THr SourHwEsrERN Nrrunel¡sr 38(2):127 -135 Juxe 1993 DISTRIBUTION AND ABUNDANCE OF COUES DEER AND CATTLE IN MICHILIA BIOSPHERE RESERVE, MEXICO C¡,n¡,os GeltNoo-LEAL, ANGEL¿s Mon¡¡.¡s G nNo M¡,NuEL WEBE,R R. Centre Jor Applied Conseruatíon Biology, Facttlty ol Forestry,2357 Main Mall, Uniuersity ol Brittsh Columbia, Vancouuer, B. C., Canada, V6T-l24 (CG), Ins'ituto de Eatogía a'c "';';i,!.:;:,"í:;f{;:::';í;t,\;ííf,"'Apdo posrar612, A¡srn¡cr-Patterns of habitat usc and relative abundancc by Coues whire-tailed deer (Odocoileus uirginianus couesi) and cattle wcre assessed by pellet group and dropping counrs in and around l\{ichilia Biosphere Rcserve, Durango, Mexico. Decr and cattle use habitats more than expncted by chance only in sites where either specics was more abundant. Both spccics showed high habitar breadrh and consequently had high habitat overlap. Deer abundance, as indicated by deer pcllet-groups, was higher in some parts of the Reserve, particularly in the Reserve Core. Cattle abundance, as indicared by droppings, was high outside and in some si¡es inside the Reserve. Lowest cattle abundance was recorded in the Reserve Core. Numbers of deer pellet-groups and cattle droppings were negarively correlated. The data suggest that ovcrgrazing by cattle and hunting strongl.v influence distriburion and abundance of deer. R¿sutr¿BN-Efectuamos una evaluación de los patrones de uso de habitat y abundancia relariva por el venado de Coucs y cl ganado vacuno dentro y ccrca de la Reserva de la Biósfera de la Michiiía, Durango México, a través del contco de excrementos. Se documentaron preferencias de habitar sólo en los sitios con mas abundancia de venado o de ganado. Documentamos amplitud de habitar aha para ambas cspecies y consecuentemente hubo una sobreposición de habitats alta. La abundancia de .venados, dcterminada a través de los grupos de excrementos, fué mayor en algunas partes de la Reserva, particularmente en la zbna núcleo. La abundancia de ganado fuó alta tanto fuera como dentro de la Reserva. La abundancia mas baja de ganado se registró en la zona núcleo. Encontramos una correlación negativa cntrc los excrcmcntos de ganado y los de venado. Los patrones documentados sugieren que el sobrepastorco del ganado y la cacería, cstán influenciando fuertemente la distribución y abundancia dcl vcnado de Coues. A main objeaive of a biological reserve (i.e., national park, sanctuary, biosphüre reserve) is to protect and conserve the biological diversity of a region. Early management .policies in reserves included avoiding direct intervention or control- ling the action of factors perceived as negative (i.e., fire and predators). However, several studies have shown that to cope with the dynamic state of natural populations and communities, constant monitoring and an active approach to managing unexpected changes are required. Thus, biolog- ical reserves can be used as valuable tools to com- pare the behavior of managed and unmanaged biological systems. Michilia Biosphere Reserve was established in 1975 in the Western Sierra Madre, Durango. Mexico (Fig. 1). The reserve is located in rhe southern portion of the distributional range of Coues deer, the most widespread subspecies ol white-tailed deer in Mexico. Coues deer inhabir the Western Sierra Madre from Arizona and Ner+, Mexico to northern Jalisco. Because of the xeric conditions of this area, this subspecies seems ro be more vulnerable to droughts and to overgraz- ing by cattle than other subspecies (Anthony, 1 976; Anthony and Smith, 1977; Brown and Henrv. 1981';.j¡own,1984; Smith, 1984; H. L. Shorr, in litt.). There have been reductions in local dis-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THr SourHwEsrERN Nrrunel¡sr 38(2):127 -135 Juxe 1993
DISTRIBUTION AND ABUNDANCE OF COUES DEERAND CATTLE IN MICHILIA BIOSPHERE
RESERVE, MEXICO
C¡,n¡,os GeltNoo-LEAL, ANGEL¿s Mon¡¡.¡s GnNo M¡,NuEL WEBE,R R.
Centre Jor Applied Conseruatíon Biology, Facttlty ol Forestry,2357 Main Mall,Uniuersity ol Brittsh Columbia, Vancouuer, B. C., Canada, V6T-l24 (CG),
Ins'ituto de Eatogía a'c
"';';i,!.:;:,"í:;f{;:::';í;t,\;ííf,"'Apdo posrar612,
A¡srn¡cr-Patterns of habitat usc and relative abundancc by Coues whire-tailed deer (Odocoileusuirginianus couesi) and cattle wcre assessed by pellet group and dropping counrs in and around l\{ichiliaBiosphere Rcserve, Durango, Mexico. Decr and cattle use habitats more than expncted by chanceonly in sites where either specics was more abundant. Both spccics showed high habitar breadrh andconsequently had high habitat overlap. Deer abundance, as indicated by deer pcllet-groups, was higherin some parts of the Reserve, particularly in the Reserve Core. Cattle abundance, as indicared bydroppings, was high outside and in some si¡es inside the Reserve. Lowest cattle abundance was recordedin the Reserve Core. Numbers of deer pellet-groups and cattle droppings were negarively correlated.The data suggest that ovcrgrazing by cattle and hunting strongl.v influence distriburion and abundanceof deer.
