Direct epithelial–stromal interaction in corneal wound healing: Role of EMMPRIN/CD147 in MMPs induction and beyond Eric E. Gabison a, b, c, * , Eric Huet a , Christophe Baudouin c , Suzanne Menashi a a CNRS UMR 7149, Universite´ Paris 12, Cre ´teil, France b Department of Ophthalmology, AP-HP, Ho ˆpital Bichat, APHP, Paris, France c UMRS968, INSERM UPMC, Vision Institute, Paris, France abstract In the cornea, the epithelium and the underlying stroma are separated by the basement membrane and Bowman’s layer. The disruption of these anatomical barriers during wound healing represents a key step which initiates tissue remodeling through the modification of the epithelial–stromal interactions (ESI). Diffusible cytokines are generally viewed as central modulators in the bidirectional communi- cation between these epithelial and stromal compartments and their implication in all stages of the wound healing process has been an active area of research for many years. Our studies which aimed to explore mechanisms of matrix degradation in pathological corneal wound healing have shown that EMMPRIN, a glycoprotein expressed on corneal epithelial cell surface, can induce matrix metal- loproteinase (MMP) production and myofibroblasts differentiation after direct interaction with corneal fibroblasts. EMMPRIN appears therefore as a potential mediator of ESI by direct cell–cell contact which represents a new mechanism for dysregulated MMPs’ induction observed in corneal ulcerations. These direct epithelial–stromal interactions (direct-ESI) can occur when delayed epithelial healing prevents regeneration of the basement membrane and allows the two cell types to come into close proximity. We propose that prevention of these interactions through inhibition of EMMPRIN may represent a promising therapeutic strategy in the inhibition of MMP induction in ulceration. Ó 2008 Elsevier Ltd. All rights reserved. Contents 1. Introduction ....................................................................................................................... 20 2. Background: cytokine mediated epithelial–stromal interactions (ESI) in the cornea ........................................................ 20 2.1. ESI in physiology: questioning basement membrane permeability ................................................................... 20 2.2. ESI in fibrosis ................................................................................................................ 21 2.3. ESI in ulcerations ............................................................................................................ 21 3. EMMPRIN as a mediator of direct epithelial–stromal interactions ........................................................................ 22 3.1. EMMPRIN in pathology ........................................................................................................ 22 3.2. Role of EMMPRIN in the induction of MMPs through direct-ESI in corneal ulcerations ................................................ 23 3.3. EMMPRIN and myofibroblast differentiation ..................................................................................... 24 4. Interplay between EMMPRIN, IL1 and TGFb in corneal stromal remodeling: melts versus fibrosis ............................................ 25 5. Protection against direct-ESI mediated corneal melts: hypothesis ........................................................................ 28 5.1. Corneal basement membrane and the Bowman layer as barriers against direct-ESI ................................................... 28 5.2. Additional role for stromal cells’ apoptosis as a dynamic barrier to the healing epithelium? ....................... .................... 30 6. Conclusions and future directions ..................................................................................................... 31 Acknowledgments .................................................................................................................. 31 References ......................................................................................................................... 31 * Corresponding author. UMRS968, INSERM UPMC, Vision Institute, Paris, France. E-mail address: [email protected] (E.E. Gabison). Contents lists available at ScienceDirect Progress in Retinal and Eye Research journal homepage: www.elsevier.com/locate/prer 1350-9462/$ – see front matter Ó 2008 Elsevier Ltd. All rights reserved. doi:10.1016/j.preteyeres.2008.11.001 Progress in Retinal and Eye Research 28 (2009) 19–33

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

lable at ScienceDirect
Progress in Retinal and Eye Research 28 (2009) 19–33
Contents lists avai
Progress in Retinal and Eye Research
journal homepage: www.elsevier .com/locate/prer
Direct epithelial–stromal interaction in corneal wound healing: Roleof EMMPRIN/CD147 in MMPs induction and beyond
Eric E. Gabison a,b,c,*, Eric Huet a, Christophe Baudouin c, Suzanne Menashi a
a CNRS UMR 7149, Universite Paris 12, Creteil, Franceb Department of Ophthalmology, AP-HP, Hopital Bichat, APHP, Paris, Francec UMRS968, INSERM UPMC, Vision Institute, Paris, France
* Corresponding author. UMRS968, INSERM UPMCE-mail address: [email protected] (E.E. Gabison).
1350-9462/$ – see front matter � 2008 Elsevier Ltd.doi:10.1016/j.preteyeres.2008.11.001
a b s t r a c t
In the cornea, the epithelium and the underlying stroma are separated by the basement membrane andBowman’s layer. The disruption of these anatomical barriers during wound healing represents a keystep which initiates tissue remodeling through the modification of the epithelial–stromal interactions(ESI). Diffusible cytokines are generally viewed as central modulators in the bidirectional communi-cation between these epithelial and stromal compartments and their implication in all stages of thewound healing process has been an active area of research for many years. Our studies which aimed toexplore mechanisms of matrix degradation in pathological corneal wound healing have shown thatEMMPRIN, a glycoprotein expressed on corneal epithelial cell surface, can induce matrix metal-loproteinase (MMP) production and myofibroblasts differentiation after direct interaction with cornealfibroblasts. EMMPRIN appears therefore as a potential mediator of ESI by direct cell–cell contact whichrepresents a new mechanism for dysregulated MMPs’ induction observed in corneal ulcerations. Thesedirect epithelial–stromal interactions (direct-ESI) can occur when delayed epithelial healing preventsregeneration of the basement membrane and allows the two cell types to come into close proximity.We propose that prevention of these interactions through inhibition of EMMPRIN may representa promising therapeutic strategy in the inhibition of MMP induction in ulceration.
� 2008 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202. Background: cytokine mediated epithelial–stromal interactions (ESI) in the cornea . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
2.1. ESI in physiology: questioning basement membrane permeability . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 202.2. ESI in fibrosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 212.3. ESI in ulcerations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
3. EMMPRIN as a mediator of direct epithelial–stromal interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 223.1. EMMPRIN in pathology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 223.2. Role of EMMPRIN in the induction of MMPs through direct-ESI in corneal ulcerations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 233.3. EMMPRIN and myofibroblast differentiation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
4. Interplay between EMMPRIN, IL1 and TGFb in corneal stromal remodeling: melts versus fibrosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 255. Protection against direct-ESI mediated corneal melts: hypothesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 28
5.1. Corneal basement membrane and the Bowman layer as barriers against direct-ESI . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 285.2. Additional role for stromal cells’ apoptosis as a dynamic barrier to the healing epithelium? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
6. Conclusions and future directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .31Acknowledgments . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
, Vision Institute, Paris, France.
All rights reserved.

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3320
1. Introduction
The cornea, first and most powerful lens of the eye, is a trans-parent anterior ocular tissue that has been extensively used asa model for studying wound healing because of its remarkablyorganized structure. It is composed of three distinct layers, anepithelium, a stroma, and an endothelium separated by threedifferent structures, the epithelial basement membrane, Bowman’slayer, between the epithelium and the stroma and Descemet’smembrane between the stroma and the endothelium. Separationof these tissue compartments is considered to be essential forcorneal physiology and abnormalities in the regeneration of theirrespective limiting membranes following injury have beendemonstrated to be associated with pathological wound healingthat ranges from corneal haze (scar tissue) to ulceration andperforation, resulting in a loss of transparency and vision. Thesecomplications are thought to be triggered by the epithelial–stromalinteractions (ESI) that follow the disruption of the epithelialbasement membrane (Matsubara et al., 1991; Fini et al., 1992;Stramer et al., 2003).
Reconstitution of the transparent corneal stroma in woundhealing depends on the fine regulation of the biosynthetic activitiesof post-lesion fibroblasts and on matrix degradation, attributedessentially to proteolytic enzymes belonging to the matrix metal-loproteinase (MMP) family (Sivak and Fini, 2002). There is nowincreasing evidence that ESI may have a significant impact on theproduction and the localization of these enzymes observed duringphysiological wound healing and in their persistent upregulationobserved in pathological situations such as ulcerations (Wong et al.,2002).
ESI can be mediated through soluble factors such as cytokinesand growth factors or through cell–matrix interaction where thecellular behavior of the migrating cell is modified by the newenvironment (Wilson et al., 2001). As most studies in thisfield have focused on the role of cytokines, ESI have became byextension the study of the biological effects of cytokineproduced by one cellular compartment (stroma or epithelium) onthe cells from the other compartment. In this review weintroduce the concept of ESI in the cornea through direct cell–cell contact, which we refer to as direct-ESI and discuss themechanisms underlying such interactions and their conse-quences in the wound healing process in which they mayoccur.
The notion of direct-ESI was first introduced in the cancerfield when EMMPRIN was identified as an MMPs’ inducer presenton the cell surface of tumor cells which can activate stromal cellsthrough direct contact and signal them to increase MMPs’production (Toole, 2003). Recently accumulated data also advo-cate a role for EMMPRIN in modulating MMPs’ expression duringnon-tumoral pathological situations as well as in normal tissueremodeling and differentiation (Muramatsu and Miyauchi, 2003;Gabison et al., 2005a; Weiss et al., 2007). Our own workidentified EMMPRIN in the normal corneal epithelium anddemonstrated its ability to activate corneal fibroblasts in culturethrough direct contact with epithelial cells so to trigger bothmyofibroblasts differentiation and MMPs’ production, thus sug-gesting a role for EMMPRIN as a key mediator of direct-ESI duringcorneal wound healing (Gabison et al., 2005b; Huet et al.,2008a,b).
After a brief review on the implications of ESI mediated bydiffusible cytokines, viewed as central modulators of the responseto corneal injury, we focus more particularly on the role of EMM-PRIN in promoting corneal stroma remodeling through direct-ESIand hypothesizes on the different clinical situations where theseinteractions may occur and on the natural defenses that exist forthe prevention of such interactions.
2. Background: cytokine mediated epithelial–stromalinteractions (ESI) in the cornea
2.1. ESI in physiology: questioning basementmembrane permeability
Accumulated evidence based on immunohistochemical analysisof corneal tissue sections or on in vitro experimentation suggestscomplex interplay between cytokines and their receptors in kera-tocytes (corneal stromal cells) and epithelial cells. Differentpatterns of cytokines/receptors expression have been described (Liand Tseng, 1995) which suggest the existence of paracrine signalstransmitted from mesenchyme to epithelia or vice versa. Somecytokines or growth factors, such as IL1 or PDGF, are exclusivelyexpressed by epithelial cells under physiologic conditions, whiletheir respective receptors are predominantly expressed by stromalcells, suggesting epithelial to mesenchymal communication(Wilson et al., 1999a,b; Daniels and Khaw, 2000; Wilson et al.,2001). Others, including TGFb and basic-FGF, are expressed in bothepithelial and stromal cells, while keratinocyte growth factor (KGF)and hepatocyte growth factor (HGF) are predominantly expressedby fibroblasts and selectively stimulate corneal epithelial cellproliferation through the activation of their cognate receptors. HGFalso promotes epithelial cells’ motility and inhibits their terminaldifferentiation (Wilson et al., 1993; Wilson et al., 1999a,b). As anadditional level of complexity, these cytokines and growth factorsare also involved in feedback mechanisms where the activation oftheir receptor in a target cell can stimulate the production ofanother growth factor which may in turn cross-talk to the originalcells in reciprocal epithelial–mesenchymal interactions (Sotozonoet al., 1994; Honma et al., 1997; Weng et al., 1997; Imanishi et al.,2000; Carrington and Boulton, 2005).
The existence of such elaborate system for epithelial–stromalcommunication through cytokines raises a fundamental question asto whether it necessitates basement membrane (BM) disruption.The review of published reports suggests that this may depend onthe cytokine in question. Elegant studies blocking KGF receptorfunction in the skin by the expression of a dominant-negative KGFreceptor mutant resulted in disorganized atrophic epidermis andreduced keratinocyte steady state proliferation (Werner et al.,1994).This implies that in tissue homeostasis, KGF can exert its biologicaleffects on epithelial cells through intact BM, although it remains tobe seen whether the same also applies to KGF effects in the cornea.Conversely, BM seems to be impermeable to the epithelial cellderived IL1, TGFb and PDGF as these cytokines were shown to besequestrated in the epithelial compartment under homeostasisconditions (Weng et al., 1997; Kim et al., 1999). The intracellularlocalization of IL1 alpha, due to the lack of signal peptide forextracellular secretion, may be one explanation for its retention inthe unwounded epithelium which suggests that it requires epithe-lial lesion to be released (Wilson et al., 2001; Stapleton et al., 2008).The findings that keratocyte apoptosis can result from superficialepithelial injury without disruption of the BM provide support forBM permeability to released IL1 (Wilson et al., 2002).
Like IL1, PDGF and TGFb expressed by the corneal epithelial cellsdo not exert their proliferation, chemotaxis and differentiationeffects on stromal cells under homeostasis conditions. The mech-anisms preventing both these growth factors to reach the stromaare likely to be related to their heparin binding property whichwould favor attachment and sequestration within the heparinsulfate proteoglycans present in the BM (Gohring et al., 1998;Eming et al., 1999). Accordingly, comparative studies of woundedand unwounded corneas revealed intense staining of PDGF andTGFb in areas of intact epithelial basement membrane (Kim et al.,1999), while stromal subepithelial staining was only detected inareas of BM disruption (Wilson et al., 2001).

