
20 (27.8) 74 (51.4) 70 (48.6)14 (28.6) 51 (52.0) 47 (48.0)
χ2 = 0.01; df = 1; p = 0.92
14 (29.8) 48 (51.0) 46 (49.0)6 (17.1) 42 (60.0) 28 (40.0)
χ2 = 1.29; df = 1; p = 0.25
6 (25.0) 25 (52.1) 23 (47.9)8 (57.1) 9 (32.1) 19 (67.9)
χ2 = 2.84; df = 1; p = 0.09

Table 4ANCOVA analysis of COMT gene expression in the PFC.
Brain region Variable F df p
FPC (n = 56) Diagnosis (suicides vs. controls) 15.29 1 b0.001⁎⁎⁎
Val158Met 0.55 2 0.58Gender 0.24 1 0.63Diagnosis × Val158Met 5.45 2 0.007⁎⁎
Diagnosis × Gender 0.09 1 0.76OFC (n = 34) Diagnosis (suicides vs. controls) 13.77 1 0.001⁎⁎
Val158Met 1.11 2 0.34Gender 3.29 1 0.08Diagnosis × Val158Met 2.11 2 1.40Diagnosis × Gender 0.11 1 0.74
⁎⁎ p b .01.⁎⁎⁎ p b 0.001.
182 L. Du et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
samples (Pivac et al., 2011), which might also have contributed to thedifferences observed across studies.
Although the COMT Val158Met variant has been shown to havemajor effects on enzyme activity, its actions on COMTmRNA expressionlevels in the FPC andOFC of depressed suicides have not previously beenexamined. It has been shown that a haplotype that contained the Valallele was associated with reduced COMT expression in normalhuman brain (Bray et al., 2003), and it was also demonstrated that theVal158Met genotype was positively related to COMT gene expressionin the cerebellum (Dempster et al., 2006; Zhu et al., 2004). However,this polymorphism in the dorsolateral prefrontal cortex was unrelatedto major depression, schizophrenia, and bipolar disorder, (Chen et al.,2004; Tunbridge et al., 2004). In the present investigation the COMTVal158Met variant was related to COMT mRNA expression levels inthe controls, but not in suicide brains. Indeed, among those that diedby suicide who carried the homozygous Met/Met alleles, higher COMTmRNA expression was evident than in those who carried Val/Met andVal/Val genotypes, whereas in controls Met/Met carriers expressedlower COMT mRNA. This is in agreement with previous findings(Abdolmaleky et al., 2006) indicating that in suicide samples (15 bipolarand 7 schizophrenia suicides) expression of membrane-boundcatechol-O-methyltransferase (MB-COMT is predominantly expressedin the brain; Tenhunen et al., 1994) was higher than that in controls,possibly owing to hypomethylation of the MB-COMT promoter. In thisparticular study the Met homozygosity was accompanied by approxi-mately two-foldmore COMTmRNA in those that died by suicide relativeto controls. It is uncertain whether the higher COMT mRNA expressionin suicide samples of the present study was due to hypomethylationof the COMT promoter. However, given that depression and suicideare associated with considerable distress, it might be significant that
Fig. 1.Normalized COMTmRNAexpression in FPCof depressed suicide victims and controlsubjects with genotypes Met/Met (control: n = 10 vs. suicide: n = 6), Met/Val (control:n = 11 vs. suicide: n = 12), and Val/Val (control: n = 6 vs. suicide: n = 11). *p b 0.05;**p b 0.01.
greater stress was associated with lower methylation of the Val158 al-lele in peripheral blood mononuclear cells of Val/Val individuals(Ursini et al., 2011).
In the current study, we detected an alteration of COMT mRNAexpression in the PFC of suicide victims, but significant associationswere not observed in the COMT Val158Met genotypic distribution orallelic frequencies between suicide completers and controls. The lackof an association may be due to small sample size. It is also possiblethat no associations may indicate that the Val158Met polymorphism ismost likely not a major player in the genetic susceptibility to suicide.Our results agree with previous reports that the levels of COMT mRNAexpression were elevated in schizophrenia, bipolar disorder and suicide(Abdolmaleky et al., 2006; Matsumoto et al., 2003), suggesting thatincreased dopamine degradation in the PFC plays a role in relationwith suicide.