R¿sutr¿BN-Efectuamos una evaluación de los patrones de uso de habitat y abundancia relariva porel venado de Coucs y cl ganado vacuno dentro y ccrca de la Reserva de la Biósfera de la Michiiía,Durango México, a través del contco de excrementos. Se documentaron preferencias de habitar sóloen los sitios con mas abundancia de venado o de ganado. Documentamos amplitud de habitar ahapara ambas cspecies y consecuentemente hubo una sobreposición de habitats alta. La abundancia de.venados, dcterminada a través de los grupos de excrementos, fué mayor en algunas partes de la Reserva,particularmente en la zbna núcleo. La abundancia de ganado fuó alta tanto fuera como dentro de laReserva. La abundancia mas baja de ganado se registró en la zona núcleo. Encontramos una correlaciónnegativa cntrc los excrcmcntos de ganado y los de venado. Los patrones documentados sugieren queel sobrepastorco del ganado y la cacería, cstán influenciando fuertemente la distribución y abundanciadcl vcnado de Coues.
A main objeaive of a biological reserve (i.e.,national park, sanctuary, biosphüre reserve) is toprotect and conserve the biological diversity of aregion. Early management .policies in reservesincluded avoiding direct intervention or control-ling the action of factors perceived as negative(i.e., fire and predators). However, several studieshave shown that to cope with the dynamic stateof natural populations and communities, constantmonitoring and an active approach to managingunexpected changes are required. Thus, biolog-ical reserves can be used as valuable tools to com-pare the behavior of managed and unmanagedbiological systems.
Michilia Biosphere Reserve was established in1975 in the Western Sierra Madre, Durango.Mexico (Fig. 1). The reserve is located in rhesouthern portion of the distributional range ofCoues deer, the most widespread subspecies olwhite-tailed deer in Mexico. Coues deer inhabirthe Western Sierra Madre from Arizona and Ner+,Mexico to northern Jalisco. Because of the xericconditions of this area, this subspecies seems robe more vulnerable to droughts and to overgraz-ing by cattle than other subspecies (Anthony, 1 976;Anthony and Smith, 1977; Brown and Henrv.1981';.j¡own,1984; Smith, 1984; H. L. Shorr,in litt.). There have been reductions in local dis-

[llr*'',: ¡l¿:t:ii.
128Th¡ Sout hu,e slern l\' aluroi t sl vol J¡. n,, -
Michilia BiosphereReserve,Durango, Mexico
SAMPLING AREAS
l"o" AN ¡t"t' ñ-
f] xovrlrn¿n..- RESEN\¡E BOL\DAR}
- SECOI{DAR]' ROAD
Ftc. 1-Map of the stud; area in Durango. N4exico showing the location of sampling areas in \,lichili;,Biosphere Reserve and vicinity. Areas were sampled during May (FL, PLl ,CL1 ,CLz, PL2. and PL3), Junc(CL3, CL4, CL5, and CL6), and November 1987, (CL7, CL8, CL9, and CL10). At least 24() plots rr'eresampled in each area. FL : Federal Land (Reserve core); PL : Private Land. CL : C)ommunal Land.
tribution (Anthony and Smith, 1977), populationdeclines (Anthony, 197ó; Brown and Henrl', 19$l ;
H. L. Short, in litt.), and local extinctions (Brou'nand Henry, 1981).
Beginning in the 1950s, research has shou'nrhat deer in Mexico were becoming rare in placeswhere they were once abundant (Baker, 1958;Leopold,1959; Baker and Greer.1962). Al-though Coues deer have a wide distribution inMexico. there is an absolute lack of informationon their status, and, like many other Ntlexicansubspecies, rhey are iikely endanga"ed (Galindo-Leal, 1992¡.
N{ichilia Reserve provided an opportunitv toexamine the impact ol human activities on Couesdeer populations. Managemenl practices (i.e..livestock production, logging, and hunting), varywidely in the types of land tenure that constirur€the reserve, ranging from almost no human ac-
tiviries in the Reserve Core to heavy livestockgrazing and selective logging in other areas.
We had two objectives: to assess the status o[Coues deer populations in Michilia BiosphereReserve and neighboring areas. and to comparerhe patrerns of deer distribution and abunciancewrth those ol cattle in areas thar differ in theircatrle management prac'lices and srocking rares.
Sruov Aan¡ ¡No Mrruoos-N4ichilia Brosphere
Reserve (MAB UNESCIO) (23"1 0'-3Lr'N. 1C).1"05'
20'W) encompasses 420 km:rof dr)' and tenrperate oak-pine forest in the \\'estern Sierra N{adrr in southe:'¡Durango. Mexico. Aititude ranges from 2.20(r to 2.85(i
m. Annual precipiraLion averages from 5(l to 70 cnr.
concentrated between June and Ocrober. N4ean renr-
peratures fluctuare from 17.4 to 20.4'C. The reserveincludes three different types ol'land tenure: Federal(FL), Communal (Ejidos: CL), and Private land (PL)Neighboring ranches are communal or private. Themain vegetation type of the area is dry temperare oak-pine forest. We distinguished four major veeetaiir,tassociations or habirat ttpes: oak foresi. pinr fc,re:,:.
rnanzani¡a (.4rctostaph",,b.t pungens) sl-rrubland. and
grassland. Oak foresr u'as subdivided inio pure oak.
oakjuniper. oak-prne. and oak-manzanira. I'rnc íoresr
was subdivided i¡it¡r pine-oak and pure pine srands
\Ve surveyed deer pellet-groups and carrlc ciro¡'-pines. During N{av 1987. u,e sampled (rr = 1.44()
sampling units) six areas: the Rescrve Core (FL). rhree
in the buffer area (PL1, CL1, and CL2), and th'o area-<
in the vicinity (PL2 and PL3), The second and thirdsurvevs were concentrared in Communal Land. Thertook plac¿durjng Julv (CL3 CL4. CL5. and CLó: r: 1.240 sampling units) and November íClL- CL8.
