Dual role of the MgtC virulence factor in host and non-host environments Cécile Rang, 1 Eric Alix, 1 Christine Felix, 1 Annie Heitz, 2 Lena Tasse 1 and Anne-Béatrice Blanc-Potard 1 * 1 INSERM U431, Avenir Team, Faculté de Médecine, Avenue J. F. Kennedy, 30908 Nîmes Cedex 02, France. 2 Centre de Biochimie Structurale, INSERM U554 – CNRS UMR5048, Université Montpellier I, 15 Avenue, Charles Flahault, 34060 Montpellier Cedex, France. Summary MgtC is required for intramacrophage replication of intracellular pathogens and growth in low Mg 2+ medium. A link between these two phenotypes has been proposed due to putative Mg 2+ deprivation inside phagosome. MgtC is part of a family of pro- teins that share a conserved N-terminal transmem- brane domain and a variable C-terminal domain. A combination of predictive and experimental approaches indicates that the Salmonella MgtC C-terminal domain is cytoplasmic, adopts a fold also found in metal transporters and RNA interacting domain, and does not bind Mg 2+ . MgtC homologues from diverse g-proteobacteria, including the extracel- lular pathogens Yersinia pestis, Photorhabdus lumi- nescens and Pseudomonas aeruginosa, have been expressed in a Salmonella DmgtC strain. The Y. pestis MgtC fully replaced the Salmonella MgtC whereas P. luminescens or P. aeruginosa MgtC complemented only in low Mg 2+ medium, thus disso- ciating for the first time the two MgtC-related phe- notypes. In addition, we identified single amino acids changes that prevent or promote MgtC role in macrophages without affecting MgtC role in low Mg 2+ culture. A Salmonella DmgtC strain showed elongated and autoaggregated bacteria in low Mg 2+ medium but not in macrophages. Taken together our results suggest that MgtC has a dual role when bacteria localize in macrophages or low Mg 2+ environment. Introduction The success of intracellular pathogens requires a precise adaptive response to conditions encountered in the host milieu. MgtC is a virulence factor of intracellular patho- gens that is involved in adaptation to low Mg 2+ environ- ment (Blanc-Potard and Lafay, 2003). MgtC was first described in Salmonella enterica serovar Typhimurium (S. Typhimurium) where it is required for intramacrophage survival and long-term systemic infection in mice (Blanc- Potard and Groisman, 1997; Lawley et al., 2006). MgtC is also a critical factor for the intramacrophage survival of other intracellular bacterial pathogens, Mycobacterium tuberculosis, Brucella suis and Burkholderia cenocepacia (Buchmeier et al., 2000; Lavigne et al., 2005; Maloney and Valvano, 2006). MgtC has been shown to be required for growth in liquid medium for all these intracellular pathogens in a strikingly similar manner at Mg 2+ concen- tration below 50 mM (Blanc-Potard and Groisman, 1997; Buchmeier et al., 2000; Lavigne et al., 2005; Maloney and Valvano, 2006). The role of MgtC in macrophages might be related to a rate-limiting Mg 2+ concentration inside the phagosome. Mg 2+ plays a major role in biochemical func- tions, being a cofactor of numerous enzymes and partici- pating to membrane stability, and Mg 2+ depletion from phagosome could be used by the host to limit growth of intracellular pathogens. In agreement with this hypoth- esis, the macrophage survival defect of a S. Typhimurium mgtC mutant or a B. suis mgtC mutant can be partially suppressed by adding Mg 2+ to the cell culture medium (Blanc-Potard and Groisman, 1997; Lavigne et al., 2005). In S. Typhimurium, expression of mgtC is controlled by external Mg 2+ concentration through the PhoQ sensor kinase which activates the PhoP transcription factor (Garcia Vescovi et al., 1996). High level of mgtC expres- sion is observed when Mg 2+ concentration is in the micro- molar range (Garcia Vescovi et al., 1996; Tao et al., 1998). The hypothesis of a low Mg 2+ environment in pha- gosomes is supported by the fact that mgtC is highly induced in macrophages (Smith et al., 1998), being among the genes most highly induced in a microarray study (Eriksson et al., 2003). However, interpretation of these results has been reconsidered by the recent finding that cationic peptide could mimic the induction effect driven by limiting Mg 2+ and might be relevant signal to induce PhoP-regulated genes in macrophages (Bader Accepted 23 November, 2006. *For correspondence. E-mail [email protected]; Tel. (+33) 4 66 02 81 47; Fax (+33) 4 66 02 81 48. Molecular Microbiology (2007) 63(2), 605–622 doi:10.1111/j.1365-2958.2006.05542.x First published online 14 December 2006 Journal compilation © 2006 Blackwell Publishing Ltd No claim to original French government works

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Dual role of the MgtC virulence factor in host andnon-host environments
Cécile Rang,1 Eric Alix,1 Christine Felix,1
Annie Heitz,2 Lena Tasse1 andAnne-Béatrice Blanc-Potard1*1INSERM U431, Avenir Team, Faculté de Médecine,Avenue J. F. Kennedy, 30908 Nîmes Cedex 02, France.2Centre de Biochimie Structurale, INSERM U554 –CNRS UMR5048, Université Montpellier I, 15 Avenue,Charles Flahault, 34060 Montpellier Cedex, France.
Summary
MgtC is required for intramacrophage replication ofintracellular pathogens and growth in low Mg2+
medium. A link between these two phenotypes hasbeen proposed due to putative Mg2+ deprivationinside phagosome. MgtC is part of a family of pro-teins that share a conserved N-terminal transmem-brane domain and a variable C-terminal domain.A combination of predictive and experimentalapproaches indicates that the Salmonella MgtCC-terminal domain is cytoplasmic, adopts a fold alsofound in metal transporters and RNA interactingdomain, and does not bind Mg2+. MgtC homologuesfrom diverse g-proteobacteria, including the extracel-lular pathogens Yersinia pestis, Photorhabdus lumi-nescens and Pseudomonas aeruginosa, have beenexpressed in a Salmonella DmgtC strain. TheY. pestis MgtC fully replaced the Salmonella MgtCwhereas P. luminescens or P. aeruginosa MgtCcomplemented only in low Mg2+ medium, thus disso-ciating for the first time the two MgtC-related phe-notypes. In addition, we identified single aminoacids changes that prevent or promote MgtC role inmacrophages without affecting MgtC role in lowMg2+ culture. A Salmonella DmgtC strain showedelongated and autoaggregated bacteria in low Mg2+
medium but not in macrophages. Taken together ourresults suggest that MgtC has a dual role whenbacteria localize in macrophages or low Mg2+
environment.
Introduction
The success of intracellular pathogens requires a preciseadaptive response to conditions encountered in the hostmilieu. MgtC is a virulence factor of intracellular patho-gens that is involved in adaptation to low Mg2+ environ-ment (Blanc-Potard and Lafay, 2003). MgtC was firstdescribed in Salmonella enterica serovar Typhimurium (S.Typhimurium) where it is required for intramacrophagesurvival and long-term systemic infection in mice (Blanc-Potard and Groisman, 1997; Lawley et al., 2006). MgtC isalso a critical factor for the intramacrophage survival ofother intracellular bacterial pathogens, Mycobacteriumtuberculosis, Brucella suis and Burkholderia cenocepacia(Buchmeier et al., 2000; Lavigne et al., 2005; Maloneyand Valvano, 2006). MgtC has been shown to be requiredfor growth in liquid medium for all these intracellularpathogens in a strikingly similar manner at Mg2+ concen-tration below 50 mM (Blanc-Potard and Groisman, 1997;Buchmeier et al., 2000; Lavigne et al., 2005; Maloney andValvano, 2006). The role of MgtC in macrophages mightbe related to a rate-limiting Mg2+ concentration inside thephagosome. Mg2+ plays a major role in biochemical func-tions, being a cofactor of numerous enzymes and partici-pating to membrane stability, and Mg2+ depletion fromphagosome could be used by the host to limit growth ofintracellular pathogens. In agreement with this hypoth-esis, the macrophage survival defect of a S. TyphimuriummgtC mutant or a B. suis mgtC mutant can be partiallysuppressed by adding Mg2+ to the cell culture medium(Blanc-Potard and Groisman, 1997; Lavigne et al., 2005).
In S. Typhimurium, expression of mgtC is controlled byexternal Mg2+ concentration through the PhoQ sensorkinase which activates the PhoP transcription factor(Garcia Vescovi et al., 1996). High level of mgtC expres-sion is observed when Mg2+ concentration is in the micro-molar range (Garcia Vescovi et al., 1996; Tao et al.,1998). The hypothesis of a low Mg2+ environment in pha-gosomes is supported by the fact that mgtC is highlyinduced in macrophages (Smith et al., 1998), beingamong the genes most highly induced in a microarraystudy (Eriksson et al., 2003). However, interpretation ofthese results has been reconsidered by the recent findingthat cationic peptide could mimic the induction effectdriven by limiting Mg2+ and might be relevant signal toinduce PhoP-regulated genes in macrophages (Bader
Accepted 23 November, 2006. *For correspondence. [email protected]; Tel. (+33) 4 66 02 81 47; Fax(+33) 4 66 02 81 48.
Molecular Microbiology (2007) 63(2), 605–622 doi:10.1111/j.1365-2958.2006.05542.xFirst published online 14 December 2006
Journal compilation © 2006 Blackwell Publishing LtdNo claim to original French government works

et al., 2005). In addition, direct measurement of Mg2+ con-centration at early times of infection indicates a concen-tration in the millimolar range (Martin-Orozco et al., 2006).Analysis of the relationship between the role of MgtC inlow Mg2+ culture and its role in macrophages offers analternative way to address this fundamental aspect of theintracellular microbial environment.
The function of MgtC remains unknown but its role inadaptation to Mg2+ limitation suggests that MgtC has arole in Mg2+ uptake. In S. Typhimurium, mgtC is cotrans-cribed with mgtB, encoding a inner-membrane Mg2+
transporter (Snavely et al., 1991). The mgtC gene is notessential for membrane insertion or transport function ofMgtB (Tao et al., 1995) and MgtC, predicted as an inner-membrane protein, does not appear itself as a cationtransporter (Moncrief and Maguire, 1998). A recent studybased on overexpression of MgtC in Xenopus laevisoocytes further confirmed that MgtC does not directlytransport Mg2+ (Günzel et al., 2006). An alternativehypothesis is that MgtC might act by sequestering Mg2+
and/or by facilitating transport by specific transporters.MgtC does not exhibit similarity with proteins of known
function but belongs to a large bacterial protein family(Blanc-Potard and Lafay, 2003). Genes encoding MgtC-like proteins are found in a limited number of eubacterialgenomes and phylogenetic analysis suggests that mgtChas been acquired by horizontal gene transfer repeat-edly throughout bacterial evolution (Blanc-Potard andLafay, 2003). Alignment of MgtC-like proteins as well ashydrophobicity pattern clearly defines two domains: ahydrophobic N-terminal part of the protein, highly con-served in all MgtC-like proteins, and a soluble C-terminalpart of the protein that is much more variable, but spe-cifically conserved in a subgroup of proteins, includingMgtC-like proteins from intracellular pathogens (Blanc-Potard and Lafay, 2003). Cumulatively, domain organi-zation and sequence evolution of MgtC suggested thatthe C-terminal domain might have a key role in MgtCfunction.
In the present study, we have used predictive andexperimental approaches to characterize the SalmonellaMgtC C-terminal domain and we have explored the MgtCprotein family to better understand MgtC function and therelationship between adaptation to Mg2+ deprivation andintramacrophage survival. Using a complementationapproach in a Salmonella mgtC null mutant, we haveinvestigated the role of conserved residues in MgtCfunction, and we have analysed MgtC homologuesfrom diverse g-proteobacteria, including extracellularpathogens. In addition, we have investigated bacterialmorphology both in low Mg2+ liquid medium andmacrophages. Our results indicate that MgtC has a dualrole which differs when bacteria are in low Mg2+ culture orwithin macrophages.
Results
Sequence comparison and fold-recognition
PSI-BLAST searches identified numerous sequencesshowing significant similarities to MgtC from Salmonella(Blanc-Potard and Lafay, 2003). However, none has aknown structure or precisely annotated biochemical func-tion. We searched for structural similarities using fold-recognition techniques and secondary structure predictionfor both N-terminal and C-terminal domains of MgtC (http://www.infobiosud.cnrs.fr/bioserver/MGTC/suppl.html).