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 21
Hence, intact BM displays selective permeability to the variousdiffusible cytokines derived from either side of the BM and whichhave been extensively shown to cross-talk during wound healing.The question of their role in corneal homeostasis remains howeveran open one.
2.2. ESI in fibrosis
Based on empirical observations of surgical outcome, refractivesurgery procedures have progressively evolved towards minimi-zation of ESI long before the fundamental mechanisms of theseinteractions have been evoked. Indeed, the subepithelial fibrosis(haze) and refractive regression clinically observed in high myopicpatients after photorefractive keratectomy (PRK) have led to thedevelopment of laser in situ keratomileusis (LASIK) which attenu-ates the influence of the epithelium on the treated stroma, thusallowing surgery of higher level of myopia with minimal fibrosisand regression. In agreement with a protective role of BM, the PRKprocedure which involves photoablation through the BM and theanterior stroma is associated with the occurrence of subepithelialfibrosis in the treated area, while LASIK which maintains the BMbarrier is devoid of such complication, except at the flap margincorresponding to the areas where the BM is broken, allowing localepithelial–stromal interaction. Other surgical procedures havesince been developed, including EPILASIK and LASEK, where theepithelium together with its intact BM is replaced back on thetreated stroma following photoablation, thus minimizing ESI (Nettoet al., 2005).
Experimental investigations strengthen the notion that ESImediate fibrotic repair in the cornea. In a co-culture system,Nakamura et al. (2002) demonstrated that injured corneal epithe-lial cells secreted soluble factor that crossed a membrane imper-meable to cells to stimulate corneal fibroblasts’ proliferation,myodifferentiation and collagen gel contraction. These and otherstudies implicated TGFb as the major cytokine involved in ESImediated fibrotic repair (Nakamura, 2003). Playing a pivotal role inwound healing, TGFb is a potent stimulator for connective tissueformation as it increases ECM synthesis and downregulates ECMdegradation. TGFb is also known to exert its actions through thedifferentiation of fibroblasts into myofibroblasts, aSMA containingcontractile cells that are essential for ECM remodeling (Jester andHo-Chang, 2003). The implication of TGFb was further demon-strated by the inhibition of the fibrotic reaction after topical (Jesteret al., 1997) or systemic (Mita et al., 1998) administration of anti-TGFb antibody following corneal injury. Connective tissue growthfactor (CTGF) has also been implicated in the fibrotic process.Although it cannot induce myofibroblast differentiation on its own,it has been able to mediate some of TGFb effects (Daniels et al.,2003a,c; Garrett et al., 2004).
The protective role of the BM against ESI mediated fibroticresponse has been recently demonstrated in elegant in vivostudies. In an animal model of PRK where defective basementmembrane regeneration was obtained experimentally, Netto et al.(2006) showed increased epithelial-derived TGFb signaling andmyofibroblast generation. Conversely, Javier et al. (2006) demon-strated that faster BM regeneration/maturation was associatedwith inhibition of corneal scarring in an animal model of LASEK.Additional studies from Fini’s group demonstrated the linkbetween maturation of the BM after wounding in vivo and theprogressive disappearance of activated myofibroblasts from theanterior stroma (Stramer et al., 2003). These observations wereconfirmed by the same authors using an artificial cornea culturesystem in vitro where the formation of an endogenous BM wasfound to prevent fibrotic marker expression through the inhibitionof TGFb release to the stromal compartment (Zieske et al., 1994;Stramer et al., 2003).
The role of the BM in modulating ESI goes beyond its barrierfunction preventing TGFb diffusion towards the stroma. Recently,BM was shown to inhibit the production of TGFb by epithelial cellsin a way that depended on its composition and complex structure(LaGier et al., 2007). Consistent with these findings, TGFb levels intears were shown to be lower after LASEK which preserves BM, ascompared to PRK (Lee et al., 2002). Furthermore, in vivo studieshave shown that TGFb signaling through the Smad pathway isactivated in epithelial cell when BM is disrupted (Zieske et al., 2001;Hutcheon et al., 2005) while in vitro studies have shown inhibitionof this pathway when epithelial cells are seeded on mature BMfrom amniotic membrane (Tseng et al., 1999; Kawakita et al., 2005).
Hence, the maturation of the BM during wound healing not onlyresult in physical hindrance to epithelial pro-fibrotic cytokines butalso inhibits their production by the regenerating corneal epithelialcells, thus representing an efficient antifibrotic protectionmechanism.
2.3. ESI in ulcerations
A common initiating event in corneal ulcerations of variousetiologies including tear film anomalies, toxic reactions, neuro-trophic or metabolic disorders is a delay in epithelial closureleading to persistent epithelial defect (PED) (Ma and Dohlman,2002). In the absence of healing, PED progresses towards basementmembrane disruption which is prerequisite for stromal thinning(Matsubara et al., 1991; Fini et al., 1996). Proteinases such as thematrix metalloproteinases and the serine proteases of the plas-minogen–plasmin system are known to be overexpressed and havebeen implicated as mediators in the pathogenesis of this cornealdisorder, tilting the ECM remodeling towards proteolysis (Slanskyet al., 1969; Berman et al., 1980; Berman et al., 1983; Berman, 1993;Geerling et al., 1999). Epithelial-derived MMP-9, which is known tobe increased at the edges of corneal ulceration (Fini et al., 1992;Gabison et al., 2003), is of special interest in this process as inaddition to degrading basement membrane components, thisenzyme has been shown to delay epithelial wound closure (Danielset al., 2003a,b,c). The inhibition of MMP-9 activity may thereforeappear tempting in the management of corneal epithelial defects,but the wound healing of MMP-9 deficient mice, althoughaccelerated, resulted in defective restoration of their cornealtransparency following stromal injury (Mohan et al., 2002). Thisdemonstrates the complexity in understanding the physiopatho-logical functions of these proteinases and highlights the difficulty intargeting their activity in the treatment of corneal ulceration.
The difficulty in determining MMP functions in vivo is furtherillustrated by the findings that the proteolytic enzymes detected incorneal ulcerations do not differ from those required for tissueremodeling associated with physiological wound healing. Thus, thesame metalloproteinases MMP-1, MMP-2, MMP-3 and MMP-14 areknown to be induced in the same subepithelial region in bothchronic corneal ulcerations and during wound healing that followsPRK. However these enzymes which are rapidly repressed after PRKpersist in chronic ulcerations and can still be detectable severalweeks to months following the initial corneal lesion (Gordon et al.,1980; Matsubara et al., 1991; Girard et al., 1993; Riley et al., 1995; Luet al., 1999; Daniels et al., 2003a,b,c; Gabison et al., 2003). Thissuggests that it is not the nature of the enzymes but rather theirderegulation which alters the proteolytic balance towards excessivedegradation of the extracellular matrix.
Numerous studies have attempted to understand the mecha-nisms involved in the excessive production of MMPs in cornealulcerations. Epithelial-derived IL1 is thought to play a central roleas it is known to upregulate the expression of several MMPs,including the interstitial collagenase MMP-1 (Sivak and Fini, 2002).It also potentiates chemotactic and angiogenic effects through the

Fig. 1. Scheme of EMMPRIN structure. EMMPRIN contains an extracellular domaincomposed of two Ig loops with three Asn-linked oligosaccharides and short singletransmembrane domain (TM) and a cytoplasmic domain (Cyt). The first Ig domain isrequired for counter-receptor activity, involved in MMP induction.
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3322
induction of IL-8 in corneal resident cells and may therefore playa major role in the progression of a persistent epithelial defecttowards stromal melting, inflammatory cell infiltration and neo-vascularization, features of corneal ulcerations (West-Mays et al.,1997).
TGFb which favors ECM accumulation through inhibition of IL1-mediated MMP synthesis (Girard et al., 1991; West-Mays et al.,1999) and increased matrix synthesis is known to also be secretedunder the same conditions as IL1 during wound healing and couldtherefore antagonize IL1 effects on ECM. This may suggest that inchronic corneal ulcerations, associated with excess proteolysis,other mechanisms may exist which favor IL1 effects. The autocrineloop of IL1 in keratocytes described by the group of Fini mayrepresent such a mechanism which can potentiate proteolysiswithin the stroma (West-Mays et al., 1997). IL1 binding to itsreceptor on the surface of stromal fibroblasts, which do not nor-mally produce IL1, has been shown to activate both collagenaseexpression and the IL1 autocrine loop. Thus, when keratocytes areexposed to the cytokine following epithelial injury they can enterinto a self-perpetuating loop that can potentially amplify theproduction of metalloproteinases. Interestingly, the IL1 autocrineloop can also be stimulated by proteolytically degraded fragmentsof fibronectin, following binding to the a5b1 integrin receptor(Berman, 1994). Support for the importance of IL1-mediated effectswas provided by the use of IL1-receptor antagonist (IL1-ra) whichinhibited collagenase production in vitro (Jung et al., 2007) andimproved prognosis of corneal ulceration in animal models (Mooreet al., 2002; Xue et al., 2003; Yamada et al., 2003). The inhibition ofcollagenase obtained with IL1-ra was however not complete, sug-gesting the existence of other factors involved in this process (Junget al., 2007). EMMPRIN, which is known to induce MMPs throughcell–cell interactions may participate along with IL1 in theincreased matrix degradation observed in corneal ulceration.
3. EMMPRIN as a mediator of directepithelial–stromal interactions
The possibility that direct-ESI can modulate proteinaseproduction by stromal cells was suggested by the group of Gross(Johnson-Muller and Gross, 1978) about 30 years ago. However, theimportance of these interactions was recognized much later withthe realization that the stromal cells at the vicinity of tumor cellswere the origin of the overexpressed MMPs often observed incancer tissues. EMMPRIN (Extracellular Matrix MetalloPRoteinaseInducer) was then identified (Kataoka et al., 1993; Biswas et al.,1995) back in the 1990s as a transmembrane glycoprotein enrichedon tumor cells of epithelial origin and which is responsible for thisinduction of MMPs in neighboring stromal cells through directtumor–stromal interactions. EMMPRIN can greatly increase thecell’s proteolytic potential, as in addition to inducing a variety ofMMPs including MMP-1 (interstitial collagenase), MMP-2 (gelati-nase A), MMP-3 (stromelysin 1), it has no effect on their physio-logical inhibitors, TIMP-1 and TIMP-2 (Toole, 2003). EMMPRIN wasalso shown to increase expression of the plasminogen activationsystem, including urokinase and its receptor, hence furtherincreasing its proteolytic potential in the stroma (Quemener et al.,2007).
EMMPRIN, also commonly referred to as basigin or CD147,belongs to the immunoglobulin superfamily, cell surface proteinsthat are generally involved in cell recognition. The proposed role forEMMPRIN in cell–cell interaction and its ability to induce MMPs’production in neighboring cells implies the existence of a counter-receptor on the opposing cell, although as yet none has beenidentified. It has been suggested that EMMPRIN may serve as itsown counter-receptor (Sun and Hemler, 2001), thus stimulatingMMPs via homophilic interactions through the extracellular
domains of EMMPRIN in neighboring cells. These extracellularregions contain two immunoglobulin (Ig) like domains with threeconserved N-glycosylation sites that are variably glycosylated.Glycosylation was shown to determine its MMP stimulatingactivity, as the purified deglycosylated EMMPRIN not only failed toinduce MMP activity but also antagonized the activity of the nativemolecule (Sun and Hemler, 2001). The transmembrane domain andthe short cytoplasmic domain are thought to be implicated ininteractions between EMMPRIN and other molecular partnerswithin the membrane such as integrins, cyclophilins and others(Nabeshima et al., 2006; Yurchenko et al., 2006), although thesignificance of such interactions or their involvement in signaltransduction is yet to be determined (Fig. 1).
In addition to regulating MMPs, other functions are increasinglyascribed to EMMPRIN. These are evidenced in the variety ofdevelopmental defects observed in the EMMPRIN null mouse. Thesemutants are small and usually unable to undergo implantation. Theembryos that implant survive past birth, but the offsprings aresterile, demonstrating deficiencies in spermatogenesis and fertil-ization. In addition, these null mice are blind due to defective retinaldevelopment and have faulty sensory and memory functions (Iga-kura et al., 1996; Igakura et al., 1998; Muramatsu and Miyauchi,2003). Experimental challenges to the adult EMMPRIN knockoutmouse are still missing, but the implication of EMMPRIN in severalphysiopathological situations in which there is a close correlationbetween EMMPRIN and proteolysis nonetheless suggests a majorrole of EMMPRIN in tissue remodeling (Huet et al., 2008a,b).
3.1. EMMPRIN in pathology
As EMMPRIN was first identified in tumor cells where it isparticularly enriched, its role in cancer progression through MMPs’induction was initially emphasized (Yan et al., 2005). Immunohis-tochemical analysis detected EMMPRIN mainly at the periphery ofinvasive tumor clusters, corresponding to the leading edge of tumorinvasion and compatible with the concept that EMMPRIN plays