CL9. CLI0: ¡; : 1 .24A sarnpling unirs. Fis. I i. Be caust
ol the iou' cielrsi¡ies recorded ¡n the ia,.; i\'.r survc\:most of rhe analvs!s included iníc¡rna;iolr ¡r¡l¿tt¡rei ;1,
4A+ffJIIE I
\ó)PL¡ r/ rclto
.h\ {
St' \----r-----19E
|_.--0 5 ¡Obn
Junc 1993
FL PLT CLl CLz PLz PL3
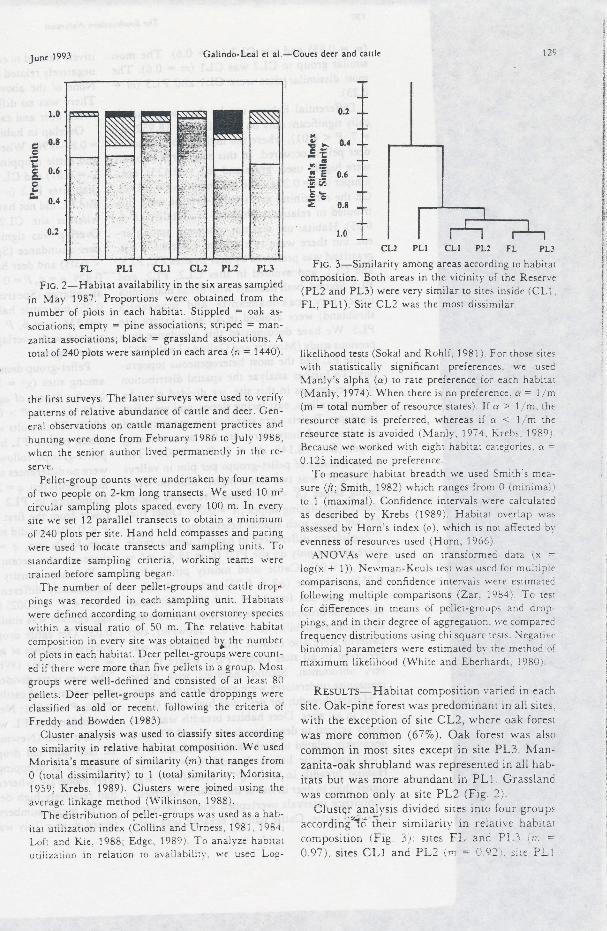
Frc. 2-Habitat availability in the six areas sampled
in May 1987. Proportions were obuined from the
number of plots in each habitat. Stippled = oak as-
sociarions; empty : pine associations; striped : man-zanita associations; black = grassland associalions .{roral of 240 plots were sampled in each area (n : t 440).
the first surveys. The latrer surveys were used to verify
patterns of relative abundance of catrle and deer- Gen-
eral observarions on cattle management Practices and
hunting were done from February 1986 to Julv 1988,
when the senior author lived permanently in the re-
tt?1i,.,-*.ouo counts were undertaken by four teams
of two people on 2-km long transects. We used 10 micircular sampling plots spaced every 100 m. In every
sire u'e set 12 parallel transects to obrain a minimumof 240 plors per site. Hand held compasses and pacing
were used ro locate transeds and sampling unils. Tc,
standardize sampling criteria. working leams were
¡rained before sampling began.
The number of deer peller-groups and cattle droFF
pings rvas recorded in each sampling unit. Habitatswere deñned according to dominant oversrorey species
wirhin a visual ratio of .50 m. The relarive habitatcomposition in everv sire was oblained b¡ the numberof plors in each habirat. Deer pellet-grouPs were count-
ed if there were more than five pellets in a group. Mosrgroups were well-defined and consisted of at least 80
pellers. Deer pellet-groups and cattle droppings u'ere
classified as old or recent. follos¡ing the criteria of
Freddy and Bowden (1983).
Cluster analysis was used to classify sites according
to similarity in rclative habitat composirion. We used
Morisita's measure of similarity (m) that ranges from0 (rotal dissimilarity) to I (total similarity; Morisira,1959; Krebs, 1989). Clusters were joined using the
average }inkage method (\\¡ilkinson, 1988).
The distribution of pellet-groups r+'as used as a hab-
i¡ar utiiization index (Coliins and tlrness, 1981, 1984r
Lofi and Kie. 1986: Edge. 1989). To anaivze habirat
uriiizarion in reiarion to availabiliir'. u'e useci Log-
Galindo-Lcal et al.-Coucs deer and cattlc 129
1.0
xi¡ !\
IEtrba-!s¿
- 0.tcb o.e
& o.l
cL2 PLI CLI PLz FL
F¡c. 3-Similarity among areas according ro habitarcomposirion. Both areas in the vicinity of rhe Reserve(PL2 and PL3) were very similar to sites inside (CL1,FL, PL1). Site CL2 was the most dissimilar.
likelihood tesrs (Sokal and Rohlf. I 981 ). For those sites
with statistically significant preferences. we used
Manly's alpha (a) to rate preferencc for each habitat(Manly, 1974). When there is no preference. a = 1/m(m = total number of resource srates). If a > 1rlm. theresource state is preferred. whereas if a < 1/m the
resource state is avoided (Manly, 1974; K¡ebs, 1989).
Because we u'orked with eight habitat categories. a =0.125 indicated no preference.
To measure habitat breadth we used Smith's mea-
sure (/; Smith, 1982) u'hich ranges from 0 (minimal)to 1 (maximal). Confidence intervals r+'ere calculatcdas described by Krebs (1989). Habitat overlap was
assessed by Horn's index (o), which is not affected b¡'
evenness of resources used (Horn, 1966).
ANOVAs were used on transformeii data 1x :log(x + 1)). Newman-Keuis test was used for mulriplecomparisons, and confrdence intervals were esrtmated
following multiple comparisons (Zar. 1984). To tesr
for differences in means of pellet-groups and drop-pings, and in their degree of aggregation. u,e compared
frequency distributions using chi square tests. .i.\eqativc
binomial parameters were estimated bv the merhod of
maximum likelihood (White and Eberhardt. 1980).