The N-terminal domain (residues 1–140) matcheshelical proteins embedded in membranes with no clearsequence similarity, thus preventing detection of a similartopology as well as a putative function. The C-terminaldomain (residues 140–231) showed weak but significantcompatibilities with small proteins adopting a babbabtopology, with a four-stranded b-sheet flanked on one sideby two a-helices. This fold, not associated with any par-ticular class of function, is found in Heavy-Metal Associ-ated domain (Pfam PF00403), RNA recognition motif(RRM) domain (Pfam PF00076) or the amino acid bindingACT regulatory domain (Pfam PF01842). This fold-recognition is further confirmed by the overall quality ofthe derived three-dimensional (3D) models showinghydrophobic and conserved residues mostly buried.However, despite a global structural similarity, nosequence motif appeared conserved at any face of themodels precluding any identification of a functional site.
Location of Salmonella MgtC C-terminal domain
The prediction of topology of Salmonella MgtC proteinusing five predictive programs gave controversial models.All programs predicted a periplasmic N-terminal end, butfollowed by four to five transmembrane domains in theN-terminal region. Therefore, the C-terminal domain ispredicted either in the periplasmic or the cytoplasmicspace. Experimental determination of the location of theC-terminus of MgtC was done by fusing either the PhoAprotein (periplasmic reporter) or LacZ protein (cytoplasmicreporter) at position W226 of the MgtC protein. Both fusionproteins were produced as shown by Western analysisusing MgtC antibodies (not shown). The C-terminal PhoAfusion gave low PhoA activity (average of 14 units)whereas the C-terminal LacZ fusion gave a high LacZactivity (average of 1219 Miller units), indicating that theC-terminal domain of MgtC is cytoplasmic. It has beenshown that reliable topology models of inner-membraneproteins can be achieved based on a combination oftopology prediction and experimental determination of thelocation of the protein’s C-terminus (Drew et al., 2002;Rapp et al., 2004). Hence, combination of our results withtopological predictions suggests that the N-terminal
606 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
domain has five transmembrane segments. This model isfurther supported by two additional PhoA fusions in pre-dicted periplasmic loops at position G63 and Q111 thatgave high PhoA activity (average of 862 units and522 units respectively).
Nuclear magnetic resonance (NMR) spectroscopyanalysis of the soluble Salmonella MgtC C-terminaldomain
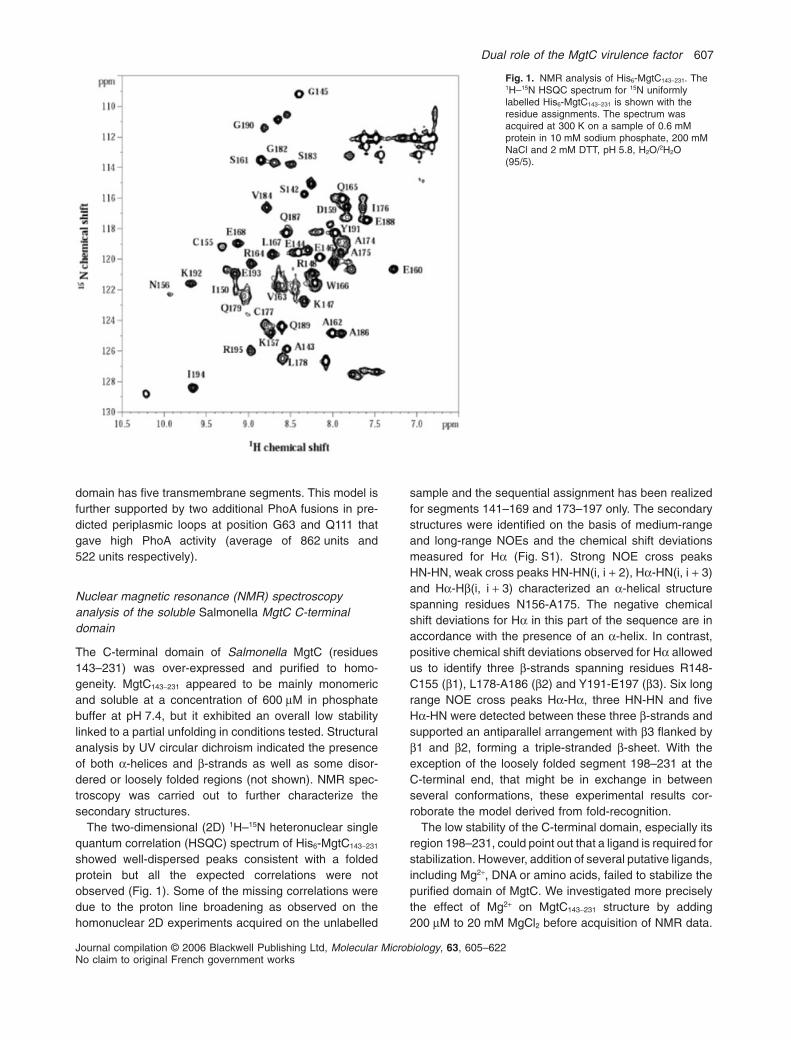
The C-terminal domain of Salmonella MgtC (residues143–231) was over-expressed and purified to homo-geneity. MgtC143-231 appeared to be mainly monomericand soluble at a concentration of 600 mM in phosphatebuffer at pH 7.4, but it exhibited an overall low stabilitylinked to a partial unfolding in conditions tested. Structuralanalysis by UV circular dichroism indicated the presenceof both a-helices and b-strands as well as some disor-dered or loosely folded regions (not shown). NMR spec-troscopy was carried out to further characterize thesecondary structures.
The two-dimensional (2D) 1H–15N heteronuclear singlequantum correlation (HSQC) spectrum of His6-MgtC143-231
showed well-dispersed peaks consistent with a foldedprotein but all the expected correlations were notobserved (Fig. 1). Some of the missing correlations weredue to the proton line broadening as observed on thehomonuclear 2D experiments acquired on the unlabelled
sample and the sequential assignment has been realizedfor segments 141–169 and 173–197 only. The secondarystructures were identified on the basis of medium-rangeand long-range NOEs and the chemical shift deviationsmeasured for Ha (Fig. S1). Strong NOE cross peaksHN-HN, weak cross peaks HN-HN(i, i + 2), Ha-HN(i, i + 3)and Ha-Hb(i, i + 3) characterized an a-helical structurespanning residues N156-A175. The negative chemicalshift deviations for Ha in this part of the sequence are inaccordance with the presence of an a-helix. In contrast,positive chemical shift deviations observed for Ha allowedus to identify three b-strands spanning residues R148-C155 (b1), L178-A186 (b2) and Y191-E197 (b3). Six longrange NOE cross peaks Ha-Ha, three HN-HN and fiveHa-HN were detected between these three b-strands andsupported an antiparallel arrangement with b3 flanked byb1 and b2, forming a triple-stranded b-sheet. With theexception of the loosely folded segment 198–231 at theC-terminal end, that might be in exchange in betweenseveral conformations, these experimental results cor-roborate the model derived from fold-recognition.
The low stability of the C-terminal domain, especially itsregion 198–231, could point out that a ligand is required forstabilization. However, addition of several putative ligands,including Mg2+, DNA or amino acids, failed to stabilize thepurified domain of MgtC. We investigated more preciselythe effect of Mg2+ on MgtC143-231 structure by adding200 mM to 20 mM MgCl2 before acquisition of NMR data.
Fig. 1. NMR analysis of His6-MgtC143-231. The1H–15N HSQC spectrum for 15N uniformlylabelled His6-MgtC143-231 is shown with theresidue assignments. The spectrum wasacquired at 300 K on a sample of 0.6 mMprotein in 10 mM sodium phosphate, 200 mMNaCl and 2 mM DTT, pH 5.8, H2O/2H2O(95/5).
Dual role of the MgtC virulence factor 607
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

Modulation of Mg2+ concentration in the sample yield indis-tinguishable 2D 1H–15N HSQC spectra indicating that thesolution-state conformation is identical in the presence orabsence of Mg2+ (not shown). This result indicate that theisolated C-terminal domain of MgtC does not bind Mg2+.
Design of a complementation approach to test MgtCfunction
To conduct functional analysis on the entire MgtC proteinand to better understand the relationship between low Mg2+
adaptation and intramacrophage survival, we have usedan approach based on the complementation of a Salmo-nella mgtC null mutant. Previous studies on S. Typhimu-rium mgtC mutants have been carried out with polarinsertional mutants that lack both mgtC and mgtB (Blanc-Potard and Groisman, 1997; Moncrief and Maguire, 1998).We have constructed a non-polar DmgtC mutant (seeExperimental procedures). This mgtC null strain, NM14,exhibit similar phenotypes as those reported for previousmutants, i.e. is similarly defective for growth in low Mg2+
liquid medium (Fig. S2) and for replication in J774 mac-rophages (not shown). In addition, the growth of NM14 isidentical to the one of the wild-type strain in high Mg2+ liquidmedium (Fig. S2). NM14 has been used in complementa-tion experiments to address the function of SalmonellaMgtC mutated proteins and MgtC homologues from otherg-proteobacteria. All MgtC proteins were similarlyexpressed from a low copy-number plasmid under the S.Typhimurium regulatory sequences (pNM11 vector, seeExperimental procedures), and are therefore induced inlow Mg2+ liquid medium and macrophages.
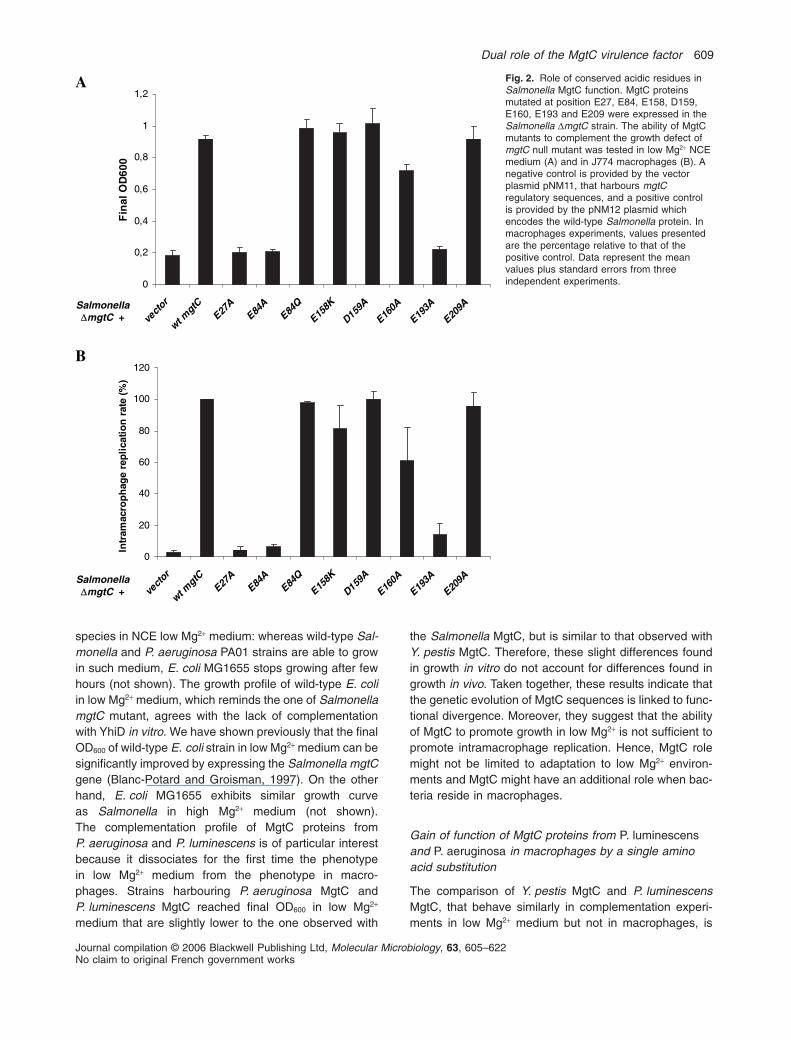
Role of acidic residues in Salmonella MgtC function
The hypothesis of MgtC being a Mg2+ binding protein wasfurther tested by a mutational analysis of acidic residues.Negatively charged acidic residues have been implicatedin Mg2+ binding by proteins that use Mg2+ as a ligand or acofactor and an acidic cluster contributes to Mg2+ sensingby the PhoQ protein (Chamnongpol et al., 2003). We havecarried out site-directed mutagenesis on five acidic resi-dues conserved among MgtC-like proteins, and two resi-dues of an ‘EDE’ cluster (Fig. S3). We have analysedwhether expression of mutated proteins complementedthe DmgtC strain NM14 for growth in low Mg2+ medium.We identified three mutants (E27A, E84A, E193A) thatfailed to complement NM14 in low Mg2+ medium, and onemutant (E160A) that grows slightly less than wild-type,whereas other mutants complemented as well as the wild-type protein (Fig. 2A). All complemented strains grewsimilarly to wild-type strain in high Mg2+ medium (notshown). As shown in Fig. 2B, the macrophage replicationrate correlated with the growth in low Mg2+ medium.Western blot analysis using anti-MgtC antibodies failed to
detect MgtC proteins with E27A and E193A substitutions,suggesting a main structural role for these residues, butE84A mutation retained protein expression (data notshown). However, a protein carrying a mutation E84Qbehaves similarly to the wild-type protein (Fig. 2A), sug-gesting that the negative charge of E84 is not importantfor correct MgtC function. Hence, no putative Mg2+ bindingsite was identified among the acidic residues tested. Wecannot exclude a role of other acidic residues that werenot included in the mutational analysis.
