&p.1:Abstract The distribution of the vagal nerve was stud- ied in whole-mount specimens and serial sections of chick embryos after retinoic acid treatment. White Leg- horn chick embryos were treated at stage 15 either with 1 μg all-trans retinoic acid (n=11), or with the solvent di- methylsulphoxide (sham-operated embryos, n=8). Eight embryos served as normal controls. At stage 34 all 27 embryos were examined with a dissecting microscope. In order to reveal the vagal patterning, the hearts were re- moved and whole-mount stained with the HNK-1 anti- body. In three hearts of the retinoic acid-treated group a morphologic intracardiac anomaly – a double outlet right ventricle – was found. To explore in depth the vagal nerve distribution in the heart, a separate set of hearts of retinoic acid embryos (n=5), sham-operated (n=4) and control embryos (n=5), was devised solely for serial sec- tioning and staining with the HNK-1 antibody. All hearts of retinoic acid-treated embryos showed a disturbed va- gal nerve distribution both over the surface of the heart and within the heart wall. The vagal patterning was not altered in the sham-operated embryos compared to con- trols. It is concluded that retinoic acid disturbs the devel- opment of vagal nerve patterning regardless of the con- current presence of intracardiac malformations. The mechanism and functional implications remain to be in- vestigated. &kwd:Key words Heart development · Vagal nerve · Immunohistochemistry · Retinoic acid&bdy: Introduction During embryonic development a widespread migration of neural crest cells gives rise to neuronal cell popula- tions of the autonomic nervous system. Sensory innerva- tion of the heart in the chick comes from the nodose gan- glion of the vagus nerve (Walkey and Bower 1981). This ganglion originates from the nodose placode, one of the epibranchial placodes (Narayan and Narayan 1980; D’Amico-Martel and Noden 1983). A morphologic study of the development of cardiac innervation by the vagus nerve (Kuratani and Tanaka 1990) in whole-mount speci- mens of chick embryos stained with anti-neurofilament protein antibody indicated that the final configuration of the vagus nerve is established at stage 34 (day 8 of incu- bation). The vagal branches can be classified in into two categories: (1) the branches that are primarily related to the pharyngeal arch system; (2) the intestinal arboriza- tion derivatives that are associated primarily with the primitive gut. The branchial portion of the vagus nerve consists of the superior cardiac branch innervating the truncus arteriosus of the heart. The intestinal portion of the vagus nerve consists of the sinal branch, pulmonary branches as well as the recurrent nerve and the other in- testinal branches. In normal development of the sympa- thetic innervation to the chick heart, nerves are first seen in the heart at stage 36, day 10 of incubation (Higgins and Pappano 1979; Kirby et al. 1980). In order to study abnormal development of the heart including the autonomic nervous system, animal models are needed. In a previous study an animal model was generated using all-trans retinoic acid (Broekhuizen et al. 1992). All-trans retinoic acid has an essential role in normal development but in excess leads to malforma- tions. Congenital malformations have been described af- ter therapeutic use of retinoids and retinol derivatives, for acnae and psoriasis, during the first trimester of ges- tation (Lammer et al. 1985). These malformations com- prise cardiovascular and facial defects. A single adminis- tration of all-trans retinoic acid given at an early stage in the chick embryo induces outflow tract anomalies, a spe- M.L.A. Broekhuizen · J.W. Wladimiroff Department of Obstetrics and Gynecology, University Hospital Rotterdam, Dr. Molewaterplein 40, 3015 GD Rotterdam, The Netherlands A.C. Gittenberger-de Groot · M.J. Baasten · R.E. Poelmann ( ✉ ) Department of Anatomy and Embryology, Leiden University, Medical Center, P.O. Box 9602, 2300 RC Leiden, The Netherlands Tel.: 31-71-5276660; Fax: 31-71-5276680&/fn-block: Anat Embryol (1998) 197:391–397 © Springer-Verlag 1998 ORIGINAL ARTICLE &roles:Monique L.A. Broekhuizen Adriana C. Gittenberger-de Groot · Mieke J. Baasten Juriy W. Wladimiroff · Robert E. Poelmann Disturbed vagal nerve distribution in embryonic chick hearts after treatment with all-trans retinoic acid &misc:Accepted: 22 January 1998

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

&p.1:Abstract The distribution of the vagal nerve was stud-ied in whole-mount specimens and serial sections ofchick embryos after retinoic acid treatment. White Leg-horn chick embryos were treated at stage 15 either with 1µg all-trans retinoic acid (n=11), or with the solvent di-methylsulphoxide (sham-operated embryos, n=8). Eightembryos served as normal controls. At stage 34 all 27embryos were examined with a dissecting microscope. Inorder to reveal the vagal patterning, the hearts were re-moved and whole-mount stained with the HNK-1 anti-body. In three hearts of the retinoic acid-treated group amorphologic intracardiac anomaly – a double outlet rightventricle – was found. To explore in depth the vagalnerve distribution in the heart, a separate set of hearts ofretinoic acid embryos (n=5), sham-operated (n=4) andcontrol embryos (n=5), was devised solely for serial sec-tioning and staining with the HNK-1 antibody. All heartsof retinoic acid-treated embryos showed a disturbed va-gal nerve distribution both over the surface of the heartand within the heart wall. The vagal patterning was notaltered in the sham-operated embryos compared to con-trols. It is concluded that retinoic acid disturbs the devel-opment of vagal nerve patterning regardless of the con-current presence of intracardiac malformations. Themechanism and functional implications remain to be in-vestigated.