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 23
a role in tumor–stroma interaction (Caudroy et al., 1999). Theassumption that EMMPRIN may be implicated in tumor progres-sion has been strengthened by the demonstration of high levels ofEMMPRIN in numerous malignant tumors including bladder, skin,lung and breast carcinoma, and lymphoma (Polette et al., 1997;Bordador et al., 2000; Thorns et al., 2002). It was also shown tocorrelate with tumor progression (Sameshima et al., 2000; Zuckeret al., 2001; Kanekura et al., 2002) and in some cases and was alsoassociated with poor prognosis (Kanekura et al., 2002; Davidsonet al., 2003a,b; Rosenthal et al., 2003; Ishibashi et al., 2004). Therole of EMMPRIN in the induction of MMPs was confirmed bystudies on a wide range of tumors showing that EMMPRINexpression level correlated with the degree of MMP expression bystromal fibroblasts (Davidson et al., 2003a). However, experimentalstudies have demonstrated that in addition to increasing invasionthrough proteinase induction, EMMPRIN induces several othermalignant properties associated with cancer. These include thestimulation of angiogenesis by the upregulation VEGF expression(Tang et al., 2005) as well as the stimulation of cell survivalsignaling, including Akt, Erk and FAK, through the increasedproduction of the pericellular polysaccharide hyaluronan (Tooleand Slomiany, 2008). These effects of EMMPRIN would be furtheraccentuated by its ability to induce its own expression by positivefeedback regulatory mechanism (Tang et al., 2004).
In addition to cancer, EMMPRIN has been implicated in manyother pathological processes. Its upregulation has been identified intissues such as lung injury (Foda et al., 2001), rheumatoid arthritis(Konttinen et al., 2000), chronic liver disease (Shackel et al., 2002),heart failure (Spinale et al., 2000) and atherosclerosis (Major et al.,2002). The induction of certain MMPs (e.g., MMP-1, MMP-2, andMMP-3) as well as of uPA has been shown to be central in the tissuedestruction associated with these diseases. These same proteinaseshave been detected in the subepithelial layer in corneal ulcerations/perforations of multiple origin (Sivak and Fini, 2002) suggestingthat they may be induced by direct-ESI in the cornea, mediated byepithelial cell EMMPRIN.
Fig. 2. Induction and co-localization of EMMPRIN and MMP-2 in the stroma at the epithelialulcerations (left panel) and in the ulcerated area (right panel) were subjected to double-labrabbit anti-human MMP-2 pAb antibodies (red) and counterstained with DAPI (blue). Mergeulcerated area in the subepithelial stroma whereas MMP-2 staining was weak without co-loacellular layer of Bowman. Reprinted with modification from Gabison et al. (2005b) with p
3.2. Role of EMMPRIN in the induction of MMPs through direct-ESIin corneal ulcerations
Studies into the pathogenesis of corneal ulceration, such asthose associated with rheumatoid arthritis, Sjogren syndrome(Riley et al., 1995), alkali burn injury (Fini et al., 1998) or iatrogeniccorneal melts (Gabison et al., 2003), revealed a massive induction ofMMP-1, MMP-2 and MMP-3 in the subepithelial layer of the cornealstroma (Gordon et al., 1980; Wagoner and Kenyon, 1989; Majoret al., 2002). Soluble factors such as cytokines, growth factors andhormones have been long considered the major physiologicalregulators of MMPs responsible for this excessive production, butthe high levels of MMP confined to the epithelial–stromal interfaceled us to hypothesize that EMMPRIN may represent an additionalmode for focal MMP induction through direct interaction betweenepithelial cells and fibroblasts in the cornea.
In the healthy cornea, EMMPRIN was identified by immuno-histochemistry mainly in the epithelium where it is particularlyconcentrated in the basal cells, while it is barely detected in theunwounded stroma. In ulcerated corneas, EMMPRIN is markedlyincreased in the subepithelial stroma and was associated with aninduction of MMP-2 (Fig. 2) as well as MMP-3 (not shown) atprecisely the same regions of the stroma. This co-localization,which is also clearly demonstrated in isolated fibroblasts in vitrowhere both EMMPRIN and MMP-2 are localized within the samemembrane microdomains (Fig. 3), would be in accord with the roleof EMMPRIN as an inducer of these proteinases (Gabison et al.,2005a,b).
In order to evaluate the role of EMMPRIN in this proteinaseinduction, we developed a cell culture system where cornealepithelial cells and fibroblasts were allowed to come into directcontact, forming a clear interface between the two cell types.Double-labeled confocal immunohistochemistry of this co-culturerevealed a very strong staining of MMP-2 and EMMPRIN which wasrestricted to the fibroblasts along the border with the epithelialcells, as was observed in ulcerated corneas and is suggestive of
–stromal boundary in chronic corneal ulcerations. Tissue sections at a distance from theeled confocal immunohistochemistry using mouse anti-human EMMPRIN (green) andd staining of EMMPRIN and MMP-2 revealed induction of EMMPRIN and MMP-2 in thecalization with EMMPRIN in the non-ulcerated area. Note the absence of staining in theermission from the American Society for Investigative Pathology.

Fig. 3. Double-labeled confocal microscopy of EMMPRIN and MMP-2 in corneal fibroblasts in culture. EMMPRIN (green) and MMP-2 (red) are co-localized (arrows) and restricted toperipheral patchy structures (yellow).
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3324
direct cell–cell contact as the major mechanism of their induction(Fig. 4). Although a paracrine regulation of MMPs involving cyto-kines could also be involved to some extent in this co-culturesystem, it would be expected to result in a more generalized andhomogenous staining, as secreted cytokines, diluted in the culturemedium, would not be compatible with a localized induction.
To further define the implication of direct cell contact in thisinduction, another cell culture system was developed wherecorneal fibroblasts were incubated with purified epithelial cellmembranes, thus limiting the influence of cytokines that maybe secreted by epithelial cells. These EMMPRIN-containingmembranes stimulated both MMP-1 and MMP-2 and also inducedEMMPRIN’s own expression in corneal fibroblasts. The direct role ofEMMPRIN in this system was confirmed by the inhibition of theMMPs’ induction in the presence of a blocking antibody to EMM-PRIN (Gabison et al., 2005a,b). These experiments demonstrate thatdirect cellular contact mediated by the EMMPRIN present on thecell membrane can regulate both MMP and EMMPRIN expression infibroblasts and suggest that EMMPRIN may be a key factor in theexcessive tissue degradation which ensues direct-ESI.
The positive feedback regulation of EMMPRIN, already reportedin the tumor context where EMMPRIN induces its own expression(Tang et al., 2004), may account for the marked increase in its levelsobserved in the ulcerated stroma. This could represent a mecha-nism for amplifying EMMPRIN’s effects with the potential topropagate MMP induction deeper in the stroma. EMMPRIN mayalso be upregulated by cytokines (Gabison et al., 2005a,b). Inepithelial cells it is upregulated by both EGF and TGFb as well as bylow cell density which has previously been shown to regulate thefunctional expression of TGFb receptors in corneal epithelial cells(Wang et al., 2002). In fibroblasts, however, EMMPRIN’s levelpositively responded to both TGFb and IL1 with little effect of EGF.
On the basis of these results the following scenario may bepostulated on the possible events that follow corneal lesionsinvolving direct epithelio-stromal interaction. The decrease inepithelial cell density and the accumulation of cytokines known tobe secreted during wound healing, such as TGFb, EGF and IL1,
would increase EMMPRIN levels in both epithelial cells and fibro-blasts in the wounded area. This can in turn induce fibroblasts, bydirect interaction, to increase their own level of EMMPRIN. Theresultant EMMPRIN amplification may lead to excessive MMPrelease and stromal destruction in chronic situations where healingis delayed and direct epithelio-stromal interaction persists.
3.3. EMMPRIN and myofibroblast differentiation
While normal corneal stromal cells, the keratocytes, appearquiescent under normal tissue homeostasis, injury to the corneainduces a series of phenotypic changes which result in theirtransformation into myofibroblasts (Jester et al., 1995). These cellscontrol the deposition and organization of extracellular matrix incorneal wounds and are responsible for corneal wound contraction.It has been suggested that this transformation process and thesubsequent physiological activity of myofibroblasts that ultimatelydetermine the final outcome of healing process, resulting in normalrepair, fibrosis or ulceration of the cornea (Fini, 1999). The differ-entiation of fibroblast to myofibroblast can be identified by theexpression of the actin isoform typical of vascular smooth-musclecells, aSMA, which characterizes the myofibroblasts both in vitroand in vivo and which is accountable for the contractile activities ofthe myofibroblasts (Jester et al., 1999; Jester and Ho-Chang, 2003).Among the cytokines, TGFb is generally considered to be the majorfactor in the differentiation of fibroblasts into myofibroblasts(Desmouliere et al., 2005). We recently have presented evidencethat EMMPRIN, in addition to inducing proteases production, canalso actively participate in the process of myofibroblasts differen-tiation (Huet et al., 2008a,b).
Several observations suggested an association between EMM-PRIN overexpression and myofibroblasts phenotype: (1) theexpression of EMMPRIN increased in low density cultures, condi-tion known to favor myofibroblasts differentiation (Masur et al.,1996; Petridou et al., 2000; Folger et al., 2001), (2) EMMPRIN wasshown to be an early target of TGFb, the main cytokine implicatedin myofibroblasts differentiation (Verrecchia et al., 2001), (3) both