Identification of MgtC homologues that complement theSalmonella DmgtC mutant in low Mg 2+ medium but notin macrophages
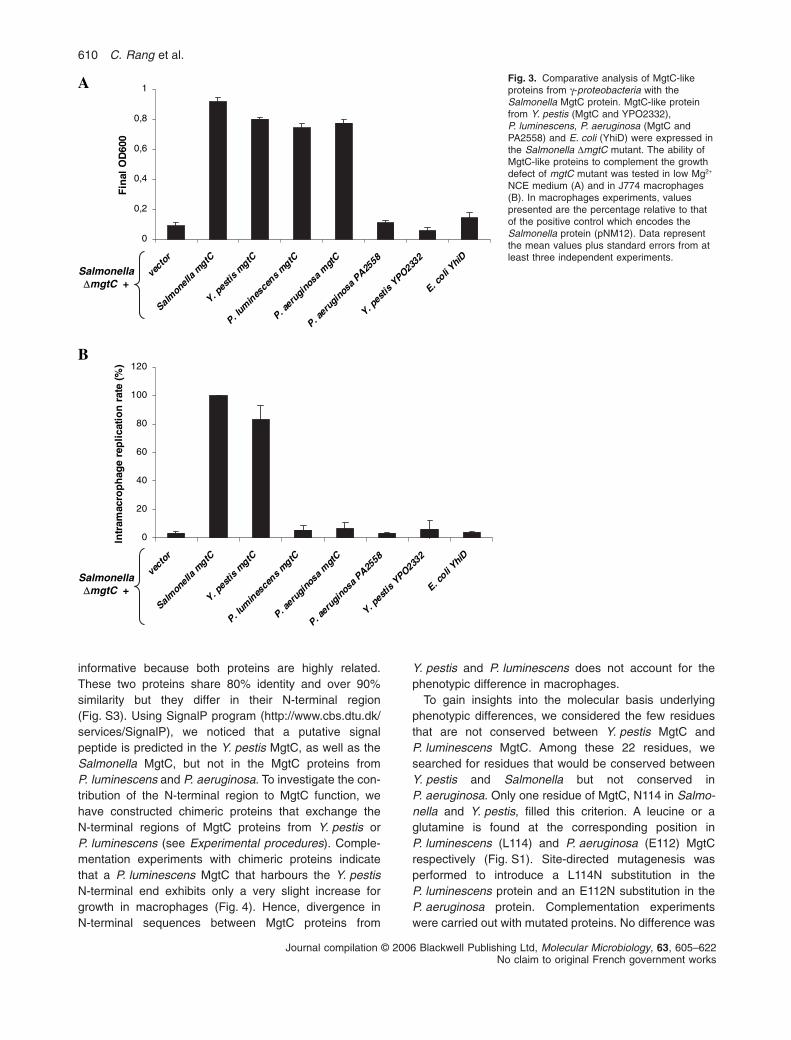
The phylogenetic analysis of ‘MgtC-like’ proteins identifieda cluster of proteins that exhibit conserved residues in theC-terminal domain (Blanc-Potard and Lafay, 2003). Todetermine whether the phylogenetic clustering of MgtCsequences was linked to functional evolution, we havestudied MgtC-like proteins from diverse g-proteobacteria(Fig. S3). MgtC homologues from Photorhabdus lumine-scens and Pseudomonas aeruginosa are closely relatedto the Salmonella protein and belong to the same phylo-genetic group. P. aeruginosa encodes two MgtC-like pro-teins, PA4635, referred as P. aeruginosa MgtC, andPA2558. In contrast, the MgtC-like protein of Escherichiacoli (YhiD) is distantly related from the one of Salmonellaand does not belong to the phylogenetic cluster. Yersiniapestis is of particular interest because it harbours twoMgtC-like proteins, one related to the Salmonella protein(YPO1660, referred as Y. pestis MgtC) and one related tothe E. coli protein (YPO2332).
The diverse mgtC gene homologues were expressed inthe Salmonella mgtC null mutant strain NM14 and theirability to complement the growth defect in low Mg2+
medium (Fig. 3A) and in J774 macrophages (Fig. 3B) wasanalysed. Using the same heterologous expressionsystem, we have previously reported that expression ofMgtC protein from M. tuberculosis partially complementeda Salmonella mgtC mutant for both phenotypes (Alixet al., 2006). Results clearly show three phenotypicclasses: (i) Y. pestis MgtC complements both phenotypes;(ii) P. aeruginosa MgtC and P. luminescens MgtC comple-ment the phenotype in low Mg2+ medium but not in mac-rophages; (iii) P. aeruginosa PA2558, Y. pestis YPO2332and E. coli YhiD do not complement any phenotype. Allstrains grew similarly to wild-type strain in high Mg2+
medium (not shown). These results are partially consis-tent with the phylogeny, because the Y. pestis MgtC isclosely related to the Salmonella MgtC protein whereasY. pestis YPO2332, E. coli YhiD and, to a lesser extendP. aeruginosa PA2558, are only distantly related. Theseresults are also consistent with the growth of bacterial
608 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
species in NCE low Mg2+ medium: whereas wild-type Sal-monella and P. aeruginosa PA01 strains are able to growin such medium, E. coli MG1655 stops growing after fewhours (not shown). The growth profile of wild-type E. coliin low Mg2+ medium, which reminds the one of SalmonellamgtC mutant, agrees with the lack of complementationwith YhiD in vitro. We have shown previously that the finalOD600 of wild-type E. coli strain in low Mg2+ medium can besignificantly improved by expressing the Salmonella mgtCgene (Blanc-Potard and Groisman, 1997). On the otherhand, E. coli MG1655 exhibits similar growth curveas Salmonella in high Mg2+ medium (not shown).The complementation profile of MgtC proteins fromP. aeruginosa and P. luminescens is of particular interestbecause it dissociates for the first time the phenotypein low Mg2+ medium from the phenotype in macro-phages. Strains harbouring P. aeruginosa MgtC andP. luminescens MgtC reached final OD600 in low Mg2+
medium that are slightly lower to the one observed with
the Salmonella MgtC, but is similar to that observed withY. pestis MgtC. Therefore, these slight differences foundin growth in vitro do not account for differences found ingrowth in vivo. Taken together, these results indicate thatthe genetic evolution of MgtC sequences is linked to func-tional divergence. Moreover, they suggest that the abilityof MgtC to promote growth in low Mg2+ is not sufficient topromote intramacrophage replication. Hence, MgtC rolemight not be limited to adaptation to low Mg2+ environ-ments and MgtC might have an additional role when bac-teria reside in macrophages.
Gain of function of MgtC proteins from P. luminescensand P. aeruginosa in macrophages by a single aminoacid substitution
The comparison of Y. pestis MgtC and P. luminescensMgtC, that behave similarly in complementation experi-ments in low Mg2+ medium but not in macrophages, is
Fig. 2. Role of conserved acidic residues inSalmonella MgtC function. MgtC proteinsmutated at position E27, E84, E158, D159,E160, E193 and E209 were expressed in theSalmonella DmgtC strain. The ability of MgtCmutants to complement the growth defect ofmgtC null mutant was tested in low Mg2+ NCEmedium (A) and in J774 macrophages (B). Anegative control is provided by the vectorplasmid pNM11, that harbours mgtCregulatory sequences, and a positive controlis provided by the pNM12 plasmid whichencodes the wild-type Salmonella protein. Inmacrophages experiments, values presentedare the percentage relative to that of thepositive control. Data represent the meanvalues plus standard errors from threeindependent experiments.0
0,2
0,4
0,6
0,8
1
1,2
evtcor
wt
Ctgm
A72E
A48E
Q48E
K851E
A951D
A061E
A391E
A902E
00
6D
Ol
ani
F
0
20
40
60
80
100
120
rotcev
tgmt
w
C
E
A72E
A48E
Q48K851
E D1A95
E
A061 E
A391 E
A902
)%(
etar
noit
acil
per
eg
ah
por
ca
mart
nI
A
B
SalmonellaΔmgtC +
Salmonella
ΔmgtC +
Dual role of the MgtC virulence factor 609
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
informative because both proteins are highly related.These two proteins share 80% identity and over 90%similarity but they differ in their N-terminal region(Fig. S3). Using SignalP program (http://www.cbs.dtu.dk/services/SignalP), we noticed that a putative signalpeptide is predicted in the Y. pestis MgtC, as well as theSalmonella MgtC, but not in the MgtC proteins fromP. luminescens and P. aeruginosa. To investigate the con-tribution of the N-terminal region to MgtC function, wehave constructed chimeric proteins that exchange theN-terminal regions of MgtC proteins from Y. pestis orP. luminescens (see Experimental procedures). Comple-mentation experiments with chimeric proteins indicatethat a P. luminescens MgtC that harbours the Y. pestisN-terminal end exhibits only a very slight increase forgrowth in macrophages (Fig. 4). Hence, divergence inN-terminal sequences between MgtC proteins from
Y. pestis and P. luminescens does not account for thephenotypic difference in macrophages.
To gain insights into the molecular basis underlyingphenotypic differences, we considered the few residuesthat are not conserved between Y. pestis MgtC andP. luminescens MgtC. Among these 22 residues, wesearched for residues that would be conserved betweenY. pestis and Salmonella but not conserved inP. aeruginosa. Only one residue of MgtC, N114 in Salmo-nella and Y. pestis, filled this criterion. A leucine or aglutamine is found at the corresponding position inP. luminescens (L114) and P. aeruginosa (E112) MgtCrespectively (Fig. S1). Site-directed mutagenesis wasperformed to introduce a L114N substitution in theP. luminescens protein and an E112N substitution in theP. aeruginosa protein. Complementation experimentswere carried out with mutated proteins. No difference was
Fig. 3. Comparative analysis of MgtC-likeproteins from g-proteobacteria with theSalmonella MgtC protein. MgtC-like proteinfrom Y. pestis (MgtC and YPO2332),P. luminescens, P. aeruginosa (MgtC andPA2558) and E. coli (YhiD) were expressed inthe Salmonella DmgtC mutant. The ability ofMgtC-like proteins to complement the growthdefect of mgtC mutant was tested in low Mg2+
NCE medium (A) and in J774 macrophages(B). In macrophages experiments, valuespresented are the percentage relative to thatof the positive control which encodes theSalmonella protein (pNM12). Data representthe mean values plus standard errors from atleast three independent experiments.