&kwd:Key words Heart development · Vagal nerve ·Immunohistochemistry · Retinoic acid&bdy:
Introduction
During embryonic development a widespread migrationof neural crest cells gives rise to neuronal cell popula-tions of the autonomic nervous system. Sensory innerva-tion of the heart in the chick comes from the nodose gan-glion of the vagus nerve (Walkey and Bower 1981). Thisganglion originates from the nodose placode, one of theepibranchial placodes (Narayan and Narayan 1980;D’Amico-Martel and Noden 1983). A morphologic studyof the development of cardiac innervation by the vagusnerve (Kuratani and Tanaka 1990) in whole-mount speci-mens of chick embryos stained with anti-neurofilamentprotein antibody indicated that the final configuration ofthe vagus nerve is established at stage 34 (day 8 of incu-bation). The vagal branches can be classified in into twocategories: (1) the branches that are primarily related tothe pharyngeal arch system; (2) the intestinal arboriza-tion derivatives that are associated primarily with theprimitive gut. The branchial portion of the vagus nerveconsists of the superior cardiac branch innervating thetruncus arteriosus of the heart. The intestinal portion ofthe vagus nerve consists of the sinal branch, pulmonarybranches as well as the recurrent nerve and the other in-testinal branches. In normal development of the sympa-thetic innervation to the chick heart, nerves are first seenin the heart at stage 36, day 10 of incubation (Higginsand Pappano 1979; Kirby et al. 1980).
In order to study abnormal development of the heartincluding the autonomic nervous system, animal modelsare needed. In a previous study an animal model wasgenerated using all-trans retinoic acid (Broekhuizen etal. 1992). All-trans retinoic acid has an essential role innormal development but in excess leads to malforma-tions. Congenital malformations have been described af-ter therapeutic use of retinoids and retinol derivatives,for acnae and psoriasis, during the first trimester of ges-tation (Lammer et al. 1985). These malformations com-prise cardiovascular and facial defects. A single adminis-tration of all-trans retinoic acid given at an early stage inthe chick embryo induces outflow tract anomalies, a spe-
M.L.A. Broekhuizen · J.W. WladimiroffDepartment of Obstetrics and Gynecology,University Hospital Rotterdam, Dr. Molewaterplein 40,3015 GD Rotterdam, The Netherlands
A.C. Gittenberger-de Groot · M.J. Baasten · R.E. Poelmann (✉)Department of Anatomy and Embryology, Leiden University,Medical Center, P.O. Box 9602, 2300 RC Leiden,The NetherlandsTel.: 31-71-5276660; Fax: 31-71-5276680&/fn-block:
Anat Embryol (1998) 197:391–397 © Springer-Verlag 1998
O R I G I N A L A RT I C L E
&roles:Monique L.A. BroekhuizenAdriana C. Gittenberger-de Groot · Mieke J. BaastenJuriy W. Wladimiroff · Robert E. Poelmann
Disturbed vagal nerve distribution in embryonic chick heartsafter treatment with all-trans retinoic acid
&misc:Accepted: 22 January 1998

cific spectrum of double outlet right ventricle (Broekhui-zen et al. 1992, 1995; Bouman et al. 1995).