The sample size in the present study was admittedly small, but thefrequencies of the COMT Val158Met genotype were similar to that of aprevious report in healthy East European controls (Nedic et al., 2010).This said, suicide is a complex behavior that likely involves multiplegenes and gene × environment interactions, and hence it can beexpected that any single variable would have small to modest geneticeffects in relation to completed suicide (Kendler, 2010). Although thepresent study, likemany association studies, has limited power to detectsmall effects, it may nevertheless contribute to future (meta-) analysesthat combine several studies in an effort to overcome difficulties relatedto small sample sizes.
In addition, the alteration of COMT mRNA levels in suicide brainsmight also be caused by altered mRNA expression and/or degra-dation rates owing to other factors, such as methylation levels andtranscriptional/post-transcriptional regulation, as well as that mRNAexpression do not necessarily reflect the final protein levels/expression.
Summarizing, the present study revealed that COMT mRNA expres-sion levels were significantly higher in the FPC and OFC of depressedindividuals that died by suicide than among nondepressed individualsthat died as a result of factors other than suicide. The increased COMTexpression might be associated with lower dopaminergic activity inthese regions, which could potentially have been related to depres-sion/suicide. Several studies have indeed suggested an associationbetween dopaminergic hypoactivity and suicide. For example, decreaseddopaminergic neurotransmission has been reported in depressivedisorders (see reviews by Dailly et al., 2004; Dunlop and Nemeroff,2007; Pitchot et al., 2001) and reduced dopamine turnover wasobserved in the basal ganglia of depressed suicides (Bowden et al.,1997) and CSF of depressed suicide attempters (Sher et al., 2006). Tobe sure, being correlational, the present data, like those of earlierstudies, do not speak to whether dopamine plays a causal role insuicide/depression. Moreover, it remains to be established whetherthe altered COMT mRNA expression in the present study was relatedto depression or was unique to individuals that died by suicide. Never-theless, the data available are in line with the view that dopaminefunctioning plays a role in relation to depression/suicide (Berton et al.,2006; Nestler and Carlezon, 2006).
Acknowledgments
This researchwas supported by an OntarioMental Health Foundationoperating grant to LD, HA, and ZM and by the European Union GrantNo. FP6, BNEII No. LSHM-CT-2004-503039 to MP. HA holds a CanadaResearch Chair. We thank Yue He and Jill Chatt for their technical assis-tance. The authors report no biomedical financial interests or potentialconflicts of interest.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.pnpbp.2013.12.016.

183L. Du et al. / Progress in Neuro-Psychopharmacology & Biological Psychiatry 50 (2014) 178–183
References
Abdolmaleky HM, Cheng KH, Faraone SV, Wilcox M, Glatt SJ, Gao F, et al. Hypomethylationof MB-COMT promoter is a major risk factor for schizophrenia and bipolar disorder.Hum Mol Genet 2006;15:3132–45.
American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders.4th ed. Washington, DC: American Psychiatric Press; 1994.
Anisman H, Du L, Palkovits M, Faludi G, Kovacs GG, Szontagh-Kishazi P, et al. Serotoninreceptor subtype and p11 mRNA expression in stress-relevant brain regions of suicideand control subjects. J Psychiatry Neurosci 2008;33:131–41.
Austin MC, Janosky JE, Murphy HA. Increased corticotropin-releasing hormone immuno-reactivity in monoamine-containing pontine nuclei of depressed suicide men. MolPsychiatry 2003;8:324–32.
Baca-Garcia E, Diaz-Sastre C, Ceverino A, Perez-Rodriguez MM, Navarro-Jimenez R,Lopez-Castroman J, et al. Suicide attempts among women during low estradiol/lowprogesterone states. J Psychiatr Res 2010;44:209–14.
Baune BT, Hohoff C, Berger K, Neumann A, Mortensen S, Roehrs T, et al. Association of theCOMT val158met variant with antidepressant treatment response in major depression.Neuropsychopharmacology 2007;33:924–32.