0
0,2
0,4
0,6
0,8
1
rotcev
Smla
leno
alm
Ctg
Y
ep.
sit s
mgtC
.Pl
mu
nie
ecsns
gm
tC Ctgm
asonigurea.PP. a
igure
on
asP
8552A
Yp.
seit s
YP2
O3
23
.Eco
ilY
ihD
00
6D
Ol
ani
F
0
20
40
60
80
100
120
rotcev llenomlaS
ag
mtC Ctg
msitsep.Y
.Pl
mu
nie
ecsns
gm
tC
P.
igurea
n
aso
mtgC
a.P
igure
Pason
A
8552
Y
itsep.
sPYO
2332
E
oc.
l
ihYi
D
)%(
etar
noit
acil
per
eg
ah
por
ca
mar t
nI
Salmonella
ΔmgtC +
A
B
Salmonella
ΔmgtC +
610 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
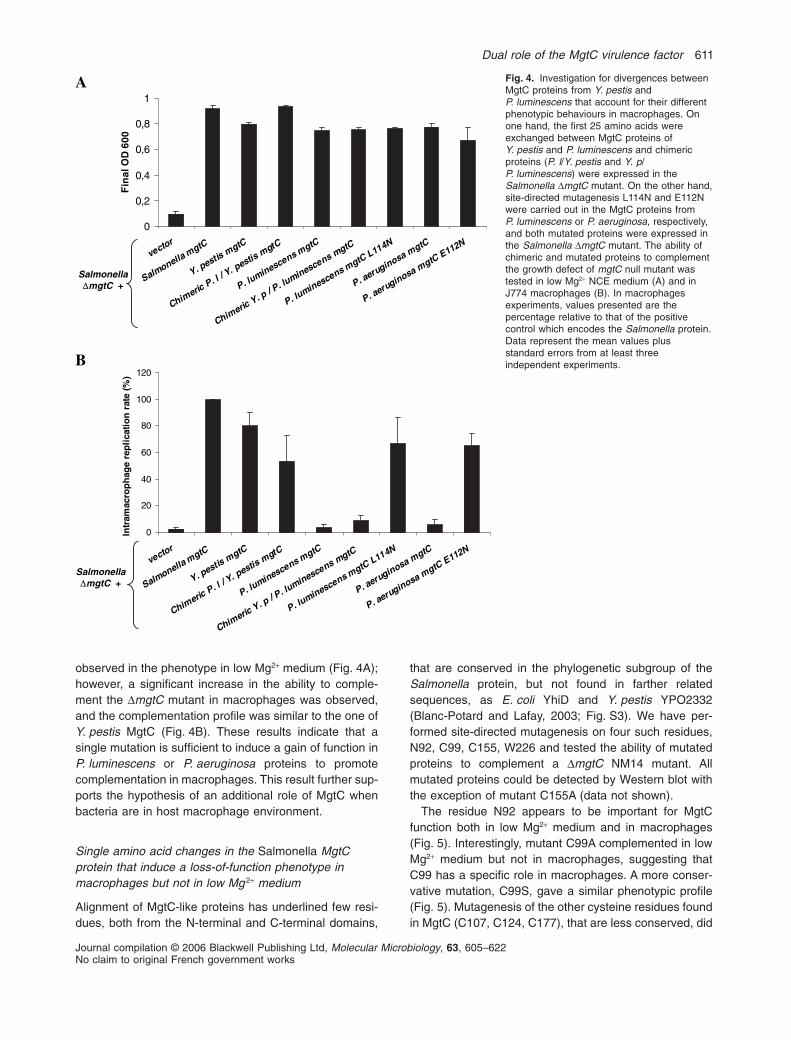
observed in the phenotype in low Mg2+ medium (Fig. 4A);however, a significant increase in the ability to comple-ment the DmgtC mutant in macrophages was observed,and the complementation profile was similar to the one ofY. pestis MgtC (Fig. 4B). These results indicate that asingle mutation is sufficient to induce a gain of function inP. luminescens or P. aeruginosa proteins to promotecomplementation in macrophages. This result further sup-ports the hypothesis of an additional role of MgtC whenbacteria are in host macrophage environment.
Single amino acid changes in the Salmonella MgtCprotein that induce a loss-of-function phenotype inmacrophages but not in low Mg 2+ medium
Alignment of MgtC-like proteins has underlined few resi-dues, both from the N-terminal and C-terminal domains,
that are conserved in the phylogenetic subgroup of theSalmonella protein, but not found in farther relatedsequences, as E. coli YhiD and Y. pestis YPO2332(Blanc-Potard and Lafay, 2003; Fig. S3). We have per-formed site-directed mutagenesis on four such residues,N92, C99, C155, W226 and tested the ability of mutatedproteins to complement a DmgtC NM14 mutant. Allmutated proteins could be detected by Western blot withthe exception of mutant C155A (data not shown).
The residue N92 appears to be important for MgtCfunction both in low Mg2+ medium and in macrophages(Fig. 5). Interestingly, mutant C99A complemented in lowMg2+ medium but not in macrophages, suggesting thatC99 has a specific role in macrophages. A more conser-vative mutation, C99S, gave a similar phenotypic profile(Fig. 5). Mutagenesis of the other cysteine residues foundin MgtC (C107, C124, C177), that are less conserved, did
Fig. 4. Investigation for divergences betweenMgtC proteins from Y. pestis andP. luminescens that account for their differentphenotypic behaviours in macrophages. Onone hand, the first 25 amino acids wereexchanged between MgtC proteins ofY. pestis and P. luminescens and chimericproteins (P. l/Y. pestis and Y. p/P. luminescens) were expressed in theSalmonella DmgtC mutant. On the other hand,site-directed mutagenesis L114N and E112Nwere carried out in the MgtC proteins fromP. luminescens or P. aeruginosa, respectively,and both mutated proteins were expressed inthe Salmonella DmgtC mutant. The ability ofchimeric and mutated proteins to complementthe growth defect of mgtC null mutant wastested in low Mg2+ NCE medium (A) and inJ774 macrophages (B). In macrophagesexperiments, values presented are thepercentage relative to that of the positivecontrol which encodes the Salmonella protein.Data represent the mean values plusstandard errors from at least threeindependent experiments.
0
0,2
0,4
0,6
0,8
1
vector
Salmonella
mgtC
Y. pestismgtC
Chimeric
P. l / Y. pestismgtC
P. luminescens mgtC
Chimeric
Y. p / P. luminescens mgtC
P. luminescens mgtCL114N
P. aeruginosa mgtC
P. aeruginosa mgtC
E112N
00
6D
Ol
ani
F
0
20
40
60
80
100
120
vector
Salmonella
mgtC
Y. pestisgmtC
Chiemric
P. l / Y. pestismgtC
P. luminescens mgtC
Chimeric
Y. p / P. luminescens mgtC
P. luminescens mgtCL114N
P. aeurginosa mgtC
P. aeruginosa mgtC
E112N
)%(
etar
noit
acil
per
eg
ah
por
ca
mart
nI
B
A
Salmonella
ΔmgtC +
Salmonella
ΔmgtC +
Dual role of the MgtC virulence factor 611
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
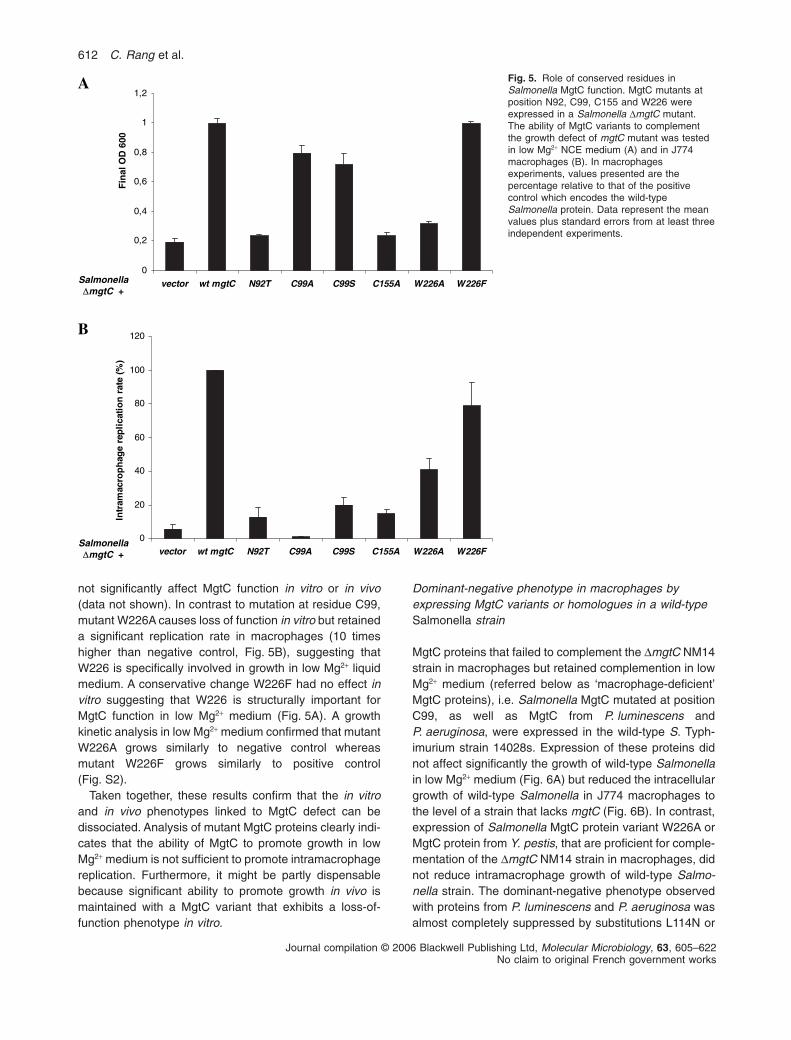
not significantly affect MgtC function in vitro or in vivo(data not shown). In contrast to mutation at residue C99,mutant W226A causes loss of function in vitro but retaineda significant replication rate in macrophages (10 timeshigher than negative control, Fig. 5B), suggesting thatW226 is specifically involved in growth in low Mg2+ liquidmedium. A conservative change W226F had no effect invitro suggesting that W226 is structurally important forMgtC function in low Mg2+ medium (Fig. 5A). A growthkinetic analysis in low Mg2+ medium confirmed that mutantW226A grows similarly to negative control whereasmutant W226F grows similarly to positive control(Fig. S2).
Taken together, these results confirm that the in vitroand in vivo phenotypes linked to MgtC defect can bedissociated. Analysis of mutant MgtC proteins clearly indi-cates that the ability of MgtC to promote growth in lowMg2+ medium is not sufficient to promote intramacrophagereplication. Furthermore, it might be partly dispensablebecause significant ability to promote growth in vivo ismaintained with a MgtC variant that exhibits a loss-of-function phenotype in vitro.
Dominant-negative phenotype in macrophages byexpressing MgtC variants or homologues in a wild-typeSalmonella strain
MgtC proteins that failed to complement the DmgtC NM14strain in macrophages but retained complemention in lowMg2+ medium (referred below as ‘macrophage-deficient’MgtC proteins), i.e. Salmonella MgtC mutated at positionC99, as well as MgtC from P. luminescens andP. aeruginosa, were expressed in the wild-type S. Typh-imurium strain 14028s. Expression of these proteins didnot affect significantly the growth of wild-type Salmonellain low Mg2+ medium (Fig. 6A) but reduced the intracellulargrowth of wild-type Salmonella in J774 macrophages tothe level of a strain that lacks mgtC (Fig. 6B). In contrast,expression of Salmonella MgtC protein variant W226A orMgtC protein from Y. pestis, that are proficient for comple-mentation of the DmgtC NM14 strain in macrophages, didnot reduce intramacrophage growth of wild-type Salmo-nella strain. The dominant-negative phenotype observedwith proteins from P. luminescens and P. aeruginosa wasalmost completely suppressed by substitutions L114N or
Fig. 5. Role of conserved residues inSalmonella MgtC function. MgtC mutants atposition N92, C99, C155 and W226 wereexpressed in a Salmonella DmgtC mutant.The ability of MgtC variants to complementthe growth defect of mgtC mutant was testedin low Mg2+ NCE medium (A) and in J774macrophages (B). In macrophagesexperiments, values presented are thepercentage relative to that of the positivecontrol which encodes the wild-typeSalmonella protein. Data represent the meanvalues plus standard errors from at least threeindependent experiments.
0
0,2
0,4
0,6
0,8
1
1,2
vector wt mgtC N92T C99A C99S C155A W226A W226F
00
6D
Ol
ani
F
0
20
40
60
80
100
120
vector wt mgtC N92T C99A C99S C155A W226A W226F
)%(
etar
noit
ac il
pe r
eg
ah
por
ca
mart
nI
Salmonella
ΔmgtC +
B
A
Salmonella
ΔmgtC +
612 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

E114N respectively (Fig. 6B). The inhibitory properties of‘macrophage-deficient’ MgtC proteins in wild-type Salmo-nella suggest formation of non-functional MgtC com-plexes or competition for a functional interaction withanother protein. Inhibitory effects are probably exacer-bated by the fact that ‘macrophage-deficient’ MgtC pro-teins are encoded by multicopy genes whereas theendogenous MgtC is encoded by a single gene copy.