Cardiovascular anomalies in rat and mouse embryosfollowing retinoic acid treatment also include outflowtract septation defects (Taylor 1981; Pexieder et al. 1992;Yasui et al. 1995). Another experimental model exists inwhich congenital malformations are described that re-semble those caused by retinoids – the neural crest abla-tion model (Kirby and Waldo 1995). Neural crest abla-tion studies revealed that a part of the vagal neural crest,the “cardiac neural crest”, contributes to the formation ofthe outflow tract and to the cardiac ganglia (Kirby 1987).Experiments with retinoic acid receptors and retinoidsmay influence neural crest differentiation (Chambon1993; Lee et al. 1995). Therefore, we hypothesize thatretinoic acid has an effect on the neural crest that resultsboth in outflow tract anomalies and disturbed vagal dis-tribution of the heart.
The objective of the present study was to establishwhether a difference in nerve branching patients existsbetween hearts of controls, sham-operated and retinoic-acid treated embryos. Stage 34 was selected because thevagal nerve should have reached its final arborizationpattern (Kuratani and Tanaka 1990). The hearts werestudied with the HNK-1 antibody (Luider et al. 1993), animmunohistochemical marker that can be used to followneuronal differentiation.
Materials and methods
Fertilized white Lighorn chick eggs were incubated (blunt end up)at 38° C and staged according to Hamburger and Hamilton (1951).The material was subdivided into three groups: (1) embryos treat-ed with a solution of all-trans retinoic acid and dimethylsulphox-ide (n=11), (2), embryos treated with only the solvent dimethyl-suphoxide (sham-operated embryos, n=8); (3) control embryos(n=8). In an earlier study (Broekhuizen et al. 1992) we showedthat no differences were found between sham embryos in whichthe egg was merely opened and then closed without further treat-ment, and those that received the solvent.
At stage 15 (day 21/2 of a 21-day incubation period) each em-bryo, with the exception of the controls, was exposed by creating awindow both in the shell and in the overlying membranes. Either asolution containing 1.0 µg all-trans retinoic acid or only the sol-vent 2% dimethylsulphoxide was deposited on the vitelline mem-brane of the embryo, using a Hamilton syringe (Broekhuizen et al.1992). Subsequently, the window was sealed with tape and the eggreincubated.
At stage 34 (day 8 of incubation) the hearts of the embryoswere removed and subjected to whole-mount staining with HNK-1. To determine the presence of a cardiac anomaly, these heartswere serially sectioned for morphologic evaluation.
Immunohistochemistry on whole hearts
The hearts were perfusion-fixed with phosphate buffered 4% para-formaldehyde for 10 min. After 24 h immersion fixation in thesame fixative, the hearts were processed for whole-mount stainingwith HNK-1. The HNK-1 epitope is markedly encountered in dif-ferentiated neurons as well as transiently in other sites of the em-bryo. However, the differences in staining intensity, being less in-tense in the non-neuronal areas and cells, make it useful for thestudy of neuronal differentiation.
The hearts were first dehydrated in 70% through 100% alcoholand treated with a solution of 3% hydrogen peroxide (H2O2) in100% methanol to block endogenous peroxidase activity. The nextday the hearts were rehydrated and rinsed with phosphate-bufferedsaline (PBS). After washing in PBS containing 0.05% Tween-20(PBST) the hearts were incubated with the primary antibody dilut-ed 1/10 in PBST containing 1% Ovalbumine (PBST-OVA) over-night. The primary antibody was directed against the HNK-1 epi-tope (American type tissue connection, Abo and Balch 1981). Thespecimens were washed in PBS and PBST for several hours, andincubated with the secondary antibody, peroxidase-labelled rabbitanti-mouse (Dako P260) diluted 1/200 in PBST-OVA overnight.After washing in PBS and TRIS-maleate buffer the hearts werepreincubated in diaminobenzidine (DAB). After thorough wash-ing, the hearts were developed with DAB-H2O2, rinsed with PBSand stored alcohol in 70%. The hearts were examined under a dis-secting microscope; in particular, the arterial trunk and the base ofthe heart with the distribution of nerve branches were studied.
Morphology of whole-mount-stained hearts
To determine the morphology of the whole-mount-stained hearts,with emphasis on the presence or absence of a ventricular septaldefect, stained hearts were embedded in paraffin, and sectioned se-rially. The 5-µm-thick sections were counterstained with Mayer’shematoxylin/eosin. However, this material was not suitable tostudy the detailed nerve patterning in deeper layers, due to poorpenetration of antibodies. Therefore, a separate set of hearts wasdevided for direct HNK-1 staining in serial sections.