Rr,sulrs-Habitat composition varied in each
site. Oak-pine forest r+'as predominant in all sites.
with the exception of site CL2, where oak forestwas more common (677"). Oak forest *'as also
common in most sites except in site PL3. Man-zanita-oak shrubiand was rePresented in all hab-itats but was more abundant in PLl. Grassland\e'as common onl-v at site PLZ (Fig. 2).
Clustgr analysis divided sites inlo four groups
accordingld iheir similariti in relaiive habrrat
composirion (Frg. 3;: srtes FL and PL3 (n: :0.97). sites CLl and PL2 (n¡ : 0.c)2). si'.e PLi
PL3
tiI¡t

130The Soulhwestcrn N oturajist vol. -18. no.2
(2 : 0'85) and site cLZ (n - 0'ó)' The most itivelv related ro carlle abundancr (r: tl.7ó). anclsimilar group ro CL2 was CLl (n: 0.6). The ".g^;;;;;i",la,o¿..rabundance (¡ : _0.-i.lr
illl,,ot"t-ttar sites were cLt and pL3 (m = None orin. "0.".".""-.i#i;i"lT."rlr,o."",
Diflcrentiar habirar use by deer was sratisri- *x;:::;oo:flL*.. in habitar ur.'"atn t.-
cally significant only on site FL (G: rg.952,dJ. ouerlap in'iut,ru, use was high in alr sjres (o: 7 ' P < 0'01)' where the highest abundance of = o.a+-o.boj. w;h.n o'ry recenr deer p,e¡er-gr.upsdeer peliets occu'ed In this site, three habirars and cartle droppings were considered. sires pL1were highly used: grassland (a1 : 0.423), o_ak- (o:0.67)^"á'éL¡ (o : 0.75) had ic,*.e,. oue.rapjuniper (a2:0.2Aa), and oak forest (a3 : Oi.llZt thu¡, ,1,., ,ii-t, = 0.80) and pL¡ (¿ : 0.89)on the remaining areas pellet-groups were dis- site FL did no, hru. uny recenr carrie ciroppines.tributed in relation to habitat a-vailability (P > whercas ,ir. clz racked recenr deer perets0'05)' Habitat use was not consistent
"-áng ur- overlap was significantly correlated r'ith borheas, bur there wcre two evident put,a.rr.. ñir.r, d.., ubund,.o.r.."isp.urr.run r:0.913. n : 6. J,grassland was preferred by deer insites FL, pL1, < 0.05) ;ril..r';:;;;",^u.."0,n'tÁ;:u;*o" , :and clr, and avoided in sites cL2, pL2, and 0.g2g,;:6,i:0.05). in conrr-asr. neirher carrrePL3. Second, both habitats wirh densem anzunjtu abundance (Spearman ¡ : _0.31 4. n : 6, p .>understory (oak-manzanita and manzanira 0.05) nor .u.,i.i"¡iru, breadrh (Spearman r :shrubland) were avoided in ajl sites except site _0.334,, : 6,¡ , O.OS) were posirir,el' c.r--PL3' we have documented this auoidance in a reiared with overlap. In fact. borh relarionshipsprevious study (Morales and Galindo _Leal,1()g7). *.r. n.gutiu..
--
site FL had the most htteroge::ous ropogra- p.¡¡sllgroup density was significantrv difl-erenrphy. To further analyze the spátiat distrituiion among ri;Jü.-: 20.22. d.f. : 4, p < 0.r)05).of pellet-groups in this site, *t ai"¡¿.d-pl"rr uy !1t trrl j.g..J.r
aggregation u.as similar (x: :location: valleys, east facing slopes, and *err_fa.- 0.7g, d.f. J +, p > 0.05) (Whitc and Eberhardi.ing slopes Pelle¡s were significantlv concenrrated t9a0). "The
,.*jt, *... similar lor borh olci ancjin valleys and there were very feu'on east-facing recenr peliers. FL had the hrghesr abundance o1slopes (G: t8'148' dr' : z, p < 0'001). Th; p.il.,-g.o;p;, to,,.*,.. bv plr and c,-l..rherem-ean number of peilet-groups per plor in valleys were no differences among these three sires (Ne*,,(X:0.47) was two times higúer thun on *.rr_ man_Keulsres¡,g: 2.51 ,p> 0.0-5) pelier-groupfacing slopes (X : O.Z4)._"IJ !". times higher abundance ;n 'ítZ
and pL3 u,as significanrirthan on east-facing slopes (x : 0.09). orhe. sites lower from the firsr group (ANo\;A. ¡ : 9.0:.did nor show such spatiar heterogeneitl',' p < 0.001) but did nor difler among rhem (Nern.-Significant differences r1,laUr_tat use by s¿11¡s *^r-K;;; ;;r;,;: 0.85, p > 0.0s) Finalr.were evident only on site PL2 (G : rc'41 ' d.t. : there were no ..".n, deer peller-gr.ups rrr cL2.1 , P < 0.05). Cattle used mostli, pine 1ai : The mean densirl. ol cattle d.roppings r\,as ais(,0'258), oak-pine (a2:0.23), grassrand ia3 : ,ig"in.r;,|; d'i'u;;r, among sites (y: : 21.8i.0.226), and oak foresr (a4 : O.lpl). On other d.f : 4.¡ I O.OOZI (Fig. a). and therr desree c,fsites'cartledroppingsweredisrributeciinrelarion uggr.gu,ion*lr"r*tt"r(x-:7.3r,a¡:4.¡t;,ro habi¡ar availabirit.v (p > 0.05). However, tu,o o.os).-cll¿ .utu. a.oppi,.,i, ,ir,,,.,.j ,,*_,.r'.u",,,parrer-ns were arso evicienr. Flrst, oek foresr *,as differenr degrees or ugg..g^,ion (x: : 12.2<). ¿.ipreferred in ail sites excepr pL3, where ir was : 5, p < oi.o¡) (white anci Eberhardr. 19g0)very uncommon' second, habitats with dense The sites *,er" diuided inro three groups accorci-manzanita understorey were avoided in mosr sites. ing ro densiry of cattle droppings (ANO\;A. ,tr :Habitat breadth for deer was hígh (Ít: 0.87_ i¡laJ, p .^iio"ó,,T.*_^.,_Keuis tesr). I)ensirv0'97) in all sites' Deer habitat ¡teaarn was pos- of droppings in FL was significantr' iou,e¡ rhanirively related ro deer ab.undance (r:0.71)'and in or¡er.ii., 1N.*-un_Keurs rests. p < 0.05).negatively related to both cartle abundance (r : In facr, orrty ota affi;;;.r. ;;"rO ,"'' O,--0.6) and catrle habitat breadth lr = _o i?r
Habitat breadth i, ;";'. üit1 ;; t ;l lrl 3:HT:.:i,3::l'x;:..Sffi ??"J,,r, Fijhigher than in deer in five of the six sites ruápr.á. had rerariver' high densities.confidence intervals overlapped widely beri"een The me¿n ¡¡rmbers of deer peile¡-gr<;ups ob-deer anrj cattie. cattie habirát breadrh was po.s- ta.ined during Jurl u,as i).0: (cr i * ¡r.¡r1, ,, :

June I 993
g 0.1
¿€oJE
Eo:ü!9 o.l
Galindo-Lcal cr al.*Coucs deer and c¿ttlc
T,r¡lr l-Results of Pearson corrclations betu,eendcer pcllcr-groups and cattle droppings in the rhreesampling pcriods. Recent and old pellets and droppingswcre analyzed scparately.