Bacterial morphology by fluorescent microscopy in lowMg 2+ medium and macrophages
Overnight shaken cultures of Salmonella DmgtC mutantgrown in low Mg2+ medium showed very little turbidity andmost bacteria are found as a ring at the air–liquid interfaceas well as aggregates at the bottom of tubes. Such aggre-
gative phenotype was not observed with wild-type strainor when the DmgtC mutant is grown in high Mg2+ medium.The growth curve of the DmgtC mutant in low Mg2+
medium shows a growth rate similar to the one of wild-type strain for the first 4–6 h followed be a growth arrestand a slight decrease of OD600 (Blanc-Potard and Grois-man, 1997; Fig. S2). To evaluate the bacterial morphologyassociated to clumping we have used strains expressingconstitutively the fluorescent DsRed protein (see Experi-mental procedures). Morphology analysis indicates thatDmgtC mutant, but not wild-type strain, exhibits cell elon-gation as well as autoaggregation in low Mg2+ medium(Fig. 7). In addition, heterogeneity of DsRed distributionas well as low fluorescence of some bacteria at 18 hprobably denotes bacterial death. Filamentation and selfassociation of the DmgtC strain, detected 6–8 h after dilu-
Fig. 6. Dominant-negative phenotype inmacrophages due to overexpression of‘macrophage-deficient’ MgtC proteins in awild-type Salmonella strain. Salmonella MgtCmutants C99A, C99S and W226A, as well asMgtC proteins from P. luminescens,P. aeruginosa and Y. pestis were expressed ina Salmonella wild-type strain. The growth ofdifferent strains was tested in low Mg2+ NCEmedium (A) and in J774 macrophages (B). Apositive control is provided by the pNM12plasmid which encodes the wild-typeSalmonella protein. In macrophagesexperiments, values presented are thepercentage relative to that of the positivecontrol. Data represent the mean values plusstandard errors from at least threeindependent experiments.
0
0,2
0,4
0,6
0,8
1
1,2
wt C99A C99S W226A wt L114N wt E112N wt
00
6D
Ol
ani
F
0
20
40
60
80
100
120
wt C99A C99S W226A wt L114N wt E112N wt
)%(
etar
noit
ac il
per
eg
ah
por
ca
mart
n I
A
B
Salmonella mgtC P. luminescens mgtC
Y. pestis mgtC
Salmonella mgtC P. luminescens mgtC
Y. pestis mgtC
wtSalmonella +
P. aeruginosa mgtC
P. aeruginosa mgtC
wtSalmonella +
Dual role of the MgtC virulence factor 613
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
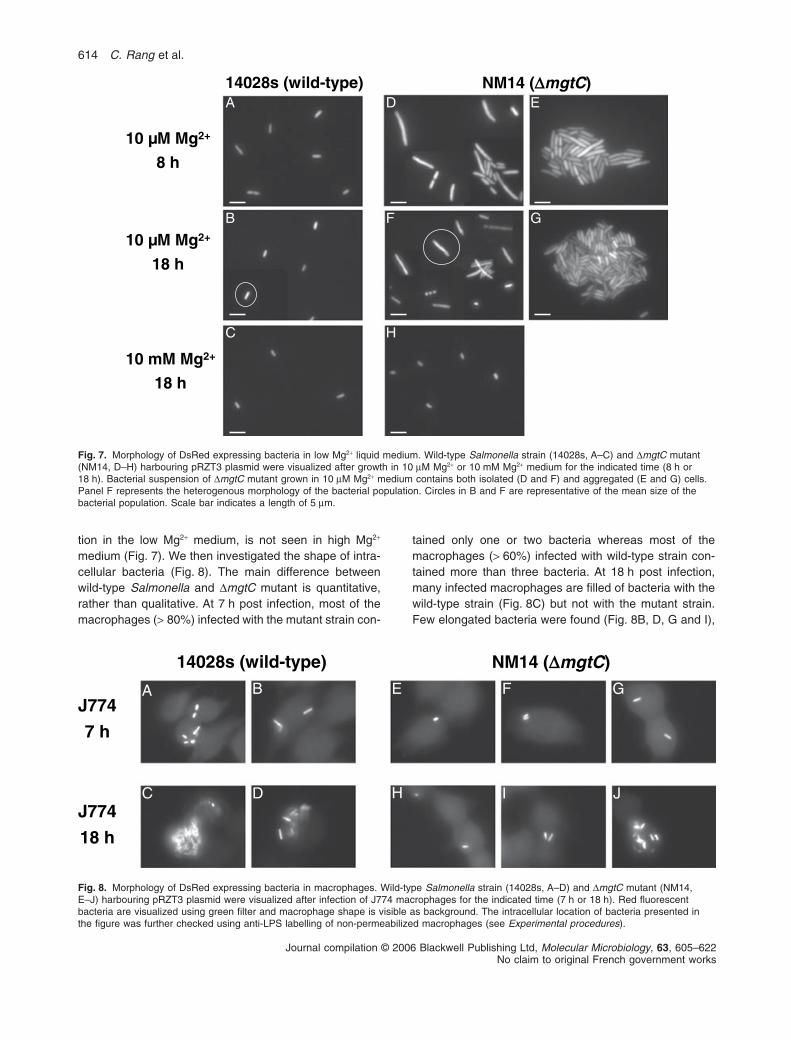
tion in the low Mg2+ medium, is not seen in high Mg2+
medium (Fig. 7). We then investigated the shape of intra-cellular bacteria (Fig. 8). The main difference betweenwild-type Salmonella and DmgtC mutant is quantitative,rather than qualitative. At 7 h post infection, most of themacrophages (> 80%) infected with the mutant strain con-
tained only one or two bacteria whereas most of themacrophages (> 60%) infected with wild-type strain con-tained more than three bacteria. At 18 h post infection,many infected macrophages are filled of bacteria with thewild-type strain (Fig. 8C) but not with the mutant strain.Few elongated bacteria were found (Fig. 8B, D, G and I),
)epyt-dliw(s82041 (41MN D Ctgm )
gMMµ01 +2
h8
gMMµ01 +2
h81
gMMm01 +2
h81
A
B
C
E
G
H
D
F
Fig. 7. Morphology of DsRed expressing bacteria in low Mg2+ liquid medium. Wild-type Salmonella strain (14028s, A–C) and DmgtC mutant(NM14, D–H) harbouring pRZT3 plasmid were visualized after growth in 10 mM Mg2+ or 10 mM Mg2+ medium for the indicated time (8 h or18 h). Bacterial suspension of DmgtC mutant grown in 10 mM Mg2+ medium contains both isolated (D and F) and aggregated (E and G) cells.Panel F represents the heterogenous morphology of the bacterial population. Circles in B and F are representative of the mean size of thebacterial population. Scale bar indicates a length of 5 mm.
)epyt-dliw(s82041 (41MN D Ctgm )
477J
h7
477J
h81
A
C
FB
JD
GE
H I
Fig. 8. Morphology of DsRed expressing bacteria in macrophages. Wild-type Salmonella strain (14028s, A–D) and DmgtC mutant (NM14,E–J) harbouring pRZT3 plasmid were visualized after infection of J774 macrophages for the indicated time (7 h or 18 h). Red fluorescentbacteria are visualized using green filter and macrophage shape is visible as background. The intracellular location of bacteria presented inthe figure was further checked using anti-LPS labelling of non-permeabilized macrophages (see Experimental procedures).
614 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

but they were not significantly longer nor more frequent inDmgtC mutant. When bacterial replication takes place inmacrophages infected with DmgtC mutant (Fig. 8J), bac-teria do not exhibit elongated or aggregated phenotype.Hence, the intracellular morphology of the DmgtC straindoes not seem to correlate with the bacterial shapeobserved in low Mg2+ liquid medium.
Discussion
Evidence for a dual role of MgtC
Previous studies on mgtC mutants have indicated thatthe growth defect in macrophages correlated with agrowth defect in low Mg2+ medium, suggesting a caus-ative link between the two phenotypes (Blanc-Potardand Groisman, 1997; Buchmeier et al., 2000; Lavigneet al., 2005). However, we have now identified MgtChomologues from P. luminescens and P. aeruginosa thatfunctionally replace the Salmonella MgtC protein in lowMg2+ medium but not in macrophages. In addition, asimilar phenotypic profile was found with a SalmonellaMgtC mutated protein at position C99. These resultsindicate for the first time a lack of correlation between invitro and in vivo phenotypes and suggest that the role ofMgtC in macrophages does not simply reflect adaptationto a low Mg2+ environment. Moreover, we describe a Sal-monella MgtC variant, mutated at position W226, thatexhibited the opposite phenotypic profile, i.e. failed topromote growth in low Mg2+ medium but still retainedfunction in macrophages. Cumulatively, these resultsindicate that the ability of MgtC to favour bacterialgrowth in macrophages is unexpectedly mainly indepen-dent from the ability of MgtC to favour bacterial growth
in low Mg2+ culture. This finding suggests that MgtC hasa dual role depending on whether bacteria reside in hostmacrophages or liquid culture.
Dual role of MgtC in relation to structural organization
Sequence alignment and hydrophobicity pattern clearlydefine two domains in MgtC. A combination of experimen-tal determination of the C-terminus location and predictivetopological models indicates that the Salmonella MgtChas five transmembrane domains in the N-terminal regionand a cytoplasmic C-terminal domain (Fig. 9). The dualrole of MgtC cannot be attributed to functionally indepen-dent structural domains because essential residues forboth in vitro and in vivo phenotypes are found in theN-terminal domain (E84 and N92). However, subdomainsof MgtC might have specific roles (Fig. 9). Indeed, a sub-domain from the N-terminal part might be specificallyimportant for function in host (see below) whereas theC-terminal end of the protein might have a specific role inlow Mg2+ medium. The role of the C-terminal end in vitro isbased on the identification of W226A substitution thatexhibits a loss-of-function phenotype in low Mg2+ culturebut is still able to promote growth in macrophages. W226is likely to have a structural role determinant for properfunction. The precise function of MgtC in low Mg2+ adap-tation remains unknown. The soluble C-terminal domain ispredicted to adopt a babbab topology based on structuralsimilarities by fold-recognition. This fold, that has beenpartially confirmed by NMR spectroscopy, is found inmetal binding domain. However, transport and electro-physiological studies demonstrate that MgtC does notdirectly transport Mg2+ (Moncrief and Maguire, 1998;
Fig. 9. Schematization of the MgtCtopological domains and position of residuesimportant for function in low Mg2+ cultureand/or in macrophages. The topologicalmodel of MgtC (five transmembrane helices,TM1 to TM5, and a cytoplasmic C-terminaldomain) is derived from a combination oftopology prediction and experimentaldetermination of C-terminus location.Secondary structure of the C-terminal domaincharacterized by NMR is indicated in plainlane. The remaining predicted secondarystructure, based on fold-recognition, isindicated in dotted line. a-Helices areschematized by rectangles and b-strands byarrows. The position of amino acids that areimportant for function based on the analysisof Salmonella MgtC, as well asP. luminescens and P. aeruginosa MgtC, areindicated.
msalpireP
msalpotyC
N
5MT4MT3MT2MT1MTrennI
enarbmem
622W
99C
29N
β1
β2
β3
48E
411N
C
niamodlanimret-N
niamodlanimret-C
eloR ortivni dna ovivni
ylniameloR ovivni
ylniameloR ortivni
α1
α2
β4
Dual role of the MgtC virulence factor 615
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

Günzel et al., 2006). Our present data further indicate thatMgtC does not bind Mg2+, because Mg2+ addition does notchange the structure of the C-terminal domain and noputative Mg2+ binding residues were identified among con-served negatively charged amino acids. The predictedfold of MgtC C-terminal domain is also found in conservedregulatory domains, RRM and ACT domains, that bindRNA, proteins or amino acids. This finding, together withthe cytoplasmic location of the C-terminal domain, promptus to propose that the role of MgtC in a low Mg2+ environ-ment might be linked to a yet uncharacterized regulatoryfunction. In addition, examination of bacterial morphologyindicates that MgtC prevents cell elongation and autoag-gregation in low Mg2+ medium. The role of MgtC in lowMg2+ medium might not be directly linked to Mg2+ uptakebut might be linked to side-effects of Mg2+ deprivation.Magnesium is important for membrane stability and hasbeen involved in the modification of LPS and formation offlagella (Gibbons et al., 2005; O’Shea et al., 2005).Hence, MgtC might act indirectly on bacterial surfacemodification and/or bacterial division in low Mg2+ medium.