Immunohistochemistry in serial sections
To explore in depth the vagal distribution in the heart, a separateset of hearts of retinoic acid-treated embryos (n=5), sham-operated(n=4) and control embryos (n=5), was selected for serial section-ing and staining of the slides with HNK-1.
Embryos were fixed in ethanol-acetic acid for 24–72 h and em-bedded in paraffin. Serial sections of 5 µm were incubated in 1%H2O2/methanol to inhibit endogenous peroxides. Both incubationwith the HNK-1 antibody (undiluted) and the secondary rabbit-an-ti-mouse immunoglobulin conjugated with peroxidase, diluted 100times, were performed for 1 h at room temperature. The tissue sec-tions were stained with DAB-H2O2, rinsed with PBST for 5 min atroom temperature and briefly counterstained with Mayer’s hema-toxylin.
392
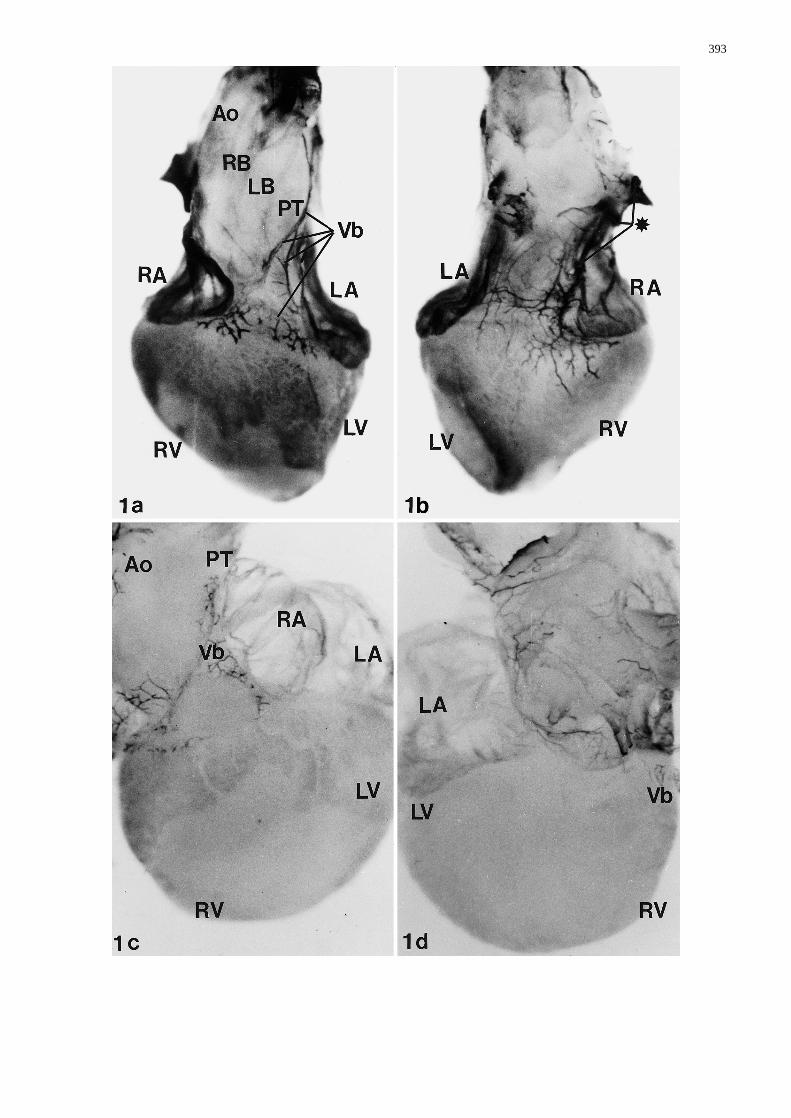
Fig. 1a–d Whole-mount staining with HNK-1 antibody. a Anteri-or view of a normal heart (stage 34). The great vessels are in nor-mal position. The pulmonary orifice is positioned in front of theaortic orifice. The ascending aorta is very short, the aortic archand the right and left branchiocephalic artery originating directlydistal to the aortic orifice. A superficial branch of the vagal nerve(Vb), descends laterally along the left side of the pulmonary trunk(PT). Nerve branches cover the shoulders of the ventricles (Vb),close to the atrioventricular sulcus (RB/LB right/left brachioce-phalic artery, RA/LA right/left atrium, RV/LV right/left ventricle,Ao aorta, PT pulmonary trunk). b Dorsal view of same heart as a.Two ganglia (*) can be distinguished from which cardiac branchesarise that communicate. c Anterior view of abnormal heart of astage 34 embryo after treatment with all-trans retinoic acid. Theaorta and brachiocephalic arteries are displaced to the right. Fur-thermore, there is a left juxtaposition of the atria. This heart wasdiagnosed as double outlet right ventricle. The vagal branch (Vb)over the pulmonary trunk (PT) is very thin and incomplete. Thedistribution over the ventricles is inconspicuous. d Dorsal view ofthe same heart as c. The thin branches (Vb) reduced in number arealso observed dorsally&/fig.c:
393

394

Results
Morphology of whole-mount-stained hearts
A study of sections showed no septation defects of theheart in 8 out of 11 embryos treated with all-trans retinoicacid. The remaining three hearts showed a rightward-posi-tioned arterial trunk in combination with a subaortic ven-tricular septal defect and was thus diagnosed as doubleoutlet right ventricle. All hearts of the controls (n=8) andfive hearts of sham-operated embryos were normal, whichincluded also the arterial trunk. Three hearts of the sham-operated embryos displayed a muscular ventricular septaldefect in combination with a normally positioned arterialtrunk.