Fig. 4. Induction and co-localization of EMMPRIN and MMP-2 in corneal fibroblasts at the interface with corneal epithelial cells in an in vitro co-culture system. Fibroblasts (f) wereadded to cultured epithelial sheet and allowed to attach (A), proliferate, and migrate around the epithelial cells (B, C), forming after 24–48 h a clear interface between the two typesof cells (C). The slides were immunostained with DAPI (D and H, blue), anti-EMMPRIN/CD147, HIM6 mAb (E and I, green), and anti-MMP-2 pAb (F and J, red) and analyzed byconfocal microscopy. (G and K) Overlapping staining of EMMPRIN and MMP-2 in yellow. (H–K) Higher magnifications of the epithelial/fibroblast interface shown in D–G,respectively. Original magnifications: �10 (D–G); �60 (H–K). Reprinted from Gabison et al. (2005b) with permission from the American Society for Investigative Pathology.
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 25
EMMPRIN and aSMA, the myofibroblast marker, were co-inducedat the point of maximal contraction in collagen lattice assay,a widely used functional test for myofibroblast differentiation(Huet et al., 2008a,b).
We therefore questioned the implication of EMMPRIN in theexpression of aSMA and in the contraction of collagen gels todetermine whether it was merely a marker or an active participantin the differentiation process. We first observed that increasingEMMPRIN expression by cDNA transfection or by treatment withexogenously added recombinant EMMPRIN resulted in an upre-gulation of aSMA expression. EMMPRIN also increased thecontractile properties of the treated fibroblasts, as demonstrated bythe immunohistochemical appearance of aSMA stress fibers and bythe accelerated contraction of fibroblasts embedded collagenlattices. This effect of EMMPRIN on myofibroblast differentiationwas further supported by the observed inhibitory effect of EMM-PRIN-siRNA on TGFb induced aSMA expression and collagen gelcontraction (Huet et al., 2008a,b) (Fig. 5).
The mechanism by which EMMPRIN promotes myofibroblastdifferentiation remains to be elucidated. However, the possibilitythat EMMPRIN’s effect may involve an MMP-dependant activationof latent TGFb, leading in turn to an active TGFb-mediated aSMAinduction was ruled out, as the presence of MMPs’ inhibitors hadno effect, further implying that this function of EMMPRIN in
myofibroblast differentiation is unrelated to its MMP inducingactivity.
The implications of these in vitro observations were examinedin corneal ulcerations both in human (Fig. 6) and in an alkali burnmouse model (Fig. 7). We frequently observed co-localization ofEMMPRIN and aSMA in the same fibroblasts of the ulcerated region,which suggests that the EMMPRIN overexpressing fibroblasts in thesubepithelial stroma could correspond to myofibroblasts andsupport a role for EMMPRIN in their differentiation in vivo.However, when examining the different regions of the ulceratedarea we also observed the presence of different subpopulations offibroblasts in terms of the relative expression of EMMPRIN andaSMA, suggesting the implication of different pathways for aSMAinduction in vivo. The next section discusses in more details theimplication of EMMPRIN with respect to different cytokines,namely TGFb and IL1, in the differentiation of myofibroblasts andtheir consequences on tissue remodeling.
4. Interplay between EMMPRIN, IL1 and TGFb in cornealstromal remodeling: melts versus fibrosis
Although cytokines have been considered as the main regula-tors of the wound healing process, it appears from the literaturethat it is the interplay between these cytokines and other

Fig. 5. EMMPRIN induces myofibroblast differentiation. Treatment with recEMMPRIN induced aSMA expression in HTK corneal fibroblasts in culture (A) and collagen latticeembedded fibroblasts contraction (B). Inhibition of EMMPRIN by siRNA decreased aSMA expression (C) and collagen lattice contraction (D). Adapted from Huet et al. (2008b).
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3326
mediators rather than the mere presence or absence of cytokinesthat plays a decisive role in regulating the initiation, progressionand resolution of wounds. TGFb for example is a potent stimulatorof connective tissue formation as it increases both the expressionof ECM components and the expression of protease inhibitors
Fig. 6. Confocal immunohistochemistry of EMMPRIN and aSMA in human chronic cornealTissue sections adjacent to the ulcerated area were stained with DAPI (blue), subjected to doaSMA mAb (B, C, E, F; green) and analyzed by confocal microscopy. Merged staining of EMMcorresponds to an area immediately adjacent to the perforation where the staining of bothfication. Note the co-localization of EMMPRIN and aSMA in the subepithelium in F (yellow).et al. (2008b).
(Verrecchia and Mauviel, 2007). By contrast, interleukin-1 mainlyprovokes tissue degradation through the upregulation of matrixdegrading proteinases (Girard et al., 1991). Since both cytokines aresecreted during injury by epithelial cells and were both describedto be present under the same circumstances, it is likely that
ulcerations showing induction and co-localization at the epithelial–stromal boundary.uble labeled immunohistochemistry using anti-EMMPRIN pAb (A, C, D, F; red) and anti-
PRIN and aSMA (C, F) revealed co-induction of EMMPRIN and aSMA. The square in CEMMPRIN and aSMA is particularly strong, presented in D, E and F at higher magni-
Original magnification: �20 (A, B, C); �40 (D, E, F). Scale bar 50 mm. Reprint from Huet

Fig. 7. Confocal immunohistochemistry of EMMPRIN and aSMA in normal and wounded mouse cornea. In normal cornea (A, B, C), EMMPRIN (red) is expressed mainly in theepithelium and the endothelium. aSMA (green) was not detected. In ulcerated mouse cornea (D, E, F), EMMPRIN and aSMA are induced in the stroma (D and E) where they are co-localized (yellow, F). Bar 50 mm.
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 27
another level of regulation exists which determines the outcome.In vitro studies suggest that when added together, TGFb effectoutbalance that of IL1 resulting in the inhibition of collagenaseexpression (Mauviel et al., 1993). Extrapolation of these in vitroresults to situations involving cytokine mediated ESI in vivo wouldargue for an overriding effect of TGFb, promoting myofibroblastsdifferentiation and matrix accumulation and leading to a fibroticoutcome.
When direct-ESI are involved, such as in chronic corneal ulcer-ation, then EMMPRIN would also enter into play and influence theoutcome of the healing process. Epithelial EMMPRIN is a potentinducer of EMMPRIN expression in fibroblasts through direct cell–cell contact. The increased EMMPRIN in the stroma would thenexert its effects on tissue remodeling by promoting both myofi-broblast differentiation and upregulation of proteinases, thusincreasing localized proteolysis. This EMMPRIN effect, which wouldprevail in chronic situations where healing is delayed and direct-ESI persist, may account for the presence of aSMA containingmyofibroblasts in ulcerated corneas where matrix degradationpredominates (Huet et al., 2008a,b).
Our findings that both TGFb and IL1 upregulate EMMPRINexpression in corneal fibroblasts represent an added complexity inthe interplay between these two cytokines in respect to tissueremodeling. The fact that the expression of EMMPRIN is inhibitedby blocking antibody to TGFb in low density migrating cells furthersuggests that EMMPRIN is constitutively under TGFb regulation(Huet et al., 2008a,b). This observation appears paradoxal sinceTGFb and EMMPRIN, although both inducing myofibroblastsdifferentiation, have opposing effect on ECM remodeling. This isclearly shown in Fig. 8 comparing the effects of TGFb and EMMPRINon the same corneal fibroblasts cell line, HTK (Jester et al., 2003)under identical conditions. While both TGFb and EMMPRINincrease aSMA, the differentiated fibroblasts that ensue produce
more MMP-1 under EMMPRIN’s effect while this proteinase isgreatly inhibited by TGFb, which in addition increases the expres-sion of its inhibitor TIMP-1. Furthermore, although EMMPRIN isknown as an MMP inducer, Fig. 8 shows its ability to also greatlyincrease the expression of the urokinase type plasminogen acti-vator (uPA) in the corneal fibroblasts, as we recently reported incancer cells (Quemener et al., 2007), expanding its proteolyticpower in the corneal stroma. Like MMP-1, urokinase is alsoinhibited by TGFb.
It is tempting therefore to speculate on the existence of severaltypes of myofibroblasts with different and even opposing effectson the ECM: those with ECM synthetic phenotype present in mostfibrotic tissues and driven more by TGFb, and those found inulcerated tissues having an increased degradative potential due toa greater EMMPRIN effect. This suggests that aSMA expressionby fibroblasts, which has been generally used as a marker offibrosis, may be uncoupled from their ability to accumulate ECM.Indeed, aSMA-containing myofibroblasts with ‘‘degradative’’phenotype, expressing high levels of matrix degrading protein-ases, are abundant in ulcerated corneas (Figs. 6 and 7). They arealso localized in the stroma surrounding many cancer tissues withthe potential to promote tumor invasion through tissue degra-dation (Kataoka et al., 2003; Jodele et al., 2006). Thus, the cellularpathways leading to the expression of aSMA may be differentfrom those of tissue remodeling and the acquisition of myofi-broblasts phenotype, characterized by the expression of aSMA,may be more representative of the contractile role of these cells asaccumulated evidence indicates that the main role of the smooth-muscle like features of myofibroblasts is in the force generationwhich results in wound contraction (Desmouliere et al., 2005).The effect of myofibroblasts on matrix remodeling may thendepend, among other factors, on the levels of TGFb and EMMPRIN,tilting the balance towards synthesis or degradation respectively.
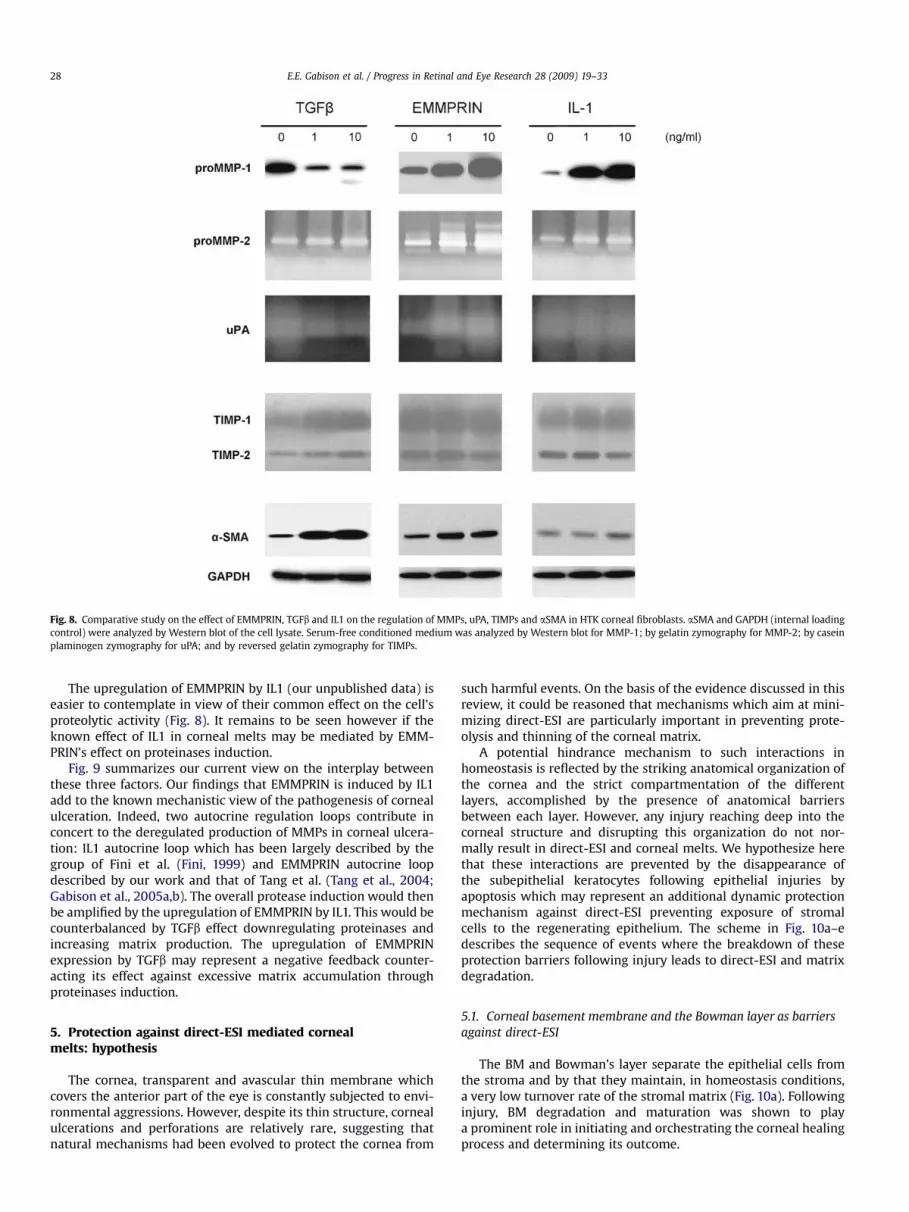
Fig. 8. Comparative study on the effect of EMMPRIN, TGFb and IL1 on the regulation of MMPs, uPA, TIMPs and aSMA in HTK corneal fibroblasts. aSMA and GAPDH (internal loadingcontrol) were analyzed by Western blot of the cell lysate. Serum-free conditioned medium was analyzed by Western blot for MMP-1; by gelatin zymography for MMP-2; by caseinplaminogen zymography for uPA; and by reversed gelatin zymography for TIMPs.
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3328
The upregulation of EMMPRIN by IL1 (our unpublished data) iseasier to contemplate in view of their common effect on the cell’sproteolytic activity (Fig. 8). It remains to be seen however if theknown effect of IL1 in corneal melts may be mediated by EMM-PRIN’s effect on proteinases induction.
Fig. 9 summarizes our current view on the interplay betweenthese three factors. Our findings that EMMPRIN is induced by IL1add to the known mechanistic view of the pathogenesis of cornealulceration. Indeed, two autocrine regulation loops contribute inconcert to the deregulated production of MMPs in corneal ulcera-tion: IL1 autocrine loop which has been largely described by thegroup of Fini et al. (Fini, 1999) and EMMPRIN autocrine loopdescribed by our work and that of Tang et al. (Tang et al., 2004;Gabison et al., 2005a,b). The overall protease induction would thenbe amplified by the upregulation of EMMPRIN by IL1. This would becounterbalanced by TGFb effect downregulating proteinases andincreasing matrix production. The upregulation of EMMPRINexpression by TGFb may represent a negative feedback counter-acting its effect against excessive matrix accumulation throughproteinases induction.
5. Protection against direct-ESI mediated cornealmelts: hypothesis
The cornea, transparent and avascular thin membrane whichcovers the anterior part of the eye is constantly subjected to envi-ronmental aggressions. However, despite its thin structure, cornealulcerations and perforations are relatively rare, suggesting thatnatural mechanisms had been evolved to protect the cornea from
such harmful events. On the basis of the evidence discussed in thisreview, it could be reasoned that mechanisms which aim at mini-mizing direct-ESI are particularly important in preventing prote-olysis and thinning of the corneal matrix.
A potential hindrance mechanism to such interactions inhomeostasis is reflected by the striking anatomical organization ofthe cornea and the strict compartmentation of the differentlayers, accomplished by the presence of anatomical barriersbetween each layer. However, any injury reaching deep into thecorneal structure and disrupting this organization do not nor-mally result in direct-ESI and corneal melts. We hypothesize herethat these interactions are prevented by the disappearance ofthe subepithelial keratocytes following epithelial injuries byapoptosis which may represent an additional dynamic protectionmechanism against direct-ESI preventing exposure of stromalcells to the regenerating epithelium. The scheme in Fig. 10a–edescribes the sequence of events where the breakdown of theseprotection barriers following injury leads to direct-ESI and matrixdegradation.
5.1. Corneal basement membrane and the Bowman layer as barriersagainst direct-ESI
The BM and Bowman’s layer separate the epithelial cells fromthe stroma and by that they maintain, in homeostasis conditions,a very low turnover rate of the stromal matrix (Fig. 10a). Followinginjury, BM degradation and maturation was shown to playa prominent role in initiating and orchestrating the corneal healingprocess and determining its outcome.