In contrast to what we had anticipated based on MgtCprotein alignment, the conserved N-terminal transmem-brane domain, rather than the variable C-terminal domain,appears to be essential for the role of MgtC inmacrophages. We have identified single amino acidschanges that prevent or promote MgtC function in mac-rophages without affecting significantly MgtC function inlow Mg2+ culture (Fig. 9). Interestingly, a subdomain thatincludes the C99 residue, located in the fourth transmem-brane helix, might play a specific role in host. Theloss-of-function phenotype in macrophages of the conser-vative substitution C99S suggests that C99 might beinvolved a disulphide bridge. The C99 residue may con-tribute to an overall conformation of MgtC or to interactionwith an as yet unidentified cofactor or protein when bac-teria are within macrophages. Interestingly, the C99amino acid appears to be on the same face of the trans-membrane helix as the N92 residue, which is important forfunction both in macrophages and low Mg2+ medium. Inaddition, overexpression of C99A or C99S MgtC mutantsin a Salmonella wild-type strain gave a dominant-negativephenotype in macrophages, but not in low Mg2+ medium.A similar phenotype was found by overexpressing MgtCproteins from P. luminescens and P. aeruginosa. One canhypothesize that MgtC function in macrophages, but notin liquid culture medium, is dependent on oligomeric MgtCcomplexes and that dominant-negative phenotype is dueto the formation of non-functional complexes containingvariant and wild-type monomers. A GxxxG motif (Seneset al., 2004) present in the third predicted transmembranedomain of MgtC could be involved in helix–helixassociation between MgtC monomers. Alternatively, thedominant-negative phenotype in macrophages could be
related to the competition of non-functional MgtC proteinsin the interaction with other partners. Interestingly, a veryrecent study has proposed that MgtC activates the hostNa+, K+, ATPase (Günzel et al., 2006). This finding isissued from Salmonella MgtC expression in Xenopusoocytes and the mechanism of activation as well as itsphysiological relevance during Salmonella infectionremains unknown. However, this putative interference ofMgtC with a host transporter might be related to the host-specific role of MgtC identified in our study. Further analy-sis will be required to characterize putative changes inMgtC protein localization and interaction with itself and/orwith other components when bacteria are in host cells.
Dual role of MgtC in relation to bacterial pathogenniches
Magnesium has been proposed as a key signal in viru-lence gene regulation inside cells through the PhoP–PhoQ two-component system (Garcia Vescovi et al.,1996; Groisman, 1998) and indirect studies based onexpression of PhoP-activated genes have indicated thatintracellular concentration might be in the micromolarrange (Garcia-del Portillo et al., 1992; Smith et al., 1998;Eriksson et al., 2003). However, other signals, as cat-ionic peptides, can activate the PhoPQ system andmight play a role for the intracellular activation of thesystem (Bader et al., 2005). A recent study, based onthe direct measurement of Mg2+, indicates that Mg2+ con-centration of the phagosome remains around 1 mM inthe early stages of infection (Martin-Orozco et al., 2006).The possibility of a drop of the phagosomal concentra-tion of Mg2+ at later times has not been experimentallytested. Our present results suggest that the role of MgtCin macrophages is not linked to Mg2+ adaptationbecause the in vivo phenotype could be dissociatedfrom the in vitro phenotype. Furthermore, analysis ofbacterial morphology by fluorescent microscopy indi-cates that a Salmonella DmgtC mutant forms elongatedand autoaggregated bacteria in low Mg2+ medium butnot in macrophages. Taken together, our results indicatethat mechanisms underlying growth inhibition of a mgtCnull mutant are different in vitro and in vivo, inferring thatthe phagosome does not necessarily constitute a lowMg2+ environment.
Interestingly, our results suggest that MgtC might haveevolved diversely depending on the replication niche ofbacterial pathogens. Analysis of mgtC mutants indicatedthan MgtC proteins from intracellular pathogens have theability to promote growth both in low Mg2+ medium and inmacrophages (Blanc-Potard and Groisman, 1997; Buch-meier et al., 2000; Lavigne et al., 2005). In the presentstudy, we have expressed in Salmonella MgtC homo-logues from the extracellular pathogens Y. pestis,
616 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

P. aeruginosa and P. luminescens. Whereas all MgtChomologues were able to functionally replace the Salmo-nella MgtC in low Mg2+ medium, the MgtC protein fromY. pestis was the only one that also could replace theSalmonella MgtC in macrophages. Yersinia species areextracellular pathogens that are able to replicate withinmacrophages, a feature that might be important at theearly stage of infection (Pujol and Bliska, 2005). Hence,the fact that Salmonella and Y. pestis encode MgtC ortho-logues is probably linked to the fact that both pathogensexperience the intramacrophage niche during infection. Inagreement with this point of view, a recent report actuallyshowed that MgtC contributes to Y. pestis intramacroph-age survival (Grabenstein et al., 2006). Despite the highsimilarity between Y. pestis and P. luminescens MgtC pro-teins, the latter failed to promote intramacrophage growth.However, sequence comparison of both proteins allowedus to identify a single amino acid change that induces again-of-function in macrophages for MgtC homologuesfrom P. luminescens or P. aeruginosa. This resultsuggests that a simple mutational event could probablydrive MgtC to evolve to gain- or loss-of-function inmacrophages. The adaptation to a Mg2+ deprived environ-ment might have a physiological role for extracellularpathogens because it has been proposed thatP. aeruginosa may encounter Mg2+ limitation in the earlystages of infection (Guina et al., 2003). However, thenatural niches that might be deprived in low Mg2+ areunknown. It is of particular interest that MgtC (PA4635) isa highly induced membrane protein of P. aeruginosa afterMg2+ starvation, suggesting a major role of this protein insuch stress conditions (Guina et al., 2003). Analysis ofP. aeruginosa and P. luminescens mgtC null mutantswould be important to better understand the contributionof adaptation to Mg2+ limitation to the virulence of extra-cellular pathogens.
Concluding remarks
In conclusion, our results indicate that MgtC role in mac-rophages does not coincide with MgtC role in low Mg2+
culture because we have shown for the first time that thephenotypes associated to MgtC defect in vivo and in vitrocan be separated. Our results further suggest that MgtCrole in macrophages, but not in low Mg2+ medium, mightrequire protein oligomerization and/or interaction withother components. The role of MgtC in macrophagesmight involve modulation of the activity of host proteins,as Na+, K+, ATPase (Günzel et al., 2006). In an evolution-ary point of view, MgtC might have had as primary role toallow adaptation to low Mg2+ conditions, a feature thatcould play a role in the virulence of some extracellularpathogens, and an additional role might have evolved inpathogens that survive inside macrophages.
Experimental procedures
Bacterial strains, growth conditions, plasmids andprimers
Bacterial strains used in this study are listed in Table 1. AllS. Typhimurium strains are derived from wild-type 14028s(with the exception of SB300#A and NM153 that are derivedfrom SL1344). E. coli DH5a (Hanahan, 1983) was used ashost in cloning experiments. Plasmids were introduced intoSalmonella by electroporation using a Bio-Rad apparatusand into E. coli by chemical transformation using standardprocedures. Plasmids used in this study are listed in Table 1and primers are listed in supplementary Table S1. Bacteriawere grown in Luria Broth (LB) supplemented with100 mg ml-1 ampicillin to maintain plasmid DNA. Kanamycinand chlorenphenicol were used at concentration of 25 mg ml-1
and 10 mg ml-1 respectively. Growth in low Mg2+ liquidmedium was carried out in NCE-minimal medium (Maloy,1990) supplemented with 0.1% casamino acids, 38 mM glyc-erol and 10 mM MgCl2. Growth in high Mg2+ liquid mediumwas carried out by adding 10 mM MgCl2.
Structure modelling
Sequence comparisons were performed using PSI-BLAST atthe NCBI server. The fold compatibility between MgtCC-terminal domain and Protein Data Bank entries was analy-sed using the meta-server @TOME (Douguet and Labesse,2001). Three-dimensional models were built using MODELLER
(Sali and Blundell, 1993) for the top-ranking structuralalignments. The programs PROSA (Sippl, 1993) and Verify 3D(Eisenberg et al., 1997) were used to evaluate the quality ofthe theoretical models.
Location of Salmonella MgtC C-terminal domain
Five topology prediction methods were used, TMHMM 2.0(http://www.cbs.dtu.dk/services/TMHMM/), HMMTOP 2.0(http://www.enzim.hu/hmmtop/), MEMSAT (http://bioinf.cs.ucl.ac.uk/psipred/ucsd.edu/memsat.html), TOPPRED 2.0 (http://www.sbc.su.se/~erikw/toppred2/) and TMPRED (http://www.ch.embnet.org/software/TMPRED_form.html).
PhoA fusion proteins were constructed using the pPho1reporter plasmid that harbours the phoA gene cloned intopET21(+) and regulated by a T7 promoter (Roth and Hager-hall, 2001; Throne-Holst, 2001); the PhoA protein encoded bypPho1 is missing the N-terminal export signal sequence (first21 amino acids). Polymerase chain reaction (PCR) fragmentsencoding part of MgtC were amplified using primers phoA1and phoA2 (MgtCW226), phoA4 (MgtCG63) or phoA5 (MgtCQ111)and pEG9094 plasmid as a template, and were cloned at theKpnI and BamHI sites of pPho1. The resulting plasmids,pNM6, pNM9 and pNM40, were transformed by electropora-tion into NM153. NM153 is a SB300#A derivative, thatexpresses an arabinose inducible T7 polymerase (McKinneyet al., 2002), in which a phoN::Tn10 insertion has been trans-duced to avoid endogenous phosphatase activity. PhoAassays were performed as previously described (Manoil,1991) from culture grown in 0.2% arabinose.
Dual role of the MgtC virulence factor 617
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

For the MgtCW226–LacZ fusion, a mgtC PCR fragmentamplified using primers mgtC-lacZ-F and mgtC-lacZ-5R (thatincludes a AvrII restriction site) was first cloned at the BamHIand PstI sites of pUHE21–2lacI q. The resulting plasmid wasused to clone a lacZ DNA fragment missing nine codons atthe 5′ end at AvrII and HindIII restriction sites. This lacZ DNAfragment was colony PCR amplified using E. coli MG1655 astemplate and 4140 and 4141 primers (Lee et al., 2004). Theresulting plasmid, pNM43, was transformed by electropora-tion into the Salmonella NM153 strain. The b-galactosidaseassays were performed as described previously from culturegrown in 1 mM IPTG (Miller, 1972). As control, the lacZ geneof pNM43 was replaced by the phoA gene using a phoA PCRfragment amplified with primers 4143-AvrII and 4144 (Lee
et al., 2004). The resulting plasmid, that harbours aMgtCW226–PhoA fusion, gave similar phosphatase activity aspNM6 plasmid (data not shown). Expression of proteinfusions from pNM6 and pNM43 plasmids was checked byWestern blot analysis using polyclonal antibodies raisedagainst the C-terminal domain of the MgtC protein.
Overexpression and purification of a His-taggedC-terminal domain of MgtC
A PCR fragment encoding the C-terminal part of SalmonellaMgtC was amplified using mgtC-AEG-Bam and mgtC-R-Hindprimers and cloned in the pQE30 vector (Qiagen). The result-
Table 1. Bacterial strains and plasmids used in this study.