Immunohistochemistry in whole hearts
Examining hearts of control and sham-operated embry-os, whole-mount-stained for HNK-1, revealed a compa-rable staining pattern. A superficial branch of the vagalnerve descends laterally along the left side of the pulmo-nary trunk to the base of the heart. Nerve branches crossthe atrioventricular sulcus and spread over the shouldersof the ventricles towards the ventral surface of the heart(Fig. 1a). The dorsal view (Fig. 1b) displays gangliafrom which cardiac branches arise and communicate,thus forming a nerve plexus that innervates the dorsal(posterior) surface of the ventricles.
Hearts of all-trans retinoic-acid-treated embryosshowed an abnormal branching. Laterally, the vagalbranch over the pulmonary trunk was very thin and in-conspicuous. The number of branches over the ventricleswas reduced. Moreover, the complexity of the networkwas less intricate than in normal hearts (Fig. 1a, c). Thereduction of branches was also seen dorsally (Fig. 1d).This impaired vagal distribution was seen in all embryostreated with all-trans retinoic acid regardless of the pres-ence of intracardiac malformations.
Immunohistochemistry in serial sections
HNK-1 immunoreactivity was observed in many regionsof the heart, including endocardium, epicardium and
valve tissue. In this study the neuronal distribution is de-scribed in the heart of five control and four sham-operat-ed embryos and of five retinoic- acid-treated embryos.Control and sham-operated embryos showed comparableHNK-1 positive neuronal cells and branches in the sub-epicardial mesenchyme and in particular the atrioventric-ular sulcus. Other branches penetrate the myocardiumand accompany the coronary arteries. Ganglia are foundat predetermined sites around the aorta and pulmonaryartery. Two large branches (dorsal and ventral) and onesuperficial smaller branch (left lateral) originate from thebranchial part of the vagus nerve and can be seen justabove the level of the semilunar valves (Fig. 2a). At thelevel of the aorto-pulmonary septum the branches fusetogether (Fig. 3a) and form a ventral network over theventricles. These data correlate and extend the whole-mount data presented earlier.
The hearts of three out of five retinoic-acid-treatedembryos were diagnosed as double outlet right ventricle.The other two hearts showed no septation defects. Allfive hearts showed a basically similar distribution ofHNK-1-positive neuronal cells. The staining pattern dif-fered from hearts of control and sham-operated embryos.Just above the level of the semilunar valves, branches(dorsal and ventral) could be distinguished (Fig. 2b).Contrary to the normal heart of a control embryo (Fig.3a), the branches do not connect at the level of the aorto-pulmonary septum (Fig. 3b). In addition the diameters ofthe vascular walls of the aorta and pulmonary artery areless than in the control and sham-operated embryos.
Discussion
The present study concerns the immunohistochemistry ofmyocardial vagal nerve innervation. The origin of the au-tonomic nervous system from the rhombencephalic neu-ral crest (Narayan and Narayan 1980) showed the originof the proximal ganglionic neurons and satellite cells inthe glossopharyngeal and vagal nerves. Neural crest ab-lation studies have revealed that the vagal innervation ismediated via the cardiac ganglia and can be identified inthe outflow tract of the heart at stage 27 (day 5 of incu-bation; Kirby et al. 1980; Kirby and Stewart 1983). Fur-thermore, the normal morphologic development of cardi-ac innervation by the vagal nerve (Kuratani and Tanaka1990) showed two categories in which the vagal nervecan be subdivided. The branchial part, consisting of thesuperior cardiac branch innervating the truncus arterio-sus of the heart, was also seen in our study of whole-mount-stained hearts of sham-operated and control em-bryos. Moreover, the intestinal portion of the vagusnerve was clearly seen in the separate set of stained seri-ally sectioned hearts of sham-operated and control em-bryos. In particular, the sinal branch innervating the dor-sal wall of the right atrium and ventricle is of impor-tance.