Benedetti F, Colombo C, Pirovano A,Marino E, Smeraldi E. The catechol-O-methyltransferaseVal (108/158) Met polymorphism affects antidepressant response to paroxetine in anaturalistic setting. Psychopharmacology (Berl) 2009;203:155–60.
Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ, et al. Essential role ofBDNF in themesolimbic dopamine pathway in social defeat stress. Science 2006;311:864–8.
Bowden C, Cheetham SC, Lowther S, Katona CL, Crompton MR, Horton RW. Reduceddopamine turnover in the basal ganglia of depressed suicides. Brain Res 1997;769:135–40.
Bray NJ, Buckland PR, Williams NM, Williams HJ, Norton N, Owen MJ, et al. A haplotypeimplicated in schizophrenia susceptibility is associated with reduced COMT expressionin human brain. Am J Hum Genet 2003;73:152–61.
Calati R, Porcelli S, Giegling I, Hartmann AM, Moller HJ, De Ronchi D, et al. Catechol-o-methyltransferase gene modulation on suicidal behavior and personality traits:review, meta-analysis and association study. J Psychiatr Res 2011;45:309–21.
Chen J, Lipska BK, Halim N, Ma QD, Matsumoto M, Melhem S, et al. Functional analysis ofgenetic variation in catechol-O-methyltransferase (COMT): effects onmRNA, protein,and enzyme activity in postmortem human brain. Am J Hum Genet 2004;75:807–21.
Crisafulli C, Calati R, De Ronchi D, Sidoti A, D'Angelo R, Amato A, et al. Genetics of suicide,from genes to behavior. Clin Neuropsychiatry 2010;7:141–8.
Currier D, Mann JJ. Stress, genes and the biology of suicidal behavior. Psychiatr Clin NorthAm 2008;31:247–69.
Dailly E, Chenu F, Renard CE, Bourin M. Dopamine, depression and antidepressants.Fundam Clin Pharmacol 2004;18:601–7.
Daniels JK, Williams NM, Williams J, Jones LA, Cardno AG, Murphy KC, et al. No evidencefor allelic association between schizophrenia and a polymorphism determining highor low catechol O-methyltransferase activity. Am J Psychiatry 1996;153:268–70.
Dempster EL, Mill J, Craig IW, Collier DA. The quantification of COMT mRNA in postmortem cerebellum tissue: diagnosis, genotype, methylation and expression. BMCMed Genet 2006;7:10.
Du L, Faludi G, Palkovits M, Demeter E, Bakish D, Lapierre YD, et al. Frequency of longallele in serotonin transporter gene is increased in depressed suicides. Biol Psychiatry1999;46:196–201.
Duman RS, Monteggia LM. A neurotrophic model for stress-related mood disorders. BiolPsychiatry 2006;59:1116.
Dunlop BW, Nemeroff CB. The role of dopamine in the pathophysiology of depression.Arch Gen Psychiatry 2007;64:327–37.
Dwivedi Y, Mondal AC, Rizavi HS, Conley RR. Suicide brain is associated with decreasedexpression of neurotrophins. Biol Psychiatry 2005;58:315–24.
Gonda X, Fountoulakis KN, Harro J, Pompili M, Akiskal HS, Bagdy G, et al. The possiblecontributory role of the S allele of 5-HTTLPR in the emergence of suicidality. JPsychopharmacol 2011;25:857–66.
Hong J, Shu-Leong H, Tao X, Lap-Ping Y. Distribution of catechol-O-methyltransferaseexpression in human central nervous system. Neuroreport 1998;9:2861–4.
Hosák L. Role of the COMT gene Val158Met polymorphism in mental disorders: a review.Eur Psychiatry 2007;22:276–81.
Huotari M, SanthaM, Lucas LR, Karayiorgou M, Gogos JA, Mannisto PT. Effect of dopamineuptake inhibition on brain catecholamine levels and locomotion in catechol-O-methyltransferase-disrupted mice. J Pharmacol Exp Ther 2002;303:1309–16.
Jasinska AJ, Lowry CA, Burmeister M. Serotonin transporter gene, stress and raphe–rapheinteractions: a molecular mechanism of depression. Trends Neurosci 2012;35:395–402.