Cattle DeerAII PL3sites excluded
Muy
July
Novembcr
recenl
oldrecent
oldrecent
old
recent -0.33 -0.52old -0.71 -0.90*
recent -0-99*old -0.24
recent -0.75old 0.12F¡c. 4-Deer pellet-group densiry in sites with dif_
ferenr densities of cattle droppings. All peller_groupsand droppings werc used regardless of age, FL = Fed_eral Land (Reserve core); PL : private Land, CL =Communal Land. A minimum of 240 plots were sam_pled in each location.
1240). During November, the mean number ofpellet-groups was 0.02 (C.L + 0.009, n : 1f,4}).Neither the means, nor the degree of aggregationof these two surveys were significantly different(x'? : 0.+3, dl = Z, P > 0.05). Borh estimareswere similar to densities documented in May insites CL2, PLZ and PL3. During July, peller-group densities were higher in sites CL3 (X :0.03, + 0.023, n : 520) and CL6 1.i : o.Or +0.03, n : 240) These two sires were in relativeproximity. In site CL5 (n: 2ae only two peller_groups were found and none were found al CL4ft = 24A),In November, the highest density wasfound at CL-9 (N : 0.04, + 0.035, n = 240).Sites CL7 (f, : 0.02, n : 280) and CL8 (X :0.01, n : 480) had intermediate densities. Fi_nally, in site CL10 only one pellet-group waslound (n :24A).
The mean numbers of deer pellet-grc,bps andcattle droppings were negatively relared. This re_lationship was evident during the three surveys(Ma1', July, and November). Correlation, *...statistically significant during May when site pL3was excluded and during July (Table 1).
Pellet-groups were clumped in all sites (all X?values ) 290.0, dl. - 239, P < 0.01) excepr pL3where thev were randomly distributed (Xr:237.0.dJ. : 239, P > 0.05). Degree of aggregation (sz¡;.l') was sronglv correlated wirh the mean densityof recent peller-groups (Spearman r = 1.0, n :5, P: 0.01).
Cartie droppings were clumped in all sites (all
1' 'alu.r > 290.0. d.í. : 239, P < 0.01) exceprCLt ix: = 226.0. cil : 239, P > 0.05) There
r P < 0.05.
was no relationship between degree of aggrega_tion of recent cattle droppings and thcii mJ.ndensity (Spearman r : 0.308, n : S, p > 0.05).
D¡scussloN-We will ñrst examine the doc_umented parterns of abundance and d.istributionof Coues deer, and then presenr some porenrialexplanations to account for them.
Habitat Composition-The classification ofhabitat composition by cluster analysis closell,matched the geographical and aititudinal distri_bution of sites. Sites FL and pL3 were borh athigh altitudes and relatively close ro each orher.Sites CL1 andPL2 were both ar high ro mocierarealtitudes and were also ciosc to each other. SirePL1 was located in a moderale to high alrirudeand was close to FL and CL1. Sire CL2 r+,as arthe lowest altitude. Its closest neighboring sire'was CLl (Fig. 1). Habitat.ornporiiion folüweda gradient of precipitation and temperarure.where, in general, oak forest occupied lou,er areaswith lower precipirarion and higher rempera_turcs, and pine forests were found ar higher alti_tudes.
Habitar preferences by deer were significanronly in site FL, where rhe highesr densities ofpeliets were found. Oak-pine foresr \^,as pre-ferred, and pine-oak forest was avoided. Brown(1984) found that Coues deer in Arizona alsopreferred oak forests over coniferous foresrs. Fur-ther anaiysis on this site showed that peller-groupswere concenrrated primarily in valleys and sec_ondarily in west-facing slopes. They were almosrabsent from gFl,l,-J4rng slopes, which had a uni-queand strikinglv differenr plan', species composirronand physiognomr'. Dominanr species ori east-f al-ing slopes were Ptnu.c lumix¡Lrzti. ()uercu: ur¡)ct!..(). crr¡ssi..ícitc. ano Aracsi.aphyict: l¡oii¡'oi.tt;. Th;:

r32
plant association is patchy in places with littlesoil and almosr no ground vegerarion. Therefore ,
in this site (FL), deer distribution was patchy aswcll. They were highly abundanr in some habi-tats and almost absent in others.