In our study, retinoic-acid-treated embryos showed anunderdevelopment of cardiac branches by an abnormal
395
Fig. 2 Serial sections of HNK-1 antibody-stained hearts; crosssection above level of semilunar valves of a normal heart (a) and aheart with double outlet right ventricle after retinoic acid treatment(b). a In this normal heart ganglia are found at predetermined sitesaround the aorta and pulmonary artery (dorsal, ventral, lateral). bAfter retinoic acid treatment only branches (Vb – dorsal, ventral)can be distinguished in this abnormal heart&/fig.c:
Fig. 3 Serial sections of HNK-1 antibody-stained hearts; crosssection at the level of the aorto-pulmonary septum of a normalheart (a) and a heart with double outlet right ventricle after retino-ic acid treatment (b). In this heart (b) there is no connection be-tween dorsal and ventral branches. In addition, the diameters ofthe vascular walls of the aorta and pulmonary artery are dimin-ished compared to (a) &/fig.c:

staining pattern and a reduction of vagal branches. Usinga similar whole-mount nerve-staining technique, Kura-tani et al. (1991) have encountered alterations in the pat-terning of cardiac nerve branches after ablation of thecardiac neural crest. The nerve trunks of the glossopha-ryngeal and vagus nerves were poorly developed and of-ten consisted of several thin bundles as in our retinoic-acid-treated embryos. Yet, what is the implication of thedisrupted vagal patterning seen after retinoic-acid treat-ment?
We are confronted with the unsolved problem thateven though a vagal branching network exists both in theretinoic-acid-treated and untreated embryos at stage 34,an active function has not yet been determined. In a pre-vious study altered hemodynamics were encountered instage 34 retinoic-acid-treated embryos (Broekhuizen etal. 1995).
A decrease in cardiac contraction force was found inthese embryos. Preliminary data in the stage 24 (day 4 ofincubation, before vagal innervation is seen) chick em-bryo displayed malfunction of the embryonic heart com-plementary with structural changes of the myocardiumafter retinoic acid (Broekhuizen 1996). Heart rate wasdecreased in these embryos, without compensatory in-crease of stroke volume, suggesting both pacemaker andcontractile dysfunction (Broekhuizen 1996).
Although the mechanism has yet to be resolved, it isproposed that retinoic acid has two effects on the devel-oping heart. An indirect effect of all-trans retinoic acidon cardiac function may be through an abnormal vagalinnervation after a disturbance of the migration of cellsfrom the neural crest. This is supported by our findingsof diminished vagal patterning. Recent experiments inwhich neural crest cell tracing and retinoic acid treat-ment are combined, point towards a diminished neuralcrest population after retinoic acid (R.E. Poelmann andA.C. Gittenberger-de Groot, unpublished results). Theeffect on the neural crest may be through cytotoxicity(Jelínek and Kistler 1981), or result from alterations inregion-specific signals necessary for homing and differ-entiation of neural crest cells (Hart et al. 1990). Our ex-periments cannot rule out a direct effect of retinoic acidon the differentiation of the cardiomyocytes and the sur-rounding interstitium. This is suggestive from work byPexieder et al. (1995), who showed that all-trans retinoicacid can modify cardiac contractility. Furthermore,Ostádalová et al. (1995) showed an inotropic effect of in-creasing concentration of Ca2 in the fetal rat heart withretinoic-acid-induced malformations. Wiens et al. (1992)found that retinoic acid has multiple effects on growthand differentiation of cardiac myocytes, including an in-hibition of cell proliferation, developmental of heart con-tractions, and delay in α-actin synthesis. An interactionof retinoic acid and contractile proteins could cause adisturbance in myofibrillogenesis or myofibril arrange-ment and result in myocardial dysfunction accompaniedby abnormal cardiac looping.