Strain or plasmid Description or phenotype Reference or source
S. enterica serovar Typhimurium strains14028s Wild type E.A. GroismanEG9527 mgtCB9527::MudJ Garcia Vescovi et al. (1996)EG9785 mgtA9926::MudCam E.A. GroismanEG10575 corA52::Tn10 E.A. GroismanMA6987 ilvI3305::Tn10dTac-cat/pKD46 Uzzau et al. (2001)MA6944 phoN52::Tn10Tc L. BossiSB300A#1 Arabinose-inducible T7 RNA polymerase McKinney et al. (2002)NM14 DmgtC This studyNM80 DmgtC mgtA9926::MudCam corA52::Tn10 This studyNM153 phoN52::Tn10Tc arabinose-inducible T7 RNA polymerase This study
PlasmidspBR322 ApR TcR reppmb1 Bolivar et al. (1977)pPho1 phoA fusion vector Throne-Holst (2001)pUHE21-2lacIq ApR reppmb1 lacIq Soncini et al. (1995)pEG9094 ApR reppmb1 mgtC+ Blanc-Potard and Groisman (1997)pRZT3 TetR DsRed.T3 A.M. van der SarpNM2 MgtC143-231 into pQE30 This studypNM6 MgtCW226–PhoA This studypNM9 MgtCG63–PhoA This studypNM11 pBR322/mgtC regulatory sequences This studypNM12 pNM11/mgtC Salmonella This studypNM12 E27A Mutation Salmonella MgtC E27A This studypNM12 E84A Mutation Salmonella MgtC E84A This studypNM12 E84Q Mutation Salmonella MgtC E84Q This studypNM12 E158K Mutation Salmonella MgtC E158K This studypNM12 D159A Mutation Salmonella MgtC D159A This studypNM12 E160A Mutation Salmonella MgtC E160A This studypNM12 E193A Mutation Salmonella MgtC E193A This studypNM12 E209A Mutation Salmonella MgtC E209A This studypNM12 N92T Mutation Salmonella MgtC N92T This studypNM12 C99A Mutation Salmonella MgtC C99A This studypNM12 C99S Mutation Salmonella MgtC C99S This studypNM12 C155A Mutation Salmonella MgtC C155A This studypNM12 W226A Mutation Salmonella MgtC W226A This studypNM12 W226F Mutation Salmonella MgtC W226F This studypNM14 pNM11/mgtC Y. pestis (YPO1660) This studypNM15 pNM11/Y. pestis YPO2332 This studypNM17 pNM11/mgtC P. luminescens This studypNM17 l114N Mutation MgtC P. luminescens L114N This studypNM19 pNM11/mgtC P. aeruginosa (PA4635) This studypNM19 E112N Mutation MgtC P. aeruginosa E112N This studypNM20 pNM11/P. aeruginosa PA2558 This studypNM23 pNM11/E. coli yhiD This studypNM40 MgtCQ111-PhoA This studypNM43 MgtCW226–LacZ This studypNM71 mgtC Y. p/P. luminescens This studypNM72 mgtC P. l/Y. pestis This study
618 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

ing plasmid, pNM2, encoding His6-MgtC143-231 was transferredto E. coli strain M15/pREP4 (Qiagen). The resulting strainwas grown to mid-exponential phase in LB Amp Kan contain-ing 1 mM IPTG. The cells were collected by centrifugation(5000 r.p.m., 10 min, 4°C) and were resuspended in 10 ml of20 mM NaPO4 (pH 7.4), 500 mM NaCl, 100 mM imidazole,2 mM B-mercaptoethanol. Cells were lysed by adding 10 mglysozyme, 20 ml of PMSF (7 mg ml-1), followed by sonication.The cell fraction was loaded onto a Ni-NTA agarose column(Qiagen) and eluted with 20 mM NaPO4 (pH 7.4), 500 mMNaCl, 300 mM imidazole, 2 mM B-mercaptoethanol.Fractions containing His-tagged MgtC were identified bySDS-PAGE. Gel filtration was performed on Superdex75 (Pharmacia) as a final step of purification in 20 mMNaPO4 (pH 7.4), 200 mM NaCl, with or without 2 mMb-mercaptoethanol. The protein purity was verified onSDS-PAGE.
NMR analysis of MgtC C-terminal in presence orabsence of Mg 2+
NMR samples were prepared in 10 mM NaPO4, 200 mMNaCl and 2 mM DTT. The pH of the samples was 5.8 and theconcentration was 0.6 mM. NMR data were acquired at300 K on a Bruker Avance 500 MHz and 600 MHz NMRspectrometers, both equipped with cryogenic triple-resonance probes. Resonance assignments were obtainedfrom 2D 1H–15N HSQC and 3D 15N NOESY-HSQC (Sattleret al., 1999) using uniformly 15N-labelled His6-MgtC143-231 andfrom 2D 1H-1H TOCSY and 2D 1H-1HNOESY using unlabelledHis6-MgtC143-231. NMR data were processed using BrukerXWINNMR v 3.0 or Gifa v 5.0 (Pons et al., 1996).
For magnesium titration, MgCl2 was added to the sample ata concentration ranging from 200 mM to 20 mM. A 2D 1H–15NHSQC was acquired after each Mg2+ addition at 300 K on a200 mM solution of uniformly 15N-labelled MgtC at pH 6.5 in20 mM sodium phosphate, 200 mM NaCl and 2 mM DTT.
Construction of a non-polar DmgtC Salmonella strain
A chromosomal deletion in the mgtC gene was constructedusing lRed mediated site-specific recombination (Datsenkoand Wanner, 2000). The kanamycin resistance gene ofplasmid pKD13 (Datsenko and Wanner, 2000) was amplifiedvia PCR using primers pp352 and pp353. The resulting PCRproduct was used to replace most of the coding sequence ofmgtC in strain MA6897 (Uzzau et al., 2001) by homologousrecombination catalysed by the lRed recombinase encodedby plasmid pKD46. The kanamycin resistance cassetteinserted at the chromosomal mgtC locus was transferred byP22 transduction into a wild-type 14028s strain. Thetemperature-sensitive plasmid pCP20 encoding FLP recom-binase (Datsenko and Wanner, 2000) was transformed in theresulting strain to allow loss of the kanamycin resistancecassette leaving behind a single FRT site. The chromosomaldeletion was verified by colony PCR using primers mgtC-5Fand mgtC-3R followed by sequencing. The resulting strain(NM14) exhibits similar phenotypes both in low Mg2+ mediumand macrophages as EG9527, a mgtCB::MudJ mutant thathas a polar effect on mgtB (Blanc-Potard and Groisman,
1997). Furthermore, a triple mutant DmgtC corA::Tn10mgtA::MudCam (NM80) is able to grow on LB plates.This phenotype confirms that MgtB is functional in the pre-sence of the DmgtC mutation because a triple mutantcorA–mgtA–mgtCB– requires 10 mM Mg2+ for growth (Hmielet al., 1989).
Heterologous expression of MgtC-like proteins
MgtC-like proteins from diverse g-proteobacteria have beenexpressed in Salmonella under the mgtC regulatorysequences from S. Typhimurium. We have first amplified byPCR a 570 bp sequence upstream of mgtC that includestranscriptional regulatory sequences (Lejona et al., 2003)using primers pr-mgtC-for-SalI and pr-mgtC-rev-SphI andpEG9094 plasmid as template. To ensure the presence ofefficient translational regulatory sequences, the PCR frag-ment also harbours the first 11 nucleotides of mgtC, thatincludes a perfect Shine-Dalgarno concensus sequence, fol-lowed by an SphI site, that includes the ATG start codon. Theresulting PCR product was cloned at the SalI and SphI sitesof pBR322 plasmid to produce pNM11 plasmid. SalmonellamgtC gene was amplified using primers mgtCforwSphInewand mgtCrevEcoRI and pEG9094 as template, and clonedinto pNM11 at SphI and EcoRI sites to produce pNM12. MgtCgene homologues from Y. pestis, P. luminescens, E. coli andP. aeruginosa were amplified using primers indicated inTable S1 and cloned into pNM11 at SphI and EcoRI sites.Y. pestis mgtC and YPO2332 genes were amplified fromCO92 chromosomal DNA. P. luminescens mgtC gene wasamplified from TT01 chromosomal DNA. The E. coli yhiDgene was amplified by colony PCR from MG1655 strain,P. aeruginosa mgtC and PA2558 genes were amplified bycolony PCR from PA01 strain.
Site-directed mutagenesis
The pNM12 plasmid, that harbours mgtC gene from S. Typh-imurium, served as template for introduction of mutations inthe Salmonella MgtC protein. Site-directed mutagenesis wasconducted using the Quickchange® II kit (Stratagene)according to manufacturer’s instructions. Sets of primerswere designed for each engineered mutation including suffi-ciently long flanking regions. Sequencing was conducted toconfirm the presence of the desired mutation and absence ofadditional mutations in the mgtC gene. The levels of mutantMgtC proteins were determined by Western blot analysisusing polyclonal antibodies raised against the C-terminaldomain of the MgtC protein. Site-directed mutagenesis ofP. luminescens and P. aeruginosa MgtC proteins was per-formed on the pNM17 and pNM19 plasmids respectively.
Construction of chimeric proteins that exchangeN-terminal end
We have exchanged the N-terminal end between MgtCproteins of P. luminescens and Y. pestis. A SacI site wascreated by site-directed mutagenesis (Quickchange® II kit,Stratagene) at position +95 and +62 in the mgtCgene of P. luminescens and Y. pestis, respectively,
Dual role of the MgtC virulence factor 619
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

using mgtCphluSac1 and mgtCphluSac2 primers andmgtCYPSac1and mgtCYPSac2 primers respectively. Thesemutagenesis did not change the amino acid sequence ofMgtC proteins. Chimeric genes were constructed by using aunique PstI site located in the pBR322 plasmid. The Y.p/P.luminescens chimeric gene was created by replacing the
1.4 kb PstI–SacI fragment of the Y. pestis mgtC gene with thecorresponding PstI–SacI fragment from the P. luminescensmgtC gene. Similarly, the P.l/Y. pestis chimeric gene wascreated by replacing the 1.4 kb PstI–SacI fragment of theP. luminescens mgtC gene with the corresponding PstI–SacIfragment from the Y. pestis mgtC gene. The resulting chi-meric proteins have exchanged the first 20 residues (the 34following residues are identical between P. luminescens andY. pestis).
Complementation experiments in S. Typhimurium
The S. Typhimurium NM14 strain (DmgtC) was transformedby electroporation with plasmids encoding MgtC mutated pro-teins or MgtC proteins from other bacteria. NM14 was alsotransformed by pNM11 that harbours only the mgtC promoteras negative control and by pNM12 that harbours the wild-typeSalmonella mgtC gene as positive control. Two phenotypeswere tested in complementation experiments: growth in mag-nesium deprived medium and intramacrophage survival.Growth in Mg2+ deprived medium was performed in NCEmedium supplemented with 10 mM MgCl2 (Blanc-Potard andGroisman, 1997). Overnight cultures of the different strainswere diluted 1:100 in NCE medium containing 10 mM Mg2+
and incubated at 37°C. Their optical densities at 600 nm(OD600) were measured after 18 h-incubation. As control,growth of strains was systematically tested in high Mg2+
medium. The rate of intramacrophage replication after 18 hinfection was performed in J774 mouse macrophages essen-tially as described previously (Blanc-Potard and Groisman,1997). Briefly, macrophages were seeded at a density of5 ¥ 105 in 24-well plates and grown in DMEM media supple-mented with 10% (v/v) fetal bovine serum at 37°C, 5% CO2.Bacteria were added to the wells at macrophage-to-bacteriaratio of 1:10. Bacterial phagocytosis was allowed to proceedfor 25 min before washing three times with sterile PBS andadding fresh DMEM media supplemented with 20 mg ml-1
gentamicin. Macrophages were lysed by using 1% TritonX-100 and the number of viable bacteria that survived gen-tamicin treatment was determined by subsequent platingonto LB agar plates. Replication rate was determined as theratio between the number of bacteria at time 18 h and thenumber of internalized bacteria after 25 min phagocytosis.
Examination of bacterial morphology by fluorescencemicroscopy
Salmonella wild-type strain 14028s and DmgtC mutant NM14were transformed with the pRZT3 plasmid that encodesDsRed protein. Plasmid pRZT3 (A.M. van der Sar, pers.comm.) results from the cloning of a 0.9 kb PvuII fragmentfrom pGMDs3 (van der Sar et al., 2003), containing theDsRed.T3 gene under control of the lac promoter, in theEcl136II site of pME6031 (Heeb et al., 2000). To examine
in vitro morphology, overnight cultures were diluted 1:100 inNCE medium containing 10 mM or 10 mM Mg2+ and incubatedat 37°C for 8 h or 18 h. For fluorescence microscopy, 1 ml ofbacterial culture was placed on a microspcope slide coatedwith a thin layer of 1% agarose in PBS and a coverslip wasplaced on top. To examine in vivo microscopy, J774 macroph-ages grown on coverslip in 12-well plates were infected withDsRed expressing bacteria for 7 h or 18 h. After infectioncells were washed three times with PBS and fixed with 3%paraformaldehyde for 20 min. Fixed cells were washed withPBS and were processed for LPS labelling without cell per-meabilization to specifically label remaining extracellularbacteria. Immunofluorescence labelling of LPS was per-formed using a polyclonal rabbit antiserum and a fluoresceinisothiocyanate-labelled anti-rabbit serum (Sigma) as second-ary antibody. Fluorescent bacteria were visualized using aLeica DM IRB microscope and images were acquired withMetavue Software. DsRed expression was visualized usinggreen filter whereas LPS labelling was visualized using redfilter. Intracellular bacteria showed only red fluorescence (nogreen fluorescence).
Acknowledgements
We thank L. Bossi, A. Filloux, E.A. Groisman, L. Hederstedt,J. McKinney, A.M. van der Sar for providing bacterial strainsand plasmids and E. Carniel, A. Givaudan for providing chro-mosomal DNA. We thank L. Bossi for help in the constructionof DmgtC mutant, A. Gastebois and A. Bordes for contributionto macrophage experiments, and A. Rostan for preliminaryexperiments with DsRed expressing bacteria. We thankseveral members of the platform RIO (CBS, Montpellier): G.Labesse for comparative modelling analysis and helpful dis-cussions, Y. Bessin and N. Declerck for biochemical analysis,and C. Roumestand for preliminary NMR analysis. We thankD. O’Callaghan for critical reading of the manuscript. Thiswork is supported by the Avenir program of Inserm. C.R. is anInserm young researcher fellow. E.A. is supported by Insermand Région Languedoc-Roussillon. A.B.B.P. is supported byCANAM.