Wc did nor find significant habitat preferencesin the remaining sites. Low dcnsities in otherareas may obscure habitat preferences, however;trcnds indicated that manzanita shrubland wasavoided in all sites excepr PL3. Manzanita shrub-.land provides potential prorection, but often is toodense and may restrain movements. In addition,this habitar provides almost no other food besidesmanzanita. Cattle also avoided manzanita shrub-land.
Borh deer and cattle preferred grassland andoak forest to pine forest and manzanita shrub-land. Deer, however, preferred grassland in siteswhere cattle were ieast abundant, and avoided irr+'here cattle were mosl abundant. The oniy ex-ception was site PL3. Here. deer avoided grass-land in the absence of cattle. We believe tharavoidance of grassland in this sire u.as due mainll.to high hunting pressure in this unprotected ranch(Galindo-Leal, pers. obser.). Two further obser-vations support this explanation. Firs¡, this wasthe only site where deer preferred manzanitashrubland, which provides porenrial protectivecover. Second, deer were very scarce in this sire,likely due to the high hunting pressure.
Habitat Breadth and Ouerlap-High values olhabitat breadth, both for deer and cartje, indicaredthat both species use mosl habitats extensivelywith a resuiting high overlap. Relationshipsamong habitar overlap, habitar breadth, andabundance differed for both species. For deer, allthree measures of habitar use were positively cor-relared. For cattle, habitat bread¡h and abun-dance were positively correlated. However, hab-itat overlap was not correlated with either.
Abundance-The three sites with higher pellet-group densities were all inside the reserve (FL.PL1, and CL1). Densities were from four to seventirnes higher than ourside. These sites have dif-ferent types of iand tenure. Site FL is federalland and has been granted protection throughinaccessibility and vigilance. Cattie have beenpartially excluded since 1975 and more acrivelysince 198ó. ln sites PL1 (private land) and CL1(communal land). cattle are rotated seasonall¡..reducing overgrazing. Inside the reserve, all si¡esr¡,ilh extremelv ]orr densiries of cieer weie coni-munai ianci (CLz. CL4, CL5. and CLICI). Ir,
The Sout hwestern Natu¡a!íst vol. 38, nr, l
some of these sites (i.e., CL2), stocking rarcs arerelatively high and cartle are kepr in large lencedareas throughout the year, resultinq in exrremeovergrazing (Galindo-Leal, pers. obser.). Grassesand forbs are almost absent and there is lirtlecover in the undcrstory. Site CL2 had the lou,esrdeer abundance.
Outside the reserve, in site PL2 (private land).more attention bv land-or.r.ners is given to cartlemanagement. Rotation of cattle and recenr con-strucion of drinking rroughs have benefiteci bothcattle and deer. In contrast, site PL3 is an aban-doned ranch where very fell cattie and cieer werefound. Lack of vigilance in this s¡te resulrs inillegal hunting. Finally. there were no sires u,hereboth deer and cattle were a1 high densrrres.
Patterns of distribution and abundance of deerdocumented in this study seem to be largeiv de-termined by three main factors. These facrr,rs arenot mutually exclusive and are onl1. proposedhere as hypotheses ro be tesied rn future rn'ork.
Habítat-Similarities in habirar compositionamong sites reffected habirat changes along analtitude gradienr. In the lorvesr altitude sjre (CL2)(2,200-2,300 m), oak íorest. (Quercus gn:ea) con-stituted a verv large component. rr,hereas borhpine-oak and oak-pine forests rtere not verv u'eiirepresented. In higher altitude sires (FL, PL1.PLZ) (2,400-2,800 m), oak-pine and pine-oakforests are betrer represented. This habitar gra-dient seems determined by increasing rainfall anddecreasing temperarur€ with higher ahirudeSimilar altitudinal gradients in vegetation largeir,determined by rainfall and temperature are com-mon in manl' desert mounrain ranges in thesouthwestern United States (Shreve, j915) andnorthern Mexico. Therefore, high and moderarcaltitudes likely are more produc¡ive habiLar lorwhite-taiied deer (Knipe. 1977).
CattLe Grazing-There are large differences rncattle managemenr policies in the sires. dependingon land tenure and affluence. The olr'ners of pri-vale ranches (PL1, PL2) do nor rel-r. on carileproduction as rheir main source of income. Ther,also provide more drinking faciliries and suppl,food during winter. ln con¡rast. for Ejidararic_rs(owners of communal iand), the number of cartleowned by an individual represenrs economit se-curitv and status, and is the main sourcc of in-come. e¿t¡le<,n communal F,iido lands (CLl - 1C\are provicied n'rih ieu'er cirinking iaciiriie. ancare given litrle íoocj dur-ing \4¡rnlei. Hrqirer oe¡isll,oí cows in CL2 has resulreci in an errrener,

Junc 1993 Galindo-Lcal ct al.-Coucs dar and cttic 133
Biosphere Rescrve (Hastings and Turner. i9ó5;Rogers, 1982; Humphrev, 1987). Some aurhr¡rshave predicted that these influences will increaseas the region continues in a long-term rrend to-ward greater aridit,v (Hastings and Turner, 19ó5;Humphrey, 1987). Brown and Henrv (1981)documented an increasing incidence and severityof drought in southcentral Arizona since 1950,anal predicted continued elimination of Coues deerpopulations from the western extremiries of therange. There is no information abour climaticchanges in Mexico, but similar trends are likell'.
In addition 10 the factors discussed above, direcrhuman activities have also strongly influenced thepatrerns of distribution and abundance docu-mented. In sire PL3, u,here deer prelerred man-zanita shrubland and avoided grassland in rheabsence of cattle, hunring could be largelv re-sponsible for these patterns. Sites u'irh highercattle stocking rates have also higher levels ofhuman activilies (more hunting, caprure of fau'ns.harassment b,v dogs), which negativelr,affecr deerpopulations. The fact that deer populations arerelativelv higher insidc the reserve seems to be
related to the interrelation ol the three facrorsdiscussed above.