The dilemma of form and function relationships in anembryonic heart is difficult to address. Furthermore, it
has to be taken into account that embryonic functions areoften different from those in the neonate and adult. It isknown that the heart at an early stage can already reactto various substances that later in development are auto-nomic nervous system transmitters. Higgins and Pappano(1981) showed that as early as stage 24 the ventricle ofthe chick embryo responded to β-adrenergic agonistswith an increase in cardiac contraction force, before ad-renergic innervation occurs. Injection of a β-adrenergicagonist, isoproterenol, and epinephrine produces an in-crease in embryonic vascular resistance, presumably sec-ondary to an increase in vascular tone (Clark et al. 1985).Beta receptors are present in embryonic vessels and havea binding affinity to isoproterenol (Clark et al. 1984).Therefore, it cannot be excluded that the hemodynamicchanges seen after retinoic acid at stage 24 are in partvessel wall-mediated.
Despite the fact that until now the vagal innervationhas only been proven to be functional at day 12 of incu-bation (stage 38) there is no evidence yet available to ex-clude the possibility that vagal fibers seen first in the day4/5 (stage 25) embryo chick heart do not have a regulato-ry function. Pappano (1990) states that there is no evi-dence yet available to exclude the possibility that the va-gal fibers seen first at stage 25 are sensory in nature.
In conclusion, a difference in vagal nerve branchingover the hearts was found between control and sham-op-erated versus retinoic-acid-treated embryos at stage 34. Itis likely that the relevance of a disturbed vagal nerve dis-tribution of the embryonic heart after retinoic acid treat-ment is associated with altered cardiac function. Al-though we assume that several mechanisms provide anexplanation of cardiac dysfunction it appears that retino-ic acid affects both myocardial and vascular function andneural crest cell distribution simultaneously. Firstly, a di-rect effect of retinoic acid on the cardiac myocytes couldresult in myocardial dysfunction accompanied by abnor-mal cardiac looping culminating in morphologically ab-normal hearts. Secondly, a relationship is postulated be-tween impaired vagal branching pattern and an alteredfunction of the heart through a still unexplained impairedparasympathetic pathway, that needs confirmation by de-tection of functional neurotransmitter proteins and theirreceptors. Alternatively, disturbed sensory innervationcould already be functional in early embryonic stages.Thirdly, we suggest that a changed vascular tone shouldalso be considered. Research using an experimental set-up that combines blocking agents and functional studiesof the heart and vascular system is currently being car-ried out.
&p.2:Acknowledgements This study was supported by a grant fromthe Netherlands Heart Foundation and the Netherlands Organiza-tion for Scientific Research, grant 900-516-096.
396

References
Abo T, Balch CM (1981) A differentiation antigen of human NKand K cells identified by a monoclonal antibody (HNK-1). JImmunol 127:1024–1029
Bouman HCG, Broekhuizen MLA, Baasten MJ, Gittenberger-deGroot AC, Wenink ACG (1995) Spectrum of looping distur-bances in stage 34 chicken hearts after retinoic acid treatment.Anat Rec 243:101–108
Broekhuizen MLA (1996) Interaction between hemodynamics andmorphology in normal and abnormal cardiac development.PhD Thesis, Erasmus University, Rotterdam, The Netherlands
Broekhuizen MLA, Wladimiroff JW, Tibboel D, Poelmann RE,Wenink ACG, Gittenberger-de Groot AC (1992) Induction ofcardiac anomalies with all-trans retinoic acid in the chick em-bryo. Cardiol Young 2:311–317
Broekhuizen MLA, Bouman GA, Mast F, Mulder PGH, Gitten-berger-de Groot AC, Wladimiroff JW (1995) Hemodynamicchanges in HH stage 34 chick embryos after treatment withall-trans retinoic acid. Pediatr Res 38:342–348
Chambon P (1993) The molecular and genetic dissection of theretinoid signalling pathway. Gene 135:223–228
Clark EB, Feldman R, Hu N (1984) Paradoxical receptor mediatedvasoconstriction in the stage 24 chick embryo (abstract).Pediatr Res 18:121
Clark EB, Hu N, Dooley JB (1985) The effect of isoproterenol oncardiovascular function in the stage 24 chick embryo. Teratol-ogy 31:41–47