Fig. 9. Interplays between EMMPRIN, IL1 and TGFb and their consequences in ECM remodeling.
E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 29
Perturbation in the proper regeneration of the BM after injuryand therefore in the restoration of the separation barriers betweenthe epithelium and the stroma was shown to be associated withincreased proteolysis in the stroma. Indeed, clinical evidencesuggests that delayed epithelial resurfacing after injury, whichusually precedes stromal ulceration, is often accompanied bydissolution and deficient regeneration of the BM through a mech-anism which is yet to be identified (Fig. 10c). The consequence of
Fig. 10. Direct epithelial–stromal interactions: preventions and consequences. Schemati
delayed epithelial healing was demonstrated in a mouse modelwhere epithelial debridement initially preserving the integrity ofthe BM followed by BM degradation when healing was delayed bymore than 24 h. Under these circumstances, the BM exposed by thedebridement is thought to be cleaved by proteinases derived fromthe migrating epithelial cells (Sta Iglesia and Stepp, 2000).Furthermore, epithelial-derived MMPs were shown to further delaywound healing and to potentiate BM degradation (Fini et al., 1996).
c diagram representing the sequence of events leading to direct-ESI in the cornea.

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3330
Hence, delayed epithelial repair would facilitate direct-ESI by thedestruction of the BM, the immediate subepithelial barrier.
Bowman’s layer, a 8–15 mm thick band composed of an acellularextracellular matrix, represents an additional protective barrieragainst direct-ESI. Studies on the role of Bowman’s layer are lack-ing, probably because of the absence of this structure in thecommonly used animal models for corneal ulcerations, such asrodents and rabbit. This species variation which appears to be withno apparent biological consequence led to a debate regarding itsbiological role in corneal physiopathology. The group of Wilson donot consider Bowman’s layer as an essential structure with a crucialbiological function, in view of the evidence that the practice of PRKin human, which cuts through the Bowman layer, is not normallyassociated with complications following wound healing, despitethe fact that this structure is not regenerated following surgery(Wilson and Hong, 2000; Obata and Tsuru, 2007). Others, includingthe group of Jester, suggested that Bowman’s layer may providebiomechanical rigidity as well as define the corneal shape andcurvature (Morishige et al., 2007). However, until studies becomeavailable to either confirm or dispute such assumptions, theconcept of direct-ESI as a mechanism of proteases induction confersto Bowman’s layer a potential role as a barrier preventing suchinteractions. Thus, although the degradation of the BM is recog-nized to be a prerequisite in the pathogenesis of corneal ulceration(Matsubara et al., 1991), it may not be sufficient, requiring anadditional step of Bowman’s layer damage to allow direct-ESI andproviding a further delay in the transition from a PED to a cornealulceration (Fig. 10d).
Amniotic membrane transplantation is routinely used toimprove healing and prevent stromal melts and is generallythought to serve as a replacement of the damaged basementmembrane. In this context, the amniotic membrane was shown notonly to accelerate wound closure but also to inhibit MMPs’production and activity (Heiligenhaus et al., 2005; Takahashi et al.,2007). The assumption that the prevention of stromal melts asso-ciated with amniotic membrane transplantation resides solely inthe replacement of the missing BM is however challenged by invitro studies with organ cultures showing that the BM componentsof these amniotic membranes are in fact degraded prior toepithelial colonization of the transplanted membrane and thesynthesis of a new BM (Li et al., 2006). It is tempting to suggest thatthe acellular stroma, also contained within the amnioticmembranes, may provide an additional advantage by serving asa barrier against direct-ESI. Indeed, histological examinations haveclearly shown the persistence of an acellular zone even monthsfollowing transplantation. Hence, the benefits of grafting amnioticmembrane may lie not only in replacing the degraded BM but alsoin providing a Bowman layer like structure to serve as a barrieragainst direct-ESI (Seitz et al., 2006).
In addition to corneal ulceration, other clinical conditionssupport the concept that the breakdown of these protectingbarriers and direct-ESI are the basis of stromal melts. These includefor example epithelial ingrowth, a complication of LASIK whichinvolves a postoperative invasion of surface epithelial cells underthe corneal flap (Melki and Azar, 2001). Little is known on thephysiopathology of this complication which remains in most casesmild and self-limiting (Netto et al., 2005). However, in some severecases, a delay in surgical removal can be associated with flap melt(Castillo et al., 1998), presumably related to the persistence of theepithelial ingrowth within the stroma allowing direct-ESI. Epithe-lial plugs within the stroma can also result after radial keratotomy,a surgical procedure involving deep stromal incisions. Theabnormal BM and the absence of Bowman’s layer regenerationfollowing such surgery may favor direct-ESI and explain certainwound healing defects observed with this technique (Durand et al.,1992; Maguen et al., 1997; Ljubimov et al., 1998). Keratoconus (KC)
is another clinical situation where the rupture of BM and Bowman’slayer is a consistent early characteristic, associated with enhancedactivity of variety of proteases and where direct contact betweenepithelial and stromal cells has been observed (Kenney et al., 1997;Morishige et al., 2007). The increased EMMPRIN levels observed inthe stroma of KC samples, in particular at areas of rupture of bothBM and Bowman’s layer (Seppala et al., 2006), provides furthersupport for the concept of an EMMPRIN-mediated direct-ESI asa mechanism for the MMPs’ induction responsible for this type ofcorneal dystrophy.
5.2. Additional role for stromal cells’ apoptosis as a dynamic barrierto the healing epithelium?
Traumatic or surgical epithelial–stromal injuries, even thoughinvolving disruption of both the basement membrane and Bow-man’s layer, only rarely lead to ulcerations unless delayed epithelialclosure occurs, suggesting an additional mechanism by whichdelayed healing promotes stromal degradation.
Following epithelial wounds, a disappearance of keratocytes isobserved in the vicinity of the injured epithelial cells creating anacellular region between corneal epithelium and stromal cells.Dohlman et al. (1968) have already reported the phenomenon inthe late 1960s but the mechanism involved was only later identifiedby Wilson et al. (2001) as an apoptosis of the keratocytes inresponse to epithelial signals mediated by cytokines such as thesoluble IL1 and the FasLigand. The authors suggested that the mainfunction of the keratocyte apoptosis is to inhibit viruses infectingthe epithelium from spreading into the stroma. They also suggestedthat apoptosis may initiate proliferation in neighboring fibroblastsand promote wound healing (Wilson, 2002). We propose here thatthe disappearance of the keratocytes from the anterior stroma mayalso serve to prevent direct-ESI once the BM and Bowman’s layerare disrupted (Fig. 10b).
Investigations into the chronological sequence of the healingresponse following epithelial injury demonstrated that epithelialhealing consistently precedes the repopulation by stromal cells inthe area of initial cell death (Zieske et al., 2001; Hutcheon et al.,2005). This asynchronous cellular response following injury createsan acellular area between regenerating epithelium and stroma,representing a dynamic barrier which may prevent direct-ESIunder normal healing conditions. Physiological healing thenfollows with epithelial resurfacing, stratification and BM matura-tion, coinciding with the disappearance of activated myofibroblastsfrom the anterior stroma and the recovery of their semi-quiescentstate (Fini and Stramer, 2005). Hence, during physiological woundhealing, direct contact between epithelial and ‘‘post-apoptotic’’stromal cells is averted by the restoration of the BM as an anatomicepithelial–stromal barrier.
By contrast, situations associated with delayed epithelial heal-ing, as encountered in persistent epithelial defect, often lead tocorneal ulceration. In such circumstances, direct-ESI are regularlyobserved, in particular at the edges of the ulceration (Gabison et al.,2003, 2005a,b). This prompts us to suggest that delayed epithelialwound closure would favor direct-ESI by allowing corneal fibro-blasts to regain and repopulate the anterior stroma following theinitial apoptotic phase and to come into direct contact with theepithelium (Fig. 10e). Epithelial EMMPRIN would then activatefibroblasts through direct cell–cell interaction and signal them toincrease their production of MMPs. In addition, as EMMPRIN alsoinduces its own production in neighboring cells, it can propagatethis MMP stimulation deeper in the stroma. The combined prote-olysis promoting effects of EMMPRIN with those of IL1 duringdirect-ESI would then further amplify proteolysis and stromal melt.
Hence, the BM, Bowman’s layer and fibroblasts’ apoptosisfollowing epithelial injury may represent both anatomical and