We thank The Ecologi, Group at Universiivof British Columbia for comrnents and supoonduring data analysis. Fieldwork was supportedby Consejo Nacional de Ciencia y Tecnología(CONACYT 22OCCOR88O554) and lnstifutc,de Ecología./MAB-UNESCO Program for theBiosphere Reserves in Mexico. We thank J. Be -
nitez, R. List (UAM-I), S. Franco (SEDUE-Hidalgo), F. Romo (SEDUE-Durango). J. AMoreno, G. Moreno, and C. Streeter (Ejido San
Juan) for their voluntarv help during fieldu'ork.We thank E. Gutiérrez from Las MargaritasRanch (PL2), Eduardo De la Peña from La PeñaRanch (PL1), and the people of the Ejido de San
Juan Michis (CL1, CL2). especiallv J. A. It4o-reno and E. Contreras, for their cooperat;on andsupport. L. Jamieson and R. List made correc-tions to the manuscripr, This studf is part of a
joint project by the three authors to restore Couesdeer popularions in the Western Sierra N{adre.Mexico.
i.,;*._Llr¿xerun¡ C¡r¿¡-r
ANTHoi-!'. R. C. 197ó. Inñuence of drougnr ancj c)ei!on numbers ol desert deer. J. \\¡iidl. hfer:::.. 4i'1 40- 1 ¿"i.
overgrazcd area and extremely few deer, com-
pared with site CLI where less alfluent Ejidatariosown fewer cor,r's.
These resuhs suggest that high stocking rates,
such as those found on site CL2, affect deer neg-
arivel\'. Studies of Coues deer in Arizona have
arrived at similar conclusions. Brown (1984)
found a negative correlation between cattle and
dcer abundance. Smith (1984) found improvedfawn production and survival when cattle num-bers were reduced and precipitation was high.Removal o[ herbaceous cover by cattle increaseswater loss rates, decreasing the effectiveness ofvegeration in retaining summer precipitation. Be-cause rainfall is lower and temperatures are high-er ar iower altitudes, overgrazing is particularlydetrimental to deer at lower elevations (Brou'nand Henry, 1981). Previous studies in site PL1(N'lorale s and Galindo-Leal, 1987) also suggested
thar both hunting and cattle grazrng are majornegarive influences in the distribution and abun-dance of Coues deer.
Cattle ma1' affec1 deer directly (behavior andcomperition) or indirectly (shared diseases andhabitar changes). Grazing, in particular, mav in-fluence deer food supply (Morales, 1985), andhabitat composition and sructure. Grazing andtrampling b1, cattle may affect understory vege-
ration. reducing protective cover for r¡'ildlife. Con-sequentl-v. deer may change their habitat utili-zation patterns to avoid areas r.t'ith littleunderstorl'. In Texas, white-tailed deer preferredrangelands that were put to rest periodically(Reardon et al., 1978). Lack of protective coveris especially detrimental for fau'ns, which dependon cover for protection (Carroli and Brou'n, 1977;Mackie. 1981; Lofi et al., 1987). Mo{:rate andhigh cattle stocking rates significantlv reduce cov-er for dee r and maf increase predation on fawns(Loft et al., 1987). This seems to be rrue in thisstudl' area, where all prorective cover providedby dry grasses (up to 80 cm) is removed in over-grazed areas (Garcia. 1 98ó). Ir{ore studi' is need-ed to test these ideas to further understand cattleand deer interactions (Weber et al., 1991).
Eventually, the effect of cattle grazing on sur-vival and regeneration of seedlings and saplingswill inffuence the relative abundance o[ dominantpiant species (Mackie. 1981 ; Brou'n, 1984). Sinceihe turn of the centur,v. both climatic changes andcatrie overgrazing in the sourhu,esre¡'n UnitedSiarcs irave been responsible for maior changes
rn plani communiries similar to those in N'lichiiia

ANTHoNv, R. G., exo N. S. Sv¡r¡¡. 1977. Ecrlógicalrelationship between mulc deer and white_taileddeer in southwestern Arizona. Ecol. Monogr., 47:art 111
B¡,x¿n, R. H. 1958. The furure of wildlife in norrh_ern Mexico-a problcm in conscrvarion education.Trans. 23rd. N. Amer. Wildl. Nat. Res. Conf.,1:5ó8-575.
Bex¡n, R. H., eNo J. K. Gnrr,n. 1962. Mammalsof the Mexican state of Durango. Mich. State Univ.Mus., Biol. Scries., 2:25-154.
BnowN, D. E., aNp R. S. HnNnr.. 19g1. On relicroccurrences of white-railed deer within the Sonorandesen in Arizona. Sour.hwestern Nat., 26:147_1 52.
Bnowx, M. 1984. Habitat selection bv Coues whjre_railed deer in relarion to grazing inrensiry. pp. 1_7, in Deer in the southwest: a workshop (p. RKrausman and N. S. Smith, eds.). Arizona Coop.Res. Unir and Univ. of Arizona, Tucson.
C,r,nnoll, B. K., ,tNo D. L. BnowN. 1977. Facrorsaffecring neonatal far.n survival in southern-centralTcxas, 1971-1973. J. Wildl. Mgmr., 41:63_69.
Cor-r-rNs, W. B., aNo P. J. UnNrss. j991. Habitatpreferences of mule deer as rated by pellet-groupdistributions. J. Wildl. Mgmr., 45:969-972.
1984. The pellet-group census technique asan indicator of relative habitar use: response to Le-opold et al. Wildl. Soc. Bull., 12:327.
Eoc¿, W. D. 1989. Determining elk distributionwith peller groups and telemetrl' techniques. J.Wildl. Mgmt., 53:621 -624.