Jeon HJ, Kang ES, Lee EH, Jeong EG, Jeon JR, Mischoulon D, et al. Childhood trauma andplatelet brain-derived neurotrophic factor (BDNF) after a three month follow-up inpatients with major depressive disorder. J Psychiatr Res 2012;46:966–72.
Käenmäki M, Tammimäki A, Myöhänen T, Pakarinen K, Amberg C, Karayiorgou M, et al.Quantitative role of COMT in dopamine clearance in the prefrontal cortex of freelymoving mice. J Neurochem 2010;114:1745–55.
Karege F, Vaudan G, Schwald M, Perroud N, La Harpe R. Neurotrophin levels in postmortembrains of suicide victims and the effects of antemortem diagnosis and psychotropicdrugs. Brain Res 2005;136:29–37.
Kendler KS. Genetic and environmental pathways to suicidal behavior: reflections of agenetic epidemiologist. Eur Psychiatry 2010;25:300–3.
Kia-Keating BM, Glatt SJ, Tsuang MT. Meta-analyses suggest association between COMT,but not HTR1B, alleles, and suicidal behavior. Am J Med Genet 2007;144B:1048–53.
Kim B, Kang ES, Fava M, Mischoulon D, Soskin D, Yu BH, et al. Follicle-stimulatinghormone (FSH), current suicidal ideation and attempt in female patients withmajor depressive disorder. Psychiatry Res 2013;210:951–6.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-timequantitative PCR and the 2−xΔΔCT method. Methods 2001;25:402–8.
Mann JJ. Neurobiology of suicidal behaviour. Nat Rev Neurosci 2003;4:819–28.Massat I, Souery D, Del-Favero J, Nothen M, Blackwood D, Muir W, et al. Association
between COMT (Val158Met) functional polymorphism and early onset in patientswith major depressive disorder in a European multicenter genetic associationstudy. Mol Psychiatry 2004;10:598–605.
Matsumoto M, Weickert CS, Beltaifa S, Kolachana B, Chen J, Hyde TM, et al. CatecholO-methyltransferase (COMT) mRNA expression in the dorsolateral prefrontal cortexof patients with schizophrenia. Neuropsychopharmacology 2003;28:1521–30.
Nedic G, Nikolac M, Borovecki F, Hajnsek S, Muck-Seler D, Pivac N. Association study of afunctional catechol-O-methyltransferase polymorphism and smoking in healthyCaucasian subjects. Neurosci Lett 2010;473:216–9.
Nestler EJ, Carlezon Jr WA. The mesolimbic dopamine reward circuit in depression. BiolPsychiatry 2006;59:1151–9.
Ono H, Shirakawa O, Nushida H, Ueno Y, Maeda K. Association between catechol-O-methyltransferase functional polymorphism and male suicide completers.Neuropsychopharmacology 2004;29:1374–7.
Pandey GN. Neurobiology of adult and teenage suicide. Asian J Psychiatry 2011;4:2–13.Pitchot W, Reggers J, Pinto E, Hansenne M, Fuchs S, Pirard S, et al. Reduced dopaminergic
activity in depressed suicides. Psychoneuroendocrinology 2001;26:331–5.Pitchot W, Reggers J, Pinto E, Hansenne M, Ansseau M. Catecholamine and HPA axis
dysfunction in depression: relationship with suicidal behavior. Neuropsychobiology2003;47:152–7.
Pivac N, Pregelj P, Nikolac M, Zupanc T, Nedic G, Muck Seler D, et al. The associationbetween catechol-O-methyltransferase Val108/158Met polymorphism and suicide.Genes Brain Behav 2011;10:565–9.
Poulter MO, Du L, Weaver IC, Palkovits M, Faludi G, Merali Z, et al. GABAA receptorpromoter hypermethylation in suicide brain: implications for the involvement ofepigenetic processes. Biol Psychiatry 2008;64:645–52.
Poulter MO, Du L, Zhurov V, Palkovits M, Faludi G, Merali Z, et al. Altered organization ofGABAA receptor mRNA expression in the depressed suicide brain. Front Mol Neurosci2010;3:3.