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 31
dynamic protection mechanisms against direct-ESI in homeostasisand physiological wound healing. It is possible that in response toinjury, the subepithelial fibroblast apoptosis may compensate thelack of Bowman’s layer in certain animal species and in post-PRKhuman corneas. However, the question of whether, in situations ofpersistent epithelial defect, the absence of Bowman’s layer mayconfer increased susceptibility to corneal melts remains to beaddressed.
6. Conclusions and future directions
The implication of MMP induction in the pathogenesis ofcorneal ulceration is well documented but their inhibition in theclinic has not yet proven its efficiency in this context. Indeed, MMPinhibition appears to confront several challenges, mainly due to thediversity of functions which are increasingly attributed to theseproteinases both in physiological and pathological situations. Thismeans therefore that the potential advantage which could beobtained by their inhibition would be counterbalanced by the lossof their physiological functions necessary for the normal woundhealing process. The example of MMP-9, which appears as anattractive target due to its major implication in the initiating stepsin corneal ulceration, illustrates this point as its inhibition couldalso result in corneal opacification, as suggested from observationsin the MMP-9 deficient mice following PRK (Mohan et al., 2002).The demonstration of corneal neovascularization following PRK inthe MMP-7 knockout mice also suggests that the inhibition of thisenzyme in clinical situations will be associated with undesired sideeffects (Kure et al., 2003). These examples demonstrate thecomplexity in specifically targeting pathological functions of theseenzymes.
The identification of EMMPRIN-mediated direct-ESI as a newmechanism of MMP induction during pathological wound healingopens new avenues for MMP inhibition.
Targeting EMMPRIN, which induces MMPs that are specificallyincreased during pathological situations involving direct-ESI, mayrepresent an attractive new strategy for selective MMP inhibition.Furthermore, inhibiting EMMPRIN would inhibit MMPs even beforethey are produced, contrary to the synthetic inhibitors which aim atinhibiting their activity after secretion and activation. The thera-peutic inhibition of EMMPRIN appears therefore promising insituations of excess extracellular matrix degradation such ascorneal ulcerations, keratoconus and other chronic wound healingassociated with stromal melts.
Acknowledgments
The authors acknowledge the support of the Fondation deL’Avenir (study ET5-400 and ET8-489).
References
Berman, M., Leary, R., Gage, J., 1980. Evidence for a role of the plasminogen acti-vator–plasmin system in corneal ulceration. Investig. Ophthalmol. Vis. Sci. 19,1204–1221.
Berman, M., Manseau, E., Law, M., Aiken, D., 1983. Ulceration is correlated withdegradation of fibrin and fibronectin at the corneal surface. Investig. Oph-thalmol. Vis. Sci. 24, 1358–1366.
Berman, M.B., 1993. Regulation of corneal fibroblast MMP-1 collagenase secretionby plasmin. Cornea 12, 420–432.
Berman, M.B., 1994. Regulation of corneal fibroblast MMP-1 secretion by cytocha-lasins. Cornea 13, 51–57.
Biswas, C., Zhang, Y., DeCastro, R., Guo, H., Nakamura, T., Kataoka, H., Nabeshima, K.,1995. The human tumor cell-derived collagenase stimulatory factor (renamedEMMPRIN) is a member of the immunoglobulin superfamily. Cancer Res. 55,434–439.
Bordador, L.C., Li, X., Toole, B., Chen, B., Regezi, J., Zardi, L., Hu, Y., Ramos, D.M., 2000.Expression of EMMPRIN by oral squamous cell carcinoma. Int. J. Cancer 85,347–352.
Carrington, L.M., Boulton, M., 2005. Hepatocyte growth factor and keratinocytegrowth factor regulation of epithelial and stromal corneal wound healing.J. Cataract Refract. Surg. 31, 412–423.
Castillo, A., Diaz-Valle, D., Gutierrez, A.R., Toledano, N., Romero, F., 1998. Peripheralmelt of flap after laser in situ keratomileusis. J. Refract. Surg. 14, 61–63.
Caudroy, S., Polette, M., Tournier, J.M., Burlet, H., Toole, B., Zucker, S., Birembaut, P.,1999. Expression of the extracellular matrix metalloproteinase inducer(EMMPRIN) and the matrix metalloproteinase-2 in bronchopulmonary andbreast lesions. J. Histochem. Cytochem. 47, 1575–1580.
Daniels, J.T., Geerling, G., Alexander, R.A., Murphy, G., Khaw, P.T., Saarialho-Kere, U.,2003a. Temporal and spatial expression of matrix metalloproteinases duringwound healing of human corneal tissue. Exp. Eye Res. 77, 653–664.
Daniels, J.T., Khaw, P.T., 2000. Temporal stimulation of corneal fibroblast woundhealing activity by differentiating epithelium in vitro. Investig. Ophthalmol. Vis.Sci. 41, 3754–3762.
Daniels, J.T., Limb, G.A., Saarialho-Kere, U., Murphy, G., Khaw, P.T., 2003b. Humancorneal epithelial cells require MMP-1 for HGF-mediated migration on collagenI. Investig. Ophthalmol. Vis. Sci. 44, 1048–1055.
Daniels, J.T., Schultz, G.S., Blalock, T.D., Garrett, Q., Grotendorst, G.R., Dean, N.M.,Khaw, P.T., 2003c. Mediation of transforming growth factor-beta(1)-stimulatedmatrix contraction by fibroblasts: a role for connective tissue growth factor incontractile scarring. Am. J. Pathol. 163, 2043–2052.
Davidson, B., Givant-Horwitz, V., Lazarovici, P., Risberg, B., Nesland, J.M., Trope, C.G.,Schaefer, E., Reich, R., 2003a. Matrix metalloproteinases (MMP), EMMPRIN(extracellular matrix metalloproteinase inducer) and mitogen-activated proteinkinases (MAPK): co-expression in metastatic serous ovarian carcinoma. Clin.Exp. Metastasis 20, 621–631.
Davidson, B., Goldberg, I., Berner, A., Kristensen, G.B., Reich, R., 2003b. EMMPRIN(extracellular matrix metalloproteinase inducer) is a novel marker of pooroutcome in serous ovarian carcinoma. Clin. Exp. Metastasis 20, 161–169.
Desmouliere, A., Chaponnier, C., Gabbiani, G., 2005. Tissue repair, contraction, andthe myofibroblast. Wound Repair Regen. 13, 7–12.
Dohlman, C.H., Gasset, A.R., Rose, J., 1968. The effect of the absence of cornealepithelium or endothelium on the stromal keratocytes. Investig. Ophthalmol. 7,520–534.
Durand, L., Monnot, J.P., Burillon, C., Assi, A., 1992. Complications of radial kera-totomy: eyes with keratoconus and late wound dehiscence. Refract. CornealSurg. 8, 311–314.
Eming, S.A., Yarmush, M.L., Krueger, G.G., Morgan, J.R., 1999. Regulation of the spatialorganization of mesenchymal connective tissue: effects of cell-associated versusreleased isoforms of platelet-derived growth factor. Am. J. Pathol. 154, 281–289.
Fini, M.E., 1999. Keratocyte and fibroblast phenotypes in the repairing cornea. Prog.Retin. Eye Res. 18, 529–551.
Fini, M.E., Cook, J.R., Mohan, R., 1998. Proteolytic mechanisms in corneal ulcerationand repair. Arch. Dermatol. Res. 290 (Suppl.), S12–S23.
Fini, M.E., Girard, M.T., Matsubara, M., 1992. Collagenolytic/gelatinolytic enzymes incorneal wound healing. Acta Ophthalmol. Suppl., 26–33.
Fini, M.E., Parks, W.C., Rinehart, W.B., Girard, M.T., Matsubara, M., Cook, J.R., West-Mays, J.A., Sadow, P.M., Burgeson, R.E., Jeffrey, J.J., Raizman, M.B., Krueger, R.R.,Zieske, J.D., 1996. Role of matrix metalloproteinases in failure to re-epithelializeafter corneal injury. Am. J. Pathol. 149, 1287–1302.
Fini, M.E., Stramer, B.M., 2005. How the cornea heals: cornea-specific repairmechanisms affecting surgical outcomes. Cornea 24, S2–S11.
Foda, H.D., Rollo, E.E., Drews, M., Conner, C., Appelt, K., Shalinsky, D.R., Zucker, S.,2001. Ventilator-induced lung injury upregulates and activates gelatinases andEMMPRIN: attenuation by the synthetic matrix metalloproteinase inhibitor,Prinomastat (AG3340). Am. J. Respir. Cell Mol. Biol. 25, 717–724.
Folger, P.A., Zekaria, D., Grotendorst, G., Masur, S.K., 2001. Transforming growthfactor-beta-stimulated connective tissue growth factor expression duringcorneal myofibroblast differentiation. Investig. Ophthalmol. Vis. Sci. 42, 2534–2541.
Gabison, E.E., Chastang, P., Menashi, S., Mourah, S., Doan, S., Oster, M., Mauviel, A.,Hoang-Xuan, T., 2003. Late corneal perforation after photorefractive keratec-tomy associated with topical diclofenac: involvement of matrix metal-loproteinases. Ophthalmology 110, 1626–1631.
Gabison, E.E., Hoang-Xuan, T., Mauviel, A., Menashi, S., 2005a. EMMPRIN/CD147, anMMP modulator in cancer, development and tissue repair. Biochimie 87,361–368.
Gabison, E.E., Mourah, S., Steinfels, E., Yan, L., Hoang-Xuan, T., Watsky, M.A., DeWever, B., Calvo, F., Mauviel, A., Menashi, S., 2005b. Differential expression ofextracellular matrix metalloproteinase inducer (CD147) in normal and ulcer-ated corneas: role in epithelio-stromal interactions and matrix metal-loproteinase induction. Am. J. Pathol. 166, 209–219.
Garrett, Q., Khaw, P.T., Blalock, T.D., Schultz, G.S., Grotendorst, G.R., Daniels, J.T.,2004. Involvement of CTGF in TGF-beta1-stimulation of myofibroblast differ-entiation and collagen matrix contraction in the presence of mechanical stress.Investig. Ophthalmol. Vis. Sci. 45, 1109–1116.
Geerling, G., Joussen, A.M., Daniels, J.T., Mulholland, B., Khaw, P.T., Dart, J.K., 1999.Matrix metalloproteinases in sterile corneal melts. Ann. N.Y. Acad. Sci. 878,571–574.
Girard, M.T., Matsubara, M., Fini, M.E., 1991. Transforming growth factor-beta andinterleukin-1 modulate metalloproteinase expression by corneal stromal cells.Investig. Ophthalmol. Vis. Sci. 32, 2441–2454.
Girard, M.T., Matsubara, M., Kublin, C., Tessier, M.J., Cintron, C., Fini, M.E., 1993.Stromal fibroblasts synthesize collagenase and stromelysin during long-termtissue remodeling. J. Cell Sci. 104 (Pt 4), 1001–1011.