Fn¡,npv, D. J.,,,rwo D. C. Bowpr,N. 1983. Efficacvof permanenr and temporary peller plots in Juni-per-Pinyon woodland. J. Wildl. \{gmr., 47:512-51 6.
Ge¡-¡xoo-L¿,r1, C. 'i992. Ove¡esrimarion of deerdensiries in \ztichilia Biosphere Resen'e, Durango,Mexico. Sourhweslern Nat-., 3j :2O9 -ZIZ.
G,u.ct¡, G. O. 198ó. Contribución al conocimientrrde la auroecología d.e Bouteloua gracili.s en el surestedel Esrado de Durango, México. Unpubl. M.S.thesis. Universidad Michoacana SalNicolás de Hi-dalgo. lvfichoacán.
HrsnHcs, J. R., aNo R. M. Tunx¿¡t. 1965. Thechanging mile, Univ. Ari¿ona Press, Tucson, Ari-zona.
Honx, H. S. 1966. Measuremenr of .,overlap" incomparative ecological studies. Amer. Nat., 100:419-424.
Huvpnney, R. R. 1987. 90 years and 535 miles:vegeration changes along the Mexican border. Univ.New lvlexico Press, Alburquerque, New Mexico.
KN¡rE, T. 1977. The Ariznna whitetail deer. SpecialReport N. 6.. Arizona Game and Fish Dep'"., phoe -nix. Arizona.
Knnss, C. J. 198S. Ecological merhodologv. Harperand Rou,. New Yorl:.
L'tqg:-o. a. S 1959 Wildlife of Mexico. Unrr
Thc Southu¡es tern l\i aturohst vol. 38, no 2
California Prcss, Berkeley and Los Angeles, Cai-ifornia -
Lorr, E. R., J W. M¿Nxr, J. G Krr. ¡xo R. CBf,nrnav. I987. Influence o[ cartlc srocking raLton the srrucrural pro6le of deer hiding cover .J.Wildl. Mgmt., 51 :655-664,
Lorr, E. R., lNp J. G. Krr. 'l 988 Comparison olpcllet-group and radio triangularion methods forassessing deer habitar use. J. lVildl. Mgmr., 52:524-527.
M,rcxte, R. J. 1981. Interspecific relarionships. pp.487-507, ¿n Mule and Black-raiied deer ol NonhAmerica (O. C. Wallm<.¡, ed.). Wildliic lr{anagc_ment Instirute Book, Unir,. Nebraska press. Li¡r_coin.
M,,rNl-y. B.F.J. 1971. A model for cerrarn ry,pes olselection experimcnts. Biomerrics. 30..291 _294.
Mon,rr¡.¿s, A. 1985. Analisis cuantira¡ivo de las dierasdel ganado vacuno !' e] venado cola blanca en LzMichiiia. Durango. Unpubl. B.S. rhesrs. Facuitacde Ciencias, Universidad Nacional Aurónoma deMéxico, México.
N4oRALEs A. ,,rxo C. G¡LtNoo-L¡.c,I_. 1.)E: . DisiriLru-ción espacial y abundancia del venado cola blancaen la Sierra Madre Occidenral. Duranso. Memc,,rias del \/ Simposio sobre Fauna Srivesrre . Unjve ¡_
sidad Nacional Autónoma de lv{éxico. lvléxico. 3_l¡339.0.
MOn¡srr¡, M. 1959. \4easuring of inrerspecifii.a:-sociation and simiiariry berween communjrjes. lv{em.Fac. Sci. Kyushu Univ. Ser. E (Biol.), 3:05*80
RrennoN, P. O., L. B. M¿nnr¡_1. AND C. A T.rr,¡-on.JR. 1978. Whire-¡ailed deer preferences and hunrer.success under various grazing syslems. J. RanqeMgmt., 31:40-42.
Roc¿ns, G. F. 1982, Then and nou.: a photographirh.istory of vegetation change in the cenrral Grea,Basin Deserr. Univ. Utah Press.. Salr Lake Cirr.L, ta h.
SHnr,v¿, F. 1915. The vegetation of a cjest.rr moun_rain range as conditioned b,v ciimarir f acrors Car-negie lnst. 14¡ashington. Pubi. ir-.2j.. J. B. Li¡,-pincotr Companv Pres:
Sv¡rg. E. P. 1982. Niche b¡eadth. resourc.e ar.¿ii_ability, and inference. Ecologr'. 63:'1615- 1ógi.
Svlrx, N. S. 1984. Reproducrion in Coues u.hirr_tailed deer relative to droughr and carrle srockinqrates. Pp. 13-20, in Deer in the sourhu,esr: a +,ork-shop (P. R. Krausman and N. S. Smirh. eds.). Ar-izona Coop. Res. Unir and Univ_ of Arizona. Tus-con.
Soxal, R. R., arn F. J. Ronr-r. 19g1. Bromer"rl .
Second ed. W- H. Frecman and Companr.. S:r¡;Frarrct¿:c. Californr a.
W¿r¿n. M-. .{. \.fon¡LEs. AND C. G¡ltroiu]-r¡r1991 Aciaprive managemenr ol Coues clerr .r i)L;-rango is joini nranaqemeni of catrit arrd oerr íe¡-
134
Junc 199.j Galindo.Leal er al.-Coues decr and c¿ttlc 135
sible? Pp. 80, in The biologv of deer (D. R. Brown,ed.). Springer Verlag, New York.
Ws¡r¿, G. C., ,rNp L. E. E¡nnn,rnpr. 1980. Sra-ristical analysis of decr and elk pcllcr-group data.J. Wildl. Mgmr., 44:121-131.
Wll-xlNsoN, L. 1988. SYSTAT: rhe s1'stem for sta-tistics. SYSTAT, Inc., Evansron. Illinois.
Zan, J. H. 1984. Biostatistical analysis. 2nd ed.
Prcntice Hall Inc., Ncw Jcrscy.
Related Documents