D’Amico-Martel A, Noden DM (1983) Contributions of placodaland neural crest cells to avian cranial peripheral ganglia. Am JAnat 166:445–468
Hamburger V, Hamilton HL (1951) A series of normal stages inthe development of the chick embryo. J Morphol 88:49–92
Hart RC, McCue PA, Ragland WL, Winn KJ, Unger ER (1990)Avian model for 13-cis retinoic acid embryopathy: demonstra-tion of neural crest related defects. Teratology 41:463–472
Higgins D, Pappano AJ (1979) A histochemical study of the on-togeny of catecholamine-containing axons in the chick embryoheart. J Mol Cell Cardiol 11:661–668
Higgins D, Pappano AJ (1981) Developmental changes in the sen-sitivity of the chick embryo ventricle to β-adrenergic agonistduring adrenergic innervation. Circ Res 245–253
Jelínek R, Kistler A (1981) Effect of retinoic acid upon the chickembryonic morphogenetic systems. The embryotoxicity range.Teratology 23:191–195
Kirby ML (1987) Cardiac morphogenesis−recent research advanc-es. Pediatr Res 21:219–224
Kirby ML, Stewart D (1983) Neural crest origin of cardiac gangli-on cells in the chick embryo: identification and extirpation.Dev Biol 97:433–443
Kirby ML, Waldo KL (1995) Neural crest and cardiovascular pat-terning. Circ Res 77:211–215
Kirby ML, McKenzie JW, Weidman TA (1980) Developing inner-vation of the chick heart: a histofluorescence and light micro-scopic study. Anat Rec 196:333–340
Kuratani S, Tanaka S (1990) Peripheral development of the avianvagus nerve with special reference to the morphological inner-vation of heart and lung. Anat Embryol 182:435–445
Kuratani SC, Miyagawa-Tomita S, Kirby ML (1991) Developmentof cranial nerves in the chick embryo with special reference tothe alterations of cardiac branches after ablation of the cardiacneural crest. Anat Embryol 183:501–514
Lammer EJ, Chen DT, Hoa RM, Agnish WD, Benke PJ, Curry CJ,Fernhoff PM, Grix AW, Lott IT, Richard JM, Sun SC (1985)Retinoic acid embryopathy. N Engl J Med 313:837–841
Lee YM, Osumi-Yamashita N, Ninomiya Y, Moon CK, Erikson U,Eto K (1995) Retinoic acid stage-dependently alters the migra-tion pattern and identity of hindbrain neural crest cells. Devel-opment 121:825–837
Luider TM, Bravenboer N, Meijers C, Kamp AWM van der, Ti-bboel D (1993) The distribution and characterization of HNK-1 antigens in the developing heart. Anat Embryol 188:307–316
Narayanan CH, Narayanan Y (1980) Neural crest and placodalcontribution in the development of the glossopharyngeal-vagalcomplex in the chick. Anat Rec 196:71–82
Ostádalová I, Pexieder T, Ostádal B, Kolár F (1995) Inotropic ef-fect of increasing concentration of Ca2+ in the fetal rat heartwith retinoic acid-induced malformations. Pediatr Res 38:892–895
Pappano AJ (1990) Development of cholinergic neuroeffectortransmission in the avian heart. Implications for regulatorymechanisms. Ann N Y Acad Sci 588:131–136
Pexieder T, Pfizenmaier Rousseil M, Prados-Frutos JC (1992) Pre-natal pathogenesis of the transposition of great arteries. In:Vogel M, Bühlmeyer K (eds) Transpostion of the great arteries25 years after Rashkind Baloon Septostomy. Steinkopff,Darmstadt, pp 11-27
Pexieder T, Blanc O, Pelouch V, Ostádalová M, Milerová M,Ostádal B (1995) Late fetal development of retinoic acid in-duced transposition of great arteries: morphology, physiologyand biochemistry. In: Clark EB, Markwald RR, Takao A (eds)Developmental mechanisms of heart disease. Futura, Armark,NY, pp 297–307
Taylor IM (1981) Some early effects of retinoic acid on the younghamster heart. In: Pexieder T (ed) Mechanisms of cardiac mor-phogenesis and teratogenesis. Raven Press, New York, pp151–161
Walkey GK, Bower AJ (1981) The distal vagal ganglion of the hen(Gallus domesticus). A histological and physiological study. JAnat 132:95–115
Wiens DJ, Mann TK, Fedderson DE, Rathmell WK, Franck BH(1992) Early heart development in the chick embryo: effects ofisoretinoin on cell proliferation, α-actin synthesis, and devel-opment of contractions. Differentiation 51:105–112
Yasui H, Nakazaw M, Morishima M, Miyagawa-Tomita S, Mom-ma K (1995) Morphological observations on the pathogeneticprocess of transposition of the great arteries induced by retino-ic acid in mice. Circulation 91:2478–2486
397
Related Documents