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–3332
Gohring, W., Sasaki, T., Heldin, C.H., Timpl, R., 1998. Mapping of the binding ofplatelet-derived growth factor to distinct domains of the basement membraneproteins BM-40 and perlecan and distinction from the BM-40 collagen-bindingepitope. Eur. J. Biochem. 255, 60–66.
Gordon, J.M., Bauer, E.A., Eisen, A.Z., 1980. Collagenase in human cornea: immu-nologic localization. Arch. Ophthalmol. 98, 341–345.
Heiligenhaus, A., Li, H.F., Yang, Y., Wasmuth, S., Steuhl, K.P., Bauer, D., 2005.Transplantation of amniotic membrane in murine herpes stromal keratitismodulates matrix metalloproteinases in the cornea. Investig. Ophthalmol. Vis.Sci. 46, 4079–4085.
Honma, Y., Nishida, K., Sotozono, C., Kinoshita, S., 1997. Effect of transforminggrowth factor-beta1 and -beta2 on in vitro rabbit corneal epithelial cellproliferation promoted by epidermal growth factor, keratinocyte growth factor,or hepatocyte growth factor. Exp. Eye Res. 65, 391–396.
Huet, E., Gabison, E.E., Mourah, S., Menashi, S., 2008a. Role of EMMPRIN/CD147 intissue remodeling. Connect. Tissue Res. 49, 175–179.
Huet, E., Vallee, B., Szul, D., Verrecchia, F., Mourah, S., Jester, J.V., Hoang-Xuan, T.,Menashi, S., Gabison, E.E., 2008b. Extracellular matrix metalloproteinaseinducer/CD147 promotes myofibroblast differentiation by inducing alpha-smooth muscle actin expression and collagen gel contraction: implications intissue remodeling. FASEB J. 22, 1144–1154.
Hutcheon, A.E., Guo, X.Q., Stepp, M.A., Simon, K.J., Weinreb, P.H., Violette, S.M.,Zieske, J.D., 2005. Effect of wound type on Smad 2 and 4 translocation. Investig.Ophthalmol. Vis. Sci. 46, 2362–2368.
Igakura, T., Kadomatsu, K., Kaname, T., Muramatsu, H., Fan, Q.W., Miyauchi, T.,Toyama, Y., Kuno, N., Yuasa, S., Takahashi, M., Senda, T., Taguchi, O.,Yamamura, K., Arimura, K., Muramatsu, T., 1998. A null mutation in basigin, animmunoglobulin superfamily member, indicates its important roles in peri-implantation development and spermatogenesis. Dev. Biol. 194, 152–165.
Igakura, T., Kadomatsu, K., Taguchi, O., Muramatsu, H., Kaname, T., Miyauchi, T.,Yamamura, K., Arimura, K., Muramatsu, T.,1996. Roles of basigin, a member of theimmunoglobulin superfamily, in behavior as to an irritating odor, lymphocyteresponse, and blood–brain barrier. Biochem. Biophys. Res. Commun. 224, 33–36.
Imanishi, J., Kamiyama, K., Iguchi, I., Kita, M., Sotozono, C., Kinoshita, S., 2000.Growth factors: importance in wound healing and maintenance of trans-parency of the cornea. Prog. Retin. Eye Res. 19, 113–129.
Ishibashi, Y., Matsumoto, T., Niwa, M., Suzuki, Y., Omura, N., Hanyu, N., Nakada, K.,Yanaga, K., Yamada, K., Ohkawa, K., Kawakami, M., Urashima, M., 2004. CD147and matrix metalloproteinase-2 protein expression as significant prognosticfactors in esophageal squamous cell carcinoma. Cancer 101, 1994–2000.
Javier, J.A., Lee, J.B., Oliveira, H.B., Chang, J.H., Azar, D.T., 2006. Basement membraneand collagen deposition after laser subepithelial keratomileusis and photo-refractive keratectomy in the leghorn chick eye. Arch. Ophthalmol. 124,703–709.
Jester, J.V., Barry-Lane, P.A., Petroll, W.M., Olsen, D.R., Cavanagh, H.D., 1997. Inhibi-tion of corneal fibrosis by topical application of blocking antibodies to TGF betain the rabbit. Cornea 16, 177–187.
Jester, J.V., Ho-Chang, J., 2003. Modulation of cultured corneal keratocyte phenotypeby growth factors/cytokines control in vitro contractility and extracellularmatrix contraction. Exp. Eye Res. 77, 581–592.
Jester, J.V., Huang, J., Fisher, S., Spiekerman, J., Chang, J.H., Wright, W.E., Shay, J.W.,2003. Myofibroblast differentiation of normal human keratocytes and hTERT,extended-life human corneal fibroblasts. Investig. Ophthalmol. Vis. Sci. 44,1850–1858.
Jester, J.V., Petroll, W.M., Barry, P.A., Cavanagh, H.D., 1995. Expression of alpha-smooth muscle (alpha-SM) actin during corneal stromal wound healing.Investig. Ophthalmol. Vis. Sci. 36, 809–819.
Jester, J.V., Petroll, W.M., Cavanagh, H.D., 1999. Corneal stromal wound healing inrefractive surgery: the role of myofibroblasts. Prog. Retin. Eye Res. 18, 311–356.
Jodele, S., Blavier, L., Yoon, J.M., DeClerck, Y.A., 2006. Modifying the soil to affect theseed: role of stromal-derived matrix metalloproteinases in cancer progression.Cancer Metastasis Rev. 25, 35–43.
Johnson-Muller, B., Gross, J., 1978. Regulation of corneal collagenase production:epithelial–stromal cell interactions. Proc. Natl. Acad. Sci. U.S.A. 75, 4417–4421.
Jung, J.C., Huh, M.I., Fini, M.E., 2007. Constitutive collagenase-1 synthesis throughMAPK pathways is mediated, in part, by endogenous IL-1alpha during fibroticrepair in corneal stroma. J. Cell. Biochem. 102, 453–462.
Kanekura, T., Chen, X., Kanzaki, T., 2002. Basigin (CD147) is expressed on melanomacells and induces tumor cell invasion by stimulating production of matrixmetalloproteinases by fibroblasts. Int. J. Cancer 99, 520–528.
Kataoka, H., DeCastro, R., Zucker, S., Biswas, C., 1993. Tumor cell-derived collage-nase-stimulatory factor increases expression of interstitial collagenase, stro-melysin, and 72-kDa gelatinase. Cancer Res. 53, 3154–3158.
Kataoka, H., Tanaka, H., Nagaike, K., Uchiyama, S., Itoh, H., 2003. Role of cancer cell–stroma interaction in invasive growth of cancer cells. Hum. Cell 16, 1–14.
Kawakita, T., Espana, E.M., He, H., Hornia, A., Yeh, L.K., Ouyang, J., Liu, C.Y.,Tseng, S.C., 2005. Keratocan expression of murine keratocytes is maintained onamniotic membrane by down-regulating transforming growth factor-betasignaling. J. Biol. Chem. 280, 27085–27092.
Kenney, M.C., Nesburn, A.B., Burgeson, R.E., Butkowski, R.J., Ljubimov, A.V., 1997.Abnormalities of the extracellular matrix in keratoconus corneas. Cornea 16,345–351.
Kim, W.J., Mohan, R.R., Mohan, R.R., Wilson, S.E., 1999. Effect of PDGF, IL-1alpha,and BMP2/4 on corneal fibroblast chemotaxis: expression of the platelet-derived growth factor system in the cornea. Investig. Ophthalmol. Vis. Sci. 40,1364–1372.
Konttinen, Y.T., Li, T.F., Mandelin, J., Liljestrom, M., Sorsa, T., Santavirta, S.,Virtanen, I., 2000. Increased expression of extracellular matrix metal-loproteinase inducer in rheumatoid synovium. Arthritis Rheum. 43, 275–280.
Kure, T., Chang, J.H., Kato, T., Hernandez-Quintela, E., Ye, H., Lu, P.C., Matrisian, L.M.,Gatinel, D., Shapiro, S., Gosheh, F., Azar, D.T., 2003. Corneal neovascularizationafter excimer keratectomy wounds in matrilysin-deficient mice. Investig.Ophthalmol. Vis. Sci. 44, 137–144.
LaGier, A.J., Yoo, S.H., Alfonso, E.C., Meiners, S., Fini, M.E., 2007. Inhibition of humancorneal epithelial production of fibrotic mediator TGF-beta2 by basementmembrane-like extracellular matrix. Investig. Ophthalmol. Vis. Sci. 48,1061–1071.
Lee, J.B., Choe, C.M., Kim, H.S., Seo, K.Y., Seong, G.J., Kim, E.K., 2002. Comparison ofTGF-beta1 in tears following laser subepithelial keratomileusis and photo-refractive keratectomy. J. Refract. Surg. 18, 130–134.
Li, D.Q., Tseng, S.C., 1995. Three patterns of cytokine expression potentially involvedin epithelial–fibroblast interactions of human ocular surface. J. Cell. Physiol. 163,61–79.
Li, W., He, H., Kuo, C.L., Gao, Y., Kawakita, T., Tseng, S.C., 2006. Basement membranedissolution and reassembly by limbal corneal epithelial cells expanded onamniotic membrane. Investig. Ophthalmol. Vis. Sci. 47, 2381–2389.
Ljubimov, A.V., Alba, S.A., Burgeson, R.E., Ninomiya, Y., Sado, Y., Sun, T.T.,Nesburn, A.B., Kenney, M.C., Maguen, E., 1998. Extracellular matrix changes inhuman corneas after radial keratotomy. Exp. Eye Res. 67, 265–272.
Lu, P.C., Ye, H., Maeda, M., Azar, D.T., 1999. Immunolocalization and gene expressionof matrilysin during corneal wound healing. Investig. Ophthalmol. Vis. Sci. 40,20–27.
Ma, J.J., Dohlman, C.H., 2002. Mechanisms of corneal ulceration. Ophthalmol. Clin.North Am. 15, 27–33.
Maguen, E., Alba, S.A., Burgeson, R.E., Butkowski, R.J., Michael, A.F., Kenney, M.C.,Nesburn, A.B., Ljubimov, A.V., 1997. Alterations of corneal extracellular matrixafter multiple refractive procedures: a clinical and immunohistochemical study.Cornea 16, 675–682.
Major, T.C., Liang, L., Lu, X., Rosebury, W., Bocan, T.M., 2002. Extracellular matrixmetalloproteinase inducer (EMMPRIN) is induced upon monocyte differentia-tion and is expressed in human atheroma. Arterioscler. Thromb. Vasc. Biol. 22,1200–1207.
Masur, S.K., Dewal, H.S., Dinh, T.T., Erenburg, I., Petridou, S., 1996. Myofibroblastsdifferentiate from fibroblasts when plated at low density. Proc. Natl. Acad. Sci.U.S.A. 93, 4219–4223.
Matsubara, M., Zieske, J.D., Fini, M.E., 1991. Mechanism of basement membranedissolution preceding corneal ulceration. Investig. Ophthalmol. Vis. Sci. 32,3221–3237.
Mauviel, A., Qiu Chen, Y., Dong, W., Evans, C.H., Uitto, J., 1993. Transcriptionalinteractions of transforming growth-factor-beta with pro-inflammatory cyto-kines. Curr. Biol. 3, 822–831.
Melki, S.A., Azar, D.T., 2001. LASIK complications: etiology, management, andprevention. Surv. Ophthalmol. 46, 95–116.
Mita, T., Yamashita, H., Kaji, Y., Obata, H., Yamada, H., Kato, M., Hanyu, A.,Suzuki, M., Tobari, I., 1998. Effects of transforming growth factor beta oncorneal epithelial and stromal cell function in a rat wound healing modelafter excimer laser keratectomy. Graefe’s Arch. Clin. Exp. Ophthalmol. 236,834–843.
Mohan, R., Chintala, S.K., Jung, J.C., Villar, W.V., McCabe, F., Russo, L.A., Lee, Y.,McCarthy, B.E., Wollenberg, K.R., Jester, J.V., Wang, M., Welgus, H.G.,Shipley, J.M., Senior, R.M., Fini, M.E., 2002. Matrix metalloproteinase gelatinaseB (MMP-9) coordinates and effects epithelial regeneration. J. Biol. Chem. 277,2065–2072.
Moore, J.E., McMullen, T.C., Campbell, I.L., Rohan, R., Kaji, Y., Afshari, N.A., Usui, T.,Archer, D.B., Adamis, A.P., 2002. The inflammatory milieu associated withconjunctivalized cornea and its alteration with IL-1 RA gene therapy. Investig.Ophthalmol. Vis. Sci. 43, 2905–2915.
Morishige, N., Wahlert, A.J., Kenney, M.C., Brown, D.J., Kawamoto, K., Chikama, T.,Nishida, T., Jester, J.V., 2007. Second-harmonic imaging microscopy ofnormal human and keratoconus cornea. Investig. Ophthalmol. Vis. Sci. 48,1087–1094.
Muramatsu, T., Miyauchi, T., 2003. Basigin (CD147): a multifunctional trans-membrane protein involved in reproduction, neural function, inflammation andtumor invasion. Histol. Histopathol. 18, 981–987.
Nabeshima, K., Iwasaki, H., Koga, K., Hojo, H., Suzumiya, J., Kikuchi, M., 2006.EMMPRIN (basigin/CD147): matrix metalloproteinase modulator and multi-functional cell recognition molecule that plays a critical role in cancerprogression. Pathol. Int. 56, 359–367.
Nakamura, K., 2003. Interaction between injured corneal epithelial cells andstromal cells. Cornea 22, S35–S47.
Nakamura, K., Kurosaka, D., Yoshino, M., Oshima, T., Kurosaka, H., 2002. Injuredcorneal epithelial cells promote myodifferentiation of corneal fibroblasts.Investig. Ophthalmol. Vis. Sci. 43, 2603–2608.
Netto, M.V., Mohan, R.R., Ambrosio Jr., R., Hutcheon, A.E., Zieske, J.D., Wilson, S.E.,2005. Wound healing in the cornea: a review of refractive surgery complica-tions and new prospects for therapy. Cornea 24, 509–522.
Netto, M.V., Mohan, R.R., Sinha, S., Sharma, A., Dupps, W., Wilson, S.E., 2006.Stromal haze, myofibroblasts, and surface irregularity after PRK. Exp. Eye Res.82, 788–797.
Obata, H., Tsuru, T., 2007. Corneal wound healing from the perspective of kerato-plasty specimens with special reference to the function of the Bowman layerand Descemet membrane. Cornea 26, S82–S89.