Roy A. Gene–environment interaction and suicidal behavior. In: Dwivedi Y, editor.The neurobiological basis of suicide. Boca Raton (FL): CRC Press; 2012. [Chapter 10.Available from: http://www.ncbi.nlm.nih.gov/books/NBK107212/].
Ryding E, LindströmM, Träskman-Bendz L. The role of dopamine and serotonin in suicidalbehaviour and aggression. Prog Brain Res 2008;172:307–15.
Schosser A, Calati R, Serretti A, Massat I, Kocabas NA, Papageorgiou K, et al. The impact ofCOMT gene polymorphisms on suicidality in treatment resistant major depressivedisorder—a European multicenter study. Eur Neuropsychopharmacol 2012;22:259–66.
Shehzad Z, DeYoung CG, Kang Y, Grigorenko EL, Gray JR. Interaction of COMT val158metand externalizing behavior: relation to prefrontal brain activity and behavioralperformance. Neuroimage 2012;60:2158–68.
Sher L, Mann JJ, Traskman-Bendz L, Winchel R, Huang YY, Fertuck E, et al. Lower cerebro-spinal fluid homovanillic acid levels in depressed suicide attempters. J Affect Disord2006;90:83–9.
Snyder SH. Serotonin, cytokines, p11, and depression. Proc Natl Acad Sci U S A 2011;108:8923–4.
Supriyanto I, Sasada T, Fukutake M, Asano M, Ueno Y, Nagasaki Y, et al. Association ofFKBP5 gene haplotypes with completed suicide in the Japanese population. ProgNeuropsychopharmacol Biol Psychiatry 2011;35:252–6.
Tenhunen J, Salminen M, Lundstrom K, Kiviluoto T, Savolainen R, Ulmanen I. Genomicorganization of the human catechol O-methyltransferase gene and its expressionfrom two distinct promoters. Eur J Biochem 1994;223:1049–59.
Tovilla-Zarate C, Juarez-Rojop I, Ramon-Frias T, Villar-Soto M, Pool-Garcia S, Medellín BC,et al. No association between COMT val158met polymorphism and suicidal behavior:meta-analysis and new data. BMC Psychiatry 2011;11:151–9.
Tsai SJ, Hong CJ, Liou YJ. Recent molecular genetic studies and methodological issues insuicide research. Prog Neuropsychopharmacol Biol Psychiatry 2011;35:809–17.
Tunbridge EM. The catechol-O-methyltransferase gene: its regulation and polymorphisms.Int Rev Neurobiol 2010;95:7–27.
Tunbridge E, Burnet PW, Sodhi MS, Harrison PJ. Catechol-o-methyltransferase (COMT)and proline dehydrogenase (PRODH) mRNAs in the dorsolateral prefrontal cortexof schizophrenia, bipolar disorder and major depression. Synapse 2004;51:112–8.
Tunbridge EM, Harrison PJ, Weinberger DR. Catechol-O-methyltransferase, cognition, andpsychosis: Val158Met and beyond. Biol Psychiatry 2006;60:141–51.
Turner CA, Watson SJ, Akil H. The fibroblast growth factor family: neuromodulation ofaffective behavior. Neuron 2012;76:160–74.
Ursini G, Bollati V, Fazio L, Porcelli A, Iacovelli L, Catalani A, et al. Stress-related methylationof the catechol-O-methyltransferase Val 158 allele predicts human prefrontal cognitionand activity. J Neurosci 2011;31:6692–8.
WHO. Suicide prevention. www.who.int/mentalhealth/prevention/suicide/suicidepreven,2012.
Witte V, Flöel A. Effects of COMT polymorphisms on brain function and behavior in healthand disease. Brain Res Bull 2012;88:418–28.
Zhu G, Lipsky RH, Xu K, Ali S, Hyde T, Kleinman J, et al. Differential expression of humanCOMT alleles in brain and lymphoblasts detected by RT-coupled 5′ nuclease assay.Psychopharmacology (Berl) 2004;177:178–84.
Related Documents