References
Alix, E., Godreuil, S., and Blanc-Potard, A.B. (2006) Identi-fication of a single nucleotide polymorphism specific tothe Haarlem genotype in the mgtC virulence gene ofMycobacterium tuberculosis. J Clin Microbiol 44: 2093–2098.
Bader, M.W., Sanowar, S., Daley, M.E., Schneider, A.R.,Cho, U., Xu, W., et al. (2005) Recognition of antimicrobialpeptides by a bacterial sensor kinase. Cell 122: 461–472.
Blanc-Potard, A.B., and Groisman, E.A. (1997) The Salmo-nella selC locus contains a pathogenicity island mediatingintramacrophage survival. EMBO J 16: 5376–5385.
Blanc-Potard, A.B., and Lafay, B. (2003) MgtC as ahorizontally-acquired virulence factor of intracellular bacte-rial pathogens: evidence from molecular phylogeny andcomparative genomics. J Mol Evol 57: 479–486.
Bolivar, F., Rodriguez, R.L., Greene, P.J., Betlach, M.C.,Heyneker, H.L., and Boyer, H.W. (1997) Construction and
620 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

characterization of new cloning vehicles. II. A multipurposecloning system. Gene 2: 95–113.
Buchmeier, N., Blanc-Potard, A.B., Ehrt, S., Piddington, D.,Riley, L., and Groisman, E.A. (2000) A parallel intraphago-somal survival strategy shared by Mycobacterium tubercu-losis and Salmonella enterica. Mol Microbiol 35: 1375–1382.
Chamnongpol, S., Cromie, M., and Groisman, E.A. (2003)Mg2+ sensing by the Mg2+ sensor PhoQ of Salmonellaenterica. J Mol Biol 325: 795–807.
Datsenko, K.A., and Wanner, B.L. (2000) One-step inactiva-tion of chromosomal genes in Escherichia coli K-12 usingPCR products. Proc Natl Acad Sci USA 97: 6640–6645.
Douguet, D., and Labesse, G. (2001) Easier threadingthrough web-based comparisons and cross-validations.Bioinformatics 17: 752–753.
Drew, D., Sjostrand, D., Nilsson, J., Urbig, T., Chin, C.N., deGier, J.W., and von Heijne, G. (2002) Rapid topologymapping of Escherichia coli inner-membrane proteins byprediction and PhoA/GFP fusion analysis. Proc Natl AcadSci USA 99: 2690–2695.
Eisenberg, D., Luthy, R., and Bowie, J.U. (1997) VERIFY3D:assessment of protein models with three-dimensionalprofiles. Methods Enzymol 277: 396–404.
Eriksson, S., Lucchini, S., Thompson, A., Rhen, M., andHinton, J.C. (2003) Unravelling the biology of macrophageinfection by gene expression profiling of intracellular Sal-monella enterica. Mol Microbiol 47: 103–118.
Garcia Vescovi, E., Soncini, F.C., and Groisman, E.A. (1996)Mg2+ as an extracellular signal: environmental regulation ofSalmonella virulence. Cell 84: 165–174.
Garcia-del Portillo, F., Foster, J.W., Maguire, M.E., and Finlay,B.B. (1992) Characterization of the micro-environment ofSalmonella typhimurium-containing vacuoles within MDCKepithelial cells. Mol Microbiol 6: 3289–3297.
Gibbons, H.S., Kalb, S.R., Cotter, R.J., and Raetz, C.R.(2005) Role of Mg2+ and pH in the modification of Salmo-nella lipid A after endocytosis by macrophage tumourcells.? Mol Microbiol 55: 425–440.
Grabenstein, J.P., Fukuto, H.S., Palmer, L.E., and Bliska,J.B. (2006) Characterization of phagosome trafficking andidentification of PhoP-regulated genes important for sur-vival of Yersinia pestis in macrophages. Infect Immun 74:3727–3741.
Groisman, E.A. (1998) The ins and outs of virulence geneexpression: Mg2+ as a regulatory signal. BioEssays 20:96–101.
Guina, T., Purvine, S.O., Yi, E.C., Eng, J., Goodlett, D.R.,Aebersold, R., and Miller, S.I. (2003) Quantitative pro-teomic analysis indicates increased synthesis of a qui-nolone by Pseudomonas aeruginosa isolates from cysticfibrosis airways. Proc Natl Acad Sci USA 100: 2771–2776.
Günzel, D., Kucharski, L.M., Kehres, D.G., Romero, M.F.,and Maguire, M.E. (2006) The MgtC virulence factor ofSalmonella enterica serovar Typhimurium activates Na+,K+-ATPase. J Bacteriol 188: 5586–5594.
Hanahan, D. (1983) Studies on transformation of Escherichiacoli with plasmids. J Mol Biol 166: 557–580.
Heeb, S., Itoh, Y., Nishijyo, T., Schnider, U., Keel, C., Wade,J., et al. (2000) Small, stable shuttle vectors based on the
minimal pVS1 replicon for use in Gram-negative, plant-associated bacteria. Mol Plant Microbe Interact 13: 232–237.
Hmiel, S.P., Snavely, M.D., Florer, J.B., Maguire, M.E., andMiller, C.G. (1989) Magnesium transport in Salmonellatyphimurium: genetic characterization and cloning of threemagnesium transport loci. J Bacteriol 171: 4742–4751.
Lavigne, J.P., O’Callaghan, D., and Blanc-Potard, A.B.(2005) MgtC is required for Brucella suis intramacrophagegrowth: a potential mechanism of adaptation to low Mg2+
environment shared by Salmonella enterica and Mycobac-terium tuberculosis. Infect Immun 73: 3160–3163.
Lawley, T.D., Chan, K., Thompson, L.J., Kim, C.C., Govoni,G.R., and Monack, D.M. (2006) Genome-wide screen forSalmonella genes required for long-term systemic infectionof the mouse. PLoS Pathog 2: e11.
Lee, H., Hsu, F.F., Turk, J., and Groisman, E.A. (2004) ThePmrA-regulated pmrC gene mediates phosphoethanola-mine modification of lipid A and polymyxin resistance inSalmonella enterica. J Bacteriol 186: 4124–4133.
Lejona, S., Aguirre, A., Cabeza, M.L., Garcia Vescovi, E., andSoncini, F.C. (2003) Molecular characterization of theMg2+-responsive PhoP–PhoQ regulon in Salmonellaenterica. J Bacteriol 185: 6287–6294.
McKinney, J., Guerrier-Takada, C., Galan, J., and Altman, S.(2002) Tightly regulated gene expression system in Salmo-nella enterica serovar Typhimurium. J Bacteriol 184: 6056–6059.
Maloney, K.E., and Valvano, M.A. (2006) The mgtC gene ofBurkholderia cenocepacia is required for growth undermagnesium limitation conditions and intracellular survivalin macrophages. Infect Immun 74: 5477–5486.
Maloy, S.R. (1990) Experimental Techniques in BacterialGenetics. Boston, MA: Jones and Bartlett Publishers.
Manoil, C. (1991) Analysis of membrane protein topologyusing alkaline phosphatase and beta-galactosidase genefusions. Methods Cell Biol 34: 61–75.
Martin-Orozco, N., Touret, N., Zaharik, M.L., Park, E., Kopel-man, R., Miller, S., et al. (2006) Visualization of vacuolaracidification-induced transcription of genes of pathogensinside macrophages. Mol Biol Cell 17: 498–510.
Miller, J.H. (1972) Experiments in Molecular Genetics. ColdSpring Harbor, NY: Cold Spring Harbor Laboratory.
Moncrief, M.B., and Maguire, M.E. (1998) Magnesium andthe role of MgtC in growth of Salmonella typhimurium.Infect Immun 66: 3802–3809.
O’Shea, T.M., Deloney-Marino, C.R., Shibata, S., Aizawa, S.,Wolfe, A.J., and Visick, K.L. (2005) Magnesium promotesflagellation of Vibrio fischeri. J Bacteriol 187: 2058–2065.
Pons, J.L., Malliavin, T.E., and Delsuc, M.A. (1996) Gifa V.4:a complete package for NMR data set processing. J BiolNMR 8: 445–452.
Pujol, C., and Bliska, J.B. (2005) Turning Yersinia pathogen-esis outside in: subversion of macrophage function byintracellular yersiniae. Clin Immunol 114: 216–226.
Rapp, M., Drew, D., Daley, D.O., Nilsson, J., Carvalho, T.,Melen, K., et al. (2004) Experimentally based topologymodels for E. coli inner membrane proteins. Protein Sci 13:937–945.
Roth, R., and Hagerhall, C. (2001) Transmembrane orienta-tion and topology of the NADH: quinone oxidoreductase
Dual role of the MgtC virulence factor 621
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works

putative quinone binding subunit NuoH.? Biochim BiophysActa 1504: 352–362.
Sali, A., and Blundell, T.L. (1993) Comparative protein mod-elling by satisfaction of spatial restraints. J Mol Biol 234:779–815.
van der Sar, A.M., Musters, R.J., van Eeden, F.J., Appelmelk,B.J., Vandenbroucke-Grauls, C.M., and Bitter, W. (2003)Zebrafish embryos as a model host for the real time analy-sis of Salmonella typhimurium infections. Cell Microbiol 5:601–611.
Sattler, M., Schleucher, J., and Griesinger, C. (1999) Het-eronuclear multidimensional NMR experiments for thestructure determination of the proteins in solution employ-ing pulsed field gradients. Prog Nucl Mag Res Sp 34:93–158.
Senes, A., Engel, D.E., and DeGrado, W.F. (2004) Folding ofhelical membrane proteins: the role of polar, GxxxG-likeand prolin motifs. Curr Opin Struct Biol 14: 465–479.
Sippl, M.J. (1993) Recognition of errors in three-dimensionalstructures of proteins. Proteins 17: 355–362.
Smith, R.L., Kaczmarek, M.T., Kucharski, L.M., and Maguire,M.E. (1998) Magnesium transport in Salmonella typhimu-rium: regulation of mgtA and mgtCB during invasion ofepithelial and macrophage cells. Microbiology 144: 1835–1843.
Snavely, M.D., Miller, C.G., and Maguire, M.E. (1991)The mgtB Mg2+ transport locus of Salmonella typhimuriumencodes a P-type ATPase. J Biol Chem 266: 815–823.
Soncini, F.C., Vescovi, E.G., and Groisman, E.A. (1995)Transcriptional autoregulation of the Salmonella typhimu-rium phoPQ operon. J Bacteriol 177: 4364–4371.
Tao, T., Snavely, M.D., Farr, S.G., and Maguire, M.E. (1995)Magnesium transport in Salmonella typhimurium: mgtAencodes a P-type ATPase and is regulated by Mg2+ in amanner similar to that of the mgtB P-type ATPase.J Bacteriol 177: 2654–2662.
Tao, T., Grulich, P.F., Kucharski, L.M., Smith, R.L., andMaguire, M.E. (1998) Magnesium transport in Salmonellatyphimurium: biphasic magnesium and time dependence ofthe transcription of the mgtA and mgtCB loci. Microbiology144: 655–664.
Throne-Holst, M. (2001) Synthesis of hemes found in heme-copper oxidases of Bacillus subtilis. PhD Thesis. Lund:University of Lund.
Uzzau, S., Figueroa-Bossi, N., Rubino, S., and Bossi, L.(2001) Epitope tagging of chromosomal genes inSalmonella. Proc Natl Acad Sci USA 98: 15264–15269.
Supplementary material
The following supplementary material is available for thisarticle online:Fig. S1. Ha chemical shift deviations from NMR analysis ofMgtC143-231.Fig. S2. Growth curves of wild-type Salmonella (14028s)and DmgtC strain (NM14) in NCE liquid medium supple-mented with 10 mM or 10 mM Mg2+.Fig. S3. Alignment of amino acid sequences of a subset ofMgtC-like proteins.Table S1. Primers used in this study.
This material is available as part of the online article fromhttp://www.blackwell-synergy.com
622 C. Rang et al.
Journal compilation © 2006 Blackwell Publishing Ltd, Molecular Microbiology, 63, 605–622No claim to original French government works
Related Documents