E.E. Gabison et al. / Progress in Retinal and Eye Research 28 (2009) 19–33 33
Petridou, S., Maltseva, O., Spanakis, S., Masur, S.K., 2000. TGF-beta receptorexpression and smad2 localization are cell density dependent in fibroblasts.Investig. Ophthalmol. Vis. Sci. 41, 89–95.
Polette, M., Gilles, C., Marchand, V., Lorenzato, M., Toole, B., Tournier, J.M., Zucker, S.,Birembaut, P., 1997. Tumor collagenase stimulatory factor (TCSF) expression andlocalization in human lung and breast cancers. J. Histochem. Cytochem. 45,703–709.
Quemener, C., Gabison, E.E., Naimi, B., Lescaille, G., Bougatef, F., Podgorniak, M.P.,Labarchede, G., Lebbe, C., Calvo, F., Menashi, S., Mourah, S., 2007. Extracellularmatrix metalloproteinase inducer up-regulates the urokinase-type plasmin-ogen activator system promoting tumor cell invasion. Cancer Res. 67, 9–15.
Riley, G.P., Harrall, R.L., Watson, P.G., Cawston, T.E., Hazleman, B.L., 1995. Collagenase(MMP-1) and TIMP-1 in destructive corneal disease associated with rheumatoidarthritis. Eye 9 (Pt 6), 703–718.
Rosenthal, E.L., Shreenivas, S., Peters, G.E., Grizzle, W.E., Desmond, R., Gladson, C.L.,2003. Expression of extracellular matrix metalloprotease inducer in laryngealsquamous cell carcinoma. Laryngoscope 113, 1406–1410.
Sameshima, T., Nabeshima, K., Toole, B.P., Yokogami, K., Okada, Y., Goya, T.,Koono, M., Wakisaka, S., 2000. Expression of EMMPRIN (CD147), a cell surfaceinducer of matrix metalloproteinases, in normal human brain and gliomas. Int.J. Cancer 88, 21–27.
Seitz, B., Resch, M.D., Schlotzer-Schrehardt, U., Hofmann-Rummelt, C., Sauer, R.,Kruse, F.E., 2006. Histopathology and ultrastructure of human corneas afteramniotic membrane transplantation. Arch. Ophthalmol. 124, 1487–1490.
Seppala, H.P., Maatta, M., Rautia, M., Mackiewicz, Z., Tuisku, I., Tervo, T.,Konttinen, Y.T., 2006. EMMPRIN and MMP-1 in keratoconus. Cornea 25, 325–330.
Shackel, N.A., McGuinness, P.H., Abbott, C.A., Gorrell, M.D., McCaughan, G.W., 2002.Insights into the pathobiology of hepatitis C virus-associated cirrhosis: analysisof intrahepatic differential gene expression. Am. J. Pathol. 160, 641–654.
Sivak, J.M., Fini, M.E., 2002. MMPs in the eye: emerging roles for matrix metal-loproteinases in ocular physiology. Prog. Retin. Eye Res. 21, 1–14.
Slansky, H.H., Gnadinger, M.C., Itoi, M., Dohlman, C.H., 1969. Collagenase in cornealulcerations. Arch. Ophthalmol. 82, 108–111.
Sotozono, C., Kinoshita, S., Kita, M., Imanishi, J., 1994. Paracrine role of keratinocytegrowth factor in rabbit corneal epithelial cell growth. Exp. Eye Res. 59, 385–391.
Spinale, F.G., Coker, M.L., Heung, L.J., Bond, B.R., Gunasinghe, H.R., Etoh, T.,Goldberg, A.T., Zellner, J.L., Crumbley, A.J., 2000. A matrix metalloproteinaseinduction/activation system exists in the human left ventricular myocardiumand is upregulated in heart failure. Circulation 102, 1944–1949.
Sta Iglesia, D.D., Stepp, M.A., 2000. Disruption of the basement membrane aftercorneal debridement. Investig. Ophthalmol. Vis. Sci. 41, 1045–1053.
Stapleton, W.M., Chaurasia, S.S., Medeiros, F.W., Mohan, R.R., Sinha, S., Wilson, S.E.,2008. Topical interleukin-1 receptor antagonist inhibits inflammatory cellinfiltration into the cornea. Exp. Eye Res. 86, 753–757.
Stramer, B.M., Zieske, J.D., Jung, J.C., Austin, J.S., Fini, M.E., 2003. Molecular mech-anisms controlling the fibrotic repair phenotype in cornea: implications forsurgical outcomes. Investig. Ophthalmol. Vis. Sci. 44, 4237–4246.
Sun, J., Hemler, M.E., 2001. Regulation of MMP-1 and MMP-2 production throughCD147/extracellular matrix metalloproteinase inducer interactions. Cancer Res.61, 2276–2281.
Takahashi, H., Igarashi, T., Fujimoto, C., Ozaki, N., Ishizaki, M., 2007. Immunohisto-chemical observation of amniotic membrane patching on a corneal alkali burnin vivo. Jpn. J. Ophthalmol. 51, 3–9.
Tang, Y., Kesavan, P., Nakada, M.T., Yan, L., 2004. Tumor–stroma interaction: positivefeedback regulation of extracellular matrix metalloproteinase inducer (EMM-PRIN) expression and matrix metalloproteinase-dependent generation ofsoluble EMMPRIN. Mol. Cancer Res. 2, 73–80.
Tang, Y., Nakada, M.T., Kesavan, P., McCabe, F., Millar, H., Rafferty, P., Bugelski, P.,Yan, L., 2005. Extracellular matrix metalloproteinase inducer stimulates tumorangiogenesis by elevating vascular endothelial cell growth factor and matrixmetalloproteinases. Cancer Res. 65, 3193–3199.
Thorns, C., Feller, A.C., Merz, H., 2002. EMMPRIN (CD 174) is expressed in Hodgkin’slymphoma and anaplastic large cell lymphoma. An immunohistochemical studyof 60 cases. Anticancer Res. 22, 1983–1986.
Toole, B.P., 2003. EMMPRIN (CD147), a cell surface regulator of matrix metal-loproteinase production and function. Curr. Top. Dev. Biol. 54, 371–389.
Toole, B.P., Slomiany, M.G., 2008. Hyaluronan, CD44 and EMMPRIN: partners incancer cell chemoresistance. Drug Resist. Updat. 11, 110–121.
Tseng, S.C., Li, D.Q., Ma, X., 1999. Suppression of transforming growth factor-betaisoforms, TGF-beta receptor type II, and myofibroblast differentiation in
cultured human corneal and limbal fibroblasts by amniotic membrane matrix.J. Cell. Physiol. 179, 325–335.
Verrecchia, F., Chu, M.L., Mauviel, A., 2001. Identification of novel TGF-beta/Smadgene targets in dermal fibroblasts using a combined cDNA microarray/promotertransactivation approach. J. Biol. Chem. 276, 17058–17062.
Verrecchia, F., Mauviel, A., 2007. Transforming growth factor-beta and fibrosis.World J. Gastroenterol. 13, 3056–3062.
Wagoner, M.D., Kenyon, K.R., 1989. Distribution of collagenase and cell types insterile ulceration of human corneal grafts. Acta Ophthalmol. Suppl. 192, 65–71.
Wang, M., Munier, F., Araki-Saski, K., Schorderet, D., 2002. TGFBI gene transcript istransforming growth factor-beta1-responsive and cell density-dependent ina human corneal epithelial cell line. Ophthalmic Genet. 23, 237–245.
Weiss, A., Goldman, S., Shalev, E., 2007. The matrix metalloproteinases (MMPs) inthe decidua and fetal membranes. Front. Biosci. 12, 649–659.
Weng, J., Mohan, R.R., Li, Q., Wilson, S.E., 1997. IL-1 upregulates keratinocyte growthfactor and hepatocyte growth factor mRNA and protein production by culturedstromal fibroblast cells: interleukin-1 beta expression in the cornea. Cornea 16,465–471.
Werner, S., Smola, H., Liao, X., Longaker, M.T., Krieg, T., Hofschneider, P.H.,Williams, L.T., 1994. The function of KGF in morphogenesis of epithelium andreepithelialization of wounds. Science 266, 819–822.
West-Mays, J.A., Cook, J.R., Sadow, P.M., Mullady, D.K., Bargagna-Mohan, P.,Strissel, K.J., Fini, M.E., 1999. Differential inhibition of collagenase and inter-leukin-1alpha gene expression in cultured corneal fibroblasts by TGF-beta,dexamethasone, and retinoic acid. Investig. Ophthalmol. Vis. Sci. 40, 887–896.
West-Mays, J.A., Sadow, P.M., Tobin, T.W., Strissel, K.J., Cintron, C., Fini, M.E., 1997.Repair phenotype in corneal fibroblasts is controlled by an interleukin-1 alphaautocrine feedback loop. Investig. Ophthalmol. Vis. Sci. 38, 1367–1379.
Wilson, S.E., 2002. Analysis of the keratocyte apoptosis, keratocyte proliferation,and myofibroblast transformation responses after photorefractive keratectomyand laser in situ keratomileusis. Trans. Am. Ophthalmol. Soc. 100, 411–433.
Wilson, S.E., Chen, L., Mohan, R.R., Liang, Q., Liu, J., 1999a. Expression of HGF, KGF,EGF and receptor messenger RNAs following corneal epithelial wounding. Exp.Eye Res. 68, 377–397.
Wilson, S.E., Hong, J.W., 2000. Bowman’s layer structure and function: critical ordispensable to corneal function? A hypothesis. Cornea 19, 417–420.
Wilson, S.E., Liu, J.J., Mohan, R.R., 1999b. Stromal–epithelial interactions in thecornea. Prog. Retin. Eye Res. 18, 293–309.
Wilson, S.E., Mohan, R.R., Hong, J., Lee, J., Choi, R., Liu, J.J., Mohan, R.R., 2002.Apoptosis in the cornea in response to epithelial injury: significance to woundhealing and dry eye. Adv. Exp. Med. Biol. 506, 821–826.
Wilson, S.E., Mohan, R.R., Mohan, R.R., Ambrosio Jr., R., Hong, J., Lee, J., 2001. Thecorneal wound healing response: cytokine-mediated interaction of theepithelium, stroma, and inflammatory cells. Prog. Retin. Eye Res. 20, 625–637.
Wilson, S.E., Walker, J.W., Chwang, E.L., He, Y.G., 1993. Hepatocyte growth factor,keratinocyte growth factor, their receptors, fibroblast growth factor receptor-2,and the cells of the cornea. Investig. Ophthalmol. Vis. Sci. 34, 2544–2561.
Wong, T.T., Sethi, C., Daniels, J.T., Limb, G.A., Murphy, G., Khaw, P.T., 2002. Matrixmetalloproteinases in disease and repair processes in the anterior segment.Surv. Ophthalmol. 47, 239–256.
Xue, M.L., Wakefield, D., Willcox, M.D., Lloyd, A.R., Di Girolamo, N., Cole, N.,Thakur, A., 2003. Regulation of MMPs and TIMPs by IL-1beta during cornealulceration and infection. Investig. Ophthalmol. Vis. Sci. 44, 2020–2025.
Yamada, J., Dana, M.R., Sotozono, C., Kinoshita, S., 2003. Local suppression of IL-1 byreceptor antagonist in the rat model of corneal alkali injury. Exp. Eye Res. 76,161–167.
Yan, L., Zucker, S., Toole, B.P., 2005. Roles of the multifunctional glycoprotein,EMMPRIN (basigin; CD147), in tumour progression. Thromb. Haemostasis 93,199–204.
Yurchenko, V., Constant, S., Bukrinsky, M., 2006. Dealing with the family: CD147interactions with cyclophilins. Immunology 117, 301–309.
Zieske, J.D., Guimaraes, S.R., Hutcheon, A.E., 2001. Kinetics of keratocyte prolifera-tion in response to epithelial debridement. Exp. Eye Res. 72, 33–39.
Zieske, J.D., Mason, V.S., Wasson, M.E., Meunier, S.F., Nolte, C.J., Fukai, N., Olsen, B.R.,Parenteau, N.L., 1994. Basement membrane assembly and differentiation ofcultured corneal cells: importance of culture environment and endothelial cellinteraction. Exp. Cell Res. 214, 621–633.
Zucker, S., Hymowitz, M., Rollo, E.E., Mann, R., Conner, C.E., Cao, J., Foda, H.D.,Tompkins, D.C., Toole, B.P., 2001. Tumorigenic potential of extracellular matrixmetalloproteinase inducer. Am. J. Pathol. 158, 1921–1928.
Related Documents









![RESEARCH ARTICLE Open Access Interleukins, laminin ......metastasis in NPC by inducing matrix metalloproteinases (MMPs) in degrading ECM proteins [5]. MMPs have the ability to digest](https://static.cupdf.com/doc/110x72/60d54502b6fa494b9302f93f/research-article-open-access-interleukins-laminin-metastasis-in-npc-by.jpg)

